 Nuria Tamayo-Estebaranz1,2
Nuria Tamayo-Estebaranz1,2 Carolina Muñoz-González3
Carolina Muñoz-González3 Ana María Gil-Valcárcel1,2
Ana María Gil-Valcárcel1,2 Paula Calvo López-Dávalos3
Paula Calvo López-Dávalos3 Andrea Martín-Vacas2*
Andrea Martín-Vacas2* Marta M. Paz-Cortés2,†Juan Manuel Aragoneses4,†
Marta M. Paz-Cortés2,†Juan Manuel Aragoneses4,†
- 1Programa de Doctorado en Ciencias de la Salud, Departamento de Medicina y Especialidades Médicas, Facultad de Medicina y Ciencias de la Salud, Universidad de Alcalá, Alcalá de Henares, Spain
- 2Facultad de Odontología, Universidad Alfonso X El Sabio, Villanueva de la Cañada, Spain
- 3Instituto de Investigación en Ciencias de la Alimentación, CSIC-UAM, Madrid, Spain
- 4Department of Dental Research, Federico Henriquez y Carvajal University, Santo Domingo Oeste, Dominican Republic
Objectives: The aim of this study was to investigate the relationship between active dentin caries (ADC), salivary biochemical parameters, and salivary microbiota composition in Spanish children and adults.
Methods: Saliva samples were collected from 80 subjects (40 adults and 40 children) divided between ADC and non-ADC. Salivary biochemical determination was performed by analysing total protein content (TPC) and total antioxidant activity (TAC) in saliva supernatants. DNA was obtained from the pellet of saliva samples using the Bacterial DNA kit and analysed with the Illumina NextSeq platform from all participants. Alpha diversity (Chao, Observed Features, Shannon and Simpson indices) and beta diversity (PCoA plot and PERMANOVA procedure) were analysed. In addition, Linear Discriminant Analysis Effect Size (LEfSe) was used to identify differential taxa between groups. All statistical analysis were performed with a 95% confidence level (p < 0.05).
Results: No significant associations were found between ADC and salivary biochemical markers in either the adult or pediatric age group, suggesting that these parameters alone may not sufficiently reflect cariogenic activity. Microbiota analysis at the phylum level did not show significant correlations with ADC; however, distinct associations appeared at the genus and species levels. In adults, several genera (Corynebacterium, Porphyromonas, Tannerella, Catonella, Filifactor, Parvimonas, and Dialister) were positively associated with ADC, reflecting a shift towards a dysbiotic microbiome composition that overlaps with periodontal and endodontic pathologies. Conversely, Haemophilus was negatively correlated with ADC, potentially indicating a protective role. At the species level, a positive correlation with ADC was found with Porphyromonas gingivalis, Porphyromonas endodontalis, Peptostreptococcus stomatis, Leptotrichia buccalis, Prevotella oris, or Corynebacterium matruchotii in the adult population. In children, microbial associations with caries were more limited, with Scardovia, a well-known acidogenic genus, positively correlated with ADC, and P. stomatis showing a negative association. Interestingly, P. stomatis exhibited opposite correlations in adults and children, possibly reflecting age-specific ecological roles. No significant differences in alpha or beta diversity were found either in adults or children participants.
Conclusions: Overall, these findings highlight a stronger and more diverse association between salivary microbiota and caries in adults compared to children. These results underscore the importance of age-specific microbial signatures in the aetiology of dental caries. The obtained differences suggest that caries development in adults may involve broader dysbiosis involving proteolytic and anaerobic organisms in addition to acidogenic species.
1 Introduction
The oral cavity is one of the most important habitats of the human body and one of the richest environments in microorganisms, consisting of bacteria, fungi, viruses, and archaea (1–3). It is formed by an ecological community of commensal, symbiotic, and pathogenic microorganisms (4) with a significant impact on both oral and general health and is a potential diagnostic indicator for various diseases such as cardiovascular disease, rheumatoid arthritis, cancer, or diabetes, among others (5–7). The oral environment constantly transforms with age, and, consequently, the microbiota also changes (8). At birth, most children do not have a colonized microbiome, and during the first two months of life, bacteria colonize the mucosal surfaces, and later, with the eruption of primary teeth, these surfaces are also colonized (8). During childhood, primary dentition exfoliates, leading to mixed dentition and finally to permanent dentition (2), with observed changes in bacterial colonization at different dentition stages (6, 9), resulting in known differences between the microbiota of children and adults (10, 11). In this regard, Ling et al. (12) observed that Actinobacteria (syn. Actinomycetota), Bacteroidetes (syn. Bacteroidota), and Fusobacteria (syn. Fusobacteriota) were overrepresented in children, whereas Firmicutes (syn. Bacillota) and Proteobacteria (syn. Pseudomonadota) were more abundant in adults.
Although oral microbiota is resistant to minor ecological changes, factors such as diet, environment, inadequate oral hygiene (13), the use of medications or prolonged use of antibiotics (8) can also induce structural changes in the microbiota (14) leading to dysbiosis of the resident microbiota (4), which can result in caries (1).
Untreated dental caries is the most prevalent condition worldwide and is estimated to affect 2.5 billion people of which 530 million are children (15). Although Streptoccocus is the main genera related to dental caries, other microorganisms have been identified in relation to the presence of caries lesions (i.e., Fusobacterium, Prevotella, Leptotrichia, Veillonella, Bifidobacterium, and Capnocytophaga species) (4). However, several important limitations in the existing studies on the oral microbiota and its association with dental caries have to be considered. Many of these investigations rely on non-omics-based techniques, which restrict the detection and characterization of non-culturable bacteria, thus providing an incomplete picture of the microbial diversity involved. Furthermore, most studies have been conducted in Asian populations, with limited data available from Spanish or broader European cohorts, making it difficult to generalize findings across different populations. In addition, there is a lack of studies that simultaneously analyse both children and adults, despite known age-related differences in oral microbiota composition. In this sense, confounding factors such as sex and age are often not adequately controlled or considered in the analysis, potentially biasing the results and limiting the reliability of observed associations.
Another aspect to consider is the site of sample collection. In this regard, saliva is a biological fluid that bathes the oral cavity, and it is essential for maintaining both oral and general health (14). It is estimated that over 700 bacterial species coexist in the oral cavity (16), among them, only two-thirds belong to in vitro cultivable species (17). The rich microbiota provides a composite representation of microorganisms from diverse niches within the oral cavity (18) and reflects local alterations of the supragingival and subgingival microbiota (4). Salivary collection is non-invasive, simple, and painless collection method, which makes it particularly suitable for studies involving children (16). Moreover, it has been reported that saliva composition parameters, such as total protein content and antioxidant capacity, are associated with the composition of the salivary microbiota (19) which may in turn influence susceptibility to oral conditions such as caries. These biochemical properties could potentially modulate the oral environment and shape microbial communities, highlighting the importance of integrating salivary biomarkers into microbiome research. These gaps underscore the need for more comprehensive, omics-based, and demographically diverse research that considers developmental stages, relevant confounding variables, and optimized sampling strategies to better understand the complex interactions between the oral microbiome and dental caries.
This study aimed to investigate the relationship between salivary microbiota composition, based on 16S rRNA gene V3–V4 amplicon sequencing, and the presence of active dentin caries (ADC), in Spanish children and adults. Additionally, the relation between salivary parameters, including pH, flow rate, total protein content, and antioxidant activity, and caries status was evaluated in both age groups.
2 Methods
A case-control, cross-sectional, observational, and analytical study was conducted, with two parallel and matched arms. The study design was approved by the Bioethics Committee of the San Carlos Clinical Hospital (code 22/334-E, 31st May 2022) complying with current Spanish and European regulations on personal data protection and the principles of the Declaration of Helsinki for research involving human subjects. Additionally, the STROBE guidelines for cross-sectional studies were followed as reporting guidelines (20) (Supplementary Table S1).
2.1 Study population
The subjects in this study were adults (n = 40) and children (n = 40) attending dental check-ups at the dental clinic of Alfonso X El Sabio University (UAX) in Madrid, Spain. Subjects aged 6–12 years were included for the paediatric population and 20–40 years for the adult population, who voluntarily agreed to participate in the study. Additionally, the main groups were subdivided into non ADC (non-ADC) and ADC categories based on their active caries index. According to the International Caries Detection and Assessment System (ICDAS), caries presence was classified in initial or enamel caries (healthy tooth according to OMS code) as ICDAS codes 1–3, or ADC as ICDAS Codes 4–6 (21). The non-ADC consisted of volunteers with non caries (ICDAS codes 0–3), while the ADC group included volunteers with active caries in dentin (ICDAS codes 4–6). This resulted in four groups of volunteers: children with non-ADC (n = 20), children with ADC (n = 20), healthy adults with non-ADC (n = 20), and adults with ADC (n = 20). All groups are balanced in terms of sex. In addition, the Community Periodontal Index (CPI) were calculated to assess oral health status of the volunteers (22, 23). Subjects with behavioural problems, smokers, systemic diseases, pregnant or during lactation, chronic medication or those who had taken antibiotics in the last three months were excluded.
Previous similar research studied 30–50 children (13, 24–27) or 46 adults (28), respectively. Due to differences in study procedure and outcomes and the preliminar feature of the research, we decided to calculate the needed sample size theoretically, based on caries prevalence. The sample size was calculated assuming an infinite reference population (average patient volume data from the dental clinic not available) and a caries prevalence of 95% (100% in the adult population, 90% in the paediatric population) (29). A random sample of 74 individuals was found sufficient to estimate, with 95% confidence and a precision of ±5 percent points. The anticipated replacement rate is projected to be 1%, to resolve possible sample losses during the collection or sampling procedure. To ensure sample homogeneity, the decision was made to increase the sample size to 80 subjects, evenly divided between adult and paediatric populations, (n = 40 in each population group) adjusted by sex. In each population group, the same subject amount of each oral health status was selected, so finally 20 children with non-ADC, 20 children with ADC, 20 adults with non-ADC and 20 adults with ADC were included.
The sampling method was non-probabilistic consecutive cases between November 2022 and January 2023, until reaching the predetermined number. Study subjects over 16 years old were initially informed verbally and given a Patient Information Sheet and an Informed Consent Form, which had to be completed before study participation. In cases where the subjects were under 12 years old, these steps were performed and authorized by their parents or legal guardians. For children aged 12 years, the child's acceptance, as well as that of the parents, was essential for participation in the study.
2.2 Health data and salivary collection
Oral and general health data were obtained directly from the study participants through oral survey by the principal investigator (N.T.-E.), including age and sex. Oral health data (caries presence and CPI) were recorded following WHO and ICDAS guidelines (21, 22). Data regarding systemic health, medication intake, smoking habits, alcohol intake and current health status were also collected to appropriately select the sample.
Saliva samples were collected at the dental clinic facilities of UAX by the study investigators between 8 and 11 am, confirming through interviews with the subjects that they maintained fasting and did not brush their teeth for at least 1 h before the collection. Individuals were instructed in the collection of samples by unstimulated saliva expectoration into collection tubes (Labbox, Barcelona, Spain) for 10 min. Immediately after saliva collection, the salivary pH was measured, and salivary flow was calculated by the difference in weight between the empty tube and the tube after spitting out the saliva and expressed as ml/min, assuming 1 g being equal to 1 ml. Immediately, salivary samples were frozen at −20°C. Subsequently, saliva samples were centrifuged (15,000 g for 15 min at 4°C), and the supernatant and pellet were aliquoted separately and stored at −80°C until analysis, to prevent potential changes in their composition and/or properties.
2.3 Biochemical analysis of salivary samples
Biochemical determination was performed by analysing total protein content (TPC) and total antioxidant activity (TAC) in saliva supernatants. TPC was measured using the commercial PierceTM BCA Protein Assay Kit (Pierce ThermoScientific, Rockford, IL, USA). Antioxidant activity (TAC) was measured using the Ferric Reducing Ability of Plasma (FRAP) assay method described by Palomar-Bonet et al. (30).
2.4 DNA extraction, sequencing, and data processing
Total DNA was obtained from the pellet of saliva samples using the Bacterial DNA kit (D3350-02, E.Z.N.A™, OMEGA bio-tek, USA), following the manufacturer's instructions. Pellets were resuspended in 100 μl TE buffer containing lysozyme (10 mg/ml), and microbial cells were lysed by mechanical disruption with glass beads (0.1 mm zirconia/silica diameter), using a FastPrep disruptor (QBioGene, Irvine, CA, USA) at a speed of 6 m/s for 30 s. DNA retained on the HiBind DNA column of the kit was eluted twice with 50 μl elution buffer. The concentration and purity of genomic DNA were measured using a NanoDrop™ ND-1000 UV spectrophotometer (Nano-Drop Technologies, Wilmington, DE, USA). DNA samples were analysed via amplicon-based metagenomic sequencing of the 16S rDNA V3–V4 region, performed by Novogen (Cambridge, UK) on an Illumina platform, to generate 250 bp paired-end reads. Bacterial taxonomy was assigned to the obtained ASVs (Amplicon Sequence Variants) by using the QIIME2 software (https://qiime2.org/). ASVs with abundance lower than 0.1% were removed. Subsequent analyses of alpha diversity (calculation of Chao, Observed Features, Shannon and Simpson indices) were performed based on the normalized data.
A beta-diversity analysis was performed, to assess differences in the multivariate microbiota composition between study groups. In order to evaluate the complexity of the community composition and compare the differences between samples, beta diversity was calculated based on weighted and unweighted unifrac distances in QIIME2 software. Cluster analysis was performed with principal component analysis (PCA), which was applied to reduce the dimension of the original variables using the ade4 package and ggplot2 package in R software (Version 3.5.3). Principal Coordinate Analysis (PCoA) was performed in the entire group to obtain principal coordinates and visualize differences of samples in complex multi-dimensional data. A matrix of weighted or unweighted unifrac distances among samples obtained previously was transformed into a new set of orthogonal axes, where the maximum variation factor was demonstrated by the first principal coordinate, and the second maximum variation factor was demonstrated by the second principal coordinate, and so on.
2.5 Statistical and bioinformatic analyses
Statistical analysis was performed XLSTAT program (version 19.01, Addinsoft, Paris, France), R Software (Version 2.15.3) and IBM® SPSS® Statistics software (version 29.0.2.0, IBM), with a confidence level of 95% (significance set at p < 0.05). Adjustment to a normal distribution was analysed with Kolmogorov–Smirnov and Shapiro–Wilk tests. Separate analysis was performed for adults and children. Intragroup differences in biochemical salivary variables were evaluated with Student's T test, as biochemical outcomes followed a normal distribution. Linear Discriminant Analysis Effect Size (LEfSe LDA Effect Size) (version 1.0), with LDA score threshold of 4, was performed for identification of genomic features. Due to non-parametric distribution of evaluated variables, differences between salivary microbiota between study groups (non-ADC and ADC) were analysed using Mann–Whitney test and the behaviour of outcomes was evaluated with Spearman correlation analysis. Regarding the beta-diversity analysis, the three-dimensional PCoA results were displayed using QIIME2 package, while the two-dimensional PCoA results were displayed using ade4 package and ggplot2 package in R software (Version 2.15.3). To study the significance of the differences in community structure (beta-diversity) between groups, the adonis (PERMANOVA) and anosim functions in the QIIME2 software were used to do analysis. To find out the significantly different species at each taxonomic level (Phylum, Class, Order, Family, Genus, Species), the R software (Version 3.5.3) was used to do MetaStat and T-test analysis.
3 Results
3.1 Sample description
Saliva samples from 40 adults and 40 children were analysed, with each age group evenly divided into ADC (n = 20) and non-ADC (n = 20) individuals. The groups were balanced by sex, and the age of both study and control samples was homogeneous within the adult (p = 0.602) and child (p = 0.192) groups (Table 1). No differences in BMI index were found (p = 0.102 and p = 0.779, respectively for adult and child population). As expected, the ADC and non-ADC groups significantly differed in the number of active caries in both children and adults (p < 0.001 for both groups). In this regard, the ADC group presented significantly higher values of dentin caries and total caries than the non-ADC group, both in adult and children population Mean CPI was found higher in ADC than non-ADC adult group (p = 0.007) but no differences were found among children (p = 0.429).
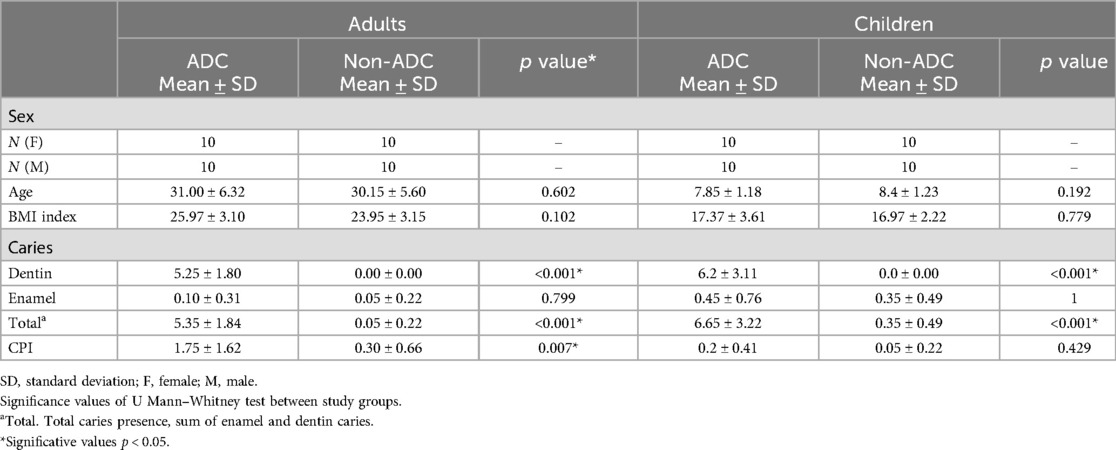
Table 1. Demographic characteristics, caries and periodontal status of the study participants.
A general visualization of the data obtained was performed, and it was observed that the samples corresponding to four individuals (three adults and one child) showed unmistakable evidence of contamination during collection, extraction, or analysis. Therefore, these samples were not considered during the statistical analysis of the metagenomic data (Supplementary Figure S1).
Regarding salivary parameters (Table 2), salivary flow was 0.6 ± 0.3 ml/min in adults and 0.6 ± 0.4 ml/min in children. These values fall within the normal unstimulated salivary flow range (typically 0.3–0.7 ml/min), indicating no apparent salivary hypofunction in either group. The salivary pH was 7.0 ± 0.3 in adults and 7.1 ± 0.3 in children, also within the expected physiological range (pH 6.2–7.6), suggesting normal acid–base balance in oral fluid. For TPC, values were 1,300.2 ± 456.7 µg/µl in adults and 1,156.3 ± 415.2 µg/µl in children. Although variability exists across individuals and collection methods, both values are within the reported reference ranges for TPC (typically between 500 and 2,000 µg/µl), indicating normal protein composition. Similarly, TAC was higher in adults (367.3 ± 138.6 µM FeSO₄) than in children (281.7 ± 136.8 µM FeSO₄). These values are consistent with ranges reported in healthy populations, although TAC may vary depending on diet, age, and oxidative stress levels. All these parameters showed no statistically significant differences in pH, salivary flow rate, TAC and TPC among the ADC level both in adult and children population (Supplementary Table S2) (p > 0.05 for all comparisons).
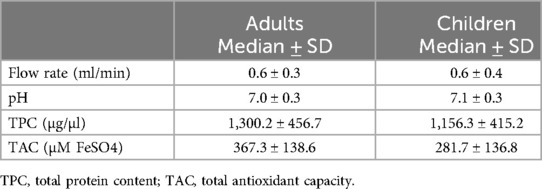
Table 2. Values of the different salivary parameters measured in this study (salivary flow rate, pH, TPC, TAC) in adults and children participants.
The results of the taxonomic profiling based on saliva samples are summarized in Figure 1. Microbial composition was examined at the phylum, genus, and species levels in both adult and child groups. To ensure clarity and comparability, only taxa with a relative abundance greater than 0.1% in the initial dataset were retained for visualization. Within this filtered subset, relative abundances were recalculated to sum to 100%. At the phylum level (Figure 1A), Firmicutes (syn. Bacillota) was the most dominant phylum in the adult group, followed by Proteobacteria (syn. Pseudomonadota) and Bacteroidetes (syn. Bacteroidota). A similar taxonomic profile was observed in children (Figure 1B), with these three phyla collectively comprising most of the oral microbial community in both groups. This composition is in accordance with previously reported profiles of the human oral microbiome (31). At the genus level, Streptococcus emerged as the most abundant genus in both groups (Figures 1C,D), reflecting its well-established dominance in the oral cavity, particularly in early biofilm formation and carbohydrate metabolism (32, 33). Other abundant genera included Prevotella, Rothia, Haemophilus, and Neisseria, all of which are considered part of the core oral microbiota and are frequently associated with mucosal surfaces and saliva (34). Intra-group analysis of alpha-diversity was conducted separetely on samples from adults and children according to ADC. No statistical significant differences (p > 0.05) were found for alpha diversity indices either for adults or children (Figure 2; Supplementary Table S3), suggesting that ADC did not influence the level of bacterial richness present in saliva samples in either children or adults. Regarding the specific taxa that differ between groups (LEfSe), certain taxa were observed to contribute to the separation of ADC groups within the adult population (Figure 3). The most discriminant taxa in the ADC group belonged to the class Clostridia, specifically the order Lachnospirales and family Lachnospiraceae, and to the phylum Fusobacteria (syn. Fusobacteriota), specifically the class Fusobacteriia and order Fusobacteriales. Additionally, the family Porphyromonadaceae and genus Porphyromonas were more abundant in this group. In contrast, the non-ADC group was associated with higher levels of the order Pasteurellales, family Pasteurellaceae, and genus Haemophilus. Regarding children, LEfSe LDA effect size did not reveal any significant results between ADC and non-ADC groups.
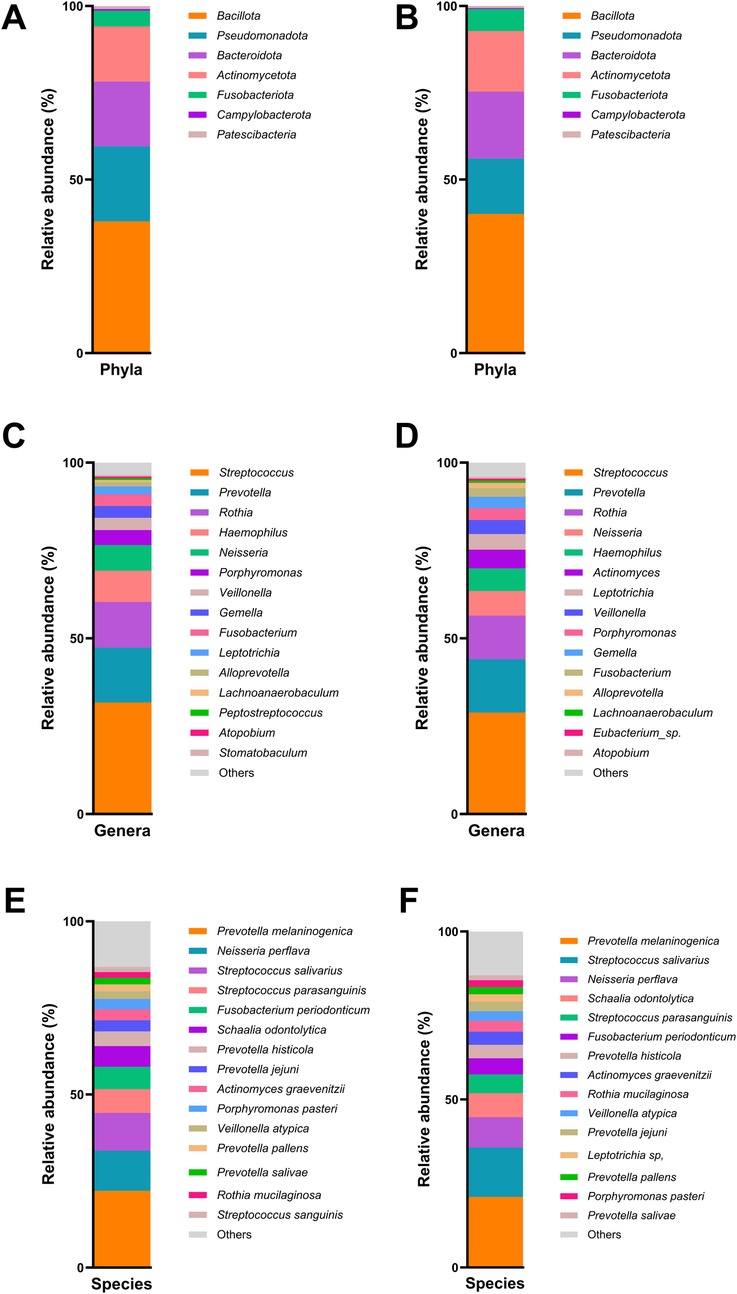
Figure 1. Relative abundance (%) of bacteria phyla in adult group (A), bacteria phyla in children group (B), bacteria genera in adult group (C), bacteria genera in children group (D), bacteria species in adult group (E), and bacteria species in children group (F).
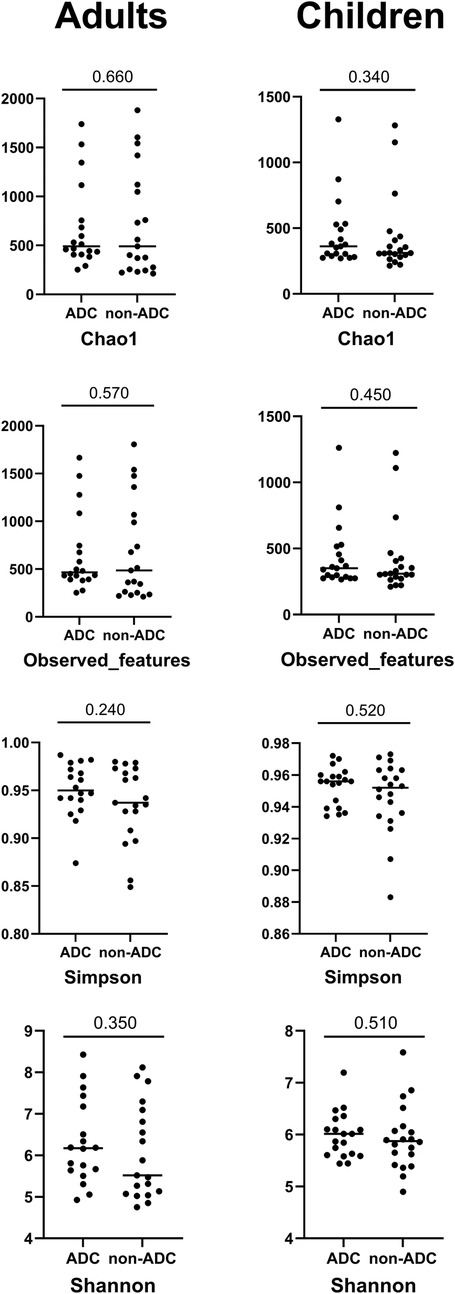
Figure 2. Alpha diversity indices between study groups (ADC and non-ADC) divided according to age categories, and significance values for Mann–Whitney test. No statistically significant differences were found between ADC and non-ADC groups either in adults or children participants in the alpha diversity indices studied.
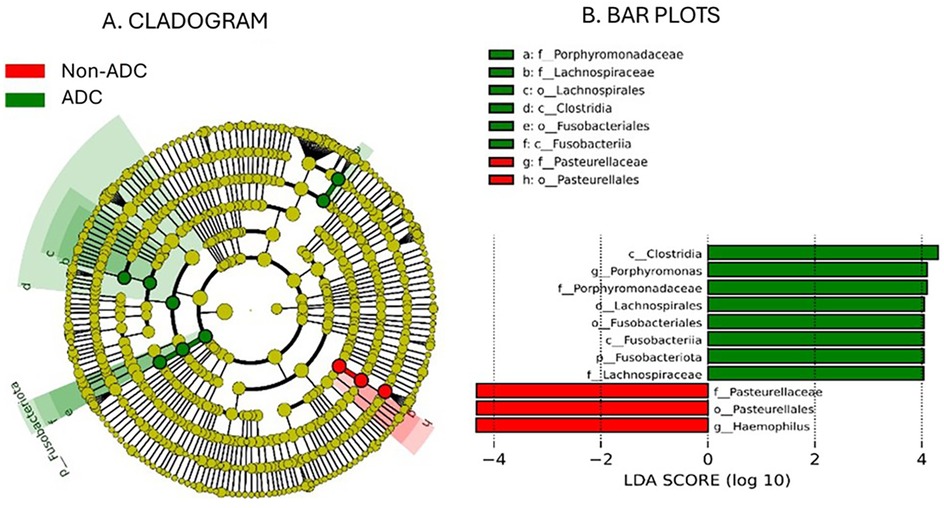
Figure 3. Cladogram analysis (A) of 16S rDNA sequences from salivary microbiota in adult group and histogram with bar plots (B) showing the significant differences in taxonomy between groups (p value < 0.05; LDA score 4.0).
In order to evaluate the complexity of the community composition and compare the differences between study groups, beta diversity was calculated. No statistically significant differences were found between ADC groups either in the adult (p = 0.168) or children (p = 0.962) groups studying beta diversity (Supplementary Table S4). A PCoA analysis was conducted to represent the obtained results in the entire group. To visualize the differences in beta diversity between groups, a PCoA based on Bray–Curtis distances (Figure 4A) and another based on Jaccard distances (Figure 4B) were performed. The obtained data indicated that the most pronounced differences were related to age groups, with less distinct clustering based on dental status.
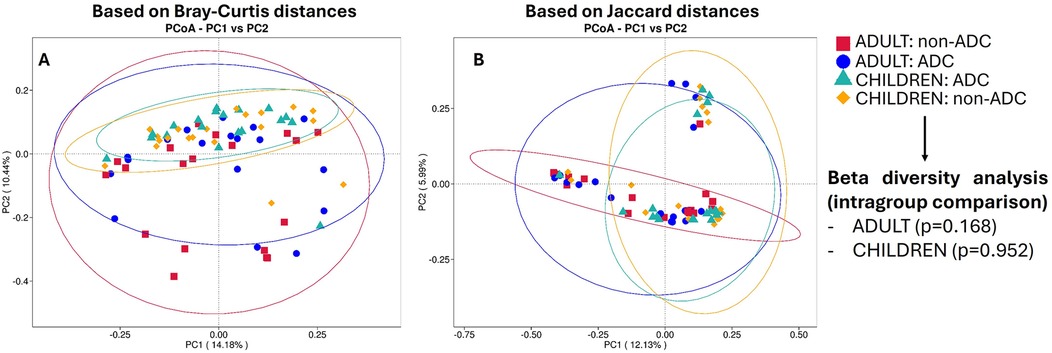
Figure 4. Pcoa plots based on bray-curtis distances (A) and on jaccard distances (B) No beta diversity differences between age groups were found (PERMANOVA p values >0.05). Notably, the most pronounced differences are related to age groups (adults or children), with less distinct clustering based on dental status (ADC or non-ADC).
3.2 Relationship between salivary parameters and active dentin caries in children and adults
Due to the sample size constraints, correlation analyses were performed to explore potential associations between salivary biochemical parameters and ADC in both adults and children.
As illustrated in Figure 5, no significant correlations were observed between ADC and any of the salivary parameters in either age group (p > 0.05 for all comparisons). However, this figure highlights notable differences in the relationship between salivary parameters and ADC in children and adults. In adults, these relationships appear weak, with only a slightly stronger negative correlation observed between salivary pH and the number of active caries, indicating that higher pH levels are associated with fewer carious lesions. In contrast, in children, the associations are somewhat stronger (but not significant) and show a different pattern: positive correlations were observed for pH, total protein content (TPC), and total antioxidant capacity (TAC), whereas flow rate exhibited a negative correlation with ADC.
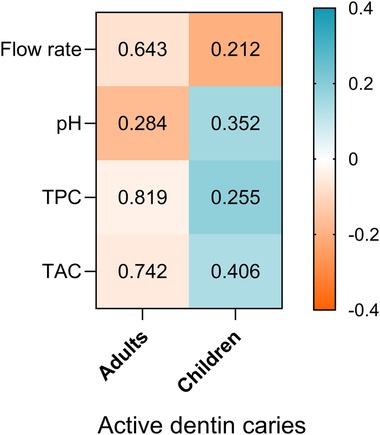
Figure 5. Heatmap of spearman correlation between salivary parameters (flow rate, pH, TPC and TAC) and ADC in adults and children participants. The direction and strength of the correlations are represented by the colour scale, where blue indicates positive correlations and orange indicates negative correlations P- values of correlation are shown inside each cell.
3.3 Relationship between salivary microbiota and active dentin caries, demographic variables, and periodontal index in children and adults
Spearman correlation analyses were conducted to investigate potential associations between salivary microbiota composition and ADC in both adults and children (Figure 6; Supplementary Table S5). Neither in adults nor children significant correlations between phylum relative abundance and ADC (p > 0.05 for all comparisons). CPI correlation with salivary microbiota composition was also analysed, finding no significant correlation between phylum relative abundance and CPI in children; on the other hand, a significant and negative relationship was found between CPI and Firmicutes (Syn. Bacillota) and Actinobacteriota (Syn. Actinomycetota)(Supplementary Table S6).
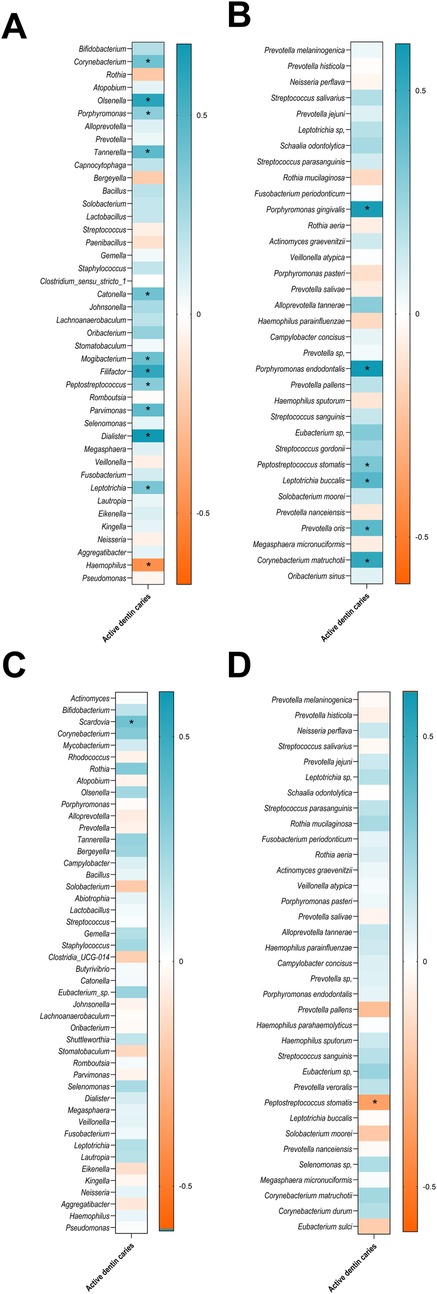
Figure 6. Heatmap of spearman correlations between ADC and the relative abundance of the oral microbiota, showing bacterial genera (A), and species in adults (B),and bacterial genera (C), and species in children (D) only taxa with a relative abundance greater than 0.1% are shown. Asterisks indicate statistically significant correlations (p < 0.05). The strength and direction of the correlations are represented by the colour scale, with blue indicating positive correlations and orange indicating negative correlations, as shown in the accompanying legend.
According to the obtained results, notable differences were observed in the association between salivary microbiota composition and the number of ADC in adults compared to children. In adults, a larger number of genera were positively correlated with ADC, including Corynebacterium, Olsenella, Porphyromonas, Tannerella, Catonella, Mogibacterium, Filifactor, Peptostreptococcus, Parvimonas, Dialister, and Leptotrichia (Figure 6A). Among species, P. gingivalis, P. endodontalis, P. stomatis, L. buccalis, Prevotella oris, and C. matruchotii were positively associated with ADC (Figure 6B). In contrast, Haemophilus was the only genus negatively correlated with ADC in adults (Figure 6A). In children, by contrast, the associations between microbiota and ADC were more limited. Only the genus Scardovia showed a significant positive correlation with the number of ADC (Figure 6C). At the species level, P. stomatis was significantly and negatively associated with ADC (Figure 6D).
Related to periodontal status, in adults many genera were positively correlated with CPI, including Bifidobacterium, Olsenella, Porphyromonas, Tannerella, Capnocytophaga, Catonella, Johnsonella, Filifactor, Parvimonas, Dialister and Leptotrichia. On the other hand, Rothia and Haemophilus were negatively correlated with CPI. Among species, P. gingivalis, P. endodontalis, P. stomatis, L. buccalis and C. matruchotii were positively and significantly correlated to CPI. In contrast, in children, the correlations were more limited, with only negative significant correlation between Porphyromonas genus, and positive correlation with Streptococcus salivarius.
4 Discussion
The main objective of this study was to evaluate the relationship between the level of ADC with salivary biochemical parameters and salivary microbiota composition in both, children and adults from Spain. In line with most of the previous work, the present study employed a cross-sectional design, which, although limited in its ability to infer causality, remains a practical and commonly used approach in oral microbiome research. Our sample size (n = 80), comprising 40 children and 40 adults evenly distributed by ADC status, is comparable to similar investigations, which have included between 30 and 50 paediatric participants or 40–50 adult subjects. In the present study, we adhered to WHO criteria to define the presence of caries and further classified them into enamel and dentin lesions according to ICDAS, allowing a more precise stratification of disease severity. To minimize confounding variables related to age, participants were selected within a narrow age range, with mean ages of approximately 8 years for children and 30 years for adults. As in many metagenomic studies, 16S rRNA gene sequencing was used to characterize the microbial communities present in saliva. All participants exhibited salivary values within normal ranges for flow rate, pH, TPC, TAC, and microbial composition.
Regarding salivary metagenomic studies, most of them have been conducted in Asian countries such as China (1, 13, 24–26, 28, 35–43) or Korea (44, 45), although they have also been carried out in European countries (9, 10, 12, 14, 16, 17, 27, 46–49), Turkey (50, 51), USA (2, 5, 23, 52, 53) and Australia (54), while only one study conducted in Spain to date (55). It is important to consider the dietary, hygienic and social differences between different races and cultures, which underlines the need to carry out population studies that describe salivary bacterial composition, helping to detect intercultural differences (56). In relation to the design of the published studies, most are cross-sectional, similar to the present study (2, 9, 14, 16, 17, 23–25, 28, 35–43, 46–50, 52, 57), although longitudinal studies have also been conducted (5, 10, 12, 13, 26, 27, 45, 51). Regarding the population characteristics, variables studied, and sample size, we find that the sample size is very heterogeneus, with some studies including only two participants (52), up to studies with 293 subjects (47). In our case, we studied a total of 80 subjects (40 children and 40 adults, evenly divided according to ADC), similar to previous studies, with 30–50 children (13, 24–27) or 46 adults (28), respectively. Regarding the age of the studied population, there is great heterogeneity, with studies analyzing ages ranging from 17 to 63 years in adults (10, 28, 35, 38, 39, 41–44, 46–49, 51) and from 3 months-18 years in children (1, 5, 9, 12, 13, 16, 24–27, 40, 42, 45, 50, 54, 57). Similar to our study, with a mean age of 30.15 ± 5.6 and 31 ± 6.3 years old for adults (non-ADC and ADC respectively) and 7.85 ± 1.18 and 8.4 ± 1.23 years old for children (non-ADC and ADC respectively), with a narrow range to avoid biases associated with age changes. Regarding the variables of interest evaluated in relation to salivary metagenomics, studies in adults have focused on caries (10, 28, 38, 39, 42, 48, 51), periodontal disease (14, 35, 44, 46, 49) or both (41, 43, 47, 52). In contrast, in children, the studies conducted evaluated caries disease (1, 5, 9, 12, 13, 16, 24–27, 40, 42, 45, 54, 57). To determine the status of oral health or disease, different diagnostic criteria and reference indices were used in published studies. In previous research the WHO criteria were employed as the diagnostic method for caries (12, 23, 24, 45), while in other studies used ICDAS method (1, 16, 27, 51).
In relation to the association between salivary biochemical parameters (salivary flow rate, pH, TPC, and TAC) and ADC in both children and adults, our analysis did not reveal any significant correlations in either age group. These findings were somewhat unexpected, given prior evidence suggesting otherwise. For instance, Pandey et al. (58) reported higher salivary pH levels in caries-free children and observed an increase in antioxidant capacity in children with active caries, irrespective of age. Consistent with our results, however, they found no significant differences in the salivary flow rate between caries-active and caries-free individuals. In contrast, Pyati et al. (59) observed significant differences in salivary flow rate, pH, and total protein levels between children with active caries and healthy controls. Similarly, a recent systematic review focusing on paediatric populations (60) supported these findings, indicating that salivary parameters such as flow rate, pH, and buffering capacity tend to decrease in children with caries, regardless of age or sex. However, the authors reported no significant changes in total protein concentration among children aged 6–12 years, which aligns with our results, especially considering the narrow age range employed in our study to minimize age-related variability.
One possible explanation for the lack of significant associations in our data could be the multifactorial nature of dental caries, which involves complex interactions between host factors, diet, oral hygiene practices, and microbiota composition. Biochemical parameters such as pH and antioxidant capacity may fluctuate over short periods and be influenced by recent food intake or stress, making them less reliable as isolated markers of caries activity. Additionally, the narrow range of biochemical values observed in our participants, most of whom exhibited values within normal physiological limits, might have limited our ability to detect statistically significant differences. It is also possible that microbial composition and ecological shifts in the biofilm play a more critical role in caries development than the salivary biochemistry alone.
Thus, we explored the relationship between salivary microbiota composition with ADC in both age groups. In the present study, we did not observe significant correlations between ADC and the relative abundance of bacterial phyla in either children or adults. These findings suggest that shifts in the salivary microbiota related to caries may occur more prominently at lower taxonomic levels, such as genus or species, rather than at the broader phylum level. This observation aligns with previous studies indicating that caries-associated microbial changes are often subtle and localized to specific taxa rather than being characterized by global shifts in major bacterial phyla (32, 61). In relation to periodontal status, although in children no statistical correlation was found, a significant and negative relationship was found between CPI and Bacillota and Actinobacteriota, according to previous research, stating that Fusobacteria and Saccharibacteria (TM7) are the most abundant phyla associated with gingivitis, while Actinobacteriota and Bacteroidetes are less frequently associated with gingivitis (35).
At the genus level, several taxa showed significant positive correlations with ADC in adults, including Corynebacterium, Porphyromonas, Tanerella, Catonella, Oribacterium, Mogibacterium, Filifactor, Parvimonas, Dialister, or Leptotrichia. Many of these genera have been associated with periodontal and endodontic infections, and their presence may reflect a microbial dysbiosis that supports both caries progression and other oral pathologies (62, 63). For example, Porphyromonas have been implicated in pulp and periapical infections and have been increasingly associated with the progression of deep carious lesions (62–67). Similarly, Filifactor and Parvimonas are anaerobic genera linked to endodontic and periodontal diseases, supporting the notion that late-stage dentin caries may share microbial signatures with these conditions. These results suggest that the adult oral microbiota harbours a more diverse array of genera associated with caries, many of which have been previously implicated in dysbiotic biofilms and periodontal or endodontic infections. The presence of multiple anaerobic and proteolytic taxa (e.g., Filifactor, Mogibacterium) in this group may reflect the complex microbial consortia contributing to caries progression beyond classical acidogenic bacteria. However, it should be noted that in our study, adults with higher ADC level also presented higher level of CPI. Thus, these findings should be confirmed in later studies. Conversely, Haemophilus showed a significant negative correlation with ADC in adults, and thus, its abundance could potentially protect from caries development. This genus is commonly associated with health-associated oral microbiota and may exert protective effects via competitive exclusion or production of neutral or alkali metabolic byproducts.
At the species level, in adult saliva, the presence of ADC was positively correlated with P. gingivalis, P. endodontalis, P. stomatis, L. buccalis, P. oris, and C. matruchotii. Interestingly, P. gingivalis, traditionally linked to periodontitis, and P. stomatis, typically associated with endodontic infections, showed positive correlations with caries in adults, suggesting potential ecological shifts in the salivary microbiome under cariogenic conditions. C. matruchotii, though traditionally considered a commensal, may contribute structurally to the development of cariogenic biofilms through its characteristic filamentous morphology and central positioning in the “hedgehog” structure of supragingival plaque (68). A positive correlation between P. gingivalis and CPI was found, according tio Kim et al. (44), who reported an increase in the group of patients with periodontitis, along with abundance of T. denticola and T. forsythia. In children, by contrast, the associations between microbiota and caries were more limited. Only the genera Scardovia, particularly known for its role in early childhood caries and acidogenic potential, was significantly and positively correlated with ADC. Scardovia wiggsiae, in particular, has been well documented as a caries-associated species in paediatric populations and has been proposed as an early colonizer in the development of severe early childhood caries (69). At the species level, P. stomatis was significantly and negatively associated with caries, suggesting a potentially protective role in the paediatric population. Interestingly, P. stomatis showed opposing correlations with caries in adults and children, positive in the former and negative in the latter. This discrepancy may reflect age-related differences in the oral microbiome's composition, maturity, and ecological dynamics. In adults, P. stomatis may participate in dysbiotic consortia involving anaerobic and proteolytic taxa that contribute to caries progression. In contrast, in children whose microbiome is less mature and more dominated by acidogenic species, P. stomatis may play a less pathogenic or even modulatory role. These divergent associations may also be influenced by differences in salivary parameters, immune responses, or microbial interactions across age groups. Further studies are needed to clarify the functional role of P. stomatis in distinct developmental contexts.
This study presents several limitations that should be considered when interpreting the results. First, the sequencing of the V3–V4 regions of the 16S rRNA gene, although widely used in microbiome studies, does not always provide sufficient resolution to reliably distinguish between closely related bacterial species. As a result, a substantial proportion of sequences could not be taxonomically classified at the species level, potentially limiting the precision of our microbial profiling. Second, the classification of participants was based solely on the presence or absence of ADC. While none of the individuals in the non-ADC group presented clinical signs of periodontitis, the ADC group included participants with varying periodontal health status, ranging from high periodontal indices to no clinical signs. This heterogeneity may function as a confounding factor, particularly considering the overlapping microbial profiles between caries and periodontal diseases. Future studies should aim to better control for periodontal status to isolate the specific microbial signatures associated with ADC. Third, the cross-sectional nature of this study prevents the establishment of causal relationships between microbiota composition, salivary biochemical parameters, and caries status. Longitudinal studies are needed to elucidate temporal dynamics and causality. Additionally, the sample size, although comparable to previous studies in this field, was relatively limited, which may reduce statistical power. Besides, due to the theoretical calculation of sample size the generalization could be reduced. Therefore, correlation analyses were employed as an exploratory approach to detect potential associations, recognizing the inherent limitations in generalizability and robustness. The statistical analysis of beta diversity lacks model adjusting of covariables as sex or age, due to the low sample size. Although that adjusted analysis would improve the quality of the interpretation and generability in a wide sample size, in our sample the sample size divided by sex and age would rise the biases (i.e., low data precision, atypical data, high confidence intervals, data distortion) of the data analysis. Besides, sampling was paired by sex in order to prevent sex influence in results. Furthermore, some aspects of the CPI index constitute limitations, as it is a population-based epidemiological index, but it is an index that measures the need for treatment associated with pathology. According to the WHO and the American Society of Pediatric Dentistry, its use is recommended for assessments in both adults and child populations. Besides, it has been used in previous research (30, 49, 70, 71).
Despite these limitations, this study has several notable strengths. Most existing salivary metagenomic studies have been conducted in Asian populations, where cultural practices, dietary habits, and oral hygiene behaviours differ significantly from those in other regions. Our study contributes novel data from a Western European population, specifically from Spain, where only limited microbiome data is currently available. This adds valuable intercultural insight into the salivary microbiota and its association with oral health. Moreover, the study was carefully designed to minimize confounding variables. Thus, adults and children, with and without ADC were balanced for sex and body mass index (BMI), and in both age groups, participants were matched by age to reduce potential age-related microbiome variability. Saliva collection followed standardized and validated protocols, ensuring consistency in sample handling and processing. Additionally, rigorous inclusion and exclusion criteria were applied, helping to control for variables such as recent antibiotic use, systemic diseases, and other factors known to influence the oral microbiome. Finally, the integration of microbiota data with salivary biochemical markers, such as TAC and total TPC, adds depth to the findings and provides a more holistic view of the oral ecosystem. The balanced study design, along with the novelty of exploring both microbiota and oxidative markers in relation to caries in adults and children, enhances the relevance and scientific value of this work.
Finally, interpreting the data obtained, significant differences were found in salivary bacterial composition in the presence of ADC, as well as periodontal disease, the most common diseases in dentistry. Furthermore, data suggest that salivary parameters do not play as important a role as salivary microbiological composition. However, the aforementioned limitations do not allow generalization to broad populations, making it necessary to conduct broad-spectrum, multicentre, and controlled studies that allow for firm conclusions and allow for extrapolation of data.
5 Conclusions
This study explored the relationship between salivary microbiota composition, salivary biochemical parameters, and ADC in both children and adults. Although no significant associations were observed between phylum-level microbial composition and ADC status, specific genera and species showed positive correlations with caries activity, particularly in adults. These included Corynebacterium, Porphyromonas, Filifactor, Olsenella, Dialister or Catonella, among other microorganisms previously associated with oral dysbiosis and disease. In children, the genus Scardovia was positively related to ADC. This supports the idea that distinct microbial profiles may underlie caries development in different age groups, which may be influenced by factors such as diet, oral hygiene habits, maturity of the oral microbiome, and host immune responses. In this sense, the specie P. stomatis showed opposing correlations with caries in adults and children, positive in the former and negative in the latter.
Despite initial hypotheses, salivary biochemical parameters, including flow rate, pH, total protein concentration, and total antioxidant capacity, did not show significant associations with ADC presence in either age group.
Taken together, these findings suggest that microbial factors may play a more significant role than salivary biochemical markers in distinguishing caries activity and suggest age-specific microbial risk markers, particularly in adults. Future studies with larger sample sizes and longitudinal designs are needed to confirm these associations and explore potential mechanistic pathways.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by CEIC Hospital Clínico San Carlos. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants' legal guardians/next of kin.
Author contributions
NT-E: Resources, Investigation, Writing – original draft. CM-G: Formal analysis, Writing – review & editing. AG-V: Writing – review & editing, Investigation, Resources. PC: Formal analysis, Writing – review & editing. AM-V: Conceptualization, Writing – review & editing, Data curation. MP-C: Project administration, Supervision, Validation, Conceptualization, Funding acquisition, Writing – original draft. JA: Validation, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by the 13th call for research projects of the Alfonso X El Sabio Foundation-Santander Bank of Spain (granted number 1.013.003).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/froh.2025.1599925/full#supplementary-material
Abbreviations
DNA, deoxyribonucleic acid; TAC, total antioxidant capacity; TPC, total protein concentration; ADC, active dentin caries; UAX, Alfonso X El Sabio University.
References
1. Xu L, Chen X, Wang Y, Jiang W, Wang S, Ling Z, et al. Dynamic alterations in salivary microbiota related to dental caries and age in preschool children with deciduous dentition: a 2-year follow-up study. Front Physiol. (2018) 9:342. doi: 10.3389/fphys.2018.00342
2. Mason MR, Chambers S, Dabdoub SM, Thikkurissy S, Kumar PS. Characterizing oral microbial communities across dentition states and colonization niches. Microbiome. (2018) 6:1–10. doi: 10.1186/s40168-018-0443-2
3. Kuramitsu HK, He X, Lux R, Anderson MH, Shi W. Interspecies interactions within oral microbial communities. Microbiol Mol Biol Rev. (2007) 71:653–70. doi: 10.1128/MMBR.00024-07
4. Belstrøm D. The salivary microbiota in health and disease. J Oral Microbiol. (2020) 12:1723975. doi: 10.1080/20002297.2020.1723975
5. Gross EL, Beall CJ, Kutsch SR, Firestone ND, Leys EJ, Griffen AL. Beyond Streptococcus mutans: dental caries onset linked to multiple species by 16S rRNA community analysis. PLoS One. (2012) 7(10):e47722. doi: 10.1371/journal.pone.0047722
6. Chattopadhyay I, Lu W, Manikam R, Malarvili MB, Ambati RR, Gundamaraju R. Can metagenomics unravel the impact of oral bacteriome in human diseases? Biotechnol Genet Eng Rev. (2023) 39:85–117. doi: 10.1080/02648725.2022.2102877
7. Agarwal J, Pandey P, Saxena SK, Kumar S. Comparative analysis of salivary microbiota in diabetic and non-diabetic individuals of north India using metagenomics. J Oral Biol Craniofac Res. (2024) 14:22–6. doi: 10.1016/j.jobcr.2023.12.003
8. Strużycka I. The oral microbiome in dental caries. Polish J Microbiol. (2014) 63:127. doi: 10.33073/pjm-2014-018
9. Crielaard W, Zaura E, Schuller AA, Huse SM, Montijn RC, Keijser BJ. Exploring the oral microbiota of children at various developmental stages of their dentition in the relation to their oral health. BMC Med Genomics. (2011) 4:1–13. doi: 10.1186/1755-8794-4-22
10. Eriksson L, Lif Holgerson P, Johansson I. Saliva and tooth biofilm bacterial microbiota in adolescents in a low caries community. Sci Rep. (2017) 7:5861. doi: 10.1038/s41598-017-06221-z
11. Mira A. Oral microbiome studies: potential diagnostic and therapeutic implications. Adv Dent Res. (2018) 29:71–7. doi: 10.1177/0022034517737024
12. Lif Holgerson P, Öhman C, Rönnlund A, Johansson I. Maturation of oral microbiota in children with or without dental caries. PLoS One. (2015) 10:e0128534. doi: 10.1371/journal.pone.0128534
13. Teng F, Yang F, Huang S, Bo C, Xu ZZ, Amir A, et al. Prediction of early childhood caries via spatial-temporal variations of oral microbiota. Cell Host Microbe. (2015) 18:296–306. doi: 10.1016/j.chom.2015.08.005
14. Belstrøm D, Constancias F, Liu Y, Yang L, Drautz-Moses DI, Schuster SC, et al. Metagenomic and metatranscriptomic analysis of saliva reveals disease-associated microbiota in patients with periodontitis and dental caries. NPJ Biofilms Microbiomes. (2017) 3:23. doi: 10.1038/s41522-017-0031-4
15. Hariyani N. Global, regional, and national levels and trends in burden of oral conditions from 1990 to 2017: a systematic analysis for the global burden of disease 2017 study. J Dent Res. (2020) 99:362–73. doi: 10.1177/0022034520908533
16. Hurley E, Barrett MP, Kinirons M, Whelton H, Ryan CA, Stanton C, et al. Comparison of the salivary and dentinal microbiome of children with severe-early childhood caries to the salivary microbiome of caries-free children. BMC oral Health. (2019) 19:1–14. doi: 10.1186/s12903-018-0693-1
17. Lazarevic V, Whiteson K, Gaïa N, Gizard Y, Hernandez D, Farinelli L, et al. Analysis of the salivary microbiome using culture-independent techniques. J Clin Bioinforma. (2012) 2:1–8. doi: 10.1186/2043-9113-2-4
18. Yamashita Y, Takeshita T. The oral microbiome and human health. J Oral Sci. (2017) 59:201–6. doi: 10.2334/josnusd.16-0856
19. López-Dávalos PC, Requena T, Pozo-Bayón MÁ, Muñoz-González C. Decreased retronasal olfaction and taste perception in obesity are related to saliva biochemical and microbiota composition. Food Res Int. (2023) 167:112660. doi: 10.1016/j.foodres.2023.112660
20. Ghaferi AA, Schwartz TA, Pawlik TM. STROBE reporting guidelines for observational studies. JAMA Surg. (2021) 156:577–8. doi: 10.1001/jamasurg.2021.0528
21. Pitts NB, Ismail AI, Martignon S, Ekstrand K, Douglas GV, Longbottom C, et al. ICCMS™ guide for practitioners and educators. ICDAS Foundation. (2014).
22. World Health Organization. Oral Health Surveys: Basic Methods. World Health Organization (2013).
23. Xiao J, Grier A, Faustoferri RC, Alzoubi S, Gill AL, Feng C, et al. Association between oral candida and bacteriome in children with severe ECC. J Dent Res. (2018) 97:1468–76. doi: 10.1177/0022034518790941
24. Luo AH, Yang DQ, Xin BC, Paster BJ, Qin J. Microbial profiles in saliva from children with and without caries in mixed dentition. Oral Dis. (2012) 18:595–601. doi: 10.1111/j.1601-0825.2012.01915.x
25. Jiang S, Gao X, Jin L, Lo EC. Salivary microbiome diversity in caries-free and caries-affected children. Int J Mol Sci. (2016) 17:1978. doi: 10.3390/ijms17121978
26. Zhu C, Yuan C, Ao S, Shi X, Chen F, Sun X, et al. The predictive potentiality of salivary microbiome for the recurrence of early childhood caries. Front Cell Infect Microbiol. (2018) 8:423. doi: 10.3389/fcimb.2018.00423
27. Simon-Soro A, Sherriff A, Sadique S, Ramage G, Macpherson L, Mira A, et al. Combined analysis of the salivary microbiome and host defence peptides predicts dental disease. Sci Rep. (2018) 8:1484. doi: 10.1038/s41598-018-20085-x
28. Jiang Q, Liu J, Chen L, Gan N, Yang D. The oral microbiome in the elderly with dental caries and health. Front Cell Infect Microbiol. (2019) 8:442. doi: 10.3389/fcimb.2018.00442
29. Cubero Santos A, Lorido Cano I, González Huéscar A, Ferrer García M, Zapata Carrasco M, Ambel Sánchez JL. Prevalencia de caries dental en escolares de educación infantil de una zona de salud con nivel socioeconómico bajo. Pediatría Atención Primaria. (2019) 21:e47–59.
30. Palomar-Bonet M, Atienza M, Hernández-Ledesma B, Cantero JL. Associations of salivary total antioxidant capacity with cortical amyloid-beta burden, cortical glucose uptake, and cognitive function in normal aging. J Gerontol Ser A. (2021) 76:1839–45. doi: 10.1093/gerona/glab034
31. Anonymous. Structure, function and diversity of the healthy human microbiome. Nature. (2012) 486:207–14. doi: 10.1038/nature11234
32. Aas JA, Paster BJ, Stokes LN, Olsen I, Dewhirst FE. Defining the normal bacterial flora of the oral cavity. J Clin Microbiol. (2005) 43:5721–32. doi: 10.1128/JCM.43.11.5721-5732.2005
33. Kreth J, Zhang Y, Herzberg MC. Streptococcal antagonism in oral biofilms: Streptococcus sanguinis and Streptococcus gordonii interference with Streptococcus mutans. J Bacteriol. (2008) 190:4632–40. doi: 10.1128/JB.00276-08
34. Dewhirst FE, Chen T, Izard J, Paster BJ, Tanner AC, Yu W, et al. The human oral microbiome. J Bacteriol. (2010) 192:5002–17. doi: 10.1128/JB.00542-10
35. Huang S, Yang F, Zeng X, Chen J, Li R, Wen T, et al. Preliminary characterization of the oral microbiota of Chinese adults with and without gingivitis. BMC oral Health. (2011) 11:1–14. doi: 10.1186/1472-6831-11-33
36. Yang F, Zeng X, Ning K, Liu K, Lo C, Wang W, et al. Saliva microbiomes distinguish caries-active from healthy human populations. ISME J. (2012) 6:1–10. doi: 10.1038/ismej.2011.71
37. Ling Z, Liu X, Wang Y, Li L, Xiang C. Pyrosequencing analysis of the salivary microbiota of healthy Chinese children and adults. Microb Ecol. (2013) 65:487–95. doi: 10.1007/s00248-012-0123-x
38. Zhou J, Jiang N, Wang S, Hu X, Jiao K, He X, et al. Exploration of human salivary microbiomes—insights into the novel characteristics of microbial community structure in caries and caries-free subjects. PLoS One. (2016) 11:e0147039. doi: 10.1371/journal.pone.0147039
39. Yang F, Ning K, Zeng X, Zhou Q, Su X, Yuan X. Characterization of saliva microbiota’s functional feature based on metagenomic sequencing. Springerplus. (2016) 5:1–10. doi: 10.1186/s40064-016-3728-6
40. Chen W, Jiang Q, Yan G, Yang D. The oral microbiome and salivary proteins influence caries in children aged 6 to 8 years. BMC Oral Health. (2020) 20:1–16. doi: 10.1186/s12903-020-01262-9
41. Sun B, Liu B, Gao X, Xing K, Xie L, Guo T. Metagenomic analysis of saliva reveals disease-associated microbiotas in patients with periodontitis and Crohn’s disease-associated periodontitis. Front Cell Infect Microbiol. (2021) 11:719411. doi: 10.3389/fcimb.2021.719411
42. Yang X, He L, Yan S, Chen X, Que G. The impact of caries status on supragingival plaque and salivary microbiome in children with mixed dentition: a cross-sectional survey. BMC Oral Health. (2021) 21:319. doi: 10.1186/s12903-021-01683-0
43. Shi C, Cai L, Xun Z, Zheng S, Shao F, Wang B, et al. Metagenomic analysis of the salivary microbiota in patients with caries, periodontitis and comorbid diseases. J Dent Sci. (2021) 16:1264–73. doi: 10.1016/j.jds.2020.12.002
44. Kim Y, Jeong J, Mun S, Yun K, Han K, Jeong S. Comparison of the oral microbial composition between healthy individuals and periodontitis patients in different oral sampling sites using 16S metagenome profiling. J Periodontal Implant Sci. (2022) 52:394. doi: 10.5051/jpis.2200680034
45. Kim B, Han D, Lee H, Oh B. Association of salivary microbiota with dental caries incidence with dentine involvement after 4 years. J Microbiol Biotechnol. (2018) 28:454–64. doi: 10.4014/jmb.1710.10028
46. Plachokova AS, Andreu-Sánchez S, Noz MP, Fu J, Riksen NP. Oral microbiome in relation to periodontitis severity and systemic inflammation. Int J Mol Sci. (2021) 22:5876. doi: 10.3390/ijms22115876
47. Belstrøm D, Fiehn N, Nielsen CH, Klepac-Ceraj V, Paster BJ, Twetman S, et al. Differentiation of salivary bacterial profiles of subjects with periodontitis and dental caries. J Oral Microbiol. (2015) 7:27429. doi: 10.3402/jom.v7.27429
48. Sundström K, Mishra PP, Pyysalo MJ, Lehtimäki T, Karhunen PJ, Pessi T. Similarity of salivary microbiome in parents and adult children. PeerJ. (2020) 8:e8799. doi: 10.7717/peerj.8799
49. Wirth R, Maróti G, Lipták L, Mester M, Al Ayoubi A, Pap B, et al. Microbiomes in supragingival biofilms and saliva of adolescents with gingivitis and gingival health. Oral Dis. (2022) 28:2000–14. doi: 10.1111/odi.13883
50. Kasimoglu Y, Koruyucu M, Birant S, Karacan I, Topcuoglu N, Tuna EB, et al. Oral microbiota and dental caries data from monozygotic and dizygotic twin children. Sci Data. (2020) 7:348. doi: 10.1038/s41597-020-00691-z
51. Gül F, Karadayı S, Yurdabakan Z, Özbek T, Karadayı B. Investigating changes in salivary microbiota due to dental treatment: a metagenomic analysis study for forensic purposes. Forensic Sci Int. (2022) 340:111447. doi: 10.1016/j.forsciint.2022.111447
52. Hasan NA, Young BA, Minard-Smith AT, Saeed K, Li H, Heizer EM, et al. Microbial community profiling of human saliva using shotgun metagenomic sequencing. PLoS One. (2014) 9:e97699. doi: 10.1371/journal.pone.0097699
53. Foxman B, Luo T, Srinivasan U, Ramadugu K, Wen A, Goldberg D, et al. The effects of family, dentition, and dental caries on the salivary microbiome. Ann Epidemiol. (2016) 26:348–54. doi: 10.1016/j.annepidem.2016.03.006
54. Dashper SG, Mitchell HL, Lê Cao K, Carpenter L, Gussy MG, Calache H, et al. Temporal development of the oral microbiome and prediction of early childhood caries. Sci Rep. (2019) 9:19732. doi: 10.1038/s41598-019-56233-0
55. Gomar-Vercher S, Cabrera-Rubio R, Mira A, Montiel-Company JM, Almerich-Silla JM. Relationship of children’s salivary microbiota with their caries status: a pyrosequencing study. Clin Oral Investig. (2014) 18:2087–94. doi: 10.1007/s00784-014-1200-y
56. Claesson MJ, Jeffery IB, Conde S, Power SE, O’connor EM, Cusack S, et al. Gut microbiota composition correlates with diet and health in the elderly. Nature. (2012) 488:178–84. doi: 10.1038/nature11319
57. Zhang C, Cheng X, Li J, Zhang P, Yi P, Xu X, et al. Saliva in the diagnosis of diseases. Int J Oral Sci. (2016) 8:133–7. doi: 10.1038/ijos.2016.38
58. Pandey P, Reddy NV, Rao VAP, Saxena A, Chaudhary CP. Estimation of salivary flow rate, pH, buffer capacity, calcium, total protein content and total antioxidant capacity in relation to dental caries severity, age and gender. Contemp Clin Dent. (2015) 6:S65–71. doi: 10.4103/0976-237X.152943
59. Pyati SA, Naveen Kumar R, Kumar V, Praveen Kumar NH, Parveen Reddy KM. Salivary flow rate, pH, buffering capacity, total protein, oxidative stress and antioxidant capacity in children with and without dental caries. J Clin Pediatr Dent. (2018) 42:445–9. doi: 10.17796/1053-4625-42.6.7
60. Martins JR, Diaz-Fabregat B, Ramirez-Carmona W, Monteiro DR, Pessan JP, Antoniali C. Salivary biomarkers of oxidative stress in children with dental caries: systematic review and meta-analysis. Arch Oral Biol. (2022) 139:105432. doi: 10.1016/j.archoralbio.2022.105432
61. Simon-Soro A, Tomás I, Cabrera-Rubio R, Catalan MD, Nyvad B, Mira A. Microbial geography of the oral cavity. J Dent Res. (2013) 92:616–21. doi: 10.1177/0022034513488119
62. Kumar A, Chordia N. Role of microbes in human health. Appl Microbiol Open Access. (2017) 3:18. doi: 10.4172/2471-9315.1000131
63. Colombo APV, Bennet S, Cotton SL, Goodson JM, Kent R, Haffajee AD, et al. Impact of periodontal therapy on the subgingival microbiota of severe periodontitis: comparison between good responders and individuals with refractory periodontitis using the human oral microbe identification microarray. J Periodontol. (2012) 83:1279–87. doi: 10.1902/jop.2012.110566
64. Siqueira JF Jr, Rôças IN. Community as the unit of pathogenicity: an emerging concept as to the microbial pathogenesis of apical periodontitis. Oral Surg Oral Med Oral Pathol Oral Radiol Endodontol. (2009) 107:870–8. doi: 10.1016/j.tripleo.2009.01.044
65. Siqueira JF Jr, Rôças IN. The microbiota of acute apical abscesses. J Dent Res. (2009) 88:61–5. doi: 10.1177/0022034508328124
66. Siqueira JF Jr, Rôcas IN. Distinctive features of the microbiota associated with different forms of apical periodontitis. J Oral Microbiol. (2009) 1:2009. doi: 10.3402/jom.v1i0.2009
67. Siqueira JF Jr, Rôças IN. Diversity of endodontic microbiota revisited. J Dent Res. (2009) 88:969–81. doi: 10.1177/0022034509346549
68. Mark Welch JL, Rossetti BJ, Rieken CW, Dewhirst FE, Borisy GG. Biogeography of a human oral microbiome at the micron scale. Proc Natl Acad Sci U S A. (2016) 113:E791–800. doi: 10.1073/pnas.1522149113
69. Tanner A, Kent RL Jr, Holgerson PL, Hughes CV, Loo CY, Kanasi E, et al. Microbiota of severe early childhood caries before and after therapy. J Dent Res. (2011) 90:1298–305. doi: 10.1177/0022034511421201
70. de Almeida CM, Petersen PE, André SJ, Toscano A. Changing oral health status of 6-and 12-year-old schoolchildren in Portugal. Community Dent Health. (2003) 20:211–6.14696739
Keywords: dentistry, microbiology, caries, oral pathology, periodontitis, metagenomic, saliva
Citation: Tamayo-Estebaranz N, Muñoz-González C, Gil-Valcárcel AM, Calvo López-Dávalos P, Martín-Vacas A, Paz-Cortés MM and Aragoneses JM (2025) Salivary microbiota profile in adult and children population according to active dentin caries: a metagenomic preliminary analysis. Front. Oral Health 6:1599925. doi: 10.3389/froh.2025.1599925
Received: 31 March 2025; Accepted: 15 July 2025;
Published: 28 July 2025.
Edited by:
Muhammed Manzoor, University of Helsinki, FinlandReviewed by:
Denis Bourgeois, Université Claude Bernard Lyon 1, FranceSigrun Eick, University of Bern, Switzerland
Copyright: © 2025 Tamayo-Estebaranz, Muñoz-González, Gil-Valcárcel, Calvo López-Dávalos, Martín-Vacas, Paz-Cortés and Aragoneses. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrea Martín-Vacas, YW1hcnR2YWNAdWF4LmVz
†These authors have contributed equally to this work