 Ella Vázquez-Domínguez
Ella Vázquez-Domínguez Brenda Solórzano-García
Brenda Solórzano-García Mariana Reyes-Vázquez1
Mariana Reyes-Vázquez1- 1Laboratorio de Genética y Ecología, Departamento de Ecología de la Biodiversidad, Instituto de Ecología, Universidad Nacional Autónoma de México, Ciudad de México, Mexico
- 2Laboratorio de Parasitología y Medicina de la Conservación, ENES-Mérida UNAM, Ucú, Yucatán, Mexico
Genetic diversity is of prime importance for biological diversity and for the long-term conservation and survival of species and populations. The conservation of hybrid individuals originating from natural hybridization is still controversial; nearly every hybrid system is different and general conservation rules are not effective. The black howler monkey (Alouatta pigra) is and endangered primate endemic to the Yucatán peninsula. It hybridizes with the mantled howler, Alouatta palliata, in a narrow contact zone located in southern Mexico. We evaluated the genetic diversity and structure patterns in natural populations of A. pigra across its distribution range in Mexico (16 localities), including the hybrid populations, and estimated the overall time of divergence of the hybrid individuals. We successfully genotyped 12 microsatellite loci and obtained a 587 bp cytochrome-b fragment alignment for 222 and 107 individuals, respectively. We identified 33 hybrids, three A. palliata and 186 non-admixed A. pigra. Our results showed the highest diversification for A. pigra in Mexico occurring at ~1.4 My, while identifying a more recent diversification event including A. palliata and hybrids (~0.56 My), supporting different diversification events along these primates’ evolutionary history. Also, divergence of hybrid individuals is not a contemporary process in the evolutionary history of these sister species, suggesting they came into contact at the zone of sympatry at least some ~0.8 My ago. Hybrids exhibited higher mitochondrial diversity in comparison with non-admixed individuals; notably, hybrids with A. pigra mitochondria exhibited both higher diversity values and number of unique haplotypes than those with A. palliata mitochondria. Comparatively, hybrid nuclear genetic diversity was higher compared with A. palliata but not with A. pigra. Regarding the A. pigra populations, we found moderate diversity values, being highest in the most conserved and continuous habitat. We also found significant genetic differentiation (five genetic clusters) and limited gene flow between them, and potential inbreeding signals, which render A. pigra prone to drift effects and increased extinction risk. Our study provides novel information regarding the historical and contemporary genetic patterns of black howler monkey and hybrid populations in Mexico, which is key for developing strategies for conserving and ensuring the survival of viable primate populations.
Introduction
Genetic diversity is crucial for the understanding of spatial, temporal, and evolutionary processes of biological diversity and of prime importance for its conservation. Additionally, disentangling if species and populations are divided into different evolutionary genetic units, as well as the structure among them, is key for the protection of wildlife (Ralls et al., 2018; Ruiz-García et al., 2019). Therefore, preserving adequate genetic variability levels is key for the long-term survival of species and populations (Allendorf et al., 2013; Ralls et al., 2018). Even though conservation efforts have focused on preserving diversity at several levels, intraspecific genetic diversity has received little attention in environmental conservation policy and wildlife management (Coates et al., 2018; Ralls et al., 2018).
Hybridization, the reproduction between individuals of different species or from otherwise genetically distinct lineages, is an important evolutionary process that provides information about the nature of species and species boundaries (Abbott et al., 2013); it is relatively frequent in nature and also a significant source of variation (Abbott et al., 2013; Sardell and Uy, 2016; Gompert et al., 2017). Evidence has shown that recent, rapidly radiating, closely related species are most likely to hybridize (Brennan et al., 2012; Lamichhaney et al., 2017). Notably, the conservation of hybrid individuals originating from natural hybridization is a controversial ongoing debate (Jackiw et al., 2015; Suárez-Atilano et al., 2024). Two main processes stimulate such debate, on the one hand hybrids can have greater fitness allowing for adaptation to novel environments (Allendorf et al., 2001; Edmands, 2007; Pacheco-Sierra et al., 2018); whereas on the other, hybridization and introgression (integration of DNA from one species into another via the backcrossing of hybrids with parental types of one or both species) can contribute to extinction, for example when endemic or rare species come into contact with non-native or more abundant species (Allendorf et al., 2001; Jackiw et al., 2015).
Knowledge and conservation of the genetic diversity of species, populations and hybrid individuals has become more and more important given the accelerated habitat loss, fragmentation, modification, and isolation of natural environments due to human activities (Moraes et al., 2017; Coates et al., 2018; Oklander and Soto-Calderón, 2024). This is particularly critical in tropical forest ecosystems, which harbor high levels of species richness, endangered species and many endemisms (Radespiel and Bruford, 2014; Oklander and Soto-Calderón, 2024). Such habitat modification leads to decreases of effective size and isolation of natural populations, limiting gene flow, reducing genetic variation and increasing genetic differentiation (Mimura et al., 2017; Coates et al., 2018). Hence, the urgency of incorporating information about the evolutionary history and population genetics of threatened plants and animals for their effective management and conservation actions.
Howler monkeys (Alouatta spp.) are among the largest non-human primates that inhabit Neotropical regions from southern Mexico to north-eastern Argentina. They are able to survive in relatively small forest remnants and to tolerate different degrees of habitat quality, given their plastic diet and activity patterns (Bicca-Marques, 2003; Solórzano-García et al., 2021). However, habitat perturbation can negatively impact their populations, losing genetic diversity and showing physiological stress and high parasitism, among others (Arroyo-Rodríguez and Dias, 2010; Cristobal-Azkarate and Dunn, 2013; Rimbach et al., 2013; Solórzano-García et al., 2021; Pozo-Montuy et al., 2024). The black howler monkey (Alouatta pigra Lawrence, 1933) (Cortés-Ortiz et al., 2015) is a diurnal arboreal primate, with sexual dimorphism (males are bigger than females), that lives in groups of 2–12 individuals and has a mostly folivorous diet (Van Belle and Estrada, 2006). It is endemic to the Yucatán peninsula, including southeastern Mexico, Belize and northern Guatemala, and is classified as Endangered by Mexican authorities (SEMARNAT, 2010) and by the IUCN (Cortés-Ortíz et al., 2020). Jointly with its sister species, the mantled howler monkey (Alouatta palliata Gray, 1849) (Cortés-Ortiz et al., 2015), they represent the northernmost distribution of howler monkeys in America.
Despite the high risk and vulnerability of A. pigra’s populations, only a few studies have examined the genetic diversity and structure of its populations albeit on a strictly local scale (James et al., 1997; Ellsworth and Hoelzer, 2006; García del Valle et al., 2005; Améndola, 2009; Van Belle et al., 2012; Baiz and Cortés-Ortiz, 2015). Alouatta pigra hybridizes with A. palliata in a narrow contact zone located in southern Mexico where their ranges overlap, composed of a highly fragmented landscape with isolated remnants of habitat immersed in an anthropogenic matrix. Many aspects of this hybridization process have been intensively studied. These studies have shown that most hybrids are multigenerational with few being the product of crosses between parentals and recent generation hybrids, based on nuclear (microsatellite loci) and mitochondrial and nuclear DNA sequences (Ho et al., 2014; Cortés-Ortiz et al., 2015, 2019; Cortés-Ortiz, 2017). Recently, Baiz et al. (2019) reported, using genomic (ddRadSeq) data, that this hybrid system is characterized by a lack of early-generation male hybrids and in which multiple forms of selection have shaped the evolution of reproductive isolation. But the genetics of the hybrid populations in comparison with the patterns of the parental species has not been specifically reported.
Our main objective in the present study was to evaluate the genetic diversity and differentiation patterns of A. pigra across its distribution range in Mexico, including the hybrid populations. We also aimed to estimate the overall time of divergence of the hybrid individuals. We assessed microsatellite loci and mitochondrial (cytochrome b) data in natural populations. Considering that habitat modification and fragmentation leads to reduced genetic diversity and increased genetic differentiation, we predicted low genetic diversity values in A. pigra populations at the local and regional level. Additionally, we expected reduced connectivity and high genetic differentiation among populations given A. pigra’s tight association with arboreal vegetation for its establishment and its limited dispersal capacity. Regarding hybrid populations, extensive introgression in both directions and a large array of admixed individuals have been reported; thus, we expected higher genetic diversity in the hybrid populations in comparison with the parentals. Finally, divergence of hybrid individuals would have started far back in the evolutionary history of these sister species (i.e. historical rather than contemporary).
Materials and methods
Sample collection
We employed a non-invasive sampling technique to obtain DNA information from fecal samples of A. pigra across its distribution in southeastern Mexico. We collected fecal samples during 2020 and 2022 from 223 individuals from 16 localities and a total of 40 groups plus four solitary individuals, including samples from hybrid individuals from 10 groups inhabiting the contact zone between A. pigra and A. palliata (Figure 1A; Supplementary Table S1). Black howler monkey groups were detected via their morning calls or by direct sighting. Fresh fecal material was collected right after deposition, preserved in dry NaCl (Oklander et al., 2004) in 50 ml tubes; they were kept at room temperature up to one week in the field, then stored at -20°C in the laboratory until DNA extraction. For each collected sample, we registered the sex and age (infant, juvenile, adult) of the individual; we also documented the group size and composition, and the geographic location. Fecal sample collection was entirely non-invasive and entailed no contact with howler monkey individuals.
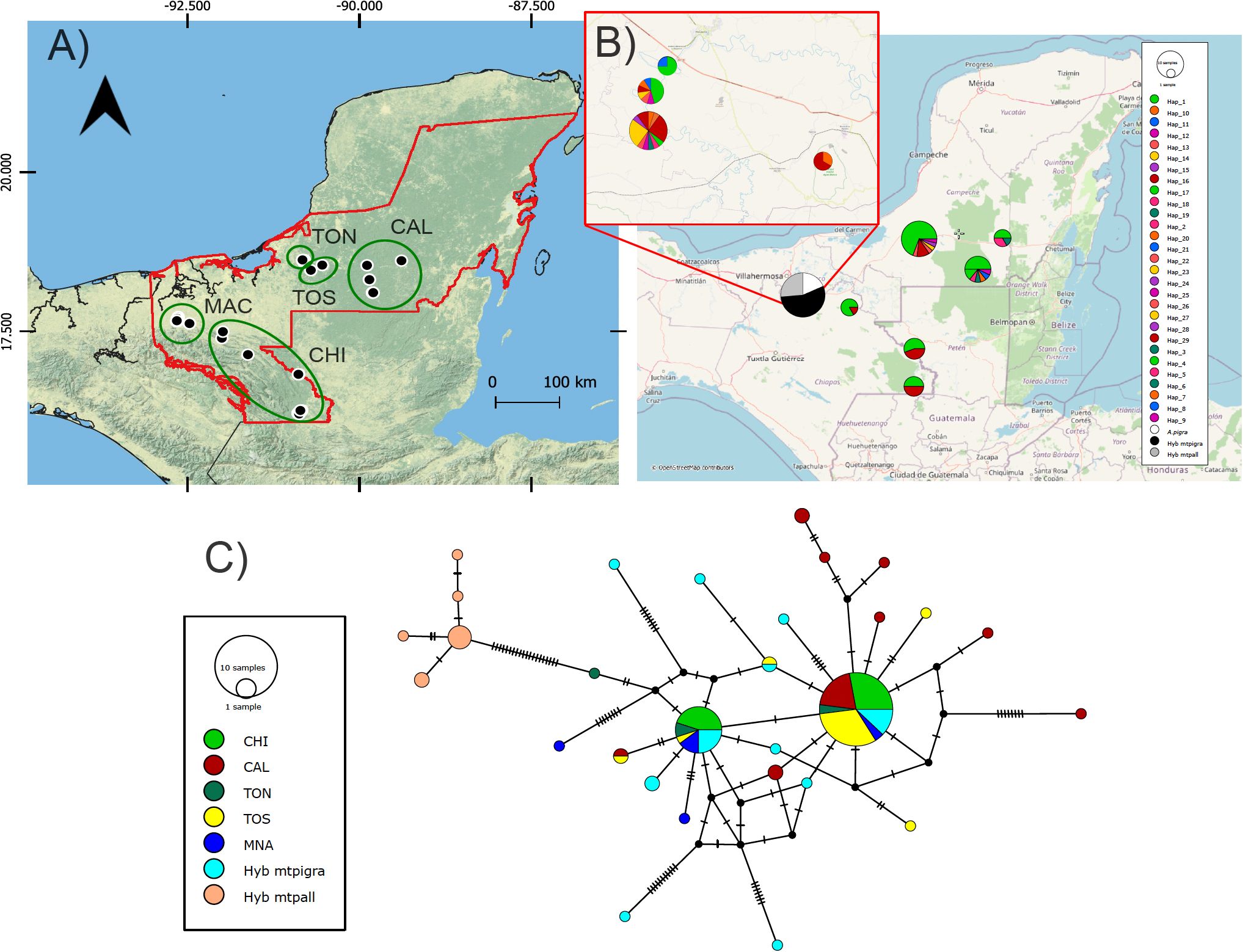
Figure 1. (A) Map of sampling of Alouatta pigra in Mexico. Native non-admixed and hybrid individuals sampled across 16 sampling localities (see Supplementary Table S1). Five genetic clusters (based on Structure; see Results) are indicated: 1) Macuspana (MAC), 2) Chiapas (CHI), 3) Tormento norte (TON), 4) Tormento sur (TOS), 5) Calakmul (CAL). Red line polygon of black howler distribution modified from Ceballos et al. (2006) (B) Pie charts depict the distribution of 29 unique mitochondrial cytochrome b haplotypes. The insert shows the localities in the contact zone in Macuspana where hybrid individuals are distributed. (C) Median-joining network based on cytochrome b haplotypes, colored by genetic cluster. MNA refers to the non-admixed A. pigra individuals from the contact zone; Hyb mtpigra, hybrids with A. pigra mitochondria; and Hyb mtpall, hybrids with A. palliata mitochondria.
Mitochondrial DNA amplification and microsatellite genotyping
DNA extraction from fecal samples was performed using the QIAamp DNA Stool Mini Kit (Qiagen) following the manufacturer’s instructions, with an overnight digestion step modification. We amplified a fragment of the mitochondrial cytochrome b gene (cyt-b) for a subset of 117 individuals, using the forward primer MVZ-03 5’ GCTTCCATCCAACATCTCAGCATGATG 3’ and the reverse MVZ-26 5’ AGATCTTTGATTGTGTAGTAGGGGT3’ (Smith and Patton, 1993). PCR conditions were initial denaturation at 94°C for 1 min, followed by 30 cycles at 94°C for 1 min, 45°C°for 1 min, 72°C for 2 min, and a post-amplification extension for 7 min at 72°C. PCR products were sent to Macrogen Korea for sequencing services. Assembling and base-calling of sequences were performed using Geneious v.5.1.7 (Biomatters); also, cyt-b sequences were translated into amino acids for accuracy evaluation, using the vertebrate mitochondrial genetic code to assess the presence of stop codons. We successfully sequenced and aligned 107 samples (587 bp; full cyt-b dataset), including both hybrid and A. pigra individuals.
We screened 12 fluorescently labelled microsatellite markers previously used in howler monkeys (Cortés-Ortiz et al., 2010, 2019) (Supplementary Table S2), some of which are diagnostic for identification of hybrid individuals. PCR reactions were carried out following the conditions described in Supplementary Table S2. To assure correct readings and identification of potential contamination, we performed 2–3 replicates based on the reproducibility and quality of each locus on the sequencing chromatograms. The replicates were done using different set of samples for each microsatellite locus, namely all samples had more than one replicate for different loci. The samples that yielded inconsistent results even after three replicates were considered as missing data. Only one sample had to be eliminated. We also included positive and negative controls in all runs. Microsatellite products were multiplexed and sent for sequencing to the UIUC Core Sequencing Facility at the University of Illinois with ROX-500 as internal size standard, in a Applied Biosystems 3730xl DNA analyzer. Allele size was determined with GeneMarker v.2.2 (SoftGenetics).
Identification of hybrid individuals
We did a preliminary identification on the field along the hybrid zone considering if they showed an A. pigra or A. palliata phenotype. Next, using the full cyt-b dataset plus four sequences from GenBank (AY065884, AY065885 and DQ875710 from A. pigra and MW452920 from A. palliata), we first built a phylogenetic tree using a Bayesian inference (BI) method, in MrBayes v.3.2.6 (Ronquist et al., 2012) and the CIPRES Science Gateway (Miller et al., 2010). BI analyses included HKY+ G as the best substitution model, two simultaneous runs of Markov chain Monte Carlo, each for four million generations, sampling trees every 4000 generations, a heating parameter value of 0.2, and a 25% burn-in. A 50% majority-rule consensus tree was constructed from the post burn-in trees. Based on the resulting tree (Supplementary Figure S1a) we classified individuals as having mtDNA of A. palliata or of A. pigra (Supplementary Table S1). Additionally, we identified diagnostic microsatellite loci (see the microsatellites section below), namely alleles characteristic of either parental species that were identified together in the same individual (A. palliata parental genotypes from Solórzano-García et al., 2021). Finally, based on the microsatellite genotypes of all individuals, we implemented a Bayesian clustering method with Structure (Pritchard et al., 2000). We applied the admixture model with independent allele frequencies in a series of 20 independent runs per K, ranging from K=1–16 in a first trial, and K=1–10 for the final one; 100,000 Markov chain Monte Carlo generations and a burn-in of 50,000 iterations. deltaK results were best for K=2 (lnP(K)= -7247.82) and K=3 (lnP(K)= -6919.52), which clearly separated the hybrid individuals; K=7 was the most informative of the differentiation across the entire distribution for both hybrids and non-hybrids (see Figure 2, Supplementary Figure S2). Hence, final identification of hybrid individuals was based on the combined information of phenotype, mitochondrial species identity, and diagnostic alleles, corroborated by the structure results.
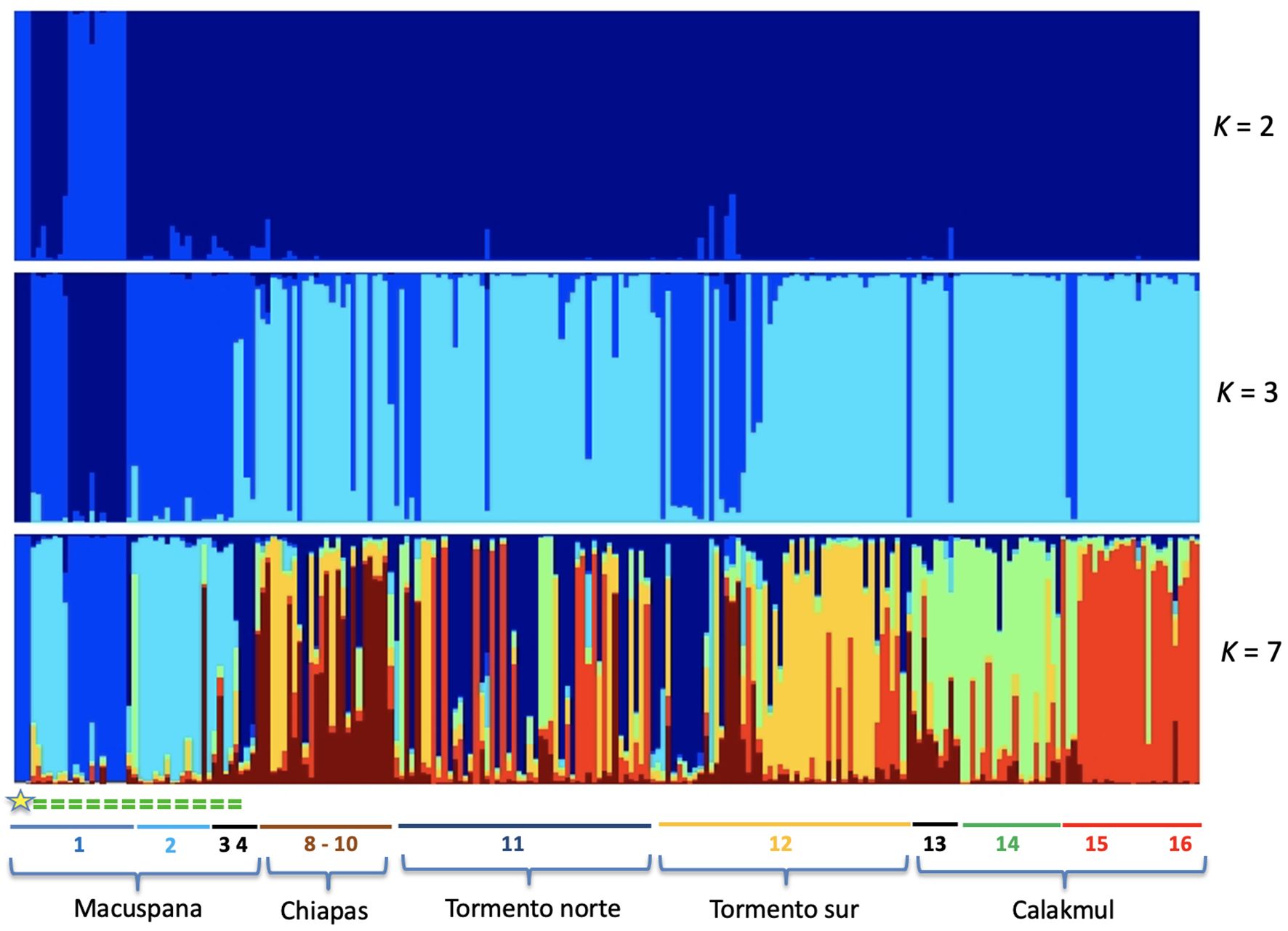
Figure 2. Genetic differentiation based on microsatellite loci for the 222 sampled individuals across 16 sampling localities (numbers 1-16; see Supplementary Table S1), including both hybrid and non-admixed Alouatta pigra individuals. Structure plots for K=2, K=3 and K=7 (K statistics shown in Supplementary Figure S2). The five genetic clusters identified are indicated as Macuspana (MAC), Chiapas (CHI), Tormento norte (TON), Tormento sur (TOS) and Calakmul (CAL). The yellow star indicates three A. palliata individuals identified (see Results) and the green dashed line depicts where hybrid individuals are distributed along the contact zone in Macuspana.
Divergence time estimation
To evaluate the overall phylogenetic relationships of our samples including outgroups, we used four species (sequences obtained from GenBank): Chiropotes albinasus (KC757393), Aotus azarai (KC757385), Ateles belzebuth (KC757386), Alouatta seniculus (HQ644333), and also included five sequences for Alouatta pigra (AY065884 and AY065885), A. palliata palliata (AY065880) and A. palliata mexicana (MW452897 and MW452920). We estimated the best fitting model of sequence evolution and performed ML phylogenetic analysis, both with PhyML 3.0 (Guindon et al., 2010), using a Nearest-Neighbor Interchange and Subtree-Prune and Regraft moves (NNI+SPR) for branch length and topology optimization. Clade support was assessed with 1,000 non-parametric bootstrap replicates and a majority-rule consensus tree was obtained with FigTree v.1.4 (Rambaut, 2018).
To have an overall approximation of the diverging times of the hybrids within both A. pigra and A. palliata clades, we used a dataset with unique haplotypes obtained in this study to estimate the time to the most recent common ancestor, TMRCA with Beast 1.10.4 (Suchard et al., 2018). We complemented this data set with sequences retrieved form GenBank of other species of Alouatta as well as other Neotropical primates as outgroup: Chiropotes albinasus, Ateles belzebuth, Alouatta seniculus, A. pigra, A. p. palliata, and A. p. mexicana. We applied four calibration points: the TMRCA for C. albinasus, At. belzebuth and A. seniculus at 19.7 (95%=17-23), 14.3 (12-17) and 6.8 million years ago (My), respectively; and the A. pigra spilt at 3.0 My (Schrago et al., 2013; Cortés-Ortiz et al., 2003; Di Fiore et al., 2015). Final estimation included the GTR+I model of evolution across all gene and codon positions, 100,000,000 generations sampled every 1000th and 20% of initial generations discarded as burn-in. Convergence and stationarity were visualized with Tracer v.1.6 (http://tree.bio.ed.ac.uk/software/tracer).
Black howler monkey mitochondrial diversity and differentiation
To provide an overall description of the mitochondrial diversity for the hybrid entities (A. palliata- and A. pigra-hybrids), as well as that of A. pigra individuals, we estimated different indices such as, the number of segregating sites (S) and haplotype (h) and nucleotide diversity (π) with DnaSP v.6 (Rozas et al., 2017); these were calculated based on the genetic clusters previously identified for the non-admixed A. pigra individuals (MNA, CHI, TON, TOS, CAL; see Results). We also estimated Tajima’s D and Fu’s FS (Fu, 1997; Tajima, 1989) neutrality tests. To infer genealogical relationships among cyt-b sequences, we built a haplotype median joining network with PopArt v.1.7 (Leigh and Bryant, 2015) and epsilon=0.
Black howler monkey nuclear genetic structure and differentiation
Microsatellites genotypes were screened for null alleles in PopGenReport v.2.0 (Adamack and Gruber, 2014); three loci showed evidence of null alleles but not consistently across individuals from the different sampling localities, thus all loci were used in further analyses. Also, loci were checked for possible departures from Hardy-Weinberg equilibrium in pegas v.0.12 (Paradis, 2010), and linkage disequilibrium was measured with a modified index of association (rbarD) (Agapow and Burt, 2001) in poppr v.2.8.3 (Kamvar et al., 2014).
To identify natural genetic clusters of A. pigra we assess genetic structuring across the study region with the online application StructureSelector (Li and Liu, 2018), a web-based software that calculates four new supervised estimators (medmedk, medmeak, maxmedk, maxmeak; Puechmaille, 2016) that can be more accurate for selecting the number of clusters in unevenly sampled datasets. For model selection we used Clumpak (Cluster Markov Packager Across K), which automates the postprocessing of results of model-based population structure analyses (Kopelman et al., 2015), testing K=1-10. Clumpak identifies sets of highly similar runs and separates distinct groups of runs that represent distinct modes in the space of possible solutions. It then identifies an optimal alignment of inferred clusters across different values of K, simplifying the comparison of clustering results across different K values.
Genetic diversity and genetic differentiation were assessed considering the hybrid and the non-admixed individuals (from the contact zone in Macuspana) and the five genetic clusters previously identified (see Results). Genetic diversity was estimated as the average number of alleles (Na), number of private alleles (Pa), and observed (Ho) and unbiased expected heterozygosity (He) in Arlequin v3.5.2.2 (Excoffier et al., 2005), while a rarefied allelic richness (Ra) was estimated with PopGenReport. Genetic differentiation between genetic clusters was calculated using pairwise RST estimated in Arlequin. We also used a Discriminant Analysis of Principal Components (DAPC) (Jombart et al., 2010), a multivariate approach that reflects the genetic variation differences between groups while minimizing variation within them, with adegenet in R v.2.1.5 (R Core Team, 2021). We also examined the distribution of the genetic variance among genetic clusters and among sampling localities within clusters, using a molecular analysis of variance (AMOVA) and pairwise RST in Arlequin. Finally, we tested for isolation by distance (IBD hypothesis) with Mantel tests; genetic distances between sampling localities were based on the estimation of D-Jost (Jost, 2008) and Edwards (Edwards, 1971) distances with mmod v.1.3.3 (Winter, 2012), while geographic distances (Euclidian) were calculated with Raster v.3.3 (Hijmans, 2020). Mantel tests were run with vegan v.2.5 (Oksanen et al., 2019) and IBD correlation plots were built with MASS v.7.3 (Venables and Ripley, 2002).
Results
Of the 223 samples we collected on the field, we obtained successful readings across all 12 microsatellite loci for 222 individuals. The samples information and genotype database for the A. pigra and hybrid individuals is available in Supplementary Table S1. Likewise, we obtained a final 587 bp cyt-b fragment alignment for 107 individuals including hybrids and non-admixed A. pigra. On the basis of the combined information of field-assigned phenotype, cyt-b sequences, diagnostic microsatellites alleles, and phylogenetic and structure results, of these 222 individuals, three were identified as A. palliata (which were not included in the genetic analyses) and 33 as hybrids, thus we analyzed a total of 186 non-admixed A. pigra samples (Supplementary Table S1).
Mitochondrial DNA diversity and divergence
Mitochondrial diversity showed high to moderate values, which differed between hybrids with A. pigra and A. palliata mitochondria: haplotype and nucleotide diversity was higher in the former than the latter (Table 1). Considering A. pigra exclusively, overall diversity values were lower than the hybrids; while per genetic cluster, the highest diversity was found in Calakmul. Tajima’s D and Fu’s Fs values were not significant, with the exception of Tormento sur, Calakmul and hybrids with A. pigra mitochondria.
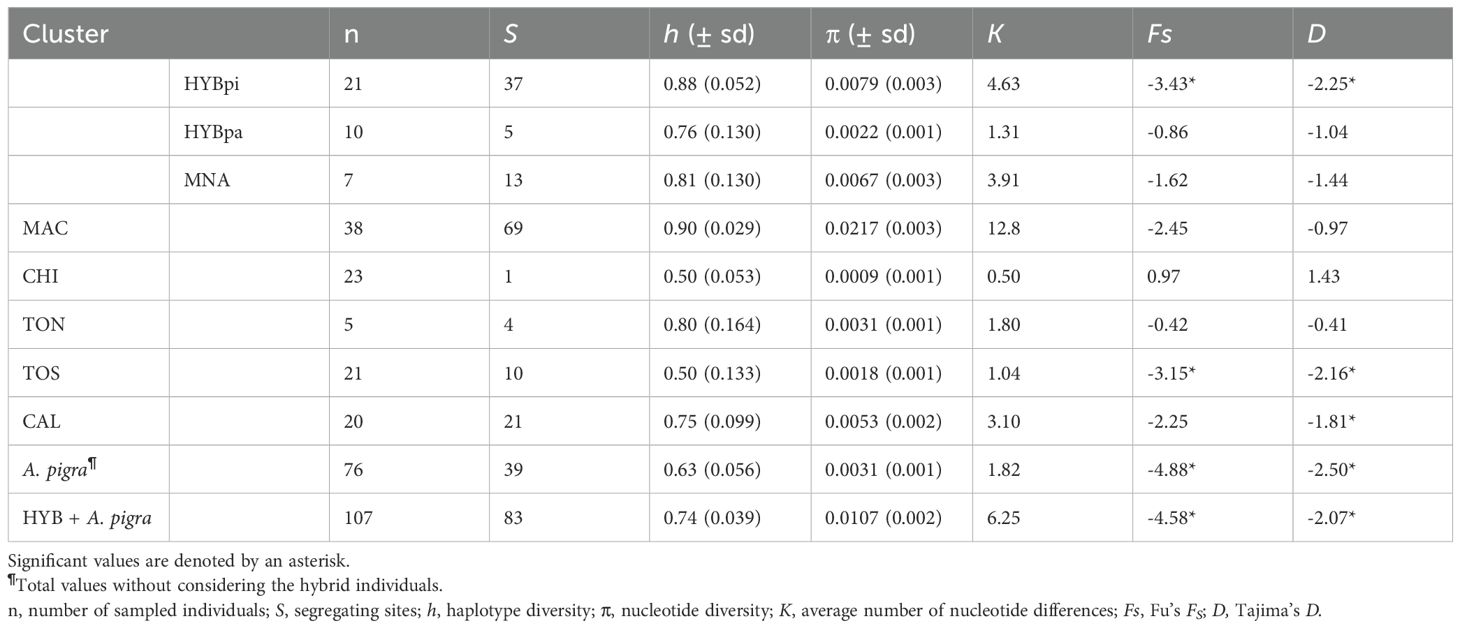
Table 1. Mitochondrial diversity values estimated for 76 Alouatta pigra and 31 hybrid individuals from five genetic clusters (MAC, CHI, TON, TOS, CAL; Structure results), including the A. pigra-hybrid (HYBpi) and the A. palliata-hybrid individuals (HYBpa), as well as the non-admixed individuals (MNA) from the contact zone, based on 587 bp cytochrome b sequences.
Phylogenetic maximum-likelihood inference results showed two major clades with high posterior support values (Supplementary Figure S1b), one comprising only hybrid individuals (10) grouping with A. palliata, and the other including all non-admixed A. pigra samples and the other 23 hybrid individuals. We obtained 29 unique haplotypes (13 from A. pigra, 5 from hybrids that had A. palliata mitochondria, 9 from hybrids with A. pigra mitochondria, and 2 shared between non-admixed and hybrid individuals (accession numbers PQ815818-PQ815846; Supplementary Table S1). The most common haplotype (hap1) was recovered in 50 individuals distributed in all the sampled localities, while the next most abundant (hap16) encompassed 20 individuals from all localities except from Calakmul. The highest number of different haplotypes was found in Macuspana, associated with the localities where hybrids are distributed, while the lowest number was in Chiapas (Figure 1B). Results from the median-joining network showed the haplotypes from the hybrids with A. palliata mitochondria clearly separated from the rest of the network. Also, the two most abundant and widespread haplotypes are shown, from which multiple unique haplotypes arise (Figure 1C); these most derived haplotypes are from Calakmul and several from A. pigra-hybrids, indicating recent divergence. Neither the phylogenetic tree nor the haplotype distribution showed a clear geographic pattern. The divergence evaluation showed the highest A. pigra diversification in Mexico near 1.4 My (95% HPD: 1.0-2.5), and an ample range of recent diverging times of the hybrids within both A. pigra and A. palliata clades (from ca. 850,000 years onwards) (Figure 3).
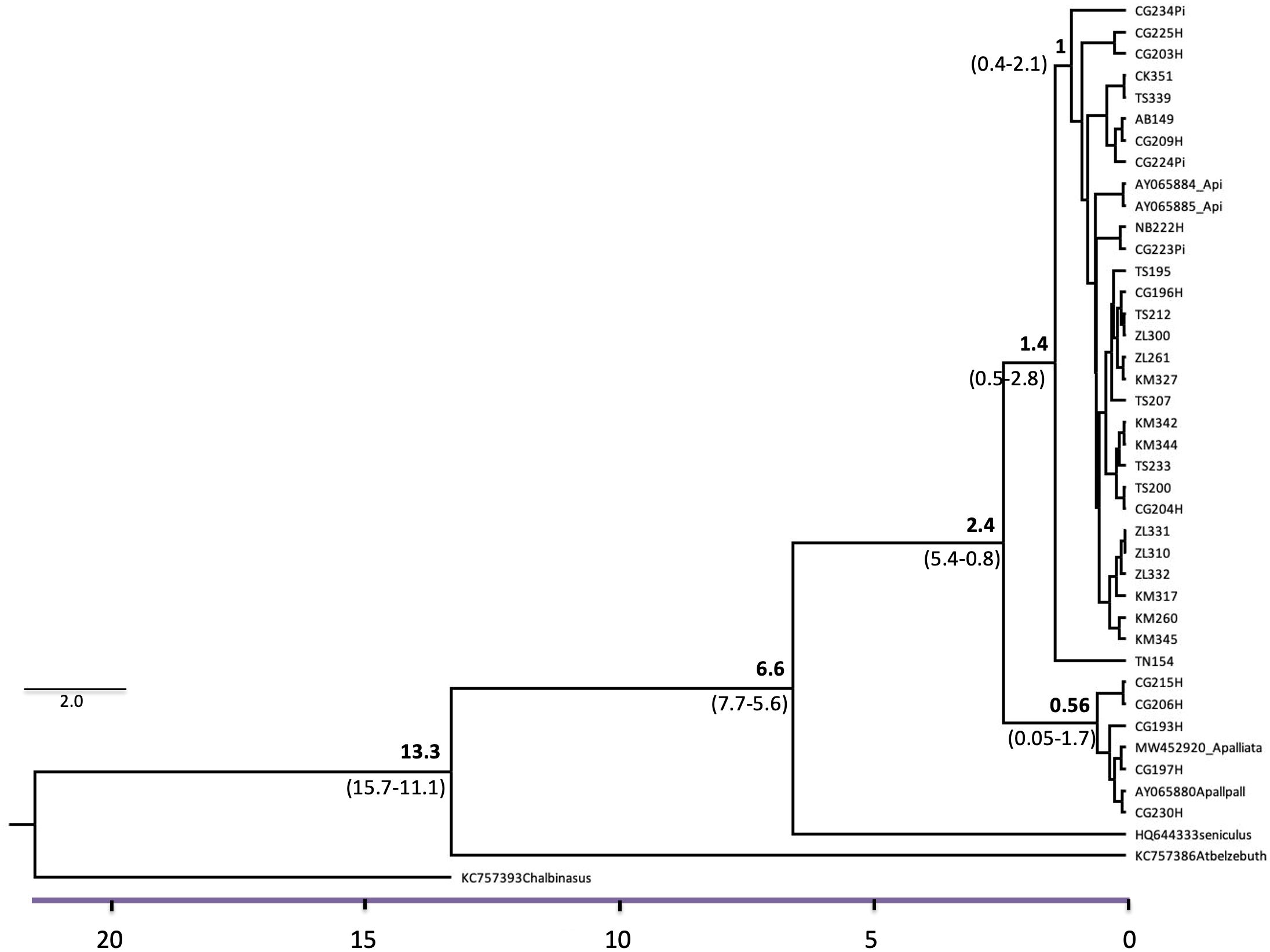
Figure 3. Divergence-time estimation (time-scale in millions of years; My), based on unique cytochrome b haplotypes of A. pigra and hybrid individuals. Chiropotes albinasus, Aotus azarai, Ateles belzebuth, and Alouatta seniculus were used as outgroups and for calibration points. Numbers above branches are mean divergence time in My with 95% HPD in parentheses).
Genetic diversity and structure (microsatellites) patterns
We amplified 12 microsatellites loci, most of which showed deviations from HWE with heterozygote deficiency. Low linkage disequilibrium (Ia=0.719, pIa=0.001; rbarD=0.066, prD=0.001) was detected between 4 loci (Supplementary Figure S3). The structuring analysis for A. pigra identified K=6 genetic clusters across the distribution, although with high admixture among them (Figure 4), namely Macuspana, Chiapas, Tormento norte, Tormento sur and Calakmul (the latter with sampling localities 15 and 16 differentiated).
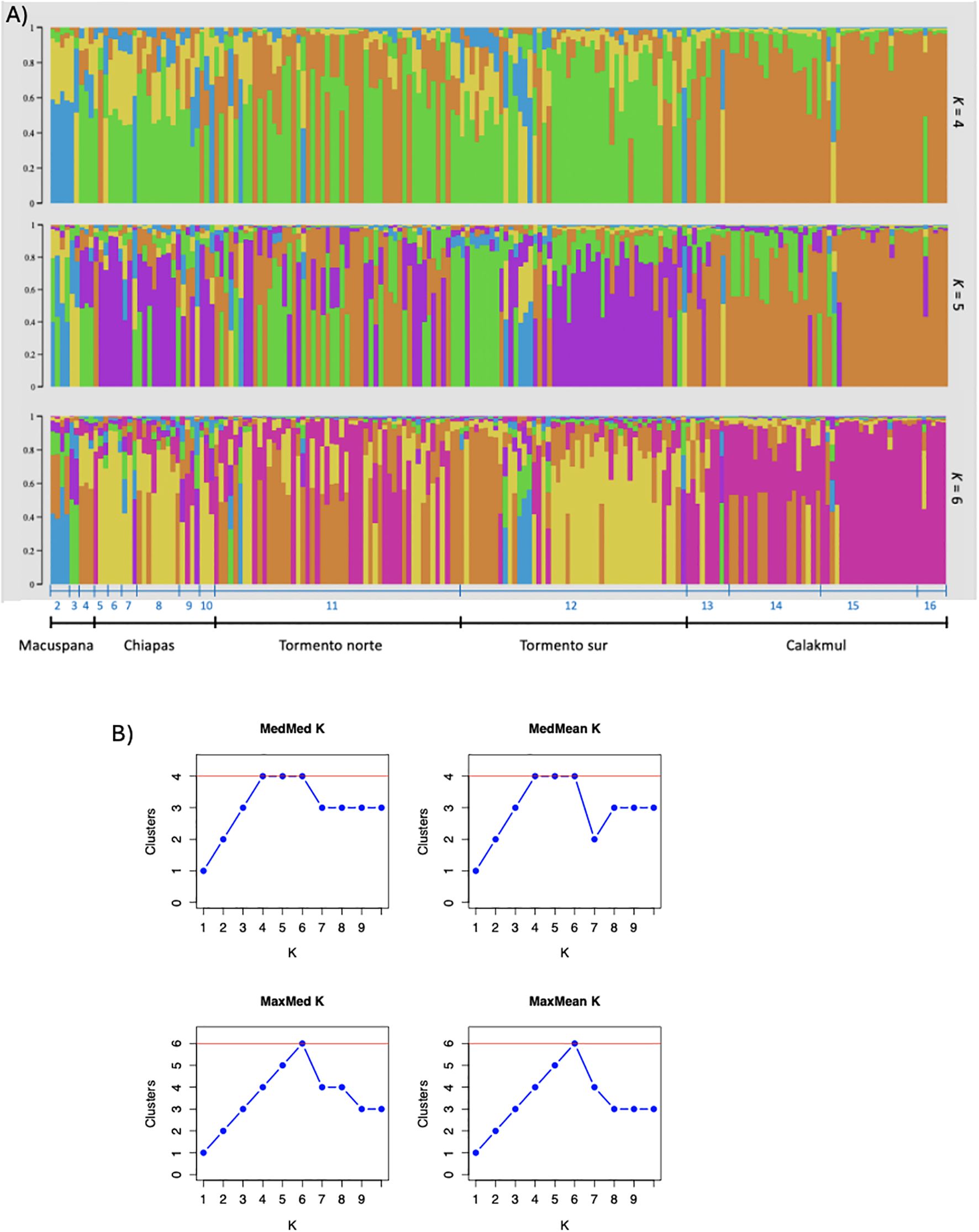
Figure 4. (A) Genetic differentiation of Alouatta pigra individuals based on microsatellite loci. Sampling localities (numbers 2-16; see Supplementary Table S1) are indicated. Structure plots for K=4, K=5 and K=6. The five genetic clusters identified are indicated as Macuspana (MAC), Chiapas (CHI), Tormento norte (TON), Tormento sur (TOS) and Calakmul (CAL). (B) Number of clusters obtained with Clumpak (Kopelman et al., 2015) with model selection based on four estimators (medmedk, medmeak, maxmedk, maxmeak; Puechmaille, 2016).
Overall genetic diversity for both hybrid individuals and for A. pigra showed high number of alleles, rarefied allelic richness, observed heterozygosity and unbiased expected heterozygosity (Table 2). Values among genetic clusters were very similar, with the highest observed heterozygosity in Calakmul and Tormento norte. Significant FIS was observed for Tormento norte and Tormento sur (Table 2). Paired genetic distance results showed significant values, indicating limited gene flow between the hybrids and all A. pigra genetic clusters, being highest with Calakmul (RST =0.378); also Calakmul showed significant distances with Tormento norte and Tormento sur (Supplementary Table S3). The DAPC analysis corroborated a clear differentiation of the hybrids from the rest of the genetic clusters (Figure 5).
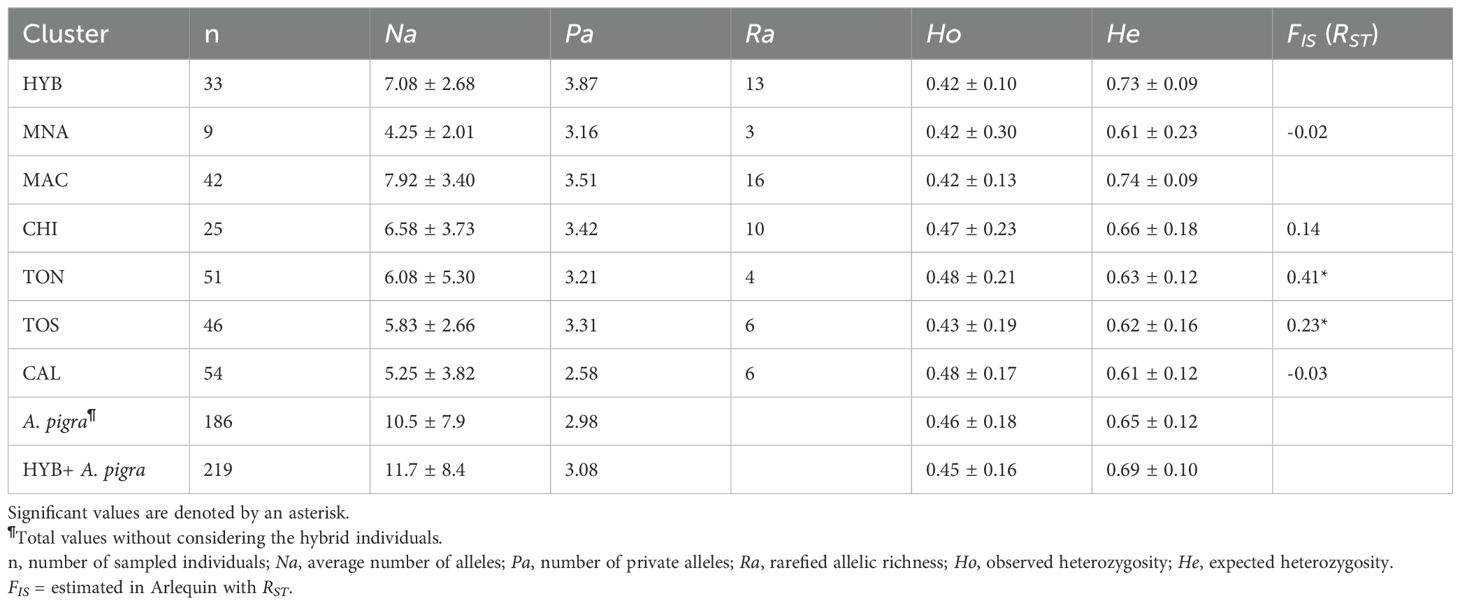
Table 2. Genetic diversity values estimated for 186 Alouatta pigra and 33 hybrid individuals from five genetic clusters (MAC, CHI, TON, TOS, CAL; Structure results), including the hybrid individuals (HYB) and the non-admixed individuals (MNA) from the contact zone.
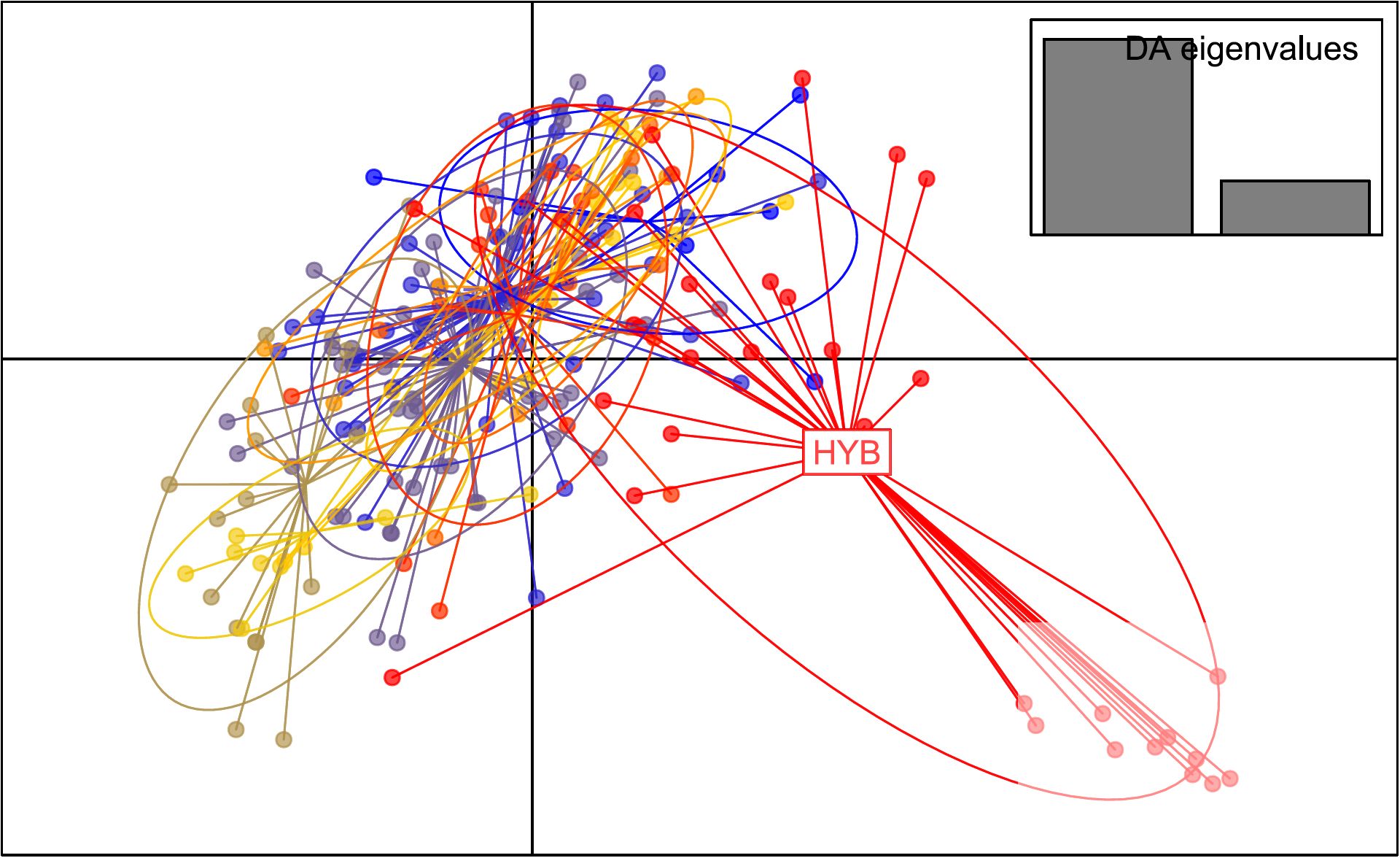
Figure 5. Discriminant Analysis of Principal Components (DAPC) based on microsatellite data including the hybrid (HYB) and the non-admixed Alouatta pigra sampled in Mexico. Colored dots indicate all the individuals sampled and analyzed in the study.
Results considering only the A. pigra data showed that the distribution of genetic variance (AMOVA) was significant (p<0.001) among populations within genetic clusters (Fsc=0.129) and within populations (Fst=0.078) (Supplementary Table S4). The Mantel test was significant (p<0.001) for both genetic distance measures (r=0.285 with D-Jost; r=0.495 with Edwards) (Supplementary Figure S4).
Discussion
Mitochondrial divergence and diversity patterns of Alouatta pigra and hybrid populations
The A. pigra and A. palliata clades each encompassed hybrid individuals with their corresponding species mitochondria, which is concordant with the historical diversification of these two species. The split between A. palliata and A. pigra has been dated between ~2.8 and ~3 My (Cortés-Ortiz et al., 2003; Doyle et al., 2021), while diversification of A. palliata in Mexico happened ~2.4 My (Solórzano-García et al., 2021). According to our results, the highest diversification for A. pigra in Mexico occurred later (1.4 My), while identifying another, more recent, diversification event including A. palliata and hybrid individuals (ca. 0.56 My). This would support different diversification events along these primates’ evolutionary history (Doyle et al., 2021). Such patterns indicate a recent colonization and population expansion of these howler monkeys through Central America northwards into Mexico, also coinciding with the hypothesis that A. pigra likely arrived first at Central America and diversified, with subsequent range reduction following the invasion of A. palliata (Ellsworth and Hoelzer, 2006). This is also in agreement with the biogeographic history hypothesis for Mesoamerican primates, namely that within species diversification was promoted by a series of demographic fluctuations in response to Pleistocene and Holocene climatic variations (Lynch-Alfaro et al., 2015; Solórzano-García et al., 2021).
On the other hand, divergence of hybrid individuals, although dated more recently in comparison with each parental species, as we predicted, is not a contemporary process in the evolutionary history of these sister species. The evolutionary outcomes when hybridization is old would be a hybrid zone that is shaped by multiple episodes of primary divergence and secondary contact, reflecting both recent and ancient hybridization (Abbott et al., 2013; Sardell and Uy, 2016; Wang et al., 2020). Indeed, our findings suggest that the two species came into contact at the zone of sympatry at least some eight hundred thousand years ago. Periods of isolation and secondary contact and reproductive isolation, jointly with particular demographic, behavioral and ecological characteristics of these primates have kept this as a narrow hybrid zone (Ho et al., 2014; Baiz et al., 2019; Cortés-Ortiz et al., 2019).
Mitochondrial haplotype and nucleotide diversity was higher in hybrids in comparison with the non-admixed individuals. In addition, the highest number of different haplotypes was found in Macuspana in the localities where hybrids are distributed. Notably, hybrids with A. pigra mitochondria exhibited both higher diversity values and number of unique haplotypes than those with A. palliata mitochondria. Cortés-Ortiz et al. (2019) surveyed populations from the hybrid zone and found that some individuals comprising nuclear (microsatellites) A. pigra alleles possessed A. palliata mitochondrial haplotypes and vice versa, implying that females of both species have contributed to hybrid offspring, but with a bias of more A. pigra females successfully producing hybrid offspring. Our diversity findings concur with hybrids resulting from multiple generations of backcrossing as well as higher successful reproduction of females with A. pigra maternal lineages than those with A. palliata maternal lineages. Such patterns can be due to a higher philopatry in A. pigra females (Ho et al., 2014; Van Belle and Di Fiore, 2022), or to mechanisms of genetic compatibility.
Regarding the black howler populations, the overall cyt-b diversity was lower than values observed in other howler monkey species, including A. palliata (Nascimento et al., 2008; Ruiz-García et al., 2017; Solórzano-García et al., 2021), as well as to that reported for other Atelids, like Ateles, Brachyteles and Lagothrix (Chaves et al., 2011; Ruiz-García et al., 2019). It is important to highlight that Chiapas had the lowest haplotype and nucleotide diversity and number of different haplotypes. This zone represents the southernmost margin of the species’ distribution, therefore A. pigra likely has reduced genetic diversity at this range edge than closer to the center; a pattern observed in many species of plants and animals (Ellsworth and Hoelzer, 2006; Lira-Noriega and Manthey, 2014).
Nuclear genetic diversity and structure patterns
Maintaining genetic diversity is key to enhance species adaptive potential and prevent extinction (Junker et al., 2020; Oklander and Soto-Calderón, 2024). Notably, contrary to our prediction of reduced genetic variation due to habitat modification and fragmentation, overall A. pigra genetic (microsatellites) diversity exhibited higher values (Ho=0.460, He=0.650) in comparison with that found in A. palliata across its distribution in Mexico (Ho=0.330, He=0.450; Solórzano-García et al., 2021) and A. caraya from Argentina and Paraguay (He=0.420; Oklander et al., 2017), which respectively represent the northernmost and southernmost distributions of this genus, reflecting a latitudinal gradient of diversity (Ellsworth and Hoelzer, 2006).
Although A. pigra genetic diversity was fairly similar among genetic clusters, the highest values were found in Calakmul, which notably represents the most conserved and continuous habitat in our study. Van Belle et al. (2012) also documented high genetic diversity (Ho=0.588) in A. pigra populations within a protected area (Palenque National Park). Regarding the genetic diversity of hybrids (Ho=0.420, He=0.730), we expected it to be higher in comparison with the parental species, which was true when compared with A. palliata but not with A. pigra. This could be the result of a series of factors, including the narrow area of the hybrid zone that only allows certain parental genotypes from adjacent populations to be admixed, along with the history of colonization of Central America and time of contact between these two species. Interestingly, significant genetic distance was observed between hybrids and all non-admixed A. pigra genetic clusters, highlighting the particular genetic configuration of these admixed populations. Furthermore, our results showed that the majority of the sampled individuals within the hybrid zone were admixed, although 27% of the genotyped individuals from this area were non-admixed, supporting the notion of ongoing influx of parental forms into the area of sympatry (Cortés-Ortiz et al., 2019).
The observed pattern of genetic differentiation based on microsatellites loci supported isolation by distance, indicating that gene flow is mainly limited by the geographic distance among localities, with closer ones sharing more genetic features that further ones. Furthermore, Calakmul conformed the most differentiated genetic cluster which not only represents the furthest localities across our sampling, but also comprises the largest continuous conserved habitat of A. pigra’s distribution in Mexico. Notably, a different genetic configuration can be observed between Tormento sur and Tormento norte, which are adjacent locations of well-preserved habitat only divided by a heavy traffic two-lined road; these results indicate that this barrier significantly hinders individual dispersal, leading to increased genetic structure between them. Thus, habitat quality, arboreal vegetation and environment modification is tightly associated with the A. pigra genetic structure gradient.
Genetics, conservation and viability
Primates perform critical ecological roles as seed dispersers and ecosystem engineers, contributing among others to maintaining the structure and regeneration of tropical forests (Melin et al., 2020). Yet, developing strategies based on genetic evidence for conserving primates and to ensure the survival of viable primate populations in the long term are still urgently needed (Junker et al., 2020). Deforestation and habitat disturbance are among the major threats of most primate species living in isolated forest fragments (Serio-Silva et al., 2015; Estrada et al., 2017), hence determining their historical and contemporary genetic patterns, as in the present study, is of prime value to assess the viability and conservation of populations.
Our study provides novel information regarding the genetic diversity and structuring of A. pigra in Mexico. It shows that most of the evaluated A. pigra maintain genetic diversity and some connectivity. Nonetheless, we also found genetic differentiation and limited gene flow among them, as well as potential inbreeding signals, all of which render A. pigra prone to drift effects and increased extinction risk. Modern human activities, habitat modification and fragmentation have drastically contributed to the current isolated distribution of both the non-admixed A. pigra and of the hybrid individuals. In accordance, some actions that should be implemented to prevent this include, but are not limited to, legal protection and landscape management throughout this primate’s range, particularly in regions outside natural protected areas. Preventing forest loss and promoting forest regeneration in the human-modified landscapes have proven effective; Alcocer-Rodríguez et al. (2021) found that forest recovery over a relatively short period could promote the recovery of the A. palliata. Landscape genetics should be applied to evaluate and understand the impact of habitat loss and fragmentation in these primates. Finally, ex-situ facilities and care centers have become crucial for reproduction programs and for the rehabilitation of primates from trafficking or environmental impacts (Estrada and Garber, 2022; Pozo-Montuy et al., 2024); confident genetic information like that provided by our study will aid to adequately perform these activities.
Importantly, hybridization has been recognized as a common phenomenon across all major primate lineages (Arnold, 2009; Cortés-Ortiz, 2017), estimating an overall prevalence of natural hybridization in more than 10% of all primate species (Zinner et al., 2011). Yet nearly every hybrid system is different and general conservation rules are not effective (Gompert and Buerkle, 2016), thus we need redefining the role of admixture in species conservation, emphasizing that protection measures are required for taxa that have experienced gene flow and introgression over the course of their evolutionary histories (Pacheco-Sierra et al., 2018; vonHoldt et al., 2018). Our findings show that hybrid individuals exhibit high genetic variability and differentiation compared to non-admixed A. pigra, suggesting that they should be considered as a significant evolutionary unit included in conservation measures (Jackiw et al., 2015: Baiz et al., 2019; Suárez-Atilano et al., 2024). Future studies with these and other primate species should consider genomic advances, which are a promising avenue for evaluating and incorporating adaptive variation and functional genetics in the conservation of wildlife (Oklander and Soto-Calderón, 2024), including hybrid entities (Suárez-Atilano et al., 2024). Genomic tools are key for the assessment of functional genetic variations associated with, for instance, diseases, environmental changes, and ex-situ conditions.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: NCBI GenBank, accession PQ815818–PQ815846. The sample data and microsatellite genotype presented in the study can be found in the article/Supplementary Material.
Ethics statement
Ethical approval was not required for the study because our fieldwork was based exclusively on the collecting of feces, with no contact whatsoever with animal individuals. Fieldwork was performed under the scientific sampling permit issued to EVD (Nº SGPA/DGVS/05372/19 and SGPA/DGVS/03783/21).
Author contributions
EV-D: Conceptualization, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing. BS-G: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Writing – review & editing, Supervision. MR-V: Conceptualization, Investigation, Methodology, Writing – review & editing. DZ-M: Data curation, Formal Analysis, Methodology, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Funding support was received from Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT-UNAM IN202819) to EV-D.
Acknowledgments
We thank Rubén Mateo and Pablo Gutiérrez for their help and support during field work, Marco Solano de la Cruz and Nancy Gálvez-Reyes for molecular laboratory assistance, and Miguel Baltazar for computational advice. Authorities from Los Tuxtlas field station, Centro de Investigación y Transferencia de Tecnología Forestal ‘El Tormento’, and Natural Protected Areas kindly granted permission for fieldwork and sample collection. Our gratitude to the people from the different localities where sampling was done, for allowing us to work on their ejidos and private properties. MR-V was supported by a postgraduate scholarship from Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCyT).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1551979/full#supplementary-material
References
Abbott R. D., Albach D., Ansell S., Arntzen J. W., Baird S. J. E., Bierne N., et al. (2013). Hybridization and speciation. J. Evol. Biol. 26, 229–246. doi: 10.1111/j.1420-9101.2012.02599.x
Adamack A. T. and Gruber B. (2014). POPGEN REPORT: simplifying basic population genetic analyses in R. Methods Ecol. Evol. 5, 384–387. doi: 10.1111/mee3.2014.5.issue-4
Agapow P. M. and Burt A. (2001). Indices of multilocus linkage disequilibrium. Mol. Ecol. Notes 1, 101–102. doi: 10.1046/j.1471-8278.2000.00014.x
Alcocer-Rodríguez M., Arroyo-Rodríguez V., Galán-Acedo C., Cristóbal-Azkarate J., Asensio N., Rito K. F., et al. (2021). Evaluating extinction debt in fragmented forests: the rapid recovery of a critically endangered primate. Anim. Conserv. 24, 432–444. doi: 10.1111/acv.12648
Allendorf F. W., Leary R. F., Spruell P., and Wenburg J. K. (2001). The problems with hybrids: setting conservation guidelines. Trends Ecol. Evol. 16, 613–622. doi: 10.1016/S0169-5347(01)02290-X
Allendorf F. W., Luikart G., and Aitken S. N. (2013). Conservation and the Genetics of Populations (Malden, MA: Wiley-Blackwell).
Améndola M. (2009). Estudio de la variabilidad genética en poblaciones de Alouatta pigra del estado de Campeche: implicaciones para la conservación (Xalapa, Ver: Instituto de Ecología, A.C).
Arnold M. L. (2009). Reticulate evolution and humans: Origins and ecology (Oxford: Oxford University Press).
Arroyo-Rodríguez V. and Dias P. A. D. (2010). Effects of habitat fragmentation and disturbance on howler monkeys: a review. Am. J. Primatol. 72, 1–16. doi: 10.1002/ajp.20753
Baiz M. D. and Cortés-Ortiz L. (2015). Intragroup genetic relatedness in two howler monkey species (Alouatta pigra and A. palliata): Implications for understanding social systems and dispersal. Am. J. Primatol. 77, 1333–1345.
Baiz M. D., Tucker P. K., and Cortés-Ortiz L. (2019). Multiple forms of selection shape reproductive isolation in a primate hybrid zone. Mol. Ecol. 28, 1056–1069. doi: 10.1111/mec.14966
Bicca-Marques J. C. (2003). “How do howler monkeys cope with habitat fragmentation?”, in Primates in Fragments: Ecology and Conservation. Ed. Marsh L. K. (Kluwer Academics/Plenum Publishers, New York), 283–303.
Brennan A. C., Barker D., Hiscock S. J., and Abbott R. J. (2012). Molecular genetic and quantitative trait divergence associated with recent homoploid hybrid speciation: a study of Senecio squalidus (Asteraceae). Heredity 108, 87–95. doi: 10.1038/hdy.2011.46
Ceballos G., Blanco S., González C., and Martínez-Meyer E. (2006). “Alouatta pigra (Mono aullador negro, saraguato negro) delimitada, con base al Atlas Mastozoológico de México,” in Distribución potencial. Catálogo de metadatos geográficos (Conabio, Mexico).
Chaves P. B., Alvarenga C. S., Possamai C., Días L. G., Boubli J. P., Strier K. B., et al. (2011). Genetic diversity and population history of a critically endangered primate, the Northern Muriqui (Brachyteles hypoxanthus). PloS One 6, e20722. doi: 10.1371/journal.pone.0020722
Coates D. J., Byrne M., and Moritz C. (2018). Genetic diversity and conservation units: Dealing with the species-population continuum in the age of genomics. Front. Ecol. Evol. 6, 1–13. doi: 10.3389/fevo.2018.00165
Cortés-Ortiz L. (2017). “Hybridization and hybrid zones”, in The International Encyclopedia of Primatology. Ed. Fuentes A. (John Wiley & Sons, Hoboken, NJ), 1–5.
Cortés-Ortiz L., Bermingham E., Rico C., Rodríguez-Luna E., Sampaio I., and Ruiz-Garcia M. (2003). Molecular systematics and biogeography of the Neotropical monkey genus, Alouatta. Mol. Phylogenet. Evol. 26, 64–81. doi: 10.1016/S1055-7903(02)00308-1
Cortés-Ortiz L., Mondragón E., and Cabotage J. (2010). Isolation and characterization of microsatellite loci for the study of Mexican howler monkeys, their natural hybrids, and other neotropical primates. Conserv. Genet. Res. 2, 21–26. doi: 10.1007/s12686-009-9124-6
Cortés-Ortiz L., Nidiffer M. D., Hermida-Lagunes J., García-Orduña F., Ragel-Negrín A., Kitchen D. M., et al. (2019). Reduced introgression of sex chromosome markers in the Mexican howler monkey (Alouatta palliata × A. pigra) hybrid zone. Int. J. Primatol. 40, 114–131. doi: 10.1007/s10764-018-0056-4
Cortés-Ortíz L., Rosales-Meda M., Marsh L. K., and Mittermeier R. A. (2020). Alouatta pigra (The IUCN Red List of Threatened Species), e.T914A17926000. Available at: https://dx.doi.org/10.2305/IUCN.UK.2020-3.RLTS.T914A17926000.en.
Cortés-Ortiz L., Rylands A. B., and Mittermeier R. (2015). “The taxonomy of howler monkeys: Integrating old and new knowledge from morphological and genetic studies,” in Howler Monkeys. Adaptative Radiation, Systematics and Morphology. Eds. Kowalewski M. M., Garber P. A., Cortes-Ortiz L., Urbani B., and Youlatos D. (Springer Science, New York), 55–84. doi: 10.1007/978-1-4939-1957-4_3.
Cristobal-Azkarate J. and Dunn J. C. (2013). “Lessons from Los Tuxtlas: 30 years of research into primates in fragments”, in Primates in Fragments: Complexity and Resilience. Eds. Marsh L. and Champan C. A. (Springer Science, New York), 75–88.
Di Fiore A., Chaves P. B., Cornejo F. M., Schmitt C. A., Shanee S., Cortés-Ortiz L., et al. (2015). The rise and fall of a genus: Complete mtDNA genomes shed light on the phylogenetic position of yellow-tailed woolly monkeys, Lagothrix flavicauda, and on the evolutionary history of the family Atelidae (Primates: Platyrrhini). Mol. Phylogenet. Evol. 82, 495–510. doi: 10.1016/j.ympev.2014.03.028
Doyle E. D., Prates I., Sampaio I., Koiffmann C., Silva W. A. Jr., Carnaval A. C., et al. (2021). Molecular phylogenetic inference of the howler monkey radiation (Primates: Alouatta). Primates 62, 177–188. doi: 10.1007/s10329-020-00854-x
Edmands S. (2007). Between a rock and a hard place: evaluating the relative risks of inbreeding and outbreeding for conservation and management. Mol. Ecol. 16, 463–475. doi: 10.1111/j.1365-294X.2006.03148.x
Edwards A. W. F. (1971). Distance between populations on the basis of gene frequencies. Biometrics 27, 873–881. doi: 10.2307/2528824
Ellsworth J. A. and Hoelzer G. A. (2006). “Genetic evidence on the historical biogeography of Central American howler monkeys”, in Primate Biogeography: Progress and Prospects. Eds. Lehman S. and Feagle J. (Springer Science, New York), 81–104.
Estrada A. and Garber P. A. (2022). Principal drivers and conservation solutions to the impending primate extinction crisis: introduction to the special issue. Int. J. Primatol. 43, 1–14. doi: 10.1007/s10764-022-00283-1
Estrada A., Garber P. A., Rylands A. B., Roos C., Fernandez-Duque E., Di Fiore A., et al. (2017). Impending extinction crisis of the world’s primates: Why primates matter. Sci. Adv. 3, e1600946. doi: 10.1126/sciadv.160094
Excoffier L., Laval G., and Schneider S. (2005). Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinf. 1, 47–50. doi: 10.1177/117693430500100003
Fu Y.-X. (1997). Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 147, 915–925. doi: 10.1093/genetics/147.2.915
García del Valle Y., Estrada A., Espinoza E., Lorenzo C., and Naranjo E. (2005). Genética de poblaciones de monos aulladores (Alouatta pigra) en hábitat continuo y fragmentado en la Selva Lacandona Vol. 2 (México: un estudio preliminar. Universidad y Ciencia), 55–60.
Gompert Z. and Buerkle C. A. (2016). What, if anything, are hybrids: enduring truths and challenges associated with population structure and gene flow. Evol. Appl. 9, 909–923. doi: 10.1111/eva.12380
Gompert Z., Mandeville E. G., and Buerkle C. A. (2017). Analysis of population genomic data from hybrid zones. Annu. Rev. Ecol. Syst. 48, 207–229. doi: 10.1146/annurev-ecolsys-110316-022652
Guindon S., Dufayard J. F., Lefort V., Anisimova M., Hordijk W., and Gascuel O. (2010). New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst. Biol. 59, 307–321. doi: 10.1093/sysbio/syq010
Hijmans R. J. (2020). raster: geographic data analysis and modeling. R package version 3.3-13. Available online at: https://cran.r-project.org/package=raster%0A.
Ho L., Cortés-Ortiz L., Dias P. A. D., Canales-Espinosa D., Kitchen D. M., and Bergman T. J. (2014). Effect of ancestry on behavioral variation in two species of howler monkeys (Alouatta pigra and A. palliata) and their hybrids. Am. J. Primatol. 76, 855–867. doi: 10.1002/ajp.22273
Jackiw R. N., Mandil G., and Hager H. A. (2015). A framework to guide the conservation of species hybrids based on ethical and ecological considerations. Conserv. Biol. 29, 1040–1051. doi: 10.1111/cobi.12526
James R. A., Leberg P. L., Quattro J. M., and Vrijenhoek R. C. (1997). Genetic diversity in black howler monkeys (Alouatta pigra) from Belize. Am. J. Phys. Anthropol. 102, 329–336. doi: 10.1002/(SICI)1096-8644(199703)102:3<329::AID-AJPA3>3.0.CO;2-Q
Jombart T., Devillard S., and Balloux F. (2010). Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 11, 94. doi: 10.1186/1471-2156-11-94
Jost L. (2008). GST and its relatives do not measure differentiation. Mol. Ecol. 17, 4015–4026. doi: 10.1111/j.1365-294X.2008.03887.x
Junker J., Petrovan S. O., Arroyo-Rodríguez V., Raesh Boonratana R., Byler D., Chapman C. A., et al. (2020). A severe lack of evidence limits effective conservation of the World’s primates. BioScience 70, 794–803. doi: 10.1093/biosci/biaa082
Kamvar Z. N., Tabima J. F., and Grünwald N. J. (2014). Poppr: an R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2, e281. doi: 10.7717/peerj.281
Kopelman N. M., Mayzel J., Jakobsson M., Rosenberg N. A., and Mayrose I. (2015). Clumpak: a program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 15, 1179–1191. doi: 10.1111/1755-0998.12387
Lamichhaney S., Han F., Webster M. T., Andersson L., Grant B. R., and Grant P. R. (2017). Rapid hybrid speciation in Darwin’s finches. Science 359, 224–228. doi: 10.1126/science.aao4593
Leigh J. W. and Bryant D. (2015). POPART: full-feature software for haplotype network construction. Methods Ecol. Evol. 6, 110–1116. doi: 10.1111/mee3.2015.6.issue-9
Li Y. L. and Liu J. X. (2018). StructureSelector: A web based software to select and visualize the optimal number of clusters using multiple methods. Mol. Ecol. Resour. 18, 176–177. doi: 10.1111/men.2018.18.issue-1
Lira-Noriega A. and Manthey J. D. (2014). Relationship of genetic diversity and niche centrality: A survey and analysis. Evolution 68, 1082–1093. doi: 10.1111/evo.2014.68.issue-4
Lynch-Alfaro J. W., Cortés-Ortiz L., Di Fiore A., and Boubli J. P. (2015). Comparative biogeography of Neotropical primates. Mol. Phylogen. Evol. 82, 518–529. doi: 10.1016/j.ympev.2014.09.027
Melin A. D., Hogan J. D., Campos F. A., Wikberg E., King-Bailey G., Webb S., et al. (2020). Primate life history, social dynamics, ecology, and conservation: Contributions from long-term research in Área de Conservación Guanacaste, Costa Rica. Biotropica 52, 1041–1064. doi: 10.1111/btp.12867
Miller M., Pfeiffer W., and Schwartz T. (2010). “Creating the CIPRES Science Gateway for inference of large phylogenetic trees”, in Proceedings of the Gateway Computing Environments Workshop, 1–8.
Mimura M., Yahara T., Faith D. P., Vázquez-Domínguez E., Colautti R. I., Araki H., et al. (2017). Understanding and monitoring the consequences of human impacts on intraspecific variation. Evol. Appl. 10, 121–139. doi: 10.1111/eva.2017.10.issue-2
Moraes A. M., Ruiz-Miranda C. R., Ribeiro M. C., Grativol A. D., Carvalho C. S., Dietz J. M., et al. (2017). Temporal genetic dynamics of reintroduced and translocated populations of the endangered golden lion tamarin (Leontopithecus rosalia). Conserv. Genet. 18, 995–1009. doi: 10.1007/s10592-017-0948-4
Nascimento F. F., Bonvicino C. R., De Oliveira M. M., Schneider M. P. C., and Seuánez H. N. (2008). Population genetic studies of Alouatta belzebul from the Amazonian and Atlantic forests. Am. J. Primatol. 70, 423–431. doi: 10.1002/ajp.20507
Oklander L. I., Marino M., Zunino G. E., and Corach D. (2004). Preservation and extraction of DNA from feces in howler monkeys (Alouatta caraya). Neotrop. Primates 12, 59–63. doi: 10.62015/np.2004.v12.571
Oklander L. I., Miño C. I., Fernández G., Caputo M., and Corach D. (2017). Genetic structure in the southernmost populations of black-and-gold howler monkeys (Alouatta caraya) and its conservation implications. PloS One 12, 1–20. doi: 10.1371/journal.pone.0185867
Oklander L. I. and Soto-Calderón I. D. (2024). Applications of primate genetics for conservation and management. Ann. Rev. Anthropol. 53, 371–395. doi: 10.1146/annurev-anthro-041422-114003
Oksanen J., Blanchet F. G., Friendly M., Kindt R., Legendre P., McGlinn D., et al. (2019). vegan: Community Ecology Package. R package version 2.5-6. Available online at: https://cran.r-project.org/web/packages/vegan/vegan.pdf.
Pacheco-Sierra G., Vázquez-Domínguez E., Pérez-Alquicira J., Suárez-Atilano M., and Domínguez-Laso J. (2018). Ancestral hybridization yields evolutionary distinct hybrids lineages and species boundaries in crocodiles, posing unique conservation conundrums. Front. Ecol. Evol. 6. doi: 10.3389/fevo.2018.00138
Paradis E. (2010). pegas: an R package for population genetics with an integrated-modular approach. Bioinformatics 26, 419–420. doi: 10.1093/bioinformatics/btp696
Pozo-Montuy G., Aguilar-Cucurachi M. S., Aureli F., Briseño-Jaramillo M., Canales-Espinosa D., Cárdenas-Navarrete A., et al. (2024). Howler monkey die-off in southern Mexico. Am. J. Primatol., e23684. doi: 10.1002/ajp.23684
Pritchard J. K., Stephens M., and Donnelly P. (2000). Inference of population structure using multilocus genotype data. Genetics 155, 945–959. doi: 10.1093/genetics/155.2.945
Puechmaille S. J. (2016). The program structure does not reliably recover the correct population structure when sampling is uneven: Subsampling and new estimators alleviate the problem. Mol. Ecol. Resour. 16, 608–627. doi: 10.1111/men.2016.16.issue-3
Radespiel U. and Bruford M. W. (2014). Fragmentation genetics of rainforest animals: Insights from recent studies. Conserv. Genet. 15, 245–260. doi: 10.1007/s10592-013-0550-3
Ralls K., Ballou J. D., Dudash M. R., Eldridge M. D. B., Fenster C. B., Lacy R. C., et al. (2018). Call for a paradigm shift in the genetic management of fragmented populations. Conserv. Lett. 11, 1–6. doi: 10.1111/conl.2018.11.issue-2
Rambaut A. (2018). FigTree v.1.4. Available online at: http://tree.bio.ed.ac.uk/software/figtree (Accessed November 27, 2024).
R Core Team (2021). R: a language and environment for statistical computing (Vienna: R Foundation for Statistical Computing). Available at: http://www.R-project.org/.
Rimbach R., Link A., Heistermann M., Gómez-Posada C., Galvis N., and Heymann E. W. (2013). Effects of logging, hunting, and forest fragment size on physiological stress levels of two sympatric ateline primates in Colombia. Conserv. Physiol. 1, cot031. doi: 10.1093/conphys/cot031
Ronquist F., Teslenko M., van der Mark P., Ayres D. L., Darling A., Höhna S., et al. (2012). MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Rozas J., Ferrer-Mata A., Sanchez-Del Barrio J. C., Guirao-Rico S., Librado P., Ramos-Onsins S. E., et al. (2017). DnaSP v6, DNA polymorphism analyses of large data sets. Mol. Biol. Evol. 34, 3299–3302. doi: 10.1093/molbev/msx248
Ruiz-García M., Albino A., Pinedo-Castro M., Zeballos H., Bello A., Leguizamon N., et al. (2019). First molecular phylogenetic analysis of the Lagothrix taxon living in southern Peru and northern Bolivia: Lagothrix lagothricha tschudii (Atelidae, Primates), a new subspecies. Folia Primatol. 90, 215–239. doi: 10.1159/000497251
Ruiz-García M., Cerón Á., Sánchez-Castillo S., Rueda-Zozaya P., Pinedo-Castro M., Gutierrez-Espeleta G., et al. (2017). Phylogeography of the mantled howler monkey (Alouatta palliata; Atelidae, Primates) across its geographical range by means of mitochondrial genetic analyses and new insights about the phylogeny of Alouatta. Folia Primatol. 88, 421–454. doi: 10.1159/000480502
Sardell J. M. and Uy J. A. (2016). Hybridization following recent secondary contact results in asymmetric genotypic and phenotypic introgression between island species of Myzomela honeyeaters. Evolution 70, 257–269. doi: 10.1111/evo.12864
Schrago C. G., Mello B., and Soares E. R. (2013). Combining fossil and molecular data to date the diversification of New World Primates. J. Evol. Biol. 26, 2438–2446. doi: 10.1111/jeb.2013.26.issue-11
SEMARNAT (2010). Norma Oficial Mexicana NOM-059-ECOL-2010, protección ambiental-especies nativas de México de flora y fauna silvestres-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-Lista de especies en riesgo (México: Diario Oficial de la Federación).
Serio-Silva J. C., Vida-García F., Alvarado-Villalobos M. A., Aristizabal J., Díaz-López H., Collado-Torres R., et al. (2015). Identity and conservation models in Balancán, Mexico: key to saving the critically endangered Mexican black howler monkey. 6th Eur. Fed. Primatol. Meet. Folia Primatol. 86, 235–386. doi: 10.1159/000435825
Smith M. F. and Patton J. L. (1993). The diversification of South American murid rodents: evidence from mitochondrial DNA sequence data for the akodontine tribe. Biol. J. Linn. Soc 50, 149–177. doi: 10.1111/j.1095-8312.1993.tb00924.x
Solórzano-García B., Zubillaga D., Piñero D., and Vázquez-Domínguez E. (2021). Conservation implications of living in forest remnants: Inbreeding and genetic structure of the northernmost mantled howler monkeys. Biotropica 53, 1163–1177. doi: 10.1111/btp.12958
Suárez-Atilano M., Pacheco-Sierra G., Vázquez-Domínguez E., Kass J. M., Paz A., and Pérez-Alquicira J. (2024). Genomic and environmental insights and conservation challenges for two hybridizing iconic crocodile species across Mexico: Crocodylus acutus and C. moreletii. Anim. Conserv. 27, 308–323. doi: 10.1111/acv.12907
Suchard M. A., Lemey P., Baele G., Ayres D. L., Drummond A. J., and Rambaut A. (2018). Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 4, vey016. doi: 10.1093/ve/vey016
Tajima F. (1989). Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123, 585–595. doi: 10.1093/genetics/123.3.585
Van Belle S. and Di Fiore A. (2022). Dispersal patterns in black howler monkeys (Alouatta pigra): Integrating multiyear demographic and molecular data. Mol. Ecol. 31, 391–406. doi: 10.1111/mec.16227
Van Belle S. and Estrada A. (2006). “Demographic features of Alouatta pigra populations in extensive and fragmented forests”, in New Perspectives in the Study of Mesoamerican Primates: Distribution, Ecology, Behavior, and Conservation. Eds. Estrada A., Garber P. A., Pavelka M. S. M., and Luecke L. (Springer, Boston, MA). doi: 10.1007/0-387-25872-8_6
Van Belle S., Estrada A., Strier K. B., and Di Fiore A. (2012). Genetic structure and kinship patterns in a population of black howler monkeys, Alouatta pigra, at Palenque National Park, Mexico. Am. J. Primatol. 74, 948–957. doi: 10.1002/ajp.2012.74.issue-10
vonHoldt B. M., Brzeski K. E., Wilcove D. S., and Rutledge L. Y. (2018). Redefining the role of admixture and genomics in species conservation. Conserv. Lett. 11, 1–6. doi: 10.1111/conl.12371
Wang M.-S., Wang S., Li Y., Jhala Y., Thakur M., Otecko N. O., et al. (2020). Ancient hybridization with an unknown population facilitated high-altitude adaptation of canids. Mol. Biol. Evol. 37, 2616–2629. doi: 10.1093/molbev/msaa113
Winter D. J. (2012). MMOD: an R library for the calculation of population differentiation statistics. Mol. Ecol. Resour. 12, 1158–1160. doi: 10.1111/j.1755-0998.2012.03174.x
Keywords: Alouatta palliata, Atelidae, hybridization, mantled howler monkey, non-human primates, tropical forests
Citation: Vázquez-Domínguez E, Solórzano-García B, Reyes-Vázquez M and Zubillaga-Martín D (2025) Population genetics and evolutionary history of the black howler monkey Alouatta pigra and hybrid populations in Mexico. Front. Ecol. Evol. 13:1551979. doi: 10.3389/fevo.2025.1551979
Received: 27 December 2024; Accepted: 08 May 2025;
Published: 03 June 2025.
Edited by:
Isabel Marques, Universidade de Lisboa, PortugalReviewed by:
Emmanuel Paul Gilissen, Royal Museum for Central Africa, BelgiumHong Wu, Yunnan University, China
Copyright © 2025 Vázquez-Domínguez, Solórzano-García, Reyes-Vázquez and Zubillaga-Martín. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ella Vázquez-Domínguez, ZXZhenF1ZXpAZWNvbG9naWEudW5hbS5teA==; Brenda Solórzano-García, YnJlbmRhX3NvbG9yemFub0B5YWhvby5jb20ubXg=
†These authors share first authorship