 Yiyang Feng
Yiyang Feng Mengyu Yang†
Mengyu Yang† Haijun Yang
Haijun Yang- School of Ecology and Environmental Science, Yunnan Key Laboratory for Plateau Mountain Ecology and Restoration of Degraded Environments, Yunnan University, Kunming, China
In this study, the community structure and diversity of macroinvertebrate benthic animals in the 18 streams, Cangshan, Dali City, Yunnan Province, were investigated in detail. The main water environment factors driving the changes in these communities were analyzed, and the effects of the wet and dry seasons were explored. There were significant differences in the characteristics of the water environment and the community structure of benthic animals between the wet and dry seasons. In particular, the precipitation, water flow, and water temperature increased in the wet season, and the species diversity and functional diversity of benthic animals were significantly greater than those in the dry season. Through the analysis of the linear mixed effect model, in the wet season, the rainfall increases greatly, leading to the acceleration of the flow velocity and the increase in turbidity; rich nutrients and suspended particles enter the water, and a relatively high water temperature provides a relatively suitable habitat. The combined effects of these factors improve the species diversity and functional diversity of benthic animals. Moreover, the wet season coincides with the agricultural farming period in the area, and the increase in organic matter and nutrients in agricultural runoff also provides food for the reproduction of some benthic animals, leading to an increase in their diversity. These results suggest that the combined effects of such significant seasonal changes and human activities (agricultural development) need to be considered when protecting and managing aquatic ecosystems in regions with distinct wet and dry seasons. This study provides a key basis for the scientific formulation of water resource management policies, which not only helps maintain the health and stability of the ecosystem of the 18 streams in Cangshan but also provides a valuable reference for the seasonal protection and management of similar water ecosystems around the world.
1 Introduction
Stream ecosystems play a crucial role in global biodiversity, providing important support for a wide range of aquatic and terrestrial organisms and offering invaluable ecological services to humanity (Chen et al., 2023; Kazikina, 2022). However, despite their significance for human progress, stream ecosystems face severe threats globally due to several factors, including climate change, land use patterns, geographical location, and human activities. These environmental pressures have profound impacts on aquatic biotic communities, altering their composition, structure, and ecological functions (Farooq and Siddiqui, 2020; Hroch and Brabec, 2022). Among these communities, benthic macroinvertebrates require special attention due to their sensitivity toward environmental changes and stressors as well as their important role in aquatic food webs (Golovatyuk et al., 2023; He et al., 2021).
Benthic organisms, such as insects, crustaceans, and worms, primarily inhabit the sediments of streambeds, feeding on various algae, organic particles, and litter (Kesti et al., 2024; Oyeku et al., 2023). They also serve as a crucial food source for fish in streams (Sun et al., 2022). Benthic organisms are important biological indicators for evaluating the health of streams, which is attributed to their quick response to changes in water quality (Varadinova et al., 2023). Investigating the diversity and structure of benthic communities can help us understand the influence of pollution and habitat destruction on stream ecosystems (Zemo et al., 2023). Such studies are vital for understanding how stream ecosystems respond to external stress, thus guiding stream conservation, utilization, and management (He et al., 2021). Given their sensitivity to environmental fluctuations, benthic macroinvertebrates are particularly susceptible to changes driven by seasonal dynamics. Therefore, understanding how seasonal variation affects their habitats and communities is essential for comprehensive ecological assessment and management.
Seasonal variation plays a fundamental role in shaping freshwater ecosystems, particularly in stream environments where hydrological and physicochemical conditions fluctuate markedly throughout the year (Gao et al., 2023; Miernik et al., 2023). In regions with distinct wet and dry seasons, these changes can lead to significant alterations in flow regimes, nutrient concentrations, temperature, and habitat stability (Liu et al., 2024). These fluctuations directly affect resource availability and reshape aquatic habitats (Okoro et al., 2024). They often result in habitat fragmentation, increased environmental stress during dry periods, and enhanced nutrient input during wet seasons. These factors profoundly influence the composition, diversity, and functionality of benthic macroinvertebrate communities (Gusmao et al., 2024). Despite increasing attention to seasonal dynamics in freshwater systems, highland streams in subtropical monsoon regions remain underrepresented in current research, particularly those with strong seasonal contrasts and complex land-use gradients (Liu et al., 2023b; Magni et al., 2023).
The 18 streams of the Cangshan Mountain region in Yunnan Province, China, represent a distinctive highland stream system characterized by steep elevation gradients, diverse land use patterns, and pronounced seasonal hydrological contrasts (He et al., 2020). These streams flow through varied ecosystems and exhibit typical regional microclimate features, with no extreme heat in summer or severe cold in winter. The climate is defined by distinct wet (May–October) and dry (November–April) seasons (Li et al., 2024). During the wet season, increased precipitation, water flow, and water temperature create dynamic and nutrient-rich aquatic habitats. In contrast, the dry season is marked by reduced rainfall, lower flow velocity, and cooler water temperatures, leading to increased environmental stress (Ni et al., 2023). Such seasonal variations have been shown to significantly affect hydrological conditions and influence the structure and diversity of benthic macroinvertebrate communities. For example, high flow and warmer water in the wet season typically support greater species richness and functional diversity, while low flow and cooler conditions in the dry season tend to limit habitat suitability and reduce biodiversity (Yang and Yang, 2019). However, it remains unclear which specific environmental drivers—such as nutrient concentrations, turbidity, dissolved oxygen, and flow velocity—are most influential in mediating these seasonal effects on benthic community composition and function (Zemo et al., 2023). Addressing this knowledge gap is essential to assess the resilience and vulnerability of highland stream ecosystems like those in Cangshan, particularly in the context of ongoing climate variability and human pressures (Li et al., 2024). The unique environmental conditions of these streams make them an ideal model system for investigating the ecological consequences of seasonal changes.
This study aims to comprehensively examine the seasonal dynamics of benthic macroinvertebrate communities in 13 representative perennial highland streams within the Cangshan region of Yunnan Province, China. Specifically, it investigates how spatiotemporal variations in key environmental drivers—including water temperature, flow velocity, and nutrient concentrations—associated with distinct wet and dry seasons influence community composition, taxonomic richness, and functional trait diversity. By integrating seasonal environmental variability with ecological responses, this study seeks to elucidate the underlying mechanisms governing biodiversity patterns in subtropical highland stream ecosystems. The outcomes are expected to contribute critical insights into the resilience and functioning of these systems under seasonal and anthropogenic pressures. Furthermore, the findings will provide a scientific foundation for evidence-based stream management and biodiversity conservation strategies, offering broader implications for comparable highland freshwater ecosystems facing similar environmental challenges globally.
2 Methods
2.1 Study area and sampling point setting
The 18 streams of the Cangshan Mountain (Figure 1) watershed are located in Dali City, Dali Bai Autonomous Prefecture, Yunnan Province, China. It spans a narrow area between 99°54’30″E to 100°12’12″E and 25°33’30″N to 25°59’48″N. Situated on the eastern slope of the Cangshan Mountain Range and the western coast of Erhai Lake, it originates from Cangshan Mountain and flows into Erhai Lake from west to east. The following 13 streams were selected for study: Wanhua stream (WHX), Yangxi stream (YX), Mangyong stream (MYX), Jinxi stream (JX), Lingquan stream (LQX), Baishi stream (BSX), Zhenhe stream (ZHX), Baihe stream (BHX), Heilong stream (HLX), Qingbi stream (QBX), Mocan stream (MCX), Tingming stream (TMX) and Yangnan stream (YNX). These 13 streams were selected because they traverse a range of land-use types—including primary forests, farmlands, villages, and roads—providing heterogeneous benthic habitats. In addition, they are the 13 perennial streams among the 18 streams in the Cangshan Mountain area, ensuring continuous flow conditions throughout the year for consistent seasonal sampling. The Shiba Xi region experiences a low-latitude plateau monsoon climate with a total length exceeding 90 km. The average annual runoff is approximately 2.331 billion m3/year, covering an area of about 352.62 km2 with an average natural water resource quantity of around 2.331 billion m3 over multiple years. The watershed includes five townships, namely Xiaguan, Dali, Yinqiao, Wanqiao, and Xizhou, with a population of approximately 380,000 people.
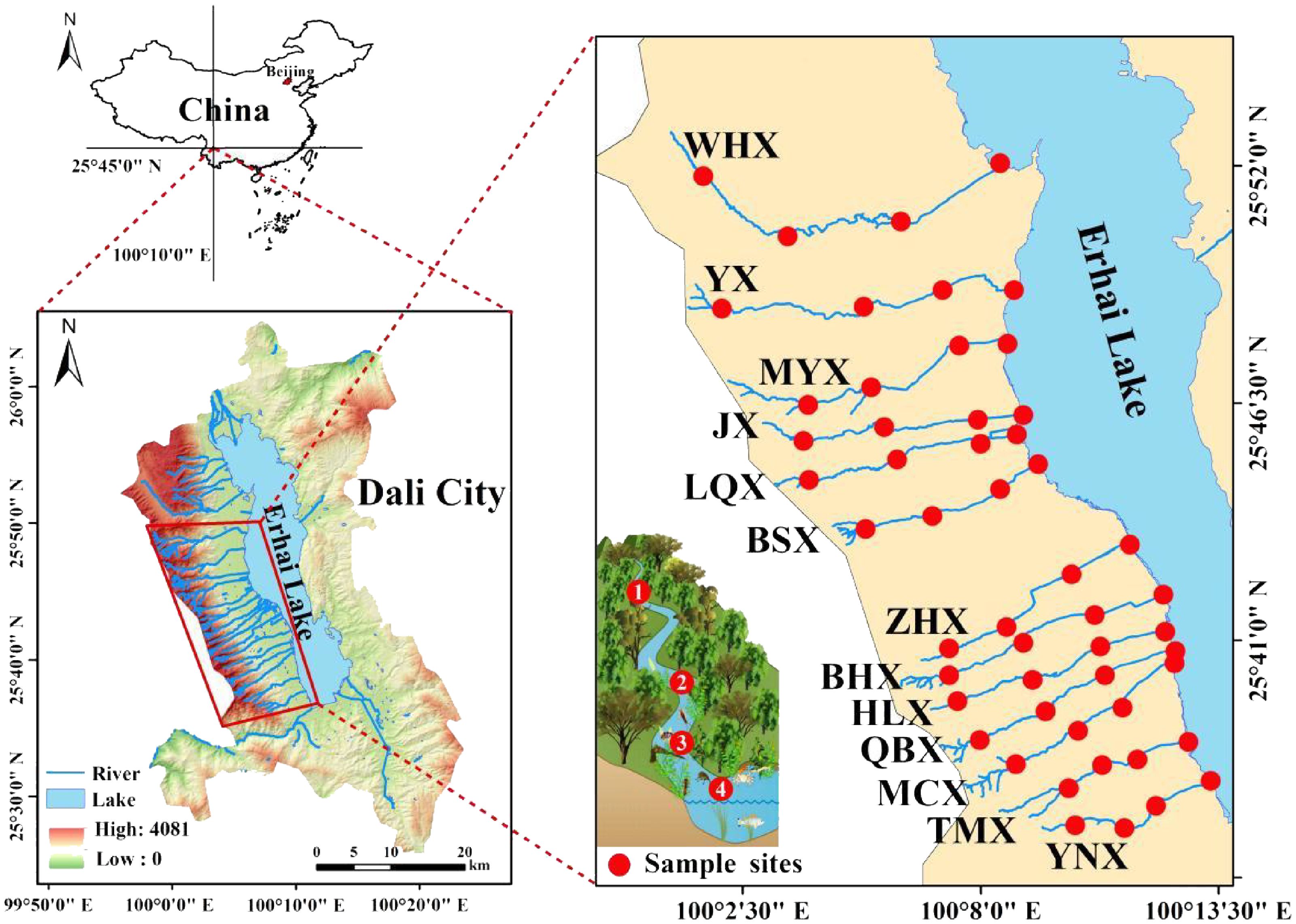
Figure 1. Geographical location and sampling point layout of the 18 streams of the Cangshan watershed.
Sampling sites were selected to capture a representative range of ecological conditions, including upstream forested reaches, midstream agricultural zones, and downstream areas influenced by human activities. Site selection also considered factors such as perennial flow, accessibility, and substrate diversity, ensuring both spatial coverage and comparability across environmental gradients (Bhaumik et al., 2023).At each site, five replicate samples of water quality and benthic macroinvertebrates were randomly collected within the sampling area to ensure representativeness and reduce spatial variability (Figure 1). In total, 52 sampling sites were established across the 13 streams, covering upstream (headwaters), midstream sections (villages and farmlands), and downstream estuaries. Sampling was conducted in July 2022 (wet season) and March 2023 (dry season).
2.2 Collection and identification of habitat factors and benthic organisms
To mitigate potential disruption to habitat data during benthic animal sample collection, a systematic approach was used. Five sampling points were selected within a 100-m stream reach centered around each transect for the comprehensive assessment of habitat conditions. Randomness was ensured by dividing the reach into equal segments and randomly selecting one sampling spot within each segment using a stratified random sampling approach. Water depth (Dep, m) was evaluated using standard rulers, whereas flow velocity (Vel, m/s), temperature (Temp, °C), DO mg/L), electrical conductivity (EC, µS/m), turbidity (Tur, NTU), and pH were measured in situ using a portable water quality analyzer (YSI EXO, USA). Furthermore, 1 L of water sample was obtained from each sampling point and quickly transported to the laboratory for post-lab analysis of total phosphorus (TP, mg/L), total nitrogen (TN, mg/L), and chemical oxygen demand (COD, mg/L). TP was determined using the ammonium molybdate spectrophotometric method after potassium persulfate digestion (HJ 670-2013); TN was analyzed via alkaline potassium persulfate digestion–UV spectrophotometric method (HJ 636-2012); and COD was measured using the dichromate reflux method (HJ 828-2017), following the protocols outlined in current national standards. The selection of environmental variables such as electrical conductivity (EC), total nitrogen (TN), and others was based on their ecological relevance and frequent use in freshwater ecological assessments. These variables are known to be sensitive to both natural variability (e.g., hydrological conditions) and anthropogenic pressures (e.g., agricultural runoff), and thus were considered suitable indicators for characterizing the physicochemical environment influencing benthic macroinvertebrate communities (Wilden et al., 2024).
Benthic macroinvertebrates were collected using Sobek nets measuring 30 × 30 cm with a pore size of 250 µm to ensure thorough coverage of their respective habitats at each sampling point. For each location, a 100-m stream section was divided into five equal segments, and one sample was randomly collected from each segment using the kick-sampling method, resulting in five replicate samples per site. The substrate retrieved by the net was thoroughly cleaned before being placed in white plastic bags labeled with unique identifiers. These samples were then preserved in 75% alcohol upon subsequent evaluation. On the collection days, in the laboratory, collected samples were carefully washed and transferred into designated white porcelain dishes. Large benthic macroinvertebrates were visually identified from these dishes and thoroughly sorted into containers corresponding to their specific sampling points. The benthic macroinvertebrates were taxonomically identified and quantified using a stereomicroscope according to relevant literature (Wang and Wang, 2025). Thus, we obtained taxonomic classification at the genus or species level.
2.3 Data statistics and analysis
Species density, dominance (species with dominance > 0.02 were defined as dominant species), indicators species (identified using Indicator Species Analysis with indicator values > 25 and P < 0.05 based on the method of Dufrêne and Legendre (1997)), Shannon–Wiener index (H’), Margalef diversity index (dM), Pielou evenness index (J), functional richness index (FRic), functional evenness index (FEve), the functional diversity index (FDiv), functional dispersion index (FDis), and Rao’s quadratic entropy index (RaoQ) were calculated using the “FD” package of R statistical software (He et al., 2021; Hu et al., 2024; Lv et al., 2016; Sun et al., 2024).
After standardizing the data, appropriate statistical tests were selected based on the data structure and sampling design. Since the same sites were sampled during the dry and wet seasons, paired t-tests were performed to compare the differences in physical and chemical indicators and benthic invertebrate community metrics between seasons. To assess differences among streams, the Kruskal–Wallis test was applied. If significant differences were detected, Mann–Whitney U tests were further conducted for pairwise comparisons (Arcifa et al., 2023; Liu et al., 2024; Varadinova et al., 2023). Pearson correlation analysis was performed to assess the correlations among all measured environmental factors (Gusmao et al., 2024; Kazikina, 2022; Miernik et al., 2023). Principal component analysis (PCA) was performed to rank the samples and identify key physicochemical gradients influencing streams during different seasons (Alves et al., 2024; He et al., 2020; Liu et al., 2023a). A linear mixed-effects model was established to determine the synergistic effect of multiple environmental factors on a specific community indicator of benthic invertebrates (Sun et al., 2024). In the model, all measured environmental variables (e.g., T, DO, Cond, Tur, pH, TN, TP, COD, Dep and Velocity) and season (wet/dry) were included as fixed effects, while stream was treated as a random effect to account for spatial variability among sampling sites. All analyses were conducted in R (Lin et al., 2022; Wang et al., 2023; Xiang et al., 2022).
All statistical analyses were performed in R version 4.3.3, using well-established packages tailored for ecological and multivariate data analysis. Prior to analysis, data standardization and normality checks were conducted using the scale(), shapiro.Test(), and qqnorm() functions. Paired t-tests were implemented using t.test(paired=TRUE), while non-parametric tests such as the Kruskal–Wallis test and Mann–Whitney U test were conducted with kruskal.Test() and wilcox.Test(), respectively. Pearson correlation coefficients among environmental variables were calculated using the cor() function, and visualized with the corrplot package. Principal Component Analysis (PCA) was performed using the prcomp() function with scaled variables, and results were visualized with ggbiplot or factoextra to aid interpretation of key environmental gradients. The linear mixed-effects model (LMM) was fitted using the lme4 package (lmer() function), where environmental variables and season were included as fixed effects, and stream identity as a random effect. Model summaries and effect significance were extracted using summary(), anova(), and the lmerTest package for p-values. Residual diagnostics were conducted using the DHARMa package to ensure model assumptions (e.g., normality, homoscedasticity) were met.
3 Results
3.1 Physicochemical properties of streams
3.1.1 Seasonal dynamics of habitat factors
The seasonal dynamics of habitat factors are shown in Figure 2. Almost all environmental factor data showed a general trend of increasing during the wet season. Additionally, the COD of LQX; EC of WHX, YX, MYX, and JX; TN levels of MYX and MCX; and TP levels in YNX exhibit significantly higher values during the dry season compared with the wet season. Notably, nearly all factors exhibited significant seasonal variations and inter-stream differences (Table 1). According to the national water quality standards for surface waters (GB 3838-2002), the measured values of DO, pH, TN, and TP mostly fell within Class II–III thresholds. This indicates that the overall water quality in the 18 streams is relatively good.
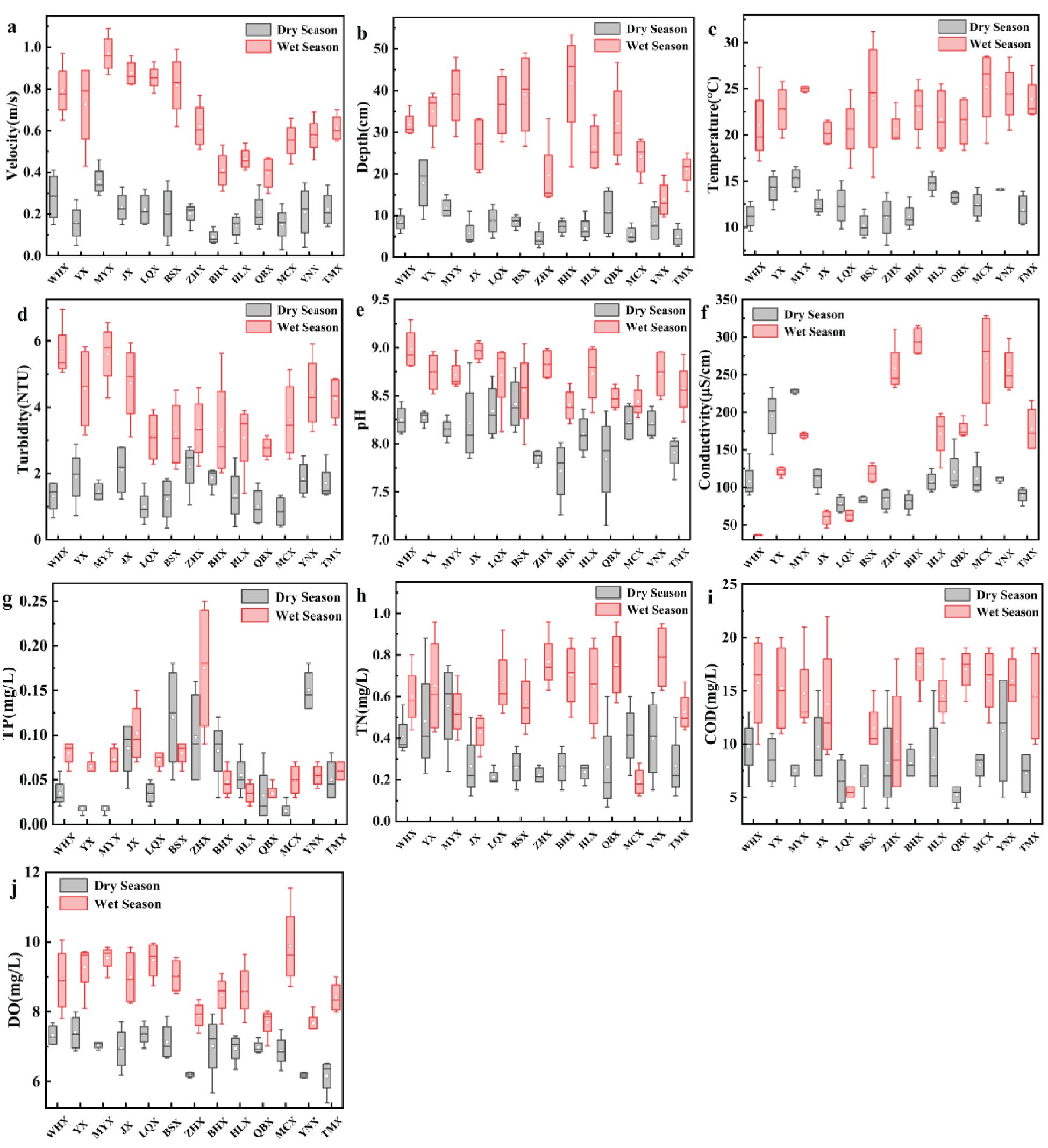
Figure 2. Seasonal dynamics of stream habitat factors: (a) velocity, (b) depth, (c) temperature, (d) turbidity, (e) pH, (f) conductivity, (g) TP, (h) TN, (i) COD, and (j) DO.
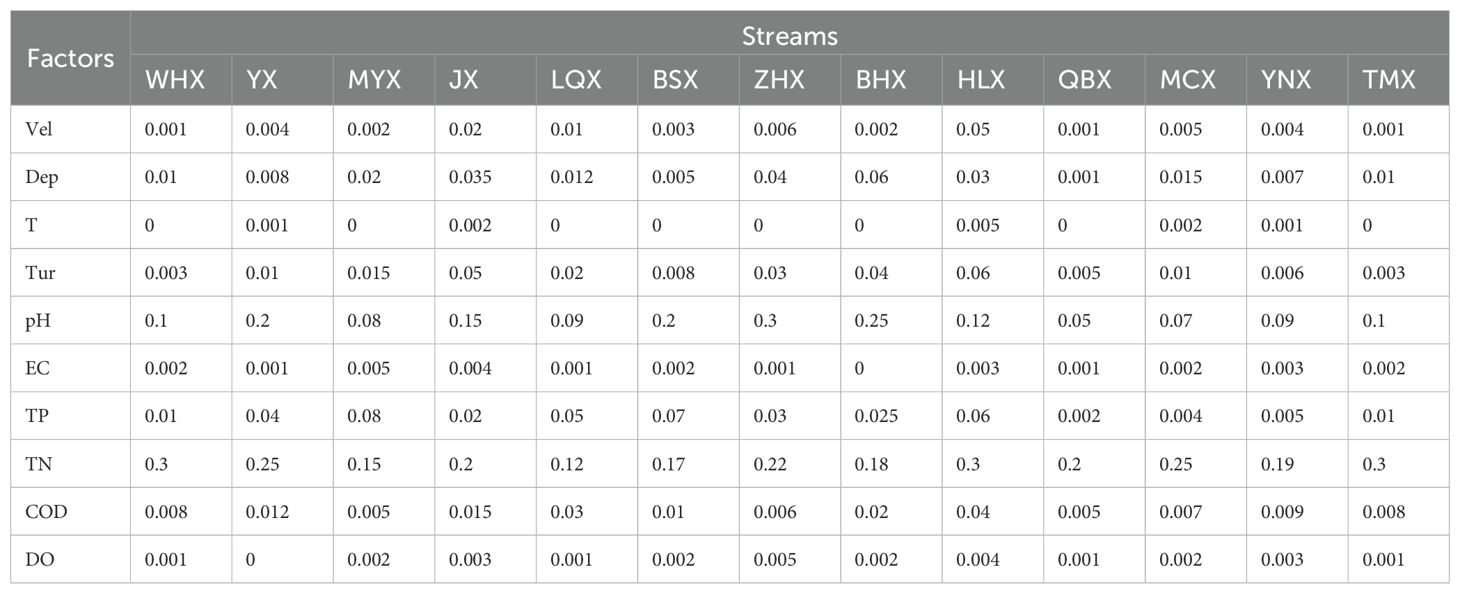
Table 1. The results of paired t-tests for environmental factors of each stream in dry and wet seasons.
3.1.2 Correlation between environmental factors
Figure 3 shows a heat map of the correlation between environmental factors. During the dry season, a significant positive correlation was found between water depth and DO (0.6). Additionally, there was a relatively strong positive correlation between DO and flow velocity (0.48) and DO and water temperature (0.47). The correlation between EC and TN was moderate and positive (0.42). Conversely, there was a stronger negative correlation between DO and water temperature (−0.45), a moderate negative correlation between TP and TN (−0.36), and a moderate negative correlation between DO and turbidity (−0.32). The correlation in the wet season was different, with EC and TP exhibiting a significant positive correlation (0.57). The correlation between flow velocity and DO was also significantly positive (0.52). Additionally, TN and turbidity also exhibited a significant positive correlation (0.50). While a positive correlation between water depth and DO was found (0.44), its strength was weaker than in the dry season. Additionally, DO and COD showed a moderate negative correlation (−0.47). The negative correlation between TP and TN remained the same as in the dry season (−0.36). However, EC and TN exhibited an opposite negative correlation (−0.36) compared with the dry season. Furthermore, EC showed moderate negative correlations with turbidity and pH (−0.35 and −0.33), respectively. To summarize, seasonal changes significantly impact the positive and negative correlations between these environmental factors.
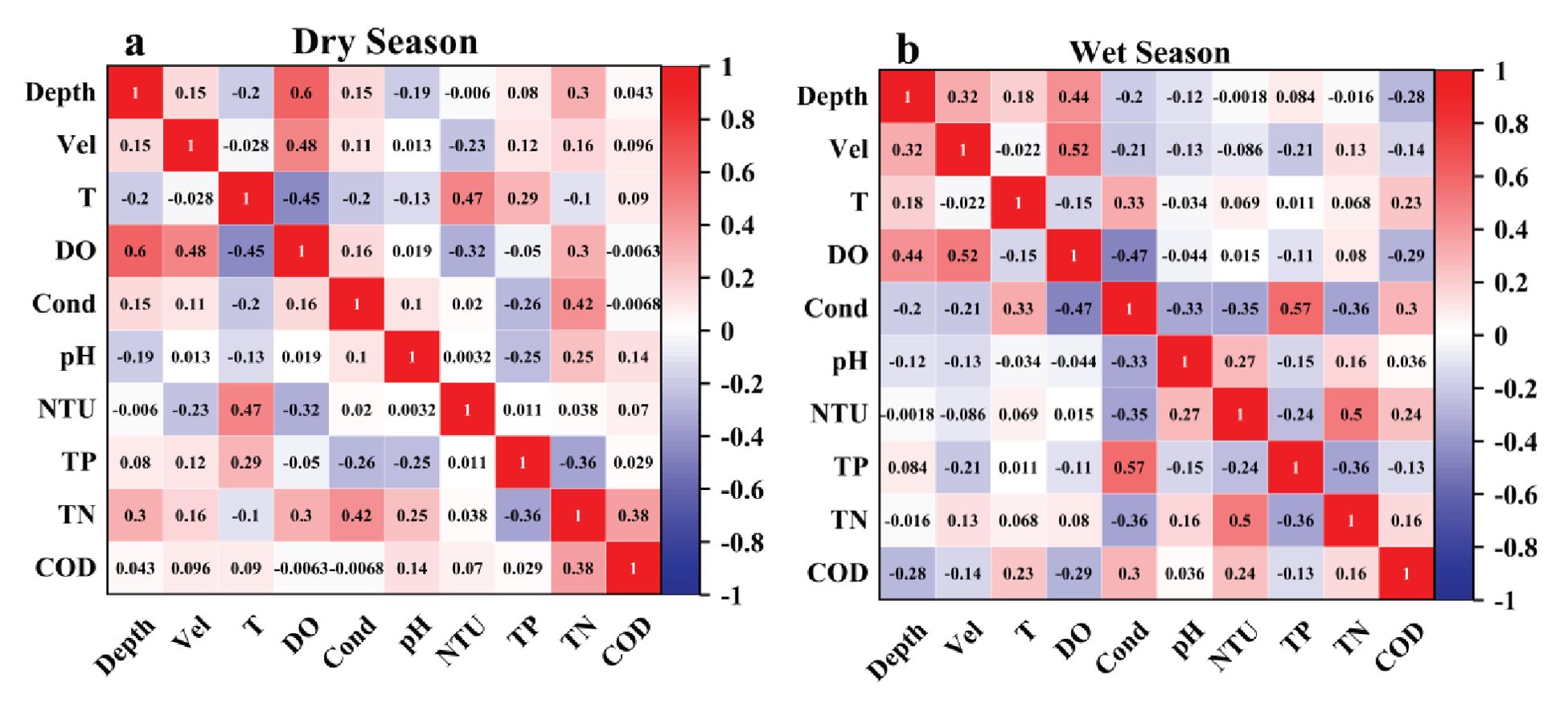
Figure 3. Heatmaps of Pearson correlation coefficients among habitat factors for (a) the dry season and (b) the wet season.
3.1.3 PCA
As shown in Figure 4, we determined the contributions of various environmental factors to principal component 1 (PC1) and principal component 2 (PC2) during dry and wet seasons. In the dry season, PC1 accounted for 27.7% of the data variability, whereas PC2 accounted for 15.9% of the variability. Together, PC1 and PC2 explained 43.6% of the total variance. Cond, TN, Dep, and Temp exhibited strong positive correlations with PC1, with Cond and TN showing longer vectors, revealing their dominant role in site-to-site variation. The primary variables for PC2 included COD and TP, as shown by their vectors pointing toward the positive direction of PC2, highlighting their significant variation across different sites. In the wet season, PC1 and PC2 accounted for 26.6% and 16.8% of the data variability, respectively. On the PC1 axis, Vel and pH exhibited strong positive correlations with PC1, whereas Cond and COD exhibited negative correlations with PC1, indicating their main directions of variation within the dataset. Conversely, on the PC2 axis in this season, water Dep and DO primarily exhibited variation in the positive direction, whereas TP and pH mainly exhibited variation in the negative direction. Comparatively, between both seasons, Temp, Cond, and Vel exhibited significant positive correlations with PC1, indicating that these factors greatly affect water quality throughout both seasons. The difference is attributed to the predominant influence of TP on PC2 during the dry season compared with its stronger negative correlations with PC1 during the wet season. Additionally, turbidity exhibited notable positive correlations with PC1, specifically during the wet season but less during the dry period. While Pearson correlation analysis provided insights into the seasonal shifts in pairwise relationships among environmental factors, PCA helped to identify the major gradients structuring stream conditions across space and time. These analyses complement each other and together provide a comprehensive understanding of seasonal environmental dynamics.
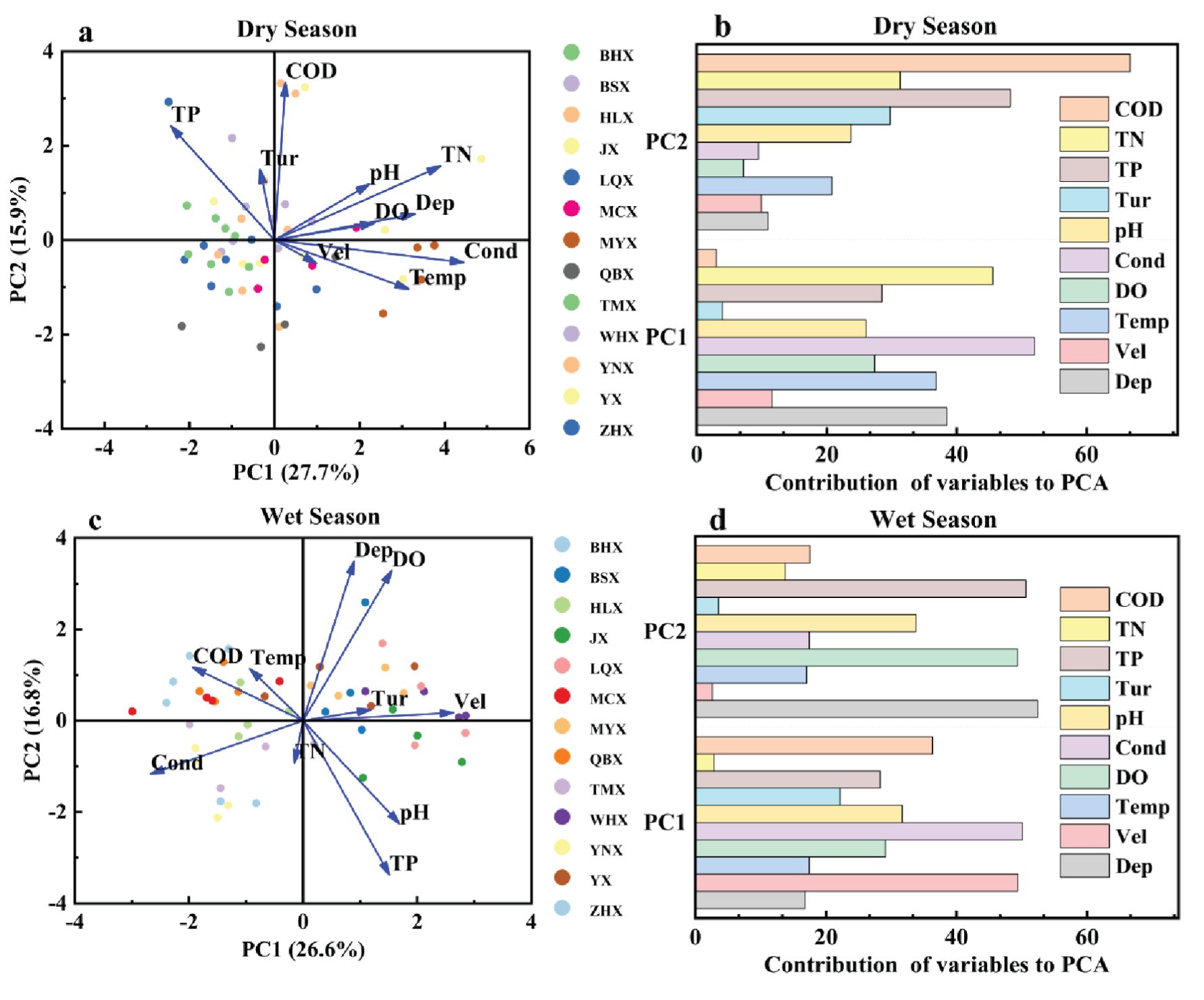
Figure 4. Principal component analysis (PCA) of habitat factors and their contributions: (a) PCA ordination plot for the dry season, (b) contribution rates of habitat factors in the dry season, (c) PCA ordination plot for the wet season, and (d) contribution rates of habitat factors in the wet season.
3.2 Composition and diversity of large benthic invertebrate communities
3.2.1 Species composition, dominant species, and indicator species
During the sampling process, 13,521 benthic invertebrates were collected, which belonged to three classes, six orders, ten families, 28 genera, and 56 taxonomic units. Table 2 shows the dominant species and their relative abundance in the 18 streams of Cangshan. In the wet season, Baetis and Gammarus were the two main dominant species in most streams, particularly in the TMX stream, where the relative abundance of Baetis was as high as 78.31%, which exceeded other streams. In the YX stream, the relative abundance of Gammarus was also relatively high, reaching 71.63%. Their dominance may be attributed to their adaptability to strong current, their generalist feeding strategies, and their reproductive efficiency under nutrient-rich and well-oxygenated conditions. Other species, including Epeorus sp., Haplotaxis, and Simuliini, became dominant species in certain specific streams. However, their abundance was generally lower than that of Baetis and Gammarus. In the dry season, Baetis and Gammarus remained the dominant species in most streams, with some streams showing a slight decrease in their relative abundance but still maintaining relatively high levels. Notably, Rheotanytarsus emerged as a key dominant species in the JX stream during the dry season (26.04%) but was absent during the wet season. This seasonal shift may reflect its adaptation to reduced flow, finer sediment, or lower dissolved oxygen typical of dry-season conditions.
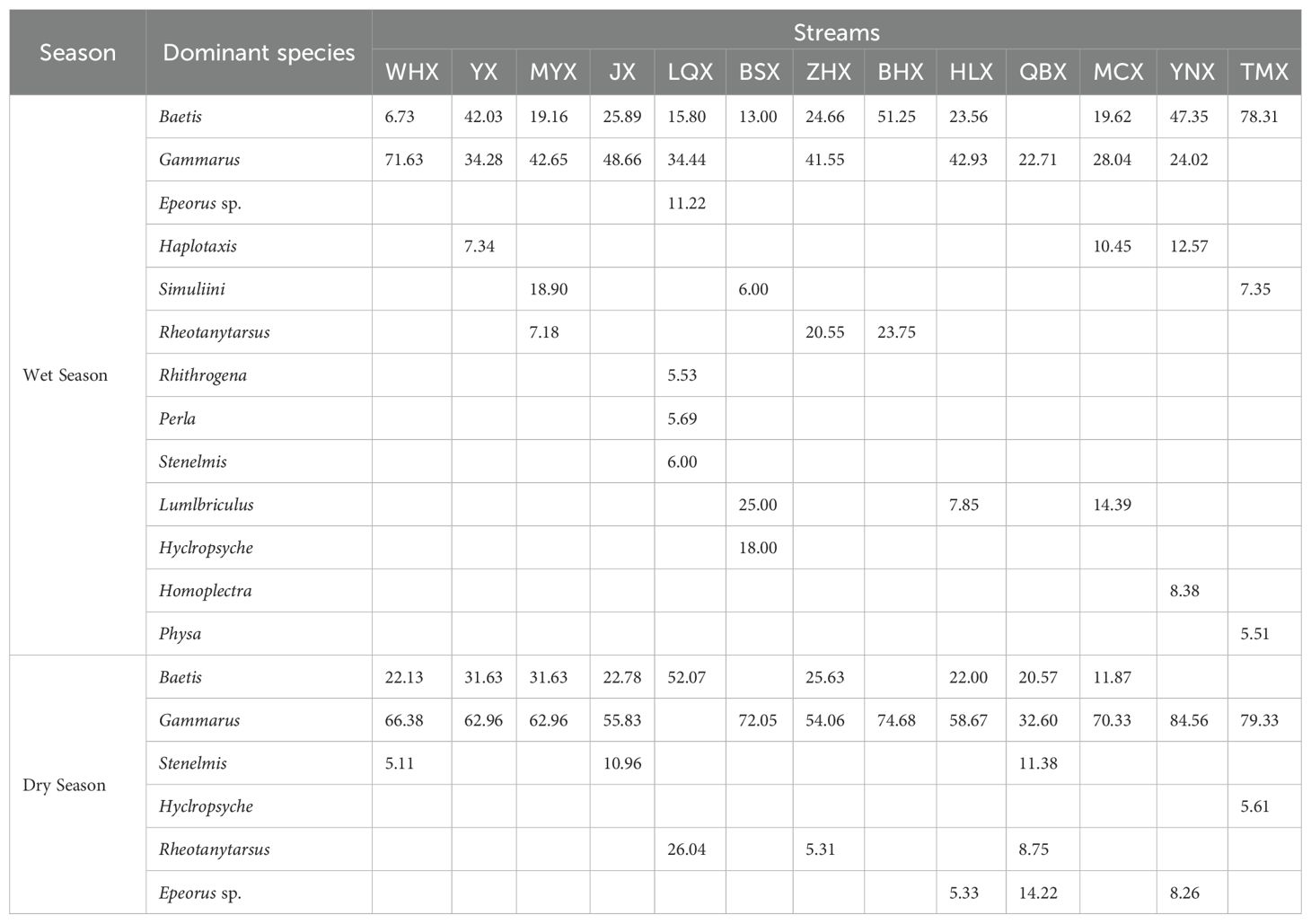
Table 2. Dominant species and their dominance in the 18 streams of the Cangshan Mountains.
Table 3 shows the seasonal indicator species. Baetis and Gammarus were indicator species in both seasons, with relatively high indicator values in LQX and YNX, respectively. Rheotanytarsus, on the other hand, was identified as a significant dry-season indicator in LQX (IV = 26.04, p = 0.044), while Lumbriculus was a wet-season indicator in BSX (IV = 25.00, p = 0.049), highlighting spatial and seasonal specificity. These distribution patterns suggest that dominant and indicator taxa are closely linked to stream-specific environmental drivers. The persistent dominance of Baetis and Gammarus across seasons indicates high ecological tolerance, whereas the appearance/disappearance of other species reflects sensitivity to seasonal environmental shifts.
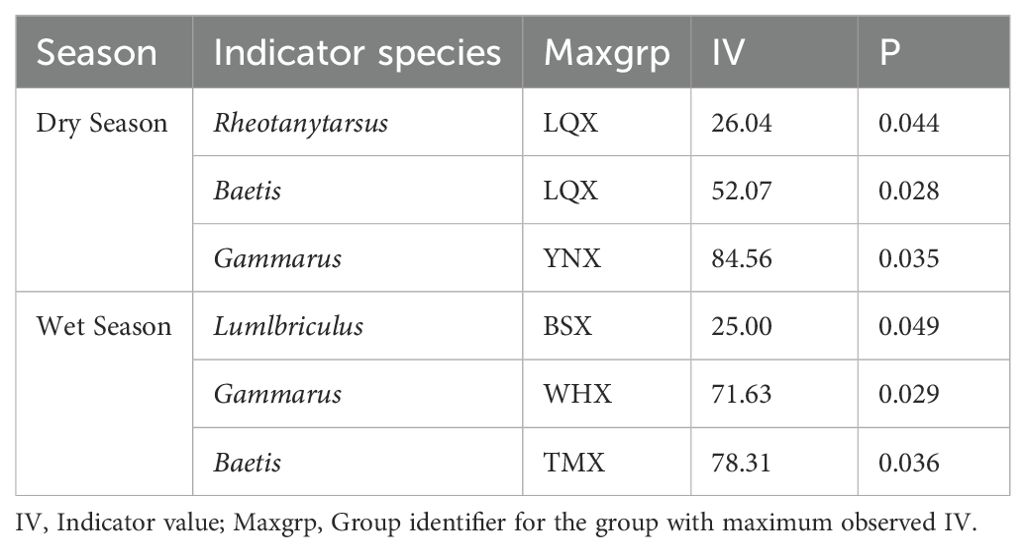
Table 3. Indicator species and their indicator values in the 18 streams of Cangshan Mountains (only species with IV>25 and P<0.05 are shown).
To summarize, the dominance of Baetis and Gammarus across streams indicates their broad ecological plasticity. In contrast, the seasonal appearance of taxa such as Rheotanytarsus illustrates species turnover shaped by hydrological variability. These findings highlight the role of environmental filtering in structuring benthic communities and reinforce the importance of linking biological patterns with habitat conditions, which aligns with the study’s objective of identifying seasonal drivers of stream biodiversity.
3.2.2 Characteristics of community structure
Figure 5 shows the community structure and diversity of sublittoral animals in the 18 streams of the Cangshan Mountains. Species abundance and density, as shown in Figures 5a, b, indicate the number of individuals in the community. In most streams, we found that species abundance and density were lower in the dry season than in the wet season. Nevertheless, some streams, namely YX and YNX, still had relatively high species abundance and density during the dry season, with species abundance reaching up to 589 and 590 ind and density reaching 327.22 and 327.78 ind/m2. This was significantly higher than the species abundance and density in BHX and MYX during the wet season. Species richness, particularly that of sensitive groups like Ephemeroptera, Plecoptera, and Trichoptera (EPT), is widely used as an indicator of stream health. Figures 5c and d show that species richness and EPT richness in most streams are higher in the wet season than in the dry season. The richness of EPT taxa across the streams ranged from 1 to 20. Nevertheless, some streams, including ZHX and BHX, have similar or slightly higher EPT richness in the dry season than in the wet season. Altogether, significant differences in community structure were observed between dry and wet seasons, with the wet season generally exhibiting better conditions. The changes in community structure are distinct among different streams.
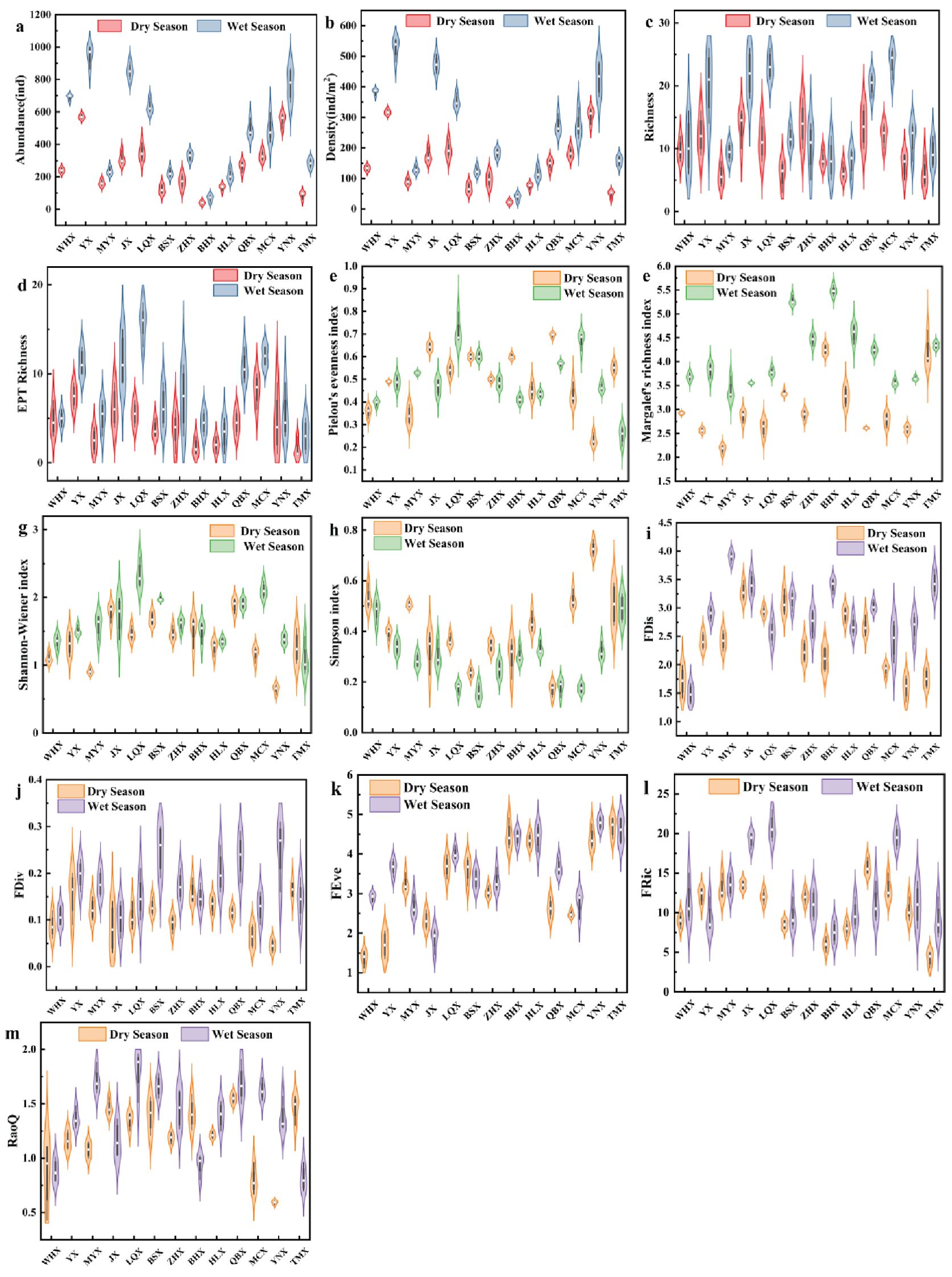
Figure 5. shows the community structure and diversity of benthic invertebrates in the two seasons of each stream catchment. (a) shows abundance, (b) shows density, (c) shows richness, (d) shows EPT Richness, (e) shows the Pielou index, (f) shows the Margalef index, (h) shows the Shannon–Wiener index, (h) shows the Simpson index, (i) shows the FDis index, (j) shows the FDiv index, (k) shows the FEve index, (l) shows the Fric index and (m) shows the RaoQ index.
3.2.3 Biodiversity of benthic communities
The functional diversity of benthic macroinvertebrate communities in the 18 streams of the Cangshan Mountains was assessed using five indices: functional dispersion (FDis), functional differentiation (FDiv), functional evenness (FEve), functional richness (FRic), and RaoQ. Correspondingly, taxonomic diversity was described using the Shannon–Wiener index (H’), Margalef diversity index (dM), and Pielou evenness index (J). Across all streams, H’ ranged from 0.23 to 2.94, dM from 1.61 to 5.72, and J from 0.12 to 0.94. Functional diversity metrics showed similar variation, with FDis ranging from 1.14 to 4.29, FDiv from 0.01 to 0.36, FEve from 1.01 to 0.54, FRic from 2.27 to 24.69, and RaoQ from 0.27 to 1.98 (Figure 5). Seasonal comparisons revealed that H’, dM, and J were generally higher in the wet season than in the dry season, indicating increased species richness, taxonomic diversity, and community evenness under wetter hydrological conditions. Notably, some streams such as JX and WHX maintained relatively high diversity and evenness values even during the dry season, based on visual trends, suggesting localized environmental stability or buffering capacity. These patterns support the interpretation that seasonal hydrological fluctuations play a critical role in shaping the structure and function of stream macroinvertebrate communities.
FDis indicates the distribution range of the functional traits of a species in the niche space. As shown in Figure 5i, most streams exhibit a higher FDis index in the wet season than in the dry season. Additionally, WHX, LQX, and HLX exhibited the opposite trend, whereas JX and BSX maintained a high FDis index in both wet and dry seasons. FDiv indicates the distribution range of species’ functional traits in the niche space. In contrast, Figure 5j shows that the values between the two seasons are not significantly distinct for most streams; all streams except TMX exhibited a slightly higher FDiv index during the wet season. FEve reflects the relative richness and even distribution of species’ functional traits. Another pattern emerged in Figure 5k, FEve indicates a more complex seasonal pattern among different streams, with MYX, JX, and BSX exhibiting a higher FEve index in the dry season. FRic indicates the size of the functional feature space occupied by the community. Unlike the previous indices, Figure 5l illustrates that most streams exhibit significant differences between the wet and dry seasons, with the functional richness index generally higher in the wet season. However, some streams, such as YX, ZHX, and QBX, have a higher FRic index in the dry season. The RaoQ index, a diversity index based on the functional trait distance between species, indicates the average level of species functional trait diversity in a community. Figure 5m, meanwhile, shows that most streams have higher RaoQ values in the wet season than in the dry season. However, WHX, JX, BHX, and TMX exhibited an opposite trend.
Taken together, the five functional diversity indices indicate that most streams exhibited higher diversity indices in the wet season, suggesting that the wet season is more conducive to the expression of functional diversity and species-level functional differentiation (Table 4). Nevertheless, each stream has its unique situation, indicating that the complexity of environmental factors, species interactions, and ecological adaptation strategies affects the functional diversity of a community.
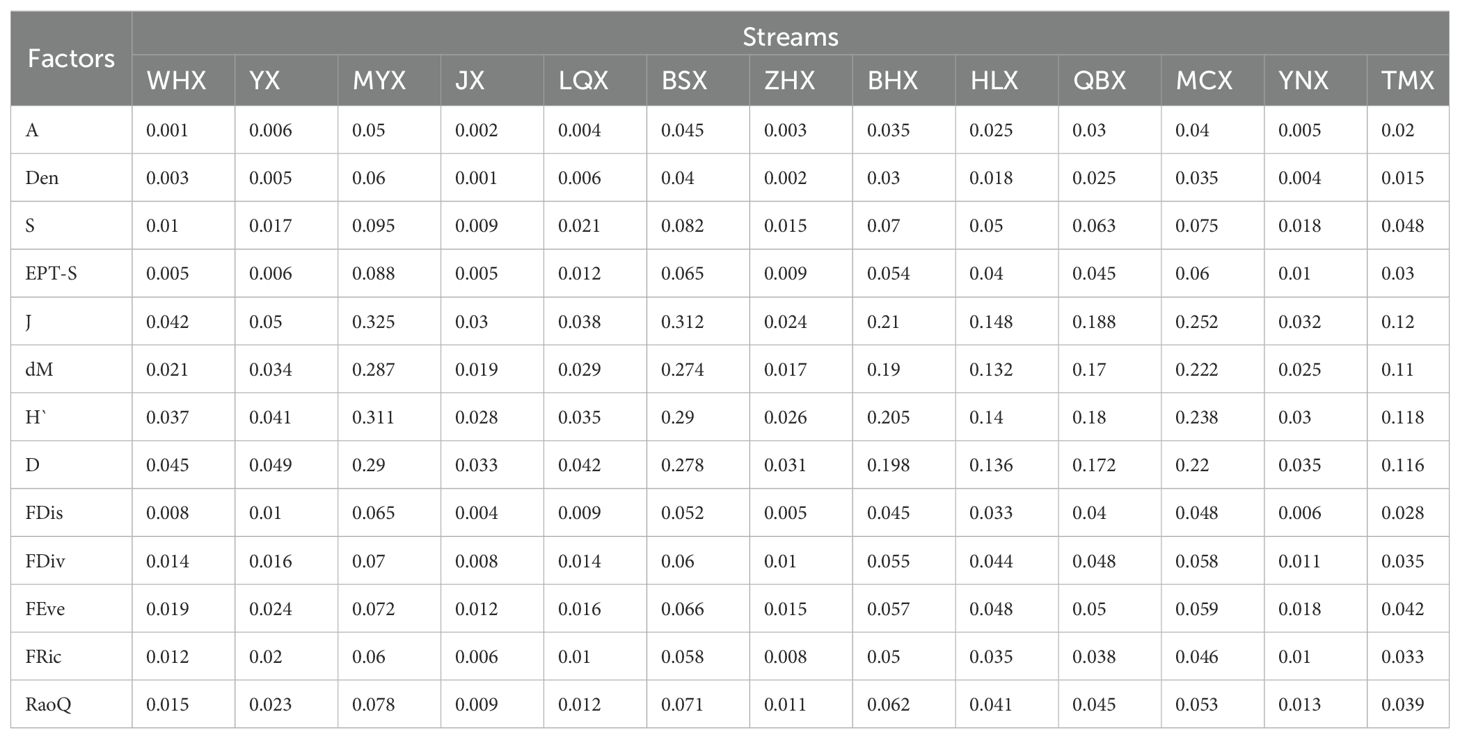
Table 4. The results of paired t-tests for biological factors of each stream in dry and wet seasons.
3.3 Correlation analysis between environmental factors and benthic fauna
Figure 6 shows the significance of the linear mixed-effects model of environmental factors and biodiversity at the 95% confidence interval. As shown in Figure 6a, in the dry season analysis, water depth, conductivity, COD, and pH exhibit significant positive effects on several ecological diversity indicators. Specifically, flow velocity and COD exhibit significant positive correlations with community structure, whereas temperature, turbidity, and water depth exhibit significant positive correlations with species diversity. Additionally, pH, conductivity, and water depth exhibit significant positive correlations with functional diversity. Notably, DO in the dry season data shows a lower correlation with multiple indicators.
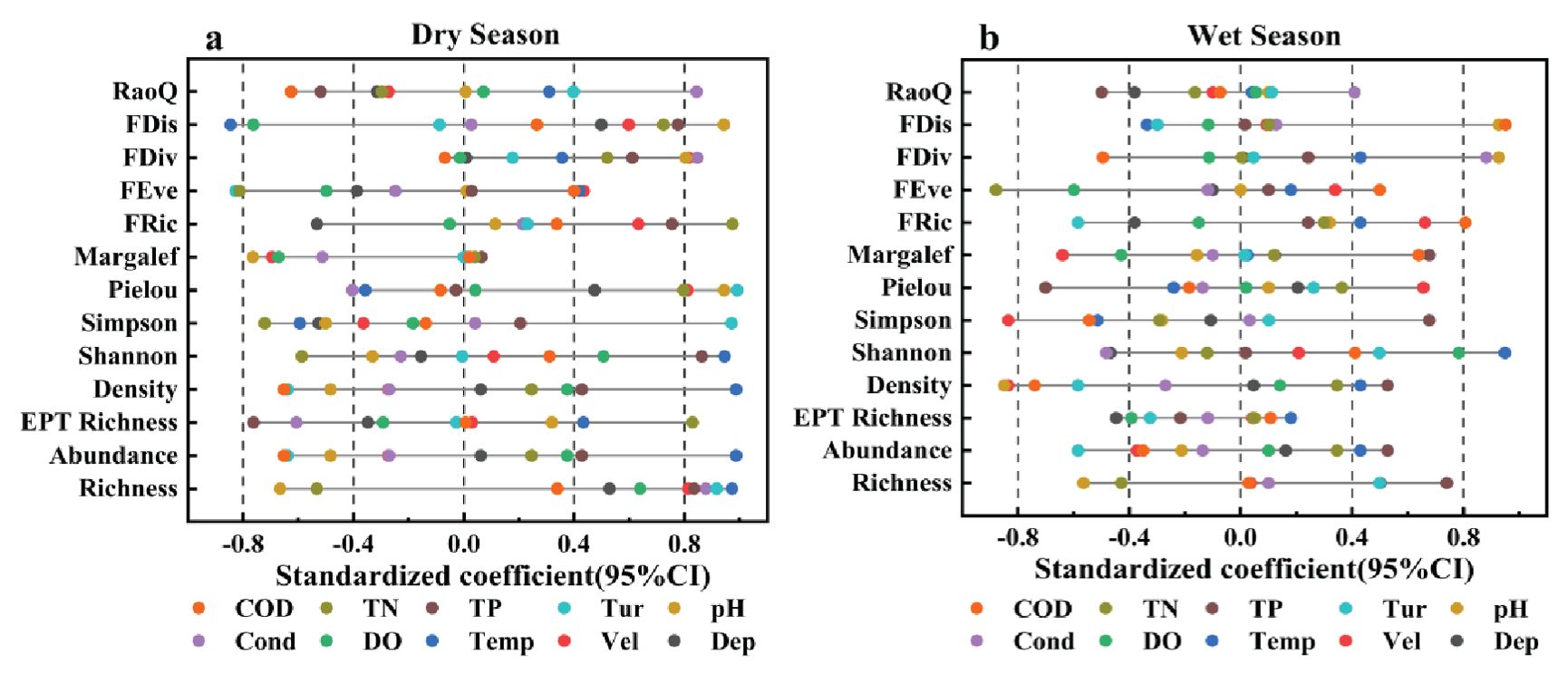
Figure 6. Correlation coefficients of the linear mixed-effects model between environmental factors and benthic community structure and diversity within a 95% confidence interval: (a) results for the dry season and (b) results for the wet season.
For the wet season, the results showed that turbidity, water temperature, flow velocity, and water depth emerge as significant positive factors affecting multiple ecological diversity indicators. Flow velocity and COD exhibit significant positive correlations with community structure, whereas turbidity, DO, water temperature, and conductivity exhibit significant positive correlations with species diversity. Additionally, TN, pH, and water depth exhibit significant positive correlations with functional diversity. Furthermore, although water temperature’s influence on ecological diversity in the wet season remains important, its influence was slightly lower than that in the dry season. Conversely, the effect of DO on biodiversity in the wet season was more significant than that in the dry season. By comparing the data of the dry and wet seasons, we found the effects of water quality parameters on ecological diversity during different seasons. The positive influence of temperature and turbidity during the dry season was more distinct. On the other hand, the wet season highlighted the importance of DO. Nevertheless, both seasons showed that pH was negatively associated with biodiversity in both seasons, suggesting that extreme pH values may adversely affect aquatic community structure. Altogether, these analyses reveal a complex relationship between biodiversity and environmental factors.
4 Discussion
4.1 Characteristics of the water environment of Cangshan streams
The 18 streams in Cangshan Mountain, located in Dali City, Yunnan Province, represent a typical plateau stream system. Their water quality and environmental characteristics are influenced by several natural and human factors. In the present study, we have performed a detailed analysis of the major factors affecting the water environment, namely water temperature, depth, turbidity, flow rate, EC, pH, TN, TP, COD, and DO. We have also reported the seasonal variations, correlations, and principal components of these factors.
We found that most environmental factors increase in the wet season than in the dry season. This may be attributed to the precipitation and changes in runoff (Alves et al., 2024; Xu et al., 2023). Particularly, indicators including COD, EC, TN, and TP exhibit significant differences between the dry and wet seasons across different streams. Streams like YX and TMX exhibited the most pronounced differences between wet and dry seasons, especially in COD and TN concentrations, which increased notably during the dry season. This pattern may be linked to lower dilution capacity, limited baseflow input, and higher pollutant accumulation due to surrounding land use (Liu et al., 2023a). In contrast, streams such as ZHX and HLX showed relatively stable water quality across seasons, possibly due to greater riparian canopy cover and reduced anthropogenic input. These contrasting patterns highlight the influence of watershed-specific factors, including land cover, elevation gradient, and streamflow regime, in mediating seasonal water quality responses (Alves et al., 2024; Baldrighi et al., 2023).
Correlation heatmap analysis showed the differences in the relationships between environmental factors in the dry and wet seasons. During the dry season, a significant positive correlation was found between water depth and DO, which decreased during the wet season. This change may be attributed to altered hydrological conditions, as increased water levels during the wet season influence the vertical mixing and diffusion of DO (Ntislidou et al., 2023). Furthermore, EC is positively correlated with TN in the dry season but is negatively correlated in the wet season. This shift highlights distinct seasonal drivers of nutrient dynamics. During the dry season, reduced flow and dilution lead to nitrogen accumulation and elevated EC, particularly in streams affected by runoff or pollution. In contrast, wet-season rainfall and surface runoff dilute solutes and redistribute nutrients, weakening or reversing the EC–TN relationship (Baldrighi et al., 2023; Kaiser et al., 2023). These changes influence benthic communities by promoting tolerant taxa under dry-season stress and enhancing biodiversity in the more favorable wet-season conditions. Understanding such patterns is key to interpreting seasonal environmental filtering effects on community structure.
Principal component analysis (PCA) was used to identify the main environmental drivers of water quality variation across seasons. In the dry season, electrical conductivity (EC) and total nitrogen (TN) were the dominant contributors to PC1, indicating that ionic concentration and nutrient accumulation are key factors shaping water chemistry under low-flow conditions. This pattern likely reflects reduced dilution and increased anthropogenic inputs, such as agricultural runoff or wastewater discharge, which are more concentrated during periods of limited hydrological connectivity (Hu et al., 2024). In contrast, PC1 in the wet season was strongly influenced by flow velocity and pH, suggesting that increased hydrodynamics and surface runoff during rainfall events become major determinants of water chemistry. For PC2, COD and TP were the primary drivers in the dry season, highlighting the influence of organic pollution and phosphorus enrichment under restricted water exchange. Meanwhile, water depth and dissolved oxygen (DO) dominated PC2 in the wet season, reflecting enhanced mixing and oxygenation due to higher water levels. These seasonal shifts indicate that the key stressors and ecological processes regulating water quality differ markedly between hydrological periods (Kesti et al., 2024; Zemo et al., 2023). Similar patterns have been reported in other subtropical stream systems, where seasonal changes in flow regime strongly modulate nutrient dynamics and physicochemical properties (Wolfe et al., 2023). While human activities such as agriculture and urbanization are known to affect seasonal water quality (Hanafi-Portier et al., 2024; Tian et al., 2020), direct land-use data were not collected in this study. However, the observed variability in nutrient concentrations among streams may be partially influenced by surrounding land use. Future studies incorporating watershed-level land cover and pollution source mapping would help clarify the specific contributions of human activities to water quality variation in the Cangshan region (Sun et al., 2022; Varadinova et al., 2023; Yang and Yang, 2019). Integrating remote sensing and GIS-based catchment analysis could further enhance spatial resolution and improve the accuracy of anthropogenic impact assessments.
To summarize, the dynamic changes in environmental factors between dry and wet seasons indicate the combined effects of natural conditions and human activities (Farooq and Siddiqui, 2020; Ntislidou et al., 2023; Xu et al., 2023; Zemo et al., 2023). Increased rainfall in the wet season has a favorable effect on water quality, increasing the dilution capacity of the water body and the rate of decomposition of organic matter, thereby reducing the concentration of pollutants (Liu et al., 2024). In the dry season, due to the decrease in precipitation and runoff, the dilution capacity of the water body decreases, and the decomposition rate of organic matter decreases; thus, the concentration of pollutants increases (Hu et al., 2024). These seasonal changes in water environmental characteristics not only affect water quality status but also have profound effects on the structure and function of benthic communities.
4.2 Characteristics of the benthic animal community structure in the 18 streams of Cangshan Mountains and its disturbance factors
The structure of the benthic community is a crucial indicator of the ecological health of a watershed. Its diversity and richness indicate the quality of the water environment and the stability of the ecological system (Magni et al., 2023; Okoro et al., 2024; Prayoga et al., 2023). The benthic community structure of the 18 streams of Cangshan Mountains represents a typical highland stream, which is influenced by various natural and human factors, such as seasonal changes, land use changes, increased agricultural activities, and other direct or indirect effects of human activities (Gao et al., 2023; Wilden et al., 2024).
A total of 13,521 benthic macroinvertebrates were collected, representing 3 phyla, 6 classes, 10 orders, 28 families, and 56 taxonomic units, indicating high community diversity. To elucidate the mechanisms driving seasonal community dynamics, we applied a trait-based approach, which links species-specific functional traits to environmental responses. This framework highlights how traits such as dispersal capacity, life cycle strategies, and habitat preferences shape resilience to environmental variability and serve as bioindicators of ecosystem health. Seasonal shifts in rainfall and temperature strongly influenced community composition. During the wet season, increased flow and lower water temperatures provided favorable habitat conditions that supported the dominance of species such as Baetis and Gammarus. These taxa possess traits such as high dispersal ability, rapid life cycles, and tolerance to moderate flow disturbance, allowing them to thrive in dynamic hydrological conditions (Arcifa et al., 2023). Baetis nymphs, for example, are agile swimmers and grazers that benefit from increased periphyton growth under higher nutrient conditions, while Gammarus species are detritivores capable of exploiting leaf litter and organic debris transported during rain events (Hroch and Brabec, 2022; Bae et al., 2023). In contrast, the dry season was characterized by reduced water levels and elevated temperatures, which led to a decline in the abundance of several sensitive taxa. However, species such as Rheotanytarsus became dominant, likely due to their adaptations to low-oxygen conditions, fine sediments, and stable microhabitats (Novichkova et al., 2023). These seasonal shifts reflect the role of species-specific functional traits in shaping community responses to environmental variability, emphasizing the importance of trait-based approaches in understanding benthic community dynamics.
Species abundance and density are usually higher in the wet season than in the dry season. This can be attributed to increased rainfall and water flow, which wash nutrients from farmland into the streams, increasing the growth of benthic animals and depositing more organic matter, thereby increasing species abundance and density (Liu et al., 2023b; Varadinova et al., 2023). Furthermore, abundant streambank vegetation during the wet season filters rainwater by trapping suspended sediments and reducing nutrient inputs (e.g., nitrogen and phosphorus) before it enters the stream, thereby improving water quality and supporting aquatic biodiversity (Lv et al., 2016; Wang et al., 2023). However, in the YX and YNX streams, species abundance and density were high during the dry season, which may be attributed to the good water supply or groundwater replenishment of these streams, providing a favorable habitat for benthic animals (Gao et al., 2022; Lin et al., 2022; Miernik et al., 2023; Xiang et al., 2022). EPT richness serves as an indicator of water quality health (Liu et al., 2024; Xiang et al., 2020; Zhang et al., 2019). In this study, most streams exhibited higher EPT richness during the wet season, likely due to improved hydrological conditions, increased oxygen availability, and reduced environmental stress. However, some streams, particularly ZHX and BHX, showed relatively high EPT richness even during the dry season. This may indicate that these streams experience lower anthropogenic pressure or possess stable microhabitats that buffer seasonal stress. ZHX, for example, is located in a relatively forested sub-watershed with minimal agricultural runoff, while BHX benefits from upstream vegetation cover and shading, which can help maintain cooler temperatures and higher oxygen levels in the dry season (Arcifa et al., 2023; Li et al., 2018; Wang et al., 2019). These findings suggest that high EPT richness in the dry season may reflect either effective watershed management or intrinsic environmental resilience. As such, EPT-based assessments can be a valuable tool not only for detecting broad seasonal trends but also for identifying streams with exceptional ecological conditions or conservation potential.
Most stream benthic organisms exhibit higher species and functional diversity indices during the wet season. This reflects the uniform distribution of species and increased richness during the wet season and more favorable environmental conditions for the expression of functional diversity and species-specific functional differentiation (Gusmao et al., 2024; Liu et al., 2024). The wet season rainfall provides suitable habitats, promoting increased biodiversity, whereas agricultural drainage during this period brings rich nutrients, indirectly increasing functional diversity (Bhaumik et al., 2023; Giampaoletti et al., 2023). During the dry season, water volume decreases and water temperature reduces rapidly; some species cannot adapt to this, leading to a decline in diversity indices (Miernik et al., 2023; Novichkova et al., 2023). Furthermore, agricultural activities decrease during the dry season. Additionally, some functional types of species may decrease due to nutrient deficiency (Varadinova et al., 2023; Wilden et al., 2024; Yang and Yang, 2019). Overall, the wet season is conducive to increased biodiversity, but the specific conditions of each stream vary, reflecting the complexity of environmental factors and species interactions.
The analysis of linear mixed effects model reveals the influence of environmental factors on biodiversity. Owing to the significant difference in environmental factors between the dry season and the wet season, biodiversity is affected differently (Magni et al., 2023; Tian et al., 2020). The model identified several statistically significant predictors, with different drivers dominating in dry and wet seasons. In the dry season, water depth and chemical variables such as EC, TN, and COD were significantly associated with reduced biodiversity, likely due to concentrated pollutants and habitat contraction under low-flow conditions (Alves et al., 2024; Arcifa et al., 2023; Kaiser et al., 2023; Kazikina, 2022). This suggests that nutrient accumulation and limited habitat heterogeneity are key stressors during dry periods. In contrast, the wet season was mainly influenced by physical variables, including flow velocity, turbidity, and water temperature, all of which were significantly linked to increased biodiversity. Enhanced hydrological activity during this season supports greater environmental heterogeneity and resource availability, which favors the coexistence of diverse benthic taxa (Ntislidou et al., 2023). These findings underscore the importance of season-specific biodiversity drivers and highlight the need for integrated ecosystem management. For example, reducing nutrient inputs during the dry season and maintaining riparian vegetation to buffer runoff in the wet season could help sustain benthic diversity (Farooq and Siddiqui, 2020; Liu et al., 2023a, 2024; Sun et al., 2022; Tian et al., 2020; Xu et al., 2023).
However, it should be noted that the observed positive effects of the wet season may not be consistent across all ecosystems. In other regions, increased rainfall can enhance surface runoff and introduce diffuse pollutants, such as pesticides, heavy metals, and excess nutrients, which may degrade water quality and reduce benthic diversity (Lv et al., 2016; Prayoga et al., 2023). Additionally, excessive flow during heavy rain events can destabilize streambeds, disrupt habitats, and displace organisms, resulting in lower abundance and diversity (Giampaoletti et al., 2023). Therefore, the ecological response to seasonal changes depends on a combination of local hydrological regimes, land use patterns, and watershed management practices. Recognizing these potential risks is essential to avoid overgeneralization and to inform site-specific conservation strategies.
In general, favorable environmental conditions in the wet season promoted the diversity and functional diversity of benthic communities, whereas unfavorable environmental conditions in the dry season limited the maintenance and growth of biodiversity. These results not only enhance our understanding of the dynamic changes in the water ecosystem in the 18 streams, Cangshan but also provide a scientific basis for formulating effective seasonal protection and management strategies for similar water ecosystems.
However, it is important to acknowledge that, while this study provides valuable insights into the seasonal dynamics of benthic biodiversity and its responses to environmental factors, it is based on sampling from only a single wet season and a single dry season, lacking true seasonal replication. This limitation constrains the extent to which the observed seasonal patterns can be generalized across different years. To enhance the robustness of such findings and more fully capture interannual variability in environmental conditions and community responses, future studies should incorporate multi-year seasonal monitoring.
5 Conclusion
This study systematically examined the seasonal dynamics of benthic macroinvertebrate communities across 18 plateau streams in the Cangshan Mountains and elucidated how seasonal shifts in environmental factors regulate community structure and biodiversity. The results indicated that water temperature, dissolved oxygen, electrical conductivity, and flow velocity were the primary drivers of benthic diversity, with their influence varying markedly between the dry and wet seasons. Enhanced hydrological conditions during the wet season supported higher diversity, while intensified environmental stress during the dry season led to simplified community structures dominated by tolerant taxa such as Rheotanytarsus, known for their adaptations to low dissolved oxygen, high organic content, and fine sediment substrates.
Moreover, the study identified distinct seasonal shifts in the dominant environmental drivers. In the dry season, biodiversity was mainly influenced by physicochemical factors such as water depth, electrical conductivity, and nutrient concentrations (e.g., TN and COD), whereas physical variables like flow velocity, water temperature, and turbidity played a more substantial role during the wet season. These seasonal contrasts significantly shaped the structure and composition of benthic communities. Notably, some streams (e.g., ZHX and BHX) sustained relatively high EPT richness even during the dry season, likely due to reduced anthropogenic disturbances and more stable hydrological and habitat conditions, such as dense riparian vegetation, cooler water temperatures from canopy shading, and relatively stable microhabitats. These spatial differences suggest that local environmental characteristics and land-use pressures are critical in determining ecological resilience under seasonal stress.
The findings offer valuable guidance for seasonal biodiversity conservation and stream ecosystem management. Targeted strategies such as controlling nutrient inputs during the dry season, preserving riparian buffer zones in the wet season, and implementing continuous hydrological monitoring are essential to maintaining ecosystem stability. However, this study was limited by the absence of direct land-use data and long-term temporal observations, which may constrain the interpretation of anthropogenic influences and interannual variability. Future research should incorporate land-use datasets and long-term monitoring to better understand how plateau stream ecosystems respond to the compounded effects of climate change and human activities, potentially through remote sensing or GIS-based land cover assessments.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
YF: Data curation, Formal analysis, Investigation, Software, Visualization, Writing – original draft, Writing – review & editing. MY: Conceptualization, Formal analysis, Investigation, Methodology, Project administration, Writing – review & editing. HC: Conceptualization, Resources, Supervision, Validation, Writing – review & editing. FT: Investigation, Project administration, Visualization, Writing – original draft. XL: Conceptualization, Methodology, Validation, Writing – review & editing. JS: Investigation, Methodology, Writing – review & editing. HY: Conceptualization, Funding acquisition, Project administration, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research is financially supported by the National Natural Science Foundation of China Youth Fund (32101310), Introduced Talent Scientific Research Start-up Fund of Yunnan University (CZ21623201) and China Postdoctoral Science Foundation (2021M692728).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alves C. M., Gomes M. A., S. Troncoso J., and Gomes P. T. (2024). Structural and functional diversity patterns of macrofaunal communities from a semi-enclosed inlet of Northeast Atlantic: The influence of environmental conditions. Estuarine Coastal Shelf Sci. 296, 108575. doi: 10.1016/j.ecss.2023.108575
Arcifa M. S., Coelho P. N., da Silva L. H. S., and Righi-Cavallaro K. O. (2023). Impacts of small dams on the phytoplankton and invertebrates in an Amazonian river. River Res. Appl. 39, 661–674. doi: 10.1002/rra.v39.4
Bae M. J., Hwang Y., Ham S. N., Kim S. Y., and Kim E. J. (2023). Community recovery of benthic macroinvertebrates in a stream influenced by mining activity: Importance of microhabitat monitoring. Environ. Res. 234, 116499. doi: 10.1016/j.envres.2023.116499
Baldrighi E., Pizzini S., Punzo E., Santelli A., Strafella P., Scirocco T., et al. (2023). Multi-benthic size approach to unveil different environmental conditions in a Mediterranean harbor area (Ancona, Adriatic Sea, Italy). PeerJ Inc 11, e15541. doi: 10.7717/peerj.15541
Bhaumik S., Mandal S., and Tripathy S. C. (2023). Unraveling the functional diversity of macrobenthic community from Prydz Bay, Indian sector of the Southern Ocean. Continental Shelf Res. 263, 105043. doi: 10.1016/j.csr.2023.105043
Chen D., Yang Z., Zeng Q., Wang W., Yan L., Zhang P., et al. (2023). Analysis of the suitable ecological flow of benthic animals in the lower reaches of Xiangjiaba Reservoir in the upper reaches of the Yangtze River based on the physical habitat model. J. Hydrology 625, 130132. doi: 10.1016/j.jhydrol.2023.130132
Dufrêne M. and Legendre P. (1997). Species assemblages and indicator species: the need for A flexible asymmetrical approach. Ecol. Monogr. 67, 345–366. doi: 10.2307/2963459
Farooq S. and Siddiqui P. J. A. (2020). Assessment of three mangrove forest systems for future management through benthic community structure receiving anthropogenic influences. Ocean Coastal Manage. 190, 105162. doi: 10.1016/j.ocecoaman.2020.105162
Gao Y., Rong L., Cao L., Li K., Lin C., Zhang Z., et al. (2022). Temporal changes in headwater streams macroinvertebrate assemblages during the snowmelt season in northeast China. Front. Environ. Sci. 10. doi: 10.3389/fenvs.2022.960254
Gao Q., Zhang Q., Zeng J., Yin Z., Liu J., Liu G., et al. (2023). Macroinvertebrate community structure, pollution tolerance, diversity and feeding functional groups in polluted urban rivers under different black and odorous levels. Ecol. Indic. 156, 111148. doi: 10.1016/j.ecolind.2023.111148
Giampaoletti J., Sbrana A., Magni P., and Gravina M. F. (2023). Macrobenthos of the tortolì Lagoon: A peculiar case of high benthic biodiversity among mediterranean lagoons. Diversity 15 (6), 783. doi: 10.3390/d15060783
Golovatyuk L. V., Nazarova L. B., Kalioujnaia I. J., and Grekov I. M. (2023). Taxonomic composition and salinity tolerance of macrozoobenthos in small rivers of the southern arid zone of the east european plain. Biology 12 (9), 1271. doi: 10.3390/biology12091271
Gusmao J. B., Rühmkorff S., Kraufvelin L., Meysick L., and Pansch C. (2024). The interplay of co-occurring ecosystem engineers shapes the structure of benthic communities – a mesocosm experiment. Front. Marine Sci. 11. doi: 10.3389/fmars.2024.1304442
Hanafi-Portier M., Samadi S., Corbari L., Boulard M., Miramontes E., Penven P., et al. (2024). Multiscale spatial patterns and environmental drivers of seamount and island slope megafaunal assemblages along the Mozambique channel. Deep Sea Res. Part I: Oceanographic Res. Papers 203. doi: 10.1016/j.dsr.2023.104198
He Y., Wang H., Zhao Y., and Cui Y. (2021). Predicting the biodiversity hotspots of macrozoobenthos in the Yangtze river basin. Ecol. Indic. 133, 108428. doi: 10.1016/j.ecolind.2021.108428
He F., Wu N., Dong X., Tang T., Domisch S., Cai Q., et al. (2020). Elevation, aspect, and local environment jointly determine diatom and macroinvertebrate diversity in the Cangshan Mountain, Southwest China. Ecol. Indic. 108, 105618. doi: 10.1016/j.ecolind.2019.105618
Hroch M. and Brabec K. (2022). Analysis of multiple-pressure pattern in rivers and its effects on the structure of macroinvertebrate communities. Limnologica 97, 126027. doi: 10.1016/j.limno.2022.126027
Hu L.-S., Zhang X.-M., Li S.-W., Li F., and Dong Y.-W. (2024). Water-sediment regulation affects community structures of macrobenthic mollusks in the Yellow River estuary and adjacent waters. Regional Stud. Marine Sci. 71, 103428. doi: 10.1016/j.rsma.2024.103428
Kaiser S., Brandt A., Brix S., Brenke N., Kürzel K., Arbizu P. M., et al. (2023). Community structure of abyssal macrobenthos of the South and equatorial Atlantic Ocean - Identifying patterns and environmental controls. Deep Sea Res. Part I: Oceanographic Res. Papers 197, 104066. doi: 10.1016/j.dsr.2023.104066
Kazikina S. M. (2022). The ecological characteristics of small rivers in the Shilka river basin by zoobenthos organisms. IOP Conf. Series: Earth Environ. Sci. 962, 012018. doi: 10.1088/1755-1315/962/1/012018
Kesti P., Strandberg U., Taipale S., Hiltunen M., Vesterinen J., Vainikka A., et al. (2024). Water quality shapes the community structure of zoobenthos in rivers but only has a minor effect on the fatty acid composition of zoobenthos and salmonids. River Res. Appl., 291-292. doi: 10.1002/rra.v40.3
Li R., Li X., Yang R., Farooq M., Tian Z., Xu Y., et al. (2024). Bioassessment of macroinvertebrate communities influenced by gradients of human activities. Insects 15 (2), 131. doi: 10.3390/insects15020131
Li K., Zhang Z., Yang H., Bian H., Jiang H., Sheng L., et al. (2018). Effects of instream restoration measures on the physical habitats and benthic macroinvertebrates in an agricultural headwater stream. Ecol. Eng. 122, 252–262. doi: 10.1016/j.ecoleng.2018.08.007
Lin C., Cao L., Xu Y., Zhang Z., Yang H., Xiang H., et al. (2022). Spatio-temporal variability (up- and down-stream and three seasons) of response patterns of macroinvertebrates to commonly applied low-head concrete weirs under reclaimed water intervention in urban river restoration. J. Cleaner Production 370, 133544. doi: 10.1016/j.jclepro.2022.133544
Liu H., Cheng W., Xiong P., Li H., Liu Z., Ai J., et al. (2023a). Temporal variation of plankton and zoobenthos communities in a freshwater reservoir: Structure feature, construction mechanism, associated symbiosis and environmental response. Ecol. Indic. 154, 110774. doi: 10.1016/j.ecolind.2023.110774
Liu Q., Liao Y., Zeng J., Tang Y., Zhang R., Liu Q., et al. (2024). Decadal changes of macrofauna community in a semi-enclosed Bay of Yueqing in East China Sea. Marine Pollution Bull. 201, 116239. doi: 10.1016/j.marpolbul.2024.116239
Liu H., Xu H., Tang Y., Wang Z., Liu Q., Zhang R., et al. (2023b). Middle-scale spatial and temporal patterns in estuaries: rocky intertidal macrobenthic communities in the hangzhou bay and zhoushan sea. Estuaries Coasts 46, 1302–1315. doi: 10.1007/s12237-023-01200-2
Lv W., Liu Z., Yang Y., Huang Y., Fan B., Jiang Q., et al. (2016). Loss and self-restoration of macrobenthic diversity in reclamation habitats of estuarine islands in Yangtze Estuary, China. Marine Pollution Bull. 103, 128–136. doi: 10.1016/j.marpolbul.2015.12.030
Magni P., Vesal S. E., Giampaoletti J., Como S., and Gravina M. F. (2023). Joint use of biological traits, diversity and biotic indices to assess the ecological quality status of a Mediterranean transitional system. Ecol. Indic. 147, 109939. doi: 10.1016/j.ecolind.2023.109939
Miernik N. A., Janas U., and Kendzierska H. (2023). Role of macrofaunal communities in the vistula river plume, the baltic sea—Bioturbation and bioirrigation potential. Biology 12 (2), 147. doi: 10.3390/biology12020147
Ni C., Chen Y., Hu X., and Feng J. (2023). Vegetation change and conservation evaluation of the cangshan erhai national nature reserve (Cangshan mountain part) in southwest China. Forests 14 (7), 1485. doi: 10.3390/f14071485
Novichkova A. A., Borisov R. R., Vorobjeva L. V., Palatov D. M., Chertoprud M. V., and Chertoprud E. S. (2023). The influence of salinity gradient and island isolation on fauna composition and structure of aquatic invertebrate communities of the shantar islands (Khabarovsk krai). Diversity 15 (2), 1198. doi: 10.3390/d15121198
Ntislidou C., Latinopoulos D., Skotida A., Giannoulis T., Moutou K., and Kagalou I. (2023). Assessment of hydrological barriers effect in river benthic fauna coupled with eDNA metabarcoding monitoring. Ecohydrology Hydrobiology 23, 389–399. doi: 10.1016/j.ecohyd.2023.04.007
Okoro C. M., Zabbey N., Hart A. I., Nwipie G. N., Ibim A. T., and Nkeeh D. K. (2024). Assessment of macrozoobenthos baseline diversity for monitoring the ecological quality of Finima Nature Park Lake. Environ. Monitoring Assess. 196, 148. doi: 10.1007/s10661-024-12325-7
Oyeku O. G., Anyaele O. O., Akindele E. O., Atobatele O. E., and Adeniyi A. V. (2023). Biological water quality of an impaired tropical river: the macrozoobenthos approach. Biologia 78, 2131–2145. doi: 10.1007/s11756-023-01346-1
Prayoga D., Hasan Z., Dewanti L. P., and Herawati H. (2023). Assesing the community structure of macrozoobenthos in cibeureum lake, tasikmalaya, west java, Indonesia. Asian J. Fisheries Aquat. Res. 25, 26–39. doi: 10.9734/ajfar/2023/v25i1655
Sun Y.-X., Li X.-X., Tan Y., Wang J., and Dong Y.-W. (2022). Microhabitat thermal environment controls community structure of macrobenthos on coastal infrastructures. Estuarine Coastal Shelf Sci. 277, 108060. doi: 10.1016/j.ecss.2022.108060
Sun C., Xia L., Zhang M., He Q., Yu N., Xiang H., et al. (2024). The impacts of different seasons on macroinvertebrate community structure and functional diversity in the Jingui River, China. Global Ecol. Conserv. 51, e02876. doi: 10.1016/j.gecco.2024.e02876
Tian G. Z., Xu Y. M., Gao Y., and Wang Y. (2020). Ecological environment of erhai and lashihai watersheds of the ancient tea horse road on the yunnan-tibet line. J. Coastal Res., 401–404. doi: 10.2112/JCR-SI115-114.1
Varadinova E., Gecheva G., Tyufekchieva V., and Milkova T. (2023). Macrophyte- and macrozoobenthic-based assessment in rivers: specificity of the response to combined physico-chemical stressors. Water 15 (2), 2282. doi: 10.3390/w15122282
Wang L., Gao Y., Han B.-P., Fan H., and Yang H. (2019). The impacts of agriculture on macroinvertebrate communities: From structural changes to functional changes in Asia's cold region streams. Sci. Total Environ. 676, 155–164. doi: 10.1016/j.scitotenv.2019.04.272
Wang S. and Wang J. (2025). Cobalt-silicon coordination-induced nonradical activation of peroxymonosulfate for enhancing the degradation of organic pollutants in real wastewater. Small 21, 2500434. doi: 10.1002/smll.202500434
Wang L., Xia L., Li J., Wan L., and Yang H. (2023). Winter dynamics of functional diversity and redundancy of riffle and pool macroinvertebrates after defoliation in a temperate forest stream. Front. Microbiol. 14, 1105323. doi: 10.3389/fmicb.2023.1105323
Wilden B., Majdi N., and Traunspurger W. (2024). Global resemblance, local divergence? – A comparison of meiobenthic invertebrate communities dwelling in ancient lakes Malawi, Ohrid and Baikal. J. Great Lakes Res. 50, 102305. doi: 10.1016/j.jglr.2024.102305
Wolfe K., Kenyon T. M., Desbiens A., de la Motte K., and Mumby P. J. (2023). Hierarchical drivers of cryptic biodiversity on coral reefs. Ecol. Monogr. 93. doi: 10.1002/ecm.1586
Xiang H., Cai Q., Li Y., Zhang Z., Cao L., Li K., et al. (2020). Sensors applied for the detection of pesticides and heavy metals in freshwaters. J. Sensors 2020, 1–22. doi: 10.1155/2020/8503491
Xiang H., Li K., Cao L., Zhang Z., and Yang H. (2022). Global patterns and drivers of coniferous leaf-litter decomposition in streams and rivers. Front. Ecol. Evol. 10, 940254. doi: 10.3389/fevo.2022.940254
Xu X., Su G., Zhang P., Wang T., Zhao K., Zhang H., et al. (2023). Effects of multiple environmental stressors on zoobenthos communities in shallow lakes: evidence from a mesocosm experiment. Animals 13 (23), 3722. doi: 10.3390/ani13233722
Yang R. and Yang D. (2019). Traditioanl settlements study on sciences of human settlements in erhai lake region. IOP Conf. Series: Materials Sci. Eng. 592. doi: 10.1088/1757-899x/592/1/012170
Zemo M. A. T., Menbohan S. F., Atchrimi B. T., Betsi W. C. N., Nwaha M., Dzavi J., et al. (2023). Effect of anthropogenic pressure on the biodiversity of benthic macroinvertebrates in some urban rivers (Yaoundé). Water 15 (13), 2383. doi: 10.3390/w15132383
Keywords: dry and wet seasons are distinct, benthic macroinvertebrates, community structure and diversity, water environmental factor, the 18 streams in Cangshan
Citation: Feng Y, Yang M, Chen H, Tang F, Liu X, Shi J and Yang H (2025) Research on the seasonal driving mechanisms of benthic macroinvertebrate communities in streams. Front. Ecol. Evol. 13:1536181. doi: 10.3389/fevo.2025.1536181
Received: 28 November 2024; Accepted: 29 April 2025;
Published: 30 May 2025.
Edited by:
Ovie Edegbene, Federal University of Health Sciences Otukpo, NigeriaReviewed by:
Raphael Ligeiro, Federal University of Pará, BrazilChipo Perseverance Mungenge, Rhodes University, South Africa
Sara El Yaagoubi, Abdelmalek Essaadi University, Morocco
Copyright © 2025 Feng, Yang, Chen, Tang, Liu, Shi and Yang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Haijun Yang, eWFuZ2hhaWp1bkB5bnUuZWR1LmNu
†These authors have contributed equally to this work