 Hui Ma1,2
Hui Ma1,2 Gang Zhu
Gang Zhu- 1Center for Mental Health, Yanshan University, Qinhuangdao, China
- 2Department of Psychiatry, The First Affiliated Hospital of China Medical University, Shenyang, China
- 3Center for Mental Health, Hebei Institute of International Business and Economics, Qinhuangdao, China
- 4Department of Psychiatry, The Third Hospital of Heilongjiang Province, Bei'an, China
Aims: In the central nerve system, neurotensin (NT), and neurotensin receptor 1 (NTR1) modulate the dopamine system. Gene variations in the dopamine system have been demonstrated to influence certain defense mechanisms, but no studies have investigated possible effect of NTR1 gene polymorphisms in the biological determination of these defenses. The present study therefore examined this link.
Methods: In 412 healthy Han Chinese, single nucleotide polymorphisms rs6090453C/G, rs6011914C/G, and rs2427422A/G of the NTR1 gene were genotyped, and the defense mechanisms were measured by the self-reporting Defense Style Questionnaire 88.
Results: Significant male-specific differences in the projective identification among the rs6090453 genotypes (p = 0.003); in the intermediate defense, reaction formation, and projective identification among the rs6011914 genotypes (p = 0.011, 0.010, and 0.011, respectively); and in the projective identification among the rs2427422 genotypes (p = 0.005) were found when the level of significance was adjusted by the Bonferroni correction. There was no significant difference in any of the defense scores among genotypes of any single nucleotide polymorphism in the total cohort or female subjects (all p > 0.017). The distributions of genotypes between the low and high score subgroups showed significant differences in the rs2427422 genotype distributions for help-rejecting complaining, regression, and projective identification (p = 0.010, 0.022, and 0.044, respectively). Significant differences were found between males and females in 10 defense mechanisms (all P < 0.05).
Conclusions: The gene variations in the NTR1 polymorphisms were involved in the biological mechanisms of intermediate defense mechanisms, and this effect was influenced by sex.
Introduction
The concept of the psychological defense mechanism is one of the cores in the psychoanalytic theory, which was first proposed by Freud (1). With the establishment and development of the personality structure model (id, ego, and superego) in the psychoanalytic school, the concept of the defense mechanism changed constantly (2). In the Diagnostic and Statistical Manual of Mental Disorders-Fourth Edition, defense mechanisms were defined as “automatic psychological processes that protect the individual against anxiety and from the awareness of internal or external dangers or stressors. Individuals are often unaware of these processes as they operate” (3). Psychological defense mechanisms represent a crucial component of individual capacity to maintain emotional homeostasis. Without them the conscious mind would be much more vulnerable to negatively charged emotional input, such as that pertaining to anxiety and sadness (4, 5).
In fact, until defense-related scales were developed, it had always been difficult to study defense mechanisms since they always worked at the unconscious level. Bond et al. developed and revised the self-reporting Defense Style Questionnaire 88 (DSQ-88) to assess possible conscious derivatives of defense mechanisms and to elicit manifestations of a subject's characteristic style of dealing with conflict, either consciously or unconsciously, based on the assumption that the subject was able to accurately comment on his/her behavior from a distance (6). The authors provided evidence for testing the reliability of the DSQ-88 by assessing psychiatric patients and healthy subjects (7–9). Since then, this questionnaire has been applied to a large body of clinical research (10–16).
The relationship of defense mechanisms with the psychopathology of mental disorders and abnormal behaviors is not been clearly understood, and in cases conflicting (17). Nevertheless, most studies have indicated strong evidence that adaptation of defense style correlated with mental health and that some diagnoses were correlated with specific defense patterns, for example, patients with depression or personality disorders tended to use more maladaptive defenses and less adaptive defenses while patients with anxiety disorders in general tended to use more immature and neurotic defenses (18, 19). Therefore, it is important and necessary to elucidate the determination of defenses.
There have been many previous studies on the social psychological factors of defense mechanisms in psychology; however, few have focused on biological factors. Moreover, the majority of previous studies focused on psychoanalytic and not statistical investigations (2). Using the data from twin-based studies, Andrews conducted a multivariate genetic analysis of three factors, including trait anxiety, locus of control and defense style, and demonstrated that defense style was substantially influenced by genetic factors and there was a significant loading (0.44) on a common genetic factor contributing to the variance of individual defenses (20). Therefore, biogenetic factors play a role in the determination of defense mechanisms. It is known that defense mechanisms are an important and enduring facet of personality (8) and that the dopamine system is closely related to personality traits (21–24). It was thus speculated that the dopamine system was associated with defenses. Two studies confirmed this hypothesis: Coming et al. demonstrated that the dopamine D2 receptor gene (DRD2) locus was the site of one of the factors that control defenses (25) and Huang et al. demonstrated that the PPP1R1B gene, encoding the dopamine- and cAMP-regulated phosphoprotein (DARPP-32), was one of the factors that was responsible for defenses (26).
In the central nervous system, neurotensin (NT) is a 13-amino acid multifunctional neurotransmitter and neuromodulator. Among the three NT receptor (NTR) subtypes, NTR1 and NTR2 are G protein-coupled receptors, and NTR1 has a much higher affinity for NT than NTR2 (27, 28). The NTR1 gene is located on the 20q13 locus and consists of four exons and three introns (29). A large amount of anatomical, physiological, pharmacological and behavioral evidence has demonstrated that NT transmission modulates central dopaminergic functions (30–33). Therefore, the genes of the NT system may contribute to defense mechanisms by regulating dopamine neurotransmission. Moreover, our team has already demonstrated an association between NTR1 gene single nucleotide polymorphisms (SNPs) and personality traits assessed by the Tridimensional Personality Questionnaire (TPQ) in Chinese Han subjects (22, 34). Therefore, we decided to test the hypothesis that the NTR1 SNPs were associated with defenses.
To our knowledge, none of the previous reports have found an association between NTR1 gene polymorphisms and defense mechanisms measured by DSQ. In this study, we investigated whether genetic variants in the NTR1 gene were associated with defenses in a large healthy Chinese Han population, and then analyzed any association by sex. The results would provide empirical evidence for the biological factors of defense mechanism. The three SNPs investigated (rs6090453, rs6011914, and rs2427422, Figure 1) were selected due to our previous research on personality traits (22, 34), copying styles (35), anxiety (36), schizophrenia (37), and alcohol dependence (38).
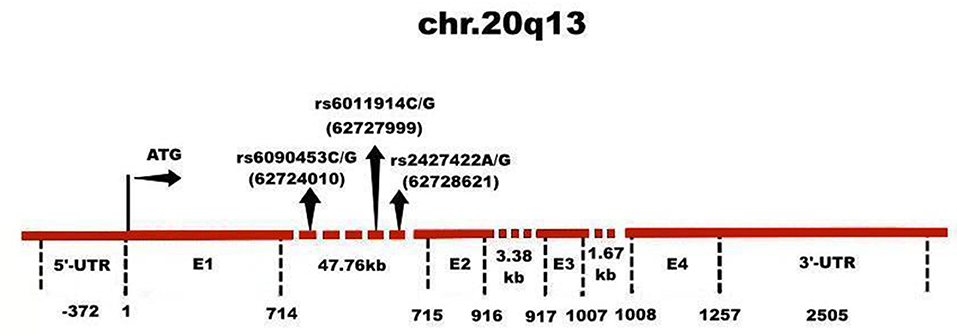
Figure 1. Genomic structure of NTR1 gene. The NTR1 gene has 4 exons (E) and 3 introns, whose sizes are indicated. The rs6090453C/G, rs6011914C/G, and rs2427422A/G polymorphisms of the NTR1 gene are located in intron 1, whose locations are indicated.
Materials and Methods
Subjects
All subjects were unrelated healthy Chinese-Han volunteers (n = 412, 196 males, 216 females) without psychiatric, neurological, or chronic physical illnesses. The age range was 19–58 years, while the mean ± standard deviation (SD) was 31.43 ± 8.16 years. There was no significant difference in age between the males and females (p = 0.187; Supplementary Table 1). The volunteers were recruited from healthy undergraduate and graduate students and staff of China Medical University, Shenyang, Liaoning Province, China, as well as healthy individuals undergoing physical examinations at the First Affiliated Hospital of China Medical University. Written informed consent was obtained from all participants. All the protocols in the present study were approved by the Ethics Committee of China Medical University (number of ethical approval: 2019-209-2).
Bond's DSQ-88 was translated and validated into the Chinese version in 1993, with no differences in meaning or content to that of the original version. The Chinese version of DSQ-88 evaluates 24 defense styles, which are classified into three factors of defense: immature or maladaptive (projection, passive aggression, acting out, help-rejecting complaining, fantasy, splitting, regression, and somatization), intermediate or neurotic (reaction formation, undoing, inhibition, withdrawal, idealization, pseudo-altruism, omnipotence, isolation, projective identification, denial, affiliation, consumption, and anticipation), and mature or adaptive (sublimation, suppression, and humor). All participants were asked to complete the questionnaire by themselves within 40 min and to check that all items had been scored. Following the completion of the questionnaire, 2 mL of venous blood from each participant was obtained for genotyping. Each defense style score was represented by the average of all the items representing the particular defense mechanism, while each defense factor score was derived from the mean of all the defense style scores belonging to the defense factor. According to the study of Bond et al. (6), a subject was considered to score high on a particular defense style and use it if his score was 0.5 SD above the mean on the particular defense. A cutting point of 0.5 SD provided the best discrimination between those who use a defense style and those who do not (6). Thus, subjects could be divided into low score (non-use) and high score (use) subgroups for each defense style based on whether the scores were above or below the cutting point.
Polymorphism Genotyping
Amplification of gene fragments containing the SNPs rs6090453, rs6011914, and rs2427422 by polymerase chain reaction and subsequent identification of genotypes by restriction fragment length polymorphism analysis were carried out as described in our previous studies (22, 34–38).
Statistical Analysis
Data are presented as the mean ± SD, frequency, or percentage. Data statistical analyses were conducted using SPSS® version 17.0 software (SPSS Inc., Chicago, IL, USA). Potential genotypic associations of the three SNPs with DSQ-scores were detected by one-way analysis of variance (ANOVA) or by the non-parametric Kruskal-Wallis test, depending on homogeneity or heterogeneity of variance of data. Separate analysis by gender was also carried out. The Hardy-Weinberg equilibrium (HWE) and the differences in genotype frequencies between the low and high score subgroups were assessed by chi-square test. The differences in defense mechanisms between male and female subjects were compared by independent sample t-test. Linkage disequilibrium (LD) and haplotype analysis of the three NTR1 gene polymorphisms were carried out using the free online software Haploview version 4.2 (http://www.broad.mit.edu/mpg/haploview). A P < 0.05 was regarded as statistically significant.
Results
HWE Results
The numbers of subjects with rs6090453 genotype CC, CG, and GG were 43, 177, and 192, respectively. The numbers of subjects with rs6011914 genotype GG, CG, and CC were 204, 176, and 32, respectively. The numbers of subjects with rs2427422 genotype GG, AG, and AA were 222, 168, and 22, respectively. The genotype distribution of the SNPs rs6090453 (χ2 = 0.054, p = 0.816), rs6011914 (χ2 = 0.496, p = 0.481), and rs2427422 (χ2 = 1.847, p = 0.174) did not deviate from the HWE in the experimental sample of 412 Chinese Han subjects (Supplementary Table 2). Moreover, for all three SNPs, the genotype frequencies were similar to those observed in other samples from the Han Chinese population (http://www.ncbi.nlm.nih.gov) (all p > 0.05). Therefore, this study sample was considered to be representative of the general Chinese Han population.
LD and Haplotype Analysis Results
As shown in Figure 2, LDs between rs6090453C/G and rs6011914C/G (D′ = 0.89, r2 = 0.69), between rs6090453C/G and rs2427422A/G (D′ = 0.92, r2 = 0.63), and between rs6011914C/G and rs2427422A/G (D′ = 0.99, r2 = 0.83) were demonstrated.
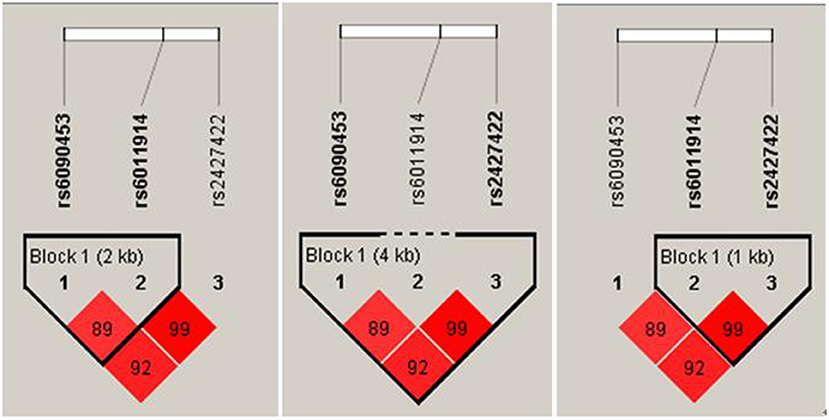
Figure 2. Linkage disequilibrium plots for the three analyzed NTR1 SNPs in healthy Chinese.
The frequencies of haplotypes composed of the alleles in the three SNPs were shown in Table 1. The haplotype blocks with low frequencies (<1%) were rejected. The two most frequent haplotypes were GGG (0.659) and CCA (0.243). None of the frequencies of these five haplotypes differed significantly between the male and female samples (all p > 0.05).
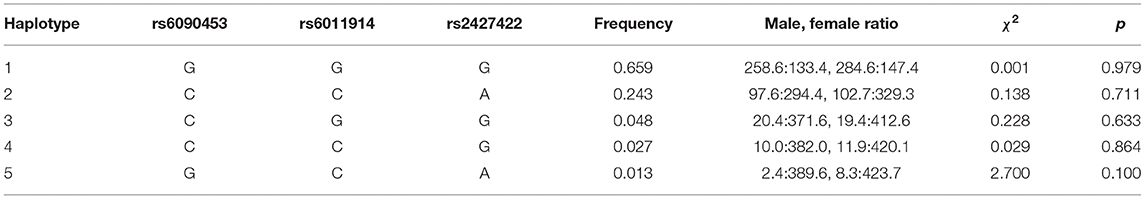
Table 1. Distribution of haplotypes for the three analyzed NTR1 SNPs between males and females.
Effects of NTR1 Gene Polymorphisms on DSQ Scores in the Total Cohort and in Male and Female Subjects
As shown in Table 2, all DSQ dimension scores among three genotypes of each SNP were compared. There was a marginal difference only in the score of help-rejecting complaining among the three genotypes of the rs2427422 polymorphism (F = 3.028, p = 0.050), but this difference was not significant when the level of significance was adjusted using Bonferroni correction [α′ = α/k, where α is the unadjusted significance level for pair-wise comparisons, k is the number of independent significance tests, and α′ (0.05/3 = 0.017) is the corrected significance level for multiple comparisons].
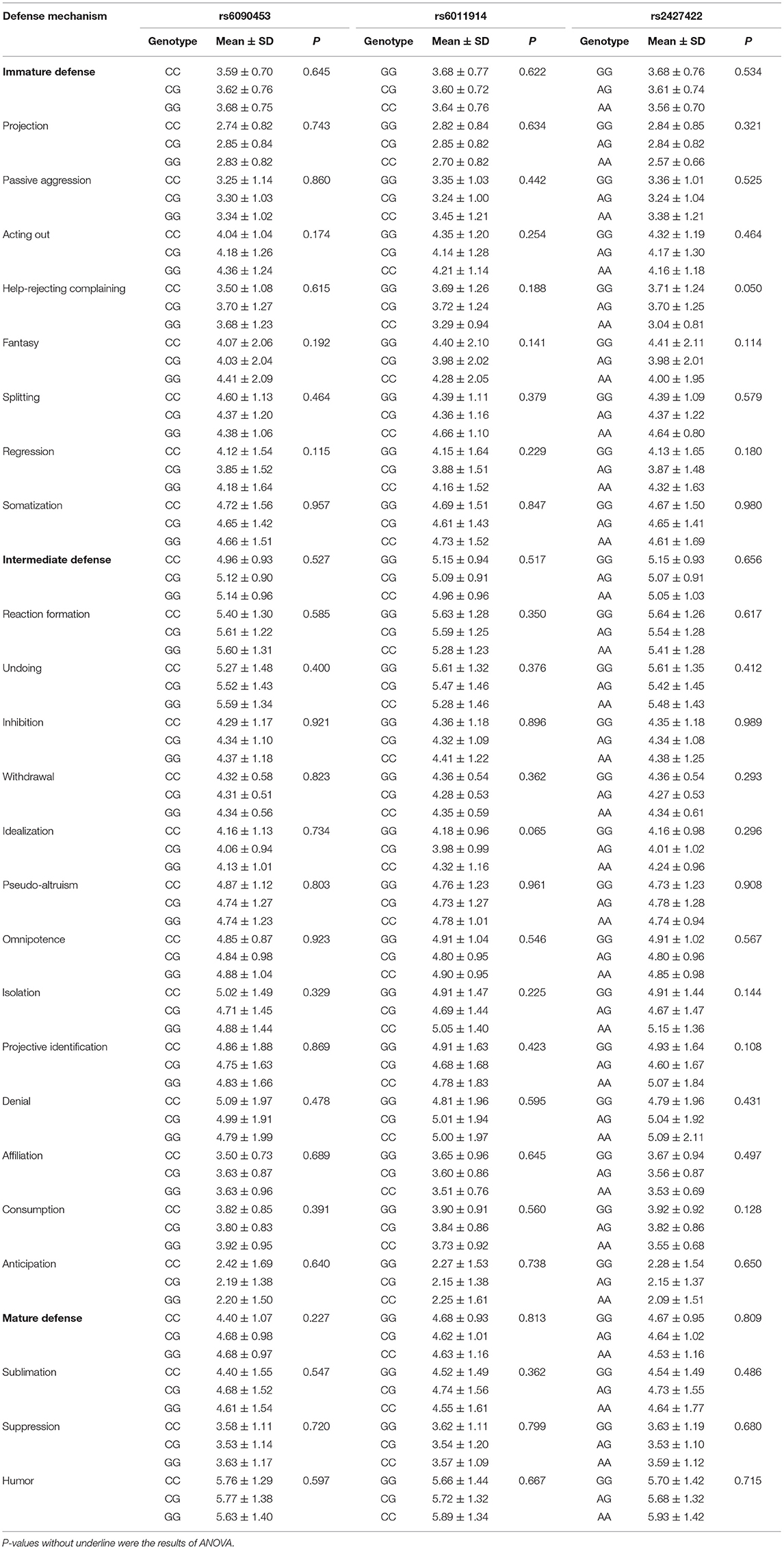
Table 2. Effects of NTR1 gene polymorphisms on DSQ scores.
The differences in all DSQ dimension scores among the genotypes of the three polymorphisms were then analyzed separately by sex. We demonstrated significant male-specific differences in the intermediate defense, reaction formation, projective identification, affiliation, mature defense, sublimation, and suppression scores among the rs6090453 genotypes (F = 3.445, 3.131, 5.982, 4.023, 3.552, 3.892, and 3.061, respectively; p = 0.034, 0.046, 0.003, 0.019, 0.031, 0.022, and 0.049, respectively); in the intermediate defense, reaction formation, undoing, and projective identification scores among the rs6011914 genotypes (F = 4.651, 4.760, 3.999, and 4.631, respectively; p = 0.011, 0.010, 0.020, and 0.011, respectively); and in the projective identification score among the rs2427422 genotypes (F = 5.433, p = 0.005; Table 3). Moreover, the male-specific differences in the projective identification score among the rs6090453 genotypes, in the intermediate defense, reaction formation, and projective identification scores among the rs6011914 genotypes, and in the projective identification score among the rs2427422 genotypes were still significant (all P < 0.017) when the level of significance was adjusted by the Bonferroni correction.
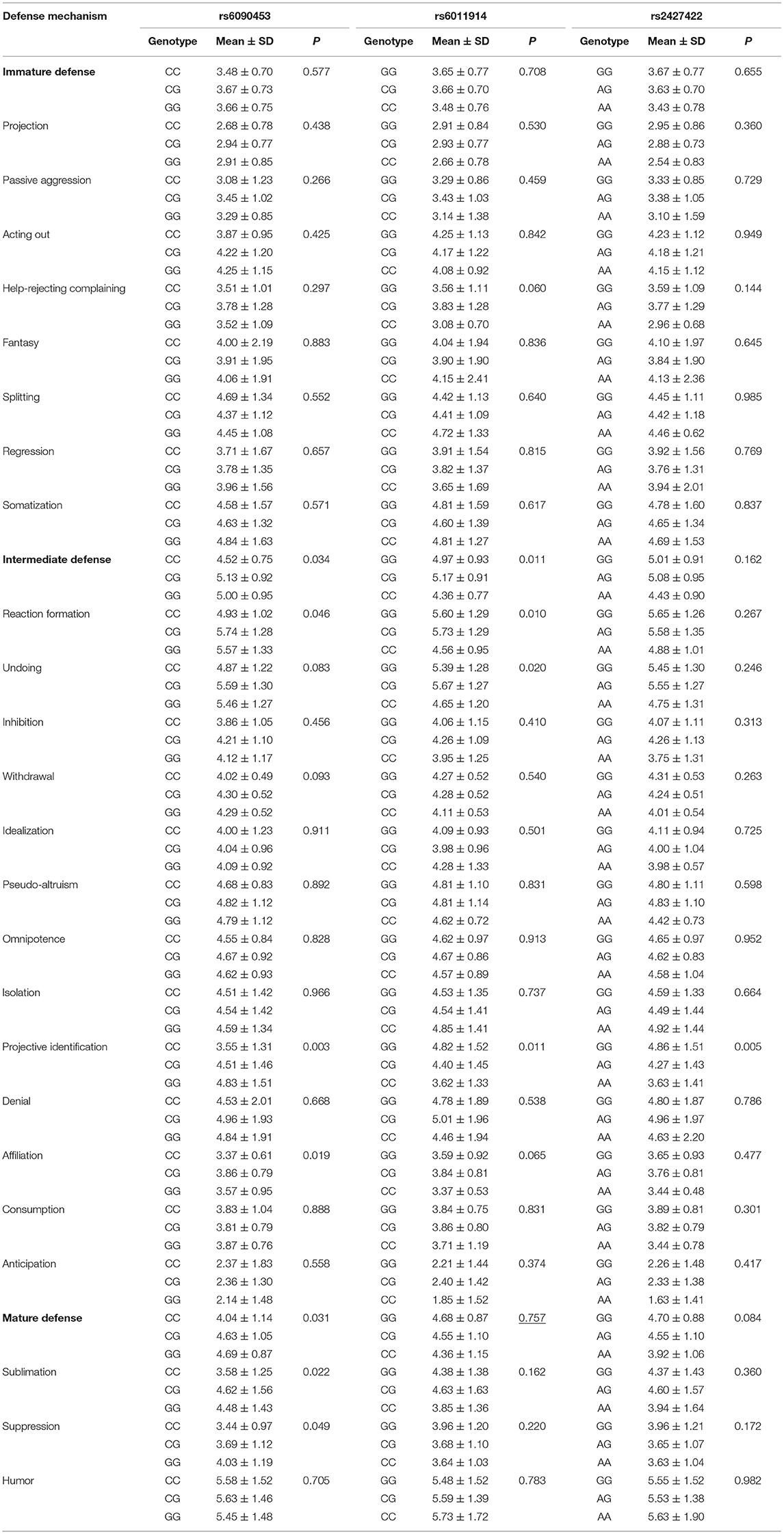
Table 3. Effects of NTR1 gene polymorphisms on DSQ scores in males.
At the same time, we demonstrated significant female-specific difference in the projective identification score among between the rs6090453 genotypes (F = 3.580, p = 0.030), and in the passive aggression, intermediated defense, and affiliation scores among the rs6011914 genotypes (F = 3.775, 3.080, and 3.273, respectively; p = 0.024, 0.048, and 0.040, respectively; Table 4). However, all these female-specific differences in defense scores among the genotypes of the three SNPs were not significant when the level of significance was adjusted by the Bonferroni correction.
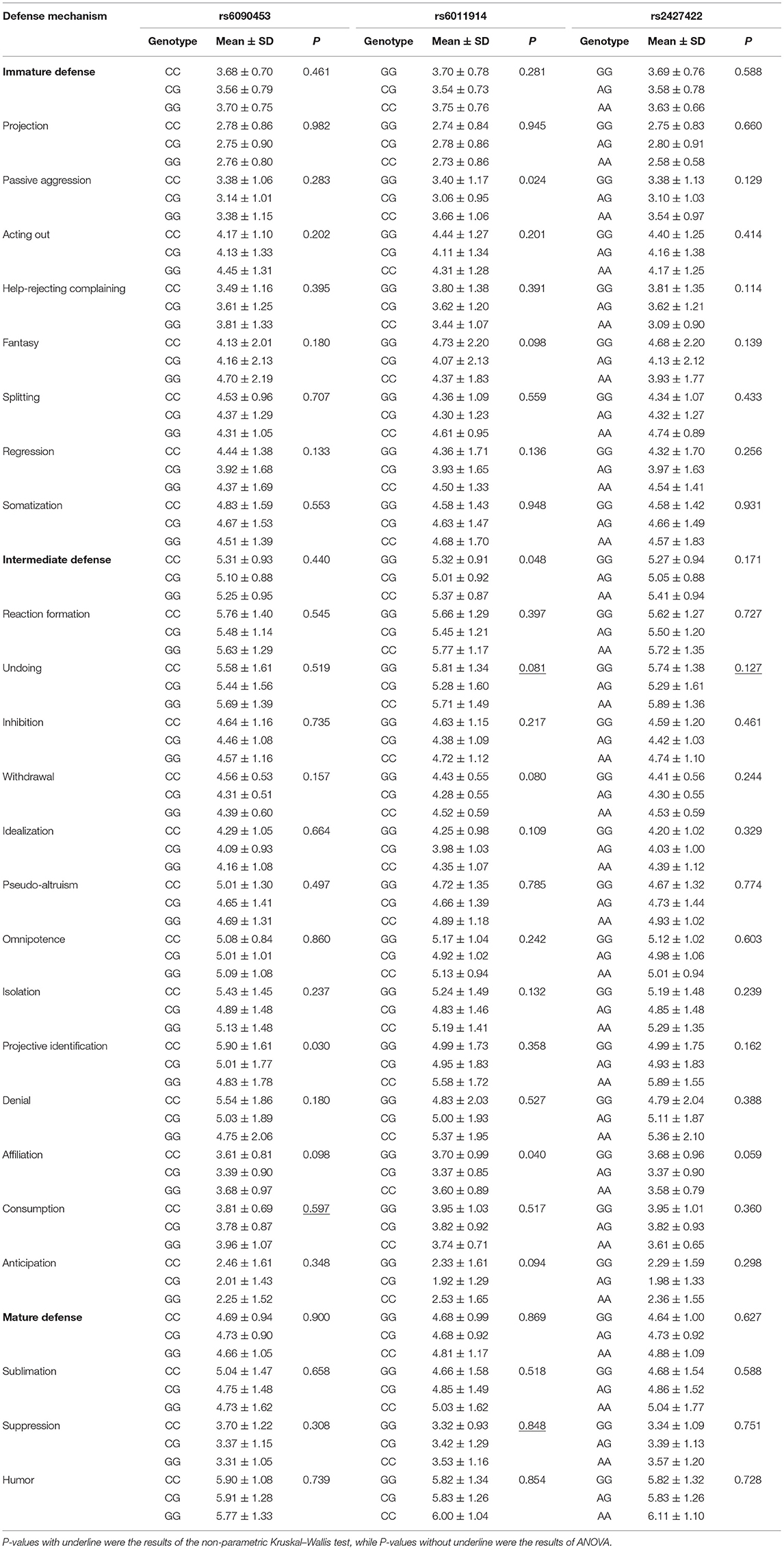
Table 4. Effects of NTR1 gene polymorphisms on DSQ scores in females.
Comparison of the Genotype Distributions Between the High and Low Score Subgroups
As shown in Table 5, the genotypes of the three SNPs between the high and low score subgroups for three defense factors and 24 defense styles were compared. We found significant differences in the rs2427422 genotype distributions between two subgroups for help-rejecting complaining, regression, and projective identification (χ2 = 9.182, 7.665, and 6.236, p = 0.010, 0.022, and 0.044, respectively).
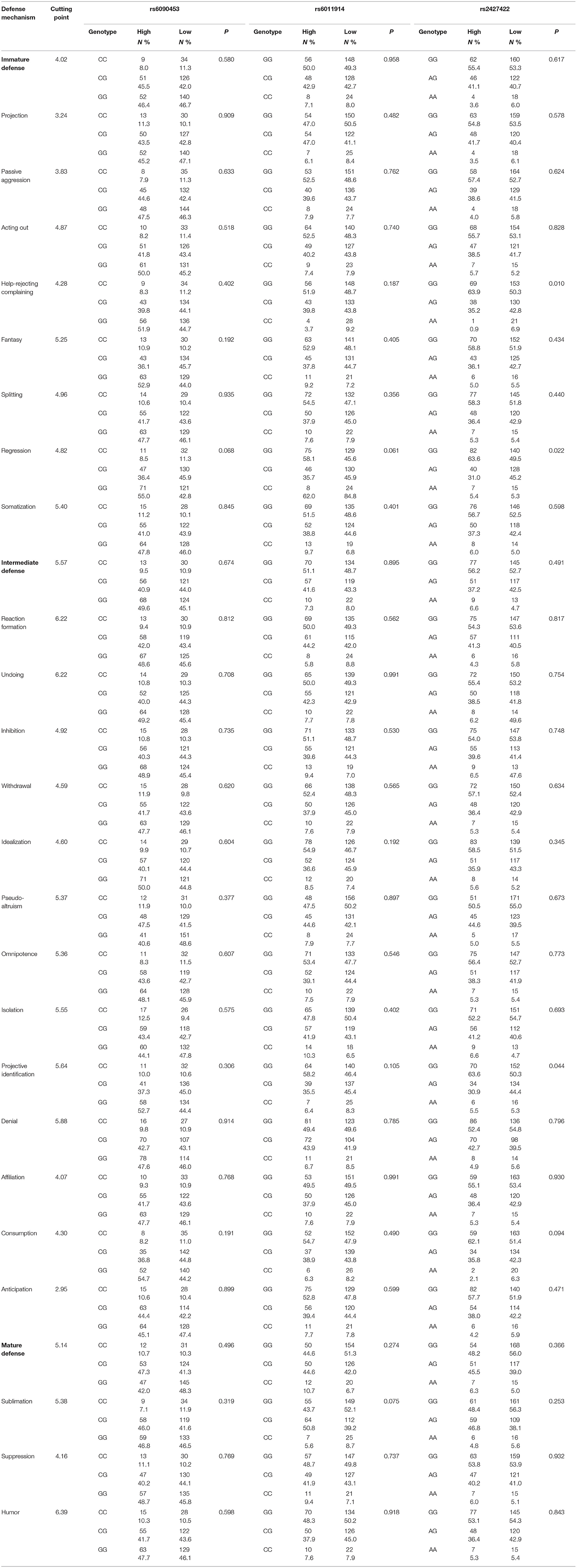
Table 5. Comparison of genotype distributions of NTR1 gene polymorphisms between high- and low-score subgroups.
Comparison of DSQ Scores Between Male and Female Subjects
When the DSQ scores were compared between male and female subjects, significant differences were found in 10 defense styles including fantasy, regression, inhibition, withdrawal, omnipotence, isolation, projective identification, sublimation, suppression, and humor (Table 6, all P < 0.05). The suppression score for males was higher than that for females, while the scores of other nine defense styles were higher for females.
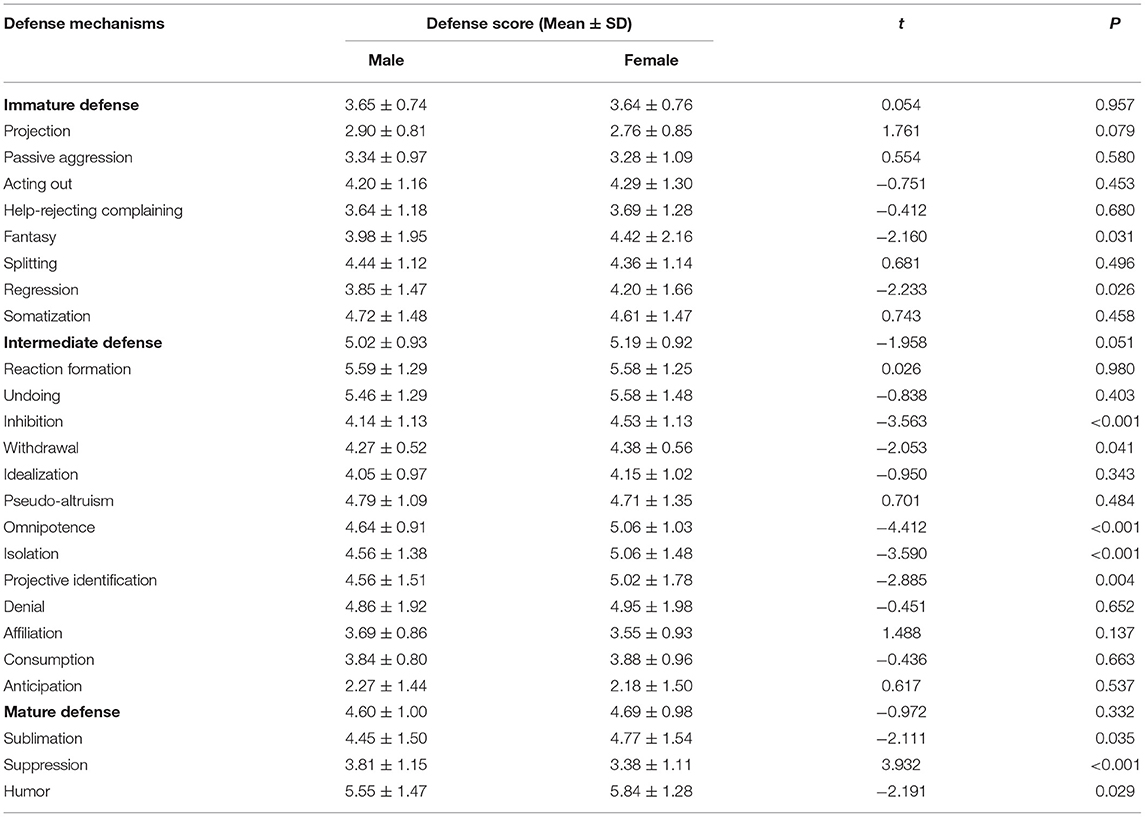
Table 6. Comparison of defense mechanisms between male and female subjects.
Discussion
The main finding of the present study was that NTR1 gene SNPs were significantly associated with two particular defense styles (reaction formation and projective identification) and intermediate defense factors to which these two defense styles belong, in healthy Han-Chinese subjects, and that this association was specific to males. On the part of an association between the NTR1 gene and DSQ-measured defenses, any relevant research reports were not be found before the present study.
Defense mechanisms have been considered to result from the interaction of biological and environmental factors (20, 26), but the research about the biogenetic basis for defenses has been rarely studied. Generally, defense mechanisms are considered to be an enduring and important dimension of personality and not just an epiphenomena of psychopathology (8). Many previous studies have demonstrated a significant association between personality traits measured by TPQ and alleles encoding dopamine receptors (DRs) and molecules mediating the synthesis, metabolism, and transport of dopamine, for example, functional variants in the D2DR (39, 40), the dopamine D4 receptor (D4DR) (40–43), the catechol-O-methyltransferase (COMT) (42, 44, 45), and the monoamine oxidase A (MAOA) genes (44, 46). Direct studies regarding the dopamine biogenetic basis of defenses measured by DSQ have been very rare. Coming et al. demonstrated that subjects with the DRD2 gene haplotype 1 tended to show a decrease in mature and an increase in neurotic and immature, defense styles compared with those without the haplotype 1, which suggested that the DRD2 locus was one of the factors controlling defense styles (25). Huang et al. demonstrated that the PPP1R1B gene, encoding DARPP-32, was one of the factors responsible for defenses, because PPP1R1B polymorphisms were found to be associated with immature defenses (26). Gene variants in the dopamine system might thus be associated with defense mechanisms.
Previous studies have concluded strong evidence for a regulatory role of NT in the dopamine system. Animal and human radioimmunoassay studies have found that NT was present in all the mammalian brain structures containing dopamine nerve cell bodies and terminals (30, 47). Moreover, NTR1 localized both pre-synaptically and post-synaptically at dopaminergic synapses expressing DRD2 (48). The anatomical overlap between NT, NTR1, dopamine, and DRD2 suggests possible interactions that regulate neurophysiological functions at the cellular level. Numerous studies have demonstrated that NT could regulate the affinity of DRs for dopamine and dopamine receptor agonists (30, 32, 49–51), in addition to its action on the excitability of dopaminergic neurons (33, 52–54). In the present study, we demonstrated that three NTR1 gene polymorphisms were significantly associated with projective identification and that the rs6011914 polymorphism was significantly associated with reaction formation in healthy Han-Chinese males. We also demonstrated significant differences in genotype distributions of the rs2427422 polymorphism between the low and high score subgroups for three defense styles: help-rejecting complaining, regression, and projective identification. These results were consistent with and further confirmed the previous study conclusion about the significant association between gene variants in the dopamine system and defense mechanisms (25, 26).
Bond reviewed the published studies about the relationships of defense styles with psychopathology and change and proposed that defense styles should become more adaptive with improvement in symptoms, but intermediate defenses tended to become stable over time (19). Recently, Hayden et al. also demonstrated that maladaptive defense mechanisms were significantly reduced during inpatient therapy and remained low until follow-up, whereas neurotic (intermediate) and adaptive defense mechanisms did not change significantly (5). In other words, intermediate defenses have been found to be a trait-like factor among the three defense factors. In a previous study, our team uncovered a particular defense style (undoing) of intermediate defense, which was demonstrated to be associated with FYN gene polymorphisms (55). In the present study, both reaction formation and projective identification, which were demonstrated to be significantly associated with NTR1 gene polymorphisms, were also classified to be intermediate defenses. Therefore, these results might account, at least partially, for the stability of intermediate defenses, and also indicate that relatively stable biogenetic factors including the dopamine system played an important role in the determination of these intermediate defenses.
Moreover, the present study found significant differences among different genotypes in six particular defense styles and intermediate and mature defenses in males, and in three particular defense styles and intermediate defense in females. Interestingly, after the Bonferroni correction, only the differences in the projective identification score among the rs6090453 genotypes; in the intermediate defense, reaction formation, and projective identification scores among the rs6011914 genotypes; and in the projective identification score among the rs2427422 genotypes were still significant in males. The association between NTR1 gene polymorphisms and defenses was thus a sex-specific result. In fact, several previous studies have demonstrated significant differences in defenses between the two genders (56–58), and in agreement, we found significant differences in 10 particular defense styles between male and female subjects in the present study. Moreover, the important effect of the interaction between gender and gene polymorphisms has already been demonstrated on individual differences in personality traits (59–61). Therefore, it was expected that our findings were mainly restricted to men. In the present study, we have demonstrated that no significant difference in the frequencies of five haplotypes composed of the alleles in the three NTR1 SNPs existed between the males and females, indicating no sex-specific difference in the haplotype frequency of NTR1 SNPs. Therefore, the present sex-specific findings may be the result of androgens, which might be involved in the effect of the NTR1 gene on neurotransmitter systems, and thus ultimately affect the biological determination of defenses. Biochemical experiments will be needed to determine if this conjecture is indeed true.
Conclusion
We investigated the associations between three NTR1 polymorphisms and defense mechanisms measured by DSQ-88 in a large healthy Chinese Han population to demonstrate possible biogenetic mechanisms affecting defenses. We found significant associations in male subjects between three SNPs and the defense style of projective identification as well as the NTR1 rs6011914 polymorphism and the defense style of reaction formation. Therefore, our results provide evidence that gene variants in the NT system can influence the formation and development of defense mechanisms by regulating the dopamine system and that this effect is affected by gender. However, this conclusion should be considered cautiously because of the limited sample size and number of studied SNPs, and the use of a self-rating scale. Further studies of these NTR1 SNPs, as well as other SNPs associated with the dopamine system in larger Chinese Han and other ethnic populations are needed to verify our results and to gain a more comprehensive understanding of the effects of NTR1 allelic variants on defenses.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics Statement
The studies involving human participants were reviewed and approved by the Ethics Committee of China Medical University. The patients/participants provided their written informed consent to participate in this study.
Author Contributions
HM and GZ: conceptualization. HM and ML: methodology. HM and JT: software, formal analysis, data curation, and visualization. ML and LZ: validation. HM and LZ: investigation. GZ: resources, supervision, project administration, and funding acquisition. HM and ML: writing—original draft preparation. HM, ML, JT, and GZ: writing—review and editing. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by a Grant from the Major Project of the Department of Science & Technology of Liaoning Province (2019JH8/10300019).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer AA declared a shared affiliation, with no collaboration, with the authors GZ and ML at the time of the review.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors thank all the volunteers for taking part in this study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyt.2021.762276/full#supplementary-material
References
1. Freud S. The Neuro-Psychosis of Defense (The Standard Edition of the Complete Psychological Works of Sigmund Freud). London: The Hogarth Press and the Institute of Psycho-Analysis (1894).
2. Mihalits DS, Codenotti M. The conceptual tragedy in studying defense mechanisms. Integr Psychol Behav Sci. (2020) 54:354–69. doi: 10.1007/s12124-020-09515-6
3. American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition (DSM-IV). Washington, DC: American Psychiatric Press (1994).
4. Bowins B. Psychological defense mechanisms: a new perspective. Am J Psychoanal. (2004) 64:1–26. doi: 10.1023/B:TAJP.0000017989.72521.26
5. Hayden MC, Mullauer PK, Beyer KJP, Gaugeler R, Senft B, Dehoust MC, et al. Increasing mentalization to reduce maladaptive defense in patients with mental disorders. Front Psychiatry. (2021) 12:637915. doi: 10.3389/fpsyt.2021.637915
6. Bond M, Gardner ST, Christian J, Sigal JJ. Empirical study of self-rated defense styles. Arch Gen Psychiatry. (1983) 40:333–8. doi: 10.1001/archpsyc.1983.01790030103013
7. Bond MP, Vaillant JS. An empirical study of the relationship between diagnosis and defense style. Arch Gen Psychiatry. (1986) 43:285–8. doi: 10.1001/archpsyc.1986.01800030103012
8. Vaillant GE, Bond M, Vaillant CO. An empirically validated hierarchy of defense mechanisms. Arch Gen Psychiatry. (1986) 43:786–94. doi: 10.1001/archpsyc.1986.01800080072010
9. Bond M, Christopher J, Gautier M, Goldenberg M, Oppenheimer J, Simand J. Validating the self-report of defense styles. J Pers Disord. (2011) 3:101–12. doi: 10.1521/pedi.1989.3.2.101
10. Bonsack C, Despland JN, Spagnoli J. The French version of the Defense Style Questionnaire. Psychother Psychosom. (1998) 67:24–30. doi: 10.1159/000012255
11. Bond M, Perry JC. Long-term changes in defense styles with psychodynamic psychotherapy for depressive, anxiety, and personality disorders. Am J Psychiatry. (2004) 161:1665–71. doi: 10.1176/appi.ajp.161.9.1665
12. Sukul YR, Birkenhager TK, van den Broek WW, Mulder PG, Bruijn JA. Trait anxiety and defensive functioning in relation to antidepressant treatment outcome. Psychopathology. (2009) 42:387–93. doi: 10.1159/000241194
13. Hyphantis T. The Greek version of the Defense Style Questionnaire: psychometric properties in three different samples. Compr Psychiatry. (2010) 51:618–29. doi: 10.1016/j.comppsych.2010.03.005
14. Zanarini MC, Frankenburg FR, Fitzmaurice G. Defense mechanisms reported by patients with borderline personality disorder and axis II comparison subjects over 16 years of prospective follow-up: description and prediction of recovery. Am J Psychiatry. (2013) 170:111–20. doi: 10.1176/appi.ajp.2012.12020173
15. Jun JY, Lee YJ, Lee SH, Yoo SY, Song J, Kim SJ. Association between defense mechanisms and psychiatric symptoms in North Korean Refugees. Compr Psychiatry. (2015) 56:179–87. doi: 10.1016/j.comppsych.2014.10.001
16. Lenzo V, Barberis N, Cannavo M, Filastro A, Verrastro V, Quattropani MC. The relationship between alexithymia, defense mechanisms, eating disorders, anxiety and depression. Riv Psichiatr. (2020) 55:24–30. doi: 10.1708/3301.32715
17. Brody S, Costa RM. Rationalization is a suboptimal defense mechanism associated with clinical and forensic problems. Behav Brain Sci. (2020) 43:e31. doi: 10.1017/S0140525X19002073
18. Waqas A, Rehman A, Malik A, Muhammad U, Khan S, Mahmood N. Association of ego defense mechanisms with academic performance, anxiety and depression in medical students: a mixed methods study. Cureus. (2015) 7:e337. doi: 10.7759/cureus.337
19. Bond M. Empirical studies of defense style: relationships with psychopathology and change. Harv Rev Psychiatry. (2004) 12:263–78. doi: 10.1080/10673220490886167
20. Andrews G. Anxiety, personality and anxiety disorders. Int Rev Psychiatry. (1991) 3:293–302. doi: 10.3109/09540269109110408
21. Li J, Ma H, Zhou H, Huang Y, Wu L, Li J, et al. Association between DARPP-32 gene polymorphism and personality traits in healthy Chinese-Han subjects. J Mol Neurosci. (2011) 44:48–52. doi: 10.1007/s12031-011-9505-7
22. Ma H, Huang Y, Zhang B, Jin L, Cong Z, Wang Y, et al. Neurotensin receptor 1 gene polymorphisms are associated with personality traits in healthy Chinese individuals. Neuropsychobiology. (2014) 69:11–8. doi: 10.1159/000356966
23. Delvecchio G, Bellani M, Altamura AC, Brambilla P. The association between the serotonin and dopamine neurotransmitters and personality traits. Epidemiol Psychiatr Sci. (2016) 25:109–12. doi: 10.1017/S2045796015001146
24. Lhommee E, Boyer F, Wack M, Pelissier P, Klinger H, Schmitt E, et al. Personality, dopamine, and Parkinson's disease: insights from subthalamic stimulation. Mov Disord. (2017) 32:1191–200. doi: 10.1002/mds.27065
25. Comings DE, MacMurray J, Johnson P, Dietz G, Muhleman D. Dopamine D2 receptor gene (DRD2) haplotypes and the defense style questionnaire in substance abuse, Tourette syndrome, and controls. Biol Psychiatry. (1995) 37:798–805. doi: 10.1016/0006-3223(94)00222-O
26. Huang Y, Li J, Ma H, Zhao X, Wang Y, Jin Q, et al. Association between PPP1R1B polymorphisms and defense mechanisms in healthy Chinese-Han subjects. J Mol Neurosci. (2013) 49:618–24. doi: 10.1007/s12031-012-9907-1
27. Kitabgi P. Targeting neurotensin receptors with agonists and antagonists for therapeutic purposes. Curr Opin Drug Discov Devel. (2002) 5:764–76. doi: 10.1016/S0149-2918(02)80052-2
28. Boules M, Shaw A, Fredrickson P, Richelson E. Neurotensin agonists: potential in the treatment of schizophrenia. CNS Drugs. (2007) 21:13–23. doi: 10.2165/00023210-200721010-00002
29. Le F, Groshan K, Zeng XP, Richelson E. Characterization of the genomic structure, promoter region, and a tetranucleotide repeat polymorphism of the human neurotensin receptor gene. J Biol Chem. (1997) 272:1315–22. doi: 10.1074/jbc.272.2.1315
30. Binder EB, Kinkead B, Owens MJ, Nemeroff CB. Neurotensin and dopamine interactions. Pharmacol Rev. (2001) 53:453–86. doi: 10.1006/phrs.2001.0891
31. St-Gelais F, Jomphe C, Trudeau LE. The role of neurotensin in central nervous system pathophysiology: what is the evidence? J Psychiatry Neurosci. (2006) 31:229–45. doi: 10.1016/j.jphysparis.2006.09.012
32. Laszlo K, Peczely L, Kovacs A, Zagoracz O, Ollmann T, Kertes E, et al. The role of intraamygdaloid neurotensin and dopamine interaction in conditioned place preference. Behav Brain Res. (2018) 344:85–90. doi: 10.1016/j.bbr.2018.01.021
33. Plach M, Schafer T, Borroto-Escuela DO, Weikert D, Gmeiner P, Fuxe K, et al. Differential allosteric modulation within dopamine D2R - neurotensin NTS1R and D2R - serotonin 5-HT2AR receptor complexes gives bias to intracellular calcium signalling. Sci Rep. (2019) 9:16312. doi: 10.1038/s41598-019-52540-8
34. Ma H, Cong Z, Zhang H, Tao J, Yang C, Song Z. Preliminary study of personality traits in Chinese lung cancer patients: modification by neurotensin receptor 1 polymorphisms. Asia Pac Psychiatry. (2017) 9:e12436. doi: 10.1111/appy.12268
35. Zhang B, Ma H, Huang Y, Li J, Wang Y, Jin Q, et al. No association of neurotensin receptor 1 gene polymorphisms with coping styles in healthy Chinese-Han individuals. Psychiatr Genet. (2013) 23:181–2. doi: 10.1097/YPG.0b013e32836130c4
36. Ma H, Huang Y, Zhang B, Li J, Wang M, Wang Y, et al. No association of neurotensin receptor 1 (NTR1) gene polymorphisms with either trait or state anxiety in healthy Chinese-Han participants. Psychiatr Genet. (2013) 23:93–4. doi: 10.1097/YPG.0b013e32835d71a5
37. Ma H, Huang Y, Zhang B, Li J, Wang Y, Zhao X, et al. Association between neurotensin receptor 1 (NTR1) gene polymorphisms and schizophrenia in a Han Chinese population. J Mol Neurosci. (2013) 50:345–52. doi: 10.1007/s12031-013-9988-5
38. Ma H, Huang Y, Zhang B, Wang Y, Zhao H, Du H, et al. Association between neurotensin receptor 1 gene polymorphisms and alcohol dependence in a male Han Chinese population. J Mol Neurosci. (2013) 51:408–15. doi: 10.1007/s12031-013-0041-5
39. Teh LK, Izuddin AF, M HF, Zakaria ZA, Salleh MZ. Tridimensional personalities and polymorphism of dopamine D2 receptor among heroin addicts. Biol Res Nurs. (2012) 14:188–96. doi: 10.1177/1099800411405030
40. Noble EP, Ozkaragoz TZ, Ritchie TL, Zhang X, Belin TR, Sparkes RS. D2 and D4 dopamine receptor polymorphisms and personality. Am J Med Genet. (1998) 81:257–67. doi: 10.1002/(SICI)1096-8628(19980508)81:3<257::AID-AJMG10>3.0.CO;2-E
41. Lee HJ, Lee HS, Kim YK, Kim SH, Kim L, Lee MS, et al. Allelic variants interaction of dopamine receptor D4 polymorphism correlate with personality traits in young Korean female population. Am J Med Genet B Neuropsychiatr Genet. (2003) 118B:76–80. doi: 10.1002/ajmg.b.10047
42. Tsai SJ, Hong CJ, Yu YW, Chen TJ. Association study of catechol-O-methyltransferase gene and dopamine D4 receptor gene polymorphisms and personality traits in healthy young Chinese females. Neuropsychobiology. (2004) 50:153–6. doi: 10.1159/000079107
43. Reist C, Ozdemir V, Wang E, Hashemzadeh M, Mee S, Moyzis R. Novelty seeking and the dopamine D4 receptor gene (DRD4) revisited in Asians: haplotype characterization and relevance of the 2-repeat allele. Am J Med Genet B Neuropsychiatr Genet. (2007) 144B:453–7. doi: 10.1002/ajmg.b.30473
44. Eley TC, Tahir E, Angleitner A, Harriss K, McClay J, Plomin R, et al. Association analysis of MAOA and COMT with neuroticism assessed by peers. Am J Med Genet B Neuropsychiatr Genet. (2003) 120B:90–6. doi: 10.1002/ajmg.b.20046
45. Hettema JM, An SS, Bukszar J, van den Oord EJ, Neale MC, Kendler KS, et al. Catechol-O-methyltransferase contributes to genetic susceptibility shared among anxiety spectrum phenotypes. Biol Psychiatry. (2008) 64:302–10. doi: 10.1016/j.biopsych.2008.03.014
46. Yu YW, Yang CW, Wu HC, Tsai SJ, Hong CJ, Chen MC, et al. Association study of a functional MAOA-uVNTR gene polymorphism and personality traits in Chinese young females. Neuropsychobiology. (2005) 52:118–21. doi: 10.1159/000087556
47. Emson PC, Goedert M, Horsfield P, Rioux F, St Pierre S. The regional distribution and chromatographic characterisation of neurotensin-like immunoreactivity in the rat central nervous system. J Neurochem. (1982) 38:992–9. doi: 10.1111/j.1471-4159.1982.tb05340.x
48. Delle Donne K T, Chan J, Boudin H, Pelaprat D, Rostene W, Pickel VM. Electron microscopic dual labeling of high-affinity neurotensin and dopamine D2 receptors in the rat nucleus accumbens shell. Synapse. (2004) 52:176–87. doi: 10.1002/syn.20018
49. Fuxe K, Von Euler G, Agnati LF, Merlo Pich E, O'Connor WT, Tanganelli S, et al. Intramembrane interactions between neurotensin receptors and dopamine D2 receptors as a major mechanism for the neuroleptic-like action of neurotensin. Ann N Y Acad Sci. (1992) 668:186–204. doi: 10.1111/j.1749-6632.1992.tb27350.x
50. Li XM, Ferraro L, Tanganelli S, O'Connor WT, Hasselrot U, Ungerstedt U, et al. Neurotensin peptides antagonistically regulate postsynaptic dopamine D2 receptors in rat nucleus accumbens: a receptor binding and microdialysis study. J Neural Transm Gen Sect. (1995) 102:125–37. doi: 10.1007/BF01276508
51. Liang Y, Boules M, Li Z, Williams K, Miura T, Oliveros A, et al. Hyperactivity of the dopaminergic system in NTS1 and NTS2 null mice. Neuropharmacology. (2010) 58:1199–205. doi: 10.1016/j.neuropharm.2010.02.015
52. Mercuri NB, Stratta F, Calabresi P, Bernardi G. Neurotensin induces an inward current in rat mesencephalic dopaminergic neurons. Neurosci Lett. (1993) 153:192–6. doi: 10.1016/0304-3940(93)90320-k
53. Farkas RH, Chien PY, Nakajima S, Nakajima Y. Properties of a slow nonselective cation conductance modulated by neurotensin and other neurotransmitters in midbrain dopaminergic neurons. J Neurophysiol. (1996) 76:1968–81. doi: 10.1152/jn.1996.76.3.1968
54. St-Gelais F, Legault M, Bourque MJ, Rompre PP, Trudeau LE. Role of calcium in neurotensin-evoked enhancement in firing in mesencephalic dopamine neurons. J Neurosci. (2004) 24:2566–74. doi: 10.1523/JNEUROSCI.5376-03.2004
55. Jin Q, Huang Y, Li X, Cong Z, Zhu G, Jiang W, et al. Association between FYN polymorphisms and defense mechanisms in healthy Chinese-Han subjects. Psychiatry Res. (2014) 220:723–4. doi: 10.1016/j.psychres.2014.07.068
56. Petraglia J, Thygesen KL, Lecours S, Drapeau M. Gender differences in self-reported defense mechanisms: a study using the new Defense Style Questionnaire-60. Am J Psychother. (2009) 63:87–99. doi: 10.1176/appi.psychotherapy.2009.63.1.87
57. Parekh MA, Majeed H, Khan TR, Khan AB, Khalid S, Khwaja NM, et al. Ego defense mechanisms in Pakistani medical students: a cross sectional analysis. BMC Psychiatry. (2010) 10:12. doi: 10.1186/1471-244X-10-12
58. Foto-Ozdemir D, Akdemir D, Cuhadaroglu-Cetin F. Gender differences in defense mechanisms, ways of coping with stress and sense of identity in adolescent suicide attempts. Turk J Pediatr. (2016) 58:271–281. doi: 10.24953/turkjped.2016.03.007
59. Konishi Y, Tanii H, Otowa T, Sasaki T, Motomura E, Fujita A, et al. Gender-specific association between the COMT Val158Met polymorphism and openness to experience in panic disorder patients. Neuropsychobiology. (2014) 69:165–74. doi: 10.1159/000360737
60. Chang CC, Chang HA, Fang WH, Chang TC, Huang SY. Gender-specific association between serotonin transporter polymorphisms (5-HTTLPR and rs25531) and neuroticism, anxiety and depression in well-defined healthy Han Chinese. J Affect Disord. (2017) 207:422–8. doi: 10.1016/j.jad.2016.08.055
Keywords: defense mechanism, defense style questionnaire, neurotensin receptor, gene polymorphism, Chinese
Citation: Ma H, Li M, Zhang L, Tao J and Zhu G (2021) Association of Neurotensin Receptor 1 Gene Polymorphisms With Defense Mechanisms in Healthy Chinese. Front. Psychiatry 12:762276. doi: 10.3389/fpsyt.2021.762276
Received: 27 August 2021; Accepted: 26 October 2021;
Published: 17 November 2021.
Edited by:
Michael Kluge, University of Leipzig, GermanyReviewed by:
Kuanjun He, Inner Mongolia University for Nationalities, ChinaAtif Adnan, China Medical University, China
Copyright © 2021 Ma, Li, Zhang, Tao and Zhu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gang Zhu, Z3podUBtYWlsLmNtdS5lZHUuY24=