 Gary Paul Nabhan
Gary Paul Nabhan Patricia Colunga-GarcíaMarín
Patricia Colunga-GarcíaMarín Daniel Zizumbo-Villarreal
Daniel Zizumbo-Villarreal- 1Desert Laboratory of Tumamoc Hill and Southwest Center, University of Arizona, Tucson, AZ, United States
- 2Retired, Mérida, México
Climate change is aggravating agricultural crop failures, and the paucity of wild food harvests for Indigenous desert dwellers in Mexico and the U.S. This food production crisis challenges ongoing efforts by Indigenous communities in obtaining greater food security, prompting them to reconsider the value of traditional Indigenous food systems in both Mesoamerica and Arid America, two adjacent centers of crop diversity. While food production strategies in these two centers share many features, the food plant diversity in the Western Mesoamerican region appears to be greater. However, a higher percentage of plants in Arid America have adapted to water scarcity, heat, and damaging radiation. The phytochemical and physiological adaptations of the food plants to abiotic stresses in arid environments offer a modicum of resilience in the face of aggravated climate uncertainties. By comparing food plant genera comprising Western Mesoamerican and Arid American diets, we detected a higher ratio of CAM succulents in the wild and domesticated food plant species in the Arid American food system. We conclude that food plant diversity in the ancestral diets of both centers can provide much of the resilience needed to advance Indigenous food sovereignty and assure food security as climate change advances.
Introduction
There is the smell of danger in the dry air: A recent analysis summarizing the U.S. Environmental Protection Agency's “Climate Change Impacts and Risk Analysis” concluded that by 2090, climate-induced impacts on agriculture and 21 other natural resource-based sectors of the economy could cost over $224 billion more per year to the U.S. economy alone, with impacts in Mexico and Canada approaching similar levels (Martinech, 2018; Nuchatelli, 2019).
It is likely that because of water scarcity, increased heat, and damaging solar radiation during flowering and fruiting—in addition to land degradation—the yields of temperate-adapted grains, vegetables and fruits will decline enough to pose food security risks for most Indigenous and campesino farmers in the Americas (Altieri and Nicholls, 2009; Nabhan, 2013, 2020). This may be due to herbaceous crops like broccoli, common beans, cowpeas, groundnuts, maize, rice, sorghum, tomato and wheat hitting temperature thresholds near flowering times (Challinor et al., 2005; Luo, 2011). In addition, climate catastrophes may disrupt or set back the valiant efforts by Indigenous communities to regain or retain food sovereignty (Peña et al., 2017; Mihesuah and Hoover, 2019). We endorse and endeavor to advance the definition of food sovereignty as stated in the 2007 Declaration of Nyéléni, released at the first global forum on food sovereignty in Mali (https://nyeleni.org/spip.php?article290) [accessed Feb 21, 2022]:
“Food sovereignty is the right of peoples to healthy and culturally appropriate food produced through ecologically sound and sustainable methods, and their right to define their food and agriculture systems. It puts the aspirations and needs of those who produce, distribute and consume food at the heart of food systems and policies rather than the demands of markets and corporations.”
At the same time, we conceded Indigenous farmers on several continents are now among the ranks of those being called “climate refugees” (Li et al., 2021), and that even their basic food security—their reliable access to a sufficient quantity of affordable, nutritious food—has been placed at further risk by climate change, neo-liberal economic globalization and the Covid-19 pandemic.
Overall, the current rates of global climate change are gravely impacting food production and security while increasing health risks for all residents of North America. Additionally, climate catastrophes and carbon emissions have generated more than $820 billion per year in additional physical and mental health care costs to U.S. and Mexican farm-based food system economies (Crimmins et al., 2016; Ebi and Hess, 2020; De Alwis and Limaye, 2021).
We are especially concerned by these climatic challenges to Indigenous communities in deserts, along coastlines or in mountainous regions who are being differentially affected by climate catastrophes and sea level rising. It is important to note the tremendous strides Indigenous peoples in the U.S. and Mexico have made in their social movements in affirming food sovereignty to gain further control over their dietary options in a globalized food system (Peña et al., 2017; Mihesuah and Hoover, 2019). We do not wish to see such initiatives compromised or disrupted by climatic stresses or by economic pressures.
Nevertheless, the combination of the Covid pandemic, high unemployment, rising gas prices, climate change and accelerating globalization pressures on Indigenous food systems have reduced dietary diversity in Indigenous pueblos at a critical time when such a variety of nutritional sources is desperately needed.
One tangible example was recently documented (Nabhan and Molina, 2021) during the 2022 phase of the Covid-19 pandemic, which was also one of the hottest, driest years suffered in recent memory by Indigenous communities in coastal Sonora, Mexico. The Comcaac, or Seri communities of coastal Sonora, Mexico, historically used between 95 and 100 wild native plants for fresh foods and beverages. However, only 12 of those species continue to be routinely harvested for food by the 1,500 inhabitants of two fishing villages today, and by far less than half of their population. Instead, they rely on an extremely limited inventory of domesticated crops such as fresh fruits and vegetables that may be found in just seven local grocery stores.
The Comcaac diet has lost 57% of the plant species richness found in their historic diet. This includes considering all purchased fruits and vegetables as well as the occasional wild foraging of native fruits, seeds, tubers and shoots. Some plant populations are no longer accessible due to the loss of aboriginal lands, while others have been destroyed altogether by land clearing. Additionally, other plant species have fallen out of use due to the loss of traditions, acculturation and shifts in livelihoods. Similar trends have occurred in other Indigenous villages. That is why we are assisting them in a three-year program where they discuss their aspirations for enhancing food sovereignty, while we provide technical assistance to help achieve food, water, and energy security.
The possibility that Indigenous cultures' own ethnobotanical resources can be used as an incentive for renewed agro-ecological development and plant domestication was first proposed in Mesoamerica by Colunga-GarcíaMarín and Zizumbo-Villarreal (1993), Colunga-GarcíaMarín et al. (2007) and Zizumbo-Villarreal et al. (2012, 2016). It prompted Mexico's activist-scholars to initiate thoughtful, thorough, and far-reaching efforts to revive the original structure and composition of the Mesoamerican farming and food plant gathering in ways that will strengthen Indigenous food sovereignty initiatives (e.g., Calvo and Rueda-Esquibel, 2015).
Similarly, Indigenous populations in the U.S. Desert Southwest have self-initiated several programs to safeguard, produce and prepare “native foods” from their own farming and wild foraging traditions to deal with food security and sovereignty issues (Edaakie and Enote, 1999; Tohono O'odham Community Action., 2010; Mihesuah and Hoover, 2019). This native farming and foraging revival began within Indigenous communities well before climate change was recognized as a stressor. Even then, it was clear that food security interventions and cultural food sovereignty affirmations could be effective in dealing with several historic issues that were both culturally devastating and economically burdensome (Kuhnlein and Receveur, 1996; Minnis, 2021).
Our objectives for this reappraisal and revival of healthful, plant-based food systems that emerged in the pre-colonial era in what is now Mexico and the Southwestern U.S. are that: (a) they can reduce crop failures from climate-related water scarcity and heat waves, and that (b) they may help restore food sovereignty to Indigenous and campesino communities in ways that reinforce their cultural identity, and reaffirm their rights to land, water, seeds and local food processing options (Peña et al., 2017).
This issue is important for the future as most of Mexico's population now dwells in hot, dry climates, where the arid food-producing landscapes dominate 60% of the national territory (Appendini and Liverman, 1994; Pontifes et al., 2018). Across much of Latin America, temperature thresholds and drought are beginning to limit the production of most maize and bean varieties (Rodríguez-De Luque et al., 2016; Stiller et al., 2021). Similar trends now occur in the Western U.S. (Nabhan, 2013).
We therefore wish to compare, contrast and, if possible, revive elements of food systems in what we call the Mesoamerican and Arid American centers of diversity. Our ultimate goal for this article is to detail how restoring the broad diversity of wild and cultivated plants once found in the ancestral diets of these areas, originated in the Archaic or preceramic period (Zizumbo-Villarreal et al., 2014, 2016) may enhance human health, economic wellbeing, food security and resilience in the face of climate change.
Geographic Context, Materials and Methods
For the purposes of this article, we compared two subareas of the centers of diversity known as Mesoamerica and Aridoamerica (Figure 1), two terms coined by Kirchhoff (2000) to describe the two cultural areas of what is now central Mexico and part of Central America, and Northern Mexico and the Southwestern U.S. This Mesoamerican ethno-ecological region refers to the megadiverse landscapes extending from central Mexico, Guatemala, El Salvador, Honduras, Nicaragua, and part of Costa Rica, where multiple cultures, languages and foodways flourished in pre-Colonial eras, elements of which continue to persist today. Here, we focus more narrowly on Western Mesoamerica along the Pacific coast of Mexico to compare with the western reaches of a considerably more arid region to its north.
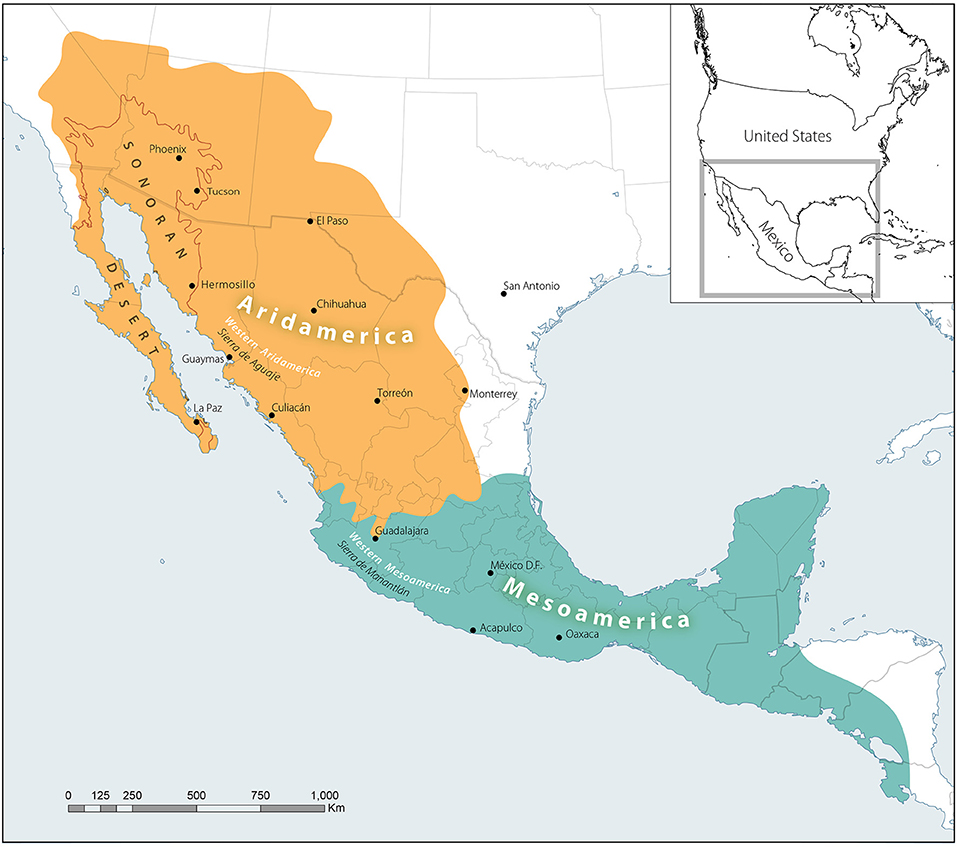
Figure 1. Aridamerican and Mesoamerican Centers of Diversity. Modified from Kirchhoff (1954, 2000); Nabhan (1985), and Nabhan et al. (2020).
The use of the term Aridoamerica is much more common among Mexican scholars than among American ones. In this paper, we have integrated Kirchoff's Oasisamerica (Kirchhoff, 1954) — with its nucleated set of irrigated agricultural communities into the larger matrix of “seasonally dry-farmed” (de temporal) communities of the cultural area which he called Aridoamerica. To distinguish the units of Kirchhoff's Oasisamerica and Aridoamerica from our re-defined secondary center of crop diversity, we use the new delineation of “Arid America”. Our new geographic delineations of these two centers were elaborated by our colleagues in Mexico and the U.S. who contributed to an agroecological research article complementary to this one (Nabhan et al., 2020), based in Nabhan (1985). It integrates floristic, vegetational, agroecological, ethnobotanical, anthropological and linguistic factors first elaborated by Hernández-Xolocotzi (2013) to find the most parsimonious fit of the boundaries of each region.
As defined here, both the Mesoamerican and Arid American centers of crop diversity fall primarily within the larger Neotropical phytogeographic region which biogeographer Rzedowski considered to be “MegaMexico” that extended into the U.S. Desert Southwest (Rzedowski, 1978). The Arid American area is a secondary, binational center of crop diversity that spans the Sonoran Desert, including Baja California's deserts; and to the east, the higher elevation Chihuahuan Desert, which includes the Zacatecas-Potosí Desert. Its biocultural food system has featured more floristic and cultural diversity than most other desert regions in the Americas (Luque-Agraz et al., 2016).
To develop a full characterization of the Arid American center of diversity, we have amplified and corrected Nabhan (1985) initial inventory of domesticated crop plants to use as a point of departure for the first published comparison of the archaic diets of Mesoamerica and Arid America (Supplementary Table S1).
First, we determined what annual and perennial crops dominated agriculture in each center during the pre-Colonial era. Next, we documented which wild native plant species likely preceded, and complemented or underpinned this domesticated crop inventory from the late pre-Invasion era, through historic “colonial” eras, rounding out the ancestral diets of “Indigenous” or “Native American” communities of Arid America and Western Mesoamerica.
To do so, we have drawn upon two ethnobotanies from each of three Arid America subregions:
(1) Baja California, Aschmann (1959) for the Cochimí of the central desert and Wilken-Robertson (2018) for the Kumeyaay of the semi-arid foothills; (2) the mainland's Sonoran Desert, including Rea (1997) for the Upper Pima of the Arizona Uplands and Felger and Moser (1985) for the hyper-Arid Gulf Coast of Sonora; and (3) the Chihuahuan Desert, including Latorre and Latorre (1977) for the Kickapoo of Coahuila and Texas, as well as Solano-Picazo (2018) for the Wixarika (Huichol) of San Luis Potosí. These have been supplemented by regional classics such as Hodgson (2001) Food Plants of the Sonoran Desert and Hernández-Sandoval et al. (1991) Plantas Utiles de Tamaulipas. These six references yield a rather comprehensive view of the wild or semi-cultivated food plants whose presence in these regions predate Spanish Invasion, with a focus on nutritionally significant genera that continue to be consumed by two or more Indigenous cultures across the entire Arid American region.
Next, we will analyze which of these wild food plants were crop relatives or congeners of the domesticated species listed in the Supplementary Table S1 inventory. Our primary hypothesis is that well before fully domesticated plants entered their traditional cuisines, Indigenous communities were already familiar with and gastronomically utilized several crop wild relatives in their archaic diets (Zizumbo-Villarreal et al., 2012; Contreras-Toledo et al., 2018; Riordan and Nabhan, 2019). Our secondary hypothesis is that these crop wild relatives have characteristics that may make them ideal food crops in a climate-changed world. Our reason for focusing more on the wild food plant species rather than on the Indigenous domesticated seeds crops of Western Mesoamerica and Arid America is to minimize the risk of cultural appropriation and legal enclosure of the particular seed stocks that fall under the “farmers' right” of specific Indigenous communities. By and large, the rejuvenation of the uses of widespread wild food plants does not pose such a risk of violating the food sovereignty of any single Indigenous community.
To compare the relative richness of crop wild relatives in a local flora of Western Mesoamerica with one from Western Arid America, we have selected the flora of the Sierra de Manantlán Biosphere Reserve on the Jalisco-Colima border (Vásquez-García, 1995) and the flora of Cañón de Nacapule in the Sierra de Aguaje, part of the Cajón del Diablo Biosphere Reserve in coastal Sonora, Mexico (Felger et al., 2017).
To further refine our retrodiction of the archaic diets in each center, we give particular attention to those documented among cultures of Arizona, Baja California and Sonora to represent the Arid American food systems, and those of Jalisco and Colima to represent the Western Mesoamerican food systems. Because early ethnographies typically lack scientific names for the wild plants included in them, we cross-referenced their regional and Indigenous folk names with updated scientific nomenclature from Martínez (1979), Inés-Olaya (1991), Moerman (1998), Hodgson (2001), and Avitia-García and Castillo-González (2002). We included domesticated foods recorded in compendia such as Burns et al. (2000), Dunmire (2004), Zizumbo-Villarreal and Colunga-GarcíaMarín (2010), and those in the Supplementary Table S1 derived from Nabhan (1985).
Results
Characterizing the Annual Food Crop Biodiversity of Arid American and Western Mesoamerican Food Systems
Of some thirty species of annual or annualized perennials domesticated as crops in the pre-Colonial era, each of the two centers of crop diversity harbors roughly the same number of these short cycle, warm-season food staples (Supplementary Table S2): 19 species for Mesoamerica and 2O for Arid America. It is abundantly clear that during pre-Invasion eras, Mesoamerican cultures domesticated far more annual food crops plants than did Arid American cultures. The mix of plant families in these two sets of regional annual crops is much the same: composites, cucurbits, grasses, legumes, as well as pseudo-cereals from the amaranth family dominate the mixes.
Many of the Arid American cultivated plants were rigorously selected by both weather and culture to be short-cycle crops, maturing with dry seeds in as little as 36 to 55 days during the monsoon season of mid-summer to early fall, to “escape” drought rather than to endure it. These crops mimic summer “ephemeral” wildflowers in the Sonoran and Chihuahuan deserts in that they quickly germinate, flower, set fruit and die before the late autumn drought period sets in. Since most of these crops utilize the C3 metabolic pathway, they require considerable soil moisture each week they are alive but reduce cumulative consumptive water use by maturing quickly. In contrast, most annual West Mesoamerican crops are facultative perennials, which persist in gardens and fields for at least seven months during the warm season, but may persist and set fruit for 12 to 15 months before senescence. Often, root fungi, other diseases, and pests terminate their growth, not climatic constraints.
Over the last four millennia, no fewer than 22 domesticated annual food crops were culturally dispersed from Mesoamerican food systems into Arid American food systems (Burns et al., 2000; Dunmire, 2004). Most of these require much more irrigation than the desert-adapted food crops like tepary beans, jack beans, little barley, and sagui or Sonoran panicgrass (Nabhan and de Wet, 1984).
Arid American cultures obtained many of their staple foods from the south via group-to-group diffusion across a Uto-Nahua linguistic continuum (Merrill et al., 2009). Probably, most domesticated annuals or annualized perennials grown for food in Arid America of the last several millennia began to diffuse into the more northern, arid region along the Western Mexican coastal trade routes beginning between 6000 and 5500 calibrated years before present (Merrill et al., 2009; Mabry1). The Las Capas site in the Western Tucson Basin has yielded three direct radiocarbon dates on maize remains between 5,700 and 4,500 calibrated years before present [Vint, 2015, 2018; Mabry (see text footnote 1, respectively)]. These are currently the oldest domesticated crop dates in the Southwest U.S./Northwest Mexico region, but archaeologists anticipate that early dates for perennial crops will eventually be reported as new genetic and archaeological methodologies are utilized in the region.
In a few cases, we can posit that because all Mesoamerican food crops adapted to the wet Neotropics did not grow well in the hot, dry lowlands of Arid America, another, more desert-adapted set of congeners of similar utility were utilized. For example, the domestication of tepary beans in Arid America appeared well after common beans, lima and runner beans were domesticated in Mesoamerica (Ford, 1981). We might hypothesize similar processes of “relay domestication” into more northerly, arid climes with Cucurbita, Solanum, Physalis, Jaltomata, among other annual crop genera were recruited to play a similar role in diets as their tropical counterparts (see Rodríguez and Spooner, 1997; Louderback and Pavlik, 2018). The broad climatic differences between the centers of diversity also influenced the prevailing plant chemical defenses in the sets of annual crops dominating each of the two cultural regions, a topic which we will address elsewhere.
These New World domesticated annuals have remained among the most important warm-season food crops in both biocultural food systems, but recent severe heat waves, prolonged drought and water scarcity associated with climate change are now impacting them in several ways, such as extremely high summer temperatures causing abortion of flowers and fruits (Altieri and Nicholls, 2009; Nabhan, 2013). This is disconcerting, since they have provided much of the calories and complex carbohydrates to their Indigenous communities for the last three millennia. In many ways, they have provided a now imperiled structural matrix, or “backbone”, of the food systems involving most cultures in Mesoamerica and Arid America up until the last several decades.
Characterizing the Perennial Food Crop Biodiversity of Historic Arid American and West Mesoamerican Food Systems
The domestication of some perennial food plants was once more difficult to discern than that of annual crops, but recent methodological advances have revealed many more domesticated perennials in North America than previously recognized (Colunga-GarcíaMarín et al., 1986; Hernández-Xolocotzi, 1993; Casas et al., 1999, 2002). Of 43 perennial food crop species, once found between the two cultural regions since pre-Invasion eras, 34 of these perennial crop species continue to be found in West Mesoamerica, while 26 perennial crop species continue to be found in Arid America (Supplementary Table S3). In general, there appears to be no sharp break, but is rather a genetic continuum from wild to domesticated in most of these perennial crops of Aridamerica and Mesoamerica than in the perennial crops from Eurasia and North Africa that were introduced during the Colonial era (Ezcurra2).
A semi-cultivated perennial, Palmer's saltgrass (Distichlis palmeri) of the Colorado River delta, fell out of management in Arid America but has since been revived (Yensen, 2008), while another cereal, foxtail millet (Setaria parviflora), has apparently disappeared altogether as a crop from both cultural regions (Callen, 1967; Austin, 2006).
Often ignored by early archaeologists seeking out the origins of domesticated crops, these plants do not exhibit morphological divergence from their wild ancestors as dramatically as annual domesticates do [Colunga-GarcíaMarín and Zizumbo-Villarreal, 1993; Casas et al., 2002; Louderback and Pavlik, 2017; Ezcurra (see text footnote 2, respectively)]. However, these resilient sets of perennial crops in Arid America and Mesoamerica survive drought and heat by deep roots tapping into deep soil moisture, by sloughing off branches during drought, or by extended dormancy. They may also sequester far more carbon in the soils of multi-cropped milpas and agroforestry-based orchards than do annual crops originating in either biocultural food system.
In contrast to the diverse perennial assemblage associated with the Mesoamerican center of diversity and its milpa fields and solar/huerta (dooryard garden) agroecosystems, only the food systems at the southernmost edge of Arid America center retain much diversity of perennial food crops. North of the Rio Soto La Marina, Rio Conchos, Rio Mayo and Rio Yaqui there were very few tree crops until Spanish introduction of Old-World fruits and nuts in the Colonial era (Burns et al., 2000; Dunmire, 2004).
Arid American perennial crops were largely limited to succulent plants utilizing the Crassulacean Acid Metabolism (CAM) pathway because of the biotic and abiotic stresses on fruit trees historically posed by highly variable, scarce rainfall as well as challenging heat, transpiration rates, and pests. These CAM succulents include agaves and cacti. Importantly, 76% of Arid American perennial crops utilize the CAM pathway, while only 66% of Western Mesoamerican crops use the CAM pathway. The fruit crops of both trees and vines using the C3 pathway are far more vulnerable to drought stress and crop failure, unless frequently irrigated.
The water-conserving CAM of cacti and succulents such as agaves allowed them to produce more edible biomass on less moisture than needed by C3 and C4 crop species from the tropics, including maize (Nobel, 2009). We predict that climate change will increasingly constrain yields of fruit and nut trees that use the C3 photosynthetic pathway and thereby suffer high transpiration rates. In our view, many fruit tree orchards in arid and semi-arid/subtropical landscapes will need to be replaced with CAM succulents such as agaves, prickly pears and columnar cacti if their farmers are to economically weather climate change.
In short, cultivators in the Arid American food system once relied on a relatively greater species richness of domesticated succulent crops, as opposed to trees and woody vines, in their cultivation of perennial food plants than did cultivators in the wetter Mesoamerican food system.
There are both agroecological and human health reasons for reviving and extending the cultivation of these CAM succulent crops in dry lands (Leach and Sobolik, 2010). First, the water-conserving Crassulacean Acid Metabolism of cacti and succulents such as agaves allowed them to produce equal tonnages of edible biomass using just half to one-sixth of the moisture needed to provide the same yields by C3 and C4 crop species from the tropics, including maize (Nobel, 2009).
Several agaves were independently brought into cultivation on well over 200,000 hectares in northwestern Arid America food systems on the edges of the Sonoran Desert. They included Agave delameteri, A. murpheyi, A. phillipsiana, A. sanpedroensis, A. verdensis and A. yavapaiensis (Fish et al., 1985; Hodgson, 2012; Hodgson and Salywon, 2013; Hodgson et al., 2018), while cultivation of a seventh species remains debated (Nabhan et al., 2019).
Toward the southeastern edges of Arid American food systems where several giant agaves used in the production of fermented pulque beverages went through the initial phases of their domestication process [Ezcurra (see text footnote 2, respectively)], these species have long been cultivated on hundreds of thousands of hectares. These magueyal plantations- with giant agaves often “alley-cropped with annuals—have included A. americana, A. angustifolia, A. mapisiga, A. lophantha and A. salmiana.” These succulent species have long been cultivated and culturally dispersed in the Chihuahuan Desert-Altiplano ecotone as much as in Mesoamerica itself (Gentry, 1982).
The same may be true with the prickly pear species and varieties that dominate the extensively managed nopaleras in the Chihuahuan Desert-Altiplano ecotone. Some appear to include hybrid clones derived from multiple lineages, including Opuntia leucotricha, O. megacantha, O. streptacantha and O. tomentosa (Griffith, 2004). Aside from this baffling array of clones that historically were all lumped into Opuntia ficus-indica, there may additional domesticated prickly pears that were separately domesticated in Arid America, such as Opuntia durangensis and possibly Opuntia robusta in the fluctuating border of Mesoamerica and Arid America (Colunga-GarcíaMarín et al., 1986).
To summarize, cultivators in the Arid American secondary center of diversity relied on a greater percentage of succulent crops than cultivators in the primary center of Mesoamerica. We hypothesize that the traditionally-processed foods and beverages from these succulent crops of Arid America comprised a greater portion of Indigenous diets than they did in the Mesoamerican center (Leach and Sobolik, 2010). They may need to do so again.
Characterizing the Wild Food Plant Biodiversity of Arid America and Western Mesoamerica
In addition to domesticated plants in both Arid American and Mesoamerican centers of crop diversity, Indigenous communities in Arid America have continued to forage for a significant number of food and beverage sources. These include over 235 wild desert plant species from at least 125 genera and 60 families on monocots and dicots (Hernández-Sandoval et al., 1991; Hodgson, 2001).
While it is beyond our current capabilities to make a similar estimate of widely used food and beverage plants from Mesoamerica, we are relatively certain that the ethnographically documented inventory of wild foods species is more numerous for Western Mesoamerica than for Western Arid America (Mapes and Basurto, 2016). If only for its greater surface area, rainfall, and floristic diversity, Western Mesoamerica most likely has far more species, genera and families of food and beverage plants.
It is likely that Supplementary Table S4 is a relatively modest underestimate of the total number of the characteristic food and beverage plants of historic Arid American food systems, since we have not included all microendemics or famine foods—only those known by two or more cultures in the region (Hernández-Sandoval et al., 1991; Hodgson, 2001; Minnis, 2021).
While 225 food and beverage plants are certainly enough to draw upon through local harvesting and intra-region trade, there are some remarkable patterns evident within this Arid American food system. For example, nearly a fourth, or 23%, of the characteristic wild food and beverage plant species in Arid America are succulents that utilize the CAM pathway for high water use efficiency photosynthesis. We do not know of any comparably high estimate for any other biocultural region in the world.
Indigenous inhabitants of the Arid American center historically drew upon a particularly high number of edible species in the genera of Agave, Amaranthus, Atriplex, Cylindropuntia, Echinocereus, Ferocactus, Opuntia, Physalis, Quercus, Randia, and Salvia. That five of these ten genera are CAM succulents and another two are drought hardy trees suggests that these food plants may help form a basis for climate-resilient food security in the future (Nabhan et al., 2020). As we will document in later discussions, these genera are particularly rich in bio-active compounds that can potentially reduce diseases and maladies of oxidative stress. In addition, the plants from five succulent genera have also been utilized to elaborate the probiotic beverages we and our colleagues have discussed in other papers (e.g., Ojeda-Linares et al., 2021).
In general, Supplementary Table S4 makes it abundantly clear that Arid American food systems before the Spanish Invasion may not have been as rich in wild species as Mesoamerican food systems. However, neither were they impoverished, or lacking a variety of plants that provided a diverse array of nutrients. This species richness of food plants is evident in 28 genera of the crop wild relatives compiled for Arid America and Northwestern Mesoamerica in another paper of ours using the same data base (Nabhan et al., in press). It determined that there are at least 28 genera and 43 wild species of crop wild relatives that have been ethnographically documented as food plants in Northwestern Mesoamerica, compared to 21 genera and 48 wild species in Arid America. There are clearly many species of crop wild relatives in each region whose use as food probably predated the presence of their domesticated congeners (Zizumbo-Villarreal and Colunga-GarcíaMarín, 2010), which could again be utilized as foods in the future (Contreras-Toledo et al., 2018; Riordan and Nabhan, 2019).
In short, certain domesticated species never really “replaced” or “made obsolete” their wild congeners as foods, especially during periods of drought or famine (Mapes and Basurto, 2016; Minnis, 2021). To this day, the popularity of the wild foods such as amaranth greens, chiltepín peppers, wild grapes and plums, and prickly pear cactus fruits in Arid America has not waned. Many of the crop wild relatives have the potential to provide more yield stability under stressful climatic conditions than do their domesticated congeners. They are also ideal to use as rootstock, trap crops for pests in hedgerows, and as pollinator attractants in orchards (Riordan and Nabhan, 2019).
Supplementary Table S5 compares the richness of crop wild relatives in two local floras, each a representative “surrogate” for its center of diversity. We compared two local floras to the crop wild relatives lists in Contreras-Toledo et al. (2018) and Riordan and Nabhan (2019). Of course the comparability of these two local floras is not geographically optimal, but the lists are taxonomically up to date, and comprehensive. One of the floras is derived from the Sierra de Manantlán of Jalisco and Colima, not far from the putative “cradle” of Mesoamerican domestication of maize and beans in the Rio Balsas watershed (Vásquez-García, 1995, with updates on file at the biosphere reserve). The other is derived from the Sierra El Aguaje of coastal Sonora, on the Sonoran Desert ecotone with semi-arid subtropical thornscrub. The Sierra del Aguaje lies within the 530,000 ha Guaymas region, and harbors roughly 700 vascular plant species (Felger et al., 2017). In contrast, the Sierra de Manantlán area—surrounding the biosphere of the same name—covers less than a fourth of the area of the Guaymas region, 140,000 ha, yet harbors at least 2,770 vascular plant species. That is nearly four times the species richness of the desert region. As one might expect, the Sierra de Manantlán in Mesoamerica is home to many more genera, 45, of wild relatives than the number of genera represented in the Sierra del Aguaje in Arid America, 17.
Remarkably, the Sierra de Manantlán conserves in situ over 330 species of crop wild relatives compared to the 30 species in the Sierra del Aguaje reserve. We tentatively project that the Mesoamerican biosphere reserve harbors ten times the number of species than the Arid American biosphere reserve, even though the latter is roughly four times larger in land area. These trends suggest three patterns: (1) Mesoamerican farmers had far more opportunities to recruit and domesticate food plant species from local wild floras; (2) the constraints on those opportunities in desert areas may have encouraged Arid American farmers to actively seek out crops first domesticated in more tropical climes; or (3) the Arid American cultivators sought to diversify the number of locally adapted landraces of the few drought-tolerant species they brought into cultivation that were derived from their own regional flora.
Discussion
As explicitly stated earlier, our goal is to detail similarities and differences in the composition of the archaic diets of Indigenous communities in two adjacent centers of crop diversity, Northwestern Mesoamerica, and Arid America. Understanding the differences in the diversity of wild and domesticated food plants in these two centers may help broaden our perspective on how to gain more food system resilience in the face of climate change, especially for Indigenous communities who have valiantly struggled to maintain their food sovereignty options.
Our results echo the distinctions between a primary and secondary center of crop diversity highlighted by many crop scientists, including Kumar (2016). As a primary center, Mesoamerica has greater domesticated crop diversity and greater diversity of crop wild relatives used as foods, with a greater species richness of wild food plants overall. As a secondary center, Arid America has relative lower crop diversity, especially for domesticated perennials and crop wild relatives. Nevertheless, the diets of Arid America harbor a high number of drought- and heat-adapted food plants, especially wild and domesticated succulents like agaves and cacti, as well as many wild “famine foods” still in use.
Our documentation and analysis indicate that currently the ancestral diets of both Arid America and Northwestern Mesoamerica have an undervalued diversity of food and beverage plants that are already cultural acceptable and accessible to many Indigenous communities. These plants also demonstrate superb adaptations to the stresses of water scarcity, heat and damaging solar radiation that will become more valuable to human wellbeing as climate change proceeds. There is an extremely high ratio of water-conserving succulent plants with CAM photosynthetic pathways compared to plants utilizing the C3 and C4 photosynthetic pathways found in archaic Arid American diets which have fallen out of daily cultural use. In contrast, more of the considerable species richness in CAM plants historically found in Mesoamerican diets remain utilized in contemporary Mexican diets to this day.
Food systems scholars express concern that the many technological advances in food harvesting, storage, processing technologies and medical care now function as disincentives for embracing or reviving the labor-intensive foraging, farming and food processing practices of the past. We do not deny that there remain many formidable social, ecological, and/or economic disincentives that keep contemporary Indigenous communities from fully reviving some of their traditions of plant food procurement.
Nevertheless, there are now many agricultural, nutritional, medical, and even spiritual or cultural reasons for doing so. The possibilities for “rebirthing,” “reviving” or “restoring” these foods and beverages derived from ancestral diets are not inevitably beyond affordability in Indigenous communities today.
This is particularly evident in communities where philanthropic or governmental subsidies for producing or harvesting healthy (including “native”) foods are offered to Indigenous communities in either Mexico or the U.S. For example, the USDA Women, Infants and Children (WIC) food program subsidizes the collection or propagation of certain native foods, as does the Comisión Nacional Forestal (CONAFOR) in Mexico (https://www.gob.mx/conafor/articulos/artesanias-sonorenses-herencia-de-los-seris?idiom=es; https://www.conafor.gob.mx/EstudiosRegionalesForestales/EstudioRegionalForestal_UMAFOR0302_PSSG.pdf.; Segura-Aguilar et al., 2020) [accessed Feb. 21, 2022].
As Colunga-GarcíaMarín et al. (2007) have proposed elsewhere, for many Indigenous communities, their most healthful and secure “future may be ancestral”. As Nabhan and colleagues have proposed, “the cultivation of many of these Indigenous foods of Arid America is needed in newly designed or renovated agroforestry systems to address the emerging food security and agricultural crises” due to climate change (Nabhan, 2020; Nabhan et al., 2020) triggered by climatic changes and the pandemic. The diversity of these regionally adapted food crops—when planted in perennial-dominated polycultures—may restore land health, especially soil moisture holding capacity, while reducing crop consumptive water use and providing yield stability in the face of climatic uncertainty (Nabhan et al., 2020).
Conclusions
We have validated our primary hypothesis that prior to when domesticated crop plants entered their food production strategies, Indigenous communities were already familiar with and gastronomically utilized numerous crop wild relatives that could be used in their archaic diets. Our secondary hypothesis is potentially valid, but remains to be fully tested, accepted, or rejected: that many of these wild congeners of crops—as opposed to domesticated crop seedstocks— have agroecological and nutritional value for enhancing Indigenous food security in a future hotter, drier world. Nervertheless, as the recent documentary film Gather (sponsored by the First Nations Development Institute) illustrates, there is already a growing movement among Native Americans to utilize community-based wild harvesting to reclaim their spiritual, political, and cultural identities through food sovereignty (https://www.nativefoodsystems.org/).
To advance their food plant options for integrating into Indigenous food sovereignty initiatives to weather climate change, we have assembled the first ethnographically documented inventory of food plant diversity in two adjacent centers of biocultural diversity in the Americas. We have concluded that the following geographic patterns in food systems emerged prior to the Spanish Invasion of Arid America and Mesoamerica, and that many of their features are still viable agroecological assets or food sovereignty strategies for agriculture and wild plant foraging today:
1. Both centers still harbor a significant diversity of plant foods with a wide variety of plant growth forms such as trees, herbs, vines, succulents, and perennials. These food plants employ a broad array of phytochemical and physiological adaptations for producing food in hot, dry climates. These adaptive strategies may become even more important to Indigenous communities as they struggle to survive the hotter, drier climates that humankind is increasingly facing with global change.
2. While the Mesoamerican center is much more floristically diverse, with greater crop diversity and species richness in wild food plants than in Aridamerica, the latter region has an unusually high percentage of endemic succulent food plants which have been elaborated into prebiotic foods and probiotic beverages for millennia. Renewed domestication, agricultural production and processing of these healthful native foods and beverages should be an explicit goal in redesigning the food systems in both centers of crop diversity.
3. Given that many Indigenous communities have taken it upon themselves to reintegrate these plants into their contemporary diets as a strategy toward achieving greater food sovereignty, agricultural scientists, educators and policymakers should offer both technical and financial support whenever requested to help them achieve their explicit desires and aspirations.
4. Far from food systems of “Mexican origin” being dominated by maize, beans, and squashes (e.g., Calvo and Rueda-Esquibel, 2015, who include these three crops in 57% of their recipes), the ancestral diets in both centers have long benefitted from employing an astonishingly broad diversity of food plants. These benefits are no less relevant to Indigenous food sovereignty today that they were historically. Furthermore, most maize varieties, as well as many beans, and cucurbits are hitting their temperature thresholds and water deficit limits in most American landscapes (Altieri and Nicholls, 2009; Nabhan, 2013). Clearly, food production and diets in the “new climatic normal” will have to employ a set of food crops far more diverse and different than those employed in conventional agriculture at this moment in time (Nabhan, 2020).
Finally, we wish to emphasize that many of the initial efforts to revive the original structure and composition of ancestral Mesoamerican and Arid American diets have already emerged from Indigenous communities themselves (e.g., Kavena, 1980; Wolfe et al., 1985; Edaakie and Enote, 1999; Mihesuah, 2005; Tohono O'odham Community Action., 2010; Patchell and Edwards, 2013; Peña et al., 2017; Mihesuah and Hoover, 2019).
Well before many of these articles or books were published by authors of Indigenous heritage, their communities in Mexico and the U.S. were quietly reintegrating traditional foods long a part of Mesoamerican and Arid American diets into their contemporary feasts and health care strategies. These community-based efforts “go far beyond” the mere achievement of food security in terms of reliable access to affordable, nutritious food.
As noted in the 2007 Declaration of Nyéléni, a broad multi-cultural coalition wishes to place “the aspirations and needs of those who produce, distribute and consume food at the heart of food systems and policies rather than the demands of markets and corporations.” We wish to support that intent by promoting participatory research and on-ground technical assistance of the kind we have begun with CONACYT support to advance climate-friendly food, water and energy solutions in two Indigenous Comcaac communities in costal Sonora, Mexico.
In essence, the revival of Mesoamerican and Arid American agricultural and foraging traditions is not a top-down or exogeneous pressure toward “reverse-engineering” dietary change, but an Indigenous “grassroots community-based movement toward true food sovereignty (Patchell and Edwards 2013; Peña et al., 2017).” Our research only validates many of the tenets that underlie this Indigenous movement; it points to the nutritional benefits of often-forgotten wild and cultivated food plants. Conserving the wild plants, protecting the traditional crop landraces, and safeguarding the traditional ecological, gastronomic, and agricultural knowledge of Indigenous Nations in Arid America and Mesoamerica will be as critical as policy reforms to foster their food justice. As climate change differentially threatens Indigenous communities in many ways, advancing and safeguarding food sovereignty and the wild food plants that may contribute to it will be paramount.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author Contributions
GN, PC-GM, and DZ-V conceived together the article in Merida, Yucatan, Mexico and did multiple revisions. GN elaborated the first draft. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We wish to thank our many Indigenous collaborators on dietary studies over the last four decades, most recently Luis Eduardo Molina of the Comcaac community of Sonora, who assisted GN with establishing baselines for monitoring dietary changes and advancing food sovereignty through time in two Indigenous villages, and the families from Zapotitlán, Jalisco, who shared their deep knowledge of their ancestral foodways with PC-G and DZ-V. We also wish to acknowledge our collaborators from many cultures including A. Mellado, E. Barnett, M. L. Astorga, M. Estrella Astorga, D. Lewis, J. Ascencio, F. Kabotie, H. Dukepoo, L. Noriega, J. Martínez-Castillo, O. Vargas-Ponce, G. Carrillo-Galván, A. Flores-Silva, I. Torres-García, A. Casas, C.J. Figueredo, S. Rangel-Landa, A. Delgado, D. Cabrera-Toledo; X. Aguirre-Dugua, L. Eguiarte, R. Bye, E. Linares, D, A. Mellado, E. Barnett, R. Felger, C. Marlett, E. Ezcurra, E. Riordan, A. Búrquez-Montijo, L. Smith Monti, E. Riordan, B. Wilder, J. Mabry, C. Khoury, T. Crews, and J. Aronson. Barbara Kuhns graciously assisted with edits of the near-final draft. GN acknowledges long-term funding from the W.K. Kellogg Foundation, and initial funding many decades ago on three grants from the National Science Foundation. GN acknowledges the W.K. Kellogg Foundation Endowment for his position at the Southwest Center which covered page charges as well as travel to Arizona for PC-GM and DZ-V, and to Yucatán for GN.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2022.840619/full#supplementary-material
Footnotes
1. ^Mabry, J. (2022). Southwest Center, University of Arizona maize archaeologist, October 5 2022. (personal communication).
2. ^Ezcurra, E. (2021). University of California Riverside Department of Botany and Plant Sciences, Desert Ecologist, September 10, 2021. (personal communication).
References
Altieri, M. A., and Nicholls, C. I. (2009). Cambio climático y agricultura campesina: impactos y respuestas adaptativas. LEISA Revista de Agroecología 14, 5–8.
Appendini, K., and Liverman, D. (1994). Agricultural policy, climate change and food security in Mexico. Food Pol. 19, 149–164.
Aschmann, H. (1959). The Central Desert of Baja California: Demography and Ecology. Iberoamericana:42. Berkeley CA; Los Angeles CA: University of California Press.
Austin, D.F. (2006). Fox-tail millets (Setaria: Poaceae)—Abandoned food in two hemispheres. Econ. Bot. 60, 143–158. doi: 10.1663/0013-0001(2006)60<143:FMSPFI>2.0.CO;2
Avitia-García, E., and Castillo-González, A. M. (2002). Taxonomía y Nomenclatura de Especies Fruticolas. Chapingo: Universidad Autónoma de Chapingo.
Burns, B. T., Drees, M., Nabhan, G. P., and Nelson, S. C. (2000). “Crop diversity among Indigenous farming cultures in the tropical deciduous forest”, in The Tropical Deciduous Forest of Alamos: Biodiversity in a Threatened Ecosystem in México, edited by R. Robichaux. Tucson AZ, University of Arizona Press. p. 152–171.
Calvo, L., and Rueda-Esquibel, C. (2015). Decolonize Your Diet: Plant-Based Mexican American Recipes for Health and Healing. Vancouver, BC: Arsenal Pulp Press.
Casas, A., Caballero, J., and Valiente-Banuet, A. (1999). Use, management, and domestication of columnar cacti in south-central Mexico: a historical perspective. J. Ethnobiol. 19, 71–95.
Casas, A., Valiente-Banuet, A., and Caballero, J. (2002). “Evolutionary trends in columnar cacti under domestication in south-central México”, in Columnar Cacti and their Mutualists. Edited by T.J. Fleming TJ. and A. Valiente-Banuet. Tucson, AZ: University of Arizona Press. p. 137–164. doi: 10.2307/j.ctv23khmrw.12
Challinor, A. J., Wheeler, T. R., Craufurd, P. Q., and Slingo, J. M. (2005). Simulation of the impact of high temperature stress on annual crop yields. Agri. Forest Meterol. 135, 180–189. doi: 10.1016/j.agrformet.2005.11.015
Colunga-GarcíaMarín, P., Hernández-Xolocotzi, E., and Castillo-Morales, A. (1986). Variación morfológica, manejo agrícola y grados de domesticación de Opuntia spp. en el Bajío Guanajuatense. Agrociencia 65, 7–49.
Colunga-GarcíaMarín, P., and Zizumbo-Villarreal, D. (1993). “Evolución bajo agricultura tradicional y desarrollo sustentable,” in Cultura y manejo sustentable de los Recursos Naturales, eds E. Leff and J. Carabias (Mexico: Miguel Angel Porrúa), 123–164.
Colunga-GarcíaMarín, P., Zizumbo-Villarreal, D., and Martínez-Torres, J. (2007). “Tradiciones en el aprovechamiento de los agaves mexicanos: una aportación a la protección legal y conservación de su diversidad biológica y cultural”, in En lo Ancestral hay Futuro: del Tequila, los Mezcales y Otros Agaves, edited by P. Colunga-García Marín, Larqué Saavedra A, Eguiarte y, L., Zizumbo-Villarreal, D. Merida, YUC: CICY-CONACYT-CONABIO-INE. p. 229–248.
Contreras-Toledo, A. R., Cortés-Cruz, M. A., Costich, D., Rico-Arce, M. L., Brehm, J. M., and Maxted, N. (2018). A crop wild relative inventory for México. Crop Sci. 58:1292–1305. doi: 10.2135/cropsci2017.07.0452
Crimmins, A., Balus, J., Gamble, J. L., Beard, C. B., Bell, J. E., Dodgen, D., et al. (2016). The Impacts of Climate Change on Human Health in the United States: A Scientific Assessment. Washington, DC: U.S. Global Change Research Program. doi: 10.7930/J0R49NQX
De Alwis, D., and Limaye, V. (2021). The costs of inaction: The economic burden of fossil fuels and climate change on health in the U.S. Available online at: https://www.preventionweb.net/publications/view/77977
Ebi, K. L., and Hess, J. J. (2020). Health risks due to climate change: Inequity in causes and consequences. Health Aff. 39, 2056–2062. doi: 10.1377/hlthaff.2020.01125
Edaakie, R., and Enote, J. (1999). Idonapshe / Let's Eat: Traditional Zuni Foods. A:shiwi A:wan Museum and Heritage Center, Zuni, N.M.
Felger, R. S., Carnahan, S. D., and Sánchez-Escalante, J. J. (2017). Oasis at the Desert Edge: Flora of Cañon de Nacapule, Sonora México. Proceedings of the Desert Laboratory Contribution No. 1, Tucson AZ: University of Arizona.
Felger, R. S., and Moser, M. B. (1985). People of the Desert and Sea: Ethnobotany of the Seri Indians. Tucson AZ, University of Arizona Press. doi: 10.2307/j.ctv1n6pvq5
Fish, S. K., Fish, P., Miksicek, C., and Madsen, J. (1985). Prehistoric agave cultivation in Southern Arizona. Desert Plant. 7, 100–112.
Ford, R. I. (1981). Gardening and farming before 1000 A.D.: Patterns of prehistoric cultivation north of México. J. Ethnobiol. 1, 6–27.
Gentry, H. S. (1982). Agaves of Continental North America. Tucson AZ: University of Arizona Press. doi: 10.2307/j.ctv1t4m2h4
Griffith, M. P. (2004). The origins of an important cactus crop, Opuntia ficus-indica (Cactaceae): new molecular evidence. Am. J. Bota. 91, 1915–1921. doi: 10.3732/ajb.91.11.1915
Hernández-Sandoval, L. E., González-Romo, C. E., and González-Medrano, F. (1991). Plantas Utiles de Tamaulipas. Ann. Inst. Biol. Serie Botan. 62, 1–38.
Hernández-Xolocotzi, E. (1993). Aspects of plant domestication in México: A personal view. Pp.733-753 in Biological Diversity in México: Origins and Distribution, edited by Ramamoorthy TP, Bye R, Fa J. New York, NY: Oxford University Press. p. 733–753.
Hernández-Xolocotzi, E. (2013). “Zonas agricolas de México”, in Xolocotzia: Obras de Efraí m Hernández-Xolocotzi, Tomo I., Chapingo, México: Universidad Autónoma de Chapingo. p. 285–300.
Hodgson, W. (2012). “Pre-Colombian agaves: living plants linking an ancient past in Arizona”, in Explorations in Ethnobiology: The Legacy of Amadeo Rea, edited by Quinlan M, Lepofsky D. Denton, TX: Society for Ethnobiology. p. 78–103.
Hodgson, W., and Salywon, A. M. (2013). Two new Agave species (Agavaceae) from central Arizona and their putative pre-Columbian domesticated origins. Brittonia. 65, 5–15. doi: 10.1007/s12228-012-9255-z
Hodgson, W. C., Salywon, A. M., and Doelle, W. H. (2018). Hohokam lost crop found: A new Agave (Agavaceae) species only known from large-scale pre-Columbian agricultural fields in Southern Arizona. System. Bota. 43, 734–740. doi: 10.1600/036364418X697445
Kirchhoff, P. (1954). Gatherers and farmers in the greater southwest: A problem in classification. Am. Anthropol. 56, 529–550. Available online at: http://www.jstor.org/stable/664319
Kirchhoff, P. (2000). “Mesoamérica,” in Dimensión Antropológica, Vol. 19. p. 15–32. Available online at: http://www.dimensionantropologica.inah.gob.mx/?p=1031
Kuhnlein, H. V., and Receveur, O. (1996). Dietary change and traditional food systems of Indigenous peoples. Ann. Rev. Nutr. 16, 417–442.
Kumar, M. (2016). Centers of diversity of crops. Biotech Articles 4, 43–59. Available online at: https://www.biotecharticles.com
Latorre, D. L., and Latorre, F. A. (1977). Plants used by the Mexican Kickapoo Indians. Econ. Bot. 31, 340–357. doi: 10.1007/BF02866886
Leach, J. D., and Sobolik, K. D. (2010). High dietary intake of prebiotic inulin-type fructans in the prehistoric Chihuahuan Desert. Br. J. Nutr. 103, 1558–1561. doi: 10.1017/S0007114510000966
Li, X., Braga-Junqueira, A., and Reyes-Garcia, V. (2021). At the crossroad of emergency: ethnobiology, climate change, and indigenous peoples and local communities. J. Ethnobiol. 41, 307–312. doi: 10.2993/0278-0771-41.3.307
Louderback, L., and Pavlik, B. (2017). Starch granule evidence for the earliest potato use in North America. Proc. Nat. Acad. Sci. 114, 7606–7610. doi: 10.1073/pnas.1705540114
Louderback, L. A., and Pavlik, B. M. (2018). Integrating modern vegetation and ethnographic data to understand dietary choices in the past. Hum. Ecol. 46, 1–12. doi: 10.1007/s10745-018-0040-9
Luo, Q. (2011). Temperature thresholds and crop production: a review. Clim. Chang. 109, 583–598. doi: 10.1007/s10584-011-0028-6
Luque-Agraz, D., Martínez-Yrizar, A., Búrquez, A., López-Cruz, G., and Murphy, A. D. (2016). Complejos Bioculturales de Sonora: Pueblos y Territorios Indígenas. Hermosillo, SON: CIAD.
Mapes, C., and Basurto, F. (2016). “Biodiversity and edible plants in México”, in Ethnobotany of México: Interactions of People and Plants in México. Edited by Lira R, Casas A, and J. Blancas. New York NY: Springer.
Martinech, J. (2018). Multi-Model Framework for Quantitative Sectoral Impacts Analysis A Technical Report for the Fourth National Climate Assessment. Washington, DC: U.S. Environmental Protection Agency.
Martínez, M. (1979). Catálogo de Nombres Vulgares y Científicos de Plantas Mexicanas. México, DF: Fondo de Cultura Económica.
Merrill, W. L., Hard, R. J., Mabry, J. B., Fritz, G. J., Adams, K. R., Roney, J. R., et al. (2009). The diffusion of maize to the southwestern United States and its impact. Proc. Nat. Acad. Sci. 106, 21019–20126. doi: 10.1073/pnas.0906075106
Mihesuah, D. (2005). Recovering Our Ancestors' Gardens: Indigenous Recipes and Guide to Diet and Fitness. Lincoln, NE: University of Nebraska Press.
Mihesuah, D., and Hoover, E. (2019). Indigenous Food Sovereignty in the United States: Restoring Cultural Knowledge, Protecting Environments and Regainibng Health. Norman, OK: University of Oklahoma Press.
Minnis, P. E. (2021). Famine Foods: Plants We Eat to Survive. Tucson AZ: University of Arizona Press. doi: 10.2307/j.ctv1k13b98
Nabhan, G., and de Wet, J. M. J. (1984). Panicum sonorum in Sonoran Desert agriculture. Econ. Bot. 38, 65–82. doi: 10.1007/BF02904417
Nabhan, G. P. (1985). Native crop diversity in Aridoamerica: Conservation of regional gene pools. Econ. Bot. 39, 387–399. doi: 10.1007/BF02858746
Nabhan, G. P. (2013). Our coming food crisis. New York Times July 21. Available online at: https://www.nytimes.com/2013/07/22/opinion/our-coming-food-crisis.html (accessed December 2, 2021).
Nabhan, G. P. (2020). Crops from U.S. food supply chains will never look nor taste the same again. Agric. Human Value. 37, 651–652. doi: 10.1007/s10460-020-10109-6
Nabhan, G. P., Colunga-García-Marín, P., and Zizumbo-Villarreal, D. (in press). Reviving Arid American and Mesoamerican diets in the face of climate change: assessing their composition and potential to advance Indigenous health and diabetes prevention. Economic Botany.
Nabhan, G. P., and Molina, L. S. (2021). Unpublished survey of fresh food plant species in grocery stores in Punta Chueca, Desemboque del Sur and Kino Viejo, Sonora accessible to the Comcaac communities. Archived at University of Arizona Desert Laboratory at Tumamoc Hill, December 2021.
Nabhan, G. P., Olmedo, J., and Pailes, M. (2019). The Huachuca agave as mescal de la mancha”: translocation without domestication of Agave parryi var. huachucensis in the Sky Islands of the U.S. and México. Desert Plant. 35, 25–42.
Nabhan, G. P., Riordan, E. C., Aronson, J., Barron-Gafford, G., Búrquez, A., Crews, T. E., et al. (2020). A new Aridamerican food system for a hotter, water-scarce world. Plants, People, Planet. 2, 627–639. doi: 10.1002/ppp3.10129
Nuchatelli, D. (2019). Climate change could cost U.S. economy billions. Yale Climate Connections. https://yaleclimateconnections.org/2019/04/climate-change-could-cost-u-seconomy-billions/ (accessed December 5, 2021).
Ojeda-Linares, C. I., Alvarez, G., Figueredo-Urbina, C. J., Torres-García, I., Islas, L. A., Vallejo, M., et al. (2021). Traditional fermented beverages: a biocultural unseen foodscape. Foods. 10, 2390. doi: 10.3390/foods10102390
Patchell, B., and Edwards, K. (2013). The role of traditional foods in diabetes prevention and management among Native Americans. Curr. Nutr. Rep. 3, 340–344. doi: 10.1007/s13668-014-0102-6
Peña, D., Calvo, L., McFarland, P., and Valle, G. R. (2017). Mexican-Origin Foods, Foodwats and Social Movements: decolonizing Perspective. Fayetteville, AR: University of Arkansas Press. doi: 10.2307/j.ctt1t89jww
Pontifes, P. A., García-Meneses, P. M., Gómez-Aíza, L., Monterroso-Rivas, A. I., and Caso-Chávez, M. (2018). Land use/land cover change and extreme climatic events in the arid and semi-arid ecoregions of Mexico. Atmósfera. 31, 355–372. doi: 10.20937/ATM.2018.31.04.04
Rea, A. M. (1997). At the Desert's Green Edge: An Ethnobotany of the Gila River Pima. Tuscon, AZ: University of Arizona Press.
Riordan, E. C., and Nabhan, G. P. (2019). Trans situ conservation of crop wild relatives. Crop Sci. 59, 2387–2403. doi: 10.2135/cropsci2019.06.0356
Rodríguez, A., and Spooner, D. (1997). Chloroplast DNA analysis of Solanum bulbocastanum and S. cardiophyllum, and evidence for the distinctiveness of S. cardiophyllum Subsp. ehrenbergii (Sect. Petota). System. Bota. 22, 31–43. doi: 10.2307/2419676
Rodríguez-De Luque, J. J., González-Rodríguez, C. E., Gourdji, S., Mason-D'Croz, D., Obando-Bonilla, D., Mesa-Diez, J., et al. (2016). Impactos socioeconómicos del cambio climático en América Latina y el Caribe: 2020–2045. Cuadernos de Desarrollo Rural. 13, 11–34. doi: 10.11144/Javeriana.cdr13-78
Segura-Aguilar, R., Shilón, J. A. H., Higuera, A. G. H., and Hernández, D. L. (2020). Estudio de Elementos Claves Mulitifactorales que Detonen el Desarollo Economico y Productivo del Sector No-Maderable. Productores Sustentables Sierra La Giganta A. C Organización Social del Sector Forestal, Baja California Sur. Available online at: https://www.conafor.gob.mx/EstudiosRegionalesForestales/EstudioRegionalForestal_UMAFOR0302_PSSG.pdf
Solano-Picazo, C. (2018). Etnobiología de Wirikuta: uso de recursos vegetales silvestres en el desierto de San Luis Potosí, México. Etnobiologia. 16, 54–77. Available online at: https://revistaetnobiologia.mx/index.php/etno/article/view/121
Stiller, A., Garrison, K., Gurdyumov, K., Kenner, J., Yasmin, F., Yates, P., et al. (2021). From fighting critters to saving lives: polyphenols in plant defense and human health. Int. J. Mole. Sci. 22, 8995.
Vásquez-García, J. A. (1995). Flora de Manantlán: Plantas Vasculares de la Reserva de la Biosfera Sierra de Manantlán Jalisco-Colima, México. Fort Worth, TX: Botanical Research Institute of Texas.
Vint, J. M. (2015). “Las Capas, AZ AA:12:111 (ASM), Introduction: background, chronology, and research orientation,” in Implements of Change: Tools, Subsistence, and the Built Environment of Las Capas, an Early Agricultural Community in Southern Arizona, ed J. M. Vint (Tuscon, AZ: Archaeology Southwest), 1–31.
Vint, J. M. (2018). “The southwest archaic in the tucson basin,” in The Archaic Southwest: Foragers in an Arid Land, ed B. Vierra (Salt Lake City, UT: University of Utah Press), 66–97.
Wilken-Robertson, M. (2018). Kumeyaay Ethnobotany: Shared Heritage of the Californias. San Diego, CA: Sunbelt Publications.
Wolfe, W. S., Weber, C. W., and Arviso, K. D. (1985). Use and nutrient composition of traditional Navajo foods. Ecol. Food Nutr. 17, 323–344. doi: 10.1080/03670244.1985.9990906
Yensen, N. P. (2008). “Halophyte uses for the twenty-first century,” in Ecophysiology of High Salinity Tolerant Plants. Tasks for Vegetation Science, Vol 40, eds M. A. Khan and D. J. Weber (Dordrecht: Springer).
Zizumbo-Villarreal, D., Colunga GarcíaMarín, P., and Flores-Silva, A. (2016). “Pre-Columbian food system in West Mesoamerica,” in Ethnobotany of México: Interactions of People and Plants in México, eds R. Lira, A. Casas, and J. Blancas (New York, NY: Springer), 67–82.
Zizumbo-Villarreal, D., Flores-Silva, A., and Colunga-GarcíaMarín, P. (2012). The archaic diet in Mesoamerica: incentive for milpa development and species domestication. Econom. Bot. 66, 328–343.
Zizumbo-Villarreal, D., Flores-Silva, A., and Colunga-GarcíaMarín, P. (2014). The food system during the formative period in West Mesoamerica. Econom. Bot. 68, 67–84.
Keywords: Arid America, climate change, desert agriculture, diabetes, Indigenous food sovereignty, Mesoamerica, traditional food systems, centers of biocultural diversity
Citation: Nabhan GP, Colunga-GarcíaMarín P and Zizumbo-Villarreal D (2022) Comparing Wild and Cultivated Food Plant Richness Between the Arid American and the Mesoamerican Centers of Diversity, as Means to Advance Indigenous Food Sovereignty in the Face of Climate Change. Front. Sustain. Food Syst. 6:840619. doi: 10.3389/fsufs.2022.840619
Received: 21 December 2021; Accepted: 29 April 2022;
Published: 20 June 2022.
Edited by:
Zakaria Kehel, International Center for Agricultural Research in the Dry Areas (ICARDA), MoroccoReviewed by:
Peter J. Jacques, University of Central Florida, United StatesDaniel Pinero, National Autonomous University of Mexico, Mexico
Copyright © 2022 Nabhan, Colunga-GarcíaMarín and Zizumbo-Villarreal. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gary Paul Nabhan, Z3BuYWJoYW5AYXJpem9uYS5lZHU=