 Letícia R. Menezes1
Letícia R. Menezes1 João Paulo L. F. Cairo2
João Paulo L. F. Cairo2 Ana Maria Costa-Leonardo3Maria Teresa Pedrosa Silva Clerici4Isabela da Costa Barreto1Bianca Suriano Francisco dos Santos1
Ana Maria Costa-Leonardo3Maria Teresa Pedrosa Silva Clerici4Isabela da Costa Barreto1Bianca Suriano Francisco dos Santos1 Alberto Arab1*
Alberto Arab1*- 1Termitolab, Centro de Ciências Naturais e Humanas, Universidade Federal do ABC (UFABC), São Bernardo do Campo, Brazil
- 2Department of Chemistry, University of York, York, United Kingdom
- 3Departamento de Biologia Geral e Aplicada, Universidade Estadual Paulista (UNESP), Rio Claro, Brazil
- 4Department of Food Technology, School of Food Engineering, University of Campinas (UNICAMP), Campinas, Brazil
Introduction: Termites rely on complex gut microbiomes to digest lignocellulosic substrates, with dietary specialization potentially shaping microbial diversity and function. However, how gut microbiota respond to dietary shifts in generalist versus specialist termites remains poorly understood, especially among neotropical species.
Methods: We evaluated microbiota composition and enzymatic function in two higher termite species with contrasting feeding strategies: the polyphagous Silvestritermes euamignathus and the litter-feeding specialist Cornitermes cumulans. Workers from four colonies per species were subjected to artificial diets differing in fiber complexity. Gut samples were analyzed via 16S rRNA gene amplicon sequencing and enzymatic assays targeting lignocellulolytic activity. Bioinformatic and statistical analyses were performed to assess diversity, taxonomic shifts, and functional predictions.
Results: Silvestritermes euamignathus maintained stable microbial composition and enzymatic activity across diets, whereas C. cumulans exhibited shifts in bacterial abundance and a marked reduction in enzymatic function when exposed to complex artificial diets. Despite some taxon-specific responses in C. cumulans, overall microbial beta diversity remained stable in both species. Functional predictions revealed broader metabolic capacity and higher CAZyme expression in the generalist species under complex diets.
Discussion: Our results suggest that microbial diversity and functional redundancy in S. euamignathus confer greater resilience to dietary perturbation, while C. cumulans displays limited plasticity consistent with dietary specialization. These findings have ecological implications, indicating that generalist termites may be better equipped to cope with environmental and resource shifts under changing climate scenarios.
1 Introduction
Termites are key decomposers in tropical ecosystems, relying on complex gut microbiomes to digest lignocellulosic materials. Due to differences in the composition of intestinal microorganisms, termites are classified into non-Termitidae termites and Termitidae, formerly called “lower termites” and “higher termites” (Carrijo et al., 2023). While non-termitid species primarily consume wood and depend on a combination of protists, bacteria, and archaea for digestion, termitids (Termitidae) depend almost exclusively on a highly diverse bacterial community. This microbial diversification has been closely linked to the evolutionary success of termitids, enabling them to exploit a wide range of ecological niches and feeding substrates (Brune, 2014; Chouvenc et al., 2021; Arab et al., 2024).
Feeding habits among termitids vary considerably, ranging from dietary specialists that primarily consume litter, grass, or soil to polyphagous generalists capable of accessing a broader range of lignocellulosic resources. For instance, the gut of wood-feeding termitids is typically enriched in Fibrobacterota and Spirochaetota, which contribute strongly to cellulolytic activity. In contrast, grass- and litter-feeding termites tend to harbor microbiomes dominated by Bacillota (formerly Firmicutes), which are better suited to processing hemicellulose (Mikaelyan et al., 2015; Menezes et al., 2018). This trophic diversity likely co-evolved with distinct microbial assemblages and enzymatic capabilities across termite lineages. In a previous study, we found that diversity of gut bacteria was higher in generalist than in specialist termitids (Moreira et al., 2018). As dietary complexity increases, a broader and more resilient repertoire of symbionts may be needed to metabolize structurally and chemically heterogeneous substrates, providing metabolic flexibility under fluctuating environmental conditions. In contrast, homogeneous diets may favor functional redundancy and lead to lower microbial diversity due to increased symbiont competition (Macke et al., 2017).
Microbiota plasticity is recognized as a critical factor in insect’s ability to cope with dietary shifts (Engel and Moran, 2013). In termites, previous studies suggest that changes in diet can lead to rapid, but often reversible, shifts in gut microbial communities (Miyata et al., 2007; Wang et al., 2016). However, most of this research has focused on a limited number of species, and little is known about the flexibility of gut microbiota and associated enzymatic traits in neotropical termites with contrasting diets.
Here, we investigate how gut microbiota structure and enzymatic function respond to dietary manipulation in two neotropical termitids: Silvestritermes euamignathus, a polyphagous species with a broad feeding repertoire, including litter, wood, and the carton nest of other termites (Mathews, 1977), and Cornitermes cumulans, a litter specialist (Coles De Negret and Redford, 1982) (Figures 1A, B). We hypothesize that (1) S. euamignathus will exhibit greater stability in both microbial composition and enzymatic activity across diets, reflecting its dietary flexibility, while (2) C. cumulans will show significant microbial shifts affecting its functional profile when subjected to different diets. By comparing microbiota diversity, taxonomic responses, and enzymatic profiles across dietary treatments, we aim to better understand the ecological and evolutionary mechanisms related to digestive resilience in termites.
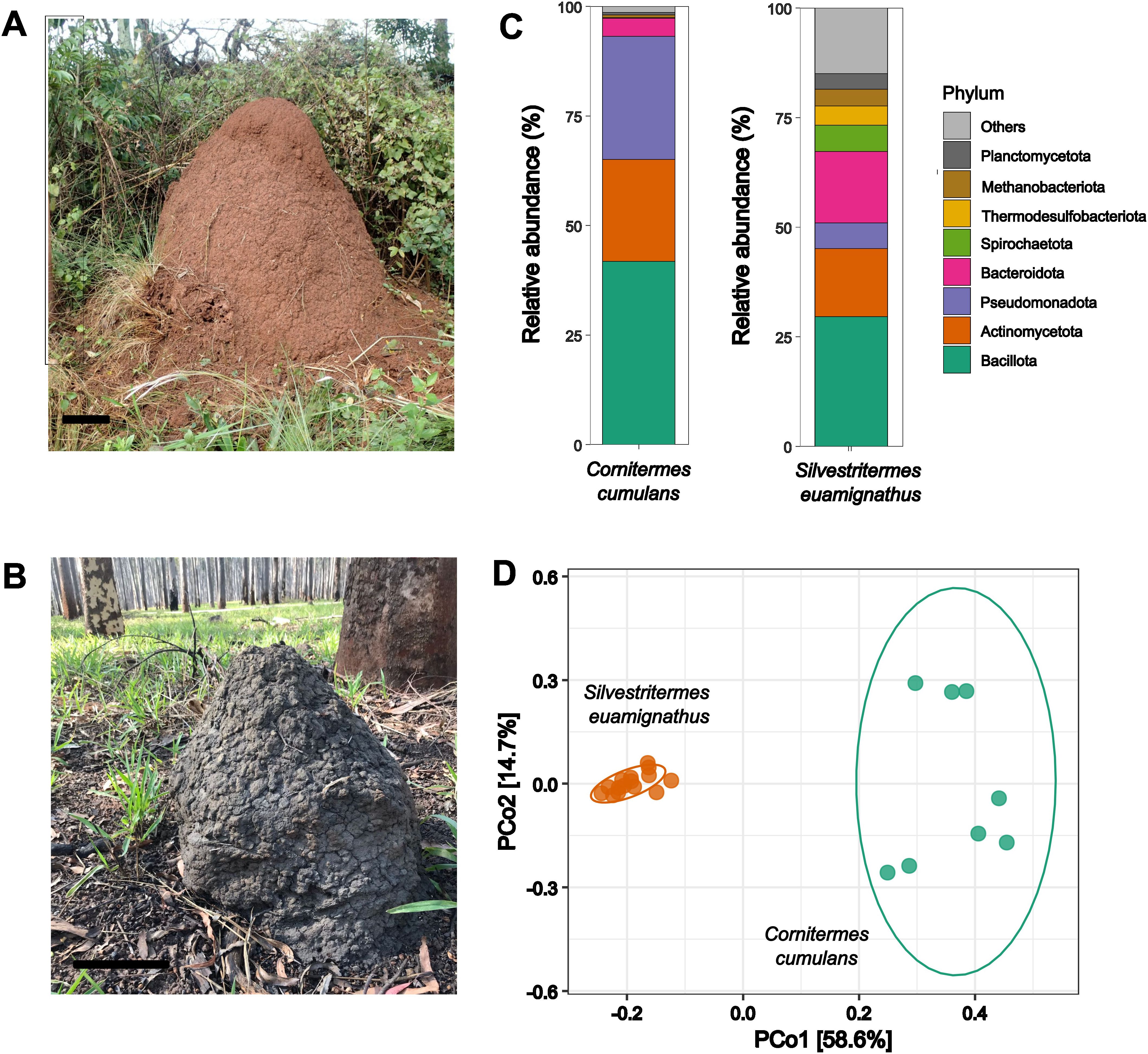
Figure 1. Epigeal nests of (A) Silvestritermes euamignathus and (B) Cornitermes cumulans. (C) Relative abundance of major bacterial phyla in the worker gut microbiota. (D) Ordination plot showing differences of gut bacterial composition between the two termite species. Scale bar= 10 cm.
2 Materials and methods
2.1 Termites and study site
Four colonies of Silvestritermes euamignathus and Cornitermes cumulans (Termitidae: Syntermitinae) were collected from pasture areas in the Campinas-SP region (22° 46’29.0”S, 47° 04’06.5”W). To maximize genetic heterogeneity and ecological independence, the colonies were collected within a ~1 km radius from a central coordinate point. This spatial scale is appropriate for these termite species, whose natural dispersal by flight is limited, ensuring that sampled colonies are genetically distinct. The nests were kept in 50 L plastic containers in a climate-controlled room at 26°C, under a 12-hour photoperiod cycle, with water and grass provided as food. Sampling was authorized by the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA), the enforcement agency of the Brazilian Ministry of the Environment (SISBIO permit no. 72676-4).
2.2 Feeding experiments
Feeding experiments were conducted using artificial diets varying in carbohydrate complexity. From each of four colonies, 50 workers and five soldiers were transferred to 10 cm Petri dishes containing sterilized, moistened sand and a 3 cm disc of analytical-grade pure agar (Sigma-Aldrich), supplemented with one of the following treatments to standardize the matrix across all diets:
Dextrose (40% dextrose + 15% agar), a monosaccharide representing a simple carbohydrate.
CreaFibe (40% CreaFibe QC 90 + 15% agar), comprising 84–88% cellulose, 12–16% hemicellulose, and <1% lignin (manufacturer’s specifications).
Arbocel® RC fine lignocellulose (40% Arbocel + 15% agar), containing 65–70% insoluble cellulose and >20% lignin (Hussein et al., 2017).
Control (15% agar only).
For each dietary treatment within each colony, we prepared 20 replicate dishes. These were distributed as follows: five dishes were used for gut microbiota sequencing, five for consumption bioassays, and ten for enzymatic assays.
After five days, surviving termites were enumerated, and their digestive tracts were dissected and stored at −22°C for enzymatic and molecular analyses (Menezes et al., 2018). An in-situ sample (50 workers per colony) was retained without experimental manipulation. Food consumption was quantified as the difference between initial and final fresh weight of diet discs, adjusted for humidity loss using control dishes (food without termites). Differences in consumption and survival among treatments were assessed via Generalized Linear Mixed Models (GLMMs) with Gaussian (consumption) and Poisson (survival) error distributions, implemented in the glmmTMB package (Brooks et al., 2017). Pairwise comparisons were conducted using estimated marginal means with the emmeans package (Lenth et al., 2023), and residual distributions were evaluated with the DHARMa package (Hartig, 2022).
2.3 DNA extraction
The entire guts of 50 workers from each colony subjected to each food treatment and from the in-situ sample were placed in 2 mL tubes containing 1 mL of lysis buffer (500 mM NaCl, 50 mM Tris-HCl, pH 8.0, 50 mM EDTA, and 4% sodium dodecyl sulfate). DNA extraction was performed using a bead-beating protocol as previously described [27]. DNA integrity was assessed via agarose gel electrophoresis (1.0% w/v), and quantification was conducted using a NanoDrop spectrophotometer by measuring absorbance at 260 nm. To optimize the amount of DNA recovered, replicate samples from each treatment were pooled prior to extraction.
2.4 16S rRNA library preparation, sequencing, and taxonomic assignment
The V4 region of the 16S rRNA gene was amplified using primers 515F (5′ TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGGTGCCAGCMGCCGCGGTAA 3′) and 806R (5′ GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGGACTACHVGGGTWTCTAAT 3′), with Illumina overhangs (underlined) (Menezes et al., 2018). Library preparation involved two PCR steps. In the first step, specific primers were used for each library with Phusion Polymerase (Thermo Scientific) and 40 ng of template DNA per sample. The amplification conditions were: initial denaturation at 98°C for 2 min, followed by 30 cycles of 98°C (30 s), 60.1°C (30 s), and 72°C (40 s), concluding with a final extension at 72°C for 5 min. The second PCR step incorporated Illumina sequencing adapters and dual index barcodes (Nextera Index Kit, Illumina) into the amplified libraries. This reaction used Phusion polymerase (Thermo Scientific) with 100 ng of purified PCR products from the first step as the template, along with indexing primers from Illumina. The amplification conditions included an initial denaturation at 98°C for 3 min, followed by 5 cycles of 98°C (30 s), 55°C (30 s), and 72°C (30 s), ending with a final extension at 72°C for 5 min. Each sample was amplified in triplicate. Pooled samples were purified using Agencourt AMPure Magnetic Beads (Beckman Coulter) and quantified with a Qubit Fluorometer 2.0 (Invitrogen). Sequencing was performed on the Illumina MiSeq platform at the Brazilian Bioethanol Science and Technology Laboratory (CTBE/CNPEM) using a MiSeq Reagent Kit V3 (600 cycles). Afterward, clean reads were processed using the DADA2 v1.16 pipeline (Callahan et al., 2016) with default parameters to generate amplicon sequence variants (ASVs). Taxonomic assignment was performed using the SILVA reference database (Quast et al., 2013). Tables generated by DADA2 were imported into phyloseq package (McMurdie and Holmes, 2013).
2.5 Microbial diversity and community structure analyses
We used R version 3.5.0 (R Core Team, 2021) to conduct analyses using different packages. Downstream analysis, including α- and β-diversity analysis was calculated using the microeco and MicrobiotaProcess packages (Liu et al., 2021; Xu et al., 2023). The ASVs table was merged with relevant metadata into a microeco object. Generalized linear mixed models (GLMM) were performed to check for overall significant differences of α-diversity estimates and the total number of ASVs among samples. We fitted multivariate generalized linear models (mvabund package) (Wang et al., 2012) to test the effects of the diet on the microbial relative abundance. Models were fitted with a negative binomial distribution using 999 bootstrap iterations. The manyglm function was used to carry out the analysis. Ordination analyses were used to evaluate microbial composition at the ASV level. We used GLMM analysis to identify ASVs that define the differences between microbiomes.
Functional predictions based on the 16S rRNA amplicon data were conducted using the Tax4Fun2 version 1.1.6 package (Aßhauer et al., 2015) and the reference profiles (Ref99NR) from the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. Similarities greater than 97% were considered orthologous KEGG groups (KOs). The output included CAZy encoded genes (https://www.cazy.org/).
2.6 Assays of lignocellulolytic activity of termite guts
The enzymatic activity against cellulose and hemicellulose substrates was evaluated using crude enzyme extracts from the gut of workers from four colonies, each subjected to different dietary treatments, following the methodology described by Franco Cairo et al. (2011). The assays aimed to assess the activity of the soluble fraction of protein extracts against both natural polysaccharides and synthetic oligosaccharides with varying monomer compositions and branching patterns. Crude protein extraction was performed using the guts of 50 workers from each colony. The samples were homogenized in 2 mL of 100 mM sodium acetate buffer (pH 5.5) and then centrifuged at 20,100 × g for 30 minutes at 4°C. The supernatant was collected, and 1 µL of Protease Inhibitor Cocktail (Anresco) was added per mL of crude extract to prevent protein degradation. Protein concentration in each extract was determined using the Bradford method (Bradford, 1976).
Each enzymatic reaction consisted of 10 µL of crude protein extract (3.2μg/μL), incubated at 37°C for 40 minutes with 40 µL of 50 mM sodium acetate buffer (pH 5.5) and 50 µL of a 0.5% specific natural polysaccharide solution (in water), performed in triplicate (Franco Cairo et al., 2011). The carbohydrate substrates used in the enzymatic assays included CMC (carboxymethyl cellulose, low viscosity) (β-1,4-carboxymethylglucan), β-glucan from barley (β-1,4-glucan), xylan from oat spelt (β-1,4-xylan), rye arabinoxylan (α-2,3-arabinose-β-1,4-xylan) from citrus, and pNP-G (4-nitrophenyl β-1,4-D-glucopyranoside). To terminate the enzymatic reactions, 100 µL of dinitrosalicylic acid (DNSA) was added, followed by heating at 99°C for 5 minutes. The resulting color change was measured at 540 nm using a TECAN M2000 plate reader. Blank reactions were prepared using the same procedure, except that DNSA was added before incubation with the protein extract. Enzymatic activity was expressed in mmol of glucose equivalents produced per mg of protein. For the assays using synthetic oligosaccharide the same enzymatic reaction was performed as described above, however using 2 mM pNP-G (4-nitrophenyl β-1,4-D-glucopyranoside) as substrate. The reaction was stopped with 100 µL of 1M sodium bicarbonate, followed by heating at 37°C for 30 minutes. The color change was analyzed at 412 nm using a TECAN M2000 plate reader. p-nitrophenyl were used for standard curve construction. Enzymatic activity was expressed in mmol of p-nitrophenyl released per mg of protein. To evaluate the effect of diet on the enzymatic activity of termite workers, a Wilks’ Lambda-type nonparametric multivariate inference was conducted using the npmv package (Ellis et al., 2017).
3 Results
3.1 The gut microbiota of Silvestritermes euamignathus and Cornitermes cumulans
We identified a total of 2,222 bacterial ASVs (508,218 reads) and 90 archaeal ASVs (18,147 reads) in the gut microbiota of S. euamignathus workers, as well as 982 bacterial ASVs (289,091 reads) and 52 archaeal ASVs (14,812 reads) in C. cumulans workers (Supplementary Tables S1, S2). The most abundant phyla—Bacillota, Actinomycetota, and Bacteroidota—accounted for 67% and 69% of sequence reads in S. euamignathus and C. cumulans, respectively (Figures 1C; Supplementary Figure S1). At the genus level, Papillibacter was the most abundant in S. euamignathus, while Acinetobacter dominated in C. cumulans (Supplementary Figure S1A). The gut microbiota composition differed significantly between these termite species (manyglm; Dev = 35.21; df = 1; p < 0.001) (Figure 1D), with only 6.3% of ASVs shared between them (Supplementary Figure S1B). Diversity indexes were not significantly different between the two termite species (Supplementary Figure S1C; Supplementary Table S3).
3.2 Diet alters bacterial abundance in C. cumulans but not in polyphagous S. euamignathus
Diet variation had no significant effect on food consumption or survival in either the litter-feeding Cornitermes cumulans or the polyphagous Silvestritermes euamignathus (Supplementary Figure S2; Supplementary Table S4). However, in C. cumulans, artificial diets led to notable shifts in both diversity (Supplementary Figures S3, S4) and relative abundance of specific bacterial taxa, particularly within the phyla Bacillota and Pseudomonadota, which varied across feeding regimes. In contrast, S. euamignathus showed no substantial changes in taxonomic abundance across diets (Figures 2A, B; Supplementary Tables S1, S2). It is worth noting that no data were available for the agar-only control in C. cumulans, as individuals from this treatment did not feed and suffered complete mortality during the assay. Despite these taxonomic fluctuations in C. cumulans, overall gut microbiota composition remained relatively stable in both species, as indicated by non-significant results from multivariate analyses (S. euamignathus: Deviance = 9.292, p = 0.068; C. cumulans: Deviance = 4.890, p = 0.092) (Figures 2C, D). These results suggest that C. cumulans exhibits some taxon-specific sensitivity to dietary changes, while maintaining a broadly stable community structure, whereas S. euamignathus displays resilience at both compositional and abundance levels under dietary stress.
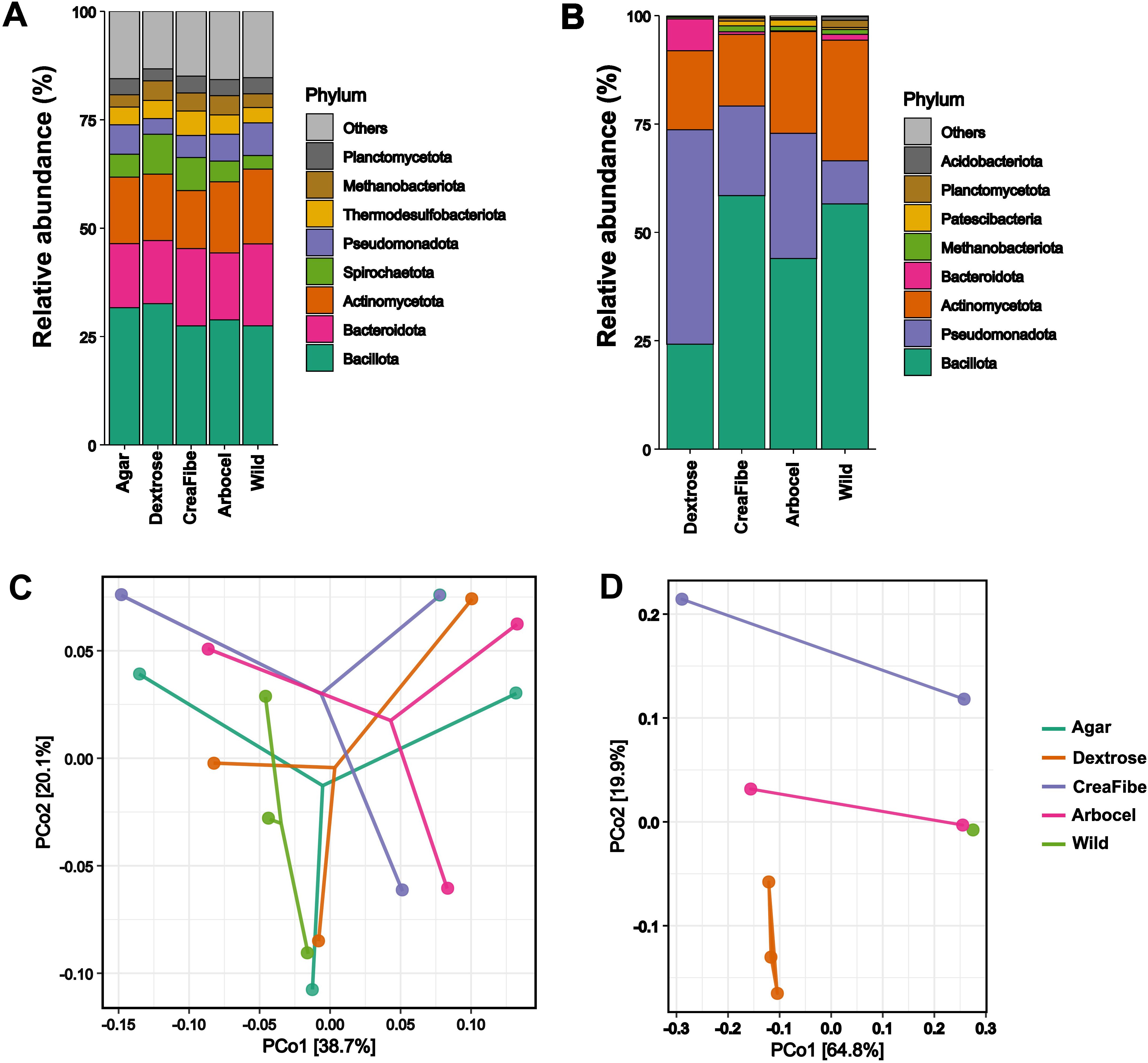
Figure 2. Effect of dietary treatments on the gut microbiota of Silvestritermes euamignathus and Cornitermes cumulans. Relative abundance of bacterial phyla in the gut microbiota of S. euamignathus (A) and C. cumulans (B) across different diets. Principal Coordinates Analysis (PCoA) based on weighted Unifrac distances showing beta diversity patterns for (C) S. euamignathus and (D) C. cumulans. Wild treatment consists in workers without experimental manipulation. We did not detect consumption of the control treatment (pure agar) by workers of C. cumulans. The agar-only control was excluded for C. cumulans due to lack of feeding and complete worker mortality during the assay.
3.3 Enzymatic responses reveal greater dietary resilience in S. euamignathus
The predicted functional inference of gut microbiota revealed differences between the two termite species in response to dietary treatments. In S. euamignathus, the dextrose diet strongly induced enzymes involved in simple sugar metabolism, including alpha-amylase (EC. 3.2.1.1) and alpha-glucosidase (EC. 3.2.1.20) (Figure 3A). However, individuals fed complex fiber-rich diets such as CreaFibe and Arbocel, or the wild treatment, showed moderate to higher profiles of ligninases (EC. 1.11.1.9 and EC. 1.11.1.15) and hemicellulases (EC. 3.2.1.10 and EC. 3.2.1.23) (Supplementary Table S5). Enzymatic activity assays also showed consistent profiles across dietary treatments, although individuals fed the dextrose diet exhibited slightly elevated activity for some substrates, retaining higher activity on β-glucan and pNP-G substrates even when exposed to artificial diets (Wilks’ Lambda = 7.91; df = 15, 36.29; p < 0.001) (Figure 3C; Supplementary Table S6).
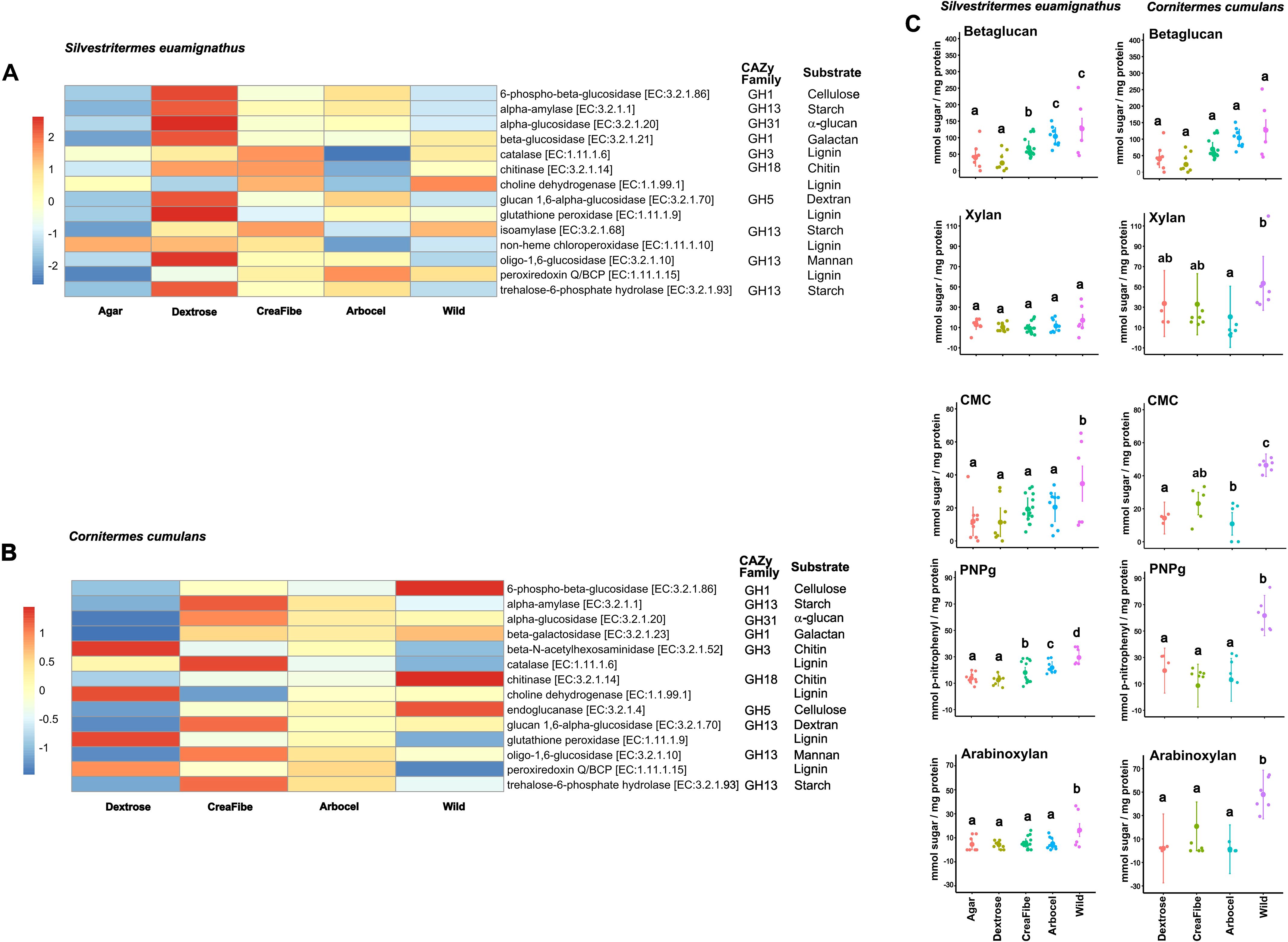
Figure 3. Functional and enzymatic responses of termite gut microbiota to dietary treatments. Heatmaps showing the relative abundance of predicted microbial enzymes related to carbohydrate and lignin degradation, inferred using Tax4Fun2 from from 16S rRNA gene profiles, for Silvestritermes euamignathus (A) and Cornitermes cumulans (B). Abundances were z-score normalized across rows to facilitate visualization of relative differences in enzyme abundance among treatments. (C) Quantification of specific enzymatic activities, expressed as mmol of sugar released per mg of protein. These values represent the activity of the soluble fraction normalized by total protein concentration. Substrates abbreviations: CMC (carboxymethyl cellulose) (β-1,4-carboxymethylglucan); β-glucan (β-1,4-glucan), Xylan (β-1,4-xylan), Arabinoxylan (α−2,3-arabinose-β-1,4-xylan), and pNP-G (4-nitrophenyl – β-1,4 -D-glucopyranoside). In the plot, the modelled means are represented by large circles, bars are the modelled 95% confidence intervals, and the model-adjusted individual response values are represented by the small dots. Different letters indicate significance at P< 0.05 (lsmeans pairwise analysis). Wild treatment consists in workers without experimental manipulation.
In contrast, Cornitermes cumulans displayed a pronounced reduction of enzymes involved in polysaccharide degradation—particularly those related to starch, cellulose, and hemicellulose—when exposed to artificial diets (Figure 3B; Supplementary Table S7). This functional suppression was reflected in significantly lower enzymatic activities against substrates such as xylan, CMC, PNPg, and arabinoxylan compared to wild-fed individuals (Wilks’ Lambda = 3.14; df = 20, 130.29; p < 0.001) (Figure 3C; Supplementary Table S8). Altogether, these results demonstrate that S. euamignathus is more resilient to dietary shifts, maintaining functional digestive enzyme activity across a range of nutritional environments. C. cumulans, in contrast, exhibits more pronounced reductions in enzymatic performance under simplified or artificial diets, suggesting a narrower metabolic scope and lower tolerance to dietary change.
4 Discussion
Our findings demonstrate that the polyphagous termite Silvestritermes euamignathus exhibits greater stability in both gut microbiota composition and enzymatic function across dietary treatments when compared to the litter-feeding specialist Cornitermes cumulans. This supports the view that high microbial diversity enhances host resilience to nutritional variability (Bertino-Grimaldi et al., 2013; Macke et al., 2017). The stability observed in S. euamignathus aligns with patterns reported in other generalist insects, where diverse microbial communities provide metabolic flexibility under novel or suboptimal diets (Shukla et al., 2016; Tinker and Ottesen, 2016). In contrast, C. cumulans exhibited pronounced shifts in the relative abundance of gut bacteria, similar to patterns seen in other termites with narrow dietary specialization (Benjamino et al., 2018), alongside a marked reduction in enzymatic activity when exposed to artificial diets. These findings suggest that the gut symbiotic system of C. cumulans is adapted to a more restricted nutritional niche, consistent with its evolutionary history as a litter specialist.
Our results build upon previous studies demonstrating that termite gut microbiomes are influenced by feeding preferences and dietary specialization (Mikaelyan et al., 2015). In wood-feeding termites, dietary shifts often lead to transient changes in microbial composition, which tend to revert once the native diet is restored (Miyata et al., 2007; Wang et al., 2016; Su et al., 2017). Here, we expand this understanding by showing that S. euamignathus, a generalist feeder, maintains both a stable microbiota and consistent enzymatic profiles across distinct dietary conditions, suggesting a flexible and functionally redundant symbiotic system. In contrast, C. cumulans exhibits reduced microbial diversity and decreased enzymatic activity when subjected to dietary changes, patterns consistent with the limited plasticity observed in other dietary specialists (Colman et al., 2012; Pérez-Cobas et al., 2015; Su et al., 2016; Macke et al., 2017). Notably, the relative abundance of Firmicutes (recently reclassified as Bacillota), a group known for its role in lignocellulose degradation in litter-feeding termites (Moreira et al., 2018), varied significantly across diets in C. cumulans. Disruption of these key symbionts under non-native diets may impair digestive efficiency and reflect the narrow functional breadth of this species’ gut microbial community.
Importantly, our data reveal that dietary complexity influences not only microbial composition but also functional gene predictions and host enzymatic responses. In S. euamignathus, complex diets (e.g., CreaFibe, Arbocel, and the wild treatment) triggered broader CAZy functional profiles and elevated expression of lignocellulolytic enzymes. These responses likely reflect adaptation to a naturally heterogeneous diet and the capacity to regulate microbial function according to substrate complexity (presence of hemicellulose and lignin). Similar flexibility in functional gene expression was observed in Odontotermes yunnanensis, a fungus-cultivating termite, where metagenomic data revealed high metabolic capacity and CAZyme diversity adapted to different plant matter (Liu et al., 2013). By contrast, C. cumulans showed functional suppression and reduced enzyme activity when fed artificial fiber-rich diets, indicating limited capacity to upregulate digestion in response to unfamiliar or compositionally complex substrates.
These findings highlight that resilience to dietary shifts is not only a matter of microbial diversity, but also of functional plasticity, the ability to activate or suppress microbial and enzymatic pathways in response to nutritional cues (Boucias et al., 2013; Benjamino et al., 2018). This adaptive flexibility may contribute to the ecological success of generalists like S. euamignathus in disturbed or changing environments.
Finally, these patterns carry important ecological and evolutionary implications. In the context of global climate change and growing anthropogenic pressures, termite species with flexible gut microbiomes may be better equipped to cope with shifts in resource availability. As plant biomass becomes more abundant or variable under warming scenarios, generalist species could expand into new habitats, aided by the metabolic versatility and functional redundancy of their symbiotic communities. In contrast, specialists like C. cumulans may be more vulnerable to environmental change, particularly if their preferred substrates become scarce—potentially disrupting decomposition processes and reshaping local species distributions (Gilbert et al., 2015; Buczkowski and Bertelsmeier, 2017). Recent comparative analyses of termite gut metagenomes have shown that, although many metabolic pathways are conserved across lineages, diet plays a central role in modulating the relative abundance and expression of carbohydrate-active enzymes (CAZymes) (Arora et al., 2022), directly influencing lignocellulose breakdown efficiency and ecological adaptability.
A methodological limitation of this study is that enzymatic activity was assessed from whole-gut extracts, which include both host-derived and microbial enzymes. In termitids, endogenous cellulases are primarily produced in the salivary glands and midgut (Ni and Tokuda, 2013), while microbial fermentation occurs mainly in the hindgut. Therefore, the enzyme activities reported here represent a combined signal from both sources, and we cannot fully separate their individual contributions. Future studies employing metatranscriptomics, host gene knockdown, or enzyme localization assays would be valuable to disentangle microbial and endogenous enzyme activity and better define their respective roles in lignocellulose degradation.
Data availability statement
The original contributions presented in the study are publicly available. Termite metagenome sequences are deposited at the NCBI Sequence Read Archive, accession PRJNA1256848.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
LM: Formal analysis, Investigation, Methodology, Writing – original draft. JC: Methodology, Writing – review & editing. AC-L: Conceptualization, Writing – review & editing. MC: Methodology, Writing – review & editing. IC: Methodology, Writing – original draft. BS: Methodology, Writing – original draft. AA: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by the São Paulo Research Foundation (FAPESP), grants # 2015/21497-6 and 2018/22839-6.
Acknowledgments
We would like to thank Brazilian Biorenewables National Laboratory (LNBR/CNPEM) NGS Sequencing Facility for generating the metagenomics sequencing data described here. We also thank Nutrassim Food Ingredients and J. RETTENMAIER Latinoamericana Ltda. for providing CreaFibe and Arbolcel, respectively, for our study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. AI was used to refine English syntax.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1625443/full#supplementary-material
References
Aßhauer K. P., Wemheuer B., Daniel R., and Meinicke P. (2015). Tax4Fun: predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 31, 2882–2884. doi: 10.1093/bioinformatics/btv287, PMID: 25957349
Arab A., Paulino L. C., Rolim M. S. L., Menezes L. R., de, Oliveira M.H. de, dos Santos R. A. C., et al. (2024). The Gut Microbiome and Lignocellulose Digestion in Constrictotermes cyphergaster (Termitidae: Nasutitermitinae): A Termite Incorporating Lichen into Its Diet. Diversity (Basel) 16, 623. doi: 10.3390/D16100623/S1
Arora J., Kinjo Y., Šobotník J., Buček A., Clitheroe C., Stiblik P., et al. (2022). The functional evolution of termite gut microbiota. Microbiome 10, 78. doi: 10.1186/s40168-022-01258-3, PMID: 35624491
Benjamino J., Lincoln S., Srivastava R., and Graf J. (2018). Low-abundant bacteria drive compositional changes in the gut microbiota after dietary alteration. Microbiome 6, 86. doi: 10.1186/S40168-018-0469-5/FIGURES/10, PMID: 29747692
Bertino-Grimaldi D., Medeiros M. N., Vieira R. P., Cardoso A. M., Turque A. S., Silveira C. B., et al. (2013). Bacterial community composition shifts in the gut of Periplaneta americana fed on different lignocellulosic materials. Springerplus 2, 609. doi: 10.1186/2193-1801-2-609, PMID: 24324923
Boucias D. G., Cai Y., Sun Y., Lietze V. U., Sen R., Raychoudhury R., et al. (2013). The hindgut lumen prokaryotic microbiota of the termite Reticulitermes flavipes and its responses to dietary lignocellulose composition. Mol. Ecol. 22, 1836–1853. doi: 10.1111/MEC.12230, PMID: 23379767
Bradford M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3, PMID: 942051
Brooks M. E., Kristensen K., Benthem K. J. ,. V., Magnusson A., Berg C. W., Nielsen A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Brune A. (2014). Symbiotic digestion of lignocellulose in termite guts. Nat. Rev. Microbiol. 12, 168–180. doi: 10.1038/nrmicro3182, PMID: 24487819
Buczkowski G. and Bertelsmeier C. (2017). Invasive termites in a changing climate: A global perspective. Ecol. Evol. 7, 974–985. doi: 10.1002/ece3.2674, PMID: 28168033
Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., and Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods, 13 (7), 581–583., PMID: 27214047
Carrijo T. F., Engel M. S., Chouvenc T., Gile G. H., Mikaelyan A., Dedeine F., et al. (2023). A call to termitologists: it is time to abandon the use of “lower” and “higher” termites. Insectes Soc. 70, 295–299. doi: 10.1007/S00040-023-00929-0/FIGURES/2
Chouvenc T., Šobotník J., Engel M. S., and Bourguignon T. (2021). Termite evolution: mutualistic associations, key innovations, and the rise of Termitidae. Cell. Mol. Life Sci. 78, 2749–2769. doi: 10.1007/s00018-020-03728-z, PMID: 33388854
Coles De Negret H. R. and Redford K. H. (1982). The biology of nine termite species (Isoptera: Termitidae) from the Cerrado of Central Brazil. Psyche (Stuttg) 89, 81–106. doi: 10.1155/1982/36726
Colman D. R., Toolson E. C., and Takacs-Vesbach C. D. (2012). Do diet and taxonomy influence insect gut bacterial communities? Mol. Ecol. 21, 5124–5137. doi: 10.1111/J.1365-294X.2012.05752.X, PMID: 22978555
Ellis A. R., Burchett W. W., Harrar S. W., and Bathke A. C. (2017). Nonparametric inference for multivariate data: the R package npmv. J. Stat. Softw 76, 1–18. doi: 10.18637/JSS.V076.I04
Engel P. and Moran N. A. (2013). The gut microbiota of insects - diversity in structure and function. FEMS Microbiol. Rev. 37, 699–735. doi: 10.1111/1574-6976.12025, PMID: 23692388
Franco Cairo J. P. L., Leonardo F. C., Alvarez T. M., Ribeiro D. A., Büchli F., Costa-Leonardo A. M., et al. (2011). Functional characterization and target discovery of glycoside hydrolases from the digestome of the lower termite Coptotermes gestroi. Biotechnol. Biofuels 4, 50. doi: 10.1186/1754-6834-4-50, PMID: 22081966
Gilbert S. F., Bosch T. C. G., and Ledón-Rettig C. (2015). Eco-Evo-Devo: developmental symbiosis and developmental plasticity as evolutionary agents. Nat. Rev. Genet. 16, 611–622. doi: 10.1038/nrg3982, PMID: 26370902
Hartig F. (2022). DHARMa: Residual diagnostics for hierarchical (multi-level/mixed) regression models. R Package 531, 532. doi: 10.32614/CRAN.package.DHARMa
Hussein S. M., Yokhana J. S., and Frankel T. L. (2017). Supplementing the feeds of layer pullets, at different ages with two different fiber sources improves immune function. Poult Sci. 96, 2718–2727. doi: 10.3382/PS/PEX051, PMID: 28371891
Lenth R. V., Bolker B., Buerkner P., Giné-Vázquez L., Herve M., Jung M., et al. (2023). Package “emmeans. Am. Statistician 34, 216–221. doi: 10.1080/00031305.1980.10483031
Liu C., Cui Y., Li X., and Yao M. (2021). Microeco: An R package for data mining in microbial community ecology. FEMS Microbiol. Ecol. 97, fiaa255. doi: 10.1093/femsec/fiaa255, PMID: 33332530
Liu N., Zhang L., Zhou H., Zhang M., Yan X., Wang Q., et al. (2013). Metagenomic Insights into Metabolic Capacities of the Gut Microbiota in a Fungus-Cultivating Termite (Odontotermes yunnanensis). PloS One 8, e69184. doi: 10.1371/JOURNAL.PONE.0069184, PMID: 23874908
Macke E., Tasiemski A., Massol F., Callens M., and Decaestecker E. (2017). Life history and eco-evolutionary dynamics in light of the gut microbiota. Oikos 126, 508–531. doi: 10.1111/OIK.03900
Mathews A. G. A. (1977). Studies on termites from the Mato Grosso State, Brazil. Brasil: Academia Brasileira de Ciências.
McMurdie P. J. and Holmes S. (2013). Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PloS One 8, e61217. doi: 10.1371/journal.pone.0061217, PMID: 23630581
Menezes L., Alvarez T. M., Persinoti G. F., Franco J. P., Squina F., Moreira E. A., et al. (2018). Food storage by the savanna termite Cornitermes cumulans (Syntermitinae): a strategy to improve hemicellulose digestibility? Microb. Ecol. 76, 492–505. doi: 10.1007/s00248-017-1128-2, PMID: 29270662
Mikaelyan A., Dietrich C., Köhler T., Poulsen M., Sillam-Dussès D., and Brune A. (2015). Diet is the primary determinant of bacterial community structure in the guts of higher termites. Mol. Ecol. 24, 5284–5295. doi: 10.1111/mec.13376, PMID: 26348261
Miyata R., Noda N., Tamaki H., Kinjyo K., Aoyagi H., Uchiyama H., et al. (2007). Influence of feed components on symbiotic bacterial community structure in the gut of the wood-feeding higher termite Nasutitermes takasagoensis. Biosci. Biotechnol. Biochem. 71, 1244–1251. doi: 10.1271/bbb.60672, PMID: 17485852
Moreira E. A., Alvarez T. M., Persinoti G. F., Paixão D. A. A., Menezes L. R., Cairo J. P. F., et al. (2018). Microbial communities of the gut and nest of the humus- and litter-feeding termite Procornitermes araujoi (Syntermitinae). Curr. Microbiol. 75, 1609–1618. doi: 10.1007/S00284-018-1567-0/FIGURES/3, PMID: 30209570
Ni J. and Tokuda G. (2013). Lignocellulose-degrading enzymes from termites and their symbiotic microbiota. Biotechnol. Adv. 31, 838–850. doi: 10.1016/j.bioteChadv.2013.04.005, PMID: 23623853
Pérez-Cobas A. E., Maiques E., Angelova A., Carrasco P., Moya A., and Latorre A. (2015). Diet shapes the gut microbiota of the omnivorous cockroach Blattella germanica. FEMS Microbiol. Ecol. 91. doi: 10.1093/femsec/fiv022, PMID: 25764470
Quast C., Pruesse E., Yilmaz P., Gerken J., Schweer T., Yarza P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/NAR/GKS1219, PMID: 23193283
R Core Team (2021). R core team (2021) (Vienna, Austria: R: A language and environment for statistical computing. R Foundation for Statistical Computing). Available online at: http://www.R-project.org (Accessed January 8, 2024).
Shukla S. P., Sanders J. G., Byrne M. J., and Pierce N. E. (2016). Gut microbiota of dung beetles correspond to dietary specializations of adults and larvae. Mol. Ecol 25, 6092–6106. doi: 10.1111/mec.13901, PMID: 27801992
Su L., Yang L., Huang S., Li Y., Su X., Wang F., et al. (2017). Variation in the gut microbiota of termites (Tsaitermes ampliceps) against different diets. Appl. Biochem. Biotechnol. 181, 32–47. doi: 10.1007/S12010-016-2197-2/FIGURES/5, PMID: 27457759
Su L. J., Yang L. L., Huang S., Su X. Q., Li Y., Wang F. Q., et al. (2016). Comparative gut microbiomes of four species representing the higher and the lower termites. J. Insect Sci. 16. doi: 10.1093/JISESA/IEW081, PMID: 27638955
Tinker K. A. and Ottesen E. A. (2016). The core gut microbiome of the American cockroach, Periplaneta americana, is stable and resilient to dietary shifts. Appl. Environ. Microbiol. doi: 10.1128/AEM.01837-16, PMID: 27590811
Wang Y., Naumann U., Wright S. T., and Warton D. I. (2012). Mvabund- an R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 3, 471–474. doi: 10.1111/j.2041-210X.2012.00190.x
Wang Y., Su L., Huang S., Bo C., Yang S., Li Y., et al. (2016). Diversity and resilience of the wood-feeding higher termite Mironasutitermes shangchengensis gut microbiota in response to temporal and diet variations. Ecol. Evol. 6, 8235–8242. doi: 10.1002/ece3.2497, PMID: 27878091
Keywords: termite gut microbiota, dietary plasticity, lignocellulose digestion, enzymatic resilience, termitidae
Citation: Menezes LR, Cairo JPLF, Costa-Leonardo AM, Clerici MTPS, da Costa Barreto I, dos Santos BSF and Arab A (2025) Dietary resilience of termite gut microbiota and enzymatic function reflects feeding strategy. Front. Ecol. Evol. 13:1625443. doi: 10.3389/fevo.2025.1625443
Received: 08 May 2025; Accepted: 15 July 2025;
Published: 01 August 2025.
Edited by:
Silvio Erler, Julius Kühn-Institut - Braunschweig, GermanyReviewed by:
Brittany Faye Peterson, Southern Illinois University Edwardsville, United StatesCristian Beza Beza, University of Minnesota Twin Cities, United States
Copyright © 2025 Menezes, Cairo, Costa-Leonardo, Clerici, da Costa Barreto, dos Santos and Arab. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alberto Arab, YWxiZXJ0b2FyYWJAZ21haWwuY29t