 Martina Carrete1*
Martina Carrete1* Fernando Hiraldo2
Fernando Hiraldo2 Pedro Romero-Vidal1,2Guillermo Blanco3
Pedro Romero-Vidal1,2Guillermo Blanco3 Dailos Hernández-Brito2
Dailos Hernández-Brito2 Esther Sebastián-González4
Esther Sebastián-González4 José A. Díaz-Luque5
José A. Díaz-Luque5 José L. Tella2
José L. Tella2
- 1Department of Physical, Chemical and Natural Systems, Universidad Pablo de Olavide, Seville, Spain
- 2Department of Conservation Biology, Estación Biológica de Doñana (CSIC), Seville, Spain
- 3Department of Evolutionary Ecology, Museo Nacional de Ciencias Naturales (CSIC), Madrid, Spain
- 4Department of Ecology, Universidad de Alicante, Alicante, Spain
- 5Endangered Conservation Consultancy, Málaga, Spain
Palms, like all plants, show coevolutionary relationships with animals that have been traditionally categorized as mutualistic (seed dispersers and pollinators) or antagonistic (seed predators). This dual perspective, however, has prevented a full understanding of their true interactions with some animal groups, mainly those that do not ingest entire fruits. One clear example is parrots, which have been described to use palm species as feeding resources, while their role as seed dispersers has been largely neglected. Here, we combined fieldwork data with information from the literature and citizen science (i.e., naturalists and nature photographers) on parrot foraging ecology worldwide to evaluate the spatial and taxonomic extent of parrot-palm interactions and to identify the eco-evolutionary factors involved. We identified 1,189 interactions between 135 parrots and 107 palm species in more than 50 countries across the six realms where palms are present as natives or introduced. Combining this information, we identified 427 unique parrot-palm interacting pairs (i.e., a parrot species interacting with a palm species). Pure antagonistic interactions (i.e., parrots just preying on seeds or eating or destroying their non-reproductive parts) were less common (5%) than mutualistic ones (i.e., parrots benefiting by partially preying on the seed or fruit or consuming the pulp of the fruit or the flower but also contributing to seed dispersal and, potentially, pollination; 89%). After controlling for phylogeny, the size of consumed seeds and parrot body mass were positively related. Seed dispersal distances varied among palm species (range of estimated median dispersal distances: 9–250 m), with larger parrots dispersing seeds at greater distances, especially large fruits commonly categorized as megafauna anachronisms (>4 cm length). Although parrot-palm interactions are widespread, several factors (e.g., social behavior, predation fear, food availability, or seasonality) may affect the actual position of parrots on the antagonism-mutualism continuum for different palm species and regions, deserving further research. Meanwhile, the pervasiveness of parrot-palm mutualistic interactions, mainly involving seed dispersal and pollination, should not be overlooked in studies of palm ecology and evolution.
Introduction
Plant-animal interactions are ubiquitous and can range from relationships that benefit both participating species (mutualisms) to interactions where only one is benefited while the other is negatively affected (antagonisms) (Mougi, 2020). However, categorizing plant-animal interactions as purely mutualistic or antagonistic is sometimes unclear. For instance, seed or pollen eaters can be considered as predators (antagonism) but also as dispersers if they move viable seeds after ingesting them or pollen further away from the mother plants (mutualism), promoting gene flow within and among plant populations (e.g., Rousset, 2004; Pérez-Méndez et al., 2016; Wandrag et al., 2018) and the colonization of new microhabitats (e.g., Janzen, 1970; Howe and Smallwood, 1982; Cousens et al., 2008). Therefore, plant-animal interactions, which have evolved over geological time and have driven both biodiversity patterns and species adaptations, should be studied considering all potential costs and benefits for both animals and plants (Montesinos-Navarro et al., 2017).
The life history of tropical trees is intimately connected to vertebrates through seed dispersal and seed predation (Levey et al., 1994). Palms (Arecaceae) are among the most abundant tree groups in tropical and sub-tropical ecosystems around the world (Pitman et al., 2001; Dransfield et al., 2008; Muscarella et al., 2020). With more than 2,400 species (Govaerts and Dransfield, 2005; Dransfield et al., 2008), palms exhibit large geographic variations in species richness, phylogenetic composition, and life forms, and serve as keystone resources for pollinators and frugivores (Terborgh, 1986; Zona and Henderson, 1989; Henderson, 2002; Onstein et al., 2017; Muñoz et al., 2019; Lim et al., 2020) but also for people by providing construction materials, fabrics, fuel, food, medicine, and ornamentals (Cámara-Leret et al., 2017). Thus, understanding factors driving palm recruitment, diversity, and distribution, which are all tightly linked to predation, dispersal, and plant regeneration, is pivotal not only to ensure their long-term persistence but also the structuring and functioning of tropical ecosystems (Eiserhardt et al., 2011).
Palms are predominantly animal dispersed (Zona and Henderson, 1989; Zona, 2006; Andreazzi et al., 2009; Muñoz et al., 2019; Dracxler and Kissling, 2021), so frugivory plays a special role in their population dynamics and evolution (Dracxler and Kissling, 2021). Although most vertebrates can disperse palm seeds, several studies provide evidence of dispersal limitations at broad and local scales among palms (Svenning, 2001, 2002; Sunderland and Morakinyo, 2002; Boll et al., 2005; Svenning and Wright, 2005; Blach-Overgaard et al., 2009, 2010). Mammals and birds have been listed as the principal dispersers for palms, in particular, tapirs, peccaries, toucans, cracids, and oilbirds, which disperse palm seeds by endozoochory (reviews in Zona and Henderson, 1989; Zona, 2006; Andreazzi et al., 2009; Muñoz et al., 2019; Dracxler and Kissling, 2021). However, palm seeds can also be effectively dispersed by stomatochory (i.e., animals flying or moving over the canopy with fruits in their beaks, mouths, or feet), such as large-fruited palm species dispersed by monkeys, rodents, or bats (e.g., Adler and Kestell, 1998; Chapman and Russo, 2007; Jansen et al., 2012; Abedi-Lartey et al., 2016; Blanco et al., 2019; Dracxler and Kissling, 2021).
Parrots (order Psittaciformes) are among those species potentially dispersing palms by stomatochory. While parrots have been traditionally considered as seed predators (Toft and Wright, 2015), recent studies have described multiple examples across the world of parrots legitimately dispersing several plant species through epizoochory, endozoocory, and stomatochory (Tella et al., 2015; Blanco et al., 2018; Hernández-Brito et al., 2021) as well as transferring pollen (i.e., acting as pollinators) after feeding on conifer cones or flowers (Fleming and Muchhala, 2008; Gleiser et al., 2017), thus challenging this paradigm. Parrots are among the richest avian orders (c. 400 extant species) and, although several species have decreasing populations in their native ranges (e.g., Olah et al., 2016; Berkunsky et al., 2017), they are still rather abundant compared with other taxa (Callaghan et al., 2021; but see Robinson et al., 2022 for caution on population estimates). This, in addition to their generalist diets and plastic feeding behaviors (Renton et al., 2015; Toft and Wright, 2015), may explain the pervasiveness of parrot-plant interactions, involving not only native but also exotic parrot and plant species. In the case of palms, a few studies have demonstrated that parrots predate on palm seeds or consume the pulp of the fruit but are also legitimate long-distance dispersers of some Neotropical species (Sazima, 2008; Prada Villalobos and Araújo Bagno, 2012; Baños-Villalba et al., 2017; Luna et al., 2018; Blanco et al., 2019; Tella et al., 2020; Silva et al., 2021). However, the large overlap between palm and parrot distributions (Provost et al., 2018; Reichgelt et al., 2018) suggests that this avian order may play a major —though overlooked— role in palm ecology worldwide (Zona and Henderson, 1989; Muñoz et al., 2019; Dracxler and Kissling, 2021).
Here, we combine fieldwork with published data and information gathered from citizen science (i.e., naturalists and nature photographers) to assess the nature of parrot-palm interactions (mutualistic or antagonistic), the eco-evolutionary factors involved, and their spatial extent across the ice-free realms of the world (sensu Olson et al., 2001), namely Nearctic, Palaearctic, Afrotropic, Indomalaya, Neotropic, Australasia, and Oceania (hereafter, these last two realms will be referred to as Australasia). As previously suggested (Montesinos-Navarro et al., 2017), we expect most interactions to be mutualistic. Moreover, given that the maximum number of interactions in an area depends on the number of interacting species (Jordano, 2016), regions with more palms (Kissling et al., 2007), and parrot species (Davies et al., 2007) such as the Neotropic, will show more interactions than regions with low numbers of species. However, although parrots may interact with palms in multiple ways, some interactions such as seed predation, defleshing, or dispersal may be limited by parrot morphology (mainly beak and body size) and palm fruit/seed size, as has been seen in other plant-frugivore pairs (Onstein et al., 2017). Thus, we predict a functional match between palm fruit size (used as a surrogate of palm seed size, see Supplementary Material) and parrot body size in all realms, regardless of whether palms were native or introduced. However, parrots can consume unripe fruits (Sebastián-González et al., 2019) and this may dismantle the functional trait matching by allowing small-sized parrots to interact with large-sized fruit palms. Moreover, as large-sized parrots are less constrained in consuming small or large fruits, they will be able to interact with more palm species than small-sized parrots, which can only interact with palms with small fruits. Accordingly, palms with smaller fruits will be able to interact with more parrot species than large-fruited palms, which will only interact with large-sized parrots. As for dispersal, since large-sized parrots are able to flight over larger areas and disperse heavier palm fruits (e.g., Tella et al., 2020) than smaller parrot species (e.g., Luna et al., 2018), we expect large-fruited palms to show longer parrot-mediated dispersal distances than small-fruited ones. Finally, despite differences in features linked to competitive ability, fecundity, and dispersal between native and introduced palms (Fehr et al., 2020), we do not expect differences in their interactions with parrots due to the generalist feeding behavior of these birds (Toft and Wright, 2015; Table 1).
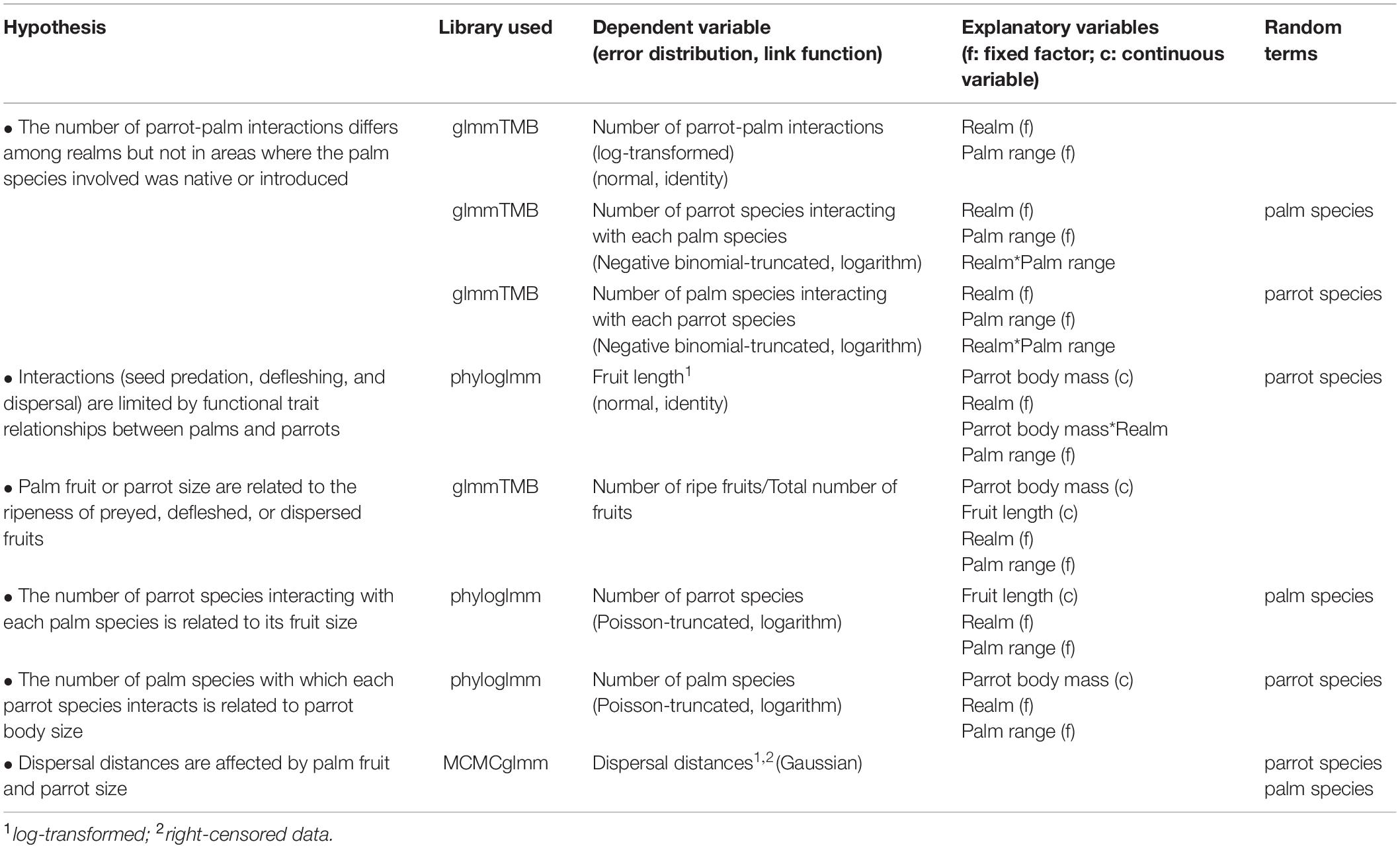
Table 1. Summary of the hypotheses and details about the models used in each case.
Materials and Methods
Parrot-Palm Interactions
Parrot-palm interactions were obtained by combining field data, bibliographic information, and citizen science. Depending on the parrot behavior, interactions were classified as seed predation (i.e., seeds or entire fruits, usually a single-seeded drupe although some genera may contain two or more seeds in each fruit; Henderson, 2002), flowers, or non-reproductive parts (i.e., bark, leaves, or petioles), defleshing (i.e., consumption of the pulp surrounding the seed, usually sweet and highly nutritive; Henderson, 2002), or dispersal (i.e., flying parrots observed carrying fruits or seeds in their beaks or feet). In all cases, both palms and parrots were identified to the species level, discarding data when this was not possible. The taxonomic names of palms and parrots, when needed, were updated following the World Checklist of palms (WFO, 2021), the World Checklist of Selected Plant Families,1 and the BirdLife Taxonomic Checklist (BirdLife International, 2020), respectively. We classified the interaction based on whether a native or introduced palm was observed using the information on botanical countries provided by the Taxonomic Databases Working Group (2001), which mostly represents countries except for some of the largest ones which are subdivided into states or provinces (Kissling et al., 2012).
Field Data
Parrot-palm interactions were collected in different fieldwork campaigns performed in 17 countries and five continents between 2012 and 2020. We actively looked for foraging groups of parrots both through roadside surveys (98 roadside surveys covering c. 57,250 km of transects, Tella et al., 2021) and a large, but unquantified, number of walking transects across a variety of biomes and habitats (Sebastián-González et al., 2019; Hernández-Brito et al., 2021). When we located parrots feeding on palm trees, we observed them with telescopes and binoculars from a distance to avoid disturbing the birds. We recorded the parrot and the palm species involved, and whether parrots were preying on the seeds and/or fruits, flowers, or non-reproductive parts, consuming the pulp of the fruits (defleshing), or dispersing them (fruits and/or seeds). When good visibility was possible, we recorded the distance from the mother tree to the perching site where the fruit or seed was dropped (exact dispersal distance) or up to where the parrot went out of sight (minimum dispersal distance) using a laser rangefinder incorporated into the binoculars (Leica Geovid 10 × 42 ×, range: 10–1,300 m). Some of these data have been previously published (Baños-Villalba et al., 2017; Luna et al., 2018; Blanco et al., 2019, Tella et al., 2020).
Bibliographic Search
We compiled interactions from a literature search of the Web of Science (WoS; July 10, 2020) including the terms: “palm*” and “comportam* aliment*” or aliment* or “ecolog* forra*” or “forra*” or feed* or forag* or food* or aliment* or diet* or “resource use” and parrot* or psittaci* or ara or parak* or macaw* or cockato* or amazon* or guacamay* or loro* or cotorr* or peri* or papagay* or periqu* or arara*. From the first list obtained (N = 9,160 papers), we discarded articles that did not include explicit information on parrot-palm interactions. Selected articles (n = 180 papers; Supplementary Table 1) were manually and meticulously screened to obtain information on the parrot and the palm species involved, the interaction type (predation of fruits or seeds, flowers, or non-reproductive parts of the palm, defleshing, or dispersal), and the location to identify the realm and whether the palm was native or introduced. We complemented this search using Academia.edu. We considered an observation as a dispersal interaction when parrots were recorded as consuming the pulp or fruits (seeds) of equal or smaller size than those previously recorded as dispersed by each species by stomatochory, following the same criterion used for palms dispersed by endozoochory (Muñoz et al., 2019). When possible, we established if the palm fruit was ripe or unripe (i.e., based on fruit color and size). Interactions cited in a paper were checked in the original study to avoid potential errors associated with misinterpretations.
Citizen Science Data
We complemented our dataset with information from wildlife naturalists and photographers. Information from wildlife naturalists was compiled from acquaintances who sent data on a non-systematic basis. Data from wildlife photographers were collected making a non-exhaustive review of several photo galleries publicly available on the internet, namely: eBird,2 WikiAves,3 The Cornell Lab of Ornithology,4 Flickr,5 Instagram,6 and Facebook.7 All the information was carefully examined and interactions were classified as predation of fruits and/or seeds, flowers, or non-reproductive parts, defleshing, or dispersal, following previous criteria. When interactions were not easy to interpret (mainly when dealing with pictures), we asked the authors for clarifications or discarded the observation. Plant and parrot species were identified when possible or the photographers were contacted for assistance in identification. The location of the sighting was recorded to assess whether the palm was native or introduced.
Statistical Analysis
We compared the number of parrot-palm interactions (log-transformed) across realms and in areas where the palm species involved were native or introduced using Generalized Linear Models (package glmmTMB; Brooks et al., 2017), applying Tukey post hoc tests (package lsmeans; Lenth, 2016). Similarly, we assessed whether the number of parrot species interacting with each palm species and vice versa varied across realms through Generalized Linear Mixed Models (palm or parrot species were used as random terms, respectively), considering if the palm was in its native or introduced range. As seed predation, defleshing, and dispersal can be limited by palm fruit size but also by parrot size, we considered whether these two variables were related (dependent variable: fruit length, log-transformed; normal error distribution, identity link function; parrot species as a random term) and if they affected the number of interacting species (dependent variables: number of parrot or palm species; truncated Poisson error distribution, log-link function; palm or parrot species as random terms, respectively). As palm fruit and parrot sizes are similar among closely related species (see Phylogenetic signal, Supplementary Figures 1, 2), we used phylogenetic generalized linear mixed models (PGLMMs) implemented in the phyloglmm package (Li and Bolker, 2019). For parrots, we used the resolved phylogeny of Jetz et al. (2012) while for palms we employed those published by Faurby et al. (2016). All models included the interaction between palm fruit and parrot sizes, the realm and whether the palm was native or introduced. We used GLM to assess whether palm fruit or parrot sizes were related to the ripeness of fruits (ripe or unripe) when they were preyed, defleshed, or dispersed, considering potential differences among realms and native and introduced palms (response variable: ripe fruits/total, binomial error distribution, logit link function; 282 parrot-palm pairs, 2,092 fruits classified as ripe or unripe). We exclusively focused on our field data to estimate the dispersal distances at which parrots dropped palm fruits or seeds, and did not include bibliographic data to avoid methodological differences. As our dispersal distances were right-censored (i.e., some corresponded to exact distances at which fruits or seeds were dropped, whereas, for others, the true distance was outside the observed sample range), we estimated the median distance at which parrots dispersed the fruits or seeds of each palm species using Kaplan–Meier survival curves. This statistical procedure, commonly used for survival analysis, does not assume underlying probability distributions and allows the inclusion of categorical variables. The low number of exact dispersal distances compared to minimum dispersal distances for most parrot species precludes the inclusion of this variable (i.e., parrot species) in the models. Thus, we estimated dispersal distances for each palm species after pooling information from all parrots dispersing them. Finally, we used MCMCglmm (Hadfield, 2010) to relate dispersal distances (log-transformed, right-censored data, cengaussian distribution) to palm fruit and parrot sizes, including fruit length and parrot body mass as covariates, and parrot and palm species as random terms. All models included the phylogenetic relationships between species (parrots or palms; Garamszegi, 2014), and were run for 100,000 iterations, preceded by a burn-in of 10,000 iterations. Estimates of parameters were stored every 25th iteration to reduce autocorrelation. The inclusion of the palm and parrot species in MCMCglmm allowed us to estimate how repeatable dispersal distances were for each group (palm or parrot species) as σparrot/(σparrot + σpalm + σresidual) and σpalm/(σparrot + σpalm + σresidual), respectively. Data on body mass, as an indicator of parrot body size, and palm fruit size were taken from the literature (Burgio et al., 2019; Kissling et al., 2019). The use of fruit length (the variable with more data across palm species) as an overall indicator of fruit and seed size was supported by its strong positive correlations across palm species with other descriptors of fruit and seed size (Supplementary Figure 3). Table 1 summarizes hypotheses tested and statistical details.
GLMs, GLMMs, and PGLMMs were compared using the Akaike Information Criteria corrected for small sample sizes (AICc), and models within 2 AICc units of the best one were considered as alternatives. Variables were considered as significant based on their associated p-value. For MCMCglmm, we tested the statistical support of the fixed effect by evaluating whether their posterior distributions (95% credible interval) overlapped with zero. Statistical analyses were conducted in R 4.1.0 (R Core Team, 2021).
Results
We recorded a total of 1,189 interactions between 135 parrots and 107 palm species around the world (Figure 1), including parrots eating or destroying the non-reproductive parts (4%), consuming flowers (11%), preying on fruits and seeds (30%), consuming the pulp (25%), and dispersing fruits or seeds (30%) (Table 2). We combined these interactions (n = 427 unique parrot-palm interacting pairs, some of them repeated in different realms; Table 2), balancing the costs and benefits for each species involved. Antagonistic interactions in which only parrots benefited while causing harm to the palm (i.e., parrots preying on seeds or eating or destroying the non-reproductive parts) totaled 20 cases (5%), while mutualist interactions (i.e., parrots benefiting by preying on the seed or consuming the pulp of the fruit or the flower but also contributing to seed dispersal and, potentially, pollination) were much more common (89%, n = 381 parrot-palm interacting pairs). The other 26 parrot-palm interacting pairs (6%) correspond to parrots defleshing and predating on palm fruits but without confident information on potential fruit dispersal. Parrot species interacting with palms ranged in body mass between 28 and 1,565 g, while the fruit size of palms interacting with parrots ranged between 0.6 and 22.5 cm.
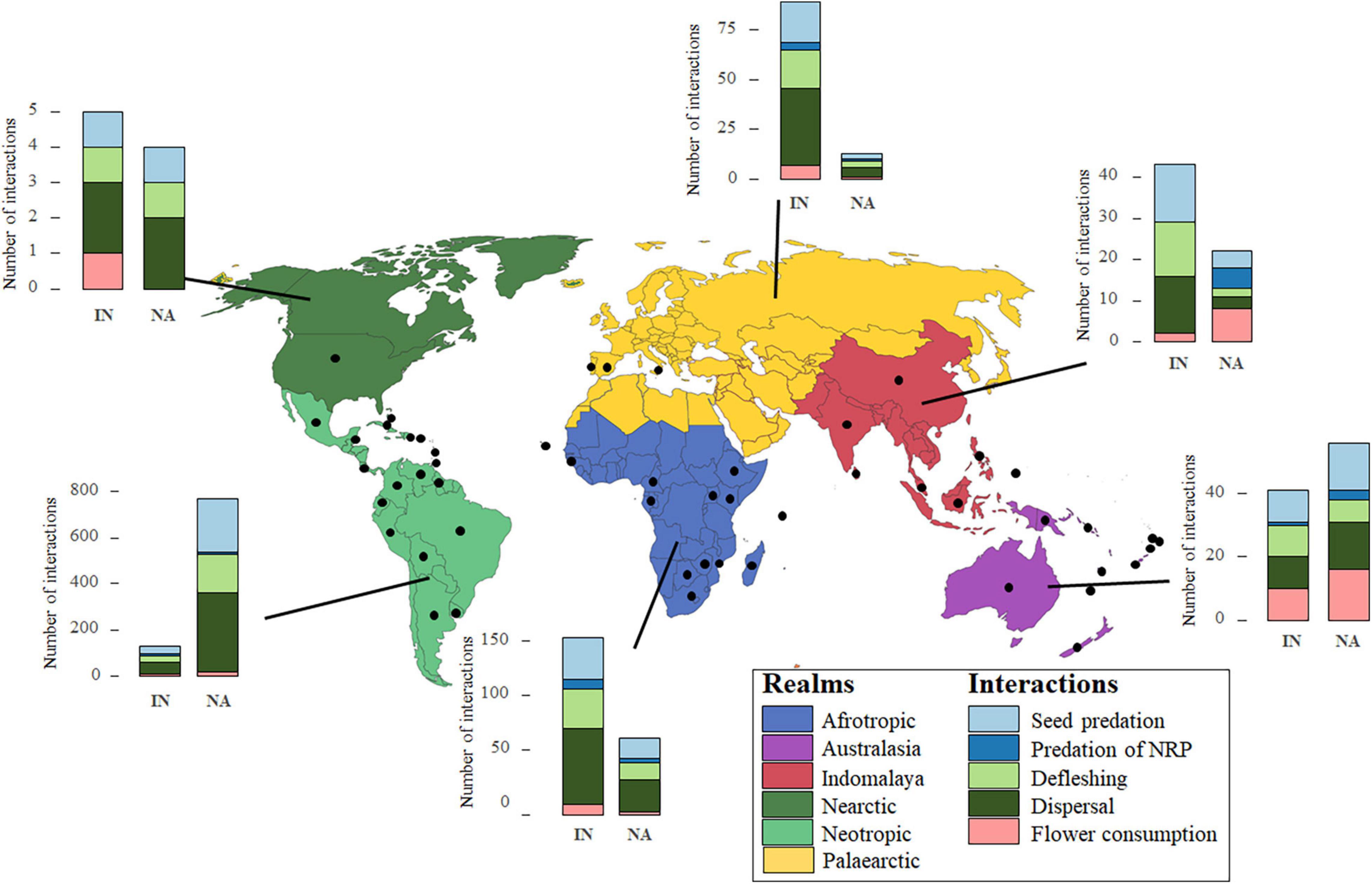
Figure 1. Number of interactions between parrots and palms across the main biogeographic realms of the world (Olson et al., 2001), considering whether the palm species was in its native (NA) or introduced (IN) range. Black points show the country of parrot-palm interactions. NRP: non-reproductive parts (bark, leaves, etc.).
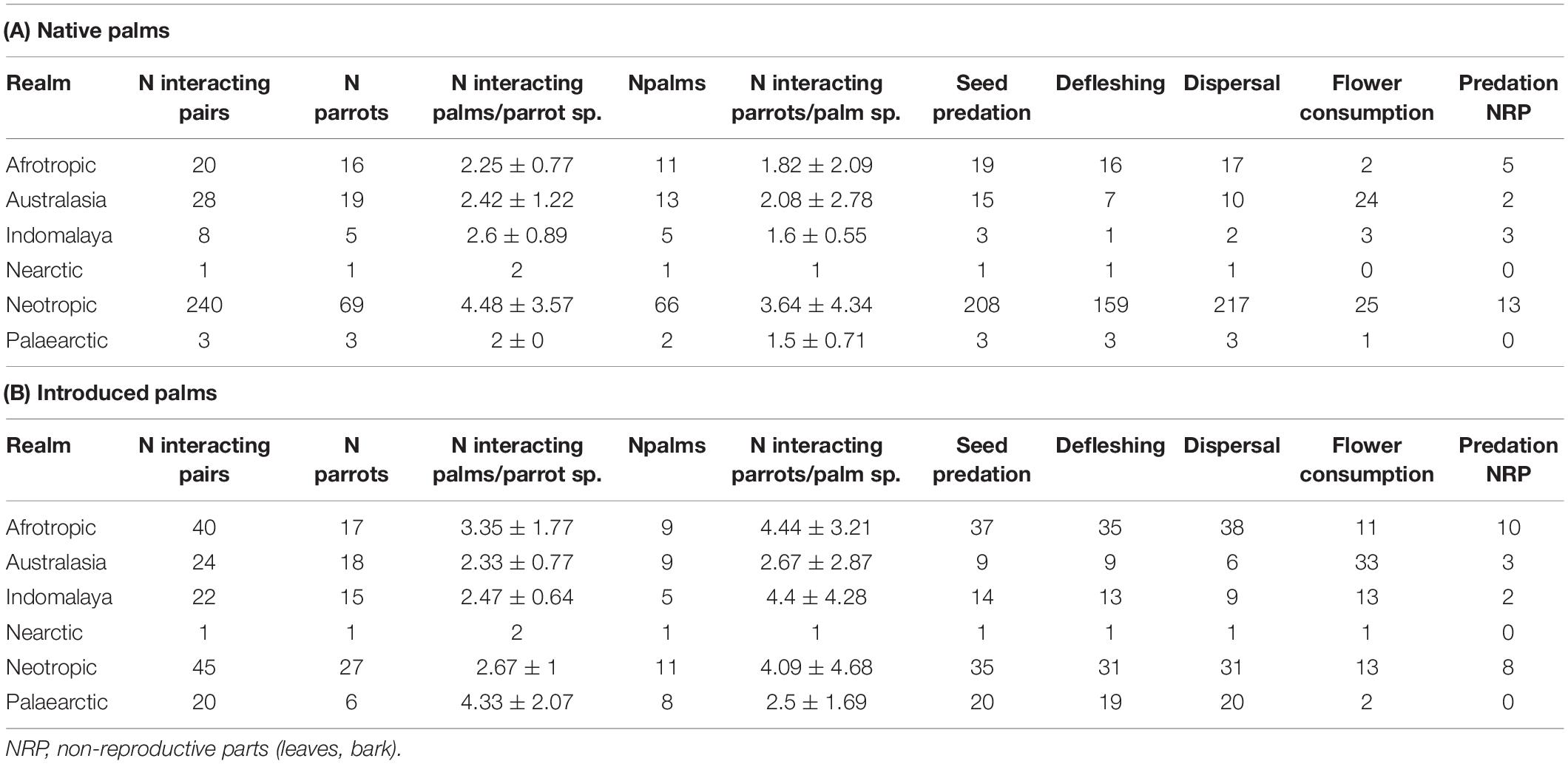
Table 2. Number of parrot and palm species (N parrots and N palms, respectively), mean (±SD) number of palms involved in interactions per parrot species and vice versa (N interacting palms/parrot sp. and N interacting parrots/palm sp. respectively) and total number of interactions recorded across realms, considering whether the palm species were native (A) or introduced (B).
The number of interactions significantly differed among realms (χ2 = 145.20, df = 5, p < 0.0001) but not between native and introduced palms (χ2 = 3.32, df = 1, p = 0.0683; Figure 1 and Supplementary Table 2). As predicted, parrot-palm interactions were more numerous in the Neotropic (comparison of the number of interactions per realm: native palms: Neotropic > Afrotropic = Australasia > Indomalaya = Nearctic = Palaearctic; introduced palms: Neotropic > Afrotropic = Australasia = Indomalaya > Palaearctic > Nearctic; Supplementary Table 3). However, the number of parrots interacting with each palm species was similar among realms and for native and introduced palms (χ2 = 7.79, df = 5, p = 0.1679, and χ2 = 3.74, df = 1, p = 0.0531, respectively; Supplementary Table 4), whereas the number of palms interacting with each parrot species did not differ in their association with the palm range (χ2 = 2.68, df = 1, p = 0.1019) but rather with the realms (χ2 = 17.25, df = 5, p = 0.0041; Supplementary Table 5). Thus, partially following our predictions, more palm species interact with each parrot species in the Neotropic while the pattern for the number of parrots interacting with each palm species was more heterogeneous (Table 2 and Figure 2).
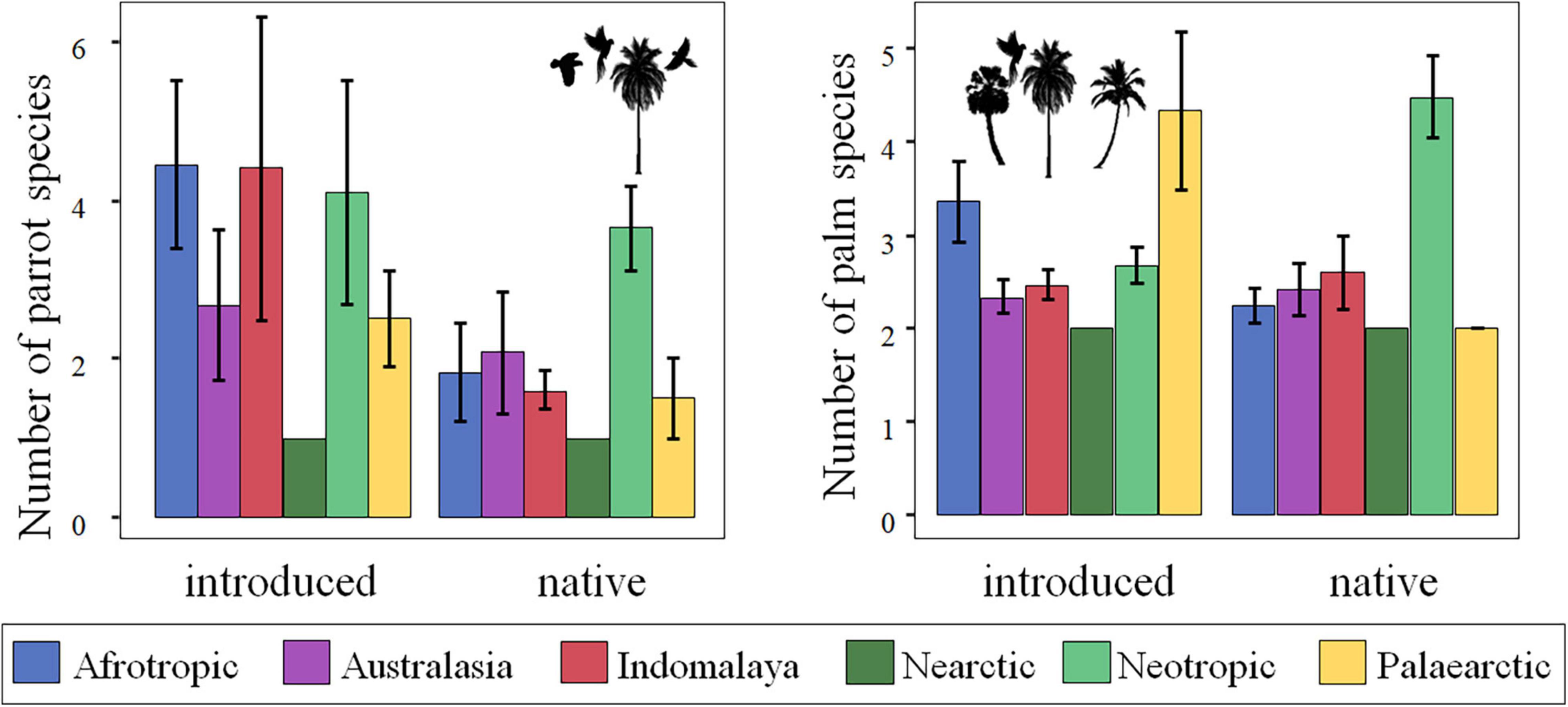
Figure 2. Mean number of parrots (±95% confidence interval) interacting with palms and number of palms interacting with parrots across the main biogeographic realms of the world, considering whether the palm species was in its native or introduced range.
Seed Predation and Defleshing
Seed predation and defleshing totaled 660 interactions and were mainly recorded in the Neotropic, in native palms (Figure 1 and Table 2). Despite differences between native and introduced palms (estimate for native palms: −0.25, SE = 0.10, χ2 = 7.14, df = 1, p = 0.0076), the proportion of ripe fruits predated were not affected by the parrot or palm fruit size (Supplementary Tables 6, 7). The ripeness of defleshed fruits did not show any significant pattern (Supplementary Tables 6, 7).
After controlling for phylogenetic relationships among species and differences among realms and for palms in their native and introduced ranges, we found a general positive relationship between palm fruit size and parrot body size (Supplementary Tables 8, 9), so that large-sized parrots predate and deflesh more large than small palm fruits while smaller parrots do the same with palms with smaller fruits (seed predation: estimate for parrot body size: 0.13, SE = 0.02, χ2 = 47.65, df = 1, p < 0.0001; defleshing: estimate for parrot body size: 0.07, SE = 0.02, χ2 = 13.60, df = 1, p = 0.0002; Figure 3 and Supplementary Figures 4, 5). Although there was no significant relationship between fruit size and the number of parrots eating or defleshing them (Supplementary Tables 10, 11 and Supplementary Figures 6, 7), only a few, large-sized parrot species were observed preying on palm species with the largest fruits (Figure 4). For instance, the mean body mass (±SD) of parrots preying on palms with fruits smaller or larger than 4 cm (the threshold to consider a fruit as megafaunal; Guimarães et al., 2008) was 567.49 g (±500.30 g) and 333.11 g (±347.46 g), respectively; Kruskal–Wallis test: p < 0.0001]. Regarding the number of palms with which each parrot species interacted, results show that, partially coinciding with our predictions, large parrots prey upon more palm species than small-sized parrots (seed predation: estimate for parrot body size: 0.23, SE = 0.10, χ2 = 4.95, df = 1, p = 0.0261; Supplementary Tables 12, 13, Figure 5, and Supplementary Figure 8). There was no relationship between parrot body mass and the number of palm species defleshed (Supplementary Tables 12, 13, Figure 5, and Supplementary Figure 9).
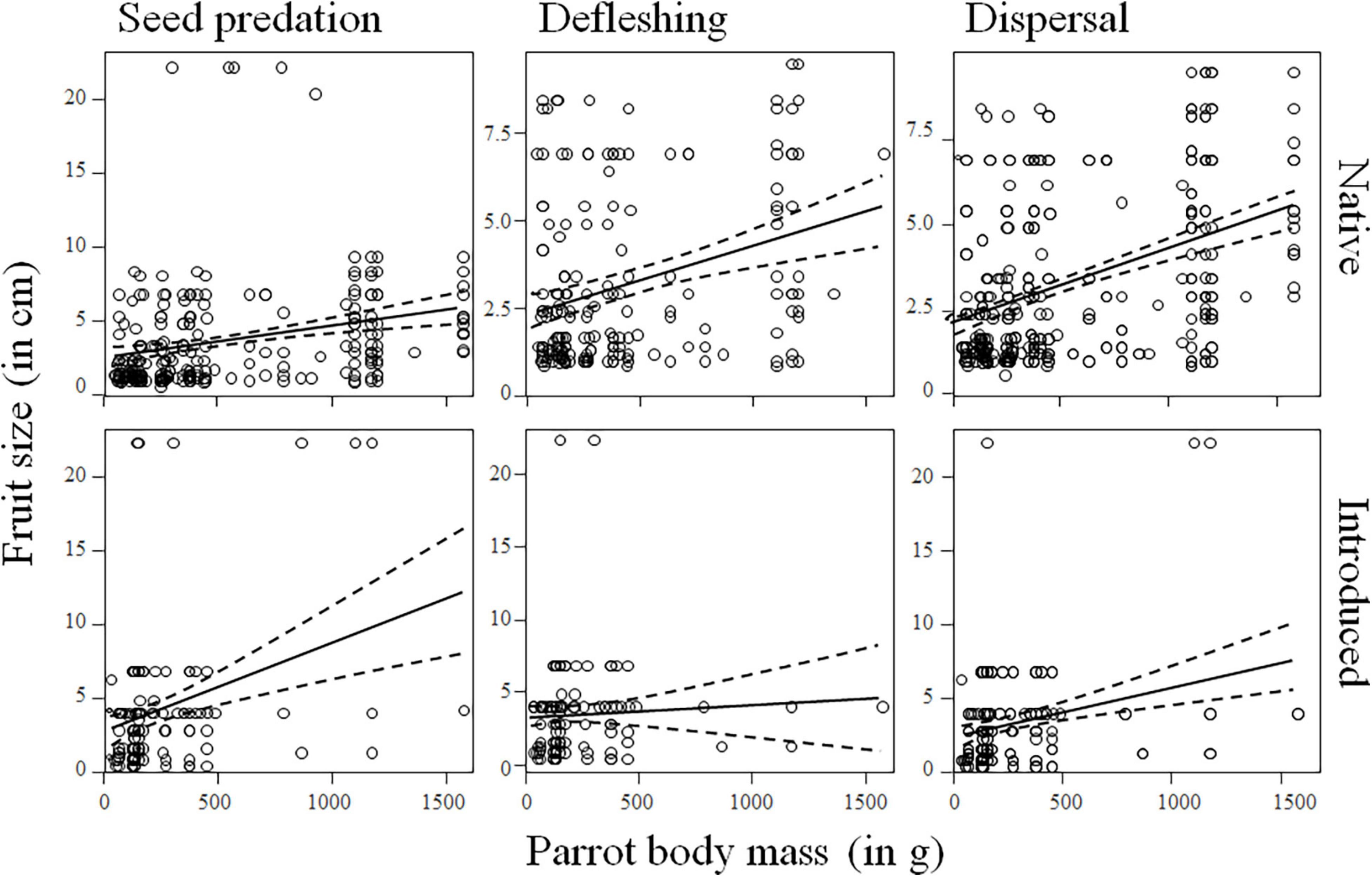
Figure 3. Relationship between palm fruit size (measured as fruit length, in cm) and the size of the parrot (measured as body mass, in g) predating, defleshing, and dispersing them. See Supplementary Figures 2, 3, and 4 for more detailed plots within each realm considered. The solid line represents the general trend across regions (dashed line: 95% confidence interval).
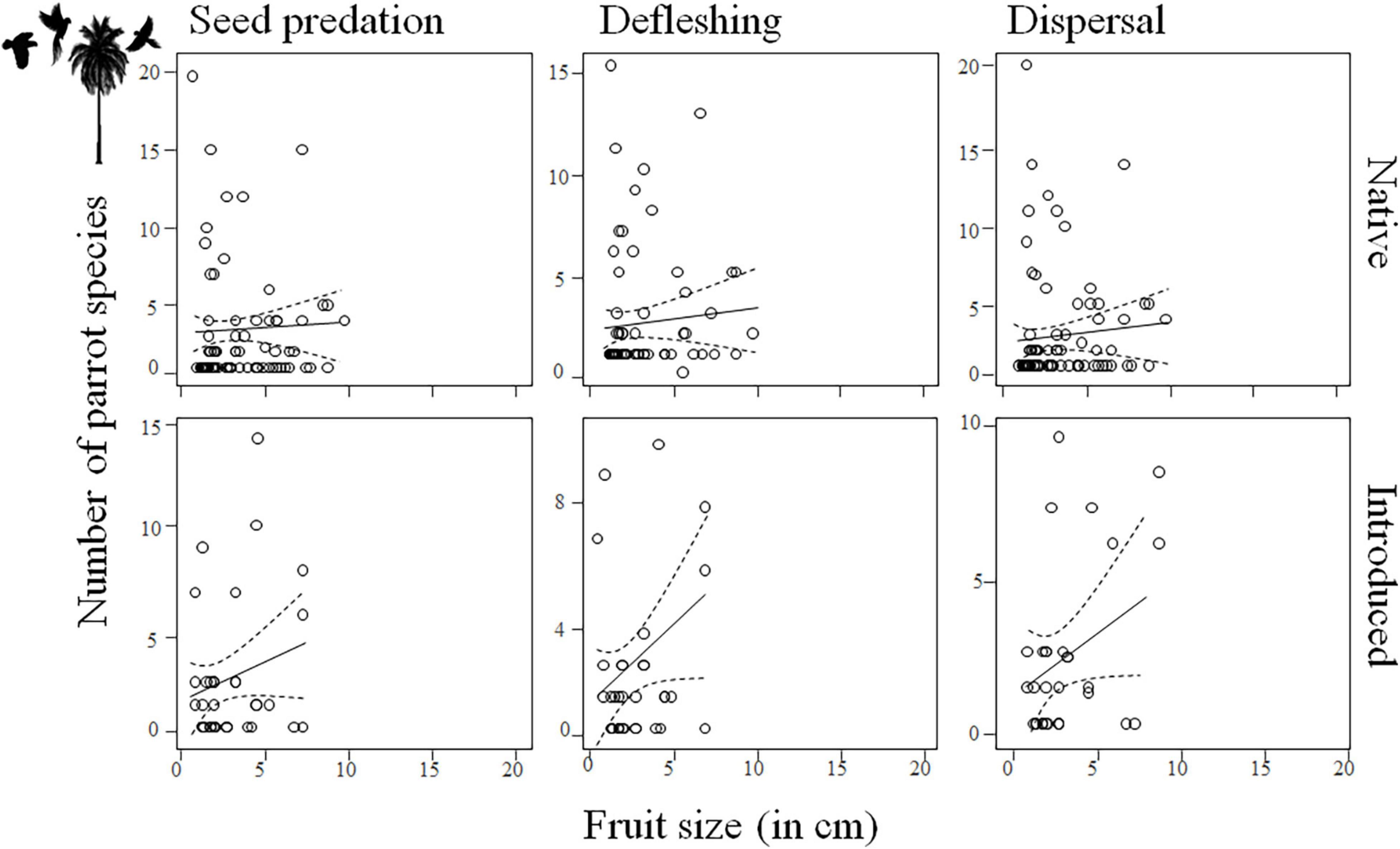
Figure 4. Number of parrot species interacting with palms (i.e., seed predation, defleshing, and dispersal) differing in fruit size (measured as fruit length, in cm) across six main realms of the world and in areas where palms were native and introduced.
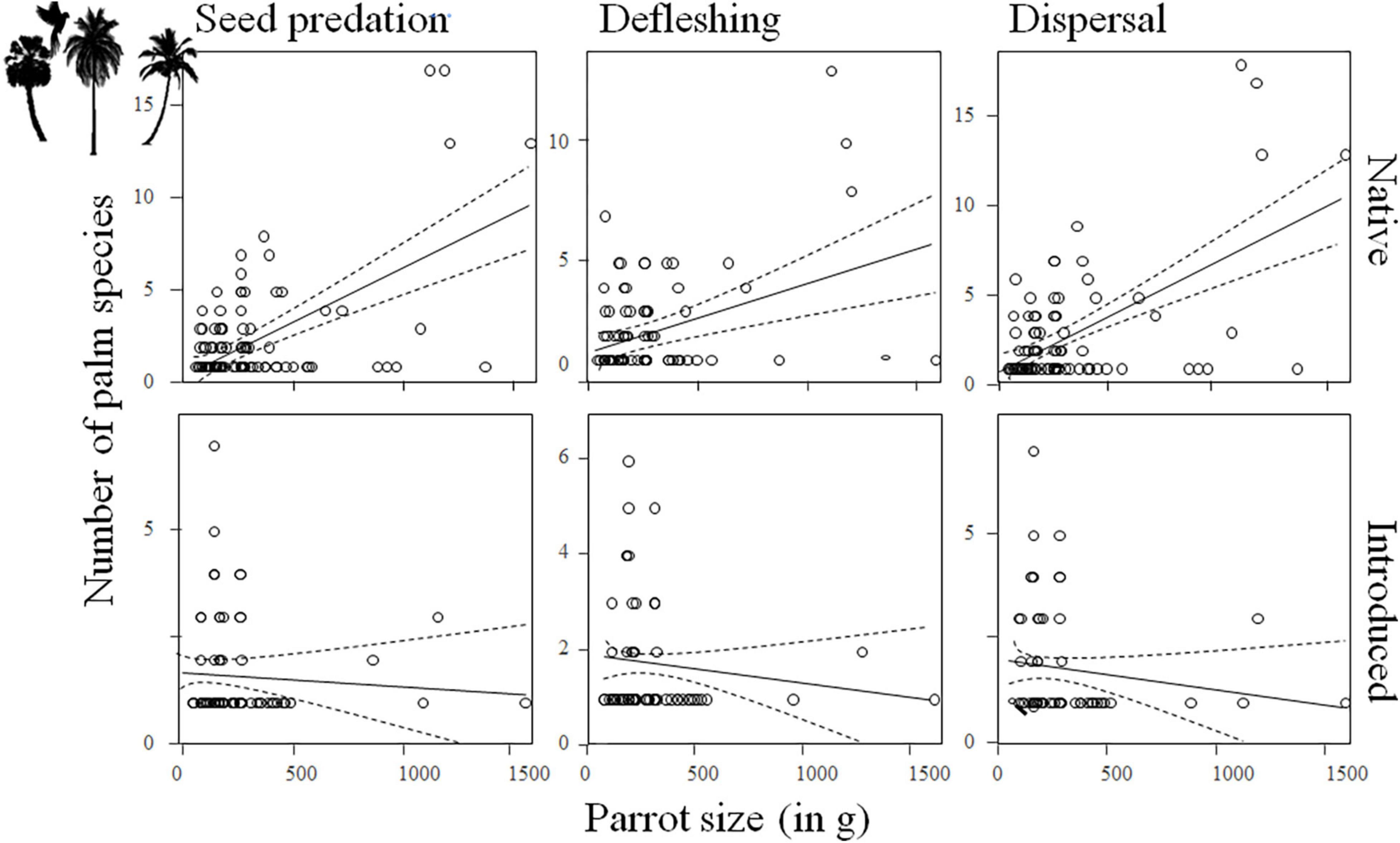
Figure 5. Number of palm species interacting with parrots (i.e., seed predation, defleshing, and dispersal) differing in size (measured as body mass) across the main biogeographic realms of the world, considering whether the palm species was in its native or introduced range.
Dispersal
Our dataset included 176 specific parrot-palm interactions corresponding to seed dispersal. However, when considering previous data from the same parrot species dispersing fruits of similar size after preying on or defleshing them, interactions involving parrots dispersing palm fruits or seeds increased to 355. Like predation, parrots tend to disperse unripe fruit of native palms more often than those of introduced palms (estimate for the proportion of ripe fruits dispersed in the native area of palms: −0.26, SE = 0.07, χ2 = 12.77, df = 1, p = 0.0004; Supplementary Tables 6, 7).
Despite the larger size of fruits in the introduced range of palms (estimate for native range: −0.07, SE = 0.04; χ2 = 3.09, df = 1, p = 0.0787) and after controlling for the phylogenetic relationship among species and potential differences among realms where dispersal events were recorded, we found a positive relationship between palm fruit and parrot size, so that large-sized parrots tend to disperse larger fruits than small-sized parrots (estimate for parrot size: 0.12, SE = 0.02, χ2 = 53.45, df = 1, p < 0.0001; Figure 3, Supplementary Figure 10, and Supplementary Tables 8, 9). Fruit size, however, did not affect the number of parrot species dispersing each palm species (Supplementary Tables 10, 11 and Figure 4), with some parrot species being able to disperse even palms with the largest fruits. We also found that large parrots dispersed more palm species than small-sized parrots in the native range of palms, but not when palms were introduced (estimate for parrot size: 0.23, SE = 0.11, χ2 = 4.01, df = 1, p = 0.0454; Supplementary Tables 12, 13, Figure 5, and Supplementary Figure 11).
During our fieldwork, we recorded 4,983 dispersal distances involving 32 parrot species and 29 palm species (20 palm species classified as native and 10 palm species classified as introduced in the area where the observation was made). Using raw data of exact and minimum dispersal distances, we obtained a median dispersal distance of 41 m (range: 1–1620 m). However, as most distances were right-censored (73%), estimated mean dispersal distances obtained using Kaplan–Meier Survival Curves were larger and rather variable among palm species (range of mean dispersal distances for the different palm species: 9–250 m; Table 3). In general, larger palm fruits were dispersed by larger parrot species (estimate for parrot size: 0.15, SE = 0.07, χ2 = 4.39, df = 1, p = 0.0362; Figure 6), although the variability was very high, and small-sized parrots were also able to move fruits up to 7 cm in length. Moreover, independent of whether palms were in their native or introduced range (95% credible interval: −0.33 to 0.27), larger parrots also dispersed larger fruits at greater distances than smaller ones (MCMCglmm, interaction parrot size*fruit size: posterior mean: −0.21, 95% credible interval: −0.35 to −0.10). The inclusion of the palm and parrot species in models allowed us to estimate how repeatable dispersal distances were for each group, indicating that repeatability for palms was high (r = 0.89, 95% credible interval: 0.68–0.96) but negligible for parrot species (r = 0.03, 95% credible interval: 0–0.24).
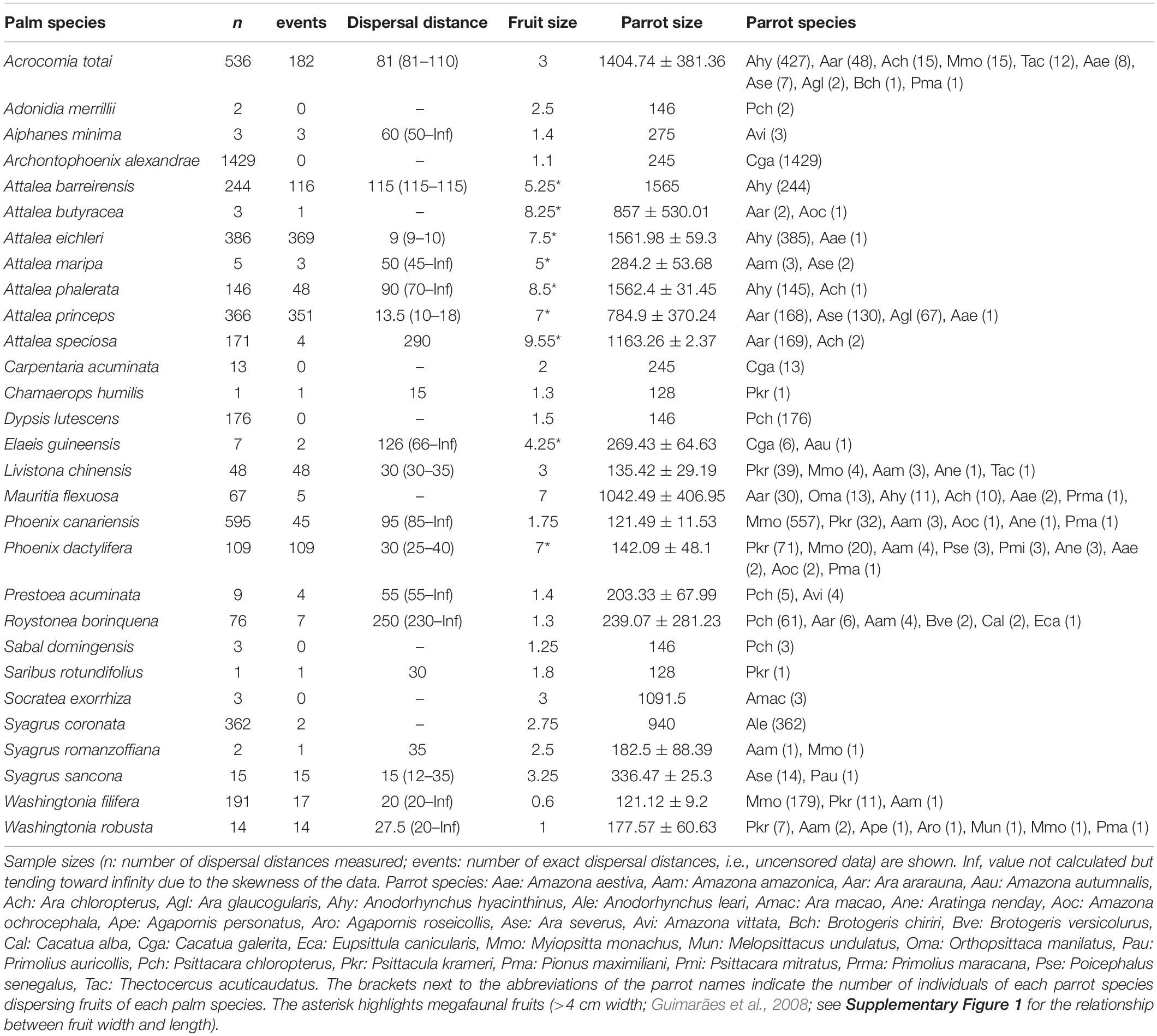
Table 3. Median distances (and 95% credible intervals) at which parrots of different sizes (mean body mass ± SD of individuals dispersing each palm species; in g) can disperse different palm species varying in their fruit size (estimated as fruit length; in cm).
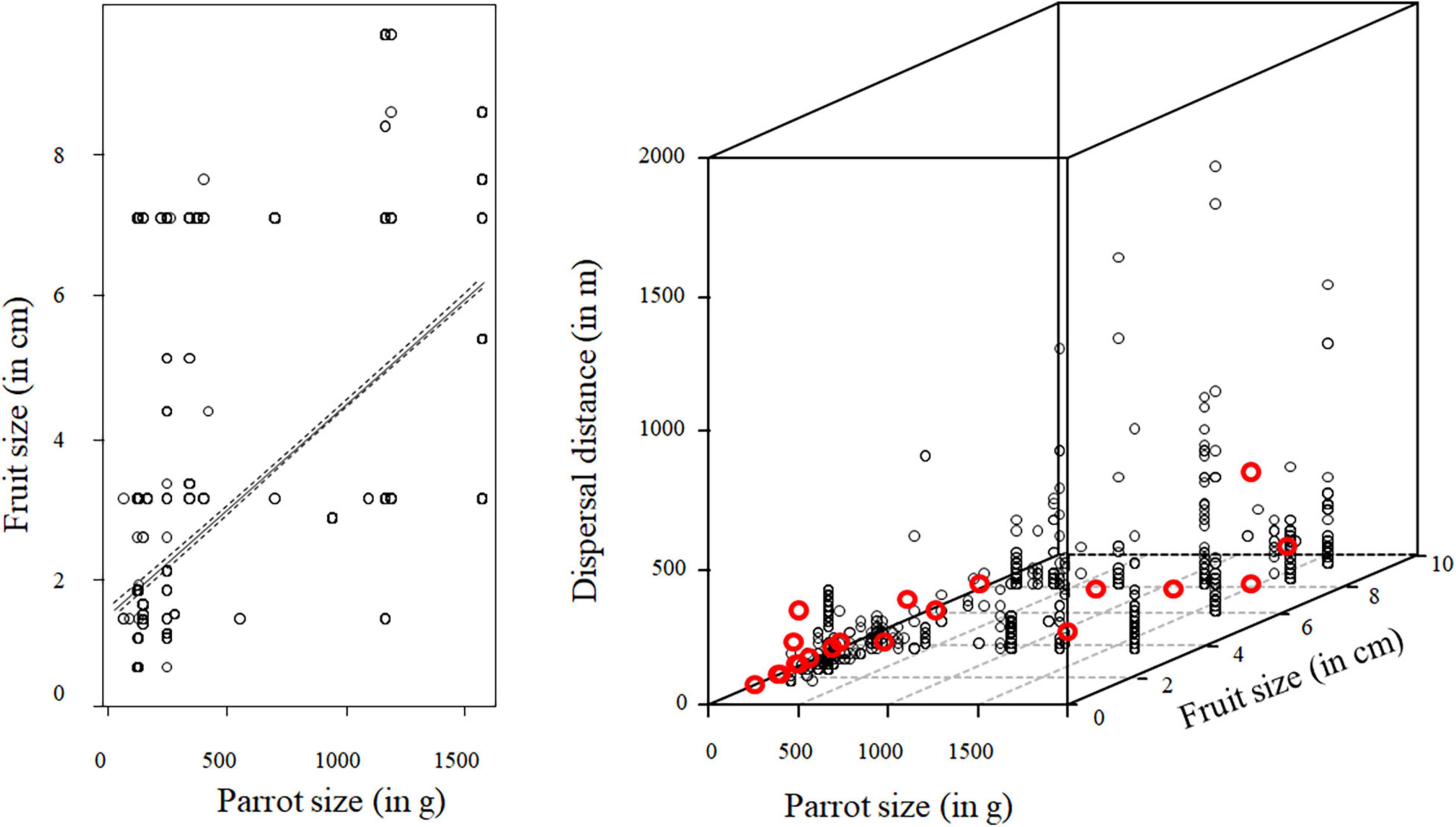
Figure 6. Relationship between the size of palm fruits (in cm) dispersed by parrots and their size (in g), and how these variables affect observed dispersal distances. Red points show the relationship among the mean dispersal distances estimated for each palm species considering the right-censored nature of data (through Kaplan Meier Survival Curves), the size of the palm fruits, and the mean size of parrots moving them (for details, see Table 3).
Discussion
Parrots have traditionally been considered as seed predators and thus plant antagonists (Toft and Wright, 2015). However, evidence has been accumulating to suggest that this role may ultimately represent a mutualistic relationship (Montesinos-Navarro et al., 2017) as, typically, a relevant proportion of seeds are viable and successfully recruited following parrot dispersal events (Blanco et al., 2016, 2020, 2021; Tella et al., 2016, 2020; Speziale et al., 2018; Bravo et al., 2020). After performing the most comprehensive compilation on parrot-palm interactions on a global scale, we show how interactions between these taxonomic groups rarely involve pure antagonistic relationships. Conversely, parrots develop mainly mutualistic interactions with palms across the world, especially when, at the same time they eat their fruits and seeds, they also disperse them. Although the overall generalist diet of parrots suggests no specificity in these interactions (but see Baños-Villalba et al., 2017 and Tella et al., 2020 for some specific macaw-palm species interactions), we found a positive relationship between fruit and parrot size (i.e., large-sized parrots predate, deflesh, and disperse larger fruits than small-sized parrots) after controlling for phylogenies. This pattern suggests that some interactions can be constrained because of morphological matching between parrots and palms, while others are favored. However, the large variability around this relationship is likely due to the flexibility in the feeding strategies of parrots, so even small species can disperse rather large fruits and vice-versa.
Although the most widely assumed interaction among parrots and palms has been seed predation (where parrots crush palm fruits and seeds with the bill), parrots can feed on fruits using strategies that may have costs but also benefits for palms (Figure 7). In most cases, parrots only consume the pulp of the fruits, without consuming the seeds (Baños-Villalba et al., 2017; Luna et al., 2018), or consume the seeds but leave a number of them undamaged and viable (Tella et al., 2020). There is increasing evidence on the viability of seeds dispersed by parrots (Blanco et al., 2016; Baños-Villalba et al., 2017; Tella et al., 2020), and even on the enhanced germination of seeds partially consumed by them (Tella et al., 2016; Speziale et al., 2018). In general, larger parrots deflesh larger fruits than smaller ones, perhaps due to the difficulty of handling small fruits while the opposite is true for small-sized parrots or simply because the beak of all parrot species is prepared to obtain pulp fragments regardless of their size. Interestingly, more parrot species consume the pulp of palms with larger fruits, maybe because the smaller ones easily ingested as a whole. Parrots are social species that usually feed in flocks. Intraspecific interactions, combined with predation fear (Howe, 1979; Laundré et al., 2010), may cause some feeding individuals to fly away to finish eating on perches located at variable distances. On these occasions, parrots carry in their feet or beaks the fruits or seeds they were eating, dispersing them by stomatochory (e.g., Sazima, 2008; Baños-Villalba et al., 2017; Blanco et al., 2019; Tella et al., 2020; Silva et al., 2021). Stomatochory allows the transportation of larger fruits than endozoochory (Blanco et al., 2016). However, there is still a constraint in the size of fruits moved (i.e., the weight of the fruit, which is related to its size), and larger parrots disperse by stomatochory larger fruits than small-sized parrots. Although more research is needed, previous work has shown that the proportion of palm fruits picked from the mother palm and dispersed by some macaw species ranges among palm species from 5% (Acrocomia totai) to 13% (Attalea phalerata) and 100% of the fruits (Attalea barreirensis, Attalea eichleri, and Acrocomia aculeata) (Tella et al., 2020; Silva et al., 2021). For smaller parrot species, these proportions are also highly variable (Sist and Puig, 1987). Moreover, parrots can accidentally or, as observed for this and other plant taxa, purposely discard a large proportion of undamaged palm fruits (11–75%) while feeding at perches distant from the mother plant (Tella et al., 2020) but also under the mother tree (Sebastián-González et al., 2019). Once on the ground, fruits can become available to other animals (e.g., insects, fish, reptiles, or mammals), which may disperse them through secondary dispersal (Blanco et al., 2019; Mittelman et al., 2021). Otherwise, if fruits remain under the mother tree or conspecifics, they can experience a high mortality rate (Janzen-Connell effect; Song et al., 2021) through parasitization by invertebrates, mainly bruchines (Fragoso et al., 2003; Dracxler et al., 2011). The pulp of palm fruits has adaptive functions such as defense against seed predators (invertebrates) and pathogens (Silvius and Fragoso, 2002; Pereira et al., 2014), so undamaged fruits may last longer under the mother tree and increase their chances of being dispersed. However, some studies show that palm fruits with injuries in the epicarp can be easily colonized by decomposing fungi, which reduces the selection of the egg-laying site by insects (Pereira et al., 2014). Moreover, defleshing facilitates germination (Nascimento et al., 2019), with dispersers removing the pulp of the fruits being more effective than those regurgitating them (Loayza and Knight, 2010; de Barros Leite et al., 2012). Thus, if defleshed fruits are moved by stomatochory far from the mother palm —under which the risk of invertebrate infection is the highest— or by other animal species, they have a higher probability of survival and germination (Fragoso et al., 2003; Comita et al., 2014; Song et al., 2021). In this sense, the large numbers of viable seeds and saplings below a high proportion of trees (31–100%) used by parrots as perching sites at variable distances from mother palms confirm their role not only as legitimate but also as frequent palm seed dispersers (Sist and Puig, 1987; Baños-Villalba et al., 2017; Tella et al., 2020). Although poorly understood, parrots could, at least theoretically, swallow the whole palm fruit directly, dispersing palm seeds by endozoochory. A few studies have shown that parrots can defecate and/or regurgitate viable seeds of different plant species including palms (Lieberman and Lieberman, 1986; Henderson, 2002; Blanco et al., 2016, 2020; Buitron-Jurado and Sanz, 2016; Bravo et al., 2020; Buitron-Jurado in Litt, unpublished). However, compared to stomatochory, endozoocory is a rather infrequent strategy (only 30 out of 11,274 records of parrots dispersing different plant species corresponded to endozoochory, and none of them were for palms; authors’ unpublished results) and deserves more research to understand its relevance for palm ecology.
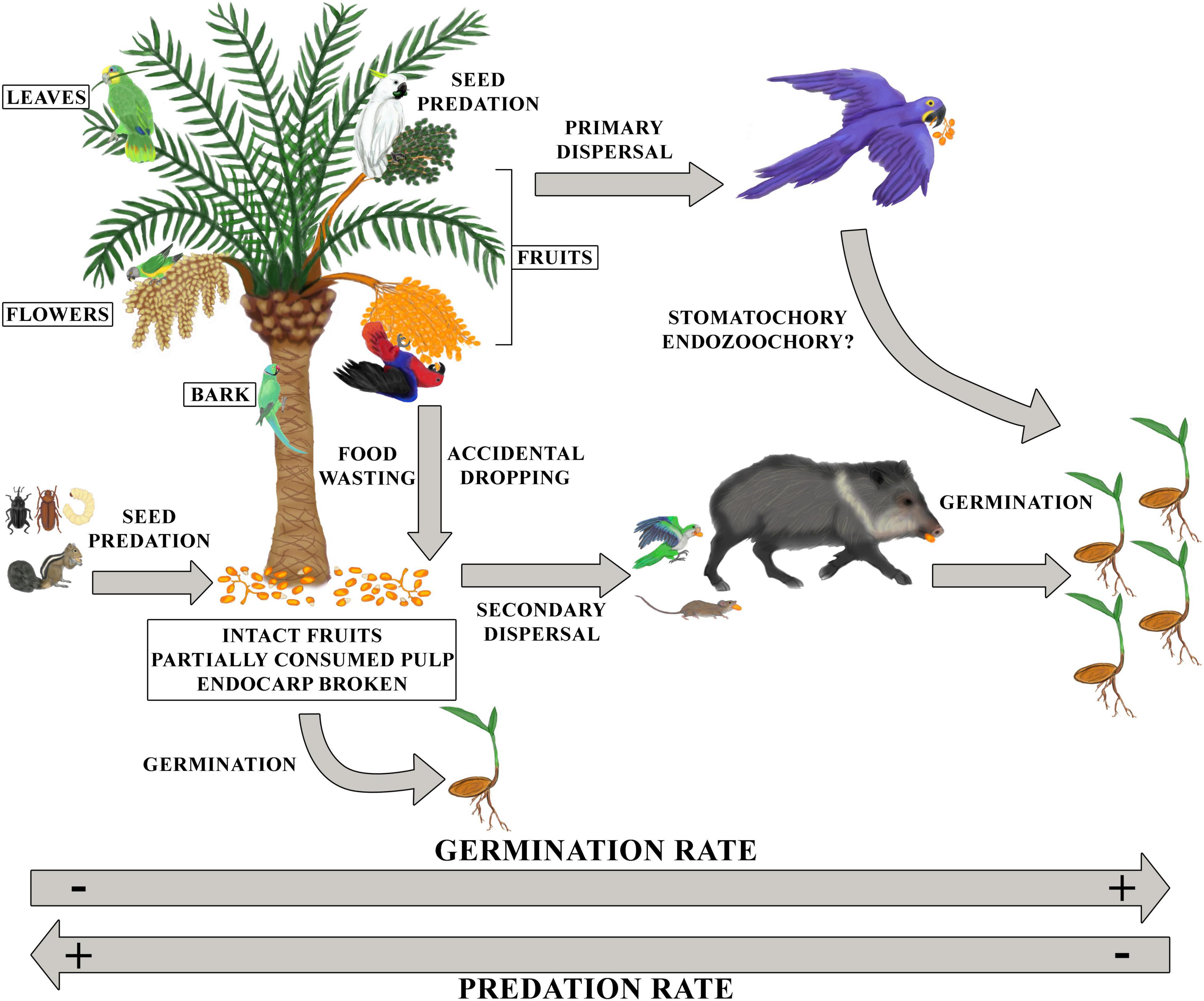
Figure 7. Although most known interactions between parrots and palms have been seed predation and consumption of vegetative parts such as leaves or bark (antagonistic interactions), parrots can also feed using strategies that favor palms (mutualistic interactions). When parrots consume only the pulp of the fruits (defleshing), without consuming the seeds or leaving some undamaged and viable, they can favor the germination of seeds that fall to the ground accidentally or when the individual wastes them (food wasting). In many cases, parrots carry the fruits or seed with their feet or beaks to consume them on perches, dispersing them (stomatochory) and favoring germination in areas far from the mother plant, where predation rates are lower. In addition, these fruits and seeds that fall to the ground, either under the mother palm or under a perch, may then be secondarily dispersed by other species. Although it is possible that parrots also swallow palm fruits and disperse them by endozoochory, this dispersal mechanism has not been described for parrots and palms. Finally, parrots may also consume palm flowers, possibly contributing to pollination. Illustrations by Dailos Hernández-Brito.
Parrots can disperse seeds at large distances (i.e., >100 m; Cain et al., 2000), especially large fruits commonly categorized as megafaunal (i.e., >4 cm width; Guimarães et al., 2008). Obtaining reliable dispersal distances is challenging, though many studies have found different approaches to directly or indirectly estimate them. The maximum dispersal distance obtained for palms was provided by Jansen et al. (2012), who recorded the movement of Astrocaryum standleyanum palm seeds over distances greater than 100 m from the source after secondary dispersal by several individuals of agoutis Dasyprocta punctata stealing each other’s seeds after burying them. However, large rodents (Dasyprocta sp. and Cuniculus sp.) usually hide or bury seeds at distances rarely exceeding 20 m from the mother tree (Franco-Quimbay and Rojas Robles, 2015; Mendieta-Aguilar et al., 2015). These seeds can be dug up and subsequently consumed by the same rodents (Janzen, 1970) or by other species such as peccaries (family Tayassuidae; Kiltie, 1981; Silman et al., 2003), or secondarily dispersed by con- or heterospecifics (Jansen et al., 2012). Our data, representing primary dispersal distances, surpass these values for some palm species (range of median dispersal distances: 9–250 m), in particular, large-fruited ones (up to at least 1,620 m for Anodorhynchus hyacinthinus dispersing Attalea barreirensis in Brazil). Importantly, dispersal distances vary among parrot species, maybe depending on the dispersed palm species, but are similar (e.g., repeatable) at the palm level. This suggests that although parrot-palm interactions are not species-specific (Blanco et al., 2019), with some exceptions like bush-layer Attalea palms (Tella et al., 2020), the ability to disperse certain palm fruits —mainly the largest ones— is restricted to a few, rather large-sized parrot species. Parrots are known for their flying abilities (i.e., they can make long-distance daily, seasonal, altitudinal, and regional movements by tracking their food resources; Renton et al., 2015) that allow them to perform movements far superior to those of large terrestrial mammals (from 10 km for medium-sized species to several tens of kilometers per day, in the case of large macaws; Adamek, 2011; Brightsmith et al., 2021), making them long-distance dispersers of the plants they use (e.g., Tella et al., 2015, 2019; Baños-Villalba et al., 2017; Blanco et al., 2021; present results). Thus, although extinct megafauna might have played a pivotal role in the dispersal of palms with large fruits, these species with presumably anachronic fruits —the so-called “megafaunal” fruits— are in fact dispersed at rather large distances by extant mammals and birds (Jansen et al., 2012; Blanco et al., 2019), such as parrots as evidenced here. Importantly, the fact that large parrots prey upon and disperse more palm species than small-sized parrots supports the primary role that this avian order can play in the structure, organization, and functioning of tropical ecosystems (Blanco et al., 2015, 2018).
Palms are mainly insect-pollinated (mainly by beetles, bees, and flies), even though they can also be pollinated by other invertebrates (e.g., crabs) and by mammals (e.g., bats and marsupials) (Barfod et al., 2011). Our data show several cases of parrots consuming palm flowers, suggesting a potential role as palm pollinators. Data on parrots consuming palm flowers are neither numerous nor detailed enough to adequately understand their significance for pollination. However, parrot feeding behavior (i.e., individuals feeding on palm flowers and moving from inflorescence to inflorescence and between different palms) may facilitate pollination through the transportation of pollen in their feathers and beaks. Indeed, lories and lorikeets have been recognized as important pollinators in Australian and Southeast Asian rainforests (Brown and Hopkins, 1995; Gelis, 2011), while species of the genera Pionites, Brotogeris, Psittacara, and Enicognathus are effective pollinators of Neotropical phanerogams and gymnosperms (Maués and Venturieri, 1997; Vicentini and Fischer, 1999; Gleiser et al., 2017). Importantly, while less frequent visitors to palm flowers than insects, parrots may make a unique contribution to palm population structure because of their capacity to move significant pollen loads per individual (Bezemer et al., 2016) and at large distances (Southerton et al., 2004; Byrne et al., 2007; Bezemer et al., 2016). This line of research deserves more attention to properly understand the relevance of parrots as palm pollinators.
Although not very frequent, parrots can develop entirely antagonistic interactions with palms when consuming their non-reproductive structures, causing fitness costs in repairing tissues (Blanco et al., 2015) and, sometimes, the death of individuals (Costion et al., 2013; Brightsmith and Cáceres, 2017; Gilardi in Litt., unpublished; authors unpublished). Palms are known to bioaccumulate sodium (Arnason et al., 1984; Brightsmith and Cáceres, 2017), a vital nutrient for vertebrates (Randall et al., 1997). Animals are regularly confronted with sodium limitations, which are usually overcome through geophagy (i.e., the deliberate consumption of clay or soil by an animal; Diamond et al., 1999). In the case of parrots, different studies show not only geophagy (Emmons and Stark, 1979; Brightsmith and Aramburu, 2004; Brightsmith and Villalobos, 2011) but also palm consumption (mainly leaves and bark; Brightsmith and Cáceres, 2017) as a source of sodium. Parrots are known consumers of plant parts rich in toxic chemicals (Gilardi and Toft, 2012), whose detoxification promotes a negative sodium balance in animals (Pehrson, 1983; Reichardt et al., 1984; Freeland et al., 1985; Jakubas et al., 1995). Therefore, parrots may have particularly high sodium requirements, which, in part, could be met by consuming the vegetative parts of palms.
Palms have been widely introduced elsewhere for ornamental purposes, food, and other uses (Byg and Balslev, 2001; Campos and Ehringhaus, 2003; Byg et al., 2006; Sosnowska and Balslev, 2009; Martins et al., 2014), resulting in the establishment and spread of many palm species into new areas (Fehr et al., 2020; Bello et al., 2021). We found that, in general, parrots interact similarly with native and introduced palms, maybe due to the generalist habits of these avian species (Christianini, 2006). Moreover, the human-induced movement of several parrot species out of their native ranges (Calzada Preston and Pruett-Jones, 2021) has allowed parrot-palm interactions for palm species introduced in the Palaearctic and Nearctic realms. Although more research is needed, these interactions may facilitate palm invasion through the dispersal of fruits, not only across human-modified but also in more natural areas across the world.
As predicted, most parrot-palm interactions have been recorded in the Neotropic, where the richness of both taxa is very high and, more importantly, the mean number of parrot and palm species overlapping their distributions is the highest (see Supplementary Figure 12). However, the scientific literature on palm ecology is also skewed toward studies performed in Neotropical countries (Supplementary Figure 12) so this result should be more deeply investigated. Muñoz et al. (2019) recently highlighted that knowledge on palm-frugivore interactions in this realm as well as in the Afrotropic is largely incomplete. Part of this gap has been filled here, by focusing on understanding the interactions that one of the most diverse and abundant avian groups has with palms. Data for Africa, Australia, and Asia are still scarce, and more research should be carried out to fully understand the role of parrots in palm ecology in these areas.
Conclusion
Parrots are undisputed seed predators. However, as we have demonstrated in this study, they frequently develop mutualistic interactions, including effective seed dispersal, facilitation of secondary dispersal and, quite possibly, pollination. The distinction between disperser and predator is not always immediately evident, and the position of parrots on the antagonist-mutualist continuum may change depending on several factors (e.g., social behavior, predation fear, food availability, seasonality), which are expected to vary among the enormous range of ecosystems they inhabit. All of these aspects should be more deeply investigated to properly comprehend how palms would respond to habitat transformations, fragmentation, and defaunation. Meanwhile, the pervasiveness of parrot-palm mutualistic interactions should not be overlooked in studies of palm ecology and evolution. This would help us to better understand the history of this plant lineage and ensure its long-term conservation. In an era where human impacts are reducing overall plant regeneration by affecting processes early in the regeneration cycle, conserving pollination and seed dispersal, to a large extent by conserving the animal species that provide these ecological functions, should become a priority in forest conservation efforts globally.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
Ethical review and approval was not required for the animal study because ethical approval not request for observational data on foraging animal.
Author Contributions
MC, FH, PR-V, GB, DH-B, ES-G, and JT conceived the ideas for this study. MC, FH, PR-V, GB, DH-B, JD-L, and JT collected the data. FH compiled the data. MC analyzed the data. MC, FH, and JT wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by Severo Ochoa Program (SVP-2014-068732), Action COST “ParrotNet” (ES1304), and Loro Parque Fundación (PP-146-2018-1). ES-G was funded by a Ramón y Cajal contract (RYC-2019-027216I) from the Spanish Ministry of Science and Innovation.
Conflict of Interest
JD-L was employed by Endangered Conservation Consultancy.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We would like to acknowledge all naturalists and photographers as well as researchers who help us to identify and complete information on parrot-palm interactions, especially G. Buitrón-Jurado, J. Gilardi, L. Ortiz-Catedral, and Y. Takahatato for their kind comments and information shared. N. Cunha, M. Paniw, and R. Rodríguez helped with the phylogenetic analysis. S. Young revised the English and two reviewers and editor have contributed with very useful comments that have improved the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.790883/full#supplementary-material
Footnotes
- ^ https://wcsp.science.kew.org
- ^ https://ebird.org/
- ^ https://www.wikiaves.com
- ^ https://www.macaulaylibrary.org/
- ^ https://www.flickr.com/
- ^ https://www.instagram.com/
- ^ https://www.facebook.com/
References
Abedi-Lartey, M., Dechmann, D. K. N., Wikelski, M., Scharf, A. K., and Fahr, J. (2016). Long-distance seed dispersal by straw-coloured fruit bats varies by season and landscape. Global Ecol. Cons. 7, 12–24. doi: 10.1016/j.gecco.2016.03.005
Adamek, K. (2011). Temporal Variation in Space and Resource Use of Macaws in the Southeastern Peruvian Amazon. Ph.D. Thesis. Austin, TX: Texas University.
Adler, G. H., and Kestell, D. W. (1998). Fates of neotropical tree seeds influenced by Spiny Rats (Proechimys semispinosus). Biotropica 30, 677–681. doi: 10.1111/j.1744-7429.1998.tb00109.x
Andreazzi, C. S., Pires, A. S., and Fernandez, F. A. S. (2009). Mamíferos e palmeiras neotropicais: interações em paisagens fragmentadas. Oecol. Bras. 13, 554–574. doi: 10.4257/oeco.2009.1304.02
Arnason, J. T., Lambert, J. D., and Gale, J. (1984). Mineral cycling in a tropical palm forest. Plant Soil 79, 211–225. doi: 10.1007/bf02182343
Baños-Villalba, A., Blanco, G., Díaz-Luque, J. A., Dénes, F. V., Hiraldo, F., and Tella, J. L. (2017). Seed dispersal by macaws shapes the landscape of an Amazonian ecosystem. Sci. Rep. 7:7373. doi: 10.1038/s41598-017-07697-5
Barfod, A. S., Hagen, M., and Borchsenius, F. (2011). Twenty-five years of progress in understanding pollination mechanisms in palms (Arecaceae). Ann. Bot. 108, 1503–1516. doi: 10.1093/aob/mcr192
Bello, C., Cintra, A. L. P., Elisa Barreto, E., Vancine, M. H., Sobral-Souza, T., Graham, C. H., et al. (2021). Environmental niche and functional role similarity between invasive and native palms in the Atlantic Forest. Biol. Invas. 23, 741–754. doi: 10.1007/s10530-020-02400-8
Berkunsky, I., Quillfeldt, P., Brightsmith, D. J., Abbud, M. C., Aguilar, J. M. R. E., Alemán-Zelaya, U., et al. (2017). Current threats faced by neotropical parrot populations. Biol. Conserv. 214, 278–287.
Bezemer, N., Krauss, S. L., Phillips, R. D., Roberts, D. G., and Hopper, S. D. (2016). Paternity analysis reveals wide pollen dispersal and high multiple paternity in a small isolated population of the bird-pollinated Eucalyptus caesia (Myrtaceae). Heredity 117, 460–471. doi: 10.1038/hdy.2016.61
BirdLife International (2020). Handbook of the Birds of the World and BirdLife International Digital Checklist of the Birds of the World. Version 5. Available online at: http://datazone.birdlife.org/userfiles/file/Species/Taxonomy/HBW-BirdLife_Checklist_v5_Dec20.zip (accessed January 12, 2022).
Blach-Overgaard, A., Svenning, J. C., and Balslev, H. (2009). Climate change sensitivity of the African ivory nut palm, Hyphaene petersiana Klotzsch ex Mart. (Arecaceae) – a keystone species in SE Africa. IOP Conf. Ser. Earth Environ. Sci. 8:012014. doi: 10.1088/1755-1315/8/1/012014
Blach-Overgaard, A., Svenning, J. C., Dransfield, J., Greve, M., and Balslev, H. (2010). Determinants of palm species distributions across Africa: the relative roles of climate, non-climatic environmental factors, and spatial constraints. Ecography 33, 380–391.
Blanco, G., Bravo, C., Chamorro, D., Lovas-Kiss, A., Hiraldo, F., and Tella, J. L. (2020). Herb endozoochory by cockatoos: is “foliage the fruit”? Austr. Ecol. 45, 122–126. doi: 10.1111/aec.12835
Blanco, G., Bravo, C., Pacífico, E., Chamorro, D., Speziale, K., Lambertucci, S., et al. (2016). Internal seed dispersal by parrots: an overview of a neglected mutualism. PeerJ 4:e1688. doi: 10.7717/peerj.1688
Blanco, G., Hiraldo, F., Rojas, A., Dénes, F. V., and Tella, J. L. (2015). Parrots as key multilinkers in ecosystem structure and functioning. Ecol. Evol. 18, 4141–4160. doi: 10.1002/ece3.1663
Blanco, G., Hiraldo, G., and Tella, J. L. (2018). Ecological functions of parrots: an integrative perspective from plant life cycle to ecosystem functioning. Emu 118, 36–49. doi: 10.1080/01584197.2017.1387031
Blanco, G., Romero-Vidal, P., Carrete, M., Chamorro, D., Bravo, C., Hiraldo, F., et al. (2021). Burrowing parrots Cyanoliseus patagonus as long-distance seed dispersers of keystone Algarrobos, Genus Prosopis, in the Monte desert. Diversity 13:204. doi: 10.3390/d13050204
Blanco, G., Tella, J. L., Díaz-Luque, J. A., and Hiraldo, F. (2019). Multiple external seed dispersers challenge the megafaunal syndrome anachronism and the surrogate ecological function of livestock. Front. Ecol. Evol. 7:328. doi: 10.3389/fevo.2019.00328
Boll, T., Svenning, J. C., Vormisto, J., Normand, S., Grandez, C., and Balslev, H. (2005). Spatial distribution and environmental preferences of the piassaba palm Aphandra natalia (Arecaceae) along the Pastaza and Urituyacu rivers in Peru. For. Ecol. Manage. 213, 175–183. doi: 10.1016/j.foreco.2005.03.020
Bravo, C., Chamorro, D., Hiraldo, F., Speziale, K., Lambertucci, S. A., Tella, J. L., et al. (2020). Physiological dormancy broken by endozoochory: austral parakeets as legitimate dispersers of Calafate (Berberis microphylla) in the Patagonian Andes. J. Plant Ecol. 13, 538–544. doi: 10.1093/jpe/rtaa041
Brightsmith, D. J., and Aramburu, R. (2004). Avian geophagy and soil characteristics in southeastern Peru. Biotropica 36, 534–543. doi: 10.1111/j.1744-7429.2004.tb00348.x
Brightsmith, D. J., and Cáceres, A. (2017). Parrots consume sodium-rich palms in the sodium-deprived landscape of the Western Amazon Basin. Biotropica 49, 921–931. doi: 10.1111/btp.12479
Brightsmith, D. J., and Villalobos, E. (2011). Parrot behavior at a Peruvian clay lick. Wilson J. Ornithol. 123, 595–602. doi: 10.1676/09-109.1
Brightsmith, D. J., Boyd, J. D., Hobson, E. A., and Randel, C. J. (2021). Satellite telemetry reveals complex migratory movement patterns of two large macaw species in the western Amazon basin. Avian Cons. Ecol. 16:14.
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/rj-2017-066
Brown, E. D., and Hopkins, M. J. G. (1995). A test of pollinator specificity and morphological convergence between nectarivorous birds and rainforest tree flowers in New Guinea. Oecologia 103, 89–100. doi: 10.1007/BF00328429
Buitron-Jurado, G., and Sanz, V. (2016). Notes on the diet of endemic Red-eared parakeet Pyrrhura hoematotis and other Venezuelan montane parrots. Ardeola 63, 357–367. doi: 10.13157/arla.63.2.2016.sc2
Burgio, K. R., Davis, K. E., Dreiss, L. M., Cisneros, L. M., Klingbeil, B. T., Presley, S. J., et al. (2019). Phylogenetic supertree and functional trait database for all extant parrots. Data Brief 24:103882. doi: 10.1016/j.dib.2019.103882
Byg, A., and Balslev, H. (2001). Diversity and use of palms in Zahamena, eastern Madagascar. Biodiv. Cons. 10, 951–970.
Byg, A., Vormisto, J., and Balslev, H. (2006). Using the useful: characteristics of used palms in south-eastern Ecuador. Environ. Dev. Sustain. 8, 495–506. doi: 10.1007/s10668-006-9051-6
Byrne, M., Elliott, C. P., Yates, C. J., and Coates, D. J. (2007). Extensive pollen dispersal in a bird-pollinated shrub, Calothamnus quadrifidus, in a fragmented landscape. Mol. Ecol. 16, 1303–1314. doi: 10.1111/j.1365-294X.2006.03204.x
Cain, M. L., Milligan, B. G., and Strand, A. E. (2000). Long-distance seed dispersal in plant populations. Ann. Bot. 87, 1217–1227. doi: 10.2307/2656714
Callaghan, C. T., Nakagawa, S., and Cornwell, W. K. (2021). Global abundance estimates for 9,700 bird species. Proc. Natl. Acad. Sci. U.S.A. 118:e2023170118. doi: 10.1073/pnas.2023170118
Calzada Preston, C. E., and Pruett-Jones, S. (2021). The number and distribution of introduced and naturalized parrots. Diversity 13:412. doi: 10.3390/d13090412
Cámara-Leret, R., Faurby, S., Macía, M. J., Balslev, H., Göldel, B., Svenning, J. C., et al. (2017). Fundamental species traits explain provisioning services of tropical American palms. Nat. Plants 3:16220. doi: 10.1038/nplants.2016.220
Campos, M. T., and Ehringhaus, C. (2003). Plant virtues are in the eyes of the beholders: a comparison of known palm uses among indigenous and folk communities of Southwestern Amazonia. Econ. Bot. 57:324. doi: 10.1663/0013-0001(2003)057[0324:pvaite]2.0.co;2
Chapman, C., and Russo, S. (2007). “Primate seed dispersal. Linking behavioral ecology with forest community structure,” in Primates in Perspective, eds C. Campbell, A. Fuentes, K. Mackinnon, M. Panger, and S. Bearder (Oxford: Oxford University Press), 510–525.
Christianini, A. V. (2006). Fecundidade, dispersão e predação de sementes de Archontophoenix cunninghamiana H. Wendl. and Drude, uma palmeira invasora da Mata Atlântica. Rev. Brasil. Bot. 29, 587–594.
Comita, L. S., Queenborough, S. A., Murphy, S. J., Eck, J. L., Xu, K., Krishnadas, M., et al. (2014). Testing predictions of the Janzen–Connell hypothesis: a meta-analysis of experimental evidence for distance and density-dependent seed and seedling survival. J. Ecol. 102, 845–856. doi: 10.1111/1365-2745.12232
Costion, C. M., Hillmann-Kitalong, A., Perlman, S., and Edwards, W. (2013). Palau’s rare and threatened palm Ponapea palauensis (Arecaceae): population density, distribution, and threat assessment. Pac. Sci. 67, 599–607. doi: 10.2984/67.4.10
Cousens, R., Dytham, C., and Law, R. (2008). Dispersal in Plants: A Population Perspective. Oxford: Oxford University Press.
Davies, R. G., Orme, C. D. L., Webster, A. J., Jones, K. E., Blackburn, T. M., and Gaston, K. J. (2007). Environmental predictors of global parrot (Aves: Psittaciformes) species richness and phylogenetic diversity. Glob. Ecol. Biogeogr. 16, 220–233. doi: 10.1111/j.1466-8238.2007.00282.x
de Barros Leite, A., Brancalion, P. H. S., Guevara, R., and Galetti, M. (2012). Differential seed germination of a keystone palm (Euterpe edulis) dispersed by avian frugivores. J. Trop. Ecol. 28, 615–618. doi: 10.1017/s0266467412000594
Diamond, J., Bishop, K. D., and Gilardi, J. D. (1999). Geophagy in New Guinea birds. Ibis 141, 181–193. doi: 10.1111/j.1474-919x.1999.tb07540.x
Dracxler, C. M., and Kissling, W. D. (2021). The mutualism–antagonism continuum in Neotropical palm–frugivore interactions: from interaction outcomes to ecosystem dynamics. Biol. Rev. 97, 527–553. doi: 10.1111/brv.12809
Dracxler, C. M., Pires, A. S., and Fernandez, F. A. S. (2011). Invertebrate seed predators are not all the same: seed predation by bruchine and scolytine beetles affects palm recruitment in different ways. Biotropica 43, 8–11. doi: 10.1111/j.1744-7429.2010.00723.x
Dransfield, J., Uhl, N. W., Asmussen, C. B., Baker, W. J., Harley, M. M., and Lewis, C. E. (2008). Genera Palmarum – The Evolution and Classification of Palms. Kew, VIC: Royal Botanic Gardens.
Eiserhardt, W. L., Svenning, J. C., Kissling, W. D., and Balslev, H. (2011). Geographical ecology of the palms (Arecaceae): determinants of diversity and distributions across spatial scales. Ann. Bot. 108, 1391–1416. doi: 10.1093/aob/mcr146
Emmons, L. H., and Stark, M. N. (1979). Elemental composition of a natural mineral lick in Amazonia. Biotropica 11, 311–313. doi: 10.2307/2387925
Faurby, S., Eiserhardt, W. L., Baker, W. J., and Svenning, J. C. (2016). An all-evidence species-level supertree for the palms (Arecaceae). Mol. Phylogenet. Evol. 100, 57–69. doi: 10.1016/j.ympev.2016.03.002
Fehr, V., Buitenwerf, R., and Svenning, J. C. (2020). Non-native palms (Arecaceae) as generators of novel ecosystems: a global assessment. Div. Dist. 26, 1523–1538. doi: 10.1111/ddi.13150
Fleming, T. H., and Muchhala, N. (2008). Nectar-feeding bird and bat niches in two worlds: pantropical comparisons of vertebrate pollination systems. J. Biogeogr. 35, 764–780. doi: 10.1111/j.1365-2699.2007.01833.x
Fragoso, J. M. V., Silvius, K. M., and Correa, J. A. (2003). Long-distance seed dispersal by tapirs increases seed survival and aggregates tropical trees. Ecology 84, 1998–2006. doi: 10.1890/01-0621
Franco-Quimbay, J., and Rojas Robles, R. (2015). Frugivory and seed dispersal of the palm Oenocarpus bataua in two regions with different conservation status. Actual. Biol. 37, 273–285.
Freeland, W. J., Calcott, P. H., and Geiss, D. P. (1985). Allelochemicals, minerals and herbivore population size. Biochem. Syst. Ecol. 13, 195–206. doi: 10.1016/0305-1978(85)90079-1
Garamszegi, L. Z. (ed.) (2014). Modern Phylogenetic Comparative Methods and their Application in Evolutionary Biology. Berlin: Springer.
Gelis, S. (2011). “A review of the nutrition of lories and lorikeets,” in Proceedings of AVES International Parrot Convention.
Gilardi, J. D., and Toft, C. A. (2012). Parrots eat nutritious foods despite toxins. PLoS One 7:e38293. doi: 10.1371/journal.pone.0038293
Gleiser, G., Lambertucci, S. A., Speziale, K. L., Hiraldo, F., Tella, J. L., and Aizen, M. A. (2017). The southernmost parakeet might be enhancing pollination of a dioecious conifer. Ecology 98, 2969–2971. doi: 10.1002/ecy.1938
Govaerts, R., and Dransfield, J. (2005). World Checklist of Palms. Richmond: Royal Botanic Gardens, 223.
Guimarães, P. R. Jr, Galetti, M., and Jordano, P. (2008). Seed dispersal anachronisms: rethinking the fruits extinct megafauna ate. PLoS One 3:e1745. doi: 10.1371/journal.pone.0001745
Hadfield, J. D. (2010). MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package. J. Stat. Soft. 33, 1548–7660.
Henderson, A. (2002). Evolution and Ecology of Palms. Bronx, NY: The New York Botanical Garden Press.
Hernández-Brito, D., Romero-Vidal, P., Hiraldo, F., Blanco, G., Díaz-Luque, J. A., Barbosa, J. M., et al. (2021). Epizoochory in parrots as an overlooked yet widespread plant–animal mutualism. Plants 10:760. doi: 10.3390/plants10040760
Howe, H. F., and Smallwood, J. (1982). Ecology of seed dispersal. Ann. Rev. Ecol. Syst. 13, 201–228.
Jakubas, W. J., Guglielmo, C. G., Vispo, C., and Karasov, W. H. (1995). Sodium-balance in ruffed grouse as influenced by sodium levels and plant secondary metabolites in quaking aspen. Can. J. Zool. 73, 1106–1114. doi: 10.1139/z95-132
Jansen, P. A., Hirsch, B. T., Emsens, W. J., Zamora-Gutierrez, V., Wikelski, M., and Kays, R. (2012). Rodents as dispersers of megafaunal seeds. Proc. Natl. Acad. Sci. U.S.A. 109, 12610–12615. doi: 10.1073/pnas.1205184109
Janzen, D. H. (1970). Herbivores and the number of tree species in tropical forests. Am. Nat. 104, 501–528. doi: 10.1086/282687
Jetz, W., Thomas, G. H., Joy, J. B., Hartmann, K., and Mooers, A. O. (2012). The global diversity of birds in space and time. Nature 491, 444–448. doi: 10.1038/nature11631
Jordano, P. (2016). Sampling networks of ecological interactions. Funct. Ecol. 30, 1883–1893. doi: 10.1111/1365-2435.12763
Kiltie, R. A. (1981). Distribution of palm fruits on a rain forest floor: why white-lipped peccaries forage near objects. Biotropica 13, 141–145. doi: 10.2307/2387716
Kissling, W. D., Baker, W. J., Balslev, H., Barfod, A. S., Borchsenius, F., Dransfield, J., et al. (2007). Quaternary and pre-quaternary historical legacies in the global distribution of a major tropical plant lineage. Glob. Ecol. Biogeogr. 16, 220–233.
Kissling, W. D., Balslev, H., John Baker, W., Dransfield, J., Göldel, B., Lim, J. Y., et al. (2019). Data from: PalmTraits 1.0, a species-level functional trait database for palms worldwide. Sci. Data 6, 1–13. doi: 10.1038/s41597-019-0189-0
Kissling, W. D., Eiserhardt, W. L., Baker, W. J., Borchsenius, F., Couvreur, T. L. P., Balslev, H., et al. (2012). Phylogenetic assemblage structure in palms. Proc. Natl. Acad. Sci. U.S.A. 109, 7379–7384.
Laundré, J. W., Hernandez, L., and Ripple, W. J. (2010). The landscape of fear: ecological implications of being afraid. Open Ecol. J. 3, 1–7. doi: 10.2174/1874213001003030001
Levey, D. J., Moermond, T. C., and Denslow, J. S. (1994). “Frugivory: an overview,” in La Selva: Ecology and Natural History of a Neotropical Rainforest, eds L. A. McDade, K. S. Bawa, H. A. Hespenheide, and G. S. Hartshorn (Chicago, IL: University of Chicago Press), 282–294. doi: 10.1002/ajp.22246
Li, M., and Bolker, B. (2019). Wzmli/Phyloglmm: First Release of Phylogenetic Comparative Analysis in lme4-verse (Version v1.0.0). Zenodo. doi: 10.5281/zenodo.2639887
Lieberman, M., and Lieberman, D. (1986). An experimental study of seed ingestion and germination in plant-animal assemblage in Ghana. J. Trop. Ecol. 2, 113–126. doi: 10.1017/s0266467400000717
Lim, J. Y., Svenning, J. C., Göldel, B., Faurby, S., and Kissling, W. D. (2020). Frugivore-fruit size relationships between palms and mammals reveal past and future defaunation impacts. Nat. Commun. 11:4904. doi: 10.1038/s41467-020-18530-5
Loayza, A. P., and Knight, T. (2010). Seed dispersal by pulp consumers, not “legitimate” seed dispersers, increases Guettarda viburnoides population growth. Ecology 91, 2684–2695. doi: 10.1890/09-0480.1
Luna, A., Romero-Vidal, P., Hiraldo, F., and Tella, J. L. (2018). Cities may save some threatened species but not their ecological functions. PeerJ 6:e4908. doi: 10.7717/peerj.4908
Martins, R. C., Filgueiras, T. S., and Albuquerque, U. P. (2014). Use and diversity of palm (Arecaceae) resources in Central Western Brazil. Sci. World J. 2:942043. doi: 10.1155/2014/942043
Maués, M. M., and Venturieri, G. C. (1997). Pollination ecology of platonia insignis mart. (Clusiaceae), a fruit tree from Eastern Amazon region. Acta Hortic. 437, 255–260. doi: 10.17660/actahortic.1997.437.29
Mendieta-Aguilar, G., Pacheco, L. F., and Roldán, A. I. (2015). Dispersión de semillas de Mauritia flexuosa (Arecaceae) por frugívoros terrestres en Laguna Azul, Beni, Bolivia. Acta Amaz. 45, 45–56. doi: 10.1590/1809-4392201400283
Mittelman, P., Dracxler, C. M., Santos-Coutinho, P. R. O., and Pires, A. S. (2021). Sowing forests: a synthesis of seed dispersal and predation by agoutis and their influence on plant communities. Biol. Rev. 96, 2425–2445. doi: 10.1111/brv.12761
Montesinos-Navarro, A., Hiraldo, F., Tella, J. L., and Blanco, G. (2017). Network structure embracing mutualism-antagonism continuums increases community robustness. Nat. Ecol. Evol. 1:1661. doi: 10.1038/s41559-017-0320-6
Mougi, A. (ed.) (2020). Diversity of Functional Traits and Interactions. Perspectives on Community Dynamics. Singapore: Springer.
Muñoz, G., Trøjelsgaard, K., and Kissling, W. D. (2019). A synthesis of animal-mediated seed dispersal of palms reveals distinct biogeographical differences in species interactions. J. Biogeogr. 46, 466–484. doi: 10.1111/jbi.13493
Muscarella, R., Emilio, T., Phillips, O. L., Lewis, S. L., Slik, F., Baker, W. J., et al. (2020). The global abundance of tree palms. Glob. Ecol. Biogeogr. 29, 1495–1514.
Nascimento, L. M., Vendruscolo, E. P., Campos, L. F. C., Garcia, L. G. C., Pires, L. L., and Seleguini, A. (2019). Pre-depulping and depulping treatments and the emergence of queen palm seeds (Syagrus romanzoffiana [Cham.] Glassman). Rev. Colomb. Ciencias Hortíc. 13, 466–472. doi: 10.17584/rcch.2019v13i3.8363
Olah, G., Butchart, S. H. M., Symes, A., Medina Guzmán, I., Cunningham, R., Brightsmith, D. J., et al. (2016). Ecological and socio-economic factors affecting extinction risk in parrots. Biodivers. Conserv. 25, 205–223. doi: 10.1007/s10531-015-1036-z
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial ecoregions of the world: a new map of life on earth. Bioscience 51, 933–938. doi: 10.1641/0006-3568(2001)051[0933:teotwa]2.0.co;2
Onstein, R. E., Baker, W. J., Couvreur, T. L. P., Faurby, S., Svenning, J.-C., Daniel Kissling, W., et al. (2017). Frugivory-related traits promote speciation of tropical palms. Nat. Ecol. Evol. 1, 1903–1911. doi: 10.1038/s41559-017-0348-7
Pehrson, A. (1983). Digestibility and retention of food components in caged mountain hares Lepus timidus during winter. Holarct. Ecol. 6, 395–403. doi: 10.1111/j.1600-0587.1983.tb01235.x
Pereira, A. C. F., Fonseca, F. S. A., Mota, G. R., Fernandes, A. K. C., Fagundes, M., Reis-Júnior, R., et al. (2014). Ecological interactions shape the dynamics of seed predation in Acrocomia aculeata (Arecaceae). PLoS One 9:e98026. doi: 10.1371/journal.pone.0098026
Pérez-Méndez, N., Jordano, P., García, C., and Valido, A. (2016). The signatures of Anthropocene defaunation: cascading effects of the seed dispersal collapse. Sci. Rep. 6:24820. doi: 10.1038/srep24820
Pitman, N. C. A., Terborgh, J. W., Silman, M. R., Núñez, P., Neill, D. A., Cerón, C. E., et al. (2001). Dominance and distribution of tree species in upper amazonian terra firme forests. Ecology 82, 2101–2117. doi: 10.1890/0012-9658(2001)082[2101:dadots]2.0.co;2
Prada Villalobos, M., and Araújo Bagno, M. (2012). Avian frugivores feeding on Mauritia flexuosa (Arecaceae) fruits in Central Brazil. Rev. Brasil. Ornitol. 20, 26–29.
Provost, K. L., Joseph, L., and Tilston Smith, B. (2018). Resolving a phylogenetic hypothesis for parrots: implications from systematics to conservation. Emu 118, 7–21. doi: 10.1080/01584197.2017.1387030
R Core Team (2021). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Randall, D., Burggren, W., and French, K. (1997). Eckert Animal Physiology: Mechanisms and Adaptations, 4th Edn. New York, NY: W. H. Freeman and Company.
Reichardt, P. B., Bryant, J. P., Clausen, T. P., and Wieland, G. D. (1984). Defense of winter-dormant Alaska paper birch against snowshoe hares. Oecologia 65, 58–69. doi: 10.1007/BF00384463
Reichgelt, T., West, C. K., and Greenwood, D. R. (2018). The relation between global palm distribution and climate. Sci. Rep. 8:4721. doi: 10.1038/s41598-018-23147-2
Renton, K., Salinas-Melgoza, A., De Labra-Hernández, M. Á, and de la Parra-Martínez, S. M. (2015). Resource requirements of parrots: nest site selectivity and dietary plasticity of Psittaciformes. J. Ornithol. 156, 73–90. doi: 10.1007/s10336-015-1255-9
Robinson, O. J., Socolar, J. B., Stuber, E. F., Auer, T., Berryman, A. J., Boersch-Supan, P. H., et al. (2022). Extreme uncertainty and unquantifiable bias do not inform population sizes. Proc. Natl. Acad. Sci. U.S.A. 119:e2113862119. doi: 10.1073/pnas.2113862119
Rousset, F. (2004). Genetic Structure and Selection in Subdivided Populations. Princeton, NJ: Princeton University Press.
Sazima, I. (2008). The parakeet Brotogeris tirica feeds on and disperses the fruits of the palm Syagrus romanzoffiana in Southeastern Brazil. Biota Neotrop. 8, 231–234. doi: 10.1590/s1676-06032008000100026
Sebastián-González, E., Hiraldo, F., Blanco, G., Hernández Brito, D., Romero-Vidal, P., Carrete, M., et al. (2019). The extent, frequency and ecological functions of food wasting by parrots. Sci. Rep. 9:15280. doi: 10.1038/s41598-019-51430-3
Silman, M. R., Terborgh, J. W., and Kiltie, R. A. (2003). Population regulation of a dominant rain forest tree by a major seed predator. Ecology 84, 431–438. doi: 10.1890/0012-9658(2003)084[0431:proadr]2.0.co;2
Silva, L. B., Pereira, G. A., Passos, P. B., and Almeida, N. M. (2021). Seed dispersal of the palm Acrocomia aculeata by the blue-and-yellow macaw (Ara ararauna). Braz. J. Biol. 83:e244697. doi: 10.1590/1519-6984.244697
Silvius, K. M., and Fragoso, J. M. V. (2002). Pulp handling by vertebrate seed dispersers increases palm seed predation by bruchid beetles in the northern Amazon. J. Ecol. 90, 1024–1032. doi: 10.1046/j.1365-2745.2002.00728.x
Sist, P., and Puig, H. (1987). Régénération, dynamique des populations et dissémination d’un palmier de Guyane française: Jessenia bataua (Mart.) Burret subsp. oligocarpa (Griseb. and H. Wendl.) Balick. Bull. Mus. Nat. Hist. Nat. B 9, 317–336.
Song, X., Lim, J. Y., Yang, J., and Luskin, M. S. (2021). When do Janzen-Connell effects matter? A phylogenetic meta-analysis of conspecific negative distance and density dependence experiments. Ecol. Lett. 24, 608–620. doi: 10.1111/ele.13665
Sosnowska, J., and Balslev, H. (2009). American palm ethnomedicine: a meta-analysis. J. Ethnobiol. Ethnomed. 5:43. doi: 10.1186/1746-4269-5-43
Southerton, S. G., Birt, P., Porter, J., and Ford, H. A. (2004). Review of gene movement by bats and birds and its potential significance for eucalypt plantation forestry. Aust. For. 67, 44–53. doi: 10.1080/00049158.2004.10676205
Speziale, K. L., Lambertucci, S. A., Gleiser, G., Tella, J. L., Hiraldo, F., and Aizen, M. A. (2018). An overlooked plant-parakeet mutualism counteracts human overharvesting on an endangered tree. R. Soc. Open Sci. 5:171456. doi: 10.1098/rsos.171456
Sunderland, T. C. H., and Morakinyo, T. (2002). Nypa fruticans, a weed in West Africa. Palms 46, 154–155.
Svenning, J. C. (2001). On the role of microenvironmental heterogeneity in the ecology and diversification of neotropical rain-forest palms (Arecaceae). Bot. Rev. 67, 1–53. doi: 10.1007/bf02857848
Svenning, J. C. (2002). Crown illumination limits the population growth rate of a Neotropical understorey palm (Geonoma macrostachys, Arecaceae). Plant Ecol. 159, 185–199.
Svenning, J. C., and Wright, S. J. (2005). Seed limitation in a Panamanian forest. J. Ecol. 93, 853–862. doi: 10.1111/j.1365-2745.2005.01016.x
Taxonomic Databases Working Group (2001). World Geographic Scheme for Recording Plant Distributions Standard. Biodiversity Information Standards (TDWG). Available online at: http://www.tdwg.org/standards/109 (accessed January 12, 2022).
Tella, J. L., Baños-Villalba, A., Hernández-Brito, D., Rojas, A., Pacífico, E., Díaz-Luque, J. A., et al. (2015). Parrots as overlooked seed dispersers. Front. Ecol. Environ. 13:338–339. doi: 10.1890/1540-9295-13.6.338
Tella, J. L., Blanco, G., Dénes, F. V., and Hiraldo, F. (2019). Overlooked parrot seed dispersal in Australia and South America: insights on the evolution of dispersal syndromes and seed size in Araucaria trees. Front. Ecol. Evol. 7:82. doi: 10.3389/fevo.2019.00082
Tella, J. L., Dénes, F. V., Zulian, V., Prestes, N. P., Martínez, J., Blanco, G., et al. (2016). Endangered plant–parrot mutualisms: Seed tolerance to predation makes parrots pervasive dispersers of the Parana pine. Sci. Rep. 6:31709. doi: 10.1038/srep31709
Tella, J. L., Hiraldo, F., Pacífico, E., Díaz-Luque, J. A., Dénes, F. V., Fontoura, F. M., et al. (2020). Conserving the diversity of ecological interactions: the role of two threatened macaw species as legitimate dispersers of “megafaunal” fruits. Diversity 12:45. doi: 10.3390/d12020045
Tella, J. L., Romero-Vidal, P., Dénes, F. V., Hiraldo, F., Toledo, B., and Rossetto, F. (2021). Roadside car surveys: methodological constraints and solutions for estimating parrot abundances across the world. Diversity 13:300. doi: 10.3390/d13070300
Terborgh, J. (1986). “Community aspects of frugivory in tropical forests,” in Frugivores and Seed Dispersal, eds A. Estrada and T. H. Fleming (Dordrecht: Dr. W. Junk Publishers), 371–384. doi: 10.1007/978-94-009-4812-9_32
Toft, C. A., and Wright, T. F. (2015). Parrots of the Wild: A Natural History of the World’s Most Captivating Birds. Oakland, CA: University of California Press.
Vicentini, A., and Fischer, E. (1999). Pollination of Moronobea coccinea (Clusiaceae) by the Golden-winged Parakeet in the central Amazon. Biotropica 31, 692–696. doi: 10.1111/j.1744-7429.1999.tb00419.x
Wandrag, E. M., Dunham, A. E., Duncan, R. P., and Rogers, H. S. (2018). Seed dispersal increases local species richness and reduces spatial turnover of tropical tree seedlings. Proc. Natl. Acad. Sci. U.S.A. 114, 10689–10694. doi: 10.1073/pnas.1709584114
WFO (2021). World Flora Online. Available online at: http://www.worldfloraonline.org (accessed July 19, 2021)
Zona, S. (2006). Additions to a Review of Animal-Mediated Seed Dispersal of Palms. Available online at: http://www.virtualherbarium.org/ (accessed January 12, 2022).
Keywords: pollination, seed dispersal, seed predation, dispersal distances, megafaunal fruits, defleshing, introduced species
Citation: Carrete M, Hiraldo F, Romero-Vidal P, Blanco G, Hernández-Brito D, Sebastián-González E, Díaz-Luque JA and Tella JL (2022) Worldwide Distribution of Antagonistic-Mutualistic Relationships Between Parrots and Palms. Front. Ecol. Evol. 10:790883. doi: 10.3389/fevo.2022.790883
Received: 07 October 2021; Accepted: 19 April 2022;
Published: 11 May 2022.
Edited by:
Gerardo Avalos, University of Costa Rica, Costa RicaReviewed by:
Cibele De Cássia Silva, Universidade Federal de Goiás, BrazilMauro Galetti, São Paulo State University, Brazil
Copyright © 2022 Carrete, Hiraldo, Romero-Vidal, Blanco, Hernández-Brito, Sebastián-González, Díaz-Luque and Tella. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martina Carrete, bWNhcnJldGVAdXBvLmVz