 Gwendolyn Peyre1*
Gwendolyn Peyre1* Enrique Zuñiga1
Enrique Zuñiga1 Andrea Cornejo Paredes2Aremi Paja Medina2
Andrea Cornejo Paredes2Aremi Paja Medina2 Jose Alfredo Sulla Torres2
Jose Alfredo Sulla Torres2 Manuel Zúñiga Carnero2
Manuel Zúñiga Carnero2 Karina Rosas Paredes2
Karina Rosas Paredes2- 1Department of Civil and Environmental Engineering, University of the Andes, Bogota, Colombia
- 2School of System Engineering, Catholic University Santa Maria, Arequipa, Peru
Andean treelines are sensitive ecotones that are being increasingly threatened by both the climatic and anthropogenic facets of global change. Although they have received substantial scientific focus over the last decades, we still miss a comprehensive socio-ecological overview at the continental scale. This mini review aims to compile existing research on Andean treelines and offer a synthetic outlook on their geography, as well as their environmental and human drivers. We conducted an extensive search of scientific literature focused principally on ISI Web of Knowledge sources and spanning over the last two decades. It is established that Andean treelines vary in elevation according to their position in the cordillera, mostly due to mountain mass effects and latitudinal gradients, and the local topography. Temperature and moisture availability are significant drivers shaping Andean treelines, but seasonality and extreme events gain in importance with increasing latitude and climate change. Moreover, human influence has transformed treeline areas for millenia, by affecting their position, composition and ecological functions. Agriculture, livestock grazing and fires are primarily responsible for stalling many Andean treelines in their upslope migration today. This mini-review sheds light on the current state of Andean treelines today and suggests future pathways with anticipated opportunities for conservation and sustainable restoration. For instance, we stress the need for future research in species and community modelling to predict composition and distribution changes in forest and non-forest vegetation under global change scenarios.
1 Introduction
The 7,500 km Andes form the longest continuous mountain chain in the world and constitute an El Dorado of natural and cultural resources. Despite showing striking North South differences, in terms of orogeny, climates and land-use among others, the Andes offer a unique perspective to study broad latitudinal mountain gradients. In this review, we investigate the socio-ecological aspects of Andean treelines, which represent the uppermost frontier for tree dominance in the landscape, separating forests from non-forests. Because of their ecological features, treelines make an excellent indicator for climate, such as minimum temperatures and snow cover, but also for biodiversity thresholds, for instance functional diversity and ecosystem complexity (Körner, 2021a). Treelines are one of the most at-risk ecotones, because the ecological integrity of their neighbouring ecosystems might not allow for successful species shifts and community recompositions at the pace of climate change. Either forests will take over non-forests, reducing them, or they will suffer biodiversity losses in the failed attempt to migrate swiftly. Under global change, the structural and functional ecology of treelines will be challenged, which in turn will likely affect human livelihoods.
Mountain treelines refer to the natural shift between montane forests and alpine ecosystems, characterized by lower vegetation such as grasslands or shrublands (Körner, 2021a). Nowadays, most treelines are in fact timberlines, which display similar ecotonal characteristics to natural treelines but typically result from complex land-use history (Paulsen and Körner, 2014; Testolin R et al., 2020). In this review and for simplicity reasons, we will refer to both natural treelines and timberlines under the common name treeline. Treelines can be categorized according to their tree dominance, for example single-dominant tree species versus multi-species, gymnosperm versus angiosperm. They can also be classified by their overall shape between forest and the adjacent ecosystem: i) abrupt, or a sharp ecotone, ii) diffuse, or a gradual transition, and iii) krummholz, or a scattering landscape combining young and old small trees (Bader et al., 2021). In the present study, we will characterize Andean treelines, assess the role of natural and human factors in shaping them today and finally discuss their evolution under global change.
2 Mountain geography
Andean treelines vary immensely in elevation along the latitudinal gradient. According to previous studies, they typically increase from 500 to 1500 m in the Patagonian Andes (Lat. 55–40°S), then gradually progress from 2000 to 3000 m in the Mediterranean Andes (Lat. 40–20°S), peak around 3500–4000 m in the Central Andes (Lat. 20-5°S) and decrease to 3000–3500 m in the Northern Andes (Lat. 5°S–10°N) (Lara et al., 2005; Luebert and Pliscoff, 2018; Peyre et al., 2021; Peyre et al., 2022; Kienle et al., 2023) (Figure 1).
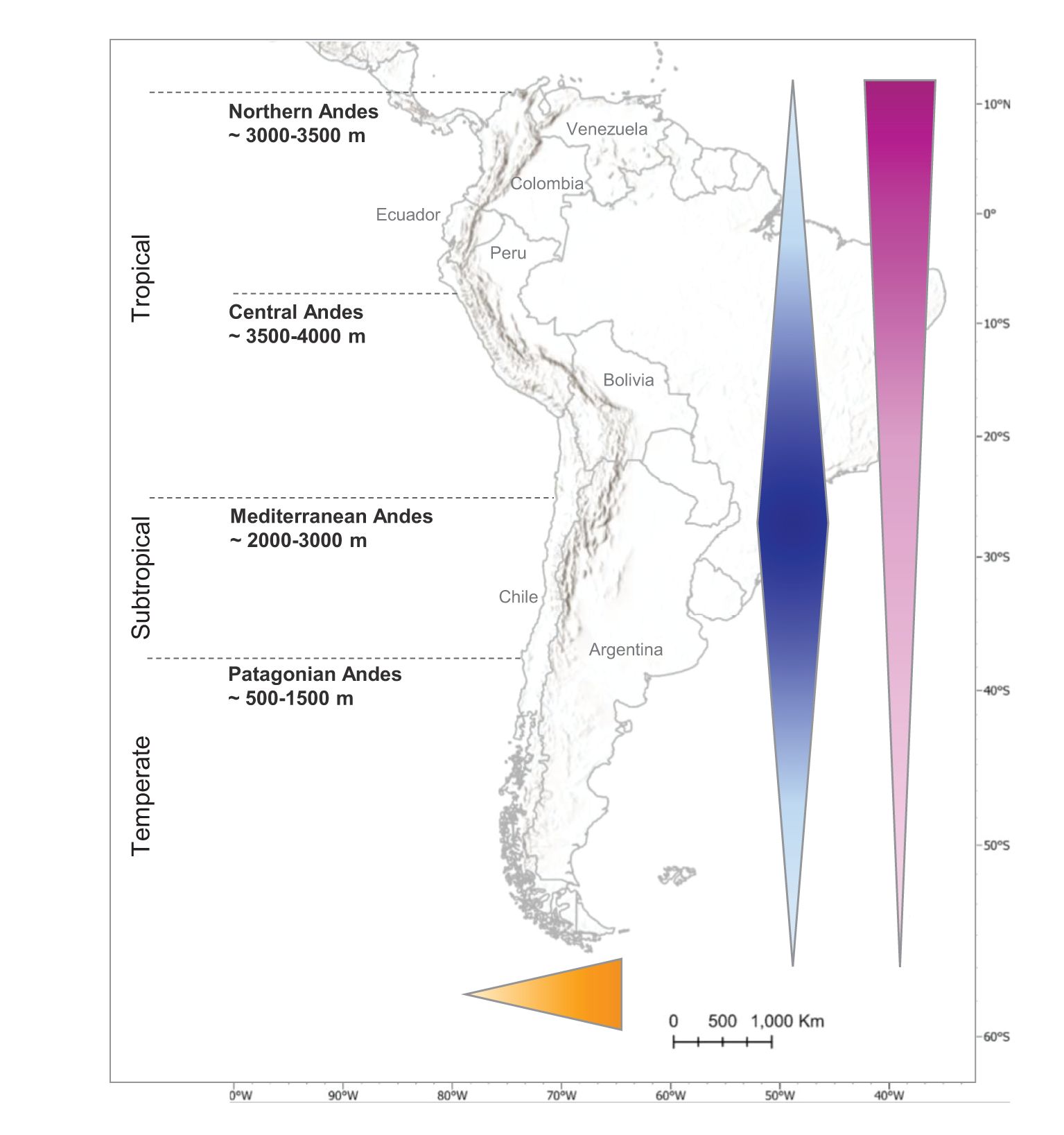
Figure 1. Andean geography and treeline elevation, influenced by two latitudinal (blue) and longitudinal (orange) mountain mass effects, as well as the temperate-tropical gradient (pink). Increasing treeline elevation is represented by a progressively larger and darker arrow.
This latitudinal hump pattern is mostly due to the mountain mass effect, a phenomenon that describes the upward shift of thermoclines, and therefore vegetation belts, toward the centre of a mountain cordillera (Körner, 2021a; Kienle et al., 2023). This effect usually increases with the spatial extent of a cordillera, which explains why it is maximized in the Andes. It is caused by increased continentality around the cordillera’s mid-point, leading to reduced cloudiness, air mass advection and overall higher temperatures, which combined, create thermal advantages for tree species (He et al., 2016; Irl et al., 2016; Kienle et al., 2023). In fact, the Andean Altiplano, forming a large high-elevation plateau circa 3500–4000 m from northern Chile to southern Peru, acts as a baseline for towering mountain peaks that present some of the highest treelines in the world. Indeed, the Polylepis forests found at 4800 m on the Sajama volcano in Bolivia are only surpassed by Juniperus treelines in Tibet (4900 m) (Kessler et al., 2014). The observed asymmetry between the Northern and Mediterranean Andes can be attributed to the Tropical-Temperate general gradient, pushing vegetation belts up in the tropics while depressing them towards higher latitudes. At the scale of the Tropical Andes and looking at nearly natural treelines only, Zelazowski et al. (2023). evidenced a clear latitudinal gradient in treeline elevation on the Amazonian side of the cordillera, with a 30–60 m increase per geographic degree towards the equator.
Another consistent mountain mass effect occurs along the longitudinal gradient, with increased continentality with distance from the coastline (Irl et al., 2016; He et al., 2023; Kienle et al., 2023). As a result, coastal mountains are usually smaller and showcase lower treelines than their inland counterparts at similar latitudes. For example, Peyre et al. (2021) observed a mean treeline at 3620 m in the western mountain chain and an equivalent 3710 m in the central chain of Colombia. When the cordillera is not subdivided longitudinally, but instead forms a high elevation plateau, such as the Altiplano, its peaks tower over said plateau, which in turn leads to rising isotherms and higher treelines (He et al., 2016).
At the local scale, geomorphology plays a significant role in driving treeline position and shape. Steep slopes, rugged terrain and unstable land-forms prone to mass movements can create topographic barriers for trees. Therefore, tree populations will only successfully establish and remain long-term confined in sheltered sites. Several authors have pointed out that Andean forests forming treelines are more likely found in valleys, protected from the natural geomorphologic disturbances on slopes, especially in the tropics (Bader and Ruijten, 2008; Toivonen et al., 2017; Urquiaga-Flores et al., 2024).
3 The role of the environment
The combined effect of the macro- and microclimates significantly conditions the survival, growth and reproduction capacities of tree species at the treeline (Hansson et al., 2021; Körner, 2021b). First and foremost, mountain ecologists acknowledge that natural treeline elevation is congruent with the 6°C isotherm and 94 growth days on average worldwide (Körner and Paulsen, 2004; Körner, 2021a). This primary physiological limitation is based on the interplay between low temperatures and carbon-use, specifically trade-offs between photosynthetic carbon, carbon available for tree growth, and carbon loss through respiration. Two prevailing hypotheses, the Growth Limitation Hypothesis and the Carbon Limitation Hypothesis, have received important scientific support today, although the former seemingly applies best to treelines (Schenker et al., 2014).
Despite many Andean treeline studies concurring with the global temperature average, there are several notable exceptions. On the one hand, Polylepis forests, remnants of dominant montane forests in the Tropical Andes of the Holocene, are usually found forming fragmented forests at high elevation today. It is not uncommon to see Polylepis treelines experience superficial temperatures lower than the global threshold, for instance i) Bendix and Rafiqpoor (2021) found forests with mean soil temperatures of the growing season of 4.5–6.0°C at 4000–4100 m in Ecuador, ii) Hoch and Körner (2005) reported others circa 4.7–5.4°C at 4810 m in Bolivia, and iii) Kessler et al. (2014) encountered treelines at 4.6–5°C at 4330–4450 m in southern Peru. Moreover, these temperatures come dangerously close to the critical thermal threshold of 5°C where biological processes and therefore tree growth become critically hindered (Körner and Paulsen, 2004; Holtmeier and Broll, 2019). On the other hand, many subtropical treelines of the Mediterranean Andes experience significantly higher temperatures than the global threshold, for example, Piper et al. (2016) reported an 8.7°C mean temperature for the growing season at several Nothofagus pumilio treelines in central Chile. Interestingly, treelines of the same species under temperate conditions in the Patagonian Andes typically experience temperatures around 6.8°C, hence closer to the global average (Fajardo and Piper, 2014). Both air and soil temperatures are decisive factors for tree growth at the treeline, because there can be striking differences at the ground and canopy levels. In fact, while their average values are chiefly comparable, around 6–7°C worldwide (Körner and Paulsen, 2004), the canopy level undergoes much higher variation and extremes than the ground level at both the daily and annual scales. These important variations, sometimes up to 20°C on the same day, can create stressful thermal conditions for the sensitive tree photosynthetic tissues (Körner and Paulsen, 2004; Richardson and Friedland, 2009).
In the humid Tropical Andes, treelines are usually comprised within the cloudy lower condensation belt, and therefore water availability is not a limiting factor (Halladay et al., 2012; Helmer et al., 2019). The protective cloud cover does not only prevent desiccation but also filters solar radiation, allowing plants to remain productive and grow tall. By contrast in the dry Tropical Andes and Mediterranean Andes, Andean forests must form close canopies, to prevent wind and solar damage and create a relatively stable microclimate for their understorey species (Körner and Paulsen, 2004; Richardson and Friedland, 2009). Many areas constrained by high evaporation and low precipitation rates, often associated with continentality, exhibit semi-arid to arid tendencies (Santini et al., 2024). The resulting treeline is often discontinuous or absent, presenting as scattered dry forests, open xeric shrublands or sparse grasslands with cacti such as Austrocedrus (Barichivich et al., 2009). In the latter case, elevation shifts in cacti distribution and composition can indicate a smoother transition from the montane to alpine belts in lieu of a treeline (obs. pers.). Despite the precipitation pattern inverting and generally increasing in the Patagonian Andes, several studies mention moisture availability and drought events as factors significantly correlated with tree growth and seedling establishment at Nothofagus treelines (Daniels and Veblen, 2004; Srur et al., 2018).
Climatic seasonality and extremes can play a key role in shaping treeline structure, although it affects tropical, subtropical and temperate treelines differently (Rehm and Feeley, 2015; He et al., 2023). Tropical treelines typically lack snow and exhibit little yearly variability in temperature and precipitation; however, strong diurnal climatic oscillations prevail. As previously established for the humid Tropical Andes, the protective cloud cover encompassing the treeline usually buffers these fluctuations and allows trees to maintain their productivity (Rehm and Feeley, 2015). However in the tropical dry Andes, these climatic oscillations can significantly impede tree survival and growth, leading to drastic anatomical and physiological plant adaptations (Young and Leon, 2007). For instance, studies show that night frost events can severely damage plant tissue and reduce tree recruitment at Andean treelines (Bader et al., 2021). Subtropical and especially temperate Andean treelines experience pronounced seasonal fluctuations in temperature and moisture availability at the yearly level. It becomes critical in the Mediterranean Andes, if a treeline receives less than 450 mm of rain during the cold season (He et al., 2023). Under long-seasonal temperature and/or water stresses can induce a state of dormancy in treeline trees and their seeds (Körner, 2021a). In case snow is present, it can create a superficial local buffer for microclimatic variations, protecting both the seedbank and seedlings until changing seasons. Combining these environmental factors helps explain the differential slope effect, where treeline position is conditioned by exposition-related characteristics. In the Andean context, treelines are typically higher on the shade slope at tropical latitudes, and on the sun slope at temperate latitudes (Hansson et al., 2021). In contrast, treelines exposed to more sun exposure at tropical latitudes, and more freeze and snow stresses at temperate latitudes, tend to display lower elevations.
4 Human landscapes
Millenia of human occupation and activities have significantly contributed to shape Andean treelines today by affecting their plant composition, structure and elevation (Sarmiento and Frolich, 2002; Young and Leon, 2007; Young et al., 2017).
Going back to the Mid Holocene, dry cold conditions could have helped alpine grasslands expand downslope to the detriment of forests, in part thanks to the spread of natural fires (Valencia et al., 2010; Bush et al., 2015). Human influence in the Andes was reported as far as 6000 yr BP in the Central Andes, where it would have increased the fire pressure and lowered alpine puna grasslands even further (Urrego et al., 2011). The following wetter and warmer climatic conditions of the Late Holocene should have favoured forests over grasslands, leading to a progressive upheaval of the treeline. However, from 4000–3000 yr BP, as human population increased and progressively sedentarized in parts of the Tropical Andes, it likely kept treeline from migrating upslope through grazing and fires (Young and Leon, 2007; Di Pasquale et al., 2008; Valencia et al., 2010).
As civilizations developed, more substantial and permanent human settlements established in the Tropical and Subtropical high Andes (Aldenderfer, 2008; Osorio et al., 2017). For example, the Aymaras and Incas were predominantly inhabiting alpine areas, such as the Titicaca lake surroundings, using punas for agricultural purposes with high-elevation crops, such as quinoa and potatoes, and small-scale livestock herding, including llama and alpaca. The long occupation and extensive use of these areas led to a gradual destruction of the local treeline, and a progressive aridification of high-elevation ecosystems in the dry Central and Mediterranean Andes. Meanwhile in the humid Tropical Andes, human settlements moved to the montane belt to escape tropical diseases and take advantage of the mild climatic conditions and fertile soils (Sarmiento and Frolich, 2002). The treeline and páramo were mostly used for agriculture and farming, with human occupation concentrating below, like the Muisca established on the Cundiboyacense plateau in Eastern Colombia. Finally, the Patagonian Andes were more scarcely occupied and local populations such as the Mapuche, favoured valleys and plain areas for permanent settlements, carrying out seasonal activities upslope.
The Andean colonization by Europeans led to the forced abandonment of many high-elevation settlements and activities, which in turn meant a progressive and secondary forest growth and a consequent upheaval of the treeline (Urrego et al., 2011; Bush et al., 2015). This trend then reversed and accelerated during the last century, as the human population increased and consequently its food and water needs. Andean landscapes have undergone unparalleled transformations since the 1950s, leading to severe biodiversity loss and degradation of ecosystem services in certain areas (Young and Leon, 2007; Rehm and Feeley, 2015). Treelines subject to historic land-use have been probably depressed hundreds of meters from their natural position today (Sarmiento and Frolich, 2002; Young et al., 2017). Many authors sustain that the historic treeline of the Tropical Andes was dominated by Polylepis forests, which were progressively cleared and fragmented (Urrego et al., 2011; Kessler et al., 2014). Today, these forests are rarely extended and instead occupy steep valleys and rocky outcrops that are difficulty attainable by fire and livestock (Toivonen et al., 2017). These findings suggest that land-use not only affected the treeline position, but also its structural shape and species composition (Young et al., 2017). It is likely that naturally diffuse treelines became either i) abrupt; ii) reforested krummholz; or ii) newly diffuse and dominated by disturbance-prone tree species such as Weinmannia and Myrsine (Bush et al., 2015; Oliveras et al., 2018).
Agriculture is vastly expanding and intensifying in many areas today, with grave environmental consequences on ecosystem destruction and pollution, but also biotic invasions (IPBES, 2019; Peyre, 2021). For example, the flower, fruit and potatoe extensive crops on the Cundiboyacense plateau subject the montane-alpine transition of their surrounding mountains to severe insect outbreaks and plant invasions, such as Ulex europaeus L. originally planted for crop fencing (Hernández et al., 2014; Peyre, 2021). The grazing of livestock is, besides agriculture, the most prevalent activity around Andean treelines and so regardless of climatic and soil conditions (Cierjacks et al., 2008; Rehm and Feeley, 2015). When livestock grazes and tramples in Andean forests, it can severely i) affect their understorey composition by selecting grazed species and bringing in exotics; as well as ii) limit tree recruitment by removing seedlings (Cierjacks et al., 2008; Urquiaga-Flores et al., 2024). A common practice to improve grazing in alpine grasslands just above the treeline consists of periodic burning to force the resprouting of palatable herbaceous leaves, remove woody vegetation and accelerate biomass cycles (Román-Cuesta et al., 2011; Bush et al., 2015). Despite few treelines being cleared for timber today, they often undergo drastic deforestation to expand livestock pastures and crops.
Treeline deforestation usually involves fire, especially if manual clearing is difficulted by the ambient moisture and high amounts of understorey vegetation (Young and Leon, 2007; Bush et al., 2015). When burning starts in neighbouring grasslands, it can affect the treeline margin by removing the protective edge and leaving the forest core open (Oliveras et al., 2018). Fire initiated inside the forest can produce severe torching anoxic episodes that affect the entire understorey (Román-Cuesta et al., 2011). The poor resilience capacity of montane Andean forests suggests long reforestation pathways for secondary succession after burning (Di Pasquale et al., 2008; Rehm and Feeley, 2015; Bader et al., 2021). Finally, a recently observed and potentially harmful trend is the use of herbicides, meant for agriculture, to purposedly depress the treeline in remote access areas (obs. pers.). This practice has been seen on multiple occasions outside protected areas in the Colombian Andes, and suggests negative consequences on both the forest biodiversity and the wellbeing of the dependent human population. However, further study is required to assess the extent and impact of this particular practice throughout the Andes.
5 Discussion
Climate change is causing rapid changes in the distribution, composition and functions of ecosystems worldwide, and its impacts are being amplified by the galloping human demographics and land-system changes (Nogués-Bravo et al., 2008; IPBES, 2019). The diverse Andean landscapes are at the forefront of environmental challenges and threatened with potentially critical socio-ecological impacts in the near future (Table 1).
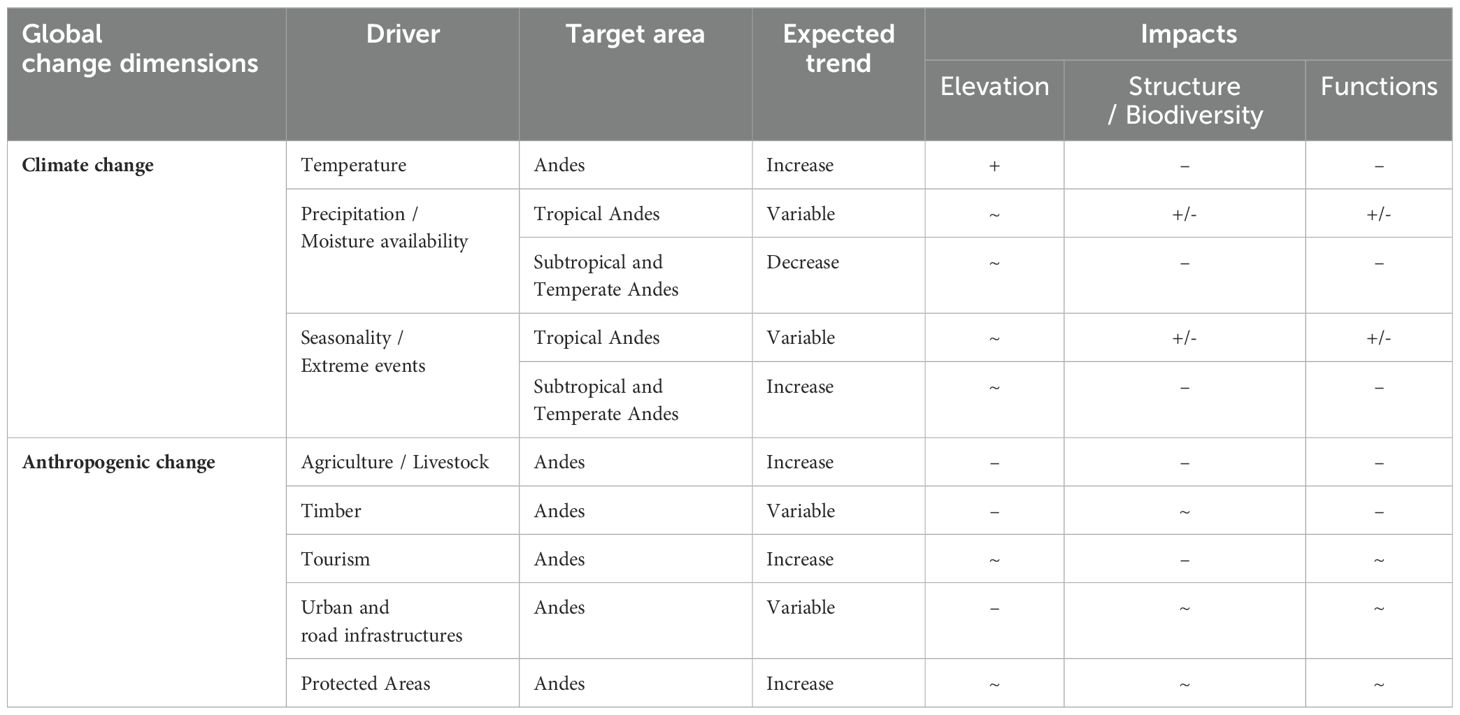
Table 1. Expected Global Change trends and impacts on the elevation (increase +, decrease -, stable ~), and ecological structure and functions (positive +, negative -, stable ~) of Andean treelines in the 21st century.
Observed trends - Global studies report a majority of treelines advancing during the 20th century, 50% according to Harsch et al (Harsch et al., 2009), and 66% according to Hansson et al (Hansson et al., 2021), although the Andes were underrepresented in both datasets. Recently, He et al (He et al., 2023). found that 70% of their studied treelines advanced at a mean rate of 1.2 m/yr between 2000–2010 alone worldwide and 3.1 m/yr in the tropics. This last finding merits further study as it contradicts the global theory that multi-species treelines (usually found in the tropics) respond slower and in a more heterogeneous manner to environmental change than single-species treelines (Harsch et al., 2009; Hansson et al., 2023). Today, evidence of Andean treeline responses to global change remains scarce, as well as geographically and methodologically heterogeneous. Examples of regional studies include i) Chacón-Moreno et al (Chacón-Moreno et al., 2021), who observed a mean 1.3 m/yr upslope migration in Venezuela since the 50s; and ii) Feeley et al (Feeley et al., 2021), who reported a 2.5–3.5 m/yr progress over 40 years in southern Peru.
Future challenges and opportunities - Andean temperatures are predicted to increase up to 5 °C during the 21st century, with the fastest warming occurring in the Central Andes (Pabón-Caicedo et al., 2020). As a result, most montane forests will undergo important thermal stress and will have to move upslope hundreds of meters to track isotherms, but very heterogeneous responses are expected (Nogués-Bravo et al., 2008; Fadrique et al., 2018; Feeley et al., 2020). Prediction for precipitation patterns in the Tropical Andes are highly variable and therefore difficult to summarize (Pabón-Caicedo et al., 2020), nevertheless, studies show cloud cover should decrease overall, which in turn will reduce their protective buffer of treeline trees and seedlings (Halladay et al., 2012; Helmer et al., 2019). For treelines strongly conditioned by moisture availability, such as many Nothofagus forests, the reduced overall precipitation and prolonged drought events will undoubtedly affect tree survival, growth and migration (Daniels and Veblen, 2004; Srur et al., 2018; Santini et al., 2024). Finally, another concern associated with increased drought events is that they are already affecting mainly humid areas, for instance in the eastern Tropical Andes along the Amazonian biome (Lewis et al., 2011). The ecological impact of these droughts on the poorly-adapted treeline biota in these areas calls for urgent study to prevent dramatic biodiversity losses.
Interestingly, Lutz et al (Lutz et al., 2013). measured the speed of treeline migration in southern Peru and compared it with core forest migration rates and the optimal speed to track climate change. Their findings showed that treelines were advancing at 0.5–2.3% of the optimal speed and 12.5–110 times slower than the core forest. This significant stalling in treeline migration refers to the grass ceiling effect and underpins the crucial importance of local biological and micro-environmental factors (Rehm and Feeley, 2015). This effect is caused by intrinsic tree species survival, growth and reproductive capacities, but mostly by the availability of suitable undisturbed sites for germination and establishment. For example, studies show that diffuse treelines might migrate faster than abrupt ones because the gradually smaller vegetation provides protective shade and moisture for seedlings (Harsch et al., 2009; Bader et al., 2021). In contrast, seedlings in abrupt transitions often present high mortality rates that can be attributed to photodamage and harsh microclimates, including wind exposure and damaging frost events (Richardson and Friedland, 2009; McIntire et al., 2016), It is also worth noting the role of biotic interactions in future treeline dynamics. For instance, dominant grasses can nurse tree seedlings by providing shade but simultaneously compete for space and available moisture (Rehm and Feeley, 2015; Bader et al., 2021). Moreover, the striking turnover in insect and bird communities at the treeline may not facilitate plant dispersal into grasslands, especially when strictly dependent plant-animal interactions are at play (Arroyo et al., 2006; Altamirano et al., 2020).
Finally, several authors emphasize the primary role of conservation strategies such as protected areas in a global change context (Lutz et al., 2013; Urquiaga-Flores et al., 2024). Despite many treelines remain (relatively) pristine in remote-access areas, most unprotected treelines today are either experiencing higher human pressure or land abandonment that can lead to secondary successions into non-analog forests (Young and Leon, 2007; Tovar et al., 2022). Under certain circumstances, ecological restoration could help counteract past human impacts on affected treelines and ease the recovery of their structure and function under a changing climate. This aspect would be particularly relevant for climate regulation, and carbon storage in particular, during the grassland to forest, soil to biomass, transition in tropical humid areas (Gibbon et al., 2010). In protected areas can use active and passive methods to effectively manage treelines and allowing them to migrate naturally under climate change, even though it might still engender carbon unbalance and habitat simplification (Michel et al., 2023). Towards that goal, predictive tools such as species and community models can play a crucial role in anticipating future distribution and composition changes, and provide useful evidence for protected area representation and future planning (Fajardo et al., 2023). To conclude, we call for more in-depth assessments of treeline dynamics and future projections at the cordillera level, and stress the need for both transformative change and sustainable conservation in Andean treeline areas.
Author contributions
GP: Conceptualization, Funding acquisition, Investigation, Resources, Writing – original draft, Writing – review & editing. EZ: Conceptualization, Writing – review & editing. ACP: Conceptualization, Writing – review & editing. APM: Conceptualization, Writing – review & editing. JAST: Conceptualization, Writing – review & editing. MZC: Writing – review & editing, Conceptualization. KRP: Conceptualization, Funding acquisition, Project administration, Resources, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors would like to thank the Research Dean Office of the Catholic University Santa Maria for their financial support with the grant 29002-R-2022.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aldenderfer M. S. (2008). “High elevation foraging societies,” in The handbook of South American archaeology. Ed. Silverman H. (Springer, New York), 131–143.
Altamirano T. A., de Zwaan D. R., and Ibarra J. T. (2020). Treeline ecotones shape the distribution of avian species richness and functional diversity in south temperate mountains. Sci. Rep. 10, 18428. doi: 10.1038/s41598-020-75470-2
Arroyo M. T., Muñoz M. S., Henríquez C., Till-Bottraud I., and Pérez F. (2006). Erratic pollination, high selfing levels and their correlates and consequences in an altitudinally widespread above-tree-line species in the high Andes of Chile. Acta Oecol. 30, 248–257. doi: 10.1016/j.actao.2006.05.006
Bader M. Y., Llambí L. D., Case B. S., Buckley H. L., Toivonen J. M., Camarero J. J., et al. (2021). A global framework for linking alpine-treeline ecotone patterns to underlying processes. Ecography 44, 265–292. doi: 10.1111/ecog.05285
Bader M. Y. and Ruijten J. J. (2008). A topography-based model of forest cover at the alpine tree line in the tropical Andes. J. Biogeogr. 35, 711–723. doi: 10.1111/j.1365-2699.2007.01818.x
Barichivich J., Sauchyn D. J., and Lara A. (2009). Climate signals in high elevation tree-rings from the semiarid Andes of north-central Chile: responses to regional and large-scale variability. Palaeogeogr Palaeoclimatol Palaeoecol. 281, 320–333. doi: 10.1016/j.palaeo.2007.10.033
Bendix J. and Rafiqpoor M. D. (2021). Studies on the thermal conditions of soils at the upper tree line in the páramo of papallacta:(Eastern cordillera of Ecuador). Erdkunde. 55, 257–276. doi: 10.3112/erdkunde.2001.03.04
Bush M. B., Alfonso-Reynolds A. M., Urrego D. H., Valencia B. G., Correa-Metrio Y. A., Zimmermann M., et al. (2015). Fire and climate: contrasting pressures on tropical Andean timberline species. J. Biogeogr. 42, 938–950. doi: 10.1111/jbi.12470
Chacón-Moreno E., Rodríguez-Morales M., Paredes D., Suárez del Moral P., and Albarrán A. (2021). Impacts of global change on the spatial dynamics of treeline in Venezuelan Andes. Front. Ecol. Evol. 9. doi: 10.3389/fevo.2021.615223
Cierjacks A., Rühr N. K., Wesche K., and Hensen I. (2008). Effects of altitude and livestock on the regeneration of two tree line forming Polylepis species in Ecuador. Plant Ecol. 194, 207–221. doi: 10.1007/s11258-007-9285-x
Daniels L. D. and Veblen T. T. (2004). Spatiotemporal influences of climate on altitudinal treeline in northern Patagonia. Ecology 85, 1284–1296. doi: 10.1890/03-0092
Di Pasquale G., Marziano M., Impagliazzo S., Lubritto C., De Natale A., and Bader M. Y. (2008). The Holocene treeline in the northern Andes (Ecuador): first evidence from soil charcoal. Palaeogeogr Palaeoclimatol Palaeoecol. 259, 17–34. doi: 10.1016/j.palaeo.2006.12.016
Fadrique B., Báez S., Duque Á, Malizia A., Blundo C., Carilla J., et al. (2018). Widespread but heterogeneous responses of Andean forests to climate change. Nature 564, 207–212. doi: 10.1038/s41586-018-0715-9
Fajardo A. and Piper F. I. (2014). An experimental approach to explain the southern Andes elevational treeline. Am. J. Bot. 101, 788–795. doi: 10.3732/ajb.1400166
Fajardo J., Lessmann J., Devenish C., Bonaccorso E., Felicísimo A. M., Fernando J. M., et al. (2023). The performance of protected-area expansions in representing tropical Andean species: past trends and climate change prospects. Sci. Rep. 13, 966. doi: 10.1038/s41598-022-27365-7
Feeley K., Martinez-Villa J., Perez T., Silva Duque A., Triviño Gonzalez D., and Duque A. (2020). The thermal tolerances, distributions, and performances of tropical montane tree species. Front. For Glob Change 3. doi: 10.3389/ffgc.2020.00025
Feeley K. J., Silman M. R., Bush M. B., Farfan W., Cabrera K. G., Malhi Y., et al. (2021). Upslope migration of Andean trees. J. Biogeogr. 38, 783–791. doi: 10.1111/j.1365-2699.2010.02444.x
Gibbon A., Silman M. R., Malhi Y., Fisher J. B., Meir P., Zimmermann M., et al. (2010). Ecosystem carbon storage across the grassland–forest transition in the high andes of manu national park, Peru. Ecosystems 13, 1097–1111. doi: 10.1007/s10021-010-9376-8
Halladay K., Malhi Y., and New M. (2012). Cloud frequency climatology at the Andes/Amazon transition: 1. Seasonal and diurnal cycles. J. Geophys Res. 117, D23102. doi: 10.1029/2012JD017770
Hansson A., Dargusch P., and Shulmeister J. (2021). A review of modern treeline migration, the factors controlling it and the implications for carbon storage. J. Mount Sci. 18, 291–306. doi: 10.1007/s11629-020-6221-1
Hansson A., Shulmeister J., Dargusch P., and Hill G. (2023). A review of factors controlling Southern Hemisphere treelines and the implications of climate change on future treeline dynamics. Agric. For Meteorol. 332, 109375. doi: 10.1016/j.agrformet.2023.109375
Harsch M. A., Hulme P. E., McGlone M. S., and Duncan R. P. (2009). Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 12, 1040–1049. doi: 10.1111/j.1461-0248.2009.01355.x
He X., Jiang X., Spracklen D. V., Holden J., Liang E., Liu H., et al. (2023). Global distribution and climatic controls of natural mountain treelines. Glob Change Biol. 29, 7001–7011. doi: 10.1111/gcb.16885
He W., Zhang B., Zhao F., Zhang S., Qi W., Wang J., et al. (2016). The mass elevation effect of the central Andes and its implications for the Southern Hemisphere’s highest treeline. Mt Res. Dev. 36, 213–221. doi: 10.1659/MRD-JOURNAL-D-15-00027
Helmer E. H., Gerson E. A., Baggett L. S., Bird B. J., Ruzycki T. S., and Voggesser S. M. (2019). Neotropical cloud forests and páramo to contract and dry from declines in cloud immersion and frost. PloS One 14, e0213155. doi: 10.1371/journal.pone.0213155
Hernández C. L., Fuentes L. S., Fajardo G. E., and Matthews D. L. (2014). A new species of Oidaematophorus (Lepidoptera: Pterophoridae) from Chingaza National Natural Park in Colombia. Trop. Lepid Res. 24, 15–21.
Hoch G. and Körner C. (2005). Growth, demography and carbon relations of Polylepis trees at the world’s highest treeline. Funct. Ecol. 19, 941-951. doi: 10.1111/j.1365-2435.2005.01040.x
Holtmeier F. K. and Broll G. (2019). Treeline research—From the roots of the past to present time. A review. Forests 11, 38. doi: 10.3390/f11010038
IPBES (2019). Global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (Bonn: UN Environment), 1148.
Irl S. D., Anthelme F., Harter D. E., Jentsch A., Lotter E., Steinbauer M. J., et al. (2016). Patterns of island treeline elevation–a global perspective. Ecography 39, 427–436. doi: 10.1111/ecog.01266
Kessler M., Toivonen J. M., Sylvester S. P., Kluge J., and Hertel D. (2014). Elevational patterns of Polylepis tree height (Rosaceae) in the high Andes of Peru: role of human impact and climatic conditions. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00194
Kienle D. R., Irl S. D., and Beierkuhnlein C. (2023). Mass elevation effect and continentality have a stronger impact on global treelines than spatial isolation. Glob Ecol. Biogeogr. 32, 1087–1097. doi: 10.1111/geb.13689
Körner C. (2021a). “Alpine treelines,” in Alpine plant life. Ed. Körner C. (Springer Cham, Basel), 141–173.
Körner C. (2021b). The cold range limit of trees. Trends Ecol. Evol. 36, 979–989. doi: 10.1016/j.tree.2021.06.011
Körner C. and Paulsen J. A. (2004). A world-wide study of high altitude treeline temperatures. J. Biogeogr. 31, 713–732. doi: 10.1111/j.1365-2699.2003.01043.x
Lara A., Villalba R., Wolodarsky-Franke A., Aravena J. C., Luckman B. H., and Cuq E. (2005). Spatial and temporal variation in Nothofagus pumilio growth at tree line along its latitudinal range (35 40′–55 S) in the Chilean Andes. J. Biogeogr. 32, 879–893. doi: 10.1111/j.1365-2699.2005.01191.x
Lewis S. L., Brando P. M., Phillips O. L., van der Heijden G. M. F., and Nepstad D. (2011). The 2010 amazon drought. Science 331, 554. doi: 10.1126/science.1200807
Luebert F. and Pliscoff P. (2018). Sinopsis bioclimática y vegetacional de Chile. Santiago Chile: Editorial Universitaria Universidad Chile. . p, 384.
Lutz D. A., Powell R. L., and Silman M. R. (2013). Four decades of Andean timberline migration and implications for biodiversity loss with climate change. PloS One 8, e74496. doi: 10.1371/journal.pone.0074496
McIntire E. J., Piper F. I., and Fajardo A. (2016). Wind exposure and light exposure, more than elevation-related temperature, limit tree line seedling abundance on three continents. J. Ecol. 104, 1379–1390. doi: 10.1111/1365-2745.12599
Michel J., Hartley I. P., Buckeridge K. M., van Meegen C., Broyd R. C., Reinelt L., et al. (2023). Preferential substrate use decreases priming effects in contrasting treeline soils. Biogeochemistry 162, 141–161. doi: 10.1007/s10533-022-00996-8
Nogués-Bravo D., Araújo M. B., Romdal T., and Rahbek C. (2008). Scale effects and human impact on the elevational species richness gradients. Nature 453, 216–219. doi: 10.1038/nature06812
Oliveras I., Román-Cuesta R. M., Urquiaga-Flores E., Quintano Loayza J. A., Kala J., Huaman V., et al. (2018). Fire effects and ecological recovery pathways of tropical montane cloud forests along a time chronosequence. Glob Change Biol. 24, 758–772. doi: 10.1111/gcb.13951
Osorio D., Steele J., Sepúlveda M., Gayo E. M., Capriles J. M., Herrera K., et al. (2017). The Dry Puna as an ecological megapatch and the peopling of South America: Technology, mobility, and the development of a late Pleistocene/early Holocene Andean hunter-gatherer tradition in northern Chile. Quat Int. 461, 41–53. doi: 10.1016/j.quaint.2017.07.010
Pabón-Caicedo J. D., Arias P. A., Carril A. F., Espinoza J. C., Borrel L. F., Goubanova K., et al. (2020). Observed and projected hydroclimate changes in the andes. Front. Earth Sci. 8. doi: 10.3389/feart.2020.00061
Paulsen J. and Körner C. (2014). A climate-based model to predict potential treeline position around the globe. Alp Bot. 124, 1–12. doi: 10.1007/s00035-014-0124-0
Peyre G. (2021). “Terrestrial biodiversity hotspots: challenges and opportunities,” in Encyclopaedia of the UN development goals: Life on Land. Eds. Leal Filho W., Azul A. M., Brandli L., Lange Salvia A., and Wall T. (Cham, Switzerland: Online: Springer Cham), 12–56. doi: 10.1007/978-3-319-71065-5_150-1
Peyre G., Montesinos D., Giraldo D., Galán de Mera A., Ruthsatz B., Luebert F., et al. (2022). VegAndes: the vegetation database for the Latin American highlands. Veg Classif Surv. . 3, 287–296. doi: 10.3897/VCS.95750
Peyre G., Osorio D., François R., and Anthelme F. (2021). Mapping the páramo land-cover in the Northern Andes. Int. J. Remote Sens. 42, 7777–7797. doi: 10.1080/01431161.2021.1964709
Piper F. I., Viñegla B., Linares J. C., Camarero J. J., Cavieres L. A., and Fajardo A. (2016). M editerranean and temperate treelines are controlled by different environmental drivers. J. Ecol. 104, 691–702. doi: 10.1111/1365-2745.12555
Rehm E. M. and Feeley K. J. (2015). The inability of tropical cloud forest species to invade grasslands above treeline during climate change: potential explanations and consequences. Ecography 38, 1167–1175. doi: 10.1111/ecog.01050
Richardson A. D. and Friedland A. J. (2009). A review of the theories to explain arctic and alpine treelines around the world. J. Sustain For. 28, 218–242. doi: 10.1080/10549810802626456
Román-Cuesta R. M., Salinas N., Asbjornsen H., Oliveras I., Huaman V., Gutiérrez Y., et al. (2011). Implications of fires on carbon budgets in Andean cloud montane forest: The importance of peat soils and tree resprouting. For Ecol. Manage. 261, 1987–1997. doi: 10.1016/j.foreco.2011.02.025
Santini L., Craven D., Rodriguez D. R. O., Quintilhan M. T., Gibson-Carpintero S., Torres C. A., et al. (2024). Extreme drought triggers parallel shifts in wood anatomical and physiological traits in upper treeline of the Mediterranean Andes. Ecol. Process. 13, 10. doi: 10.1186/s13717-024-00486-9
Sarmiento F. O. and Frolich L. M. (2002). Andean cloud forest tree lines. Mt Res. Dev. 22, 278–287. doi: 10.1659/0276-4741(2002)022[0278:ACFTL]2.0.CO;2
Schenker G., Lenz A., and Hoch KC. (2014). Physiological minimum temperatures for root growth in seven common European broad-leaved tree species. Tree Physiol. 34, 302–313. doi: 10.1093/treephys/tpu003
Srur A. M., Villalba R., Rodríguez-Catón M., Amoroso M. M., and Marcotti E. (2018). Climate and Nothofagus pumilio establishment at upper treelines in the Patagonian Andes. Front. Earth Sci. 6. doi: 10.3389/feart.2018.00057
Testolin R -, Attorre F., and Jiménez-Alfaro B. (2020). Global distribution and bioclimatic characterization of alpine biomes. Ecography 43, 779–788. doi: 10.1111/ecog.05012
Toivonen J. M., Gonzales-Inca C. A., Bader M. Y., Ruokolainen K., and Kessler M. (2017). Elevational shifts in the topographic position of Polylepis forest stands in the Andes of Southern Peru. Forests 9, 7. doi: 10.3390/f9010007
Tovar C., Carril A. F., Gutiérrez A. G., Ahrends A., Fita L., Zaninelli P., et al. (2022). Understanding climate change impacts on biome and plant distributions in the Andes: Challenges and opportunities. J. Biogeogr. 49, 1420–1442. doi: 10.1111/jbi.14389
Urquiaga-Flores E. G., Bader M. Y., and Kessler M. (2024). Contrasting topography-vegetation relationships at natural and human-influenced mountain treelines in the Peruvian Andes. Landsc Ecol. 39, 1–13. doi: 10.1007/s10980-024-02006-8
Urrego D. H., Niccum B. A., La Drew C. F., Silman M. R., and Bush M. B. (2011). Fire and drought as drivers of early Holocene tree line changes in the Peruvian Andes. J. Quat Sci. 26, 28–36. doi: 10.1002/jqs.1422
Valencia B. G., Urrego D. H., Silman M. R., and Bush M. B. (2010). From ice age to modern: a record of landscape change in an Andean cloud forest. J. Biogeogr. 37, 1637–1647. doi: 10.1111/j.1365-2699.2010.02318.x
Young K. R. and Leon B. (2007). Tree-line changes along the Andes: implications of spatial patterns and dynamics. Philos. Trans. R Soc. B: Biol. Sci. 362, 263–272. doi: 10.1098/rstb.2006.1986
Young K. R., Ponette-González A. G., Polk M. H., and Lipton J. K. (2017). Snowlines and treelines in the tropical Andes. Ann. Assoc. Am. Geogr. 107, 429–440. doi: 10.1080/24694452.2016.1235479
Keywords: Andes, ecotone, forest, global change, tree migration
Citation: Peyre G, Zuñiga E, Cornejo Paredes A, Paja Medina A, Sulla Torres JA, Zúñiga Carnero M and Rosas Paredes K (2025) Socio-ecological dynamics of Andean treelines in the 21st century. Front. Ecol. Evol. 13:1613871. doi: 10.3389/fevo.2025.1613871
Received: 17 April 2025; Accepted: 30 June 2025;
Published: 18 July 2025.
Edited by:
Rubén G. Mateo, Autonomous University of Madrid, SpainReviewed by:
Bolier Torres, Universidad Estatal Amazónica, EcuadorCopyright © 2025 Peyre, Zuñiga, Cornejo Paredes, Paja Medina, Sulla Torres, Zúñiga Carnero and Rosas Paredes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gwendolyn Peyre, Z3dlbnBleXJlOEBnbWFpbC5jb20=; Z2YucGV5cmVAdW5pYW5kZXMuZWR1LmNv