 Romain Sordello
Romain Sordello Cindy Fournier1
Cindy Fournier1- 1PatriNat Office Français de la Biodiversité (OFB)- Museum national d'Histoire naturelle (MNHN), Paris, France
- 2EDF – Recherche et Développement, Laboratoire National d’Hydraulique et Environnement (LNHE), Laboratoire National d’Hydraulique et d’Environnement, Chatou, France
Introduction: Nowadays, light pollution due to the multiplication of outdoor lightings has become a worldwide disturbance for many species. A large amount of scientific literature shows that this sensory stressor has adverse effects on several taxa. However, existing reviews tend to conclude a lack of primary research on terrestrial non-flying mammals even if these organisms are highly nocturnal and frequently exposed to artificial light.
Methods: To have a more accurate overview on the available literature, we followed the guidelines proposed by the Collaboration for Environmental Evidence to perform a comprehensive evidence synthesis, focusing on the 186 terrestrial non-flying mammal species native to Europe. We searched 2 academic databases and Google Scholar for scientific and grey literature on the impacts of artificial light. We exported 6624 citations and screened 4866 articles on titles, abstracts, and then full-texts. We coded variables and extracted results for all the included studies to build a systematic database and we synthetized the data.
Results: We collected 48 articles on 22 mammal species across 12 families, revealing a more substantial knowledge than expected. composed of 448 study units. We identified four main research hotspots that address different questions: 1/ impacts of manipulating the photoperiod; 2/ impacts of ex-situ artificial light on small mammals; 3/ impacts of in-situ outdoor lightings; 4/ impacts of global light pollution (sky glow). We described the results of the included studies in a narrative review to provide a factual overview of the findings.
Discussion: These initial results draw detrimental impacts of ALAN on circadian rhythm, behaviour, foraging or movements and ultimately occurrence, which are consistent with available literature on moonlight influence or ALAN effects on other taxa. Hence, we call for much more attention to be paid to wild terrestrial non-flying mammals in relation to this key anthropogenic driver.
1 Introduction
The emissions of artificial light at night (ALAN) have considerably increased globally in recent decades (Falchi et al., 2016; Sánchez de Miguel et al., 2022). This phenomenon is now considered one of the most disruptive anthropogenic pressures to Earth’s natural balance (Urbanski et al., 2012; Ditmer et al., 2021b). Today, the photoperiodic cycle is greatly disturbed having previously been stable for billions of years (Grubisic et al., 2019). Indeed, many species are harmfully affected by the exposure to ALAN, both flora (Bennie et al., 2016) and fauna (Sanders et al., 2021; Yang et al., 2024). Such impacts have been known for a very long time; e.g. the lethal flight-to-light behavior of insects or the delay in leaf fall (Matzke, 1936). However, it was in the 1980s that the literature began to seriously grow (Davies and Smyth, 2018; Rodrigo-Comino et al., 2021). Several reviews on this topic reveal an extensive literature for many biological groups (Svechkina et al., 2020; Falcon et al., 2020; Pérez Vega et al., 2022).
ALAN has been demonstrated to affect different levels of organization of life such as genes (Touzot et al., 2022), population (La Sorte et al., 2017), community (Grenis et al., 2023), ecosystem (Giavi et al., 2021) or landscape (Camacho et al., 2021). Various taxa are deeply investigated, particularly bats among mammals. For instance, we now know that ALAN modifies temporal pattern (Luo et al., 2021), has physiological consequences (Cravens and Boyles, 2019), alters foraging behavior (Zou et al., 2024), disturbs movements (Zeale et al., 2018), unbalances communities (Schoeman, 2016), decreases habitat suitability and connectivity (Haddock et al., 2019) and finally results in a lower diversity (Mena et al., 2022) and reduced species ranges (Azam et al., 2016). Yet, knowledge does not seem to be available to the same extent for terrestrial non-flying mammals. Indeed, systematic reviews and meta-analyses published in recent years all point to a lack or absence of articles on this group of mammals. Falcon et al (2020) published a systematic review on all species and ecosystems, whose results on mammals actually concern mainly rodents (rats, hamsters etc.) and bats, leading the author to conclude that “the potential influence of artificial nighttime light and LEDs on mammals has not been studied in depth and concerns only a limited number of species”. Svechkina et al. (2020) also conducted a systematic review on all taxa but collected no article on mammals apart from rodents, bats and two studies on wallabies. Sanders et al. (2021) performed a meta-analysis on all animals but obtained results only for rodents and birds while, Rodrigo-Comino et al. (2021) found 267 articles on animals among which 67 concern mammals, of which 44 deal with bats. Pérez Vega et al. (2022) carried out a systematic review but none of the 30 studies retrieved on mammals involved ungulates. As a result, available evidence syntheses all conclude that wild terrestrial non-flying mammals are particularly under-examined.
However, these previous reviews and meta-analyses are based on global search strategies (i.e. search strings targeting all species and ecosystems) which could partly explain this lack of results on mammals. Indeed, we are aware of relevant studies that have not been captured by existing reviews (Beier, 1995; Bliss-Ketchum et al., 2016; Ciach and Fröhlich, 2019; Berger et al., 2020). This tends to confirm that a comprehensive literature search focused specifically on terrestrial non-flying mammals - using taxonomic families and species names as searching keywords - would be necessary to better reflect the current state of knowledge.
Mammals are a highly threatened group of vertebrates worldwide (Ceballos and Ehrlich, 2002). Among 1437 mammal species assessed, 177 have lost more than 50% of their range since 1500 (Dirzo et al., 2014). Moreover, mammals are mostly nocturnal and therefore potentially very exposed to ALAN and its effects (Holker et al., 2010). The presence of humans during the day even increases the nocturnality of wild mammals (Gaynor et al., 2018). However, this promising adaptative mechanism for human-wildlife coexistence exposes animals to additional threats is since night-time is also disrupted by artificial lighting, which has negative repercussions (Procko et al., 2023). We know that the home range of most of mammals on Earth are increasingly exposed to ALAN (Duffy et al., 2015). Thus, it seems relevant to have a more comprehensive and accurate vision on the actual state of available knowledge on these species. This will enable us to assess whether the known effects on other taxa, such as bats, are also verified on terrestrial non-flying mammals, whether they are different or whether they are effectively unexplored. Systematic maps and reviews are useful tools to expose available knowledge to researchers and managers (Haddaway et al., 2016; Berger-Tal et al., 2019). Then this work will inform strategic priorities for the scientific community, policymakers, and practitioners.
Thus, we conducted an evidence synthesis using the standardized methods developed by the Collaboration for Environmental Evidence for conducting systematic maps and reviews in ecology, that allows for a high level of exhaustiveness (Livoreil et al., 2017). To further ensure maximum comprehensiveness, we concentrated solely on species native to Europe.
We addressed the following question: what evidence exists on the impacts of artificial light on terrestrial non-flying mammals native to Europe? We pursued several objectives: (1) Assess thoroughly the available literature about the impacts of ALAN of terrestrial non-flying mammals native to Europe; (2) Provide a bibliometric description of this state of knowledge; (3) Identify the main research questions that are supported by the articles (4) State factually the findings of the studies; and (5) Determine the limits, knowledge gaps and perspectives. To this end, we both a systematically mapped the evidence and provided a narrative review of the included studies.
2 Methods
The mapping process (i.e. literature search, screening, coding) was conducted according to the guidelines developed by the ‘Collaboration for Environmental Evidence’ (CEE) to conduct systematic maps. Deviations to these CEE standards are listed in the section ‘Limit of the review’. Moreover, this article conforms to ROSES reporting standards (Haddaway et al., 2018) (see Additional File 1). More detailed on the search strategy and the review process are given in Additional File 2.
2.1 Review question
Our review question is: what evidence exists on the impacts of artificial light on terrestrial non-flying mammals native to Europe?
This research question was broken down into the following elements: Population, Exposure, Comparator and Outcome (PECO) - a framework originally proposed by Richardson et al., 1995 to help researchers structure a literature review question (Richardson et al., 1995) - which are further detailed in the “Eligibility criteria” section (see Table 1). Regarding population, we considered all terrestrial non-flying mammals (i.e excluding marine mammals, bats, and humans) which are European native; which means that we also accepted studies conducted in extra-European countries as long as the species studied is native to Europe. However, we excluded introduced, domesticated or man-created (e.g. laboratory strain such as Wistar rats) species from in-situ or ex-situ populations. As regards to exposure, artificial light was considered (no natural light such as lightning, sunlight, moonlight, etc.), whether provided by indoor (e.g. light treatment in laboratory) or outdoor lightings. We accepted spatial (population exposed vs population not exposed) and/or temporal comparator (before/after exposure) as well as several artificial light treatments (e.g. different durations, compositions, amount). Finally, we included all types of outcomes on the studied population (e.g. physiological, behavioral, ecological).
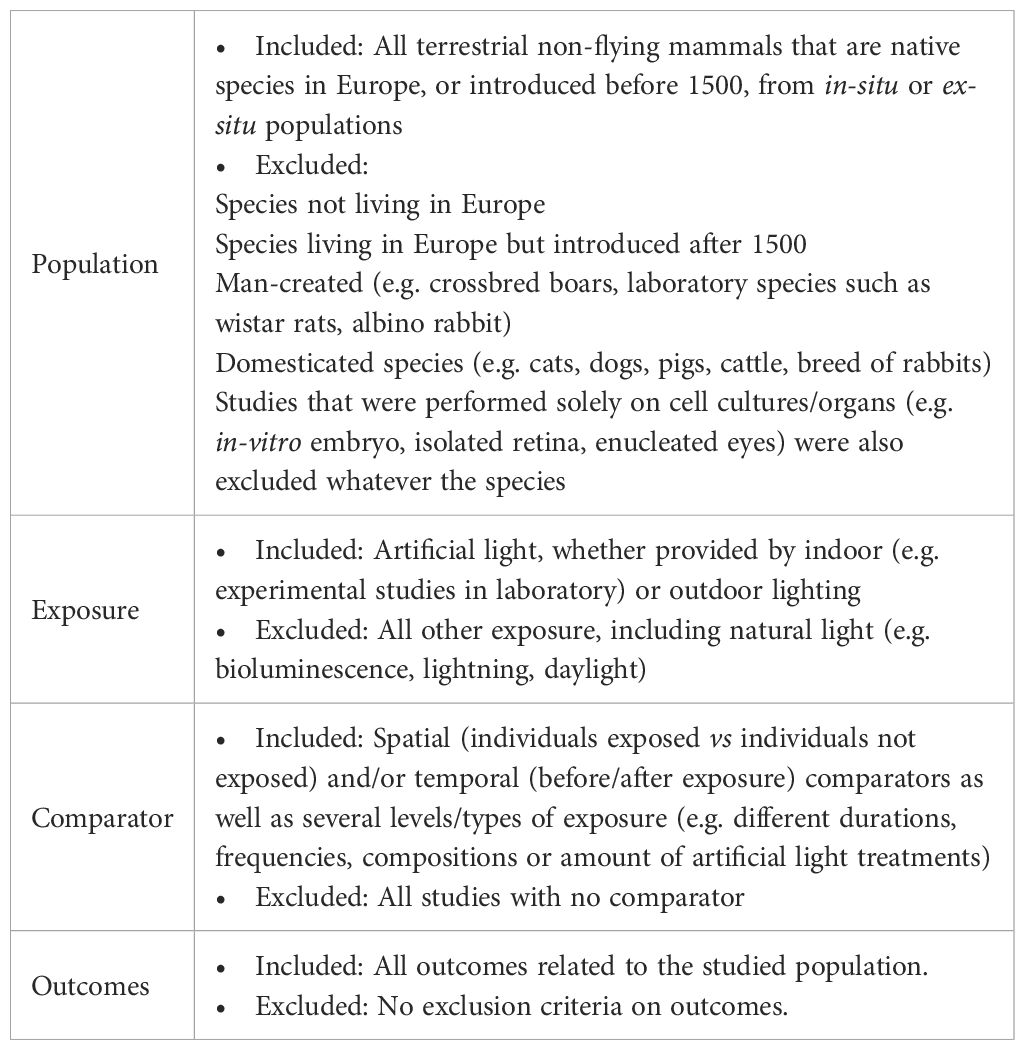
Table 1. Inclusion/exclusion criteria for the three-stage screening process.
2.2 Literature search
Searches were performed using exclusively English search terms presented below. Only studies published in English and in French were included in this systematic map, due to limited resources and the languages understood by the review team.
Our literature search was performed on the Web of Knowledge platform (Clarivate) - in the database “Web of Science Core Collection” - and in Scopus (Elsevier), that both reliable in ecology.
We first listed 186 terrestrial non-flying mammal species that are native in Europe (see Additional File 2 - Section I). Then, a search string was built associating families and species names (see Additional File 2 - Section II) and the term light*:
A first search was run on 16th June 2022 in WOSCC and 17th June 2022 in Scopus and then an updated search was performed on 28th December 2022 for both databases. All articles were merged and duplicates were identified.
2.3 Screening process
All articles were screened through a three-stage process: firstly on titles, then on abstracts, then on full-texts. Screening was performed on predefined criteria (see Table 1) according to the PECO described above. When there was any doubt regarding the presence of a relevant inclusion criterion or insufficient information, articles were retained for assessment at a later stage. Articles included after title screening, but that did not have an abstract were transferred to full-text screening.
Each screening stage was conducted by two or three screeners: CF, RS, TT on titles, CF and RS on abstracts, CF, RS, TT on full-texts. To assess the consistency of the inclusion/exclusion decisions, a Randolph’s Kappa coefficient was computed before screening commenced at each separate stage. To that end, a set of articles was randomly selected (respectively composed of 180 titles, 20 abstracts and 15 full-texts and screened by each reviewer independently (see Additional File 2 - Section III). We fixed 0.7 as an acceptable Kappa coefficient (Frampton et al., 2017). All three Kappa test were valid the first time: we obtained 0.76 for title Kappa value, 0.9 for abstract Kappa value and 0.73 for full-text Kappa value. For the three Kappa test, all disagreements were discussed and resolved before beginning the screening process. Screeners did not screen any article that they would have authored themselves.
At title screening stages, we only excluded articles that did not deal with artificial light nor mammals. At this stage we retained titles on hamsters or rabbits to treat them separately because these two taxa represented a large part of articles. After title screening, included titles were screened on abstract this time according to the full PECO criteria. The two groups of included titles dealing with rabbits or hamsters were refined on title before going to abstract screening in order to eliminate irrelevant populations (laboratory rabbits, non-native hamster species, etc.). Similarly, included titles that had no abstract were also refined according to the title again, to better consider the population (excluding species not living in Europe, domesticated species, man-created species, etc.) before going to full-text screening. At full-text screening, we added new criteria, concerning language (only documents written in French or English were included) and document type (we included journal article, chapter books or books and rejected conference object such as meeting abstracts, notes, news, editorial and all other types of documents).
2.4 Additional literature sources
To complete our search strategy, in particular to increase the probability to get grey literature: 1/We searched the web-based search engine, Google Scholar, with a simplified search string [mammal$ AND light*], using the software Publish or Perish (Harzing, 2007). Searches were performed on the 4th of January 2023 and resulted in the retrieval of 57 citations that were added to the citations retrieved by the two academic databases(WOSCC and Scopus) then screened with an identical method; 2/Backward citation chasing - the process of obtaining records cited within the reference lists of a set of articles – was undertaken relevant meta-analyses or systematic reviews on ALAN (n = 6) that were known by the review team (see Additional File 2 - section V); 3/We launched a call for articles through an experts’ networks; 4/We included relevant articles from the review team which have not been retrieved by any other sources.
2.5 Coding and data extraction
At the end of the screening process, all included articles were computed into a single systematic map database (objective 1). Then, each article was split into study units, which we will call ‘cases’; a ‘case’ was considered at the crossing point from one species, one exposure, and one outcome. Here, we considered that lights with different characteristics (color, duration, intensity, continuous/flashing, etc.) constituted different exposures. We extracted data from all cases, according to a predefined code book, to describe the study location (e.g. country, study area), the population (e.g. family name, species name, origin of individuals), the exposure, the outcome and the type of the study (e.g. observational/experimental, in-situ/ex-situ, type of comparator). We also extracted results from all observations; i.e. the effect of artificial light on the studied species that the authors measured on the considered outcome and summarized them as a negative, positive or non-significant impact. Coding and data extraction were conducted during a unique process, by two reviewers (RS, CT). Before coding, the two reviewers performed a test on 10 articles for training and discussed their coding results to assess their consistency.
2.6 Data mapping and narrative review methods
First, we exposed bibliometric results and produced tables and plots to have an overview of the available literature regarding descriptive variables (geographic distribution, chronologic distribution) and the PECO framework (objective 2). Second, we organized the studies to highlight the main research questions they address on the effect of artificial light on European terrestrial non-flying mammals, crossing several data (e.g. study context and study design, population, outcomes) (objective 3). Third, we described these knowledge clusters through a narrative synthesis; i.e. factually listing the findings of the studies avoiding any vote-counting (objective 4). Finally, knowledge gaps have been identified using the heat-tables of population-exposure-outcomes results and comparing knowledge clusters to the expected effects of ALAN on organisms (objective 5).
3 Results
Flow diagram in Figure 1 summarizes the results of the searching, screening and coding stages in adherence to CEE guidelines and ROSES reporting standards. Detailed bibliometric results of the screening process are available in Additional File 2 - Section VI and all inclusion/exclusion decisions at the three stages of the screening are available in Additional File 3. All these stages enabled us to build a systematic map database composed of 48 articles on the impact of artificial light on terrestrial non-flying mammals native to Europe, which provided 448 cases computed in Additional File 4.
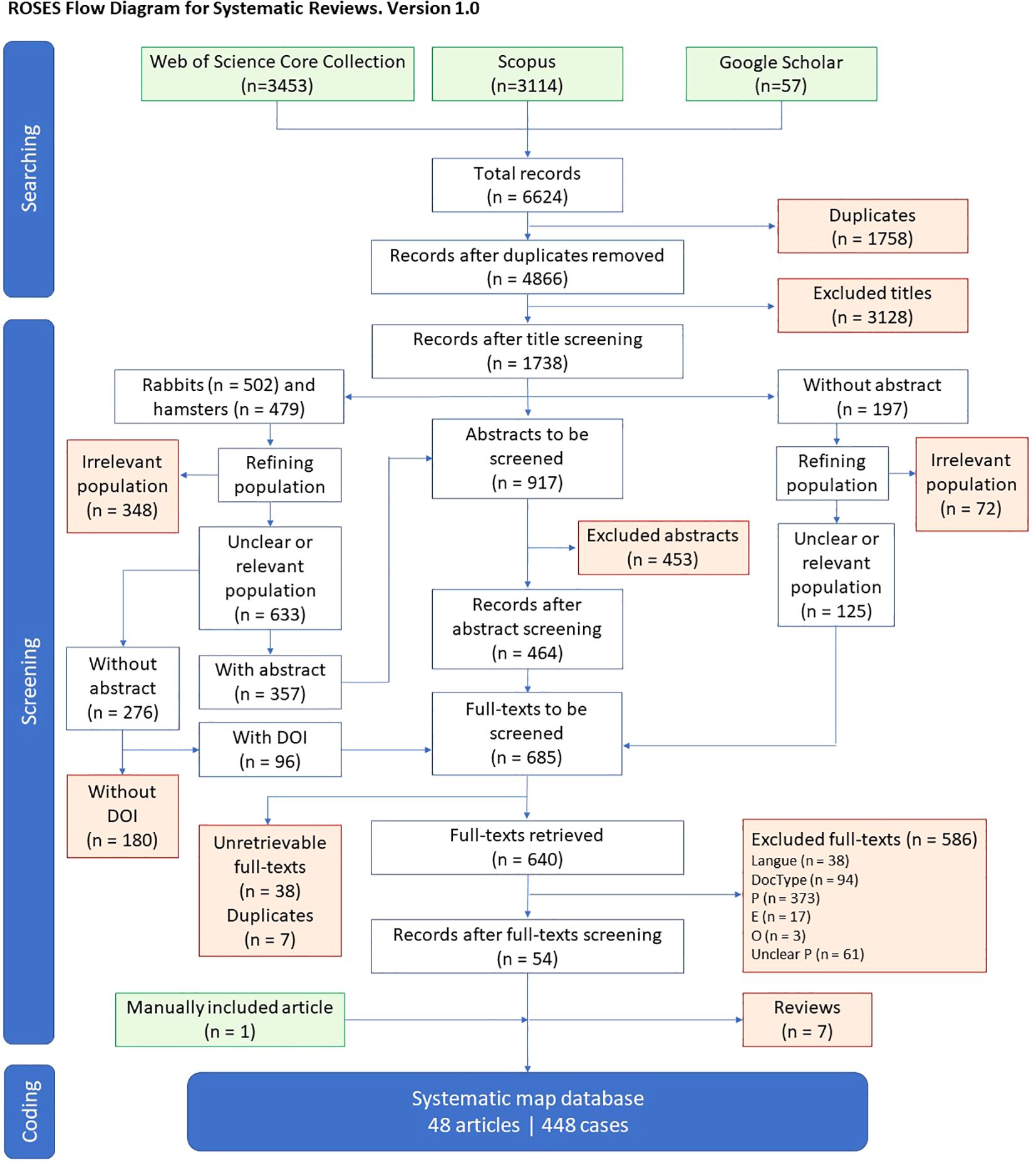
Figure 1. Flow diagram of the review process. Based on ROSES reporting standards from Haddaway et al. (2018), see https://www.roses-reporting.com.
3.1 Bibliometric features
Four articles are written in French and all the others are written in English. Only one article is a report (grey literature) and all the others are journal articles (peer-reviewed). Many different journals (33) provided articles although three journals count 3 or 4 articles (Chronobiology International, Journal of Experimental Biology, Neuroscience and Behavioral Physiology). In terms of chronologic distribution, we observe that publications have been increasing since 1975, with a maximum of 1 or 2 articles per year (Figure 2). Before 1975, 2 articles had been published, the oldest dating back to 1946 on Vulpes vulpes (Bassett, 1946). Only 2011, 2019 and 2022 stand out as special years in the whole corpus, with 5 articles published each, perhaps suggesting a kind of acceleration in the consideration of mammals in light pollution research.
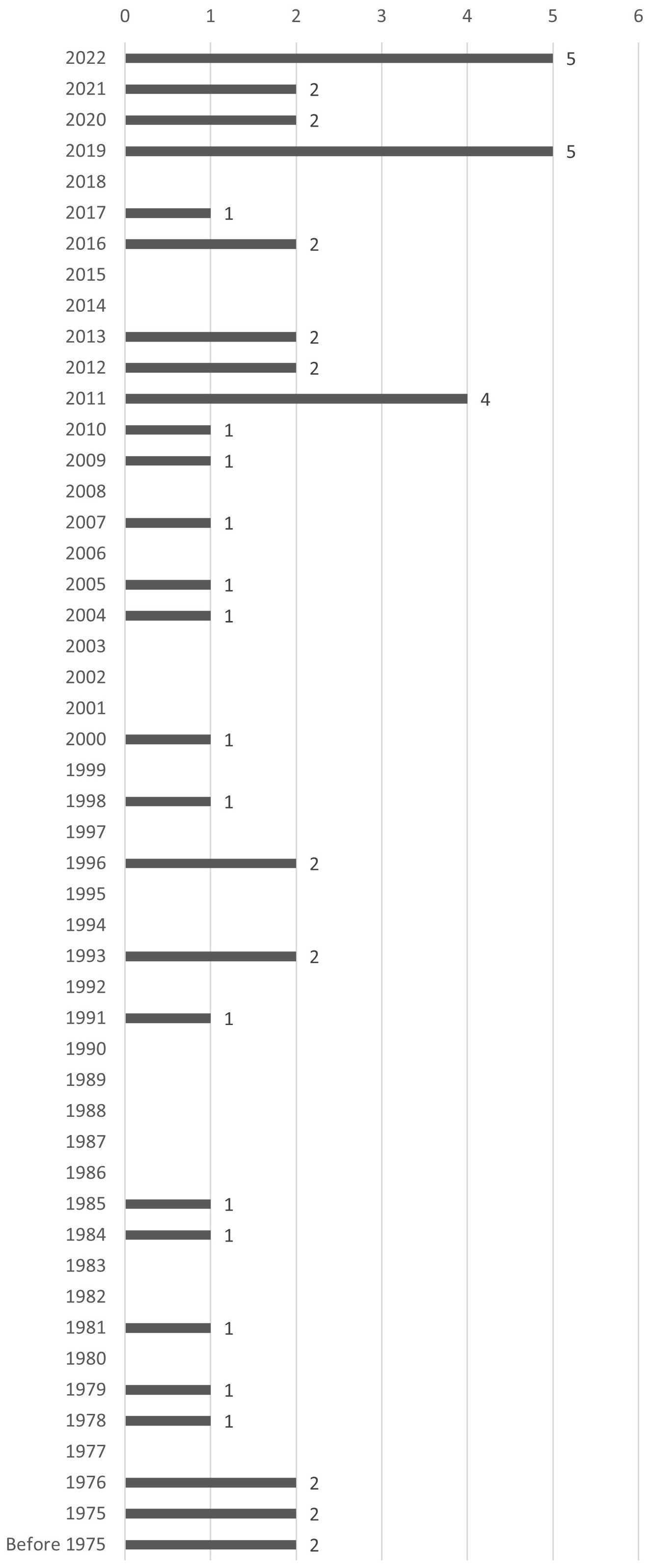
Figure 2. Chronological distribution of the included articles.
Articles cover 16 countries (included extra-European countries) and most studies were conducted in France (12 articles) and in Israel (9 articles) (Figure 3). The majority of studies were conducted in laboratory or in enclosure and few were conducted in-situ (respectively 31, 10 and 7).
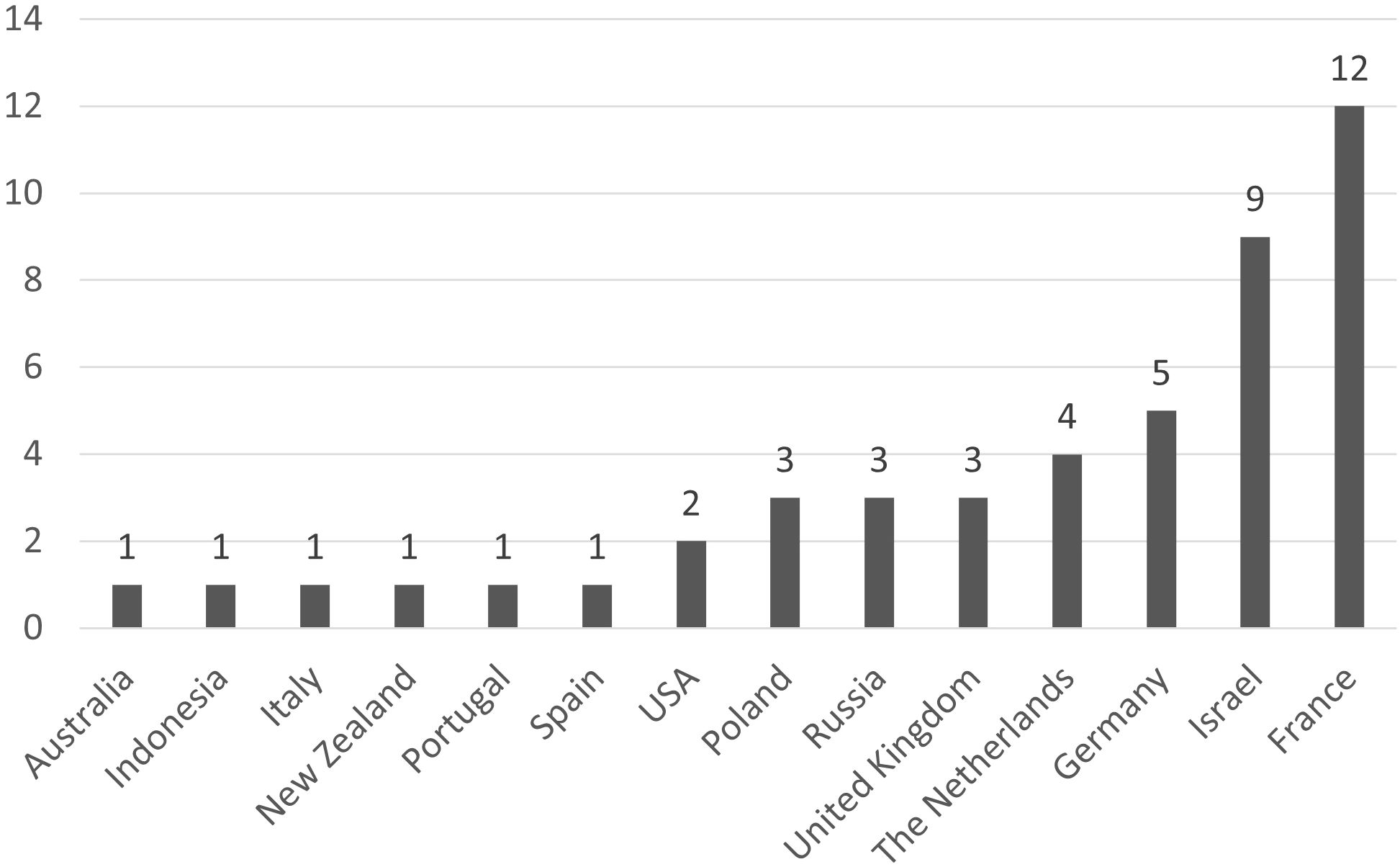
Figure 3. Geographic distribution of the included articles. The size of compartments is proportional to the number of articles for each country.
We collected articles about 12 mammal families (Figure 4, Table 2). Cricetidae and Muridae are the two most studied families, with respectively 13 articles/233 cases and 9 articles/82 cases. A total of 22/186 species (12%) are covered by at least one article. Microtus socialis (Cricetidae), Acomys cahirinus (Muridae) and Oryctolagus cuniculus (Leporidae) are the three most studied species (5 articles each). Four studies concern Erinaceus europaeus (Erinaceidae) and Sus scrofa (Suidae).
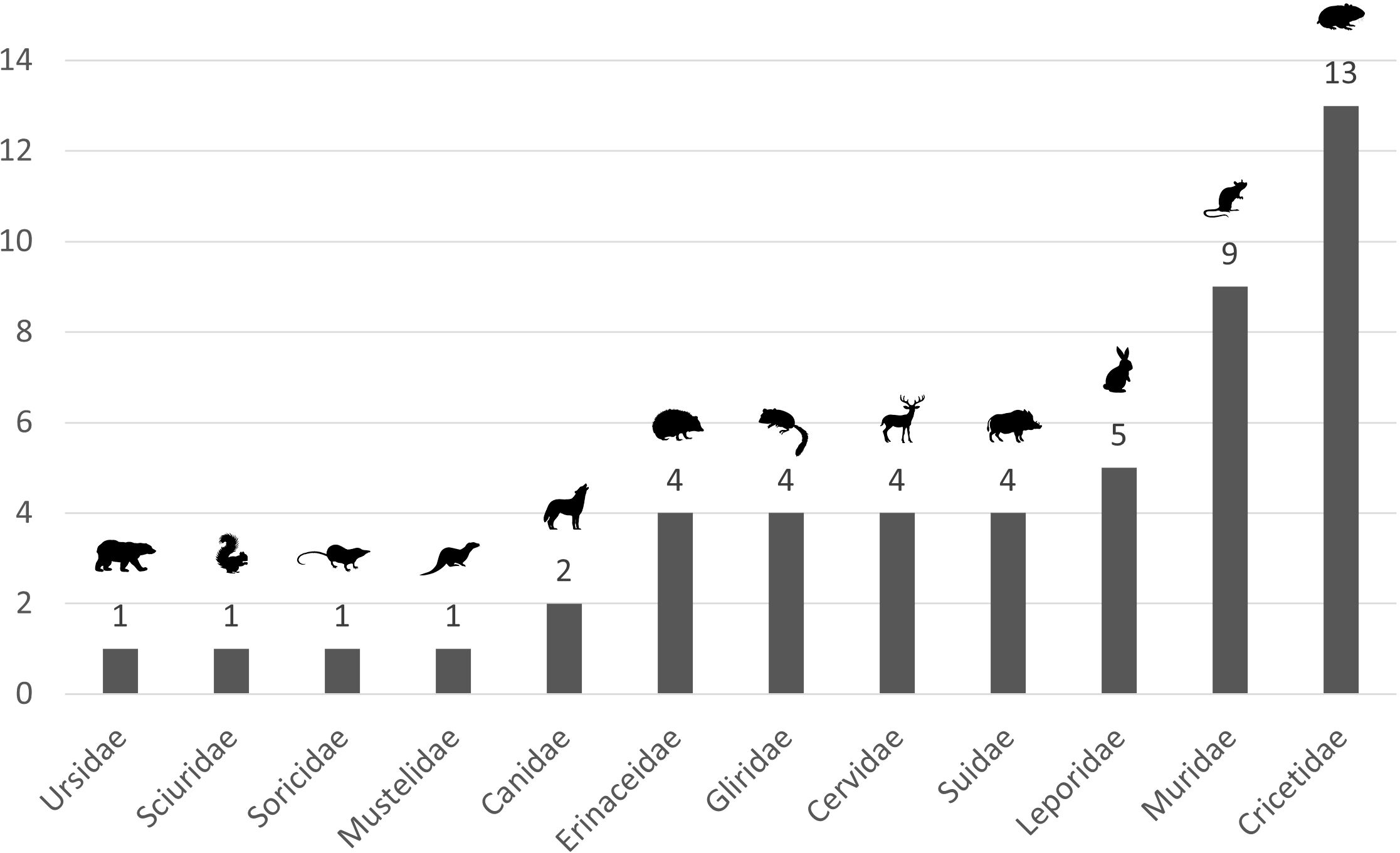
Figure 4. Proportion of articles for the 12 studied families. The size of compartments is proportional to the number of articles for each mammal family.
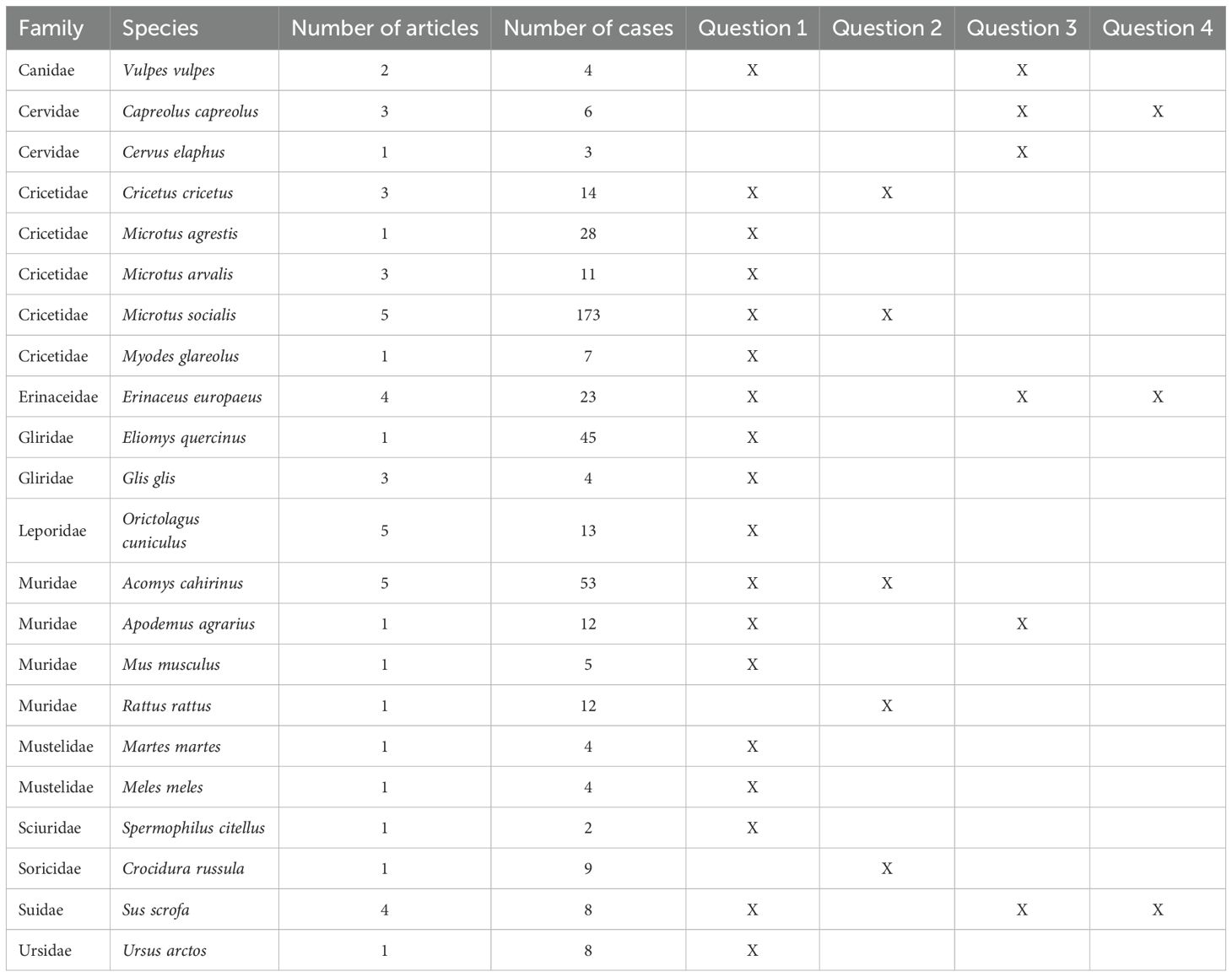
Table 2. Family and species covered by the included literature, with associated number of articles and cases and raised questions.
We observed four situations regarding the origin of the animals in the studies: 1/in 20 articles, individuals are wild animals (captured in the wild for the purpose of the experiment); 2/in 16 articles, animals did not come directly from wild (e.g. purchased from a commercial supplier, coming from breeding colonies or captive population); 3/in 11 articles, the origin of individuals was not specified; 4/1 article includes both wild and non-wild animals.
More than half of articles did not specify the source of artificial light the authors used (13 articles: not reported at all; 10 articles: only generic terms such as ‘lamps’ or ‘bulbs’). When specified, LED (light emitting diode) is the most studied technology (9 articles), followed by fluorescent lamp (tubes or bulbs) (7 articles) (Figure 5). Some other technologies are very briefly considered (metal halide, 2 articles; halogen, 2 articles; incandescent, 1 article). At last, 4 articles are counted separately since they assess the in-situ global light pollution whatever the source using satellite data or field monitoring.
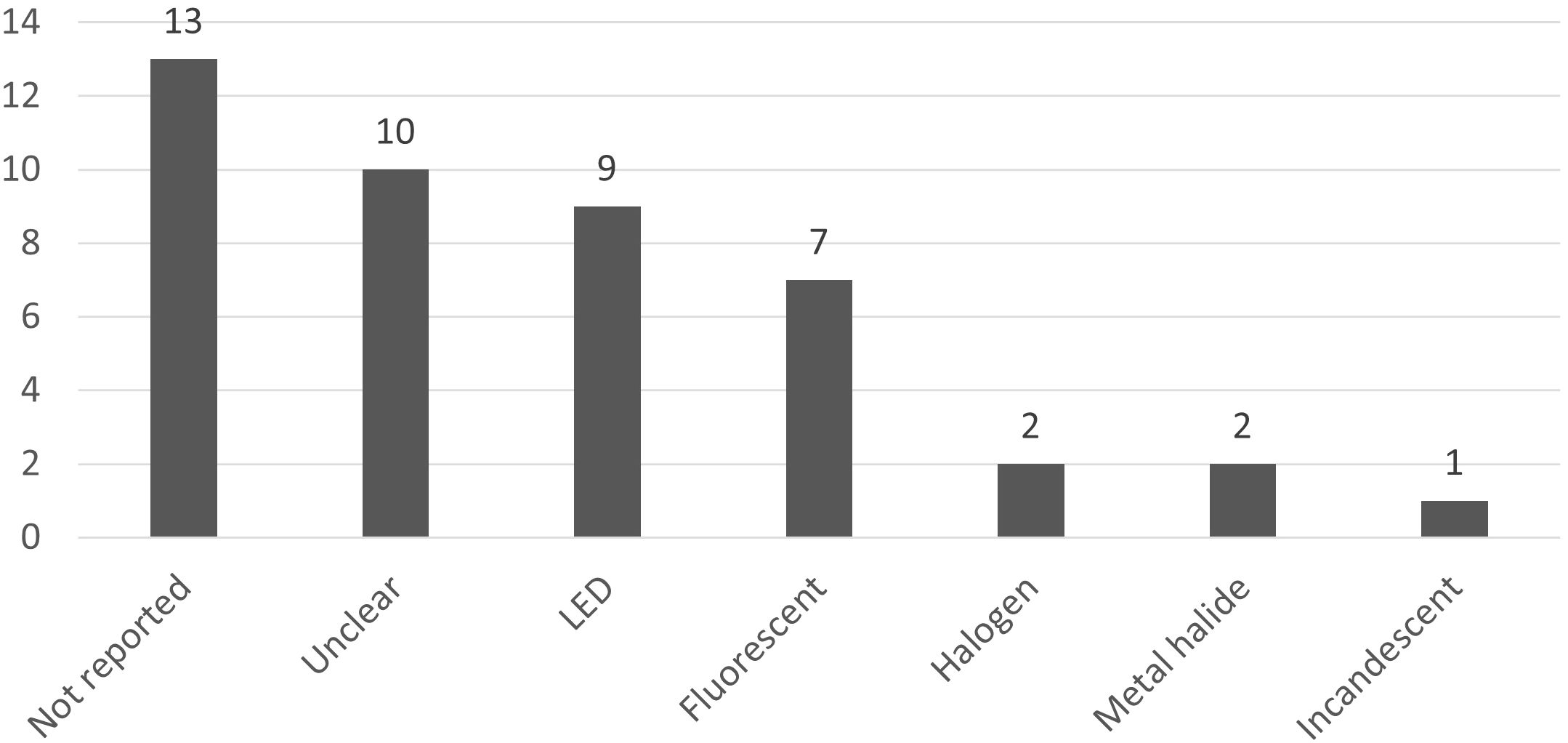
Figure 5. Proportion of articles for the studied types of lamps.
Outcomes of all the coded cases could be classified into 7 groups: activity, physiology, demography, distribution, reproduction, space use, species interactions. The very large majority of cases concern physiological outcomes (314 cases, 71%), covering a very wide range of variables monitored (e.g. body weight, urine production, hormonal rates, metabolism, oxygen consumption, sperm quality, neuron activity, cells or organs) and very diverse metrics (volume, mass, number of cells, size/diameter, rhythms of production or even qualitative issues). Although much less so, activity (97 cases, 22%) and space use (22 cases, 5%) are also studied. Activity can concern foraging (e.g. duration, frequency, schedule), ultradian/circadian/annual rhythms (e.g. diurnality index, activity on 24h cycle, daily activity, phase shifts), micro-behavior (e.g. wheel rotation) and all kinds of tasks (time spent in a room, number of visits). Space use outcomes deal with spatial organization (e.g. home range size, home range overlap) and movements (e.g. traveled distance, speed, number of stops, number of trips). Other outcomes are little studied (reproduction: 6 cases; distribution: 5 cases; demography: 3 cases; interaction: 1 case).
3.2 Research hotspots and narrative synthesis
By meeting our third objective described in the introduction, we identified four main research questions on the impacts of ALAN on European species of terrestrial non-flying mammals (Table 3) addressed by the available literature and based on specific study designs (Figure 6). This section will describe these four clusters and give a factual overview of their results.
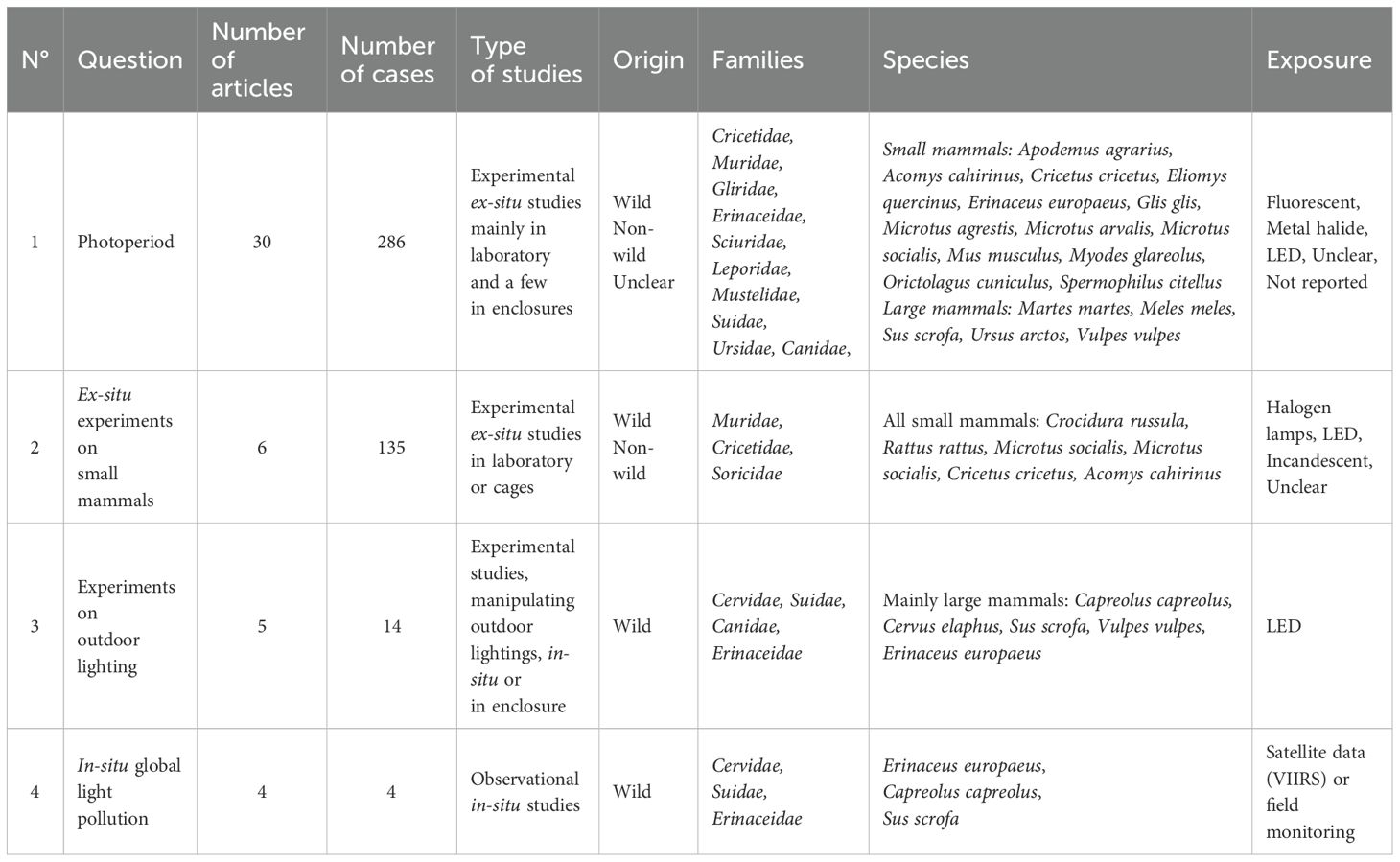
Table 3. Overview of the four main research questions.
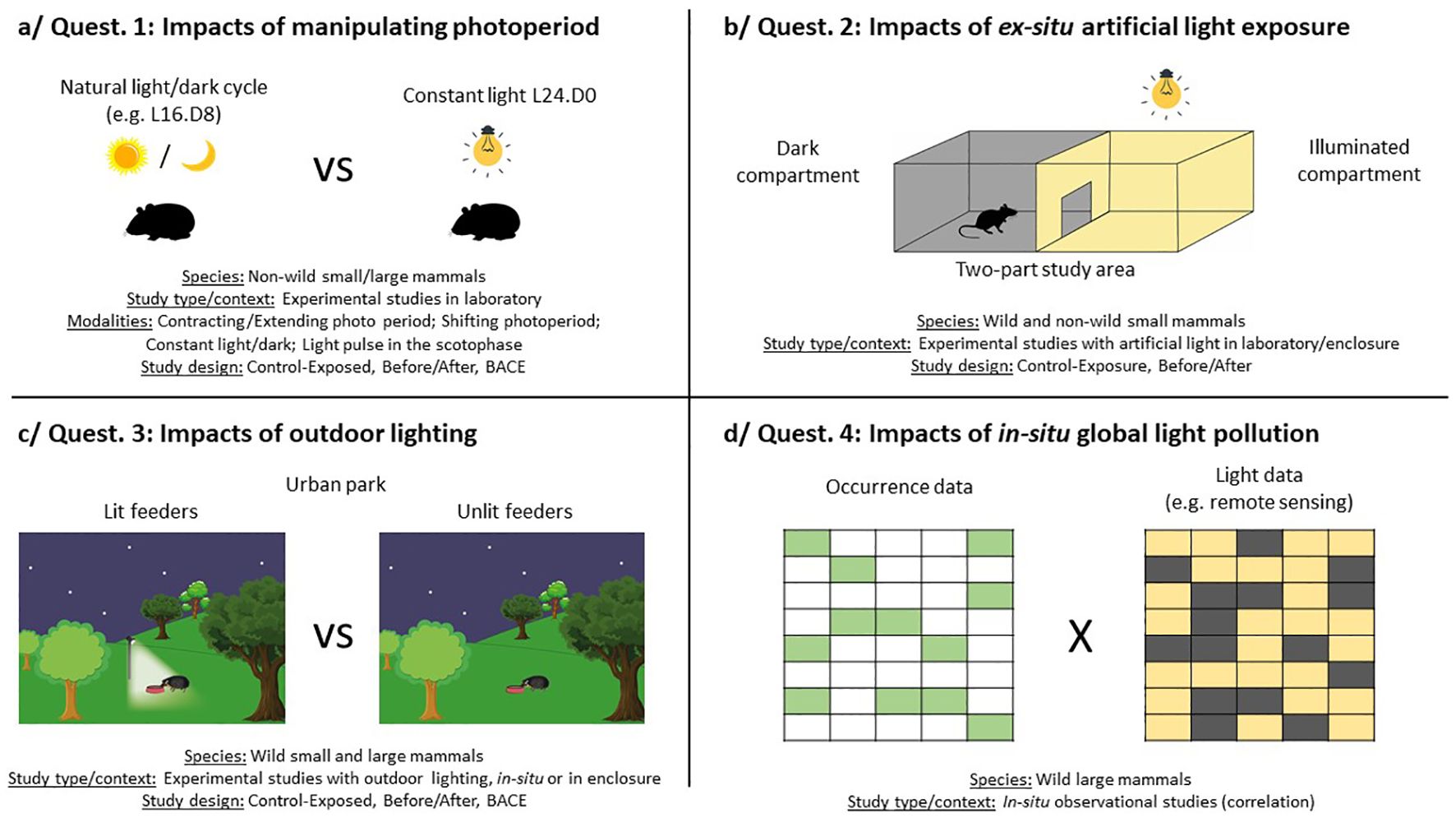
Figure 6. Main study designs for the four identified research questions.
3.2.1 Question 1: Can manipulating photoperiod in ex-situ conditions disturb small and large mammals?
A very substantial set of articles (30/48) raises the effects of modifying the photoperiod (Table 4). These articles especially deal with physiological outcomes (20 articles) and activity (12 articles), and in a very small number of articles: space use (1 article), reproduction (1 article), demography (1 article) or species interactions (1 article). They concern 10 families, covering 13 species of small mammals and 5 of large mammals. They are all experimental ex-situ studies, mainly conducted in laboratory (22 articles) (Figure 6a) and for a few in enclosures (8 articles). Source of artificial light is unclear or even not reported in more than 50% of articles but, when specified, the authors assessed the effects of fluorescent or metal halide lamps and LEDs. We can distinguish four ways in which the photoperiod is modulated: 1/photoperiod is contracted or extended (e.g. animals under short photoperiod L8.D16 are compared to animals under long photoperiod L16.D8, or vice versa); 2/photoperiod is shifted at once or progressively (e.g. from summer to winter light schedule over 6 months); 3/animals are maintained under constant illumination or under constant dark all day long; 4/a short period of light is applied at a precise moment of the scotophase (e.g. 3h first hours of the night). These artificial regimes can be either compared to each other or to natural dark/light cycle. Given the volume of articles in this knowledge cluster, we were not able to list here all the findings (all results are available in Table 4) and we decided to illustrate what these different lighting regimes can generate:
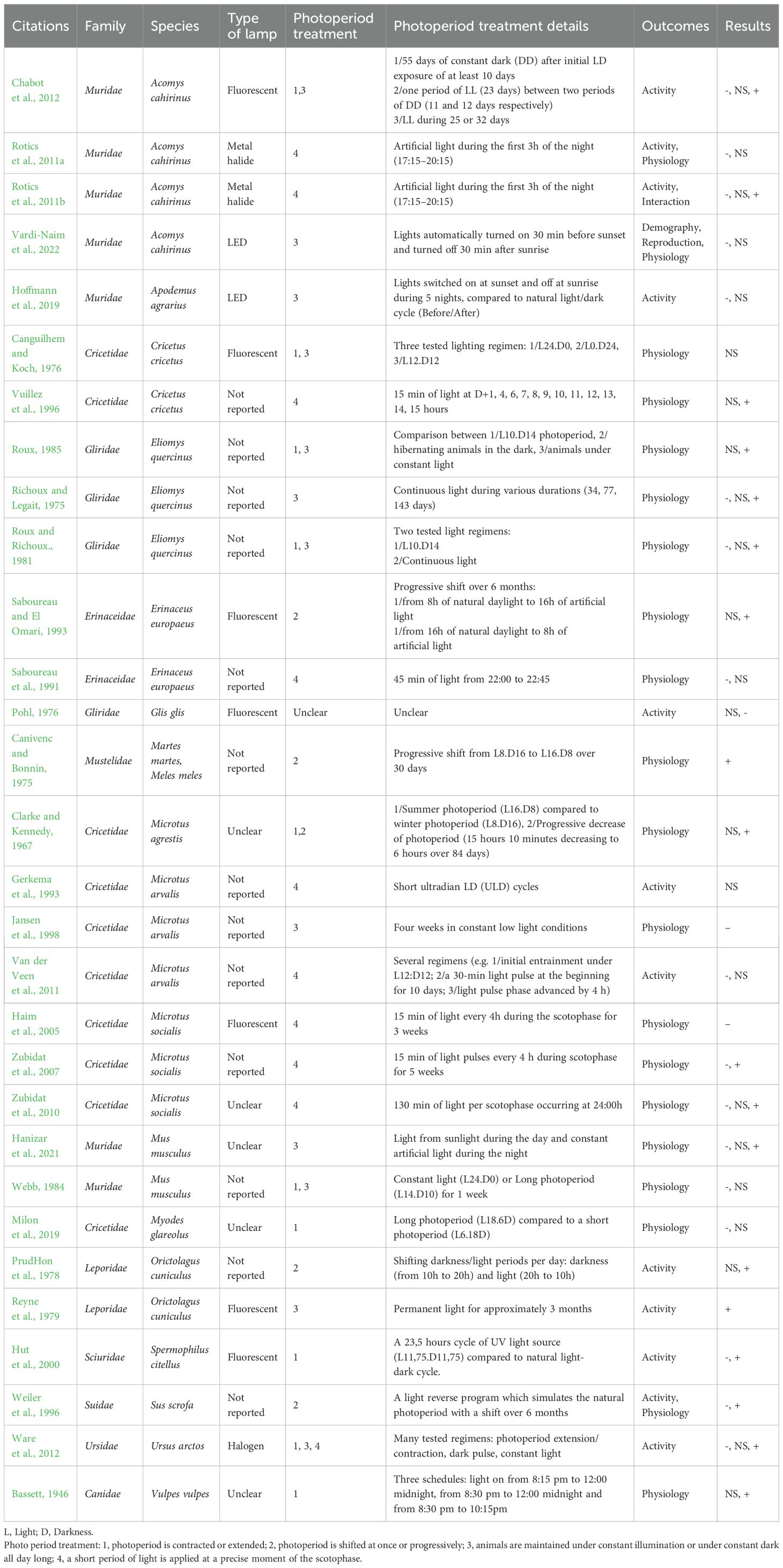
Table 4. Articles about manipulating the photoperiod (Question 1, N=30).
1/Photoperiod is contracted or extended: Ware et al. (2012) showed that photoperiod extension (+2h morning and +2h evening) for at least 2 weeks had various effect on bears’ activity (Ursus arctos) depending on the season (e.g. at fall it decreased activity whereas at spring it had no effect) (Ware et al., 2012). Exposure to a long photoperiod (L18.D6) produced neutral to negative effects on metabolism of interstitial cells (telocytes) of Myodes glareolus in comparison to a short photoperiod (L6.D18) (Milon et al., 2019). On Microtus agrestis, a long photoperiod (L16.D8), compared to a short one (L8.16D), was found to have a significant effect on body weight and specifically on weight of male and female reproductive organs (Clarke and Kennedy, 1967).
2/Photoperiod is shifted at once or progressively: Individuals of Orictolagus cuniculus submitted to a photoperiod shift in one go (from 14L.10D to 10L.14D) gradually changed the distribution of food consumption over the day to adapt to the new schedule in about four weeks; however the quantities of solid/liquid food eaten per day were not affected (PrudHon et al., 1978). A progressive reduction in light exposure, shifting from 8L:16D to 16L:8D in 30 days, impacted female reproductive organs of two mustlids (Meles meles and Martes martes) (increasing corpus luteum weight, luteal cell diameter and progesterone content of luteal tissue) (Canivenc and Bonnin, 1975). Saboureau and El Omari (1993) tested the influence of a progressive shift (from 8L.16D to 16L.8D and vice versa) on male reproductive organs of Erinaceus europaeus and found a synchronization of testicular volume, plasma testosterone concentrations and gonadotrophin concentrations with the new schedule, although values of these outcomes did not vary (Saboureau and El Omari, 1993).
3/Animals are maintained under constant illumination or under constant dark all day long: Constant illumination has been highlighted to have several types of adverse effects. For instance, this led to a false perception of summer day length by Acomys cahirinus (whatever the wavelengths), resulting in the loss of seasonality and continuous reproduction; however the number of pups seemed to be more affected by competition than by artificial light (Vardi-Naim et al., 2022). A transponder tag survey in enclosure based on wild Apodemus agrarius found that ‘warm white’ LED street lamps, switched on at sunset and off at sunrise, increased home range size of animals and affected synchrony between individuals (Hoffmann et al., 2019). Constant light produced a spontaneous increase in activity on hibernating bears Ursus arctos (Ware et al., 2012). It had detrimental effects on neuronal functioning on Microtus arvalis (Jansen et al., 1998) and decreased sperm quantity of Mus musculus although sperm motility was not affected (Hanizar et al., 2021).
4/A short period of light is applied at a precise moment of the scotophase: Rotics et al. (2011b) found that illuminating the 3 first hours of the night with yellow metal halide lamps could decrease body temperature of Acomys cahirinus (Rotics et al., 2011a). In another experiment on the same species, such a light pulse in the scotophase increased foraging during the hour after illumination and as a consequence competition for foraging between individuals (Rotics et al., 2011b). Saboureau et al. (1991) found that an interruption of the night by 45 min of light decreased rapidly and significantly the pineal melatonin content in Erinaceus europaeus but that a return to darkness for approximately 2 hours restored the usual values (Saboureau et al., 1991). Zubidat et al. (2007) also found negative effects on Microtus socialis exposed to 15 min of light pulses every 4 h during scotophase for 5 weeks, demonstrating a decrease in many physiological outcomes (e.g. oxygen consumption, body mass, rectal temperature, urine production) (Zubidat et al., 2007).
3.2.2 Question 2: May artificial light in ex-situ conditions have detrimental effects on small mammals?
A set of 6 articles addresses the effects of artificial light on wild or non-wild small mammals (Muridae, Cricetidae, Soricidae), through experimental ex-situ studies in laboratory or cages (Figure 6b). These studies give results on activity, space use and physiological outcomes (Table 5). They are based on control/exposure study design with same individuals in a two-part study area (e.g. illuminated compartment vs dark compartment [Dupont et al., 2019; Aparicio et al., 2023)] or with different individuals (e.g. individuals in an illuminated U maze vs individuals in a dark U maze [Farnworth et al., 2019)], or on before/after study design [scotophase vs photophase, e.g (Zubidat et al., 2010)].
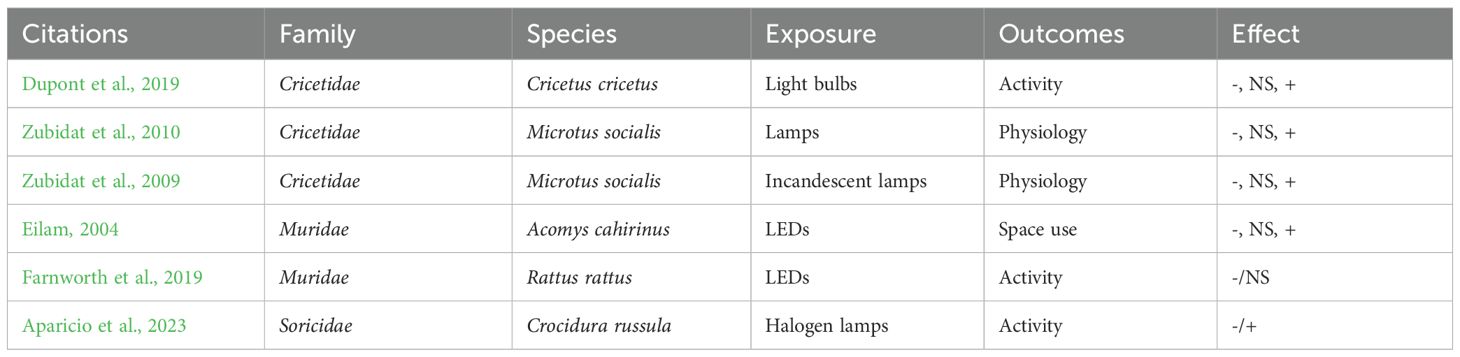
Table 5. Articles on adverse effects of artificial light on small mammals (Question 2, N=6).
Aparicio et al. (2023) showed that halogen lamps modified foraging activity of Crocidura russula; shrews came out of their shelters less if the outside was lit, leading to a reduction in the number of foraging individuals and, ultimately, less food consumed (Aparicio et al., 2023). Another study on European hamster Cricetus cricetus demonstrated that individuals generally avoided the illuminated compartment, particularly when grooming; individuals spent significantly more time in the dark compartment than in the lit one with amber LED 40 lux (Dupont et al., 2019). Eilam (2004) studied the effect of artificial light on Acomys cahirinus space use and founded that in illuminated areas spiny mice moved less frequently, traveled shorter distances, and made more direct trips (fewer stops) (Eilam, 2004). Farnworth et al. (2019) also demonstrated an avoidance mechanism of artificial light on the Rattus rattus with a significant reduction in rat movements. Under illumination, rats tended to approach the seed tray faster and reduce their total foraging time within the seed tray (Farnworth et al., 2019). Finally, two laboratory studies carried out in Israel assessed the biological effects of artificial light on Microtus socialis and revealed that it is likely to alter a number of parameters linked to urine production and oxygen consumption (Zubidat et al., 2009, Zubidat et al., 2010).
3.2.3 Question 3: Is outdoor lighting at night likely to have adverse effects on wild mammals?
A set of 5 articles gathers experimental studies, manipulating outdoor lightings at night, in-situ or in enclosure (Figure 6c), to assess the effects on diverse outcomes (distribution, activity, space use) (Table 6). All the studies deal with wild animals, mainly large mammals (Capreolus capreolus, Cervus elaphus, Sus scrofa, Vulpes vulpes and Erinaceus europaeus), using camera traps to collect occurrence and behavioral data.
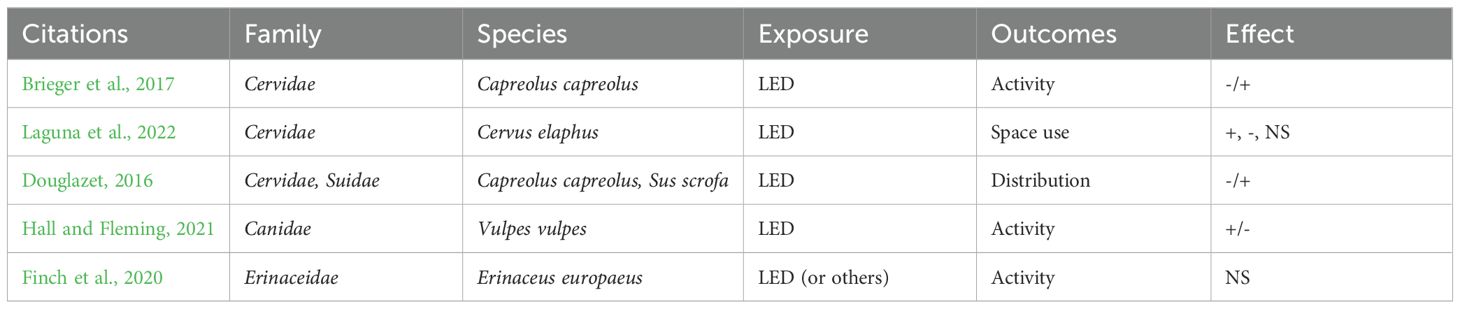
Table 6. Articles on adverse effects of artificial light on wild large mammals (Question 3, N=5).
Finch et al. (2020) assessed the effect of outdoor lightings installed at feeding stations for European hedgehogs in 33 British gardens (Finch et al., 2020). The results showed no consistent overall effect of ALAN on the presence and feeding activity of hedgehogs, nor on the circadian rhythm of their activity. Brieger et al. (2017) tested the behavioral response of roe deer to LEDs. Their results show that roe deer preferred feeders that are lit to those that are not; however, they spent less time feeding when the feeders were lit (Brieger et al., 2017). The results were similar for the two colors of light tested, “blue” (440-490 nm) and “warm white” (575-675 nm). Douglazet (2016) studied the impact of LEDs on the nocturnal movements of terrestrial non-flying mammals in woodland and showed that artificial light appears to have a slight negative effect on the presence of roe deer and a rather attractive effect on wild boar (Douglazet, 2016).
Two other studies can be directly linked to conflict between humans and wildlife, since deterrent systems are often used (e.g. to limit fox predation or roadkill). Laguna et al. (2022) tested the effect of light (flashing LEDs) and sound (ultrasound) devices simultaneously designed to deter cervids (Cervus elaphus) at the roadside (Laguna et al., 2022). Their results showed (i) a 48.96% reduction in the use of the treated zone and (ii) a 67.71% reduction in the number of times the deterrent line was crossed. However, the deterrent effect wore off after 6 to 7 weeks. Hall and Fleming (2021) conducted a study on the effectiveness of two LED lamps sold to deter red foxes from approaching farrowing sheds on farms (Foxlights® and motion activated spotlights) (Hall and Fleming, 2021). Foxlights® treatment resulted in a 12% increase in fox activity compared to the unlit treatment (control), suggesting that Foxlights® may attract rather than deter foxes. The motion-activated spotlight decreases fox activity by 5% compared to control (no lighting), suggesting a slight avoiding effect of flashing light.
3.2.4 Question 4: Does global sky glow affect mammals in-situ?
A set of 4 articles, addresses the effects of sky glow on wild mammals (Table 7). These are in-situ observational studies that look for correlation between a global light pollution (e.g. estimated from remote sensing) and diverse outcomes (attendance, activity) (Figure 6d). These studies are quite recent (2019-2022) which underlines a new consideration of this issue on mammals. Berger et al. (2020) compared light pollution levels with the activity of hedgehogs equipped with bio-loggers by monitoring their spatio-temporal behavior in three urban areas of Berlin over several weeks (Berger et al., 2020). Although hedgehogs can react very variably to environmental influences, they preferred the least lit areas, suggesting a consistent tendency for hedgehogs to avoid light pollution. Two other studies focused on the city of Krakow in Poland. Ciach and Fröhlich (2019) studied the factors governing the winter distribution of Capreolus capreolus in this urban environment and observed that light pollution was negatively correlated with the probability of species occurrence (Ciach and Fröhlich, 2019). Ciach et al. (2022) studied the influence of anthropogenic factors on the presence of Sus scrofa in this town and also highlighted a negative impact of light pollution on the attendance of wild boars (Ciach et al., 2022). Finally, Gordigiani et al. (2022) found that wild boar limited their activity on nights with high level of ambient light (Gordigiani et al., 2022). However, this study is less clear because ambient light included both moonlight and artificial lighting. Though, wild boar activity was better explained by the variation in luminosity than by the variation in the lunar day, which may highlight an additional inhibiting effect of light pollution.
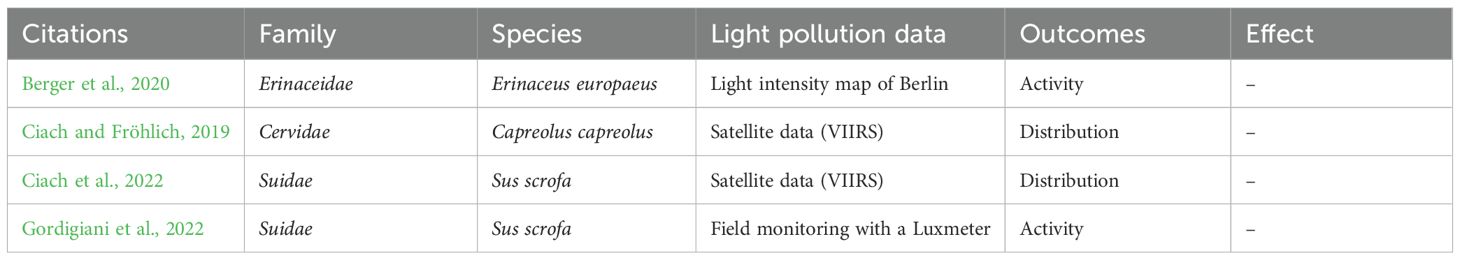
Table 7. Articles on the effects of global light pollution on mammals (Question 4, N=4).
4 Discussion
4.1 In line results with expected impacts of ALAN
First, a substantial set of included articles stress disturbances of altering dark/light cycle (e.g. shifting dark/light phases, reducing or cutting scotophase) on several outcomes. The role of photoperiodism in life synchronization is fundamental (Bradshaw and Holzapfel, 2007) and ALAN is a strong disruptor of chronobiology in many organisms including humans (Zeman et al., 2023; Lei et al., 2024). This review confirms that extending photoperiod leads to metabolic and physiological troubles in several non-flying mammal species (Clarke and Kennedy, 1967; Milon et al., 2019), including neuronal alteration (Jansen et al., 1998) and decrease in reproduction ability (Canivenc and Bonnin, 1975; Saboureau and El Omari, 1993; Hanizar et al., 2021) which could result in many cascading consequences. Cutting the scotophase (e.g. with light pulse) appears to disturb many physiological parameters of small mammals (e.g. oxygen consumption, body mass, body temperature, urine production) (Zubidat et al., 2007; Rotics et al., 2011a) and totally deleting it (i.e. constant light exposure) results in loss of seasonality, for instance regarding reproduction (Vardi-Naim et al., 2022). These results are consistent with previous studies drawing detrimental effects of modifying circadian rhythms (van Jaarsveld et al., 2019; Issad et al., 2021; Zuo et al., 2023). Exposure to ALAN is widely demonstrated to drop melatonin production (Robert et al., 2015; Le Tallec et al., 2016; Moaraf et al., 2020) - a fundamental hormone for sleep/activity balance, metabolism and health stability - and we found such a pattern in Erinaceus europaeus (Saboureau et al., 1991).
Second, included studies stress an overall light-avoidance behavior in several species, particularly small mammals such as Acomys cahirinus (Eilam, 2004), Cricetus cricetus (Dupont et al., 2019), Crocidura russula (Aparicio et al., 2023), Rattus rattus (Farnworth et al., 2019), Erinaceus europaeus (Berger et al., 2020) but also large mammals such as Capreolus capreolus (Douglazet, 2016) and Cervus elaphus (Laguna et al., 2022). Such animals avoid - or very less use - illuminated areas, come out of their shelters less or spend significantly more time in dark habitats. Light is known to have two opposite behavioral effects on fauna: attraction (e.g. flight-to-light insect behavior) vs repulsion (Farnworth et al., 2018; Elvidge et al., 2019). Mammals are precisely more likely to belong to the second group (Kott et al., 2010; Stone et al., 2012; Hoffmann et al., 2018; Rydell et al., 2021) and our review confirms this general trend for non-flying terrestrial species.
Third, in this review we highlighted that activity is lower under ALAN exposure, for instance Sus scrofa is less active when night is brighter (Gordigiani et al., 2022), Capreolus capreolus spends less time feeding when the feeders are lit (Brieger et al., 2017) while activity of Vulpes vulpes is reduced under intermittent light (Hall and Fleming, 2021). Particularly, this can delay or disturb foraging (i.e. reduced foraging time, less food consumed) (Farnworth et al., 2019; Aparicio et al., 2023). ALAN has been demonstrated to decrease activity of several taxa, including mammals (Francis et al., 2015; Zhang et al., 2020) and our results are also in line with moonlight influence. Many mammals show inhibition while moon is waxing (Prugh and Golden, 2014) resulting in a reduction of activity for small mammals (Abramsky et al., 1996; Hemami et al., 2011; Bueno A de and Motta-Junior, 2015; Hernández et al., 2021) or bats (Saldaña-Vázquez and Munguía-Rosas, 2013). We also found that artificial light is likely to disrupt mammal movements; e.g. individuals move less frequently, travel shorter distances and make more direct trips when they are exposed to ALAN (Eilam, 2004), again underlining an aversive effect consistent with previous results on bats (Voigt et al., 2020; Bhardwaj et al., 2020; Barré et al., 2023).
Finally, at a broader level, light pollution would tend to be negatively correlated with occurrence of some species, such as Sus scrofa and Capreolus capreolus (Ciach and Fröhlich, 2019; Ciach et al., 2022). Negative link between ALAN and species attendance has been documented for many taxa (Marín-Gómez et al., 2020; Luscier et al., 2023; Mathiaparanam et al., 2024). ALAN removes suitable habitats (Straka et al., 2020; Shier et al., 2020) and fragments nightscapes (Beier, 1995; Bliss-Ketchum et al., 2016) which could ultimately result in lower distribution.
4.2 Limits of the primary studies
First, we could observe that a lot of information was often missing from the articles - including, basic data such as the year of the experiment or the study area, which is a handicap for systematic mapping and would be all the more detrimental to a future systematic review. The source of the artificial light (whether it is LED, halogen, metal halide, etc.) was unknown in half of the articles, which did not mention it at all or used only generic terms such as bulbs or lamps. It should also be noted that many articles had to be rejected during the screening process because species name were not specified (only vernacular terms - such as rabbits, hamsters or voles - which unable to determine whether the species is native to Europe or not). Moreover, the origin of the individuals may remain unclear (whether they are wild animals or not), however it can be an important criterion; e.g. influencing particular behavior or habituation to a specific circadian rhythm of activity. We therefore encourage authors to provide greater details in the ‘materials and methods’ section of the articles.
Second, since we did not perform a critical appraisal on the included studies (in adherence to CEE guidelines for systematic maps), we are unable to identify limitations of primary research in terms robustness and susceptibility to bias. However, for each case we coded variables that deal with the study designs which will inform future users of the systematic map when interpreting the studies.
4.3 Knowledge gaps: implications for research
Although our searches targeted 186 species, we retrieved documents only for 22 of them (12%). Moreover, half of the species counts a unique article; only a few species, such as Microtus socialis, Acomys cahirinus or Erinaceus europaeus, have 4-5 articles each, which is still very few. Regarding mammal families, 55% (12/22) are covered by at least one article; however, the available literature did not pay any attention to some large families, such as Talpidae, Bovidae, Leporidae (only 1 species considered), Sciuridae or Mustelidae (only 1 article each). As a result, while sensory pollutants, including artificial light and anthropogenic noise, are becoming major concerns for ecological research (Dominoni et al., 2020); this review highlights a lack of primary research on terrestrial non-flying mammal species as informed in previous systematic reviews, even though our more targeted search strategy has led to more mammal studies being reported.
Regarding exposure, LEDs are more and more considered which is a good point with regard to their expansion as outdoor lightings. However, some characteristics of the exposure are not considered in studies, such as wavelength/color temperature (very few studies) or flickering. The available literature essentially deals with fundamental research on the effect of artificial light, which may be unrelated to real-life outdoor lightings and does not represent their diversity. For instance, there are no studies on the different types of lighting (enhancement, sports fields, shop windows, domestic lighting, car light on roads) or based on real lighting management (such as part-night lighting, motion activated lighting).
As far as the outcomes are concerned, ecological issues have been little studied (mainly activity such as foraging), whereas the majority of the studies are in the field of physiology and cell biology. This is a major shortcoming if we are to make a comprehensive overview on the effects of artificial light on terrestrial non-flying European mammals. Numerous outcomes should be studied, such as competition, dispersal, and studies should be carried out at the ecosystem level (e.g. prey-predator relationships, species richness). Reproduction and survival are also under-studied, even though they are essential factors in the sustainability of mammal populations. Finally, it would also be interesting to assess the variability of mammalian responses to the effects of ALAN as a function of several morphological/biological traits (e.g. dark adaptation speed, spectral color resolution) or functional traits (e.g. diet, habitat specialization) that may influence species vulnerability (Ditmer et al., 2021a). This knowledge would be of major operational interest for species conservation and lighting planification, by defining guilds of mammals that are more or less sensitive.
4.4 Limits of the review
We are aware that several points in our map and review process may limit the comprehensiveness of our results. Firstly, language restriction to English and French may constitute a limit as far as grey literature is concerned because non-English-language literature can be a source of important evidence. However, this choice is dictated by the capacity of our review team. This limitation is recurrent in systematic maps and reviews due to constraints related to resources and time (Hannah et al., 2024). Solutions to limit this gap may be 1/including non-English partners in the review team to increase multilingual capabilities, 2/using reliable machine translation applications. Secondly, we were not able to find 5.5% of retained PDFs (38/685 full-texts), which was considered as acceptable. These PDFs remained unretrievable for our institution despite all our efforts to find them (interlibrary loans, requests on Researchgate, etc.). They are listed in Additional File 3 to facilitate their search by anyone who would want to complete this review. Thirdly, we decided to exclude after title screening 180 articles dealing with undetermined rabbits that had no abstract nor DOI. We assumed that the probability that these articles did deal with European rabbit was extremely low because of the 322 rabbit articles that were screened up to full-text only 5 articles were included at the end.
5 Conclusion
This evidence synthesis provides a comprehensive overview on the impacts of night-time lighting - a key sensory anthropogenic driver - on European terrestrial non-flying mammals. Thanks to a focused search strategy we were able to capture 48 articles covering 22 species from 12 families and providing 448 cases (i.e. data crossing one species, one exposure and one outcome), which reveals that terrestrial non-flying mammals in Europe are more studied than expected. We could identify 4 research hotspots addressed by the available literature, including a substantial knowledge cluster on the influence of artificial light on chronobiology. Even if the amount of available literature is modest for some outcomes, the detrimental consequences highlighted on circadian rhythm, behavior, foraging, movement or occurrence, are consistent with the documented influence of moonlight as well as the known effects of artificial light on other taxa. These results, including the knowledge gaps, will be useful for researchers and for those commissioning research projects.
However, this evidence base needs to be consolidated to make further systematic reviews or meta-analyses possible, and to this end we encourage the undertaking of additional primary research. While small mammals are commonly used as organisms for medical studies in laboratories; research in ecology should pay more attention to wild species mammals, since they are mainly nocturnal and some of them are endangered species in Europe. Research should diversify the studied mammal species and light exposure and consider outcomes at population, landscape or ecosystem levels. We also suggest that articles should be more detailed, in particular in their ‘material and methods’ section, for a better use in evidence syntheses.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author/s.
Author contributions
RS: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing. CF: Conceptualization, Data curation, Funding acquisition, Investigation, Writing – review & editing. TT: Conceptualization, Data curation, Funding acquisition, Investigation, Writing – review & editing. CT: Data curation, Investigation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by PatriNat and EDF, as part of a partnership. PatriNat is a joint research unit funded by OFB French Biodiversity Office, MNHN National Museum of Natural History, CNRS National Scientific Research Center and IRD International Research Institute, on behalf of the French Ecology Ministry.
Acknowledgments
We want to thank Patrick Haffner (PatriNat) for having exported the list of european mammal species, Erika Beauregard (EDF) for her help in search string building process and Jade Ka (MNHN) for her help in searching the fulltexts. Equally, Joseph Langridge (FRB), a native English speaker, for his thorough proof-reading.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Correction note
A correction has been made to this article. Details can be found at: 10.3389/fevo.2025.1649381.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1544308/full#supplementary-material
Additional File 1 | ROSES form for systematic reviews (Excel document).
Additional File 2 | Search strategy and review process (Word document).
Additional File 3 | Screening process (Excel document).
Additional File 4 | Systematic map and cases databases (Excel document).
References
Abramsky Z., Strauss E., Subach A., Riechman A., and Kotler B. P. (1996). The effect of barn owls (Tyto alba) on the activity and microhabitat selection of Gerbillus allenbyi and G. pyramidum. Oecologia 105, 313–319. doi: 10.1007/BF00328733
Aparicio G., Carrilho M., Oliveira F., Mathias M. D. L., Tapisso J. T., and Von Merten S. (2023). Artificial light affects the foraging behavior in greater white-toothed shrews (Crocidura russula). Ethology 129, 88–98. doi: 10.1111/eth.13347
Azam C., Le Viol I., Julien J.-F., Bas Y., and Kerbiriou C. (2016). Disentangling the relative effect of light pollution, impervious surfaces and intensive agriculture on bat activity with a national-scale monitoring program. Landscape Ecol. 31, 2471–2483. doi: 10.1007/s10980-016-0417-3
Barré K., Thomas I., Le Viol I., Spoelstra K., and Kerbiriou C. (2023). Manipulating spectra of artificial light affects movement patterns of bats along ecological corridors. Anim. Conserv. 26, 865–875. doi: 10.1111/acv.12875
Bassett C. F. (1946). Extending the lighting range and increasing the total hours of illumination for growing fox pups. Ann. N. Y. Acad. Sci. 48, 247–253. doi: 10.1111/j.1749-6632.1946.tb31776.x
Beier P. (1995). Dispersal of juvenile cougars in fragmented habitat. J. Wildlife Manage. 59, 228. doi: 10.2307/3808935
Bennie J., Davies T. W., Cruse D., and Gaston K. J. (2016). Ecological effects of artificial light at night on wild plants. J. Ecol. 104, 611–620. doi: 10.1111/jec.2016.104.issue-3
Berger A., Lozano B., Barthel L. M. F., and Schubert N. (2020). Moving in the Dark—Evidence for an Influence of Artificial Light at Night on the Movement Behaviour of European Hedgehogs (Erinaceus europaeus). Animals 10, 1306. doi: 10.3390/ani10081306
Berger-Tal O., Greggor A. L., Macura B., Adams C. A., Blumenthal A., Bouskila A., et al. (2019). Systematic reviews and maps as tools for applying behavioral ecology to management and policy. Behav. Ecol. 30, 1–8. doi: 10.1093/beheco/ary130
Bhardwaj M., Soanes K., Lahoz-Monfort J. J., Lumsden L. F., and van der Ree R. (2020). Artificial lighting reduces the effectiveness of wildlife-crossing structures for insectivorous bats. J. Environ. Manage. 262, 110313. doi: 10.1016/j.jenvman.2020.110313
Bliss-Ketchum L. L., de Rivera C. E., Turner B. C., and Weisbaum D. M. (2016). The effect of artificial light on wildlife use of a passage structure. Biol. Conserv. 199, 25–28. doi: 10.1016/j.biocon.2016.04.025
Bradshaw W. E. and Holzapfel C. M. (2007). Evolution of animal photoperiodism. Annu. Rev. Ecology Evolution Systematics 38, 1–25. doi: 10.1146/annurev.ecolsys.37.091305.110115
Brieger F., Kämmerle J., Martschuk N., Ortmann S., and Hagen R. (2017). No evidence for a ‘warning effect’ of blue light in roe deer. Wildlife Biol. 2017, 1–5. doi: 10.2981/wlb.00331
Bueno A de A. and Motta-Junior J. C. (2015). Behavioural and morphological strategies by small savannah rodents to avoid predation. Mamm. Biol. 80, 401–408. doi: 10.1016/j.mambio.2015.05.005
Camacho L. F., Barragán G., and Espinosa S. (2021). Local ecological knowledge reveals combined landscape effects of light pollution, habitat loss, and fragmentation on insect populations. Biol. Conserv. 262, 109311. doi: 10.1016/j.biocon.2021.109311
Canguilhem B. and Koch A. (1976). Effects of temperature and light variations, and of radiothyroidectomy on body weight regulation in the European hamster (Cricetus cricetus). J. Physiol. 72 (5), 653–661.
Canivenc R. and Bonnin M. (1975). Regulation of luteal activity by light and temperature in two delayed implantation mammals, the European badger and the pine marten. J. Physiol. Paris 70, 533–538.
Ceballos G. and Ehrlich P. R. (2002). Mammal population losses and the extinction crisis. Science 296, 904–907. doi: 10.1126/science.1069349
Chabot C. C., Connolly D. M., and Waring B. B. (2012). The effects of lighting conditions and food restriction paradigms on locomotor activity of common spiny mice, Acomys cahirinus. J. Circadian Rhythms. 10, 6. doi: 10.1186/1740-3391-10-6
Ciach M., Faraś H., Fröhlich A., and Fedyń I. (2022). Contrasting effects of light and noise pollution interact with natural vegetation remnants: Human-related indicators of the habitat suitability for ungulates in the urban landscape. Ecol. Indic. 142, 109261. doi: 10.1016/j.ecolind.2022.109261
Ciach M. and Fröhlich A. (2019). Ungulates in the city: light pollution and open habitats predict the probability of roe deer occurring in an urban environment. Urban Ecosyst. 22, 513–523. doi: 10.1007/s11252-019-00840-2
Clarke J. R. and Kennedy J. P. (1967). Effect of light and temperature upon gonad activity in the vole (Microtus agrestis). Gen. Comp. Endocrinol. 8, 474–488. doi: 10.1016/S0016-6480(67)80009-1
Cravens Z. M. and Boyles J. G. (2019). Illuminating the physiological implications of artificial light on an insectivorous bat community. Oecologia 189, 69–77. doi: 10.1007/s00442-018-4300-6
Davies T. W. and Smyth T. (2018). Why artificial light at night should be a focus for global change research in the 21st century. Global Change Biol. 24, 872–882. doi: 10.1111/gcb.2018.24.issue-3
Dirzo R., Young H. S., Galetti M., Ceballos G., Isaac N. J. B., and Collen B. (2014). Defaunation in the anthropocene. Science 345, 401–406. doi: 10.1126/science.1251817
Ditmer M. A., Francis C. D., Barber J. R., Stoner D. C., Seymoure B. M., Fristrup K. M., et al. (2021a). Assessing the vulnerabilities of vertebrate species to light and noise pollution: expert surveys illuminate the impacts on specialist species. Integr. Comp. Biol. 61, 1202–1215. doi: 10.1093/icb/icab091
Ditmer M. A., Stoner D. C., and Carter N. H. (2021b). Estimating the loss and fragmentation of dark environments in mammal ranges from light pollution. Biol. Conserv. 257, 109135. doi: 10.1016/j.biocon.2021.109135
Dominoni D. M., Halfwerk W., Baird E., Buxton R. T., Fernández-Juricic E., Fristrup K. M., et al. (2020). Why conservation biology can benefit from sensory ecology. Nat. Ecol. Evol. 4, 502–511. doi: 10.1038/s41559-020-1135-4
Douglazet M. (2016). Elaboration d’un protocole d’étude de l’impact de l’éclairage artificiel sur les déplacements des mammifères terrestres nocturnes. Parc naturel régional des Causses du Quercy. 114p.
Duffy J. P., Bennie J., Durán A. P., and Gaston K. J. (2015). Mammalian ranges are experiencing erosion of natural darkness. Sci. Rep. 5, 12042. doi: 10.1038/srep12042
Dupont S. M., Guinnefollau L., Weber C., and Petit O. (2019). Impact of artificial light at night on the foraging behavior of the European Hamster: consequences for the introduction of this species in suburban areas. Rethinking Ecol. 4, 133–148. doi: 10.3897/rethinkingecology.4.36467
Eilam D. (2004). Locomotor activity in common spiny mice (Acomys cahirinuse): The effect of light and environmental complexity. BMC Ecol. 4, 16. doi: 10.1186/1472-6785-4-16
Elvidge C. K., Reid C. H., Ford M. I., Sills M., Patrick P. H., Gibson D., et al. (2019). Ontogeny of light avoidance in juvenile lake sturgeon. J. Appl. Ichthyology 35, 202–209. doi: 10.1111/jai.2019.35.issue-1
Falchi F., Cinzano P., Duriscoe D., Kyba C. C. M., Elvidge C. D., Baugh K., et al. (2016). The new world atlas of artificial night sky brightness. Sci. Adv. 2. doi: 10.1126/sciadv.1600377
Falcon J., Torriglia A., Attia D., Viénot F., Gronfier C., Behar-Cohen F., et al. (2020). Exposure to artificial light at night and the consequences for flora, fauna, and ecosystems. Front. Neurosci. 14, 602796. doi: 10.3389/fnins.2020.602796
Farnworth B., Innes J., Kelly C., Littler R., and Waas J. R. (2018). Photons and foraging: Artificial light at night generates avoidance behavior in male, but not female, New Zealand weta. Environ. pollut. 236, 82–90. doi: 10.1016/j.envpol.2018.01.039
Farnworth B., Meitern R., Innes J., and Waas J. R. (2019). Increasing predation risk with light reduces speed, exploration and visit duration of invasive ship rats (Rattus rattus). Sci. Rep. 9, 3739. doi: 10.1038/s41598-019-39711-3
Finch D., Smith B., Marshall C., Coomber F., Kubasiewicz L., Anderson M., et al. (2020). Effects of Artificial Light at Night (ALAN) on European Hedgehog Activity at Supplementary Feeding Stations. Animals 10, 768. doi: 10.3390/ani10050768
Frampton G. K., Livoreil B., and Petrokofsky G. (2017). Eligibility screening in evidence synthesis of environmental management topics. Environ. Evidence 6, 27. doi: 10.1186/s13750-017-0102-2
Francis M. J., Spooner P. G., and Matthews A. (2015). The influence of urban encroachment on squirrel gliders (Petaurus norfolcensis): Effects of road density, light and noise pollution. Wildlife Res. 42, 324–333. doi: 10.1071/WR14182
Gaynor K. M., Hojnowski C. E., Carter N. H., and Brashares J. S. (2018). The influence of human disturbance on wildlife nocturnality. Science 360, 1232–1235. doi: 10.1126/science.aar7121
Gerkema M. P., Daan S., Wilbrink M., Hop M. W., and Van Der Leest F. (1993). Phase control of ultradian feeding rhythms in the common vole (Microtus arvalis): the roles of light and the circadian system. J. Biol. 8, 151–171. doi: 10.1177/074873049300800205
Giavi S., Fontaine C., and Knop E. (2021). Impact of artificial light at night on diurnal plant-pollinator interactions. Nat. Commun. 12, 1690. doi: 10.1038/s41467-021-22011-8
Gordigiani L., Viviano A., Brivio F., Grignolio S., Lazzeri L., Marcon A., et al. (2022). Carried away by a moonlight shadow: activity of wild boar in relation to nocturnal light intensity. Mammal Res. 67, 39–49. doi: 10.1007/s13364-021-00610-6
Grenis K., Nufio C., Wimp G. M., and Murphy S. M. (2023). Does artificial light at night alter moth community composition? Philos. Trans. R. Soc. B. 378I (1892). doi: 10.1098/rstb.2022.0365
Grubisic M., Haim A., Bhusal P., Dominoni D. M., Gabriel K. M. A., Jechow A., et al. (2019). Light pollution, circadian photoreception, and melatonin in vertebrates. Sustainability 11, 6400. doi: 10.3390/su11226400
Haddaway N. R., Bernes C., Jonsson B.-G., and Hedlund K. (2016). The benefits of systematic mapping to evidence-based environmental management. Ambio 45, 613–620. doi: 10.1007/s13280-016-0773-x
Haddaway N. R., Macura B., Whaley P., and Pullin A. S. (2018). ROSES RepOrting standards for Systematic Evidence Syntheses: pro forma, flow-diagram and descriptive summary of the plan and conduct of environmental systematic reviews and systematic maps. Environ. Evidence 7, 7. doi: 10.1186/s13750-018-0121-7
Haddock J. K., Threlfall C. G., Law B., and Hochuli D. F. (2019). Light pollution at the urban forest edge negatively impacts insectivorous bats. Biol. Conserv. 236, 17–28. doi: 10.1016/j.biocon.2019.05.016
Haim A., Shanas U., Zubidad A. E. S., and Scantelbury M. (2005). Seasonality and seasons out of time—the thermoregulatory effects of light interference. Chronobiol. Int. 22, 59–66. doi: 10.1081/CBI-200038144
Hall K. J. and Fleming P. A. (2021). In the spotlight: Can lights be used to mitigate fox predation on a free-range piggery? Appl. Anim. Behav. Sci. 242, 105420. doi: 10.1016/j.applanim.2021.105420
Hanizar E., Huang Y.-S., Siswoyo T. A., and Aswan M. S. (2021). Light environment effect in the sperm and ribonucleic acid quality and body weight of male mus musculus. Open Access Macedonian J. Med. Sci. 9, 644–650. doi: 10.3889/oamjms.2021.6535
Hannah K., Haddaway N. R., Fuller R. A., and Amano T. (2024). Language inclusion in ecological systematic reviews and maps: Barriers and perspectives. Res. Synthesis Methods 15, 466–482. doi: 10.1002/jrsm.v15.3
Harzing A. W. (2007). Publish or perish. Version 7. Available online at: https://harzing.com/resources/publish-or-perish.
Hemami M.-R., Naderi G., Karami M., and Mohammadi S. (2011). Nocturnal activity of Iranian jerboa, Allactaga firouzi (Mammalia: Rodentia: Dipodidae). mamm 75, 31–34. doi: 10.1515/mamm.2010.062
Hernández M. C., Jara-Stapfer D. M., Muñoz A., Bonacic C., Barja I., and Rubio A. V. (2021). Behavioral responses of wild rodents to owl calls in an austral temperate forest. Animals 11, 428. doi: 10.3390/ani11020428
Hoffmann J., Palme R., and Eccard J. A. (2018). Long-term dim light during nighttime changes activity patterns and space use in experimental small mammal populations. Environ. pollut. 238, 844–851. doi: 10.1016/j.envpol.2018.03.107
Hoffmann J., Schirmer A., and Eccard J. A. (2019). Light pollution affects space use and interaction of two small mammal species irrespective of personality. BMC Ecol. 19, 26. doi: 10.1186/s12898-019-0241-0
Holker F., Wolter C., Perkin E. K., and Tockner K. (2010). Light pollution as a biodiversity threat. Trends Ecol. Evol. 25, 681–682. doi: 10.1016/j.tree.2010.09.007
Hut R. A., Scheper A., and Daan S. (2000). Can the circadian system of a diurnal and a nocturnal rodent entrain to ultraviolet light? J. Comp. Physiol. A. 186, 707–715. doi: 10.1007/s003590000124
Issad S. M., Benhafri N., El Allali K., Farsi H., Ouali-Hassenaoui S., and Dekar-Madoui A. (2021). Effects of prolonged night-time light exposure and traffic noise on the behavior and body temperature rhythmicity of the wild desert rodent, Gerbillus tarabuli. Chronobiology Int. 38, 415–425. doi: 10.1080/07420528.2020.1862858
Jansen K., Van der Zee E. A., and Gerkema M. P. (1998). Concurrent decrease of vasopressin and protein kinase Ca immunoreactivity during the light phase in the vole suprachiasmatic nucleus. Neurosci. Letters. 248 (2), 81–84. doi: 10.1016/S0304-3940(98)00271-7
Kott O., Šumbera R., and Němec P. (2010). Light perception in two strictly subterranean rodents: life in the dark or blue? PloS One 5, e11810. doi: 10.1371/journal.pone.0011810
Laguna E., Palencia P., Carpio A. J., Mateos-Aparicio J., Herraiz C., Notario C., et al. (2022). Evaluation of a combined and portabl e light-ultrasound device with which to deter red deer. Eur. J. Wildlife Res. 68, 50. doi: 10.1007/s10344-022-01599-2
La Sorte F. A., Fink D., Buler J. J., Farnsworth A., and Cabrera-Cruz S. A. (2017). Seasonal associations with urban light pollution for nocturnally migrating bird populations. Global Change Biol. 23, 4609–4619. doi: 10.1111/gcb.2017.23.issue-11
Lei T., Hua H., Du H., Xia J., Xu D., Liu W., et al. (2024). Molecular mechanisms of artificial light at night affecting circadian rhythm disturbance. Arch. Toxicol. 98, 395–408. doi: 10.1007/s00204-023-03647-5
Le Tallec T., Théry M., and Perret M. (2016). Melatonin concentrations and timing of seasonal reproduction in male mouse lemurs (Microcebus murinus) exposed to light pollution. J. Mammalogy 97, 753–760. doi: 10.1093/jmammal/gyw003
Livoreil B., Glanville J., Haddaway N. R., Bayliss H., Bethel A., de Lachapelle F. F., et al. (2017). Systematic searching for environmental evidence using multiple tools and sources. Environ. Evidence 6, 23. doi: 10.1186/s13750-017-0099-6
Luo B., Xu R., Li Y., Zhou W., Wang W., Gao H., et al. (2021). Artificial light reduces foraging opportunities in wild least horseshoe bats. Environ. pollut. 288, 117765. doi: 10.1016/j.envpol.2021.117765
Luscier J. D., Christopher A., Synan H., and Wilson K. B. (2023). Effects of light and noise pollution on occupancy of gray treefrogs (Hyla versicolor) and green frogs (Lithobates clamitans) in Syracuse, NY. Urban Ecosyst. 26, 941–953. doi: 10.1007/s11252-023-01329-9
Marín-Gómez O. H., García-Arroyo M., Sánchez-Sarria C. E., Sosa-López J. R., Santiago-Alarcon D., and MacGregor-Fors I. (2020). Nightlife in the city: drivers of the occurrence and vocal activity of a tropical owl. Avian Res. 11, 9. doi: 10.1186/s40657-020-00197-7
Mathiaparanam K. J., Mulder R. A., and Hale R. (2024). Anthropogenic double jeopardy: Urban noise and artificial light at night interact synergistically to influence abundance. Environ. pollut. 363, 125078. doi: 10.1016/j.envpol.2024.125078
Matzke E. B. (1936). The effect of street lights in delaying leaf-fall in certain trees. Am. J. Bot. 23, 446–452. doi: 10.2307/2436035
Mena J. L., Rivero J., Bonifaz E., Pastor P., Pacheco J., and Aide T. M. (2022). The effect of artificial light on bat richness and nocturnal soundscapes along an urbanization gradient in an arid landscape of central Peru. Urban Ecosyst. 25, 563–574. doi: 10.1007/s11252-021-01163-x
Milon A., Pawlicki P., Rak A., Mlyczynska E., Płachno B. J., Tworzydlo W., et al. (2019). Telocytes are localized to testis of the bank vole (Myodes glareolus) and are affected by lighting conditions and G-coupled membrane estrogen receptor (GPER) signaling. Gen. Comp. Endocrinol. 271, 39–48. doi: 10.1016/j.ygcen.2018.10.021
Moaraf S., Vistoropsky Y., Pozner T., Heiblum R., Okuliarová M., Zeman M., et al. (2020). Artificial light at night affects brain plasticity and melatonin in birds. Neurosci. Lett. 716, 134639. doi: 10.1016/j.neulet.2019.134639
Pérez Vega C., Zielinska-Dabkowska K. M., Schroer S., Jechow A., and Hölker F. (2022). A systematic review for establishing relevant environmental parameters for urban lighting: translating research into practice. Sustainability 14, 1107. doi: 10.3390/su14031107
Pohl H. (1976). Proportional effect of light on entrained circadian rhythms of birds and mammals. J. Comp. Physiol. 112, 103–108. doi: 10.1007/BF00612678
Procko M., Naidoo R., LeMay V., and Burton A. C. (2023). Human presence and infrastructure impact wildlife nocturnality differently across an assemblage of mammalian species. PloS One 18, e0286131. doi: 10.1371/journal.pone.0286131
PrudHon M., Goussopoulos J., and Reyne Y. (1978). Comportement alimentaire du lapin de garenne élevé en captivité. II. - Modifications induites par un décalage des périodes d’éclairement. Ann. Zootech. 27, 101–106.
Prugh L. R. and Golden C. D. (2014). Does moonlight increase predation risk? Meta-analysis reveals divergent responses of nocturnal mammals to lunar cycles(S Boutin, Ed). J. Anim. Ecol. 83, 504–514. doi: 10.1111/jane.2014.83.issue-2
Reyne Y., Goussopoulos M., and Prud'hon M. (1979). Comportement alimentaire du Lapin de Garenne élevé en captivité. III. — Etude des rythmes d’ingestion d’aliment et d’eau en lumière permanente. Ann. Zootech. 282, 159–164. doi: 10.1051/animres:19790202
Richardson W. S., Wilson M. C., Nishikawa J., and Hayward R. S. (1995). The well-built clinical question: a key to evidence-based decisions. ACP J. club 123, A12–A13. doi: 10.7326/ACPJC-1995-123-3-A12
Richoux J. P. and Legait E. (1975). Influence of constant darkness or light on the evolution of dormouse ovary during the winter period. Archives d'Anatomie, d'Histologie et d'Embryologie. 58, 67–88.
Robert K. A., Lesku J. A., Partecke J., and Chambers B. (2015). Artificial light at night desynchronizes strictly seasonal reproduction in a wild mammal. Proc. R. Soc. B-Biological Sci. 282. doi: 10.1098/rspb.2015.1745
Rodrigo-Comino J., Seeling S., Seeger M. K., and Ries J. B. (2021). Light pollution: A review of the scientific literature. Anthropocene Rev. 10 (2), 205301962110512. doi: 10.1177/205301962110512
Rotics S., Dayan T., and Kronfeld-Schor N. (2011a). Effect of artificial night lighting on temporally partitioned spiny mice. J. Mammalogy 92, 159–168. doi: 10.1644/10-MAMM-A-112.1
Rotics S., Dayan T., Levy O., and Kronfeld-Schor N. (2011b). Light masking in the field: an experiment with nocturnal and diurnal spiny mice under semi-natural field conditions. Chronobiology Int. 28, 70–75. doi: 10.3109/07420528.2010.525674
Roux M. and Richoux J. P. (1981). Effect of castration and adrenalectomy on the ultrastructure of pinealocytes in the female garden dormouse (Eliomys quercinus L.) under different conditions of lighting and temperature during the normal period of hibernation. J. Neural. Transmission. 51, 283–294.
Roux M. (1985). Ultrastructural study of the Sertoli cell in the seminiferous tubules of a hibernating animal, Eliomys quercinus L.), living in different lighting conditions. Bulletin de l'Association des Anatomistes. 69 (205), 63–75.
Rydell J., Michaelsen T. C., Sanchez-Navarro S., and Eklöf J. (2021). How to leave the church: light avoidance by brown long-eared bats. Mamm. Biol. 101, 979–986. doi: 10.1007/s42991-021-00154-x
Saboureau M. and El Omari B. (1993). Effect of light entrainment and temperature on the reproductive cycle in the male hedgehog (Erinaceus europaeus). Reproduction 99, 497–504. doi: 10.1530/jrf.0.0990497
Saboureau M., Vivien-Roels B., and Pevet P. (1991). Pineal melatonin concentrations during day and night in the adult hedgehog: Effect of a light pulse at night and superior cervical ganglionectomy. J. Pineal Res. 11, 92–98. doi: 10.1111/j.1600-079X.1991.tb00462.x
Saldaña-Vázquez R. A. and Munguía-Rosas M. A. (2013). Lunar phobia in bats and its ecological correlates: A meta-analysis. Mamm. Biol. 78, 216–219. doi: 10.1016/j.mambio.2012.08.004
Sánchez de Miguel A., Bennie J., Rosenfeld E., Dzurjak S., and Gaston K. J. (2022). Environmental risks from artificial nighttime lighting widespread and increasing across Europe. Sci. Adv. 8, eabl6891. doi: 10.1126/sciadv.abl6891
Sanders D., Frago E., Kehoe R., Patterson C., and Gaston K. J. (2021). A meta-analysis of biological impacts of artificial light at night. Nat. Ecol. Evol. 5, 74–81. doi: 10.1038/s41559-020-01322-x
Schoeman M. C. (2016). Light pollution at stadiums favors urban exploiter bats: Selected urban exploiter bats hunt insects at stadiums. Anim. Conserv. 19, 120–130. doi: 10.1111/acv.12220
Shier D. M., Bird A. K., and Wang T. B. (2020). Effects of artificial light at night on the foraging behavior of an endangered nocturnal mammal. Environ. pollut. 263, 114566. doi: 10.1016/j.envpol.2020.114566
Stone E. L., Jones G., and Harris S. (2012). Conserving energy at a cost to biodiversity? Impacts of LED lighting on bats. Global Change Biol. 18, 2458–2465. doi: 10.1111/j.1365-2486.2012.02705.x
Straka T. M., Greif S., Schultz S., Goerlitz H. R., and Voigt C. C. (2020). The effect of cave illumination on bats. Global Ecol. Conserv. 21, e00808. doi: 10.1016/j.gecco.2019.e00808
Svechkina A., Portnov B. A., and Trop T. (2020). The impact of artificial light at night on human and ecosystem health: a systematic literature review. Landscape Ecol. 35, 1725–1742. doi: 10.1007/s10980-020-01053-1
Touzot M., Lefebure T., Lengagne T., Secondi J., Dumet A., Konecny-Dupre L., et al. (2022). Transcriptome-wide deregulation of gene expression by artificial light at night in tadpoles of common toads. Sci. Total Environ. 818, 151734. doi: 10.1016/j.scitotenv.2021.151734
Urbanski J., Mogi M., O’Donnell D., DeCotiis M., Toma T., and Armbruster P. (2012). Rapid Adaptive Evolution of Photoperiodic Response during Invasion and Range Expansion across a Climatic Gradient. Am. Nat. 179, 490–500. doi: 10.1086/664709
van der Veen D. R., Saaltink D. J., and Gerkema M. P. (2011). Behavioral responses to combinations of timed light, food availability, and ultradian rhythms in the common vole (Microtus arvalis). Chronobiol. Int. 28 (7), 563–571. doi: 10.3109/07420528.2011.591953
van Jaarsveld B., Bennett N. C., Hart D. W., and Oosthuizen M. K. (2019). Locomotor activity and body temperature rhythms in the Mahali mole-rat (C. h. mahali): The effect of light and ambient temperature variations. J. Thermal Biol. 79, 24–32. doi: 10.1016/j.jtherbio.2018.11.013
Vardi-Naim H., Benjamin A., Sagiv T., and Kronfeld-Schor N. (2022). Fitness consequences of chronic exposure to different light pollution wavelengths in nocturnal and diurnal rodents. Sci. Rep. 12, 16486. doi: 10.1038/s41598-022-19805-1
Voigt C. C., Scholl J. M., Bauer J., Teige T., Yovel Y., Kramer-SChadt S., et al. (2020). Movement responses of common noctule bats to the illuminated urban landscape. Landscape Ecol. 35, 189–201. doi: 10.1007/s10980-019-00942-4
Vuillez P., Jacob N., Teclemariam-Mesbah R., and Pévet P. (1996). In Syrian and European hamsters, the duration of sensitive phase to light of the suprachiasmatic nuclei depends on the photoperiod. Neurosci. Lett. 208, 37–40. doi: 10.1016/0304-3940(96)12535-0
Ware J. V., Nelson O. L., Robbins C. T., and Jansen H. T. (2012). Temporal organization of activity in the brown bear (Ursus arctos): roles of circadian rhythms, light, and food entrainment. Am. J. Physiology-Regulatory Integr. Comp. Physiol. 303, R890–R902. doi: 10.1152/ajpregu.00313.2012
Webb S. M. (1984). Immunoreactive somatostatin in the pineal gland of different rodent species: circadian rhythm, effects of superior cervical, ganglionectomy, pineal indole administration and lighting conditions. Biomedical Research. 5 (6), 473–480. doi: 10.2220/biomedres.5.473
Weiler U., Claus L., Dehnhard M., and Hofacker S. (1996). Influence of the photoperiod and a light reverse program on metabolically active hormones and food intake in domestic pigs compared with a wild boar. Can. J. Anim. Sci. 764, 531–539. doi: 10.4141/cjas96-080
Yang Y., Liu Q., Pan C., Chen J., Xu B., Liu K., et al. (2024). Species sensitivities to artificial light at night: A phylogenetically controlled multilevel meta-analysis on melatonin suppression. Ecol. Lett. 27, e14387. doi: 10.1111/ele.14387
Zeale M. R. K., Stone E. L., Zeale E., Browne W. J., Harris S., and Jones G. (2018). Experimentally manipulating light spectra reveals the importance of dark corridors for commuting bats. Global Change Biol. 24, 5909–5918. doi: 10.1111/gcb.2018.24.issue-12
Zeman M., Okuliarova M., and Rumanova V. S. (2023). Disturbances of hormonal circadian rhythms by light pollution. Int. J. Mol. Sci. 24, 7255. doi: 10.3390/ijms24087255
Zhang F.-S., Wang Y., Wu K., Xu W.-Y., Wu J., Liu J.-Y., et al. (2020). Effects of artificial light at night on foraging behavior and vigilance in a nocturnal rodent. Sci. Total Environ. 724, 138271. doi: 10.1016/j.scitotenv.2020.138271
Zou W., Wu P., Wei X., Zhou D., Deng Y., Jiang Y., et al. (2024). Artificial light affects foraging behavior of a synanthropic bat. Integr. Zoology 19, 710–720. doi: 10.1111/1749-4877.12787
Zubidat A. E., Ben-Shlomo R., and Haim A. (2007). Thermoregulatory and endocrine responses to light pulses in short-day acclimated social voles (Microtus socialis). Chronobiology Int. 24, 269–288. doi: 10.1080/07420520701284675
Zubidat A. E., Nelson R. J., and Haim A. (2009). Photosensitivity to different light intensities in blind and sighted rodents. J. Exp. Biol. 212, 3857–3864. doi: 10.1242/jeb.033969
Zubidat A. E., Nelson R. J., and Haim A. (2010). Photoentrainment in blind and sighted rodent species: responses to photophase light with different wavelengths. J. Exp. Biol. 213, 4213–4222. doi: 10.1242/jeb.048629
Keywords: light pollution, ALAN, night-time lighting, LEDs (light emitting diodes), illumination, sensory pollutant, anthropocene, wildlife
Citation: Sordello R, Fournier C, Tanaka T and Thierry C (2025) What evidence exists on the impacts of artificial light on terrestrial non-flying mammals native to Europe? A systematic map and a narrative review. Front. Ecol. Evol. 13:1544308. doi: 10.3389/fevo.2025.1544308
Received: 12 December 2024; Accepted: 26 March 2025;
Published: 26 May 2025; Corrected: 01 July 2025.
Edited by:
Fernanda Michalski, Universidade Federal do Amapá, BrazilReviewed by:
Reinhard Arnold Klenke, Martin Luther University of Halle-Wittenberg, GermanyDani Broitman, Technion Israel Institute of Technology, Israel
Copyright © 2025 Sordello, Fournier, Tanaka and Thierry. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Romain Sordello, cm9tYWluLnNvcmRlbGxvQG1uaG4uZnI=