 Denise Jäckel
Denise Jäckel Kim G. Mortega1
Kim G. Mortega1 Gerlind U. C. Lehmann
Gerlind U. C. Lehmann Silke L. Voigt-Heucke
Silke L. Voigt-Heucke- 1Museum für Naturkunde Berlin, Leibniz Institute for Evolution and Biodiversity Science, Berlin, Germany
- 2Faculty of Life Sciences, Humboldt-Universität zu Berlin, Berlin, Germany
- 3Evolutionary Ecology, Department of Biology, Humboldt-Universität zu Berlin, Berlin, Germany
- 4Animal Behavior, Institute of Biology, Freie Universität Berlin, Berlin, Germany
Open science approaches enable and facilitate the investigation of many scientific questions in bioacoustics, such as studies on the temporal and spatial evolution of song, as in vocal dialects. In contrast to previous dialect studies, which mostly focused on songbird species with a small repertoire, here we studied the common nightingale (Luscinia megarhynchos), a bird species with a complex and large repertoire. To study dialects on the population level in this species, we used recordings from four datasets: an open museum archive, a citizen science platform, a citizen science project, and shared recordings from academic researchers. We conducted the to date largest temporal and geographic dialect study of birdsong including recordings from 1930 to 2019 and from 13 European countries, with a geographical coverage of 2,652 km of linear distance. To examine temporal stability and spatial dialects, a catalog of 1,868 song types of common nightingales was created. Instead of dialects, we found a high degree of stability over time and space in both, the sub-categories of song and in the occurrence of song types. For example, the second most common song type in our datasets occurred over nine decades and across Europe. In our case study, open and citizen science data proved to be equivalent, and in some cases even better, than data shared by an academic research group. Based on our results, we conclude that the combination of diverse and open datasets was particularly useful to study the evolution of song in a bird species with a large repertoire.
Introduction
Open science practices such as open data (OD), citizen science (CS), and data sharing may open new avenues for answering novel kinds of research questions. Open data are databases and platforms that are freely available to the public and can be reused without copyright restrictions (Kassen, 2013). Acoustic datasets, for example, are increasingly shared publicly both on museum archives, e.g., the animal sound archive at the Museum für Naturkunde Berlin (MfN)1, and in open platforms, e.g., Xeno-Canto2. Biologists are increasingly using these large open datasets of observations to answer a broad range of questions on evolution and ecology across time and space (Aplin et al., 2021; Dominguez et al., 2021). CS as a concept describes research conducted by non-professional scientists who are not institutionally employed in this field of science (as defined in the Greenbook Citizen Science; Bonn et al., 2017). The use of CS data is rapidly growing, especially in ornithology, and has been successfully used, e.g., for bird monitoring (de Camargo Barbosa et al., 2021), geographical distribution (de Jesus et al., 2021), and population trends (Neate-Clegg et al., 2020). Nevertheless, usage of CS data is not yet well established in ornithology (Weisshaupt et al., 2021) due to a lack of information and thus quality concerns about data availability, data bias, and data generation (Hochachka et al., 2012; Lukyanenko et al., 2016). This quality concern is why CS data are often not made publicly available in the sense of open science. The concerns can be overcome when different datasets are connected, in particular when CS data are used as a complement to fill in gaps (Kirchhoff et al., 2021) or when they are directly compared with data gathered by academic researchers. In an earlier study, we were able to show that citizen scientists made a surprising number of recordings of valid quality when compared to expert recordings and that differences between the recordings were mainly due to the technical quality of the equipment used. Recordings were valid to distinguish songs visually and conduct duration measurements. Frequency measurements showed significant differences depending on the cost, recentness, and brand of the smartphone. Thus, quality is multidimensional and always depends on the research question. In the current study, we conducted a quality dataset comparison based on the same parameters to ensure that open and CS data are considered as sufficiently valuable and are used more often in ornithology. To aid in establishing such a representative case study, we investigated temporal and geographical song variations, so-called dialects, in a bird species.
Studies of dialects have historically been based mostly on academic-collected data (in our case shared data) and began as early as (Poulsen, 1958), when Poulsen studied evolutionary processes leading to the formation of dialects, the shape and extent of spatial variation within a species, and the coding of dialects. Dialects have been found with academic-collected data in cetaceans (Sharpe et al., 2019), hummingbirds (Gaunt et al., 1994), parrots (Wright and Dahlin, 2018), suboscine passerines (Kroodsma et al., 2013), bats (Prat et al., 2017) and with CS data in songbirds (oscine, Searfoss et al., 2020). Song in birds is a sexually selected and culturally transmitted trait that plays a critical role, particularly in breeding ecology as dialects can restrict gene flow among conspecific populations (Baker and Mewaldt, 1978) or contribute to reproductive isolation and speciation resulting in ring species (Irwin et al., 2001). In the well-studied, mostly temperate model species, males sing to defend their territory and attract females (Catchpole and Slater, 2008). Songbirds acquire their song mainly through social learning via cultural imprinting. In this process, young males learn their song from their parents or neighbors and pass those songs onto the next generation (Mennill et al., 2018). This process can lead to copying errors and modifications over time and space, resulting in a cultural evolution of song (Lynch, 1996). Sexual selection, cultural drift, environment, and population adaptation (immigration, emigration) influence whether song types are particularly common or rare in individual songs and populations (Derryberry et al., 2011). A song type is a unique sequence of elements with defined frequencies and durations, as described by Gil and Gahr (2002). If song types are beneficial to individuals in terms of environmental adaptation, female choice – or their own fitness – some songs types persist over time (Lynch, 1996). Unbeneficial song types can be loosed or modified, which lead to geographic variation that may evolve into distinct, local dialects over time (Beecher and Brenowitz, 2005). Local dialects reduce song variation at the microgeographic level compared to the macrogeographic level, resulting in greater differences between populations than within populations (Catchpole and Slater, 2008). Investigating dialects over time and space allows insight into the processes of song evolution (Rendell and Whitehead, 2005) and improves the understanding of songs as well as repertoires and thus vocal learning (Pozzi et al., 2010). Previous dialect studies have increasingly benefited from the use of open databases (Wright and Dahlin, 2018) and CS recordings (Diblíková et al., 2019). Due to the internet and GPS-based app development (Dickinson et al., 2012), these studies generated considerable data on a spatial and temporal scale that would be unachievable for traditional scientific studies due to financial or logistical constraints (Diblíková et al., 2019). These studies showed that songs and parts of them remain stable for up to three decades (Nelson et al., 2004) or change geographically especially in bird species with small repertoires, and typically with one to three song types (Diblíková et al., 2019; Searfoss et al., 2020). Nevertheless, it is not yet known how the song or parts of it evolve over a century or Europe-wide across the entire breeding range and in a bird species with a large repertoire.
In our study, we examined the common nightingale (Luscinia megarhynchos), a species with approximately 190 song types (Hultsch and Todt, 1982) and a Europe-wide breeding range (from the Mediterranean to Central Asia in the East and Central Europe in the North; Glutz von Blotzheim, 1988). The nightingale is particularly interesting for studies on song dialects due to its complex and well-studied song behavior in the field (e.g., Kiefer et al., 2006) and in the laboratory (e.g., Hultsch and Todt, 1996). Males are territorial, return to their natal breeding area, and have been found to also occupy the same territory in consecutive years (Roth and Amrhein, 2010). Songs can be classified into four song categories (whistle, buzz, trill, and so-called “other songs,” Figure 1) as well as song types which form the song repertoire (doctoral thesis Hultsch, 1980; unpublished data). Nightingale males, as so-called open-ended learners, are among the bird species in which 1-year-old males can significantly expand and adjust their repertoire for the following breeding season, but not thereafter (Kipper et al., 2004). In this process, song types are learned in a very stereotypical way (Hultsch et al., 1999), which has the capacity to make the song repertoire very diverse with many individual song type variations (Landgraf et al., 2015). These different song types are not repeated directly, but rather after a variety of other song types, with some shared commonly and others rarely within and between individuals (Hultsch and Todt, 1989). Earlier studies have already shown that neighboring nightingales in intermediate distances share a higher proportion of songs than close or distant ones (Hultsch and Todt, 1981) due to song type adjustment to the breeding population (Kiefer et al., 2010, 2014). Based on knowledge of song ontogeny and migration behavior, temporally stable and geographically variable nightingale song can be expected and would be consistent with previous studies. Weiss (2012; unpublished data) noted in his doctoral thesis that dialects are most likely coded in the number of times a song type appears (relative frequency). Schehka (2004) found in her master thesis that common song types have been proven to be stable in their occurrence over two decades and in two countries, while she did not study rare song types. Thus, several lines of evidence suggest that stability and dialects may be occurring on a larger scale, although their extent has not been fully investigated yet. We addressed this question in our study by examining the stability (occurrence of common and rare song types), and dialects (relative frequency) on an expanded temporal (almost a century) and geographical (whole Europe) scale. Because certain parts of songs (thereafter defined as sub-categories of song) showed stability in some and coded for dialects in other species with a small repertoire, we investigated their function in the large repertoire of the nightingale. OD and CS data approaches have been used successfully for dialect research in other species with few song types (Wright and Dahlin, 2018; Diblíková et al., 2019), but never before in a songbird with so many song types, such as the nightingale. Therefore, we examined whether the datasets showed different results for the sub-categories of song, the occurrence or relative frequency of common song types to assess their comparability and derive their respective usefulness.
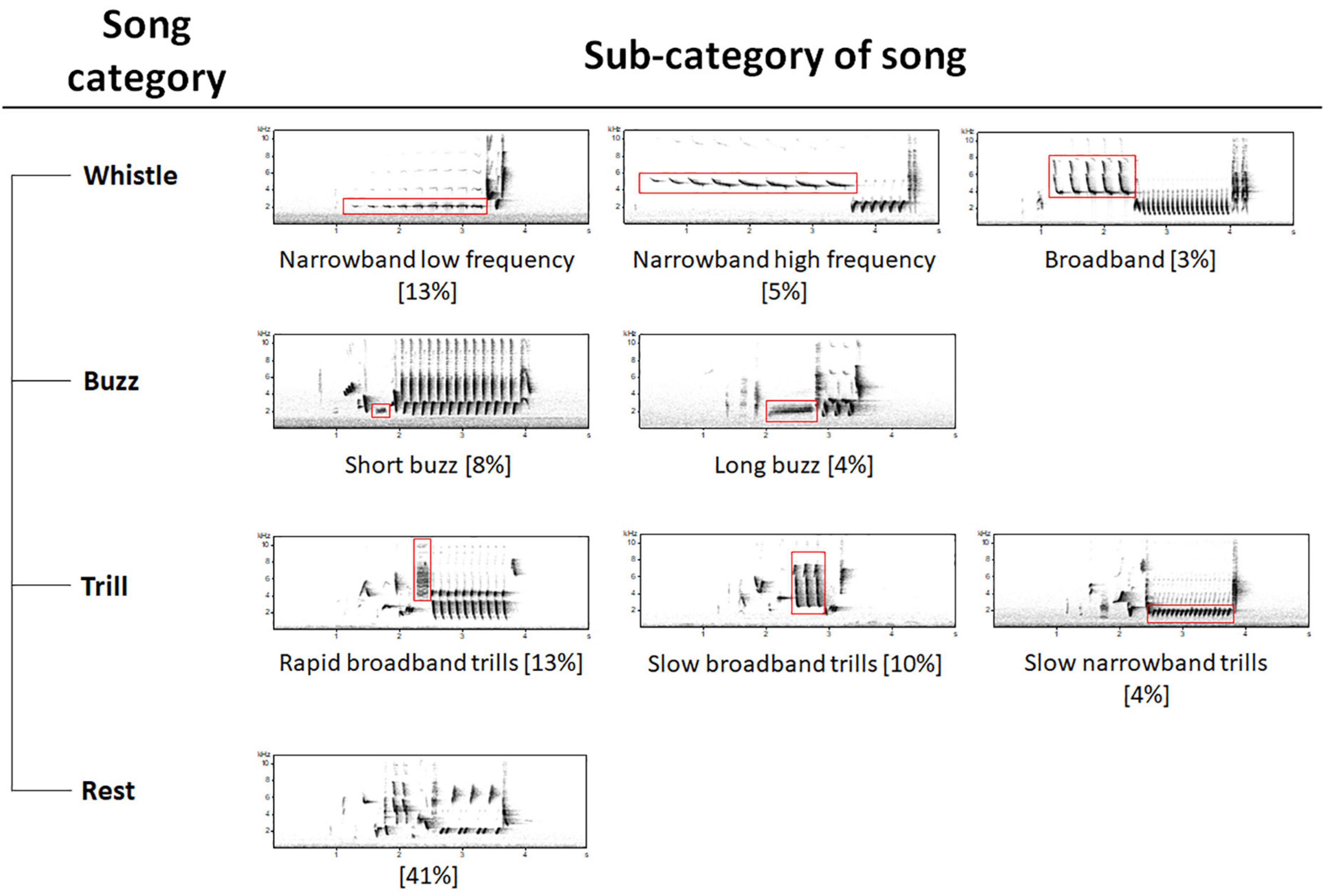
Figure 1. Decision tree for song types classification. Subdivision of whistles: narrowband (<3 kHz frequency bandwidth) low and high frequency and broadband (>3 kHz frequency bandwidth). Subdivision of buzz songs: short (<500 ms) and long (<500 ms) sections. Subdivision of trills: rapid broadband trills, slow broadband trills, and slow narrowband trills. Percentages are given in square brackets.
To investigate the potential for new avenues for research, we specifically addressed two biological and one technical hypothesis: (1) Common song types occur in a stable manner over nearly a century in the nightingale song (H1: stability). Since stable common song types have previously been found over two decades in the nightingale, we predicted stability over several decades, as has been detected in other species. (2) The relative frequency of common song types is encoded for dialects in nightingales across Europe (H2: dialects). As the relative song type frequency has been shown to differ in the nightingale between two neighboring countries, we predicted that such variance occurs also across the entire breeding range of the nightingale. (3) The sub-categories of song, occurrence, and relative frequency do not differ between the datasets (H3: datasets). Since OD, CS, and shared data have been used successfully for dialect research in other species and CS recordings were determined to be as valid as scientific recordings in an earlier comparison, we expected that all datasets used will be appropriate for nightingale dialect research.
Materials and Methods
The Nightingale Song as a Model in Song Research
The nightingale is a recognized model species in song research due to its clearly structured song with durations of about 2–4 s alternating with silent intervals (pauses) of similar length (Hultsch, 1980; unpublished data). The basal unit is an element with a characteristic frequency (bandwidth) and duration, containing pauses less than 10 ms. Up to four elements are grouped together to form syllables which are separated by pauses of 10 ms and produced once or several times repeated in a stereotypic order, form phrases. These are grouped into four sections (α, β, γ, and Ω) which can be characterized by analyzing element features and repetition rate (Hultsch, 1980; unpublished data; Figure 1). Male nightingales can produce a variable number of distinct song types whereas different individual birds can sing the same song type with minor variations (between them, or from rendition to rendition), resulting in extraordinarily large song repertoires (on average 190 with a maximum of 250 different song types; e.g., Hultsch and Todt, 1982; Kipper et al., 2004). As it is therefore challenging to study the song and with regard to the work economy, we focused on 20 song types (approx. 10% of the song types of a nightingale repertoire, Supplementary Figure 1). For this species, it is a standard procedure (e.g., Kipper et al., 2015) and has also been used in a previous dialect study of the nightingale due to the complex song (Schehka, 2004; unpublished data). We examined the ten most common and the ten rarest song types (occurred only once) from Berlin-Brandenburg within all datasets and across years. As there were 276 rare song types we selected ten of them randomly. Using these 20 song types (ten common, ten rare ones with Berlin-Brandenburg as a reference point) we analyzed their geographical occurrence across Europe. The ten most common song types were used for the temporal and dataset occurrence analysis and the relative frequency estimation for time, space and datasets.
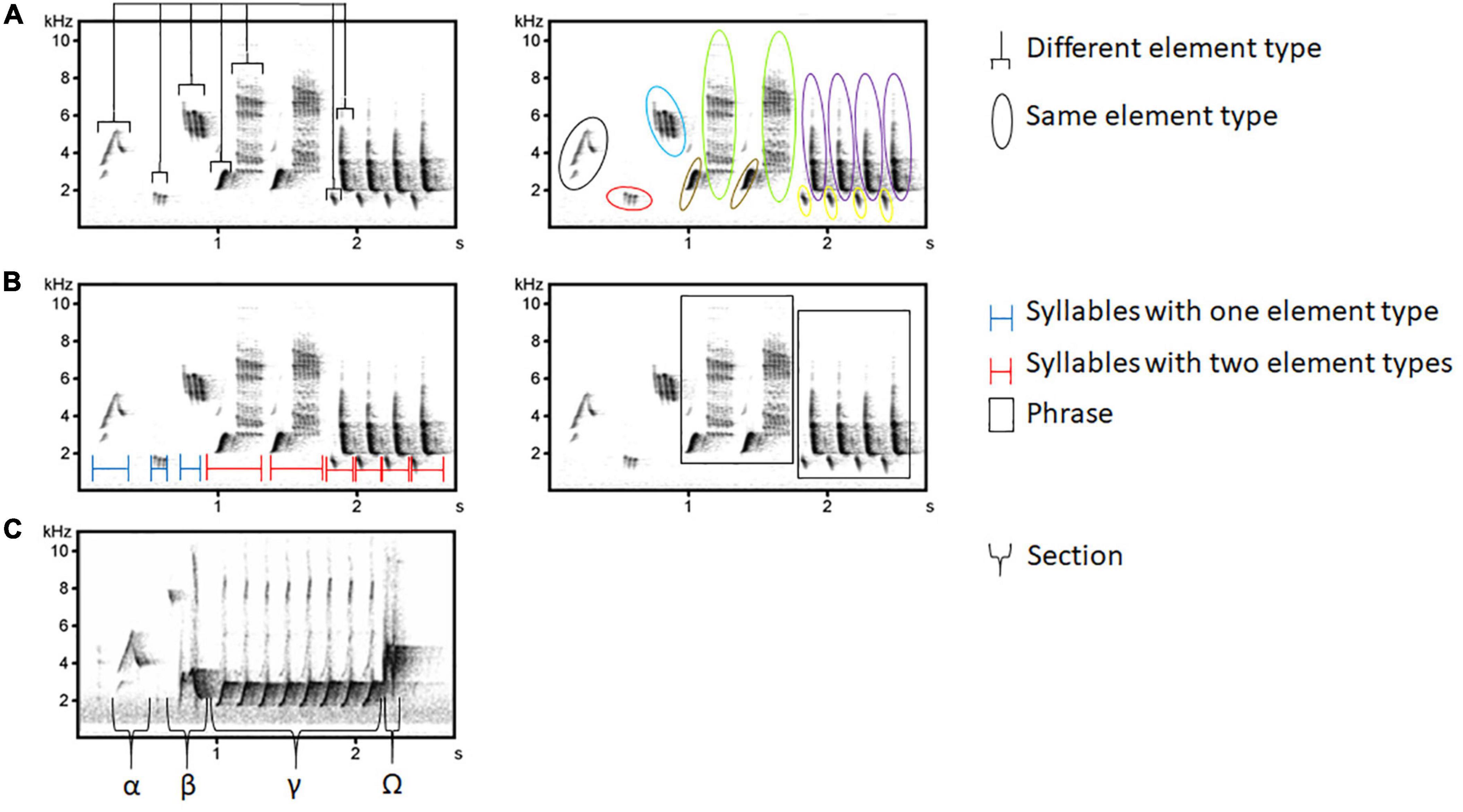
Figure 2. Basic units of a nightingale song. Elements (A) form syllables (B), which are joined together to form phrases. Several phrases form sections (C): α (quiet and short), β (louder, with melodic structure), γ (rapidly repeated syllables), and Ω (hardly any repetition, can be missing).
Databases
To unravel the potential of OD, CS, and data sharing, we used and compared four datasets (Figure 3): one open archive, two CS datasets, and one shared dataset. In total, we analyzed 14,140 nightingale songs. Since the length as well as the recording quality varies greatly, songs are a better measure to indicate the amount of data material than recording numbers. No recording originated from more than one dataset. Recording standards varied, depending on the species, research question, and context. For studies on individual song differences in the nightingale, 1-h nocturnal recordings are the standard (Kiefer et al., 2014), as they contain the most songs as well as no disturbing background noise in terms of other singing birds (bachelor thesis Jäckel, 2013; unpublished data). Only the shared recordings were standardized. The other datasets were unstandardized in terms of technique (smartphones to professional equipment), time (day and night), location (Europe-wide), and durations (few seconds to 90 min). However, standardized recordings were not required for our study, as we were working on the population rather than the individual level. Individual differences are used to define signature and not geographical variations. We cannot completely rule out the possibility that certain individuals were recorded repeatedly and therefore their songs occur multiple times in our dataset, in particular this is the case for the citizen science data. However, based on the GPS data we assume that the vast majority of recordings were not from a few individuals, but that the recordings were well distributed in space and time. To emphasize that we are not making statements about individuals, we worked at the population level.
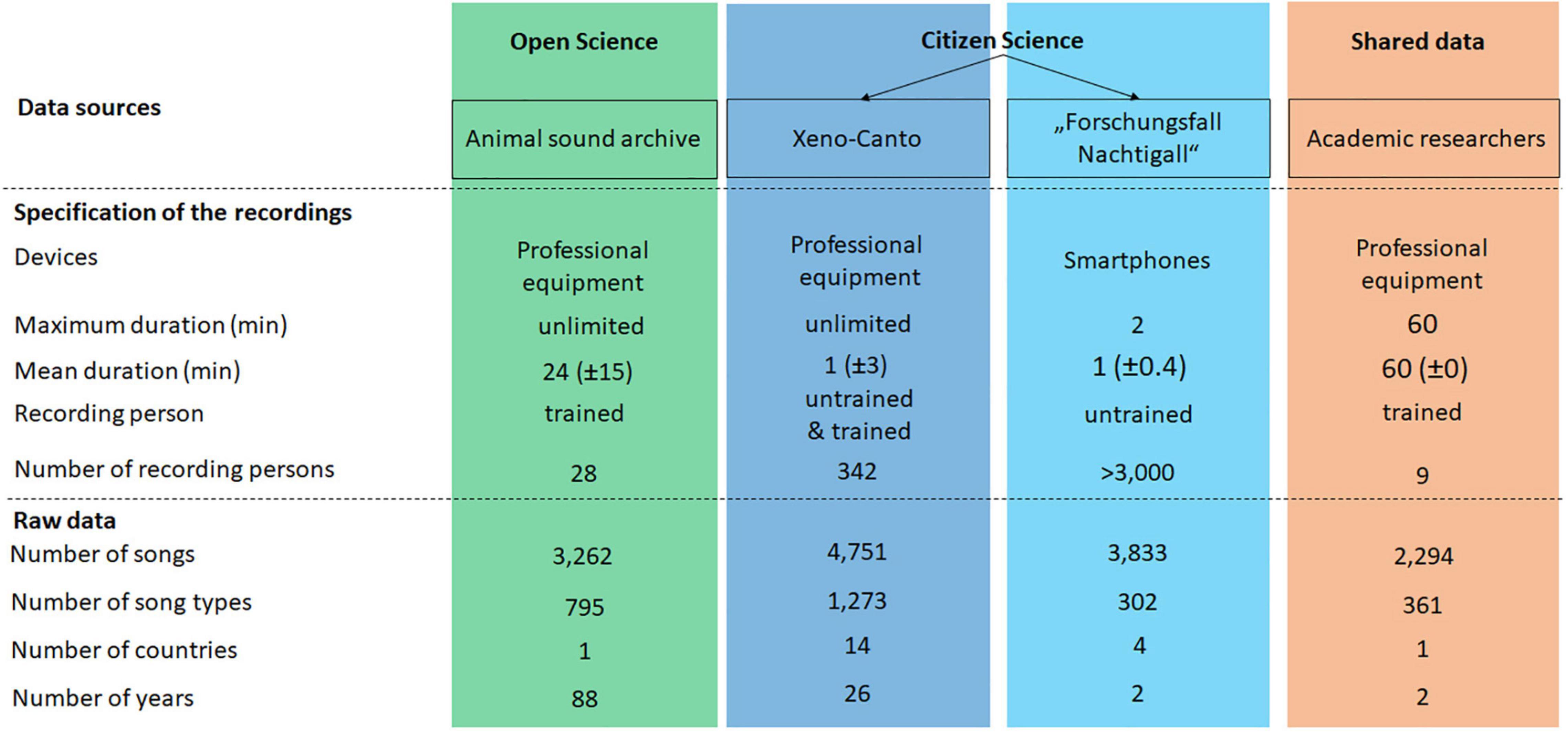
Figure 3. Overview of the data material for open science [animal sound archive (ASA)], citizen science [Xeno-Canto and “Forschungsfall Nachtigall” (FFN)] and shared data [academic researcher (AR)]. Information was separated into specification of the recordings and raw data.
We used the open animal sound archive of the MfN (hereinafter abbreviated as ASA) which contained recordings from the 1930s until 2015 of which we examined 3,262 songs from Germany. These recordings were generated by different experts with different recording devices (sampling frequency: 96 kHz; resolution: 16 bit). The data were distributed differently over the decades, with a significant decrease from 2006 onward. We analyzed 3,833 songs from a nightingale citizen science project (project name: “Forschungsfall Nachtigall,” hereinafter abbreviated as FFN; eng. “nightingale citizen science project”), led by a team of academic researchers at the MfN. These recordings were generated with diverse smartphones (sampling frequency: 44.1 kHz; resolution: 16 bit) within 2018 and 2019 mainly in Berlin. Additionally, we examined 4,751 songs from the CS website Xeno-Canto of 13 countries and spanning over 26 years. These recordings were generated by both laypeople and experts, using a wide range of technology (sampling frequency: 8–48 kHz; resolution: 16 bit). The shared dataset of 2,294 songs from Berlin was generated by academic researchers (hereinafter abbreviated as AR) in 2018 and 2019 by students of Freie Universität Berlin (Berlin, Germany) and staff of the FFN. Recordings were generated with Sennheiser ME66/K6 directional microphones (Georgsmarienhütte, Germany) connected to a Tascam Dr-40 4-Track Portable Digital Recorder or a Marantz solid-state recorder PMD660 as well as Zoom H2n recorders (sampling frequency: 44.1 kHz; resolution: 16 bit).
A European Nightingale Song Type Catalog
The high precision of song learning, copying, and delivering in male nightingales within and between individuals allows for a reliable assignment of a song to the same song type (Hultsch et al., 1999). We used Avisoft SASLab Pro 5.2 (R. Specht, Berlin, Germany) for visual inspection of songs (sampling rate = 22.05 kHz, FFT = 1024 points, Hamming-Window, overlap 93.75%). As Avisoft can only open WAV files, the MP3 and m4a recordings were converted into WAV files by using the program WaveLab 7. We started our cataloging process with recordings from Berlin-Brandenburg and extended across Germany and Europe. During the last 2 years, a single person (DJ) visually compared songs using spectrograms with catalog song types and categorized a song as either known or new. We did a self-check at the beginning to evaluate the accuracy of the cataloged song types. In this cataloging process, comparable to the nightingale song type catalog of Hultsch and Todt (1981) and Kipper et al. (2004), we based the definition of a song type on an identical γ-section. But our analysis differed from their previous catalog: (a) As we worked with different types of recordings (unknown individuals), we defined variations of a song type as all differences in the elements in the α, β, and in the Ω-section (Figure 2). Variations in the previous catalog occurred when more than three of ten elements (80% of the song type) differed in the α or β section. Variations are thus minor modifications of a song type that do not occur in the repetitive γ-section. We therefore based the structure within our song type catalog on the γ-section since any differences in that section were used to define a new song type. (b) We added all newly found song types to the catalog. For the previous catalog, which was based on 1-h song recordings of individual known nightingales, it was necessary that song types had to be sung at least twice by the same male in the same recording to be included. (c) We created our catalog on a basis of recordings across European datasets without any knowledge of the individual. Doing so, we created a European song type catalog on the population level.
Sub-Categories of Song Classification
We established previously that recordings made with qualitatively and technically different recording devices presented significantly different acoustic parameters in subsequent measurements. Therefore, we based the classification of sub-categories of song on visually distinguishable parameters, i.e., frequency ranges, durations, pauses, and element rates – and not on measuring frequencies. Sub-categories of song were extended divisions of the known four song categories of the nightingale: whistles, buzzes, trills, and “other songs” (Figure 1; Hultsch, 1980; unpublished data). “Other songs” were not further subdivided. Whistle, buzz and trill elements occur in the γ-section. Whistle elements never occur with buzz elements in the same song type simultaneously. It is possible that whistle or buzz elements are followed by trill elements in different sections, whereas these are no trill songs.
Whistles have been shown to have frequency-dependent functions (Hultsch, 1980; unpublished data) and are used for female attraction (Naguib et al., 2002). On this basis, we classified whistles according to the frequency bandwidth of the last whistle element into narrowband (<3 kHz) and broadband (>3 kHz) ones. We divided the narrowband ones corresponding to the frequency of the last whistle element into low (<4 kHz) and high (>4 kHz) whistles. Buzz songs have been found to indicate the quality of a male by the length of the buzz section (Weiss et al., 2012). We subdivided the buzz songs into ones with short (<500 ms) and long (>500 ms) sections. The relationship between frequency bandwidth and trill rate has been demonstrated to serve as an aggressive signal in nightingales (Kunc et al., 2006). In some bird species, this relationship has been found to indicate the quality of a male to females (Podos, 1996). Based on this finding, we divided the trills into rapid broadband (bandwidth of >9 kHz and an element repetition rate without visible pauses between the trills), slow broadband (bandwidth of >9 kHz and element repetition rate with visible pauses between the trills), and slow narrowband trills (bandwidth <6 kHz and element repetition rate with visible pauses between the trills).
Comparison of Time Periods
We specified four time periods (TP) for the temporal analysis. TP1 was defined by us as the 1930s, followed by TP2 (1960–1980), TP3 (1981–2000), and TP4 (2001–2019). As the recordings were not evenly distributed over time and some years were missing, we had to combine two decades as one TP (Figure 4A). See for an overview of the songs per TP and datasets in Supplementary Table 1.
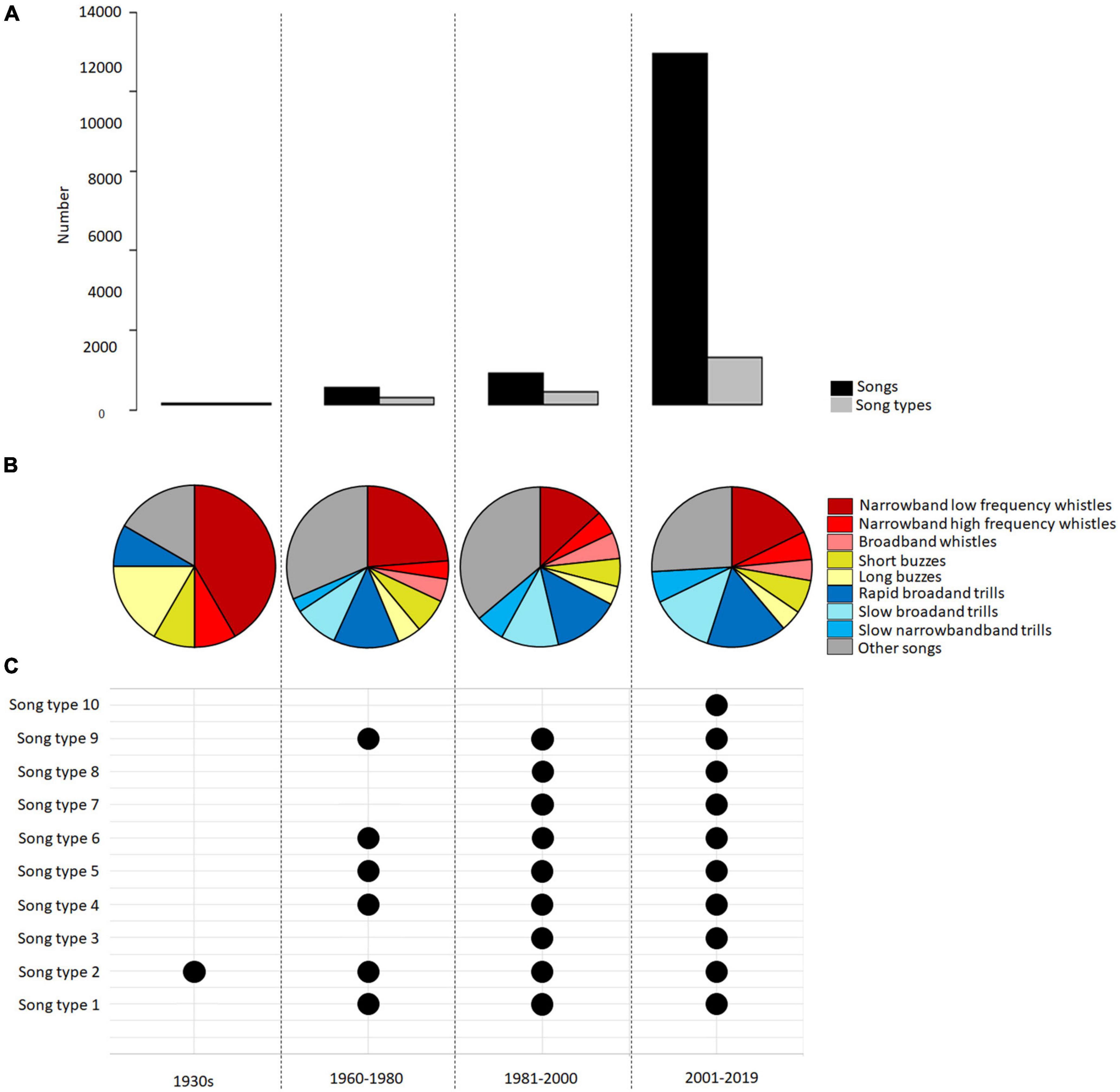
Figure 4. The structure of recordings of time period 1 (1930s), time period 2 (1960s–1980), time period 3 (1980s–2000), and time period 4 (2000s–2019). (A) Number of songs (black) and number of song types (gray). (B) Respective percentage of the nine sub-categories of song for the time periods. (C) Occurrence of the most common ten song types in Berlin spanning all time periods.
Comparison of Geographical Regions
For the geographical analysis, we selected the region Berlin-Brandenburg as a reference for a location since the majority of our recordings (58%) were generated there and both, the FFN and the AR recordings, largely originated from this region (99–100%). We did not have GPS tags of a large amount of the recordings for the countries Italy, France, Spain, and Portugal, which did not allow for a statistical method to classify the regions. We merely had the information about the respective countries but did refrain from a comparison based on national borders. Thus, due to the missing metadata, we based the geographic distribution on the approximate estimated distance from Berlin-Brandenburg and defined five geographical regions (Figure 5A). An overview of the songs per region separated according to the datasets is available in Supplementary Table 2.
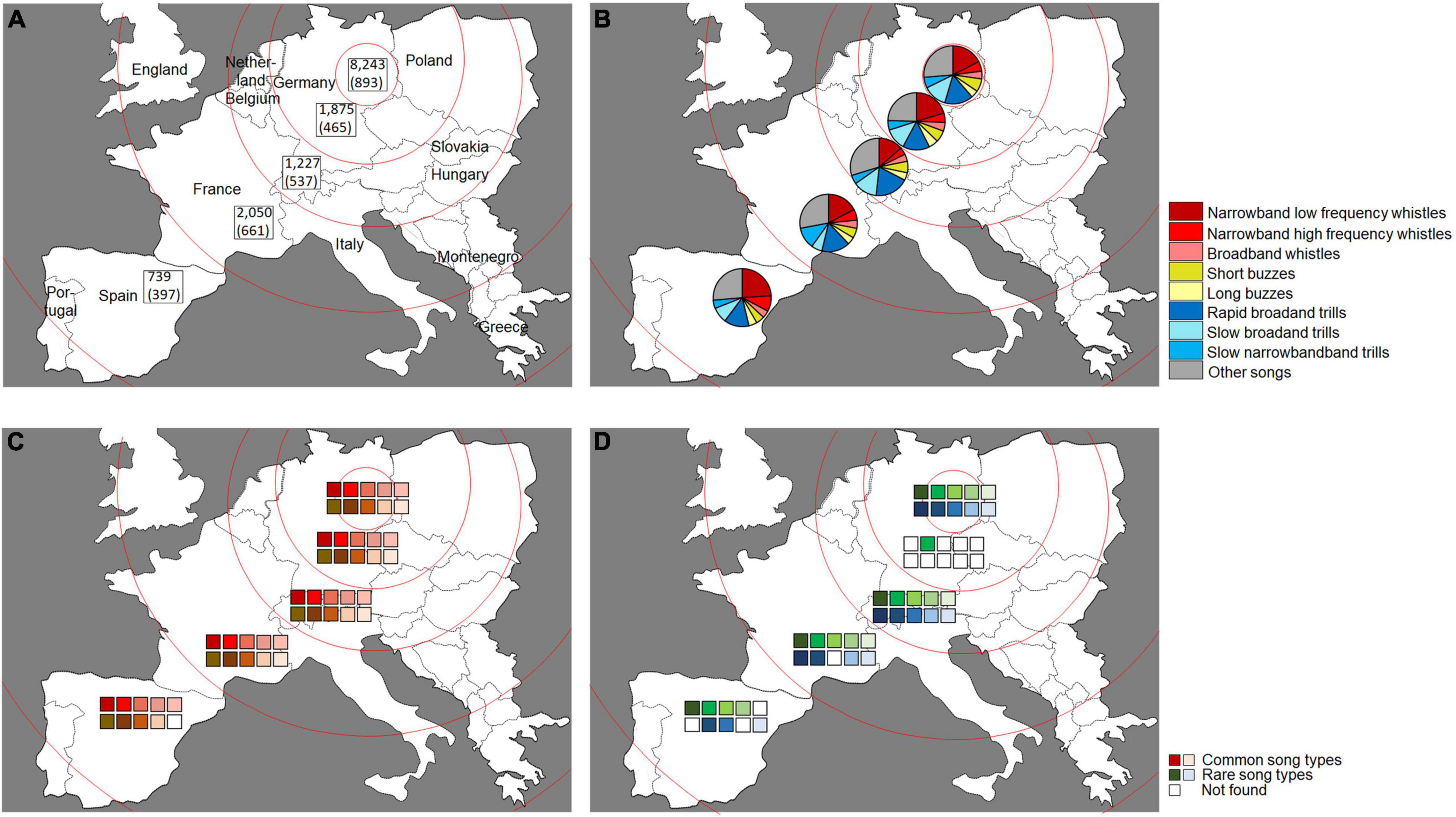
Figure 5. The distribution of the recordings across the five regions we defined. The regions were based on the approximate estimated distance from the Berlin-Brandenburg region. (A) Numbers of songs and song types (in brackets). (B) Respective percentage of the nine sub-categories of song for the other regions. (C) Occurrence of the most common ten song types from the Berlin-Brandenburg region in the other regions. The different shades of red each represent a song type. The most common song type is described by the top red left square. The tenth most common song type is the bottom red right square. A white square means that none of the song types in question were found. (D) Occurrence of the ten rare song types found only once in the Berlin-Brandenburg region in the other regions. The different shades of green and blue each represent a song type. A white square means that none of the song types in question were found.
Comparison of Different Databases
In order to compare the different datasets, we chose the same time period (TP4: 2001–2019) and region (Berlin-Brandenburg) to exclude temporal and regional differences in the analysis. We chose this period because the last recordings from the ASA were from 2015. In addition, the AR and FFN recordings were generated in 2018 and 2019. We thus had sufficient recordings to study and compare data from all datasets (AR: 2,294 songs; ASA: 1,669 songs; FFN: 3,366 songs; Xeno-Canto: 99 songs; Figure 6A).
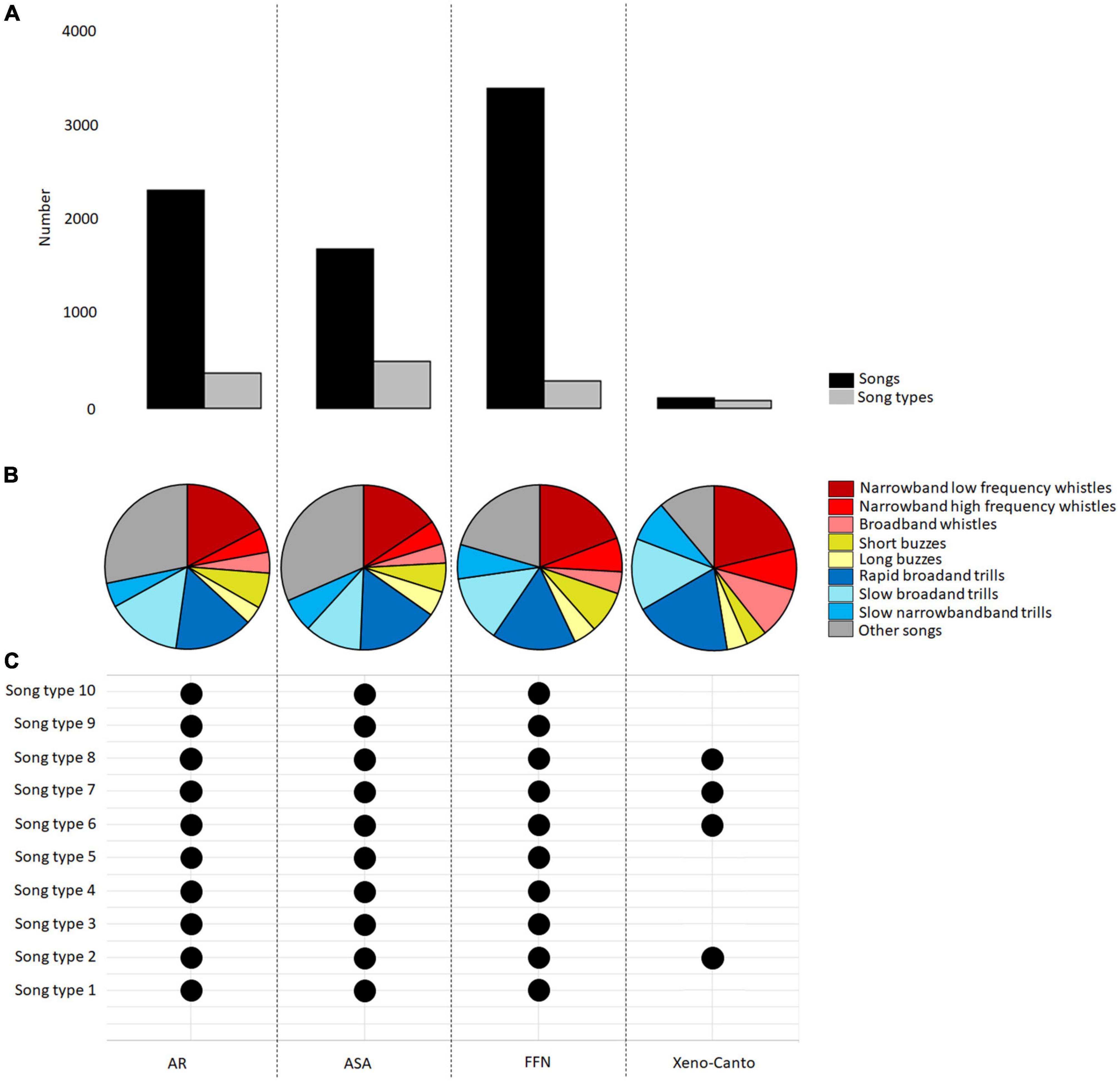
Figure 6. The structure of the recordings within the datasets (AR, academic researchers; ASA, animal sound archive; FFN, “Forschungsfall Nachtigall; Xeno-Canto) from Berlin spanning the period between 2001 and 2019. (A) Number of songs (black) and number of song types (gray). (B) Respective percentage of the nine sub-categories of song for the datasets. (C) Occurrence of the ten most common song types from Berlin in all time periods.
Statistical Analyses
For the occurrence analysis of a given song type, we recorded whether it was present or absent in the recordings that we analyzed over time, space or dataset. We defined the relative frequency from the number of times a song type appears divided by the total number of songs. Two one-sided tests (TOST) were used to test for the effects of time, space, and datasets for sub-categories of song and the relative frequency of song types. We set the upper and lower equivalence boundary at 0.5 and the statistical significance at p ≤ 0.05. All statistical analyses were performed using R version 4.0.0 (R-Team, 2020; unpublished data).
Results
European Song Type Catalog
Based on 14,140 songs from four datasets, we created a European nightingale song type catalog at the population level, which currently contains 1,868 different song types. As of September 2021, the catalog comprises 20% whistles, 12% buzz, 27% trills, and 41% so-called “other songs.” The nine sub-categories of song were significantly equivalent between the datasets in their percentages (TOST test: equivalent p-values of <0.01). The occurrences of song types differed among the four datasets. 78% of songs types appeared in only one of the datasets, 9% of songs types were found in two of the datasets, 5% of songs types occurred in three datasets and 8% of songs types were in all four datasets. Song types occurred with different frequencies in all datasets (1×: 56%; ≤10×: 30%; >100×: 1%). The rate of new song types that have not yet been sung (song type diversity) was different between the datasets (AR: each sixth song; ASA: each fourth song; FFN: each thirteenth song; Xeno-Canto: each fourth song). The agreement between the song types varied (highest between AR and FFN: 61%; lowest between Xeno-Canto and ASA: 26%; Supplementary Table 3).
Temporal Comparison of Nightingale Song
When comparing the sample years between 1930 and 2019 among all datasets, we detected that the nine sub-categories of song were distributed in a stable way with similar percentages over time (TOST test: equivalent p-values of <0.01). In most time periods, the narrowband low-frequency whistles, the rapid broadband trills, and the “other songs” accounted for the largest percentage of the nine sub-categories of song (Figure 4B and Supplementary Table 4). An analysis with regard to song type stability showed that the second most common song type in our dataset – a narrowband low-frequency whistle – occurred in all time periods. Six common song types were found in the recordings from the 60s until 2019 (Figure 4C). An analysis with regard to the relative frequency of the ten most common song types revealed that they accounted for 2% of the total number of songs and were significantly equivalent between the time periods (TOST test: equivalent p-values of <0.01).
Geographical Comparison of Nightingale Song
A comparison between the five geographic regions in all datasets showed a stable and similar distribution in the percentage of the nine sub-categories of song over space (TOST test: equivalent p-values of <0.01). The highest percentage of the nine sub-categories of song was accounted for by the low-frequency narrowband whistles, the rapid broadband trills and the “other songs” in all regions (Figure 5B and Supplementary Table 5). An analysis with regard to the stability of the ten most common song types revealed that nine of them were found in four of our geographic regions (Figure 5C). Six of the ten rare song types occurred in four regions (Figure 5D). An analysis with regard to the relative frequency of the ten most common song types showed that they accounted for 2% of the total number of songs and were significantly equivalent between the regions (TOST test: equivalent p-values of <0.01).
Comparison of Nightingale Song Among the Four Different Datasets
We found an equal distribution of the nine sub-categories of song when comparing the four datasets in the time between 2001 and 2019 from Berlin-Brandenburg (TOST test: equivalent p-values of <0.01). The low-frequency whistles had the highest percentage, followed by the rapid broadband trills and “other songs” (Figure 6B and Supplementary Table 6). Concerning the ten most common song types, four of them occurred in all datasets (Figure 6C), six of them occurred in the AR, ASA, and FFN recordings. An analysis with regard to the relative frequency of the ten most common song types revealed that they accounted for 3% of the total number of songs and were significantly equivalent between the datasets (TOST test: equivalent p-values of <0.01).
Discussion
The aim of this study was to investigate whether nightingale song is stable in terms of song type occurrence over nearly a century and whether song dialects are found in the relative frequency of song types across Europe. We also examined if sub-categories of song codes for stability or dialects. Additionally, we investigated if open, citizen science and shared data are an advantageous complement for dialect studies in a bird species with a large repertoire. Our study encompassed a period twice as large as previous studies (e.g., Grant and Grant, 1996) with recordings from the 1930s until 2019 and covered a linear geographic range of 2,652 km across 13 European countries, twice as large as previous studies (e.g., Weiss, 2012; unpublished data). In the following, we will evaluate the results from the biological (temporal stability and spatial dialects) and the technical (comparison of different datasets) perspectives.
Vocal Dialect Research Across Time and Space
Our European population-based nightingale song type catalog was three times larger than a previous one (623 song types; Weiss et al., 2014), which included songs of 271 individual nightingale males from a Berlin and 272 from a Brandenburg population. In addition, the catalogs differed in two more respects. First, we included all song types, instead of only those that appeared at least twice because otherwise that would have resulted in the loss of 1,048 song types. Second, in the definition of new song types: any differences in γ-section (our study) vs. less than 80% similarity or differences in more than three of ten elements in the α- and β-section (Weiss et al., 2014). However, we would like to emphasize that even if the total number of song types varies between the different catalogs, the results should remain unaffected. This notion is supported by the number of song types for the Berlin-Brandenburg region in two populations (623, Weiss et al., 2014) compared to multiple populations (893, our method).
Despite our large song type catalog, we found, for the first time, stability both in time (nearly a century) and space (geographical area — Europe) within the percentage of sub-categories of song and the occurrence of song types (common and rare) in the song of a bird species with a large repertoire. The stability of two frequent sub-categories of song could be explained by the fact that they may indicate the quality of a male, as low-frequency whistles and rapid broadband trills are difficult to produce as has been shown in other species (Podos, 1996; Kirschel et al., 2009), and degrade less in the nightingale habitat (Wiley and Richards, 1978). Thus, these sub-categories of song indicate stability over time and space in bird species with large (the nightingale) and small repertoires (Nelson et al., 2004) and cetaceans (Deecke et al., 2000). Temporal stability in the occurrence of song types have also been found in songbird species with a small repertoire (Grant and Grant, 1996), hummingbirds (González and Ornelas, 2009), parrots (Wright et al., 2008) and in cetaceans (Rekdahl et al., 2013), where the most common vocalizations were likewise stable. Although geographic persistence of certain song types has been noted in songbirds (Searfoss et al., 2020) and parrots (Wright et al., 2008), suggesting an important function in animal communication, we expected temporal stability and spatial drift: (a) Nightingales have been shown to adapt their song to the population from the first to the second breeding season (Kiefer et al., 2010, 2014). (b) Nightingales have been demonstrated to share song types among males of the same population to facilitate communication during interactions between males (Todt and Naguib, 2000). (c) Different song types are performed with different frequencies, while those shared with neighboring males are common (Hultsch, 1980; unpublished data). The stable occurrence of common and rare song types detected in our large dataset is consistent with previous playback experiments (Weiss, 2012; unpublished data). Moreover, this stability seems to be consistent with the nightingale behavior in the wintering grounds (Glutz von Blotzheim, 1988). Spatially separated nightingales probably interact on wintering grounds, which could lead to adaptation and stability of song types. These stable song types could have positive effects (adaptation to the environment, choice of female) and an important communicative function in social interactions, such as in cetaceans (Rekdahl et al., 2013). All this could have led to the fact that the song characteristics we studied were stable across time and space. Future studies should investigate exactly which and why these stable song types exist in nightingale song, and how they mediate communication.
In contrast to our findings on the stability of nightingale song, we observed no evidence of song dialects in the relative frequency of the ten most common song types across Europe. As Weiss (2012; unpublished data) found differences in the relative frequency of song types between populations from Berlin, Brandenburg, and France, we expected the same for Europe in general. However, the relative frequency of common song types was so low that no geographical differences could be inferred. This low relative frequency could be related to the size of our song type catalog but not to the datasets used, as they showed the same results. However, as the exact relative frequency of song types of a male can only be determined after approximately 1 h (533 songs; Kipper et al., 2004), we suggest that it is not the relative frequency of a single song type that codes for dialects, but the composition of song type delivery of a bird, i.e., repertoire or syntax (Weiss, 2012; unpublished data). This has already been suggested for orcas (Riesch et al., 2006; unpublished data) and is in line with previous bird studies showing that even species with a large repertoire recognize foreign males from short songs without the need to listen to the complete repertoire (e.g., Jaška et al., 2015; Moser-Purdy and Mennill, 2016). Since nightingales sing with immediate variety, i.e., the same song type is not sung repeatedly, but only after a certain number of songs (Hultsch and Todt, 1989), this pattern allows them to hear several song types in a short time – and thus may enable nightingales to determine whether a singer is a resident or a stranger. This result is indeed consistent with studies suggesting that song dialects are unlikely to emerge in species with a large vocal repertoire (Goodfellow and Slater, 1986; Lachlan et al., 2016; Tchernichovski et al., 2017), such as the nightingale. However, we need to stress that we in fact only looked at a rather small part of the nightingale song type catalog determined in our case study. Thus, future studies could extend the analysis to examine if potentially other common or rare song types code for dialects.
Combining Datasets for Research Over Time and Space
We evaluated OD, CS, and shared data for dialect research over time and space in direct comparison of sub-categories of song, as well as the occurrence and relative frequency of the ten most common song types in nightingale song. The similarity in the three most frequent sub-categories of song and variations in the other six sub-categories of song probably resulted from some song types occurring in only one dataset, leading to different song type diversities and thus percentage distribution. Because all ten common song types were found in the datasets, except Xeno-Canto, we suspect that the absence of the common song types was due to the number of analyzed songs. Since these 99 Xeno-Canto songs were only sufficient for finding four common song types, about three times the amount of songs is needed for a valid analysis. We showed that the datasets are not different per se, but our evaluation was influenced by other aspects.
The differences between the datasets were evident in their temporal and spatial validity for our analysis, as all exhibited either a temporal or spatial bias. Our temporal analysis was mainly conducted with the OD and the CS (especially Xeno-Canto) recordings. The spatial analysis was mainly possible with the CS recordings as these had the largest spatial distribution and the most metadata, since these were generated with an app (FFN) and Xeno-Canto requires a minimum of metadata for the recordings (Vellinga and Planqué, 2015). CS projects can provide more data with a greater geographic coverage than academic researchers alone, which are limited in number of simultaneously generated recordings due to the (mostly) stationary location and the restricted availability of (expensive) equipment (e.g., Vellinga and Planqué, 2015; Steen et al., 2021). The shared data were the least suited for our temporal and geographic analysis as they were only from 2 years from Berlin. The importance of the geographic distribution of the recordings was evident in two ways: First, song type diversity was mainly affected by the spatial distribution of the recordings and not by the number of songs and thus the size of the dataset nor the duration or the time of the recording as no song types only occur at night or day (Jäckel, 2013; unpublished data). Second, the degree of concordance of song types in each dataset was also influenced by spatial distribution, with datasets from the same region (e.g., AR and FFN) accounting for the greatest share. We conclude that to study song stability and dialects (1) the recording location is more important than recording length and that (2) many short, non-standardized OD and CS recordings may achieve the same diversity of song types within a dataset as long, standardized professional recordings. Thus, for our study using the nightingale as an example, standardized recordings were not required in terms of technology (professional equipment), time (nocturnal song), duration (1 h), or great experience of the recordist.
Our study showed that even if longer recordings would have been helpful, considering the large repertoire of the nightingale, it was more critical for our temporal and spatial analysis to have access to diverse datasets. To make access also possible for future nightingale researchers, we are in the process of making the recordings from the FFN online available in an open archive at the MfN. Currently only half of the academic researchers publish their data in open archives (Tenopir et al., 2011), as such access is not encouraged widely, which reduces the opportunities for reuse (Mills et al., 2015; Federer, 2020). Missing open science practices are particularly disadvantageous for evolution and ecology research, where datasets from longitudinal studies contribute greatly to scientific understanding but are expensive and time-consuming (Mills et al., 2015; Hughes et al., 2017). We would therefore like to encourage more song recordings to be made publicly available with sufficient metadata so that they can be effectively used in future research. Open science, and thus the availability of diverse datasets, allowed us to conduct the largest temporal and geographic study of song evolution to date. By directly comparing the datasets, we have shown that OD and CS recordings are well suited – and in some cases better suited – for studying song variation in bird species with a large repertoire than data collected by an academic research group, because they were collected over a shorter period of time and in a much smaller geographic range. Therefore, we recommend that OD and CS datasets should be used more frequently in ornithology and that song recordings should be openly accessible. Given that each dataset has different advantages and disadvantages, we further recommend combining diverse datasets for dialect research in bird species with a large repertoire.
Conclusion
In our study, we found the song of common nightingales to be stable over time and space. We did not detect evidence of dialects. However, the nightingale has a highly sophisticated syntax and a complex song, only a small part of which we studied, so further analysis is needed to rule out the absence or presence of dialects. Our results showed that non-standardized OD and CS recordings offer the potential to support ornithology and in particular dialect research. The general quality concerns about OD and CS could not be confirmed in our study. Based on our findings, when studying a bird species with a large repertoire such as the nightingale, we recommend combining different and diverse datasets as quality is multidimensional and always depends on the research question. As more and more (CS) recordings are currently being made available on open platforms (Sanderson et al., 2021; unpublished data), our study is of great use to increase trust in them. We hope to encourage fellow bird enthusiasts to share their song recordings in open repositories and promote the use of these data to the scientific community.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
Ethical review and approval were not required for the animal study because, in Germany, permission from an ethics committee is not required for such research questions.
Author Contributions
DJ, KM, and SV-H wrote the concept and design of the study and analyzed and interpreted the data. DJ and UB performed the statistical analysis and designed the figures. DJ wrote the first draft of the manuscript. KM, GL, and SV-H revised the manuscript. All authors contributed to the manuscript and revised, read, and approved the submitted version.
Funding
This publication was written as part of the project Forschungsfall Nachtigall, funded by the German Federal Ministry of Education and Research (BMBF, Förderkennzeichen 01BF1709), and Stadtnatur entdecken funded by the German Federal Ministry for the Environment, Nature Conservation and Nuclear Safety and The Leibniz Aktions Fond. The publication of this article will be funded by the Deutsche Forschungsgemeinschaft for Open Access publication costs. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank the BMBF (Förderkennzeichen: 01BF1709) for financing of the project “Forschungsfall Nachtigall” (herein: FFN) and MfN Berlin for hosting the project; Marco Walther, Vanessa Proß, Diana Trinh Hong, Solveyg Nagy, Sophie Holtz, and Sophia Wutke for their song recordings and other relevant information collected during the animal behavior course at the Freie Universität Berlin 2018 taught by Constance Scharff and Henrike Hultsch; and Constance Scharff and Henrike Hultsch for sharing these recordings with us. We would like to acknowledge the contributions of many enthusiastic citizen scientists and Sarah Darwin, Alexander Buhl, Nadja Tata, and Julia Rostin as part of the FFN. Furthermore, we are indebted to the whole “Stadtnatur entdecken” team, in particular Ulrike Sturm and Omid Khorramshahi, for the development of the app “Naturblick” and their ongoing excellent support during the project. Furthermore, we also thank Dietmar Todt for his wonderful support in this study and the kind collaboration on the song type catalog; Sarah Darwin, Ulrike Sturm, Mario Lasseck, Nicola Moczek, and Sophie Ewert for commenting on an earlier version of the manuscript; and Xeno-Canto for their efforts as an open resource for bird sounds. We are grateful to Karl-H. Frommolt, head of the animal sound archive, for sharing the recordings from over 89 years of nightingale song with us.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.778610/full#supplementary-material
Footnotes
References
Aplin, L. M., Major, R. E., Davis, A., and Martin, J. M. (2021). A citizen science approach reveals long-term social network structure in an urban parrot, Cacatua galerita. J. Anim. Ecol. 90, 222–232. doi: 10.1111/1365-2656.13295
Baker, M. C., and Mewaldt, L. R. (1978). Song dialects as barriers to dispersal in white-crowned sparrows, Zonotrichia leucophrys nuttalli. Evolution 32, 712–722. doi: 10.1111/j.1558-5646.1978.tb04624.x
Beecher, M. D., and Brenowitz, E. A. (2005). Functional aspects of song learning in songbirds. Trends Ecol. Evol. 20, 143–149. doi: 10.1016/j.tree.2005.01.004
Bonn, A., Richter, A., Vohland, K., Pettibone, L., Brandt, M., Feldmann, R., et al. (2017). Grünbuch Citizen Science Strategie 2020 für Deutschland. Cenate Sotto: Bürger schaffen Wissen (GEWISS).
Catchpole, C. K., and Slater, P. J. B. (2008). Bird Song: Biological Themes and Variations. Cambridge: Cambridge University Press.
de Camargo Barbosa, K. V., Develey, P. F., Ribeiro, M. C., and Jahn, A. E. (2021). The contribution of citizen science to research on migratory and urban birds in Brazil. Ornithol. Res. 29, 1–11. doi: 10.1007/s43388-020-00031-0
de Jesus, M. D., Zapelini, C., and Schiavetti, A. (2021). Can citizen science help delimit the geographical distribution of a species? The case of the Callistoctopus sp. (“eastern octopus”) on the Brazilian coast. Ethnobiol. Conserv. 10, 1–15. doi: 10.15451/ec2020-09-10.03-1-15
Deecke, V. B., Ford, J. K. B., and Spong, P. (2000). Dialect change in resident killer whales: implications for vocal learning and cultural transmission. Anim. Behav. 40, 629–638. doi: 10.1006/anbe.2000.1454
Derryberry, E. P., Claramunt, S., Derryberry, G., Chesser, R. T., Cracraft, J., Aleixo, A., et al. (2011). Lineage diversification and morphological evolution in a large-scale continental radiation: the Neotropical ovenbirds and woodcreepers (Aves: Furnariidae). Evolution 65, 2973–2986. doi: 10.1111/j.1558-5646.2011.01374.x
Diblíková, L., Pipek, P., Petrusek, A., Svoboda, J., Bílková, J., Vermouzek, Z., et al. (2019). Detailed large-scale mapping of geographical variation of yellowhammer Emberiza citrinella song dialects in a citizen science project. Ibis 161, 401–414. doi: 10.1111/ibi.12621
Dickinson, J. L., Shirk, J., Bonter, D., Bonney, R., Crain, R. L., Martin, J., et al. (2012). The current state of citizen science as a tool for ecological research and public engagement. Front. Ecol. Environ. 10, 291–297. doi: 10.1890/110236
Dominguez, M., Lapido, R., Gorrindo, A., Archuby, D., Correa, E., Llanos, F., et al. (2021). A citizen science survey discloses the current distribution of the endangered Yellow Cardinal Gubernatrix cristata in Argentina. Bird Conserv. Int. 31, 139–150. doi: 10.1017/S0959270920000155
Federer, L. (2020). Measuring and mapping data reuse: findings from an interactive workshop on data citation and metrics for data reuse. Harv. Data Sci. Rev. 2, 1–8. doi: 10.1162/99608f92.ccd17b00
Gaunt, S. L., Baptista, L. F., S<nchez, J. E., and Hernandez, D. (1994). Song learning as evidenced from song sharing in two hummingbird species (Colibri coruscans and C. thalassinus). Auk 111, 87–103. doi: 10.2307/4088508
Gil, D., and Gahr, M. (2002). The honesty of bird song: multiple constraints for multiple traits. Trends Ecol. Evol. 17, 133–141. doi: 10.1016/S0169-5347(02)02410-2
González, C., and Ornelas, J. F. (2009). Song variation and persistence of song neighborhoods in a lekking hummingbird. Condor 111, 633–640. doi: 10.1525/cond.2009.090029
Goodfellow, D., and Slater, P. (1986). A model of bird song dialects. Anim. Behav. 34, 1579–1580. doi: 10.1016/S0003-3472(86)80233-0
Grant, B. R., and Grant, P. R. (1996). High survival of Darwin’s finch hybrids: effects of beak morphology and diets. Ecology 77, 500–509. doi: 10.2307/2265625
Hochachka, W. M., Fink, D., Hutchinson, R. A., Sheldon, D., Wong, W. K., and Kelling, S. (2012). Data-intensive science applied to broad-scale citizen science. Trends Ecol. Evol. 27, 130–137. doi: 10.1016/j.tree.2011.11.006
Hughes, B. B., Beas-Luna, R., Barner, A. K., Brewitt, K., Brumbaugh, D. R., Cerny-Chipman, E. B., et al. (2017). Long-term studies contribute disproportionately to ecology and policy. Bioscience 67, 271–281. doi: 10.1093/biosci/biw185
Hultsch, H., Mundry, R., and Todt, D. (1999). “Learning, representation and retrieval of rule-related knowledge in the song system of birds,” in Learning, eds A. Friederici and R. Menzel (Berlin: Walter de Gruyter), 89–115. doi: 10.1515/9783110803488.89
Hultsch, H., and Todt, D. (1981). Repertoire sharing and song-post distance in nightingales (Luscinia Megarhynchos B.). Behav. Ecol. Sociobiol. 8, 183–188. doi: 10.1007/BF00299828
Hultsch, H., and Todt, D. (1982). Temporal performance roles during vocal interactions in nightingales (Luscinia megarhynchos B.). Behav. Ecol. Sociobiol. 11, 253–260. doi: 10.1007/BF00299302
Hultsch, H., and Todt, D. (1989). Memorization and reproduction of songs in nightingales (Luscinia megarhynchos): evidence for package formation. J. Comp. Physiol. A 165, 197–203. doi: 10.1007/BF00619194
Hultsch, H., and Todt, D. (1996). Discontinuous and incremental processes in the song learning of birds: evidence for a primer effect. J. Comp. Physiol. A. 179, 291–299. doi: 10.1007/BF00194983
Irwin, D. E., Bensch, S., and Price, T. D. (2001). Speciation in a ring. Nature 409, 333–337. doi: 10.1038/35053059
Jaška, P., Linhart, P., and Fuchs, R. (2015). Neighbour recognition in two sister songbird species with a simple and complex repertoire – a playback study. J. Avian Biol. 45, 151–158. doi: 10.1111/jav.00482
Kassen, M. (2013). A promising phenomenon of open data: a case study of the Chicago open data project. Govern. Inf. Q. 30, 508–513. doi: 10.1016/j.giq.2013.05.012
Kiefer, S., Scharff, C., Hultsch, H., and Kipper, S. (2014). Learn it now, sing it later? field and laboratory studies on song repertoire acquisition and song use in nightingales. Naturwissenschaften 101, 955–963. doi: 10.1007/s00114-014-1236-5
Kiefer, S., Sommer, C., Scharff, C., and Kipper, S. (2010). Singing the popular songs? nightingales share more song types with their breeding population in their second season than in their first. Ethology 116, 619–626. doi: 10.1111/j.1439-0310.2010.01774.x
Kiefer, S., Spiess, A., Kipper, S., Mundry, R., Sommer, C., Hultsch, H., et al. (2006). First-Year common nightingales (Luscinia Megarhynchos) have smaller song-type repertoire sizes than older males. Ethology 112, 1217–1224. doi: 10.1111/j.1439-0310.2006.01283.x
Kipper, S., Kiefer, S., Bartsch, C., and Weiss, M. (2015). Female calling? Song responses to conspecific call playbacks in nightingales, Luscinia megarhynchos. Anim. Behav. 100, 60–66. doi: 10.1016/j.anbehav.2014.11.011
Kipper, S., Mundry, R., Hultsch, H., and Todt, D. (2004). Long-term persistence of song performance rules in nightingales (Luscinia megarhynchos): a longitudinal field study on repertoire size and composition. Behaviour 141, 371–390. doi: 10.1163/156853904322981914
Kirchhoff, C., Callaghan, C. T., Keith, D. A., Indiarto, D., Taseski, G., Ooi, M. K., et al. (2021). Rapidly mapping fire effects on biodiversity at a large-scale using citizen science. Sci. Total Environ. 755:142348.
Kirschel, A. N., Blumstein, D. T., Cohen, R. E., Buermann, W., Smith, T. B., and Slabbekoorn, H. (2009). Birdsong tuned to the environment: green hylia song varies with elevation, tree cover, and noise. Behav. Ecol. 20, 1089–1095. doi: 10.1093/beheco/arp101
Kroodsma, D., Hamilton, D., Sánchez, J. E., Byers, B. E., Fandiño-Mariño, H., Stemple, D. W., et al. (2013). Behavioral evidence for song learning in the suboscine bellbirds (Procnias spp.; Cotingidae). Wilson J. Ornithol. 125, 1–14. doi: 10.1676/12-033.1
Kunc, H. P., Amrhein, V., and Naguib, M. (2006). Vocal interactions in nightingales, Luscinia megarhynchos: more aggressive males have higher pairing success. Anim. Behav. 72, 25–30. doi: 10.1016/j.anbehav.2005.08.014
Lachlan, R. F., van Heijningen, C. A. A., ter Haar, S. M., and ten Cate, C. (2016). Zebra finch song phonology and syntactical structure across populations and continents-a computational comparison. Front. Psychol. 7:980. doi: 10.3389/fpsyg.2016.00980
Landgraf, C., Weiss, M., and Kipper, S. (2015). Multiple song features are related to paternal effort in common nightingales. BMC Evol. Biol. 15:115. doi: 10.1186/s12862-015-0390-5
Lukyanenko, R., Parsons, J., and Wiersma, Y. F. (2016). Emerging problems of data quality in citizen science. Conserv. Biol. 30, 447–449. doi: 10.1111/cobi.12706
Lynch, A. (1996). “The population memetics of birdsong,” in Ecology and Evolution of Acoustic Communication in Birds, eds D. E. Kroodsma and E. H. Miller (New York: Cornell University Press), 181–197. doi: 10.7591/9781501736957-017
Mennill, D. J., Doucet, S. M., Newman, A. E. M., Williams, H., Moran, I. G., Thomas, I. P., et al. (2018). Wild birds learn songs from experimental vocal tutors. Curr. Biol. 28, 3273–3278. doi: 10.1016/j.cub.2018.08.011
Mills, J. A., Teplitsky, C., Arroyo, B., Charmantier, A., Becker, P. H., Birkhead, T. R., et al. (2015). Archiving primary data: solutions for long-term studies. Trends Ecol. Evol. 30, 581–589. doi: 10.1016/j.tree.2015.07.006
Moser-Purdy, C., and Mennill, D. J. (2016). Large vocal repertoires do not constrain the dear enemy effect: a playback experiment and comparative study of songbirds. Anim. Behav. 118, 55–64. doi: 10.1016/j.anbehav.2016.05.011
Naguib, M., Mundry, R., Hultsch, H., and Todt, D. (2002). Responses to playback of whistle songs and normal songs in male nightingales: effects of song category, whistle pitch, and distance. Behav. Ecol. Sociobiol. 52, 216–223.
Neate-Clegg, M. H., Horns, J. J., Adler, F. R., Aytekin, M. ÇK., and Şekercioğlu, ÇH. (2020). Monitoring the world’s bird populations with community science data. Biol. Conserv. 248:108653. doi: 10.1016/j.biocon.2020.108653
Nelson, D. A., Hallberg, K. I., and Soha, J. A. (2004). Cultural evolution of puget sound white-crowned sparrow song dialects. Ethology 110, 879–908. doi: 10.1111/j.1439-0310.2004.01025.x
Poulsen, H. (1958). The Calls of the Chaffinch (Fringilla coelebs L.) in Denmark. Dan. Ornithol. Foren. Tidsskr. 52, 89–105.
Pozzi, L., Gamba, M., and Giacoma, C. (2010). The use of Artificial Neural Networks to classify primate vocalizations: a pilot study on black lemurs. Am. J. Primatol. 72, 337–348. doi: 10.1002/ajp.20786
Prat, Y., Azoulay, L., Dor, R., and Yovel, Y. (2017). Crowd vocal learning induces vocal dialects in bats: playback of conspecifics shapes fundamental frequency usage by pups. PLoS Biol. 15:e2002556. doi: 10.1371/journal.pbio.2002556
Rekdahl, M. L., Dunlop, R. A., Noad, M. J., and Goldizen, A. W. (2013). Temporal stability and change in the social call repertoire of migrating humpback whales. J. Acoust. Soc. Am. 133, 1785–1795. doi: 10.1121/1.4789941
Rendell, L., and Whitehead, H. (2005). Spatial and temporal variation in sperm whale coda vocalizations: stable usage and local dialects. Anim. Behav. 70, 191–198. doi: 10.1016/j.anbehav.2005.03.001
Roth, T., and Amrhein, V. (2010). Estimating individual survival using territory occupancy data on unmarked animals. J. Appl. Ecol. 47, 386–392. doi: 10.1111/j.1365-2664.2010.01786.x
Searfoss, A. M., Liu, W.-C., and Creanza, N. (2020). Geographically well-distributed citizen science data reveals range-wide variation in the chipping sparrow’s simple song. Anim. Behav. 161, 63–76. doi: 10.1016/j.anbehav.2019.12.012
Sharpe, D. L., Castellote, M., Wade, P. R., and Cornick, L. A. (2019). Call types of Bigg’s killer whales (Orcinus orca) in western Alaska: using vocal dialects to assess population structure. Bioacoustics 28, 74–99. doi: 10.1080/09524622.2017.1396562
Steen, V. A., Tingley, M. W., Paton, P. W., and Elphick, C. S. (2021). Spatial thinning and class balancing: key choices lead to variation in the performance of species distribution models with citizen science data. Methods Ecol. Evol. 12, 216–226. doi: 10.1111/2041-210X.13525
Tchernichovski, O., Feher, O., Fimiarz, D., and Conley, D. (2017). How social learning adds up to a culture: from birdsong to human public opinion. J. Exp. Biol. 220, 124–132. doi: 10.1242/jeb.142786
Tenopir, C., Allard, S., Douglass, K., Aydinoglu, A. U., Wu, L., Read, E., et al. (2011). Data sharing by scientists: practices and perceptions. PLoS One 6:e21101. doi: 10.1371/journal.pone.0021101
Todt, D., and Naguib, M. (2000). Vocal interactions in birds: the use of song as a model in communication. Adv. Stud. Behav. 29, 247–296. doi: 10.1016/S0065-3454(08)60107-2
Vellinga, W. P., and Planqué, R. (2015). “The Xeno-canto collection and its relation to sound recognition and classification,” in Proceedings of the 2015 CLEF, (Toulouse: CLEF).
Weiss, M., Hultsch, H., Adam, I., Scharff, C., and Kipper, S. (2014). The use of network analysis to study complex animal communication systems: a study on nightingale song. Proc. R. Soc. B Biol. Sci. 281:20140460. doi: 10.1098/rspb.2014.0460
Weiss, M., Kiefer, S., and Kipper, S. (2012). Buzzwords in females’ ears? the use of buzz songs in the communication of nightingales (Luscinia megarhynchos). PLoS One 7:e45057. doi: 10.1371/journal.pone.0045057
Weisshaupt, N., Lehikoinen, A., Mäkinen, T., and Koistinen, J. (2021). Challenges and benefits of using unstructured citizen science data to estimate seasonal timing of bird migration across large scales. PLoS One 16:e0246572. doi: 10.1371/journal.pone.0246572
Wiley, R. H., and Richards, D. G. (1978). Physical constraints on acoustic communication in the atmosphere: implications for the evolution of animal vocalizations. Behav. Ecol. Sociobiol. 3, 69–94.
Wright, T. F., and Dahlin, C. R. (2018). Vocal dialects in parrots: patterns and processes of cultural evolution. Emu 118, 50–66. doi: 10.1080/01584197.2017.1379356
Keywords: birdsong, cultural evolution, geographic variation, Luscinia megarhynchos, repository, song dialects, vocal learning
Citation: Jäckel D, Mortega KG, Brockmeyer U, Lehmann GUC and Voigt-Heucke SL (2022) Unravelling the Stability of Nightingale Song Over Time and Space Using Open, Citizen Science and Shared Data. Front. Ecol. Evol. 10:778610. doi: 10.3389/fevo.2022.778610
Received: 17 September 2021; Accepted: 24 January 2022;
Published: 17 February 2022.
Edited by:
Renoult P. Julien, UMR 5175 Centre d’Ecologie Fonctionnelle et Evolutive (CEFE), FranceReviewed by:
Aloïs Robert, Yale University, United StatesBernard Lohr, University of Maryland, Baltimore County, United States
Copyright © 2022 Jäckel, Mortega, Brockmeyer, Lehmann and Voigt-Heucke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Denise Jäckel, RGVuaXNlLkphZWNrZWxAbWZuLmJlcmxpbg==