 Tamás Sipos1*
Tamás Sipos1* Balázs Kolics2Éva Kolics-Horváth2Tamás Donkó3Ádám Csóka3Kristóf Kozma-Bognár4András Kovács5Sándor Farkas6
Balázs Kolics2Éva Kolics-Horváth2Tamás Donkó3Ádám Csóka3Kristóf Kozma-Bognár4András Kovács5Sándor Farkas6 Katalin Somfalvi-Tóth1
Katalin Somfalvi-Tóth1 Sándor Keszthelyi1
Sándor Keszthelyi1- 1Institute of Agronomy, Hungarian University of Agriculture and Life Sciences, Kaposvár, Hungary
- 2Festetics Bioinnovation Group, Institute of Genetics and Biotechnology, Hungarian University of Agriculture and Life Sciences, Keszthely, Hungary
- 3Moritz Kaposi General Hospital, Medicopus Nonprofit Ltd., Kaposvár, Hungary
- 4Festetics Doctoral School, Institute of Agronomy, Hungarian University of Agriculture and Life Sciences, Keszthely, Hungary
- 5Department of Material Science and Chemical Engineering, University of Pannonia, Veszprém, Hungary
- 6Institute for Wildlife Management and Nature Conservation, Hungarian University of Agriculture and Life Sciences, Kaposvár, Hungary
Introduction: The yellow-legged hornet (Vespa velutina nigrithorax Lepeletier, 1836) and the European hornet (Vespa crabro L., 1758) are two social generalist predator wasps that have successfully expanded beyond their native ranges, raising ecological concerns in newly colonized regions. This study presents a comparative analysis of the two species based on morphological parameters derived from digital microscopy and micro-CT imaging.
Methods: Species were collected in Hungary and examined in order to assess differences in body size, colouration, and flight-related morphology. To determine the differences between species, 16 different morphological parameters were measured by micro-ct examination and digital microscopy. The flight parameters of the examined two species were assessed through 14 flight-related coefficients and a correlation matrix and a linear model based on these.
Results and discussion: Results from the colour analyses performed significantly darker colouration of V. v. nigrithorax compared with V. crabro. Morphological measurements confirmed that V. crabro is generally larger and more robust in body size, while V. v. nigrithorax posesses relatively longer legs and a greater wing surface area proportional to body mass. V. v. nigrithorax outperformed V. crabro in most flight-related traits, e.g. speed, manoeuvrability, balancing ability, hovering and carrying capacity. Correlation analyses supported these findings, showing stronger associations among wing parameters in V. v. nigrithorax and more uniform body-thorax interaction and more robust body structure in V. crabro. These results argue that V. v. nigrithorax’s better flight performance underlies its effective bee-hawking behaviour and competitive dominance in habitats in which the two hornet species co-occur. The study highlights how morphological adaptations contribute to predatory success and invasion potential. Our findings also confirm that in the absence of natural enemies, an adventive predator can exhibit significant ecological advantages in competitive interactions, certainly in the initial success of an invasive species.
1 Introduction
The yellow-legged hornet (Vespa velutina nigrithorax Lepeletier, 1836) (Hym.: Vespidae) and the European hornet (Vespa crabro L., 1758) (Hym: Vespidae) occupy different geographic ranges with different native fauna. As top insect predators, these two Vespa species have successfully established non-native populations far from their endemic ranges, adding to economic difficulties and environmental problems (Beggs et al., 2011; Barbet-Massin et al., 2020). Both are invasive species that permanently occupy new habitats with negative consequences for the natural environment and/or humans (Monceau et al., 2015; Otis et al., 2023). The natural range of V. crabro is temperate Eurasia, from the United Kingdom through the Iberian and Italian Peninsula to the Far East, Eastern Russia and Japan. It was accidentally introduced into eastern North America in the 1840s (Bequaert, 1932; Shaw and Weidhaas, 1956; Landolt et al., 2010; Bass et al., 2022). One of the most devastating enemies of the western honey bee (Apis mellifera L., 1758) (Hym.: Apidae), the yellow-legged hornet is currently spreading rapidly and become the most abundant Vespa species in the world (Park et al., 2024). V. v. nigrithorax is widely distributed and occupies different habitats in tropical and subtropical regions of Asia, from Pakistan throughout most of Indochina, Indonesia, and Taiwan, Southeastern Asia and the Malay Penninsula and to North America (Choi et al., 2012; Monceau et al., 2014; Lioy et al., 2023; Otis et al., 2023; Hoebeke et al., 2024). Its high invasion potential has been demonstrated by its rapid dispersal through Europe, Korea, and Japan (Robinet et al., 2017; Lioy et al., 2023; Otis et al., 2023).
The emergence and successful spread of this invasive hornet into new habitats poses a serious threat to the conservation of the native insect fauna, due to their predation and competition abilities (Snyder and Evans, 2006; Cini et al., 2018; Carisio et al., 2022). The competitive interactions can emerge among Vespa species when their habitats overlap within the same biotope. This phenomenon relates to several factors, such as overlapping nesting sites, access to resources, prey acquisition, and partially overlapping seasonal phenology (Beggs et al., 2011; Barbet-Massin et al., 2020). Each species efficiently exploits the organic resources available in its endemic range, thereby effectively occupying its ecological niche (Landolt et al., 2010; Lioy et al., 2023). However, this ecological success has also facilitated the colonization of new, previously uninhabited environments, driven by factors such as increased transportation, urbanization, and, importantly, climate change. V. crabro has historically occupied Eurasian regions (Bequaert, 1932; Shaw and Weidhaas, 1956; Snyder and Evans, 2006; Landolt et al., 2010). Whilst V. crabro is a generalist predator, it has not been reported to cause significant losses of honey bees, or environmental damages within endemic nor newly introduced ranges (Cini et al., 2018; Rome et al., 2021). The yellow-legged hornet by way of contrast has a semi-specialised diet which mainly targets honey bees and social wasps of the genus of Vespula and Polistes, and Diptera species (Laurino et al., 2019; Rome et al., 2021). Its presence in regions it has invaded raises various problems, due to its high predation of A. mellifera, causing a serious economic challenge for the beekeeping industry (Monceau et al., 2014; Rome et al., 2021; Diéguez-Antón et al., 2025).
The competition between predator and prey is an ongoing evolutionary contest, enacted via traits and strategies that evolve to maximize the chances of the predator to successfully catching food resources, and for local prey populations survive the attack and live on (Brodie and Brodie, 1999; Kwon and Choi, 2020). The problematic situation could arise when the ranges of two Vespa species begin to overlap. In such cases, the native Vespa species may experience competitive pressures, or even displacement following the arrival of a previously absent Vespa species (Snyder and Evans, 2006; Cini et al., 2018; Carisio et al., 2022; Lioy et al., 2023). However, in areas where the endemic species coexists with the ecologically similar and invasive V. v. nigrithorax, its dominance may be challenged. Previous studies have shown that V. v. nigrithorax can compete with other Vespa species within its invasive range in Asia (Choi et al., 2012; Park et al., 2024). Moreover, it has rapidly become the most common hornet species in Korea near Busan within six years (Choi et al., 2012; Lioy et al., 2023; Park et al., 2024).
In Europe, competition has also been hypothesized between V. v. nigrithorax and the native V. crabro, although direct field-based confirmation remains limited (Cini et al., 2018; Carisio et al., 2022). There are clearly visible morphological differences between the workers of the Vespa species studied for, example in terms of colour and body size (Kovac and Stabentheiner, 2012; Smith-Pardo et al., 2020; Do et al., 2022; Mattila et al., 2022) These differences are the result of adaptive processes and predation strategy linked to the biotope of the original habitat (Araújo et al., 2004; Perrard et al., 2013, 2016). Among hornet species, the flight capacity is a major driver of success, most exemplified by bee-hawking activity, a specialised predation stategy found in the case of V. v. nigrithorax (Tan et al., 2007; Sauvard et al., 2018; Cappa et al., 2021).
The morphology associated with flight affects various aspects of performance, such as lifting capacity, speed, endurance, and manoeuvrability (which refers to the ability to change the velocity and the direction of movement) (Marden, 1987; Sauvard et al., 2018). Several morphological parameters and the ratios calculated from them can be used to infer flight performance and locomotion ability (Marden, 1987; Byrne et al., 1988; Danforth, 1989; Sauvard et al., 2018; Kwon and Choi, 2020). There is currently a scarcity of data on insect morphological characteristics derived from three-dimensional micro-CT imaging (Jonsson, 2023; Toulkeridou et al., 2023). Overall, relatively few studies in entomology have employed non-invasive imaging techniques such as micro-tomographyto investigate insect anatomy. The principal advantage of this technique lies in its ability to capture detailed, comparable internal structures and morphological features without resorting to dissection or other destructive methods, thus preserving specimens in their natural state (Jonsson, 2023; Moraes et al., 2023; Toulkeridou et al., 2023).
Based on these morphological features, our present research aimed to investigate the fundamental differences in the body structure of the two Vespa species, V. crabro and the invasive V. velutina nigrithorax, to understand and describe the external morphology in more detail, and to compare the two species flying abilities by calculating several flight coefficient factors. Our work also aimed to confirm the differences in morphology related to hunting strategies of the two species using modern 3-D imaging techniques and digital microscopy. Using these data, it is possible to compare the predation success and ecological competitiveness of two generalist predators within a specific habitat by examining their morphological traits. By introducing a generalist predator into a newly occupied habitat, the study can effectively highlight the strengths and weaknesses of this predator when competing with endemic species, providing valuable insights into ecological adaptations and competitive interactions.
2 Materials and methods
2.1 Sampling
Worker hornets of the two test species were collected from Hungary on 5th of October 2023. Specimens of V. v. nigrithorax represented workers from a colony detected for the first time in Hungary, originating from Kimle (Győr-Moson-Sopron County; GPS: 47°49’38.3”N; 17°22’17.9”E). To eliminate possible variation that the external factors cause in the developmental stage of a nest, V. crabro workers from one colony were collected at the same time in Kimle (Győr-Moson-Sopron County; GPS: 47°49’43.0”N; 17°22’08.7”E). Insects were trapped with insect nets and immediately killed with ethyl acetate vapour. The collected insect material was stored in a laboratory under room-temperature conditions until the studies were begun. In order to compare the morphological characteristics of V. crabro and V. v. nigrithorax, the whole body and wing compartments were analysed. All investigated parameters were represented by averages of values taken from 15 individuals of each species, a sample size considered adequate in terms of species representation because of the fact that these insects have a haplodiploid breeding system and hence tend to be somewhat inbred anyway, and in the case of the invasive hornet here studied, populations probably initially derived from a few mated founder queens (Taylor et al., 2024). For the micro-CT imaging and for the wing parameters analysis air-dried pinned insects were used. The insects were pinned using a 0.2 mm metal needle and mounted on a polystyrene tray following the method described by Methven et al. (1995). Before pinning, wings were carefully removed from each specimen and mounted on microscope slides with Euparal mounting media to allow subsequent wing measurements.
2.2 Micro-computed tomographical analysis
A Nikon XT H 225 ST X-Ray Tomograph instrument was applied to perform the micro-CT scans. During the measurements, 160 kV accelerating voltage, and 85 µA beam current were used. For the scan, 1,250 projections were acquired with 2 frames/projection, which meant ~17 GB file size. The effective pixel size was 8 µm. During the reconstruction, scatter reduction, noise reduction, median filter, and image processing were applied to enhance the quality of the scans. The analysis was performed using Volume Graphics Studio Max 2023/1 software. The measurements were performed on prepared wasps that had been air-dried. Consequently, very thin layer thicknesses were formed, which made measurements difficult. Additionally, a thin metal needle used for fixation of the insects gave a high contrast, which caused a certain amount of shading. To eliminate this, several software algorithms were applied to reduce the shadowing and enhance the contrast. The measurements were performed both automatically and manually using Volume Graphics Studio Max 2023/1 and 3D Slicer software (version 5.6.3).
2.3 Morphometric measurements
Measurements of morphological parameters of the 15 individuals of each species were conducted using digital microscopy and micro-CT imaging techniques. A summary of the measured parameters is presented in Figure 1. Microscopic examinations were performed using a Keyence VHX-XF1 digital microscope equipped with a VH-Z20T lens (magnification range: 20× to 200×). Morphometric analyses were performed on dissected forewings and hindwings, including measurements of front wing length (e), front wing width (f), and cell surface area (g). Additionally, the total surface areas of the front wings (h1) and hind wings (h2) were determined according to the abbreviations detailed in Figure 1. To evaluate the flight capabilities and consequent predatory specializations of the examined Vespa species, various morphological parameters were measured using micro-CT imaging and analysed in 3D Slicer software (version 5.6.3). Volume measurements – specifically, dry body volume (k) and thorax volume (d2) – were calculated using the Segment Editor and Segment Statistics modules. Additionally, linear dimensions including longitudinal body length (a), leg lengths (b), head width (c), and thorax width (d1) were measured utilizing the Markups module within 3D Slicer.
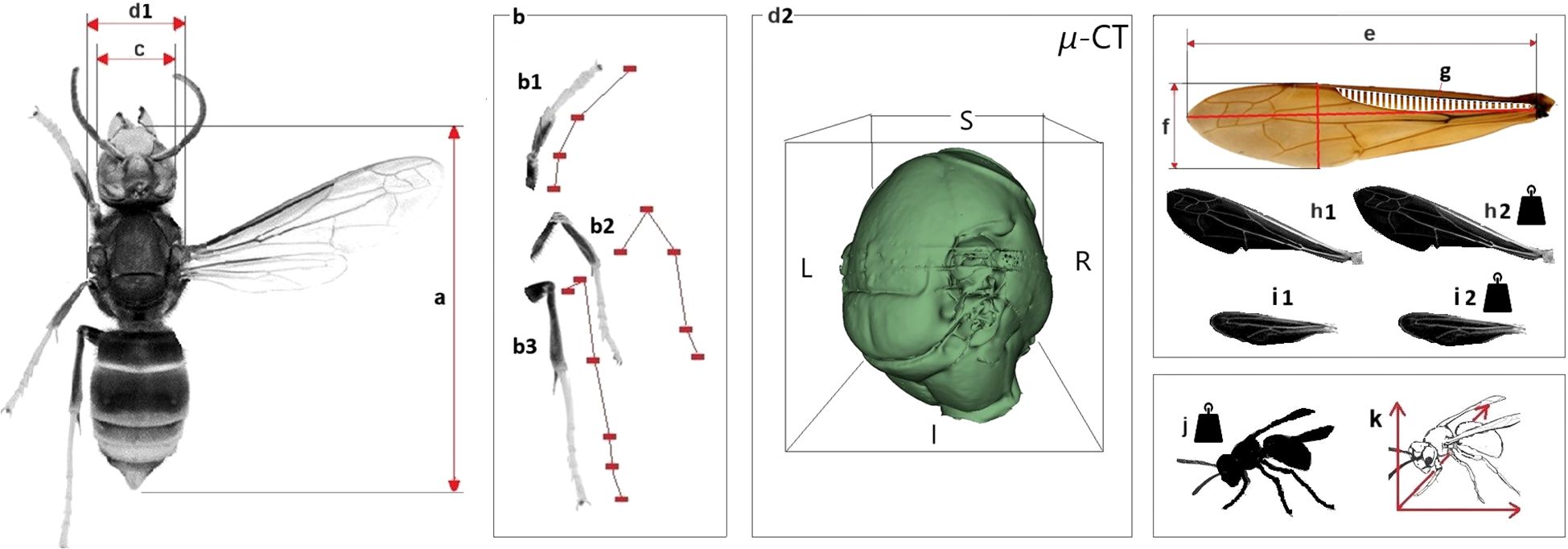
Figure 1. The morphological characteristics of the examined Vespa species. Explanation: (a): longitudinal body length (mm); (b): legs lengths (mm); (b1): front leg length; (b2): middle leg length; (b3): hind leg length; (c): head width (mm); (d1): thorax width (mm); (e): front wing length (mm); (f): front wing width (mm); (g): cell surface (mm2); (h1): front wing surface (mm2); (h2): front wing mass (mg): (i1): hind wing surface (mm2); (i2): hind wing mass (mg); (j): dry body mass (mg); (k): dry body volume (mm3); (d2): thorax volume (mm3).
The mass of various body components, including front wing mass (h2), hind wing mass (i2), and dry body mass (j), was measured using an Ohaus Explorer Semi-Micro balance (model EX125), with a precision of 0.01 mg.
To compare the estimated flight capabilities of the two species and thus estimate their hunting abilities, specific flight-related morphological ratios were calculated. The morphological parameters outlined in Figure 1 contributed to the calculation of these ratios listed in Table 1. The formulas for each ratio, along with the specific flight abilities they directly influence, are presented in Table 1.
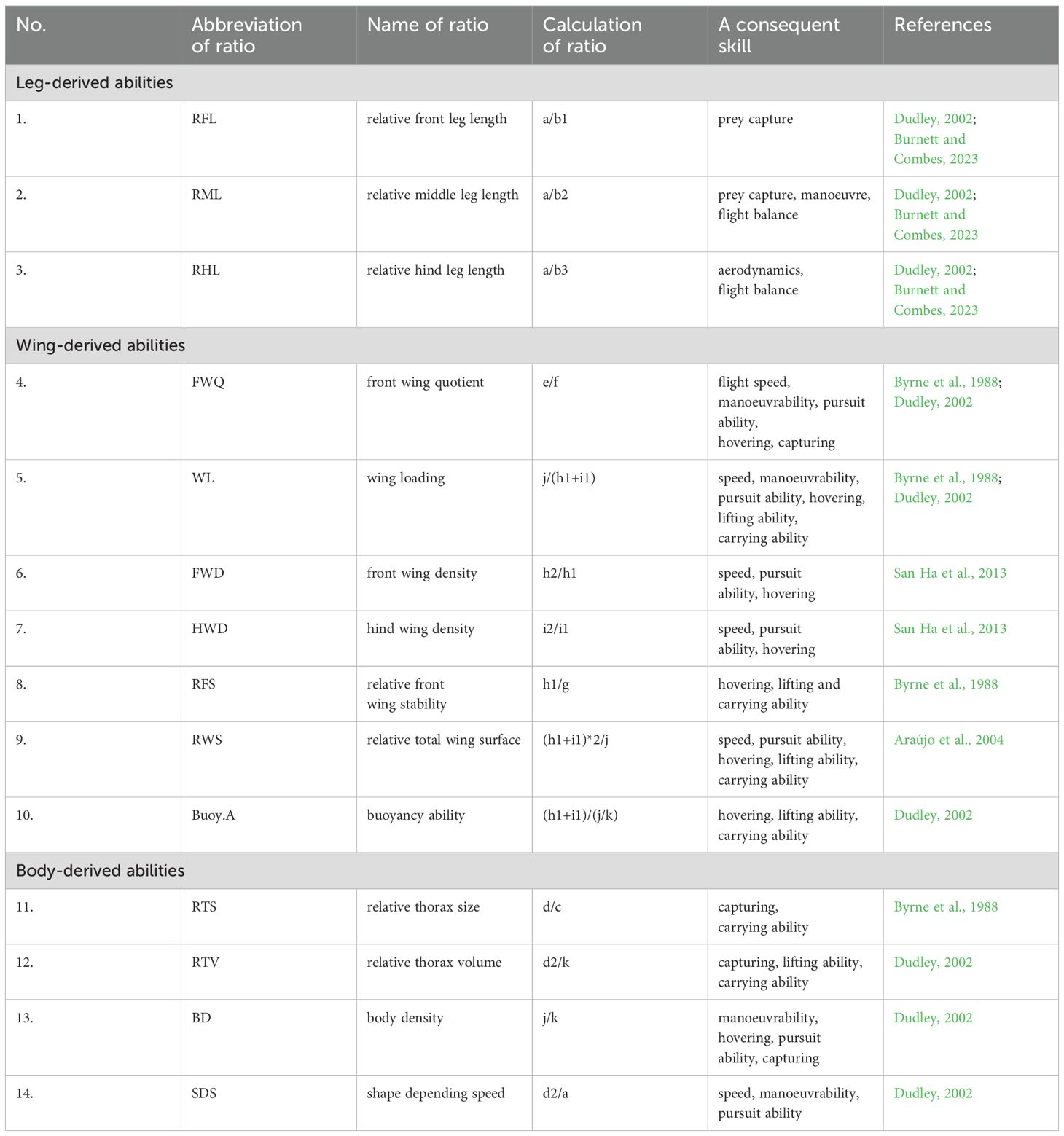
Table 1. The calculated morphometric coefficients used to estimate flight and predation characteristics of the examined two Vespa species.
2.4 Colour analysis
The MultiSpec (Version 2024.05.16 64-bit Windows) software (Biehl and Landgrebe, 2002) was used to classify different images from frontal and dorsal perspectives of each species (n=15) taken by Keyence VHX-XF1 digital microscope equipped with a VH-Z20T lens. For the precise classification of the species this was preformed by taking samples from certain parts of the images and assigned them to three different classes (bright, reddish-brown, black). The coverage values of each classes were used for the differentation of the species. The classification was done by selecting a small group of pixels from the different body parts (eg. clypeus), which is a dataset that represents the characteristics of the specified class. If an image fragment contained multiple colour tones, it was sampled in its entirely, thus minimizing the problem caused by unclassified pixels. For the classification of five samples from the image for a given class, this was done to increases the accuracy of the analysis process. The classification was performed using the Gaussian Maximum Likelihood classification algorithm. After the classification were performed, the individual classes were displayed in different colours, the image accuracy of the analysis being measured by Kappa statistics.
2.5 Data evaluation and statistical analysis
Shapiro-Wilk normality test was conducted on the dataset of colour analysis, morphological measurements and calculated parameters. The colour, morphological and calculated parameters of the two Vespa species examined were evaluated using one-way ANOVA (p < 0.05). The relationship between the morphological parameters were calculated by the Pearson-Correlation Matrix for each species and visualized on a heat map. To analyse the hunting and flight performances of each species, a linear model was created for the scoring of different calculated parameters. Given the multiple parameters that were considered in the characterisation of each skill (scouting, hunting, prey carrying as shown in Figure 2), a scoring method was developed that encompassed all the derived parameters. It was hypothesised that all the parameters presented in Figure 2, whether they influenced the flight, hunting or prey-catching properties of the species in a positive or negative direction, were considered with similar weighting. The scoring method was based on a normalisation procedure, i.e. firstly all data were sorted in ascending order of value, then the minimum value was invariably associated with 0 and the maximum value with 1, and a linear equation of a Descartes coordinate was plotted between the two points. Subsequently, all derived parameters were converted to a number ranging from 0 to 1 and aggregated as illustrated in Figure 2. The upward-pointing arrow was interpreted as a positive value, the downward one as a negative value. The scoring of different flight abilities was based on Figure 2. Statistical analyses were performed using Microsoft Excel (Office version 2016) and R statistics software version 4.2.1 (Team, 2010) applying the ‘corrplot’ (Wei and Simko, 2017) and ‘ggplot2’ packages (Wickham, 2016).
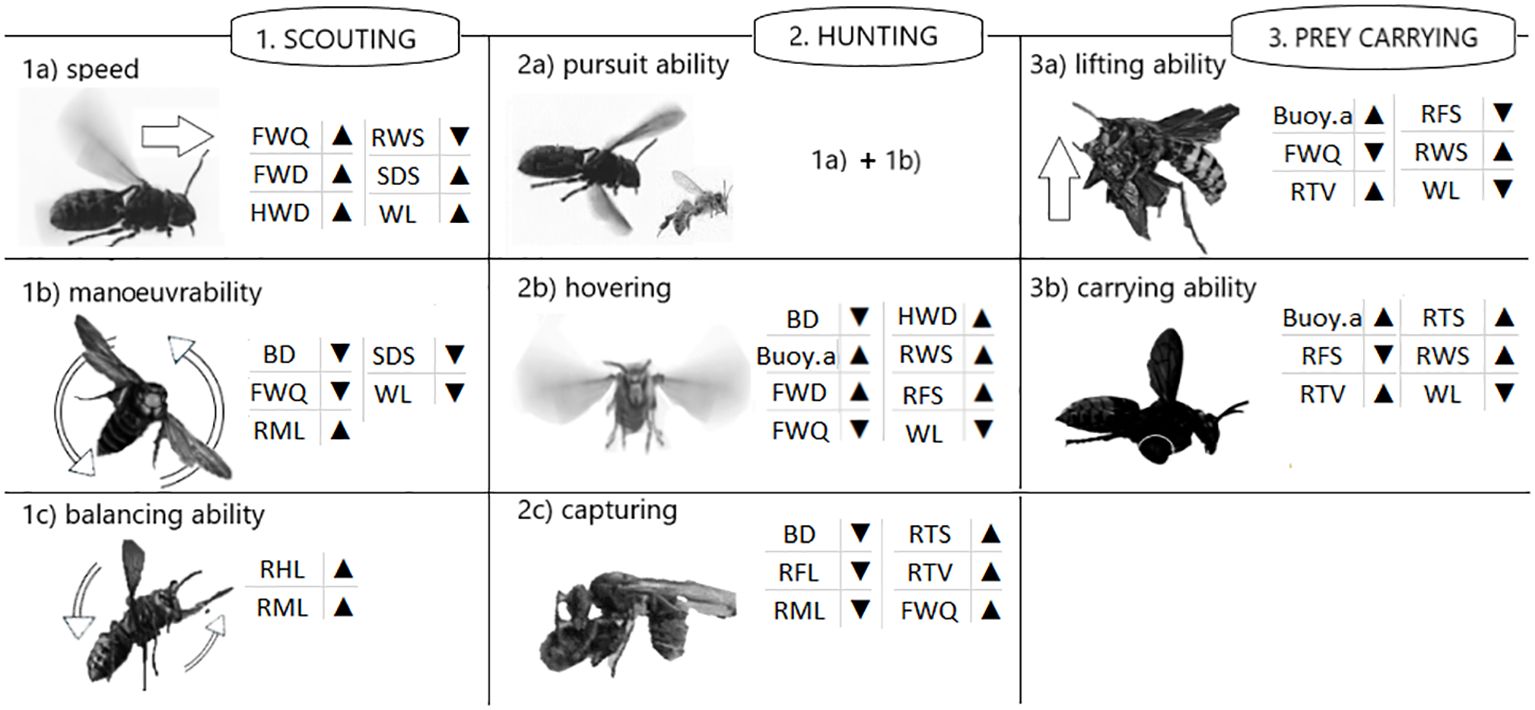
Figure 2. A conceptual illustration of the examined functional flight abilities of V. crabro and V. v. nigrithorax, based on the calculated morphological ratios presented in Table 1. The arrows indicate the contribution of calculated parameter to the surveyed abilities.
3 Results
3.1 Colour analysis
Based on analyses conducted to assess interspecific differences in colouration, various body regions were taken into consideration, the colour analysis results of one specimen of each species illustrated in Figure 3. Regarding head colouration viewed from the front, the following distinctions were observed between the two species. In V. crabro, brownish-reddish colouration dominated, covering 44 ± 0.5% of the head, while in V. v. nigrithorax, the brownish-reddish colouration averaged 18 ± 0.5%. This difference was statistically significant (df = 1; F = 19398; p < 0.001). Bright yellow tones were dominant in V. crabro, covering 30 ± 0.4% of the head, in contrast to V. v. nigrithorax, where the head was covered by bright yellow colouration at a rate of 22 ± 0.8%. Statistical analysis confirmed the difference (df = 1; F = 1173; p<0.001). Regarding black colouration, the head of V. crabro exhibited an average of 26 ± 0.5% such coverage, while V. v. nigrithorax showed an exceptionally high black colouration at 59 ± 1.1%. This disparity was also statistically significant (df = 1; F = 10622; p<0.001). The Kappa statistics values were 98.8% for V. crabro, 98.6% for V. v. nigrithorax.

Figure 3. Comparative dorsal and frontal views of Vespa velutina [left column, (A/a)] and Vespa crabro [right column, (B/b)] alongside their schematic illustrations of the color analysis (A1/a1, B1/b1).
In dorsal view, similar trends were observed in terms of colouration. In V. crabro, brownish-reddish tones dominated the back, accounting for 41 ± 1%, compared to 34 ± 0.5% in V. v. nigrithorax. The difference between the species was statistically significant (df = 1; F = 564; p<0.001). Noteworthy differences were found in the yellow colouration, where V. crabro exhibited a value similar to that of the head (31 ± 1%), whereas V. v. nigrithorax showed a significantly lower average coverage of 10 ± 0.5%. This difference in yellow coverage on the dorsal surface was confirmed through one-way ANOVA (df = 1; F = 6129; p<0.001). Concerning black colouration, V. crabro had 28 ± 0.6% of the back-coloured black, while V. v. nigrithorax displayed the highest level of black colouration at 56 ± 0.6%. The interspecific differences in black colouration on the back were statistically significant (df = 1; F = 18568; p<0.001). The Kappa statistics values of the dorsal view were high for the examined species 98,5 for V. crabro and 98,7% for V. v. nigrithorax.
3.2 Body morphological analyses
The analysis of body size measurements revealed that V. crabro exhibited larger values across all measured parameters and dimensions compared to V. v. nigrithorax. Furthermore, all measured values were found to differ significantly between the two species. The mean values, standard deviations of the measured parameters, and the results of the statistical analyses are presented in Table 2. The differences between the two species can best be determined from the numbered morphological quotients. The mean values, standard deviations of the calculated morphometric coefficients, and the results of the statistical analyses are presented in Table 3.
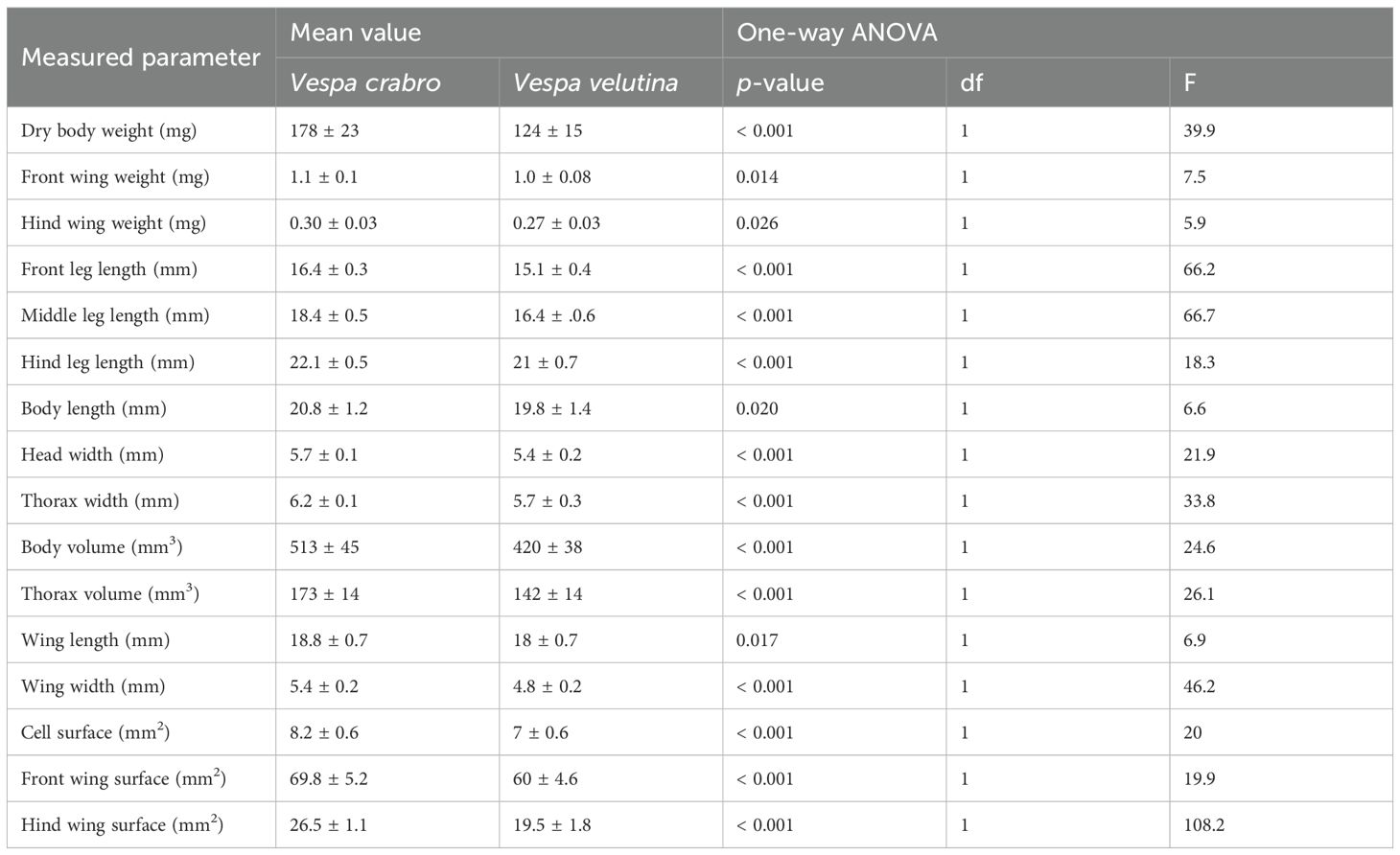
Table 2. The measured morphological parameters of Vespa crabro and Vespa velutina nigrithorax, and the findings from statistical analysis.
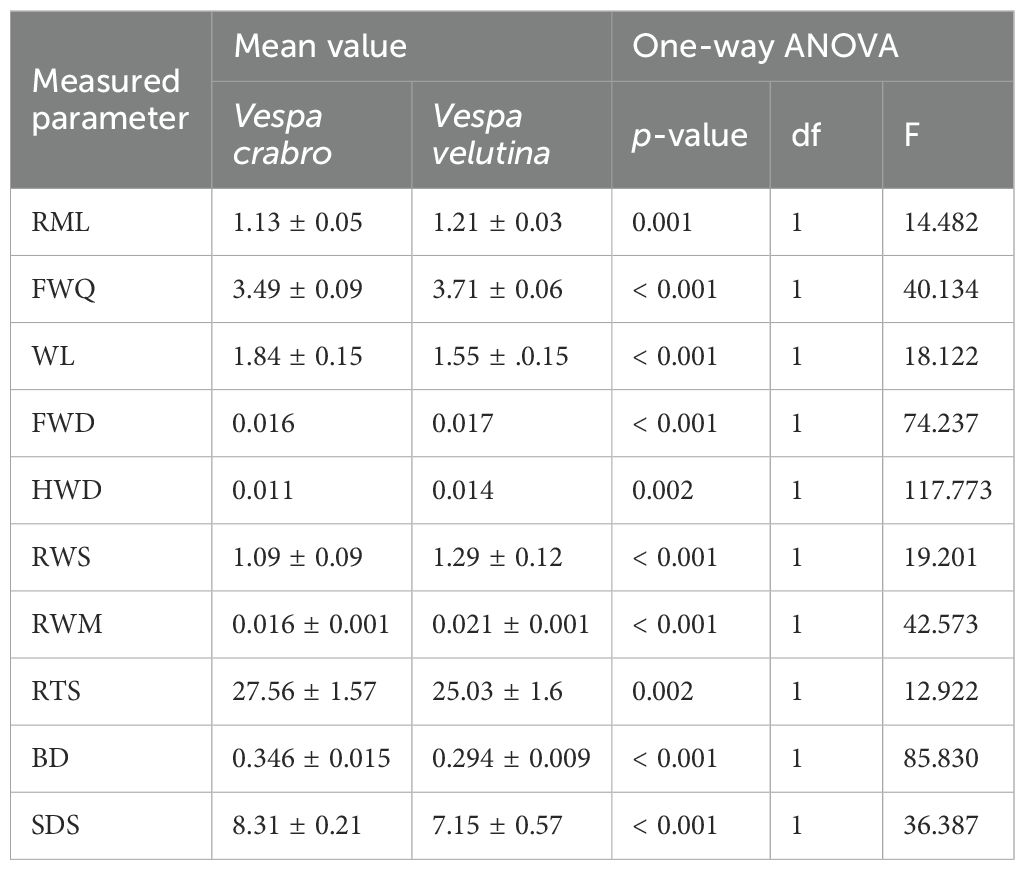
Table 3. The calculated morphometric coefficients of Vespa crabro and Vespa velutina nigrithorax and the statistical analysis outcomes.
3.2.1 Leg-derived parameters
In the case of relative front leg length (RFL) we found no significant differences (df=1; F=3.2; p=0.090), both V. crabro and V. v. nigrithorax having similar front leg proportions relative to body length. However, a significant difference was calculated in the case of relative middle leg length (RML) (df=1; F= 14.5; p=0.001). According to the coefficient, V. v. nigrithorax has proportionally shorter middle legs relative to body length compared to V. crabro. Interestingly, both species have similar parameters derived from the hind leg length (RHL) (df=1; F=0.2; p=0,676).
3.2.2 Wing-derived parameters
Regarding the front wing quotient (FWQ), a highly significant difference was observed (df=1; F=40.1; p<0.001). The higher value in FWQ indicates relatively narrower wings of V. v. nigrithorax. The wing loading (WL) was significantly different, as supported by statistical analysis (df=1; F=18.1; p<0.001). A more robust body structure in V. crabro is indicated by its elevated wing loading, as demonstrated in Table 2. Statistically significant differences were found in front wing density (FWD) (df=1; F=74.3; p=0.001) and hind wing density (HWD) (df=1; F=117.8; p=0.002), with V. v. nigrithorax exhibiting higher values in both cases. Interestingly, relative front wing stability (RFS) did not differ significantly between the species (df=1; F=1.1; p=0.314), suggesting similar wing stability. The relatively larger wings of V. v. nigrithorax, in proportion to its body mass, were reflected in the relative total wing surface (RWS), which showed a statistically significant difference between the species (df=1; F=19.2; p<0.001). It is noteworthy that a similar buoyancy number reflects comparable load-carrying abilities of the examined species, supported by the analysis (df=1; F=1.8; p=0.201).
3.2.3 Body-derived parameters
The relatively robust body structure of V. crabro can be indicated by the higherr value of relative thorax size (RTS), which was different for the two species (df=1; F=12.9; p=0.002). Curiously, the relative thorax volume (RTV) was alike in both species, indicating similar proportions of thorax-to-body volume (df=1; F=0.1; p=0.797). Body density (BD) (df=1; F= 85.84; p=0.001) and shape-dependent speed (SDS) (df=1; F= 36.4; p=0.002) were also elevated in V. v. nigrithorax, like RTS pointing to a more robust body structure.
3.3 Association between morphometric parameters
The Paired-Pearson correlation analysis illustrates on Figure 4 the strength and direction of relationships among the measured morphometric parameters of V. crabro and V. v. nigrithorax. The parameters include body dimensions (dry body weight, body volume, body length, head width, thorax width, thorax volume) leg metrics (front, middle, hind leg lengths) and wing dimensions (length of front wing, width of front wing, front and hind wing surface and weight, front wing cell surface) summarized in Table 2. The analysis reveals the correlation coefficient to be higher than 0.5 and to include the weak (0-0.499) moderate (0.5–0.599), strong (0.6–0.799) and very strong (0.8–1.00) relationships as well as the negative relationships.
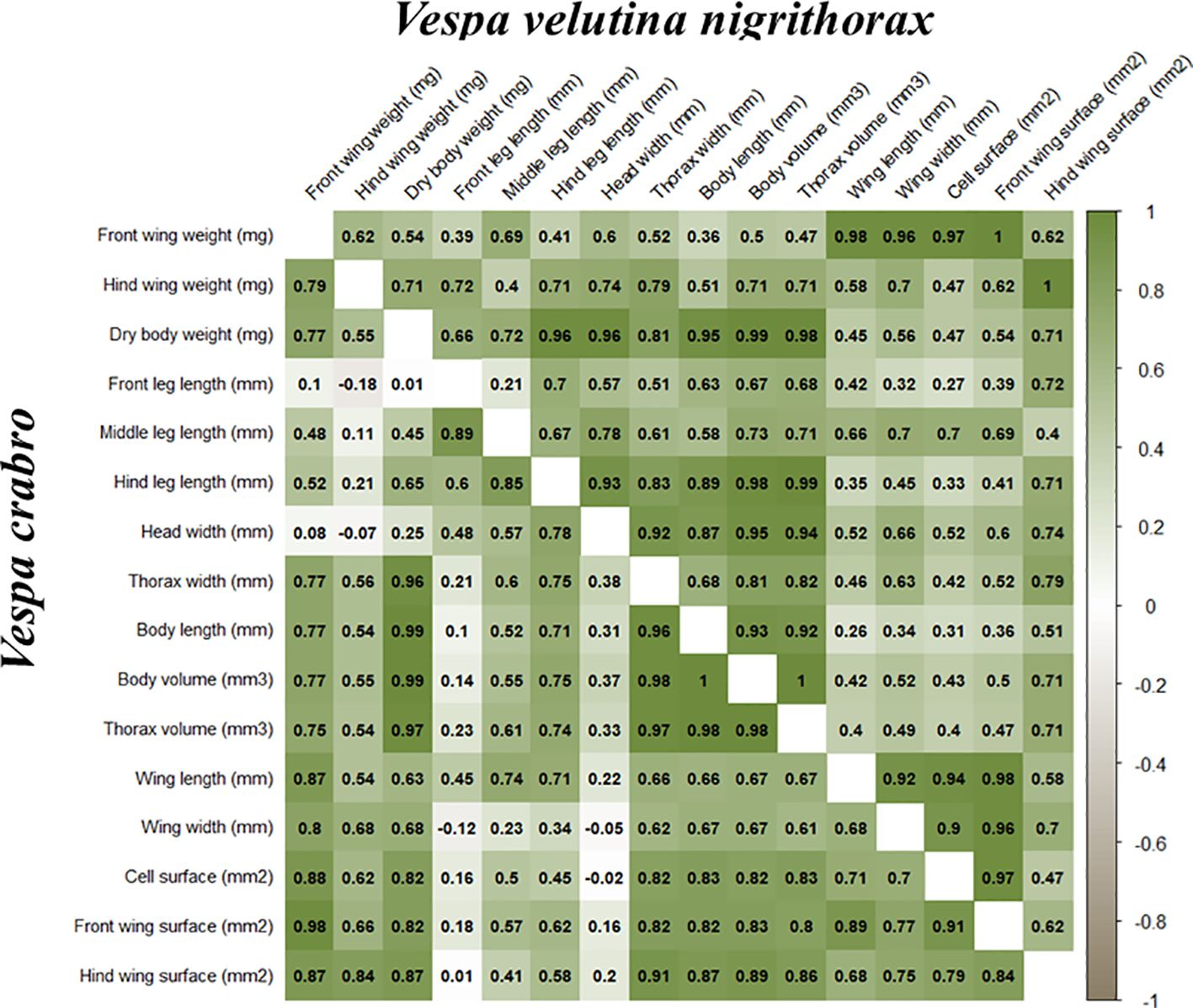
Figure 4. Pearson correlation matrices of the calculated morphological parameters for Vespa crabro and Vespa velutina nigrithorax. Positive correlations are indicated by varying shades of white to green, while negative correlations are represented by white to grey tones.
3.3.1 Correlation patterns in Vespa crabro
Regarding body dimensions, dry body weight exhibits very strong positive correlations with body volume (r=0.99), body length (r=0.99), thorax width (r=0.96), and thorax volume (r=0.97). Additionally, correlations between dry body weight and wing surface parameters are strong, with correlation coefficients ranging from 0.82 to 0.87. Body length, thorax width, thorax volume, and body volume parameters demonstrate very strong correlations (r ranging from 0.96 to 1). Body and thorax dimensions also correlate very strongly with wing surface parameters (r ranging from 0.80 to 0.91). However, head width correlations are relatively weak, showing both low positive and negative relationships (r ranging from –0.15 to 0.38) between all parameters, except the moderate correlation with front leg length (r=0.48).
Wing-related dimensions display very strong correlations; wing length strongly correlates with front wing weight (r=0.87), front wing surface area (r=0.89), and wing cell surface area (r=0.91). Additionally, front and hind wing surface parameters are highly correlated (r=0.84). Both front and hind wing surfaces show very strong correlations with their respective wing weights (r ranging from 0.80 to 0.88). The front wing weight correlates strongly with most body dimensions (r ranging from 0.75 to 0.77), except for a weak correlation observed with head width (r=0.08). Hind wing weight demonstrates moderate correlations with body metrics (r ranging from 0.54 to 0.56).
Leg-length parameters reveal moderate to strong positive correlations with overall body size and weight metrics, particularly middle and hind leg lengths (r ranging from 0.52 to 0.78). However, front leg length correlations are relatively weak, showing both low positive and negative relationships, exemplified by a notably weak negative correlation between front leg length and hind wing weight (r=-0.18), except for the moderate correlations with head width (r=0.48) and wing length (r=0.46).
3.3.2 Correlation patterns in Vespa velutina nigrithorax
Dry body weight exhibits very strong positive correlations with body volume (r=0.99), thorax volume (r=0.98), thorax width (r=0.81), and body length (r=0.95). Correlations between dry body weight and wing surface parameters (front wing surface, hind wing surface, and cell surface area) range from moderate to strong (r ranging from 0.47 to 0.71). Interestingly, correlations between body length, body width, and body surface parameters with wing length, wing width, and wing surface parameters range from weak to strong (r ranging from 0.26 to 0.79). The weakest correlation was observed between body length and wing length (r=0.26), contrasting sharply with the very strong relationship between body length and body volume (r=0.93). Furthermore, body volume, thorax volume, head width, and thorax width parameters are all very strongly intercorrelated (r ranging from 0.81 to 1). The correlations between body and thoracic parameters and wing surface dimensions range from weak to strong (r ranging from 0.40 to 0.71).
Wing dimensions display consistently strong correlations. Specifically, front wing weight correlates very strongly with front wing length (r=0.98), wing width (r=0.92), front wing surface (r = 1.00), and cell surface area (r=0.96). Additionally, front and hind wing surface parameters are moderately correlated (r=0.62). Hind wing parameters exhibit weak to strong correlations with front wing length, width, and cell surface area (r ranging from 0.47 to 0.70).
Leg-length parameters reveal considerable variability in their correlations. Front and hind leg lengths exhibit a strong correlation with each other (r=0.70), whereas front and middle leg lengths demonstrate only a weak correlation (r=0.21). Overall, leg-length parameters show strong positive correlations with body dimensions, particularly hind leg length, which strongly correlates with body weight, width, and volume parameters (r ranging from 0.83 to 0.96). Middle leg length correlations with body dimensions are moderate to strong (r ranging from 0.58 to 0.78). Moreover, a descending correlation trend like that observed in V. crabro is apparent, with front leg length showing moderate to strong correlations with body metrics (r ranging from 0.51 to 0.67). Correlations between front and hind leg lengths and wing dimensions are weak (r ranging from 0.27 to 0.45), except for a strong correlation with hind wing weight (r=0.71). Lastly, middle leg length correlates strongly with wing dimensions (r ranging from 0.66 to 0.70), but weakly with hind wing weight (r=0.40).
3.4 Linear model of flight and hunting abilities
Based on the results of the linear model (Table 4), we show how in ways that likely influence flight abilities are reflected in the scores produced by the model. In terms of scouting abilities, V. v. nigrithorax markedly outperformed V. crabro. Regarding flight speed, V. v. nigrithorax achieved an average score of 2.64, while V. crabro reached a lower score of 2.14. In terms of manoeuvrability, V. crabro performed notably worse, with an average score of -2.19. Although V. v. nigrithorax also received a negative value -0.8, its performance was better in this aspect. Furthermore, V. v. nigrithorax demonstrated more balanced flight with an average score of 1.25, compared to 0.81 achieved by V. crabro.
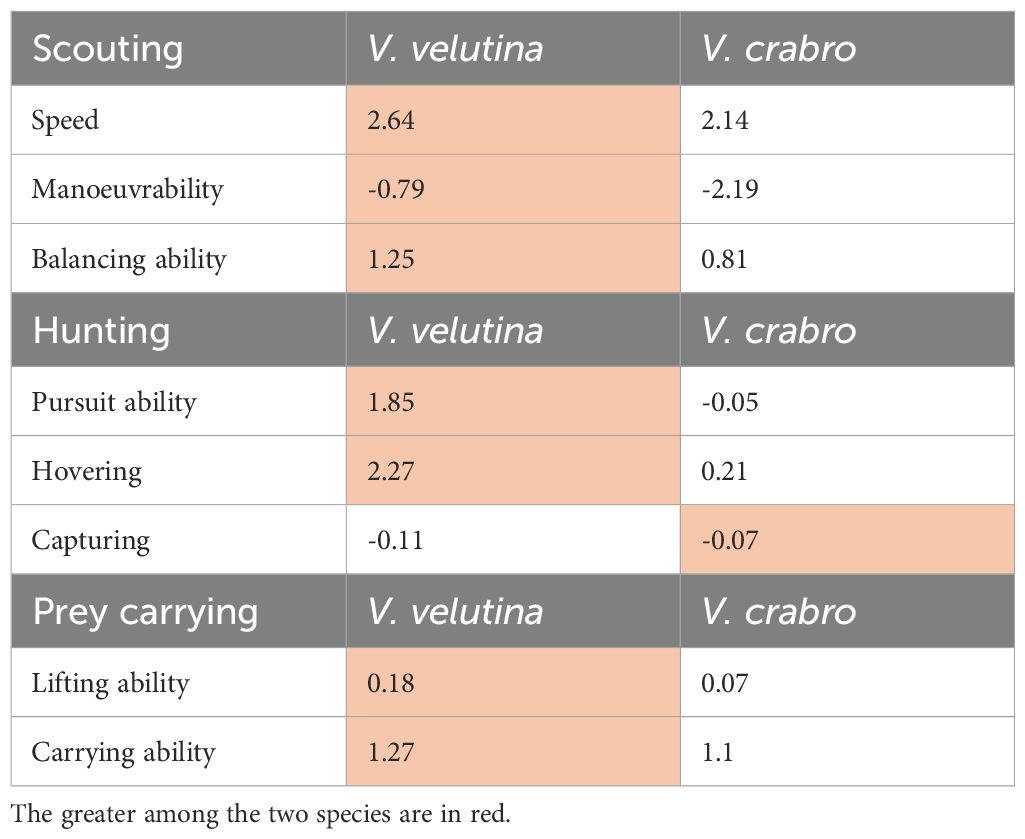
Table 4. Comparative functional flight ability scores of Vespa v. nigrithorax and Vespa crabro based on a linear model and the calculated flight related coefficients.
In the context of abilities contributing to hunting performance, pursuit ability was based on flight speed and manoeuvrability. V. v. nigrithorax outperformed V. crabro in both of these underlying traits, whilst the same trend was also observed in the case of pursuit ability. V. v. nigrithorax achieved a higher scoring of 1.85 in opposition to -0.05. The analysis revealed substantial differences in hovering ability, with V. v. nigrithorax achieving a significantly higher score of 2.27 in the linear model, compared to a score for V. crabro of 0.21. The results for prey capturing ability showed unexpected tendencies, although the differences were minor; V. crabro attained a slightly better score -0.07 than V. v. nigrithorax -0.11. Nevertheless, these values remained relatively close, indicating only a marginal difference between the two species.
In aerial predatory social insects, the ability to transport prey for the benefit of the colony is essential. For the calculation of flight related scores, we incorporated coefficients associated with thorax structure and flight capabilities. The analyzed parameters revealed only minor differences between the two species. Regarding lifting ability, V. v. nigrithorax slightly outperformed V. crabro, achieving a score of 0.18 compared to 0.07. When assessing the ability to simulate prey transport, the differences became more pronounced, with V. v. nigrithorax displaying a higher value of 1.27 compared to V. crabro of 1.1.
4 Discussion
The paradox of the Vespa genus lies in the fact that its various species are highly successful organisms within their respective native habitats, whereas in our case, both examined species have successfully colonized various regions (Monceau et al., 2015; Otis et al., 2023; Hoebeke et al., 2024). The different endemic ranges of these two species may be reflected in the very distinct colouration patterns shown, from both the frontal and dorsal views (Kovac and Stabentheiner, 2012; Do et al., 2022; Mattila et al., 2022). Our study confirmed that the dorsal colouration of V. v. nigrithorax dorsal colouration is much darker (9.5% bright; 34.3% reddish-brown; 56.2% black) than that of V. crabro, suggesting that darker colourn may relate to thermal regulation, crypsis, or mimicry (Joron, 2009; Perrard et al., 2014). Furthermore, the colouration of V.crabro displays a more vivid and variegated dorsal tone (30.9% yellow; 41% redish brown; 28.1% black) with more bright yellowish colours, which may well serve as aposematic signalling against predators (Joron, 2009; Perrard et al., 2014). The different colouration in different areas may certainly play a role in terms fo the ecological specialization and hence success of the two species. In the eastern Palaearctic, within its native range, V. v. nigrithorax does not appear to have any natural predators that exert significant population-regulating pressure. In contrast, within the western Palaearctic, the European bee-eater (Merops apiaster; L. 1758) (Aves: Meropidae), which feeds on hymenopterans, including bees and wasps, has been identified as a potential natural enemy of V. v. nigrithorax (Onofre et al., 2023). Moreover, the cryptic black colouration of V. v. nigrithorax may, in some cases, effectively conceal it from visual predators, thereby reducing its likelihood of being preyed upon (Endler, 1988; Keszthelyi et al., 2024).
The fighting ability related to the success of a predator is largely influenced by body size (Kwon and Choi, 2020). Some earlier studies support the differences here found between the examined two Vespa species, including in terms of morphological and behavioural aspects. Thus, our results reinforce the study by Kwon and Choi (2020), completed using deeper morphological analyses. In our study V. crabro was here found to be larger than V. v. nigrithorax in every aspect, mosty the body volume and dry body weight, reflecting the clearly visible different anatomical structure. The correlation analysis revealed more structural differences in the body anatomy, while V. crabro displayed more uniformly strong correlations among body and thoracic parameters, suggesting a more robust body structure. In contrast, V. v. nigrithorax exhibited stronger correlations among wing traits and leg traits suggesting the key to the success of this species is related to its flight performance. The different body structure can be rooted in terms of direct ecological pressure, thereby supporting the results of Kwon and Choi (2020) in suggesting that V. v. nigrithorax tends to avoid direct competition (fighting, nesting sites) with native hornet species including V. crabro in its Asian range.
The predatory success of these two social wasp species is fundamentally based not only in their overall body size but in their flight performance capabilities (Tan et al., 2007; Sauvard et al., 2018; Cappa et al., 2021). To gain a deeper understanding of the structural differences between V. crabro and V. v. nigrithorax, and thereby their potential predatory success, we calculated multiple flight-related morphological ratios, analying them through the linear model developed specifically to assess key differences of flight and predation abilities.
Based on the analysed coefficients, V. v. nigrithorax has relatively longer legs in proportion to its body size, which likely supports enhanced aerial balance and prey-capturing ability (Dudley, 2002; Burnett and Combes, 2023). In contrast, V. crabro showed higher values in buoyancy-ability but not significant different to that of V. v. nigrithorax in terms of the relative thorax size and volume. However, in terms of wing loading and wing surface-to-body ratio, V. v. nigrithorax appears more adapted for hovering flight and rapid manoeuvrability. The higher wing surface, density, and shape related quotients emphasize the better flight performances of V. v. nigrithorax that appear unequivocally reflected in different hunting activity, the so-called bee-hawking behaviour. These findings are further supported by our linear model analysis, which compared distinct flight abilities between the species (Figure 4; Table 4). According to this model, V. v. nigrithorax outperformed V. crabro in nearly all evaluated flight and predation-related traits as would be expected from empirical studies (Tan et al., 2007; Sauvard et al., 2018; Cappa et al., 2021; Herrera et al., 2023).
The relative success of these two distinct taxa is likely influenced by key morphological differences as revealed by our findings from multiple perspectives. Our research, has been driven by the aim to better understand how morphological traits alongside behavioural differences previously noted, can help explain their ecological performance. Specifically, we tried to examine how the generalist predator V. crabro and the semi-specialized, bee-adapted predator V. v. nigrithorax have morphologically adapted to specific predation strategies, such as bee-hawking.
5 Conclusion
Due to its ethological and ecological traits, V. v. nigrithorax appears to have gained a competitive edge over the native V. crabro in some special areas, i.e. bee-hawking. However, more empirical studies need to be preformed to ascertain whether or not V.v. nigrithorax can really affect native Vespidae species at the population-level. The invasive V. v. nigrithorax behavioural, morphological, and visual adaptations – particularly its efficiency in terms of honeybee predation – have positioned it as a major emerging threat to honeybee populations in western Palaearctic landscapes. These characteristics allow V. v. nigrithorax to become the dominant arthropod predator of honey bees in this region, regardless of the continued presence of its congener V. crabro.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
TS: Writing – original draft, Project administration, Funding acquisition, Investigation, Data curation, Supervision, Resources, Conceptualization, Writing – review & editing. BK: Methodology, Writing – original draft, Resources. ÉK-H: Writing – original draft, Resources, Methodology. TD: Writing – original draft, Resources, Investigation, Formal analysis, Data curation. ÁC: Data curation, Validation, Writing – original draft, Software. KK-B: Software, Writing – original draft, Visualization, Validation. AK: Writing – original draft, Methodology, Software, Resources, Visualization, Investigation. SF: Writing – original draft, Writing – review & editing, Data curation, Methodology, Resources, Validation, Software. KS-T: Funding acquisition, Supervision, Resources, Writing – original draft, Data curation. SK: Writing – review & editing, Conceptualization, Investigation, Supervision, Data curation, Methodology, Visualization, Funding acquisition, Writing – original draft, Project administration, Formal analysis.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Supported by the EKÖP-MATE/2024/25/D and EKÖP-MATE/2024/25/K New National Excellence Program of the Ministry for Culture and Innovation from the source of The National Research, Development and Innovation Fund. This work was supported by the Flagship Research Groups Programme of the Hungarian University of Agriculture and Life Sciences.
Acknowledgments
We thank the several independent reviewers for their insightful comments, which have made us re-think some of that we have said and have thus improved the paper.
Conflict of interest
Authors TD and ÁC were employed by Medicopus Nonprofit Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Araújo E. D., Costa M., Chaud-Netto J., and Fowler H. G. (2004). Body size and flight distance in stingless bees (Hymenoptera: Meliponini): inference of flight range and possible ecological implications. Braz. J. Biol. 64, 563–568. doi: 10.1590/S1519-69842004000400003
Barbet-Massin M., Salles J. M., and Courchamp F. (2020). The economic cost of control of the invasive yellow-legged Asian hornet. NeoBiota 55, 11–25. doi: 10.3897/neobiota.55.38550
Bass A., Needham K., and Bennett A. M. (2022). First record of Vespa crabro Linnaeus (Hymenoptera: Vespidae) in western North America with a review of recorded species of Vespa Linnaeus in Canada. Zootaxa 5154, 305–318. doi: 10.11646/zootaxa.5154.3.4
Beggs J. R., Brockerhoff E. G., Corley J. C., Kenis M., Masciocchi M., Muller F., et al. (2011). Ecological effects and management of invasive alien Vespidae. BioControl 56, 505–526. doi: 10.1007/s10526-011-9389-z
Bequaert J. (1932). A tentative synopsis of the hornets and yellow-jackets (Vespinae; Hymenoptera) of America. Entomol Am. 12, 71–138. doi: 10.5281/zenodo.16436501
Biehl L. and Landgrebe D. (2002). MultiSpec—a tool for multispectral–hyperspectral image data analysis. Comput. Geosci 28, 1153–1159. doi: 10.1016/S0098-3004(02)00033-X
Brodie E. D. III and Brodie E. (1999). Predator-prey arms races: asymmetrical selection on predators and prey may be reduced when prey are dangerous. Bioscience 49, 557–568. doi: 10.2307/1313476
Burnett N. P. and Combes S. A. (2023). Close encounters of three kinds: impacts of leg, wing and body collisions on flight performance in carpenter bees. J. Exp. Biol. 226, jeb245334. doi: 10.1242/jeb.245334
Byrne D. N., Buchmann S. L., and Spangler H. G. (1988). Relationship between wing loading, wingbeat frequency and body mass in homopterous insects. J. Exp. Biol. 135, 9–23. doi: 10.1242/jeb.135.1.9
Cappa F., Cini A., Bortolotti L., Poidatz J., and Cervo R. (2021). Hornets and honey bees: a coevolutionary arms race between ancient adaptations and new invasive threats. Insects 12, 1037. doi: 10.3390/insects12111037
Carisio L., Cerri J., Lioy S., Bianchi E., Bertolino S., and Porporato M. (2022). Impacts of the invasive hornet Vespa velutina on native wasp species: a first effort to understand population-level effects in an invaded area of Europe. J. Insect Conserv. 26, 663–671. doi: 10.1007/s10841-022-00405-3
Choi M. B., Martin S. J., and Lee J. W. (2012). Distribution, spread, and impact of the invasive hornet Vespa velutina in South Korea. Jl Asia-Pac Entomol 15, 473–477. doi: 10.1016/j.aspen.2011.11.004
Cini A., Cappa F., Petrocelli I., Pepiciello I., Bortolotti L., and Cervo R. (2018). Competition between the native and the introduced hornets Vespa crabro and Vespa velutina: a comparison of potentially relevant life-history traits. Ecol. Entomol 43, 351–362. doi: 10.1111/een.12507
Danforth B. N. (1989). The evolution of hymenopteran wings: the importance of size. J. Zool 218, 247–276. doi: 10.1111/j.1469-7998.1989.tb02536.x
Diéguez-Antón A., Escuredo O., Seijo M. C., and Rodríguez-Flores M. S. (2025). Long-term Vespa velutina nigrithorax pressure: honey bee risk survival to alien invasion. Apidologie 56, 1–16. doi: 10.1007/s13592-025-01158-z
Do Y., Park W. B., Park J. K., Kim C. J., and Choi M. B. (2022). Genetic and morphological variation of Vespa velutina nigrithorax which is an invasive species in a mountainous area. Sci. Rep. 12, 4737. doi: 10.1038/s41598-022-08756-2
Dudley R. (2002). The biomechanics of insect flight: form, function, evolution (Princeton, NJ: Princeton University Press).
Endler J. A. (1988). Frequency-dependent predation, crypsis and aposematic coloration. Philos. T R Soc. Lon. B Biol. Sci. 319, 505–523. doi: 10.1098/rstb.1988.0062
Herrera C., Williams M., Encarnação J., Roura-Pascual N., Faulhaber B., Jurado-Rivera J. A., et al. (2023). Automated detection of the yellow-legged hornet (Vespa velutina) using an optical sensor with machine learning. Pest Manag Sci. 79, 1225–1233. doi: 10.1002/ps.7296
Hoebeke E. R., Bartlett L. J., Evans M., Freeman B. E., and Wares J. P. (2024). First records of Vespa velutina (Lepeletier) (color form Nigrithorax) (hymenoptera: Vespidae) in North America, an invasive pest of domesticated honeybees. Proc. Ent Soc. Wash 126, 193–205. doi: 10.4289/0013-8797.126.2.193
Jonsson T. (2023). Micro-CT and deep learning: modern techniques and applications in insect morphology and neuroscience. Fronts Insect Sci. 3. doi: 10.3389/finsc.2023.1016277
Joron M. (2009). “Aposematic coloration,” in Encycl Insects (Cambridge, MA: Academic Press), 33–38. doi: 10.1016/B978-0-12-374144-8.00010-2
Keszthelyi S., Orsi-Gibicsár S., Pal-Fam F., Somfalvi-Tóth K., and Balog A. (2024). Colour polymorphism of cotton bollworm larvae as a function of the type of host plant providing its development. Front. Ecol. Evol. 12. doi: 10.3389/fevo.2024.1376435
Kovac H. and Stabentheiner A. (2012). Does size matter? Thermoregulation of ‘heavyweight’and ‘lightweight’wasps (Vespa crabro and Vespula sp.). Biol. Open 1, 848–856. doi: 10.1242/bio.20121156
Kwon O. and Choi M. B. (2020). Interspecific hierarchies from aggressiveness and body size among the invasive alien hornet, Vespa velutina nigrithorax, and five native hornets in South Korea. PLoS One 15, e0226934. doi: 10.1371/journal.pone.0226934
Landolt P. J., Sierra J. M., Unruh T. R., and Zack R. S. (2010). A new species of Vespula, and first record of Vespa crabro L. (Hymenoptera: Vespidae) from Guatemala, Central America. Zootaxa 2629, 61–68. doi: 10.11646/zootaxa.2629.1.4
Laurino D., Lioy S., Carisio L., Manino A., and Porporato M. (2019). Vespa velutina: An alien driver of honey bee colony losses. Diversity 12, 5. doi: 10.3390/d12010005
Lioy S., Carisio L., Manino A., and Porporato M. (2023). Climatic niche differentiation between the invasive hornet Vespa velutina nigrithorax and two native hornets in Europe, Vespa crabro and Vespa orientalis. Diversity 15, 495. doi: 10.3390/d15040495
Marden J. H. (1987). Maximum lift production during takeoff in flying animals. J. Exp. Biol. 130, 235–258. doi: 10.1242/jeb.130.1.235
Mattila H. R., Otis G. W., Billen J., Nguyen L. T., and Shimano S. (2022). Comparison of the external morphology of the sternal glands for hornets in the genus Vespa. Biology 11, 245. doi: 10.3390/biology11020245
Methven K., Jeffords M., Weinzierl R., and McGiffen K. (1995). How to collect and preserve insects (Illinois Natural History Special Publication no. 17) Champaign, IL, Illinois Natural History Survey.
Monceau K., Bonnard O., and Thiéry D. (2014). Vespa velutina: a new invasive predator of honeybees in Europe. J. Pest Sci. 87, 1–16. doi: 10.1007/s10340-013-0537-3
Monceau K., Maher N., Bonnard O., and Thiéry D. (2015). Evaluation of competition between a native and an invasive hornet species: do seasonal phenologies overlap? B Entomol Res. 105, 462–469. doi: 10.1017/S0007485315000280
Moraes S. S., Söderholm M. S., Aguiar T. M., Freitas A. V., and Sihvonen P. (2023). Micro-CT imaging in species description: exploring beyond sclerotized structures in lichen moths (Lepidoptera: Erebidae, Arctiinae, Lithosiini). PeerJ 11, e15505. doi: 10.7717/peerj.15505
Onofre N., Portugal e Castro M. I., Nave A., Cadima I. S. P., Ferreira M., and Godinho J. (2023). On the evidence of the European Bee-Eater (Merops apiaster) as a predator of the yellow-legged hornet (Vespa velutina) and its possible contribution as a biocontrol agent. Animals 13, 1906. doi: 10.3390/ani13121906
Otis G. W., Taylor B. A., and Mattila H. R. (2023). Invasion potential of hornets (Hymenoptera: Vespidae: Vespa spp.). Front. Ins Sci. 3. doi: 10.3389/finsc.2023.1145158
Park W. B., Do Y., and Choi M. B. (2024). Ecological integration of Vespa velutina Lepeletier 1836 in South Korea: a comprehensive study of co-occurrence and interaction patterns with native social wasps. Pest Manag Sci. 80, 6201–6210. doi: 10.1002/ps.8348
Perrard A., Arca M., Rome Q., Muller F., Tan J., Bista S., et al. (2014). Geographic variation of melanisation patterns in a hornet species: genetic differences, climatic pressures or aposematic constraints? PLoS One 9, e94162. doi: 10.1371/journal.pone.0094162
Perrard A., Carpenter J., Kojima J. I., and Villemant C. (2013). Phylogeny of hornets: a total evidence approach (Hymenoptera, Vespidae, Vespinae, Vespa). J. Hymenopt Res. 32, 1–15. doi: 10.3897/jhr.32.4685
Perrard A., Lopez-Osorio F., and Carpenter J. M. (2016). Phylogeny, landmark analysis and the use of wing venation to study the evolution of social wasps (Hymenoptera: Vespidae: Vespinae). Cladistics 32, 406–425. doi: 10.1111/cla.12138
Robinet C., Suppo C., and Darrouzet E. (2017). Rapid spread of the invasive yellow-legged hornet in France: the role of human-mediated dispersal and the effects of control measures. J. Appl. Ecol. 54, 205–215. doi: 10.1111/1365-2664.12724
Rome Q., Perrard A., Muller F., Fontaine C., Quilès A., Zuccon D., et al. (2021). Not just honeybees: predatory habits of Vespa velutina (Hymenoptera: Vespidae) in France. Ann. Soc. Entomol Fr 57, 1–11. doi: 10.1080/00379271.2020.1867005
San Ha N., Truong Q. T., Goo N. S., and Park H. C. (2013). Relationship between wingbeat frequency and resonant frequency of the wing in insects. Bioinspir Biomim 8, 46008. doi: 10.1088/1748-3182/8/4/046008
Sauvard D., Imbault V., and Darrouzet É. (2018). Flight capacities of yellow-legged hornet (Vespa Velutina Nigrithorax, Hymenoptera: Vespidae) workers from an invasive population in Europe. PLoS One 13, e0198597. doi: 10.1371/journal.pone.0198597
Shaw F. R. and Weidhaas J. Jr. (1956). Distribution and habits of the giant hornet in North America. J. Econ Entomol 49, 275–275. doi: 10.1093/jee/49.2.275
Smith-Pardo A. H., Carpenter J. M., and Kimsey L. (2020). The diversity of hornets in the genus Vespa (Hymenoptera: Vespidae; Vespinae), their importance and interceptions in the United States. Insect Syst. Diver 4, 2. doi: 10.1093/isd/ixaa006
Snyder W. E. and Evans E. W. (2006). Ecological effects of invasive arthropod generalist predators. Annu. Rev. Ecol. Evol. Syst. 37, 95–122. doi: 10.1146/annurev.ecolsys.37.091305.110107
Tan K., Radloff S. E., Li J. J., Hepburn H. R., Yang M. X., Zhang L. J., et al. (2007). Bee-hawking by the wasp, Vespa velutina, on the honeybees Apis cerana and A. mellifera. Naturwissenschaften 94, 469–472. doi: 10.1007/s00114-006-0210-2
Taylor B. A., Tembrock L. R., Sankovitz M., Wilson T. M., Looney C., Takahashi J., et al. (2024). Population genomics of the invasive Northern Giant Hornet Vespa mandarinia in North America and across its native range. Sci. Rep. 14, 10803. doi: 10.1038/s41598-024-61534-0
Team R. D. C. (2010). R: a language and environment for statistical computing (Austria: Vienna). Available online at: https://www.R-project.org/ (Acessed January 21, 2025).
Toulkeridou E., Gutierrez C. E., Baum D., Doya K., and Economo E. P. (2023). Automated segmentation of insect anatomy from micro-CT images using deep learning. Nat. Sci. 3, e20230010. doi: 10.1002/ntls.20230010
Wei T. and Simko V. (2017). R package “corrplot”: visualization of a correlation matrix (version 0.84). Available online at: https://github.com/taiyun/corrplot (Acessed January 21, 2025).
Keywords: colourization, comparative morphology, external morphology, flight abilities, general predators
Citation: Sipos T, Kolics B, Kolics-Horváth É, Donkó T, Csóka Á, Kozma-Bognár K, Kovács A, Farkas S, Somfalvi-Tóth K and Keszthelyi S (2025) Comparative morphological analysis of yellow-legged hornet (Vespa velutina nigrithorax) and European hornet (Vespa crabro) based on modern imaging techniques. Front. Ecol. Evol. 13:1624744. doi: 10.3389/fevo.2025.1624744
Received: 07 May 2025; Accepted: 28 July 2025;
Published: 26 August 2025.
Edited by:
Hugh David Loxdale, Cardiff University, United KingdomReviewed by:
Artúr Botond Csorba, Sapientia Hungarian University of Transylvania, RomaniaJames Carpenter, American Museum of Natural History, United States
Copyright © 2025 Sipos, Kolics, Kolics-Horváth, Donkó, Csóka, Kozma-Bognár, Kovács, Farkas, Somfalvi-Tóth and Keszthelyi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tamás Sipos, c2lwb3MudGFtYXM4QHVuaS1tYXRlLmh1