 Cesar Mayta1,2,3*
Cesar Mayta1,2,3* Matthias Schleuning4
Matthias Schleuning4 Cecilia L. López1,2,3
Cecilia L. López1,2,3 Mariana Villegas3,5Isabell Hensen1,6
Mariana Villegas3,5Isabell Hensen1,6 Silvia C. Gallegos1,2,3
Silvia C. Gallegos1,2,3- 1Institute of Biology/Geobotany and Botanical Garden, Martin Luther University Halle-Wittenberg, Halle, Germany
- 2Herbario Nacional de Bolivia, Instituto de Ecología, Universidad Mayor de San Andrés, La Paz, Bolivia
- 3Santiago de Chirca Biological Station, La Paz, Sud Yungas, Bolivia
- 4Senckenberg Biodiversity and Climate Research Centre (SBiK-F), Senckenberganlage, Frankfurt am Main, Germany
- 5Instituto de Ecología, Universidad Mayor de San Andrés, La Paz, Bolivia
- 6German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, Leipzig, Germany
Most tropical plant species depend on animals to disperse their seeds. Seed dispersal by animals allows plants to colonize new sites in deforested habitats helping to accelerate forest recovery. However, deforestation can affect the interactions between animals and plants that are crucial for forest regeneration. In this study, we analyze the differences on the composition of functional traits and on seed-dispersal interactions and functions by birds and bats between forest and deforested areas dominated by bracken fern. At eight study sites, we captured birds and bats in bracken-dominated areas and at forest in the tropical montane forest of Bolivia and analyzed their droppings. We found a similar composition of functional traits related to animal size, gape width and degree of frugivory of bats in both habitat types. Hand-wing index of birds was significantly higher in bracken compared to forest. Birds had more interaction richness and abundance in bracken than in forest, and bats had more interaction abundance in forest than in bracken. The majority of seeds dispersed by birds and bats in bracken were from pioneer species. We conclude that, although seed dispersal by birds and bats is maintained in the bracken-dominated areas, most of the dispersed seeds belong to pioneer species. Pioneer species are unlikely to establish in bracken-dominated areas, making the natural regeneration process of these deforested areas very slow.
1 Introduction
More than 90% of tropical plant species depend on animals to disperse their seeds (Jordano, 2000). Birds and bats are considered particularly important seed dispersers, as they play a key role in forest regeneration due to the number and diversity of seeds that they disperse (Jacomassa and Pizo, 2010). Seed dispersal by these animals allows plants to colonize new sites in deforested habitats helping to accelerate forest recovery (De La Peña-Domene et al., 2014). Deforestation affects the diversity and composition of animals and plants (Haddad et al., 2015) and their biotic interactions, including seed dispersal (Laurance, 2005). Given the importance of birds and bats, it is necessary to evaluate the differences in seed dispersal by birds and bats between forested and deforested areas and to analyze the consequences for forest regeneration.
Many tropical deforested regions harbor forest fragments that are surrounded by open areas often composed by bracken fern (Pteridium spp.) (Lippok et al., 2013; Curatola Fernández et al., 2015; Matongera et al., 2018). Bracken usually grows after fire (Roos et al., 2010) and can dominate the vegetation for long periods of time (Hartig and Beck, 2003). Bracken produces dense fronds and a deep litter layer that could impede the establishment of other plant species (Marrs et al., 2000), especially of pioneer species. However, bracken generates microenvironmental conditions (high humidity, low temperature, low light) that could favor the establishment of non-pioneer shade-tolerant species (Gallegos et al., 2015; López et al., 2024; Ssali et al., 2019). The dominance of bracken can last several years, hindering the forest regeneration process. The slow forest regeneration is mainly linked to the seed limitation (i.e. low seed arrival) of animal-dispersed, non-pioneer plant species (Saavedra et al., 2015; Gallegos et al., 2016; Ssali et al., 2018). This limitation could be related to the scarcity of food resources, shelters and perches for animal seed dispersers, and to differences in species composition of seed dispersers between forest and bracken-dominated areas (Saavedra et al., 2015; Gallegos et al., 2024; Mayta et al., 2024). Although there are indications of seed limitation of non-pioneer species, it is unclear whether this is due to the functional traits of seed dispersers or other characteristics in the animal-plant interaction in bracken-dominated areas.
Fragmented landscapes are characterized by the presence of different habitats such as the forest interior, the forest edges and the more disturbed open areas (Hardwick et al., 1997). These habitats usually harbor different animal communities and properties; for example, bird species richness, abundance and composition differ between forest interior and deforested areas (Maya-Elizarrarás and Schondube, 2015; Tchoumbou et al., 2020). At disturbed forest edges, birds that require specific habitat conditions and food resources may be lost (Menke et al., 2012; Jankowski et al., 2021), and generalist species gain in richness and abundance (Morante-Filho et al., 2015). In addition, as a result of deforestation and forest fragmentation, large frugivores tend to disappear (Renjifo, 1999; Markl et al., 2012), while small frugivores and omnivores seem to better tolerate such disturbances (O’Dea and Whittaker, 2007; Gomes et al., 2008). Tropical forest disturbance could also generate differences in the trait composition of bird communities between forested and deforested areas (Newbold et al., 2013; García-Morales et al., 2016). For instance, bird species of disturbed habitats tend to be smaller and have a higher capacity to flight long distances than birds of less disturbed habitats (Camargo et al., 2020; Messina et al., 2021; Falconí-López et al., 2024). On the other hand, the abundance of frugivorous bats has been shown to increase in fragmented habitats (Cleary et al., 2016), while the trait composition of bat communities can be stable (Farneda et al., 2020; Díaz-B et al., 2023). It is therefore likely that small frugivorous and omnivorous birds, as well as frugivorous bats are particularly important for seed dispersal and forest regeneration in disturbed forest habitats (Carlo and Morales, 2016; Muscarella and Fleming, 2007), but their joint effects on tropical forest regeneration remains poorly understood. Deforestation not only affects the diversity and traits of animal seed dispersers, but can also have an effect on the interaction between plants and their animal seed dispersers (González-Varo et al., 2023). In some cases, more disturbed habitats could have less pairwise bird-plant interactions (Menezes Pinto et al., 2021), however, other studies detected an opposite pattern (Menke et al., 2012; Saavedra et al., 2014). Thus, the gain or loss of seed-dispersal interactions may related to the composition of animal and plant communities in the different ecosystems.
In this study, we aim to analyze the differences in trait composition related to seed dispersal, interactions, and seed-dispersal functions of birds and bats between bracken-dominated areas and forest. To this end, we addressed the following questions: 1) Is there a difference in the functional trait composition of animal seed dispersers (birds and bats) between forest and bracken-dominated areas? We expected that birds recorded in bracken-dominated areas would have a lower body mass, smaller gape width and morphological characteristics in the wings that allow them to flight large distance than birds captured at forest (Camargo et al., 2020; Messina et al., 2021; Falconí-López et al., 2024), whereas we did not expect differences in the trait composition of bats between both habitats [(Farneda et al., 2020). 2] What are the differences in plant-animal interactions between bracken-dominated areas and forest? We expected to find more seed-dispersal interactions in bracken-dominated areas due to their high species richness and higher abundance compared to forests, and the opposite pattern for bats due to their high abundance in forest compared to bracken-dominated areas (Gallegos et al., 2024). 3) Is there a difference in the abundance and richness of seeds from plant species with different life histories between bracken-dominated areas and forests? We expected to find more seeds of pioneer species in animals captured in bracken-dominated areas than in animals captured at forests because of the expected changes in trait composition, particularly in birds. Birds with small gape width usually disperse small seeds (Wheelwright, 1985; Bovo et al., 2018), and small seeds are characteristic of pioneer species (Valio and Scarpa, 2001). Seed dispersal in bracken-dominated areas has primarily studied though the analysis of seed rain. The morphological characteristics of animal seed dispersers (birds and bats), and the characteristics and functions of animal-plant interactions in these widely distributed deforested areas remains poorly understood.
2 Materials and methods
2.1 Study area
The study was conducted in a tropical montane forest in the Bolivian Andes between 1,900 and 2,350 m a.s.l. near the town of Chulumani (16°24’37” S, 67°31’37” W). The mean annual temperature is 20.5°C, and mean annual precipitation is 1390 mm (Molina-Carpio et al., 2019). In this region, montane forests have been deforested due to logging and human induced fires (Killeen et al., 2005). The remnant forests are highly fragmented, and are restricted to the top of the mountains. Forest fragments are surrounded by deforested areas dominated mostly by bracken fern (Pteridium esculentum subsp. arachnoideum) and shrubs of Asteraceae and Melastomataceae (Lippok et al., 2013). The majority of plant species in bracken-dominated areas are dispersed by wind, however there are species such as Myrsine coriacea (Primulaceae), Gaultheria erecta (Ericaceae) and Leandra carassana (Melastomataceae) that are dispersed by birds and are common in these deforested habitats (Lippok et al., 2013). There are also some species of Solanaceae and Piperaceae that are dispersed by bats in bracken dominated areas, although they are scarce (Lippok et al., 2014; López et al., 2024). The most common species at the forest edge of the study area are Hedyosmum racemosum (Chloranthaceae), Clusia elongata (Clusiaceae) and Hieronyma fendlerii (Phyllanthaceae), among those dispersed by birds, and Piper pilirameum, P. trigonastrifolium and Vismia glaziovii among those dispersed by bats (Lippok et al., 2014).
2.2 Study design
We conducted the study at eight sites located at the borders of remnant forests separated by at least 1 km (Supplementary Figure S1). At each site, we placed eight mist nets (9 x 2.5 m) at 0.5 m from the ground. Four mist nets were set inside the forest at 20 m from the forest margin (hereafter forest), and four mist nets were set in the bracken-dominated area (hereafter bracken) at 20 m from the forest margin. From March 2020 to June 2021, we visited each site four times and captured birds and bats. Mist nets were placed for 2–3 days on each visit, from 0630 to 1830 h to capture birds, and from 1830 to 2300 to capture bats. We closed mist nets when it rained. Total sampling effort was 4861 net-hours for birds, and 2576 net-hours for bats. It should be noted that the use of mist nets favors the capture of birds and bats present in the understory (Whitman et al., 1997).
2.3 Bird, bat and seed sampling
Captured birds and bats were placed in cloth bags for 20–30 minutes to collect feces and regurgitates. Captured animals were weighed, measured, marked with color rings, identified to the species level and released. For each bird species, we measured body mass, hand-wing index and gape width, while we could only measure body mass for bats. We chose these traits because body mass is related to the amount and size of consumed fruits and seeds, as well as the distance of seed dispersal (Wotton and Kelly, 2012). Gape width determines whether a bird species can consume large, medium or small-sized seeds (Wheelwright, 1985). In addition, we recorded the degree of frugivory of bats from the EltonTraits 1.0 database (Wilman et al., 2014). Hand-wind index (HWI) was calculated using the equation HWI = 100 x (DK/LW), where DK is Kipp’s distance (the distance between the tip of the first secondary feather and the tip of the longest primary feather) and LW is wing length. HWI is related to the flight efficiency; high values are related to species that could perform long-distance flights, whereas small values are related to more sedentary species with lower dispersal abilities (Sheard et al., 2020; Falconí-López et al., 2024). Birds and bats were classified into feeding guilds according to Aguirre (2007), Aguirre et al. (2009), Billerman et al. (2022), and Herzog et al. (2016). Because we found seeds in the feces and regurgitates of omnivorous and frugivorous birds and bats (hereafter referred to as birds and bats), functional trait analysis included both guilds. The bats were identified from external morphological characteristics, so it is possible that there was some misidentification in bats of the same genus (e.g. in Carollia). However, Aguirre et al. (2009) and Dı́az et al. (2016) provide clear external morphological differences that may help to reduce misidentifications.
We collected the seeds carried by birds and bats from the cloth bags. Undamaged seeds were extracted from all droppings and regurgitates, grouped into morphotypes, counted and identified under a dissecting microscope to the lowest possible taxonomical level by comparing them with a reference collection from the study area deposited at the Santiago de Chirca Biological Station. Overall, 321 seed samples (droppings and/or regurgitates) could be collected (267 seeds from birds and 54 from bats). Each of the identified plant species was classified as pioneer or non-pioneer species according to previous studies in our study site (Gallegos et al., 2016; Lippok et al., 2013, 2014; López et al., 2024), expert knowledge, and information at Herbario Nacional de Bolivia (LPB). These data were used to calculate the abundance and species richness of plants dispersed by birds and bats in forest and bracken.
2.4 Statistical analysis
We used generalized linear mixed-effects models (GLMMs) for all the analyses. The use of mist nets favors the capture of small birds and bats, and capture efficiency may vary across environments with different vegetation structures (Jenni et al., 1996). To address this potential sampling bias, we performed the analysis of functional traits of birds and bats by randomly drawing the same number of individuals from each habitat at each site. A GLMM was fitted to each subsample, and this process was repeated 999 times. We then interpreted the results using the mean of the estimates across all GLMMs. We analyzed the differences of gape width, body mass and HWI for birds and body mass and frugivory degree for bats. Habitat type (forest and bracken) was included as the independent fixed effect, and bird or bat species, nested within site, nested within fragment, were the random effects. The models used a Gaussian distribution of errors, except for bat’s frugivory degree that used an betabinomial distribution of errors. We compared the seed dispersal interactions in separate models for bird and bats, taking interaction richness (number of unique interactions) and interaction abundance (interaction frequency) as the response variable, habitat type (forest and bracken) as the independent fixed effect, and site nested in fragment as the random effects. The models used a Gaussian distribution of errors for interaction richness and Poisson distribution of errors for interaction abundance. The detection of all the species/interactions in natural environments is difficult (Jordano, 2016; Chiu et al., 2023). The observed interactions depend on the sample completeness which difficult the comparison of datasets across different sites (Vizentin-Bugoni et al., 2016). To overcome this problem, we estimated the interaction richness with the iNEXT.link package (Chiu et al., 2023). We used the non-asymptotic coverage-based method, which relies on coverage-based rarefaction and extrapolation (Chiu et al., 2023). This method was applied at each site, within each habitat, and for each animal group. Resulting in a total of 16 estimation of interaction richness for birds and 16 for bats. Finally, we compared the abundance and species richness of pioneer and non-pioneer species found in feces and regurgitates of birds and bats captured in bracken and forest. We performed the analysis of the abundance of seeds by randomly drawing the same number of individuals from each habitat at each site. A GLMM was fitted to each subsample, this process was repeated 999 times and we interpreted the results using the mean of the estimates across all GLMMs. Species richness of pioneer and non-pioneer species was calculated for each study site. In separate models we tested the abundance of seeds, and seed species richness as the response variables for birds and for bats. In the models used to analyze seed abundance, life history (pioneer or non-pioneer), habitat (forest and bracken) and their interaction were the independent fixed effects. Bird or bat species, nested within site, nested within fragment, were the random effects. For seed species richness models, life history (pioneer or non-pioneer), habitat (forest and bracken) and their interaction were the independent fixed effects. Site nested within fragment were the random effects. The four models used a Poisson distribution of errors. All models were performed with the glmmTMB package (Brooks et al., 2017). To determine significant differences between habitats, we performed Tukey post hoc tests using the emmeans package (Russell et al., 2024). For all models, the significance of the fixed factors was determined by Likelihood Ratio Tests (LRT) between models with and without the term of interest. Additionally, we calculated marginal R2 values to assess the proportion of variance explained by the fixed factor using the r.squaredGLMM function of the MuMIn package (Barton, 2023). Overdispersion in model residuals were tested with DHARMA package (Hartig, 2024). All statistical analyses were conducted in R 4.2.2 (R Foundation for Statistical Computing, 2022). We assessed the sample completeness of plant-animal interaction using method “abundance” with the iNEXT package (Hsieh et al., 2016).
3 Results
3.1 Functional traits of birds and bats
We captured 31 bird species, 27 species (99 individuals) in bracken and 13 species (65 individuals) at forest (Supplementary Table S1). We caught 12 bat species, eight in bracken (23 individuals) and eight at forest (34 individuals) (Supplementary Table S1). In bracken, the most common bird species were Elaenia albiceps and Mionectes striaticollis; 23 individuals of both species were recorded. The most common bat species in bracken were Sturnira oporaphilum and Carollia perspicillata, of which we recorded nine and four individuals, respectively. At forest, M. striaticollis and Chiroxiphia boliviana were the most abundant bird species, of which we recorded 23 and 17 individuals, respectively. Among the bats, Carollia perspicillata and Carollia brevicauda were the most common species with 14 and 11 records, respectively.
We found no differences in body mass (X2 = 0.098, p = 0.75) and gape width (X2 = 0.10, p = 0.75) of birds between bracken and forest (Figures 1A, B; Supplementary Table S2). The hand-wing index (HWI) was significantly higher in bracken compared to forest (X2 = 43.65, p < 0.001, Figure 1C; Supplementary Table S2). For bats, we found no differences in body mass (X2 = 0.06, p = 0.80) and frugivory degree (X2 = 1.47, p = 0.22) between the two habitat types (Figures 1D, E; Supplementary Table S2). The largest bird species such as Aulacorynchus coeruleicinctis (~208 g) and Pyrrhura molinae (~ 78 g) were only captured in bracken. At forest, Turdus amaurochalinus (~ 58 g) and Pipreola frontalis (~ 48 g) were the largest bird species. Among the bats, Sturnira tildae (~20 g) and S. oporaphilium (~17 g) were the largest species in bracken and at forest, respectively.
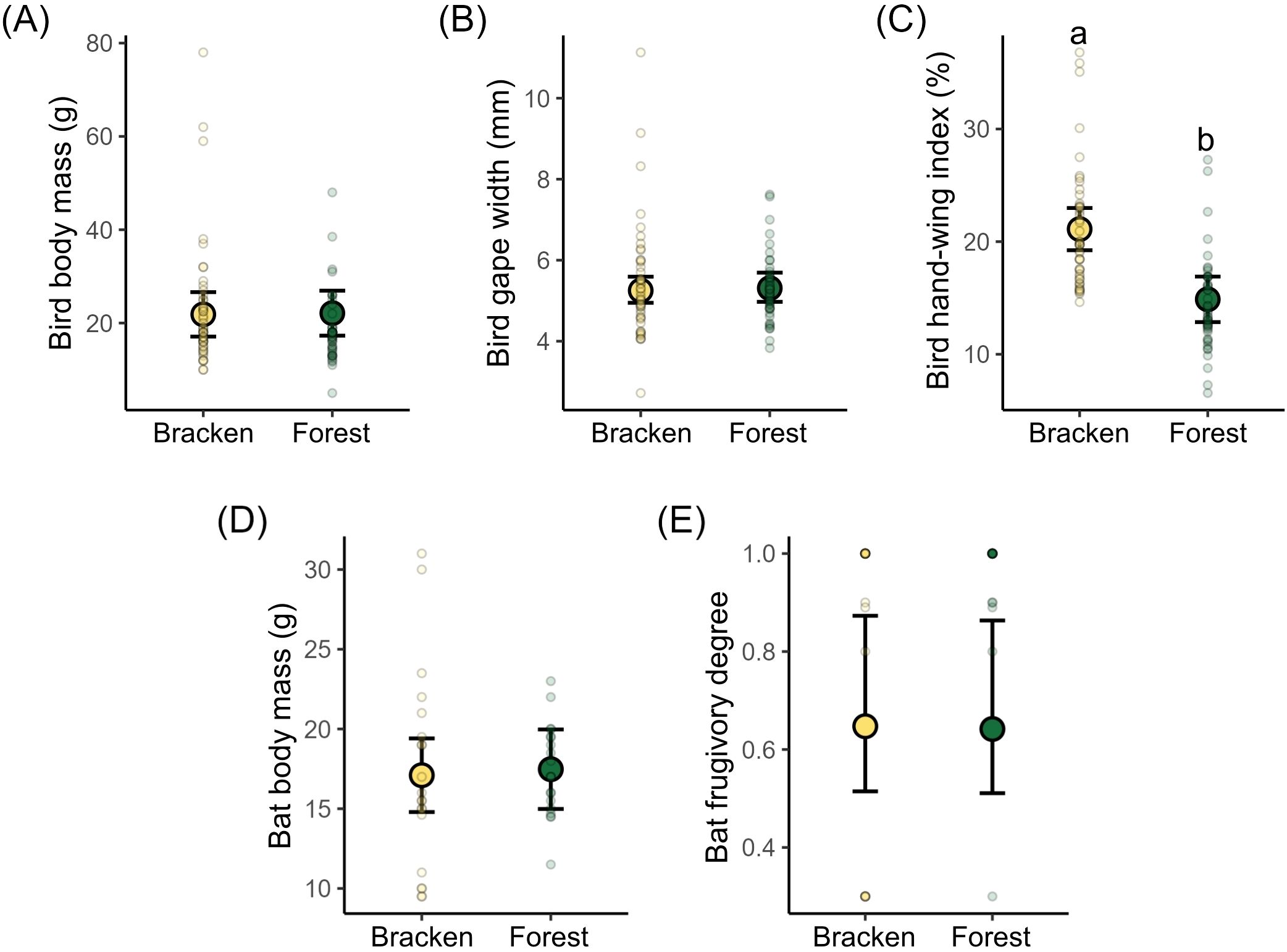
Figure 1. Comparison of community-weighted means of (A) bird body mass, (B) bird gape width and (C) bird hand-wing index between bracken and forest habitats of 31 bird species. Comparison of community-weighted means of (D) body mass and (E) frugivory degree between bracken and forest habitats of 12 bat species. Different letters indicate significant differences at level 0.05 based on post hoc Tukey comparison among habitats. Large circles with error bars denote the mean values ± 95% CI estimated by GLMMs, small circles denote the observed values at the eight sites per habitat type.
3.2 Seed-dispersal interactions
After 7,437 net-hours of sampling effort for birds and bats, we registered a total of 124 interactions between 31 species of seed dispersers (25 birds, 6 bats) and 39 plant species. Of these, 69 interactions were recorded in bracken and 55 at forest. The sampling completeness was 89% in the bracken area and 91% at the forest edge (Supplementary Figure S2). For birds we found that in bracken the interaction richness and interaction abundance were 2.50 and 2.23 times higher, respectively, compared to forest (X2 = 7.02, p = 0.008 interaction richness, X2 = 11.57, p < 0.001 interaction abundance; Figures 2A, B; Supplementary Table S3). For bats we did not find differences for interaction richness between bracken and forest (X2 = 3.47, p = 0.07; Figure 2C; Supplementary Table S3). The abundance of interactions was 2.7 times higher in bracken than in forest (X2 = 7.19, p = 0.007; Figure 2D; Supplementary Table S3).
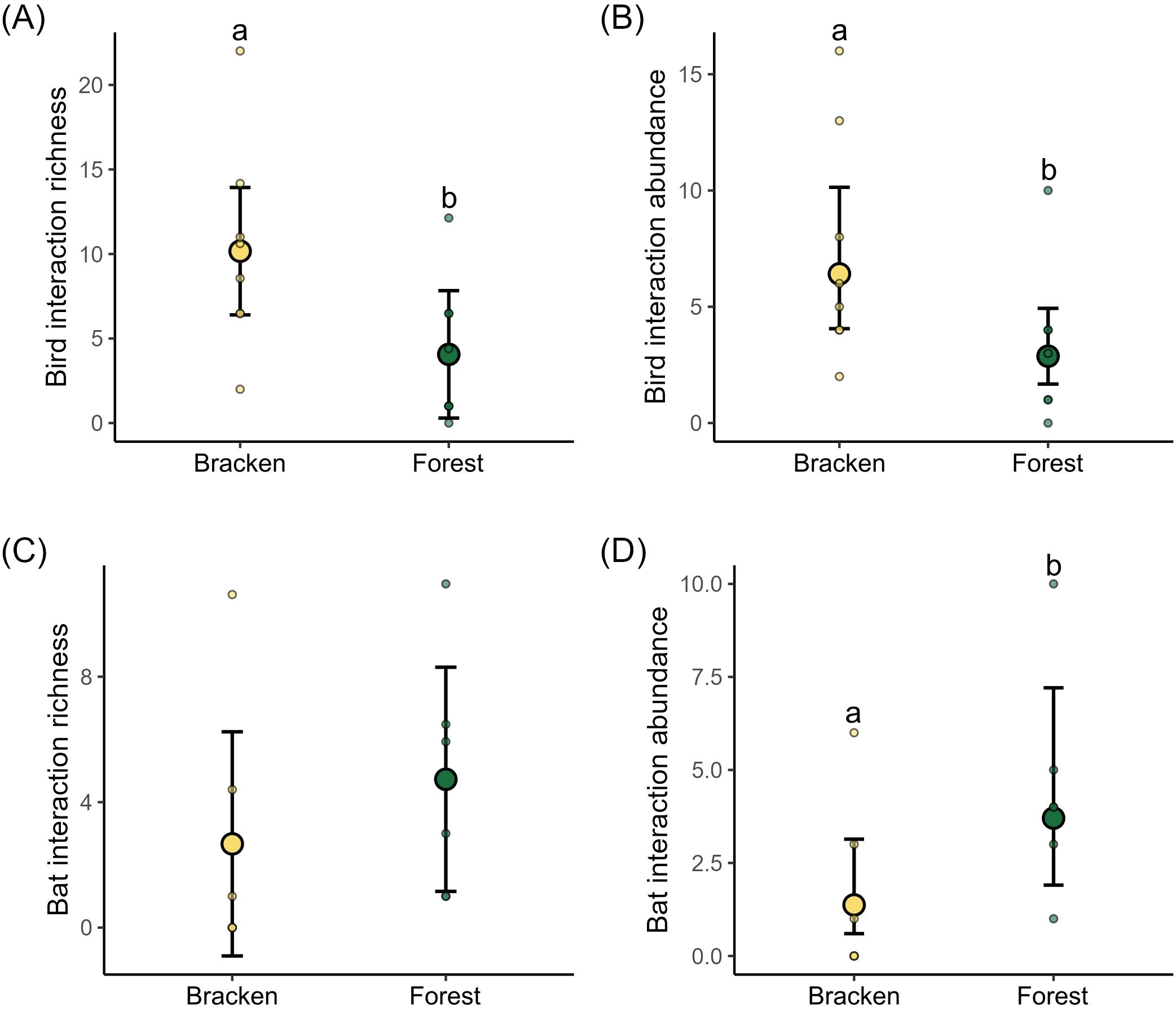
Figure 2. Difference in the (A) interaction richness and (B) abundance of birds and (C) interaction richness and (D) abundance of bats between bracken and forest habitats. Different letters indicate significant differences at level 0.05 based on post hoc Tukey comparison among habitats. Large circles with error bars denote the mean values ± 95% CI estimated by GLMMs, small circles denote the observed values at the eight sites per habitat type.
3.3 Successional status of animal-dispersed species
We found that the abundance and species richness of bird-dispersed pioneer seeds in bracken were 3.34 and 2.15 times higher, respectively, compared to forest (Figures 3A, B; Supplementary Table S4). In bracken the abundance of bird-dispersed pioneer species was 9.54 times higher than the abundance of non-pioneer species in the same treatment (X2 = 18.52, p < 0.001; Figure 3A; Supplementary Table S4). Abundance of bat-dispersed pioneer seeds in bracken was 5.12 times higher compared to forest (X2 = 5.02, p = 0.02; Figure 3C; Supplementary Table S4), and 3.47 times higher compared to non-pioneer seeds in forest (X2 = 7.80, p = 0.005; Figure 3C; Supplementary Table S4). We did not find differences in bat-dispersed richness between habitats and life history categories (Figure 3D; Supplementary Table S4). The most abundant pioneer species in feces were Gaultheria erecta (bird-dispersed), Piper elongatum (bird and bat-dispersed) and Gaultheria reticulata (bird-dispersed) (Supplementary Table S5).
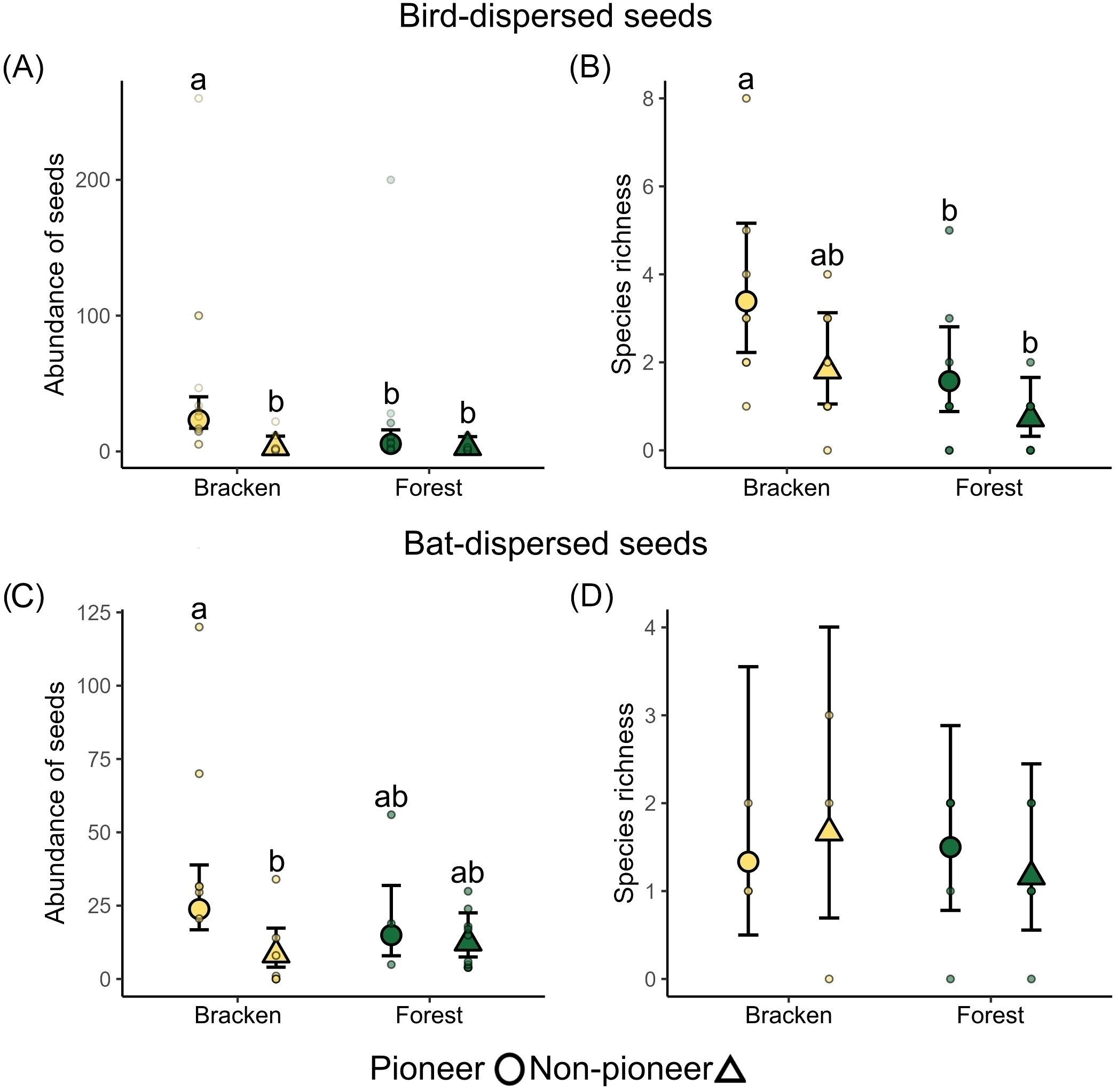
Figure 3. Differences in (A) abundance and (B) species richness of seeds found in birds and (C) abundance and (D) species richness of seeds found in bats captured in bracken and forest habitats. Different letters indicate significant differences at level 0.05 based on post hoc Tukey multiple comparison among habitats and life histories. Large circles (Pioneer) and triangles (non-pioneer) with error bars denote means ± 95% CI estimated by the GLMMs, small circles denote the observed values at each of the eight study sites.
4 Discussion
We found a similar trait composition of birds and bats in bracken and forest, except for the hand-wing index of birds that was higher in bracken compared to forest. We recorded higher interaction richness and abundance for birds in bracken compared to forest, while for bats, interaction abundance was higher in forest than in bracken. Importantly, seed-dispersing animals transported more pioneer than non-pioneer species in bracken. However, pioneer species are unlikely to become established in bracken-dominated areas due to bracken shade (Gallegos et al., 2015, 2016; Ssali et al., 2019; López et al., 2024). Although previous studies have suggested a limitation of non-pioneer seeds in bracken-dominated areas (Saavedra et al., 2015; Mayta et al., 2024), our work reveals that this limitation is linked to the morphological characteristics of dispersing animals (small animals that disperse small seeds) and the higher quantity of pioneer seeds consumed by these animals.
Deforested environments can favor the dominance of a subset of functional traits in birds and bats (Frank et al., 2017; Bovo et al., 2018). We expected birds in forests to have a higher body mass and gape width than birds in bracken, because birds in highly disturbed habitats tend to be smaller than birds in less disturbed habitats (Pavlacky et al., 2015; Bovo et al., 2018). Moreover, the loss of forested areas could lead to differences in composition of animal-seed disperser communities (Chiarello, 1999; Morante-Filho et al., 2015). For example, Gallegos et al. (2024) found differences in the composition of birds between forest and bracken areas. However, we found no differences in body mass and gape width of birds in the two habitats. The majority of birds captured in bracken and forest habitats corresponded to omnivorous and small frugivorous birds; both guilds are dominant in forest edges and deforested areas (Menezes Pinto et al., 2021) and have similar morphological characteristics. We found that birds captured in bracken had higher hand-wing index (HWI) compared to those captured in forest. Birds with high HWI could fly long distance and cross open areas and are less sensitive to habitat fragmentation than birds with low HWI (Jones et al., 2023; Falconí-López et al., 2024). It is possible that the changes in trait composition would have been more pronounced if we had compared the bracken habitats with the interior of the forest fragments. Previous studies showed that large-bodied and large-gaped bird species increase in importance towards the most conserved forest interior (Sekercioglu, 2012; Saavedra et al., 2014; Emer et al., 2018). We found no differences in the trait composition of bats. Frugivorous bats are known to tolerate habitat disturbance (Farneda et al., 2020) and these bats can move between forested and open areas, possibly in search of fruit resources from pioneer species (Muscarella and Fleming, 2007). Accordingly, in bracken areas, there are species of Piper, Vismia and Solanum (Lippok et al., 2013, 2014) that are preferentially consumed by frugivorous bats, but in low densities. Another reason for the weak differences in bird and bat communities could be our sampling method (mist netting) that tends to favor the capture of small seed dispersers, while especially large birds are more likely to escape from mist nets (Jenni et al., 1996).
The loss of forested areas affects the communities of seed-dispersing animals, plants and their interactions (Emer et al., 2020; Menezes Pinto et al., 2021). Some studies found less interactions in disturbed habitats due to the reduced abundance of seed-dispersing animals (Marjakangas et al., 2020; Menezes Pinto et al., 2021; Li et al., 2022). Contrary, we found that bracken had more bird interactions richness and abundance than forest. This could be related to the fact that in our study sites the species richness and abundance of small omnivorous/frugivorous birds are higher in bracken than in the forest (Gallegos et al., 2024). On the other hand, in bracken areas there are small-seeded/fruited plants of the families Melastomataceae and Ericaceae (Huber et al., 1984; Lippok et al., 2013; Martínez-Ramos et al., 2016) that bear fruits most of the year and are attractive to birds. For bats, we found higher interaction abundance in forest than in bracken. The abundance of frugivorous bats is related to the abundance of their food (Klingbeil and Willig, 2009). In forest there are more species and more individuals of plants of the families Piperaceae and Solanaceae than in bracken (Lippok et al., 2013, 2014). In addition, the abundance of frugivorous bats is higher in forest compared to bracken (Gallegos et al., 2024). Thus, our results for bats could be related to higher abundance of frugivorous bats and their food resources in forest compared to bracken.
We found that birds and bats present in the bracken area carried significantly more seeds of pioneer species than birds and bats captured at the forest. This could be related to the fact that the majority of birds and bats that disperse seeds into bracken area were small frugivorous and omnivorous species that mostly consume fruits with small seeds (Snow, 1981; Arteaga et al., 2006; Emer et al., 2018) a characteristic of pioneer plants (Swaine and Whitmore, 1988; Valio and Scarpa, 2001). Most of the seeds of pioneer species found in bracken were typical from deforested areas, and are rare in the forest. On the other hand, non-pioneer, shade-tolerant plant species that usually have large seeds (Thompson and Rabinowitz, 1989; Khurana and Singh, 2006) are mostly dispersed by large specialist frugivore birds (Snow, 1981; Carlo et al., 2022) and large mammals (Cramer et al., 2007), which were rare in the bracken areas. For instance, Saavedra et al. (2015) and Ssali et al. (2018) found a lack of non-pioneer and large-seeded plants in the seed rain in bracken areas of south America and Africa, respectively, suggesting that there was very little dispersal of such species from the forest. In the forest interior and forest edge of tropical mountain forests there are several species of Lauraceae, and species such as Rhamnus sphaerosperma, Symplocos arechea that have large seeds (>10 mm) and are non-pioneer species (Oosterhoorn and Kappelle, 2000; Günter et al., 2007; Lippok et al., 2014; Saavedra et al., 2015; López et al., 2024). Bracken fosters the establishment of non-pioneer species, but not of pioneer species (Gallegos et al., 2015; Ssali et al., 2019; López et al., 2024). Although pioneer species arrive into bracken areas, the recruitment of these species is hindered by the low light conditions under bracken, that is detrimental to light demanding pioneer species (Gallegos et al., 2015, 2016; López et al., 2024). In this study, we did not analyze the actual seed deposition by birds and bats. There is a possibility that animals captured in bracken from both groups are moving from one forest patch to another. However, there is also the possibility that birds perch in a plant that remains in the deforested area or that bats defecate in flight and drop seeds. Future studies can analyze aspects related to the behavior of seed dispersers, for example, analyze if birds that consume fruits in the forest are able to go out to bracken areas and deposit seeds in these deforested areas.
To gain a deeper understanding of the seed-dispersal process in the bracken area, future studies could combine different techniques to study seed-dispersal interactions. For example, the use of mist nets combined with focal observations could help to have more complete and representative data of the interactions between plants and birds (Quintero et al., 2022). We acknowledge that direct observations of plant-animal interactions in bracken areas and nearby forests could be very effective. However, one of the best methods to know which seeds birds and bats actually deliver to deforested sites is mist netting which allowed us to estimate the actual seed-dispersal functions by birds and bats. It is important to recognize that this method primarily captures small frugivores and may underestimate the role of large and other frugivorous animal groups. The characteristics of seed-dispersal interactions vary according the forest strata (Schleuning et al., 2011; Thiel et al., 2021). Our results reflect the interactions of species that actively use the understory. Additional studies using alternative methods are needed to assess the effects of communities in other forest strata and their role in forest regeneration.
Since most bird and bat species dispersed mainly pioneer species that rarely establish in bracken areas, our results suggest that seed dispersal of non-pioneer species is limiting natural regeneration dynamics in bracken-dominated areas. Thus, active restoration techniques are needed, such as planting species that can growth in the presence of bracken, reduce their dominance and facilitate the establishment of non-pioneer species (Toledo-Aceves et al., 2022). Moreover, the installation of perches increases the seed rain and seedling establishment of bird-dispersed species in bracken areas (Mayta et al., 2024). These strategies could help to reestablish seed dispersal of non-pioneer species by large animals in bracken-dominated areas, which is crucial for the maintenance and regeneration of tropical forests.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Direccion General de Biodiversidad, Ministerio de Medio Ambiente y Agua (permit: CAR/MMAYA/VMABCCGDF/DGBAP/MEG 0012/2020). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
CM: Data curation, Formal analysis, Writing – original draft. MS: Formal analysis, Writing – review & editing. CL: Data curation, Writing – review & editing. MV: Data curation, Writing – review & editing. IH: Funding acquisition, Supervision, Writing – review & editing. SG: Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors declare that the financial support for the research and publication was received from the German Research Foundation DFG (grant He 3041/23-1).
Acknowledgments
We thank Noe Taboada and his family, Mauricio Espejo, Miguel Montenegro and Valeria Rojas for field assistance. We also thank Alfredo Fuentes, Emili Jiménez, Rosenber Hurtado, Laura Moya and David Villalba for their help with plant identification. We are also very grateful to the local communities for allowing us to work on their land.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1567678/full#supplementary-material.
References
Aguirre L. F. (2007). Historia natural, distribución y conservación de los murciélagos de Bolivia. Ed. Aguirre L. F. (Santa Cruz: Editorial Centro de Ecología y Difusión Simón I. Patiño).
Aguirre L. F., Vargas A., and Solari S. (2009). Clave de Campo para la Identificación de los de Bolivia. Centro de Estudios en Biología Teórica y Aplicada. (Cochabamba) 38.
Arteaga L. L., Aguirre L. F., and Moya M. I. (2006). Seed rain produced by bats and birds in forest islands in a Neotropical Savanna. Biotropica 38, 718–724. doi: 10.1111/j.1744-7429.2006.00208.x
Barton K. (2023). MuMIn: multi-model inference tools for model selection and averaging. R package version 1. Available at: https://CRAN.R-project.org/package=MuMIn (Accessed December 6, 2024).
Billerman S. M., Keeney B. K., Rodewald P. G., and Schulenberg T. S. (2022). Birds of the world. Cornell laboratory. Available online at: https://birdsoftheworld.org/bow/home (Accessed December 3, 2023).
Bovo A. A. A., Ferraz K. M. P. M. B., Magioli M., Alexandrino E. R., Hasui É., Ribeiro M. C., et al. (2018). Habitat fragmentation narrows the distribution of avian functional traits associated with seed dispersal in tropical forest. Perspect. Ecol. Conserv 16, 90–96. doi: 10.1016/j.pecon.2018.03.004
Brooks M. E., Kristensen K., Van Benthem K. J., Magnusson A., Berg C. W., Nielsen A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.3929/ethz-b-000240890
Camargo P. H. S. A., Pizo M. A., Brancalion P. H. S., and Carlo T. A. (2020). Fruit traits of pioneer trees structure seed dispersal across distances on tropical deforested landscapes: Implications for restoration. J. Appl. Ecol. 57, 2329–2339. doi: 10.1111/1365-2664.13697
Carlo T. A. and Morales J. M. (2016). Generalist birds promote tropical forest regeneration and increase plant diversity via rare-biased seed dispersal. Ecology 97, 1819–1831. doi: 10.1890/15-2147.1
Carlo T. A., Camargo P. H. S. A., and Pizo M. A. (2022). Functional ecology of Neotropical frugivorous birds. Ornithology Res. 30, 139–154. doi: 10.1007/s43388-022-00093-2
Chiarello A. G. (1999). Effects of fragmentation of the Atlantic forest on mammal communities in south-eastern Brazil. Biol. Conserv 89, 71–82. doi: 10.1016/S0006-3207(98)00130-X
Chiu C. H., Chao A., Vogel S., Kriegel P., and Thorn S. (2023). Quantifying and estimating ecological network diversity based on incomplete sampling data. Philos. Trans. R. Soc. B: Biol. Sci. 378. doi: 10.1098/rstb.2022.0183
Cleary K. A., Waits L. P., and Finegan B. (2016). Agricultural intensification alters bat assemblage composition and abundance in a dynamic Neotropical landscape. Biotropica 48, 667–676. doi: 10.1111/btp.12327
Cramer J. M., Mesquita R. C. G., and Bruce Williamson G. (2007). Forest fragmentation differentially affects seed dispersal of large and small-seeded tropical trees. Biol. Conserv 137, 415–423. doi: 10.1016/j.biocon.2007.02.019
Curatola Fernández G. F., Obermeier W. A., Gerique A., López Sandoval M. F., Lehnert L. W., Thies B., et al. (2015). Land cover change in the Andes of southern Ecuador-Patterns and drivers. Remote Sens (Basel) 7, 2509–2542. doi: 10.3390/rs70302509
De La Peña-Domene M., Martínez-Garza C., Palmas-Pérez S., Rivas-Alonso E., and Howe H. F. (2014). Roles of birds and bats in early tropical-forest restoration. PloS One 9, 1–6. doi: 10.1371/journal.pone.0104656
Díaz-B C. A., Otálora-Ardila A., Valdés-Cardona M. C., López-Arévalo H. F., and Montenegro O. L. (2023). Bat functional traits associated with environmental, landscape, and conservation variables in Neotropical dry forests. Front. Forests Global Change 6. doi: 10.3389/ffgc.2023.1082427
Emer C., Galetti M., Pizo M. A., Guimarães P. R., Moraes S., Piratelli A., et al. (2018). Seed-dispersal interactions in fragmented landscapes – a metanetwork approach. Ecol. Lett. 21, 484–493. doi: 10.1111/ele.12909
Emer C., Jordano P., Pizo M. A., Ribeiro M. C., da Silva F. R., and Galetti M. (2020). Seed dispersal networks in tropical forest fragments: Area effects, remnant species, and interaction diversity. Biotropica 52, 81–89. doi: 10.1111/btp.12738
Falconí-López A., Mitesser O., Martin Schaefer H., Blüthgen N., Busse A., Feldhaar H., et al. (2024). Habitat niches of bird species along a recovery gradient in the Chocó tropical forest. Ecol. Indic 166, 1–9. doi: 10.1016/j.ecolind.2024.112260
Farneda F. Z., Meyer C. F. J., and Grelle C. E. V. (2020). Effects of land-use change on functional and taxonomic diversity of Neotropical bats. Biotropica 52, 120–128. doi: 10.1111/btp.12736
Frank H. K., Frishkoff L. O., Mendenhall C. D., Daily G. C., and Hadly E. A. (2017). Phylogeny, traits, and biodiversity of a neotropical bat assemblage: close relatives show similar responses to local deforestation. Am. Nat. 190, 200–212. doi: 10.5061/dryad.mb552
Gallegos S. C., Beck S. G., Hensen I., Saavedra F., Lippok D., and Schleuning M. (2016). Factors limiting montane forest regeneration in bracken-dominated habitats in the tropics. For Ecol. Manage 381, 168–176. doi: 10.1016/j.foreco.2016.09.014
Gallegos S. C., Hensen I., Saavedra F., and Schleuning M. (2015). Bracken fern facilitates tree seedling recruitment in tropical fire-degraded habitats. For Ecol. Manage 337, 135–143. doi: 10.1016/j.foreco.2014.11.003
Gallegos S. C., Mayta C., Villegas M., Ayala G. M., Naoki K., Rechberger J., et al. (2024). Habitat differences in seed-dispersing vertebrates indicate dispersal limitation in tropical bracken-dominated deforested areas. Biotropica 56, e133117. doi: 10.1111/btp.13317
García-Morales R., Moreno C. E., Badano E. I., Zuria I., Galindo-González J., Rojas-Martínez A. E., et al. (2016). Deforestation impacts on bat functional diversity in tropical landscapes. PloS One 11, 1–16. doi: 10.1371/journal.pone.0166765
Gomes L. G. L., Oostra V., Nijman V., Cleef A. M., and Kappelle M. (2008). Tolerance of frugivorous birds to habitat disturbance in a tropical cloud forest. Biol. Conserv 141, 860–871. doi: 10.1016/j.biocon.2008.01.007
González-Varo J. P., Albrecht J., Arroyo J. M., Bueno R. S., Burgos T., Escribano-Ávila G., et al. (2023). Frugivore-mediated seed dispersal in fragmented landscapes: Compositional and functional turnover from forest to matrix. Proc. Natl. Acad. Sci. U S A 120, 1–10. doi: 10.1073/pnas.2302440120
Günter S., Weber M., Erreis R., and Aguirre N. (2007). Influence of distance to forest edges on natural regeneration of abandoned pastures: A case study in the tropical mountain rain forest of Southern Ecuador. Eur. J. For Res. 126, 67–75. doi: 10.1007/s10342-006-0156-0
Haddad N. M., Brudvig L. A., Clobert J., Davies K. F., Gonzalez A., Holt R. D., et al. (2015). Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 1, 1–9. doi: 10.1126/sciadv.1500052
Hardwick K., Healey J., Elliott S., Garwood N., and Anusarnsunthorn V. (1997). Understanding and assisting natural regeneration processes in degraded seasonal evergreen forests in northern Thailand. For Ecol. Manage 99, 203–214. doi: 10.1016/S0378-1127(97)00206-5
Hartig F. (2024). Title residual diagnostics for hierarchical (Multi-level/mixed) regression models. Available online at: https://github.com/florianhartig/DHARMa/issues (Accessed October 21, 2024).
Hartig K. and Beck E. (2003). The bracken fern (Pteridium arachnoideum (Kaulf.) Maxon) dilemma in the Andes of Southern Ecuador. Ecotropica 9, 3–13.
Herzog S. K., Terril R. S., Jahn A. E., Remsen J. V., Maillard O. Z., García-Soliz V. H., et al. (2016). Birds of Bolivia. Field guide (Santa Cruz de la Sierra: Asociación Armonía). Available at: http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm (Accessed June 19, 2024).
Hsieh T. C., Ma K. H., and Chao A. (2016). iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 7, 1451–1456. doi: 10.1111/2041-210X.12613
Huber O., Steyermark J. A., Prance G. T., and Alès C. (1984). The vegetation of the Sierra Parima, Venezuela-Brazil: Some results of recent exploration. Brittonia 36, 104–139. doi: 10.2307/2806619
Jacomassa F. A. F. and Pizo M. A. (2010). Birds and bats diverge in the qualitative and quantitative components of seed dispersal of a pioneer tree. Acta Oecologica 36, 493–496. doi: 10.1016/j.actao.2010.07.001
Jankowski J. E., Kyle K. O., Gasner M. R., Ciecka A. L., and Rabenold K. N. (2021). Response of avian communities to edges of tropical montane forests: Implications for the future of endemic habitat specialists. Glob Ecol. Conserv 30, 1–12. doi: 10.1016/j.gecco.2021.e01776
Jenni L., Leuenberger M., and Rampazzi F. (1996). Capture efficiency of mist nets with comments on their role in the assessment of passerine habitat use. J. Field Ornithol 67, 263–274.
Jones H. H., Bedoya-Durán M. J., Colorado Z G. J., Londoño G., and Robinson S. K. (2023). Dietary and habitat specialization, eye size, clutch size, and aerial lifestyle predict avian fragmentation sensitivity in an Andean biodiversity hotpot. Biodivers Conserv 32, 4057–4081. doi: 10.1007/s10531-023-02682-z
Jordano P. (2000). “Fruits and frugivory,” in Seeds: the ecology of regeneration in plant communities. Ed. Fenner M. (CABI Publishing, Wallingford), 125–165. doi: 10.1079/9780851994321.0125
Jordano P. (2016). Sampling networks of ecological interactions. Funct. Ecol. 30, 1883–1893. doi: 10.1111/1365-2435.12763
Khurana E. and Singh J. S. (2006). Impact of life-history traits on response of seedlings of five tree species of tropical dry forest to shade. J. Trop. Ecol. 22, 653–661. doi: 10.1017/S0266467406003518
Killeen T. J., Siles T. M., Soria L., and Correa L. (2005). Estratificación de vegetación y cambio de uso de suelo en los Yungas y Alto Beni de la Paz. Ecología en Bolivia 40, 32–69.
Klingbeil B. T. and Willig M. R. (2009). Guild-specific responses of bats to landscape composition and configuration in fragmented Amazonian rainforest. J. Appl. Ecol. 46, 203–213. doi: 10.1111/j.1365-2664.2008.01594.x
Laurance W. F. (2005). “The alteration of biotic interactions in fragmented tropical forests,” in Biotic interactions in the tropics: their role in the maintenance of species. Eds. Burslem D., Pinard M. A., and Hartley S. E. (Cambridge University Press, Cambridge), 441–458.
Li W., Zhu C., Grass I., Vázquez D. P., Wang D., Zhao Y., et al. (2022). Plant-frugivore network simplification under habitat fragmentation leaves a small core of interacting generalists. Commun. Biol. 5, 1–10. doi: 10.1038/s42003-022-04198-8
Lippok D., Beck S. G., Renison D., Gallegos S. C., Saavedra F. V., Hensen I., et al. (2013). Forest recovery of areas deforested by fire increases with elevation in the tropical Andes. For Ecol. Manage 295, 69–76. doi: 10.1016/j.foreco.2013.01.011
Lippok D., Beck S. G., Renison D., Hensen I., Apaza A. E., and Schleuning M. (2014). Topography and edge effects are more important than elevation as drivers of vegetation patterns in a neotropical montane forest. J. Vegetation Sci. 25, 724–733. doi: 10.1111/jvs.12132
López C. L., Mayta C., Fuentes A. F., Villegas M., Jiménez E. A., Vasquez V., et al. (2024). Disentangling the roles of bracken fronds and litter on natural seedling recruitment in fire-disturbed tropical montane habitats. For Ecol. Manage 566, 122056. doi: 10.1016/j.foreco.2024.122056
Marjakangas E. L., Abrego N., Grøtan V., de Lima R. A. F., Bello C., Bovendorp R. S., et al. (2020). Fragmented tropical forests lose mutualistic plant–animal interactions. Divers. Distrib 26, 154–168. doi: 10.1111/ddi.13010
Markl J. S., Schleuning M., Forget P. M., Jordano P., Lambert J. E., Traveset A., et al. (2012). Meta-analysis of the effects of human disturbance on seed dispersal by animals. Conserv. Biol. 26, 1072–1081. doi: 10.1111/j.1523-1739.2012.01927.x
Marrs R. H., Le Duc M. G., Mitchell R. J., Goddard D., Paterson S., and Pakeman R. J. (2000). The ecology of bracken: Its role succession and implications for control. Ann. Bot. 85, 3–15. doi: 10.1006/anbo.1999.1054
Martínez-Ramos M., Pingarroni A., Rodríguez-Velázquez J., Toledo-Chelala L., Zermeño-Hernández I., and Bongers F. (2016). Natural forest regeneration and ecological restoration in human-modified tropical landscapes. Biotropica 48, 745–757. doi: 10.1111/btp.12382
Matongera T. N., Mutanga O., Dube T., and Lottering R. T. (2018). Detection and mapping of bracken fern weeds using multispectral remotely sensed data: a review of progress and challenges. Geocarto Int. 33, 209–224. doi: 10.1080/10106049.2016.1240719
Maya-Elizarrarás E. and Schondube J. E. (2015). Birds, cattle, and bracken ferns: bird community responses to a neotropical landscape shaped by cattle grazing activities. Biotropica 47, 236–245. doi: 10.1111/btp.12196
Mayta C., López C. L., Villegas M., Aguirre L. F., Hensen I., and Gallegos S. C. (2024). Bird perches and artificial bat roosts increase seed rain and seedling establishment in tropical bracken-dominated deforested areas. Restor. Ecol. 32. doi: 10.1111/rec.14197
Menezes Pinto Í., Emer C., Cazetta E., and Morante-Filho J. C. (2021). Deforestation simplifies understory bird seed-dispersal networks in human-modified landscapes. Front. Ecol. Evol. 9. doi: 10.3389/fevo.2021.640210
Menke S., Böhning-Gaese K., and Schleuning M. (2012). Plant-frugivore networks are less specialized and more robust at forest-farmland edges than in the interior of a tropical forest. Oikos 121, 1553–1566. doi: 10.1111/j.1600-0706.2011.20210.x
Messina S., Costantini D., Tomassi S., Cosset C. C. P., Benedick S., Eens M., et al. (2021). Selective logging reduces body size in omnivorous and frugivorous tropical forest birds. Biol. Conserv 256, 1–8. doi: 10.1016/j.biocon.2021.109036
Molina-Carpio J., Espinoza D., Coritza E., Salcedo F., Farfán C., Mamani L., et al. (2019). Clima y variabilidad espacial de la ceja de monte y andino húmedo. Ecología en Bolivia 54, 40–56.
Díaz M. M., Solari S., Aguirre Ludmilla L. F., Aguiar M. S., and Barquez R. M. (2016). Clave de Identificación de los Murciélagos de Sudamerica. Tucumán: Programa de conservación de los murciélagos de Argentina. 160.
Morante-Filho J. C., Faria D., Mariano-Neto E., and Rhodes J. (2015). Birds in anthropogenic landscapes: The responses of ecological groups to forest loss in the Brazilian Atlantic forest. PloS One 10, 1–18. doi: 10.1371/journal.pone.0128923
Muscarella R. and Fleming T. H. (2007). The role of frugivorous bats in tropical forest succession. Biol. Rev. 82, 573–590. doi: 10.1111/j.1469-185X.2007.00026.x
Newbold T., Scharlemann J. P. W., Butchart S. H. M., Şekercioğlu Ç.H., Alkemade R., Booth H., et al. (2013). Ecological traits affect the response of tropical forest bird species to land-use intensity. Proc. R. Soc. B: Biol. Sci. 280. doi: 10.1098/rspb.2012.2131
O’Dea N. and Whittaker R. J. (2007). How resilient are Andean montane forest bird communities to habitat degradation? Biodivers Conserv 16, 1131–1159. doi: 10.1007/s10531-006-9095-9
Oosterhoorn M. and Kappelle M. (2000). Vegetation structure and composition along an interior-edge-exterior gradient in a Costa Rican montane cloud forest. For. Ecol. managment 126, 291–307. doi: 10.1016/S0378-1127(99)00101-2
Pavlacky D. C., Possingham H. P., and Goldizen A. W. (2015). Integrating life history traits and forest structure to evaluate the vulnerability of rainforest birds along gradients of deforestation and fragmentation in eastern Australia. Biol. Conserv 188, 89–99. doi: 10.1016/j.biocon.2014.10.020
Quintero E., Isla J., and Jordano P. (2022). Methodological overview and data-merging approaches in the study of plant–frugivore interactions. Oikos 2022, 1–18. doi: 10.1111/oik.08379
Renjifo L. M. (1999). Composition changes in a subandean avifauna after long-term forest fragmentation. Conserv. Biol. 13, 1124–1139. doi: 10.1046/j.1523-1739.1999.98311.x
R Foundation for Statistical Computing (2022). A language and environment for statistical computing. Dendrochronologia (Verona) 26, 115–124. doi: 10.1016/j.dendro.2008.01.002
Roos K., Rollenbeck R., Peters T., Bendix J., and Beck E. (2010). Growth of tropical bracken (Pteridium arachnoideum): response to weather variations and burning. Invasive Plant Sci. Manag 3, 402–411. doi: 10.1614/ipsm-d-09-00031.1
Russell L., Bolker B., Buerkner P., Gine-Vazquez I., Herve M., Jung M., et al. (2024). Estimated marginal means, aka least-squares means. Am. Statistician 34, 1–101. doi: 10.1080/00031305.1980.10483031
Saavedra F., Hensen I., Beck S. G., Böhning-Gaese K., Lippok D., Töpfer T., et al. (2014). Functional importance of avian seed dispersers changes in response to human-induced forest edges in tropical seed-dispersal networks. Oecologia 176, 837–848. doi: 10.1007/s00442-014-3056-x
Saavedra F., Hensen I., and Schleuning M. (2015). Deforested habitats lack seeds of late-successional and large-seeded plant species in tropical montane forests. Appl. Veg Sci. 18, 603–612. doi: 10.1111/avsc.12184
Schleuning M., Blüthgen N., Flörchinger M., Braun J., Schaefer H. M., and Böhning-Gaese K. (2011). Specialization and interaction strength in a tropical plant-frugivore network differ among forest strata. Ecology 92, 26–36. doi: 10.1890/09-1842.1
Sekercioglu C. H. (2012). Bird functional diversity and ecosystem services in tropical forests, agroforests and agricultural areas. J. Ornithol 153, 153–161. doi: 10.1007/s10336-012-0869-4
Sheard C., Neate-Clegg M. H. C., Alioravainen N., Jones S. E. I., Vincent C., MacGregor H. E. A., et al. (2020). Ecological drivers of global gradients in avian dispersal inferred from wing morphology. Nat. Commun. 11, 1–9. doi: 10.1038/s41467-020-16313-6
Snow D. W. (1981). Tropical frugivorous birds and their food plants: A world survey. Biotropica 13, 1–14. doi: 10.2307/2387865
Ssali F., Moe S. R., and Sheil D. (2018). Tree seed rain and seed removal, but not the seed bank, impede forest recovery in bracken (Pteridium aquilinum (L.) Kuhn)-dominated clearings in the African highlands. Ecol. Evol. 8, 4224–4236. doi: 10.1002/ece3.3944
Ssali F., Moe S. R., and Sheil D. (2019). The differential effects of bracken (Pteridium aquilinum (L.) Kuhn) on germination and seedling performance of tree species in the African tropics. Plant Ecol. 220, 41–55. doi: 10.1007/s11258-018-0901-8
Swaine M. D. and Whitmore T. C. (1988). On the definition of ecological species groups in tropical rain forests. Vegetatio 75, 81–86. doi: 10.1007/BF00044629
Tchoumbou M. A., Malange E. F. N., Tiku C. T., Tibab B., Fru-Cho J., Tchuinkam T., et al. (2020). Response of understory bird feeding groups to deforestation gradient in a tropical rainforest of Cameroon. Trop. Conserv Sci. 13, 1–12. doi: 10.1177/1940082920906970
Thiel S., Tschapka M., Heymann E. W., and Heer K. (2021). Vertical stratification of seed-dispersing vertebrate communities and their interactions with plants in tropical forests. Biol. Rev. 96, 454–469. doi: 10.1111/brv.12664
Thompson K. and Rabinowitz D. (1989). Do big plants have big seeds? Am. Nat. 133, 722–728. doi: 10.1086/284947
Toledo-Aceves T., López-Barrera F., Vásquez-Reyes V., and Günter S. (2022). Restoration of tropical montane cloud forest in bracken dominated pastures: The role of nurse shrubs. For Ecol. Manage 508. doi: 10.1016/j.foreco.2022.120055
Valio I. F. M. and Scarpa F. M. (2001). Germination of seeds of tropical pioneer species under controlled and natural conditions. Rev. Bras. Botânica 24, 79–84. doi: 10.1590/S0100-84042001000100009
Vizentin-Bugoni J., Maruyama P. K., Debastiani V. J., Duarte L. da S., Dalsgaard B., and Sazima M. (2016). Influences of sampling effort on detected patterns and structuring processes of a Neotropical plant-hummingbird network. J. Anim. Ecol. 85, 262–272. doi: 10.1111/1365-2656.12459
Wheelwright N. T. (1985). Fruit size, gape width, and the diets of fruit-eating birds. Ecology 66, 808–818. doi: 10.2307/1940542
Whitman A. A., Hagan J. M., and L Brokaw N. V. (1997). A comparison of two bird survey techniques used in a subtropical forest. Condor 99, 955–965. doi: 10.2307/1370146
Wilman H., Belmaker J., Simpson J., de la Rosa C., Rivadeneira M. M., and Jetz W. (2014). EltonTraits 1.0: Species-level foraging attributes of the world’s birds and mammals Ecological Archives E095-178. Ecology 95, 2027. doi: 10.1890/13-1917.1
Keywords: forest regeneration, frugivory, mist nets, non-pioneer, pioneer, Pteridium
Citation: Mayta C, Schleuning M, López CL, Villegas M, Hensen I and Gallegos SC (2025) Differences in bird and bat traits, seed-dispersal interactions and functions between tropical montane forest and bracken-dominated areas. Front. Ecol. Evol. 13:1567678. doi: 10.3389/fevo.2025.1567678
Received: 27 January 2025; Accepted: 04 June 2025;
Published: 20 June 2025.
Edited by:
Pedro Gonçalves Vaz, University of Lisbon, PortugalReviewed by:
Marco Aurélio Pizo, São Paulo State University, BrazilOscar Enrique Murillo-García, Universidad Del Valle, Colombia
Copyright © 2025 Mayta, Schleuning, López, Villegas, Hensen and Gallegos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cesar Mayta, Y2VzYXJtYXl0YXJvY2FiYWRvQGdtYWlsLmNvbQ==