 Yannis Nio1Christelle Buchard1Franck Duval1Frédérique Mahéo1Ségolène Buzy1Anne Le Ralec2
Yannis Nio1Christelle Buchard1Franck Duval1Frédérique Mahéo1Ségolène Buzy1Anne Le Ralec2 Quentin Chesnais3Gabin Mardoc3Véronique Brault3Thierry Candresse4Ghislain Malatesta5Amélie Monteiro5Julie Jaquiéry1
Quentin Chesnais3Gabin Mardoc3Véronique Brault3Thierry Candresse4Ghislain Malatesta5Amélie Monteiro5Julie Jaquiéry1 Jean-Christophe Simon1*
Jean-Christophe Simon1*- 1INRAE, L’Institut Agro, Univ. Rennes, Unité Mixte de Recherche (UMR) 1349 IGEPP, Le Rheu, France
- 2Unité Mixte de Recherche (UMR) IGEPP, L’Institut Agro, Rennes, France
- 3INRAE, Univ. Strasbourg, Unité Mixte de Recherche (UMR) 1131 SVQV, Colmar, France
- 4Univ. Bordeaux, INRAE, Unité Mixte de Recherche (UMR) 1332 BFP, Villenave d’Ornon, France
- 5Institut Technique de la Betterave, Laon, France
Most phytophagous insect species are specialized to feed on a narrow range of host plants, typically within the same genus or family, and sometimes even on a single plant species. However, some insect taxa are able to feed on plants from different families and are therefore considered to be generalists. Nevertheless, these generalist species can sometimes form cryptic species complexes or differentiate into host-based populations or races. Moreover, the host breadth of generalists is often measured under laboratory conditions, which may not accurately reflect the more challenging natural environments they encounter, and thus may lead to an overestimation of generalism. In this study, we used a population genetics approach to test whether Myzus persicae, a highly polyphagous aphid, is composed of host-specialized populations or clones, or whether generalism is an intrinsic characteristic of the species as a whole. We sampled M. persicae over four consecutive years in northern France from a variety of host plants belonging to different botanical families. We found that populations of M. persicae in northern France were predominantly composed of superclones, namely multilocus genotypes identified in many copies. In particular, four superclones dominated M. persicae populations, occurring in high proportions on a broad range of unrelated host plants in each year of the study. Moreover, the array of characterized genotypes of M. persicae formed distinct genetic clusters, but with no clear association to specific host plants. This study shows that under natural conditions and at the very fine genetic level of a single clone, the generalist characteristic of M. persicae holds. Thus, M. persicae remains an exceptional example of a true generalist species.
Introduction
Phytophagous insects constitute one of the most diverse groups of organisms, with over half a million described species. This extraordinary diversification is largely attributed to their close associations with host plants (Futuyma and Agrawal, 2009). The majority of phytophagous insects exhibit a high degree of specialization, typically feeding on a narrow range of plants within a single genus or family, and in some cases, even a single plant species (Forister et al., 2015). Such specialization is thought to drive reproductive isolation and, ultimately, speciation (Drès and Mallet, 2002; Peccoud et al., 2009). As a result, host-plant specialization is considered the norm, while generalist species (i.e. those capable of utilizing a wide range of host plants), are relatively rare.
The evolutionary success of specialists over generalists has been the focus of extensive research, though no single explanation fully accounts for this phenomenon. Instead, it likely results from a complex interplay of ecological, evolutionary, and physiological factors. One prominent explanation involves co-evolution between plants and herbivorous insects (Ehrlich and Raven, 1964). Over evolutionary timescales, plants develop defense mechanisms, while insects evolve corresponding counter-adaptations, leading to increasingly fine-tuned interactions that promote specialization (Edger et al., 2015). Additionally, genetic trade-offs in host use may further reinforce specialization, as adaptation to a specific host plant often comes at the cost of reduced performance on alternative hosts (Futuyma and Moreno, 1988; Forister et al., 2015).
Despite the prevalence of specialization, there are notable exceptions: generalist species that thrive on a wide range of host plants spanning multiple botanical families. Examples include the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae), which feeds on over 350 plant species across 76 families (Tay et al., 2023), the whitefly Bemisia tabaci (Hemiptera: Aleyrodidae), with a host range exceeding 600 species from 74 families (De Barro et al., 2011), and the desert locust Schistocerca gregaria (Orthoptera: Acrididae), which consumes natural vegetation and a wide range of crops (Showler, 2002). The success of these generalists may stem from their broader tolerance for diverse environmental conditions, enhanced detoxification mechanisms, or flexible feeding behaviors. For instance, generalists usually have a range of detoxification enzymes that can neutralize many different plant chemicals, allowing them to feed on various plants without being harmed by their specific defenses (Govind et al., 2010; Simon et al., 2015). Genetic factors may also play a role in promoting generalism under certain conditions. Specialization often makes phytophagous insects highly dependent on the availability of their preferred host plants, which, in extreme cases, can lead to population declines or even extinction if those plants become scarce. Specialization is thus often associated with limited genetic diversity. In contrast, generalist species tend to maintain higher levels of genetic diversity, providing a broader foundation for natural selection to act upon. This genetic variability can enhance their capacity to adapt to new host plants and to fluctuating environmental conditions, increasing their resilience in unstable ecosystems such as agricultural landscapes (Hardy et al., 2020).
However, even among generalist species, differentiation can occur, leading to the formation of cryptic species complexes or host-associated populations. For instance, S. frugiperda comprises at least two genetically distinct strains: a corn strain and a rice strain. Despite being morphologically identical, these strains exhibit strong host plant preferences, with the corn strain primarily feeding on maize, sorghum, and cotton, while the rice strain specializes on rice and other grasses (Durand et al., 2022). Similarly, the pea aphid Acyrthosiphon pisum consists of at least 15 host-adapted biotypes, each specialized to feed on a restricted set of legume species (Peccoud et al., 2009, 2015). Furthermore, the host breadth of generalist insects is often assessed under controlled laboratory conditions, which may not fully capture the challenges they face in natural environments (Finlay-Doney and Walter, 2012). Factors such as predation, competition, and environmental variability can thus constrain their ability to exploit multiple hosts in the wild. As a result, truly generalist species may be even rarer than currently assumed.
The peach-potato aphid Myzus persicae has a very broad host range, encompassing more than 400 plant species, both cultivated and non-cultivated, belonging to more than 40 different botanical families (Blackman and Eastop, 2000; Bass et al., 2014). This species stands out from most other aphid species, which are typically specialized to feed on a limited number of host plants within the same plant family (Peccoud et al., 2010; Shih et al., 2023). The classification of M. persicae as a true generalist has been challenged in several studies, beginning with the identification of a tobacco-adapted form. Thus, certain populations that have specialized in feeding on tobacco exhibit distinct morphological and genetic differences from M. persicae sensu stricto (s.s.) and have been formally designated as M. persicae subsp. nicotianae (Blackman, 1987; Margaritopoulos et al., 2007). More recent genetic research, utilizing highly polymorphic markers or whole-genome sequencing, has revealed additional evidence of host-associated differentiation, with some genetic distinction observed among populations feeding on peach, pepper, and oilseed rape (Zamoum et al., 2005; Singh et al., 2021; Roy et al., 2022).
In the present study, we conducted an extensive sampling of M. persicae performed as part of a project aimed at identifying the sources of vectors and viruses responsible for the beet yellowing disease (Nio et al., 2025). Our main objective was to analyze the clonal diversity of M. persicae across different regions in northern France and on various host plants where this aphid can potentially overwinter. More specifically, we aimed to investigate whether M. persicae populations exhibited host-plant-based structuring and if so, to determine whether there is field evidence supporting the existence of clones capable of feeding on multiple plant species. By examining its genetic population structure and host-plant associations, this study was designed to assess whether M. persicae can truly be classified as a generalist.
Material and methods
Aphid sampling
Most aphid sampling was carried out in early spring (first or second week of April) for four consecutive years (2021 to 2024) on cultivated and non-cultivated plants in the northern half of France, with priority given to sugar beet production areas. M. persicae sampling was also conducted later in the season on spring and summer plants, which are annuals and not typically found during the winter months. Sampling was performed in a total of 16 French Departments, six of which (Aisne, Eure-et-Loir, Ille-et-Vilaine, Loiret, Oise, Somme) were sampled on each of the four years of the study (Supplementary Table S1). In addition, to assess the temporal and spatial distribution of M. persicae clones, we used samples collected in yellow or suction traps in the same areas as the aphid sampling on host plants. All sampled aphids were parthenogenetic females, whether winged or wingless. About 60 different plant species were inspected during these various surveys to look for aphids. Aphids of the Myzus genus were collected from 28 different plant species. Aphid species identification was primarily based on morphological criteria. In cases of ambiguity, molecular barcoding, through amplification and sequencing of the cytochrome oxidase gene fragment, was employed to confirm identification (Coeur d’acier et al., 2014).
Aphid genotyping
Aphid total genomic DNA was extracted from each individual in semi-deep well trays following the salting-out protocol (Miller et al., 1988), then suspended in 100 μL of ultrapure water. To genotype aphids, we used a set of 14 microsatellite markers developed in a previous study (Roy et al., 2022). For each DNA sample, we performed three multiplexed PCRs (two multiplexes of 5 loci and one multiplex of 4) with fluorescently labelled primers allowing us to generate the genetic profile of each individual at 14 microsatellite markers. Microsatellite loci were amplified in a final volume of 10 µL polymerase chain reaction (PCR) mix. The reaction mix included 2 µL of total DNA, 0.25–0.5 µM of each primer, 0.8 mM of a four dNTPs mixture, 2 mM MgCl2, 2 µL of PCR Buffer (Promega, Madison, USA), and 4.5 U of Taq DNA Polymerase (Promega, Madison, USA). PCR amplification was performed using an S1000 thermal cycler (Bio-Rad Laboratories) under the following cycling conditions: initial denaturation at 95°C for 5 min., followed by 30 cycles of denaturation at 95°C for 1 min., hybridization at 54°C for 1 min., and elongation at 72°C for 1 min. A final elongation step at 72°C for 10 minutes concluded the PCR. The PCR products were diluted (1.5 µL of product in 10 µL of water) and 1.5 µL of this diluted PCR product mixed with 6 µL of Hi-Di formamide (Applied Biosystems) containing 0.8% of the 500 LIZ DNA ladder (Applied Biosystems). Capillary electrophoresis was then performed using an ABI 3730 sequencer (Applied Biosystems). Alleles were automatically called using GENEMAPPER software (version 6, Applera Corp) based on the ladder. To ensure accuracy, the chromatograms of both the ladder and the PCR fragments were visually inspected to correct any potential misassignations resulting from the software’s automatic analysis. Individuals with missing data for two or more microsatellite loci were excluded from subsequent analyses. The combined information from the different loci formed the multilocus genotype (MLG).
Population genetic analyses
We used the R package “Rclone” (Bailleul et al., 2016) to assign multilocus genotypes (MLGs) to all aphids with complete genotypic data. For individuals with a single missing locus, MLGs were inferred based on the closest matching complete genotype. To assess the genetic structure of the dataset, we conducted individual-based Bayesian clustering using STRUCTURE v2.3.4 (Falush et al., 2003). To reduce potential biases from clonal redundancy, only one representative per unique MLG was included in the analysis. STRUCTURE was run for K values from 1 to 20, with 10 replicate runs per K value, using the admixture model and correlated allele frequencies. Each run consisted of a burn-in of 50,000 iterations followed by 300,000 MCMC iterations. The optimal number of clusters (K) was identified by evaluating the log-likelihood of the data [Pr(X|K)], the Delta-K method (Evanno et al., 2005), and the consistency of clustering patterns across runs, visualized using the R package “pophelper” (https://github.com/royfrancis/pophelper). After identifying the most relevant number of clusters, MLGs were assigned to a genetic cluster if their membership coefficient (q) exceeded a threshold of 0.51 using one of the STRUCTURE simulations. To assess the genetic differentiation among clusters, we used the R package “Hierfstat” (Goudet, 2005) to calculate pairwise FST values between clusters and FIS values within each cluster. The significance of these values was assessed by 1,000 bootstraps over loci as implemented in “Hierfstat”. We also assessed the genetic relationship between the different MLGs using the Cavalli-Sforza distance (Cavalli-Sforza and Edwards, 1967) calculated with the executable Population v1.2.23 (https://bioinformatics.org/populations/). A Neighbor-Joining tree based on the Cavalli-Sforza distance was built using the R package “ape” (Paradis and Schliep, 2018). Lastly, the number of different alleles between MLG pairs was calculated using a home-made script.
Results
Broad host range across diverse plant families
After confirming the taxonomic status of M. persicae, we identified individuals of this species on 25 host-plant species belonging to 10 plant families (Supplementary Table S2). Notably, M. persicae was most overrepresented on plants in the Brassicaceae family, with individuals found on seven Brassicaceae species, making it the most frequently encountered family.
Limited genetic diversity driven by dominant superclones
We were able to assign a MLG to 3,112 of the 4,676 genotyped individuals, (Supplementary Table S1) when excluding individuals with missing data at more than one locus. Most missing data resulted from PCR failures, likely due to degraded DNA in those aphids that remained in traps for up to a week before collection. A total of 412 different MLGs were found, of which 125 were represented by multiple individuals and 287 by single individuals (Figure 1). The G:N ratio (G=number of MLG and N=number of individuals) was 0.14, indicating low genetic diversity and a highly clonal structure. Four MLGs were particularly abundant, altogether representing about 56% of the individuals. These four MLGs (i.e. M1, M2, M7, M10), each with more than 200 individuals and a frequency of more than 10%, are referred to here as ‘superclones’. Forty-seven percent of aphids caught in either suction traps or yellow traps belonged to one of the four superclones, highlighting their good dispersal capacity. This was also supported by the widespread distribution of the four superclones, which were found throughout the study area (Figure 2). In addition, these superclones were consistently detected across all four years of the survey, each maintaining a comparable abundance, with M1 and M2 being overall the most dominant (Figure 3).
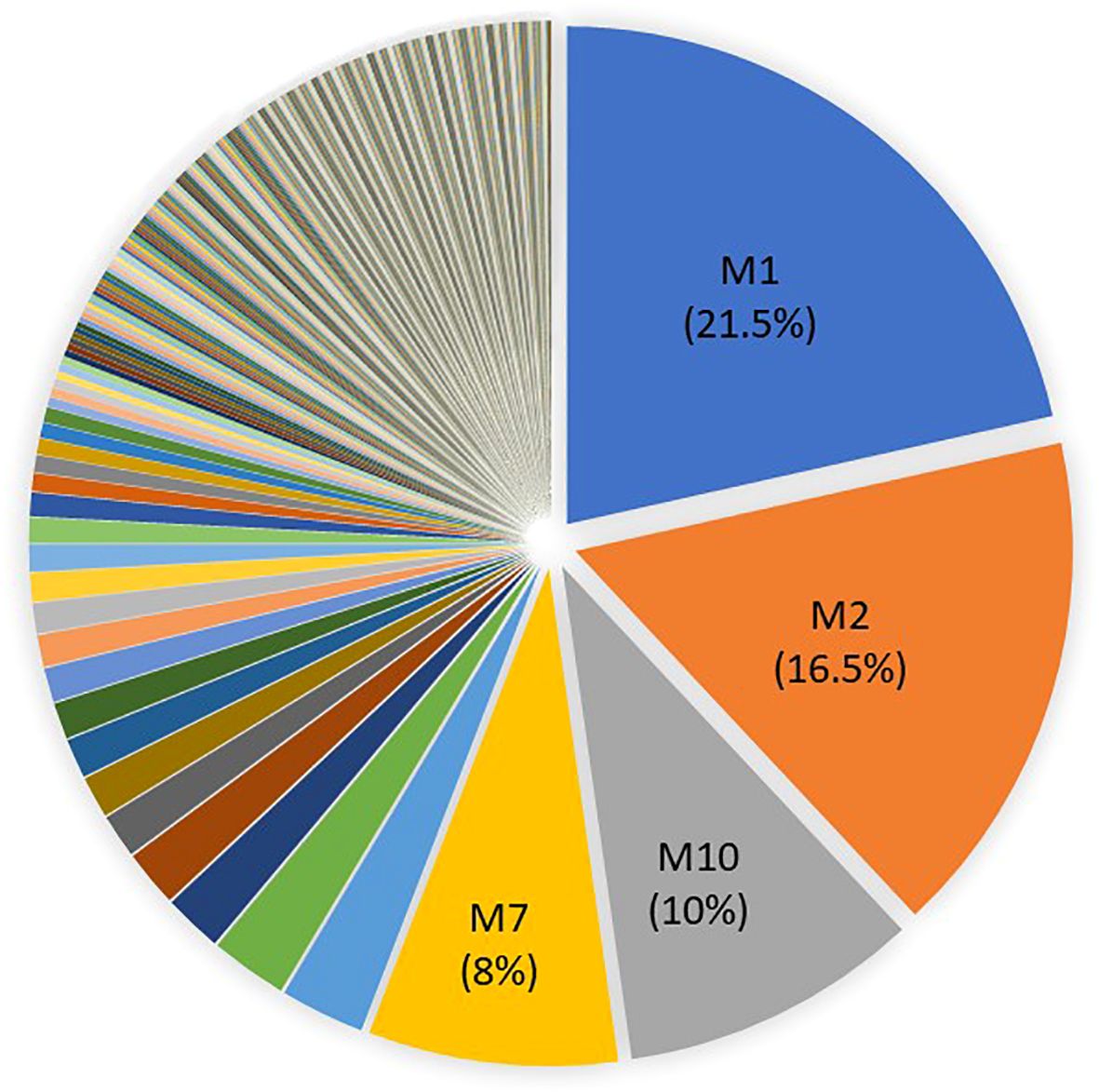
Figure 1. Frequency of the 412 multilocus genotypes (MLGs) identified in M. persicae samples. M1, M2, M7 and M10 represent the most frequent MLGs.
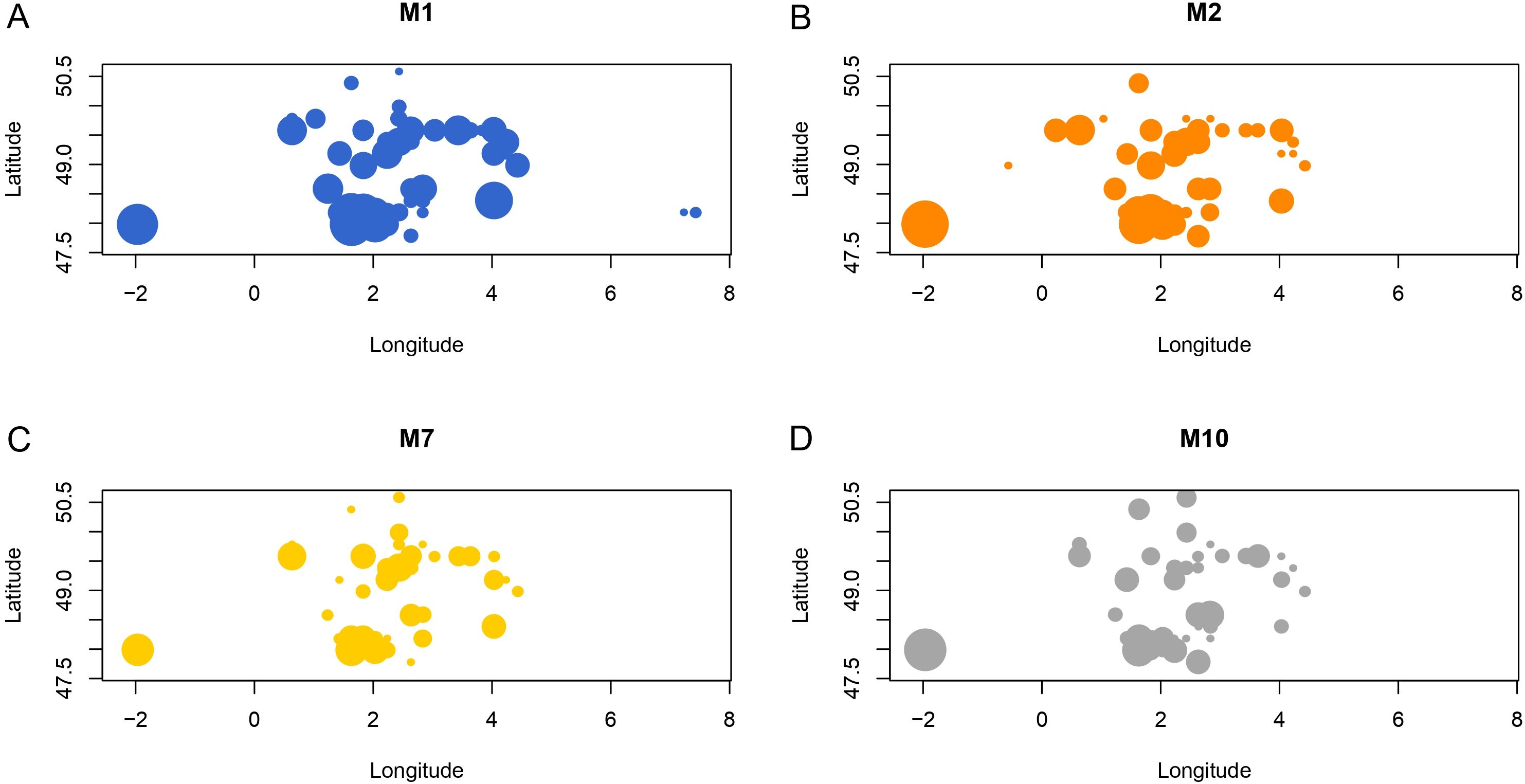
Figure 2. Geographical distribution of each of the four superclones (M1, M2, M7 and M10) of M. persicae in the sampling area covering the northern part of France. The size of the circle is proportional to the number of copies. Superclones M1 (A), M2 (B), M7 (C) and M10 (D) were represented by 621, 477, 234 and 281 copies, respectively.

Figure 3. Proportion of the four superclones of M. persicae across years in the collected samples.
Population structure analysis revealed a strong influence of clonal reproduction
To assess the extent of population structuration, we first used STRUCTURE to determine the most likely number of clusters (K). Although the simulations were carried out with a long burn-in, the results for replicates for the same K value did not systematically converge to the same outcome, as reflected by the variability of the LnP(D) for certain K values (Supplementary Figure S1). This might be partly caused by the violation of the model assumption due to the extent of asexual reproduction in the sampled populations. Nevertheless, the DeltaK suggested K=4 and K=9 as the number of clusters that best explain the structure of our dataset (Supplementary Figure S1). Given that an MLG can be represented by several copies from different geographical origins, it proved challenging to make the link between geography and cluster membership. Instead, we have represented the clusters on the tree based on the Cavalli-Sforza distance (Figure 4). At K=9, we observed that the MLGs grouped in the same cluster by STRUCTURE also tended to group together in the tree. On the other hand, at K=4, STRUCTURE grouped MLGs that were far apart in the genetic tree (Supplementary Figure S2), suggesting that K=4 explained the structure of our dataset less effectively. The tree also revealed that the four major superclones belong to different clades, consistent with the STRUCTURE analyses (black arrows in Figure 4). Interestingly, some lineages (those belonging to Cluster 2) showed longer branch lengths than the majority of MLGs.
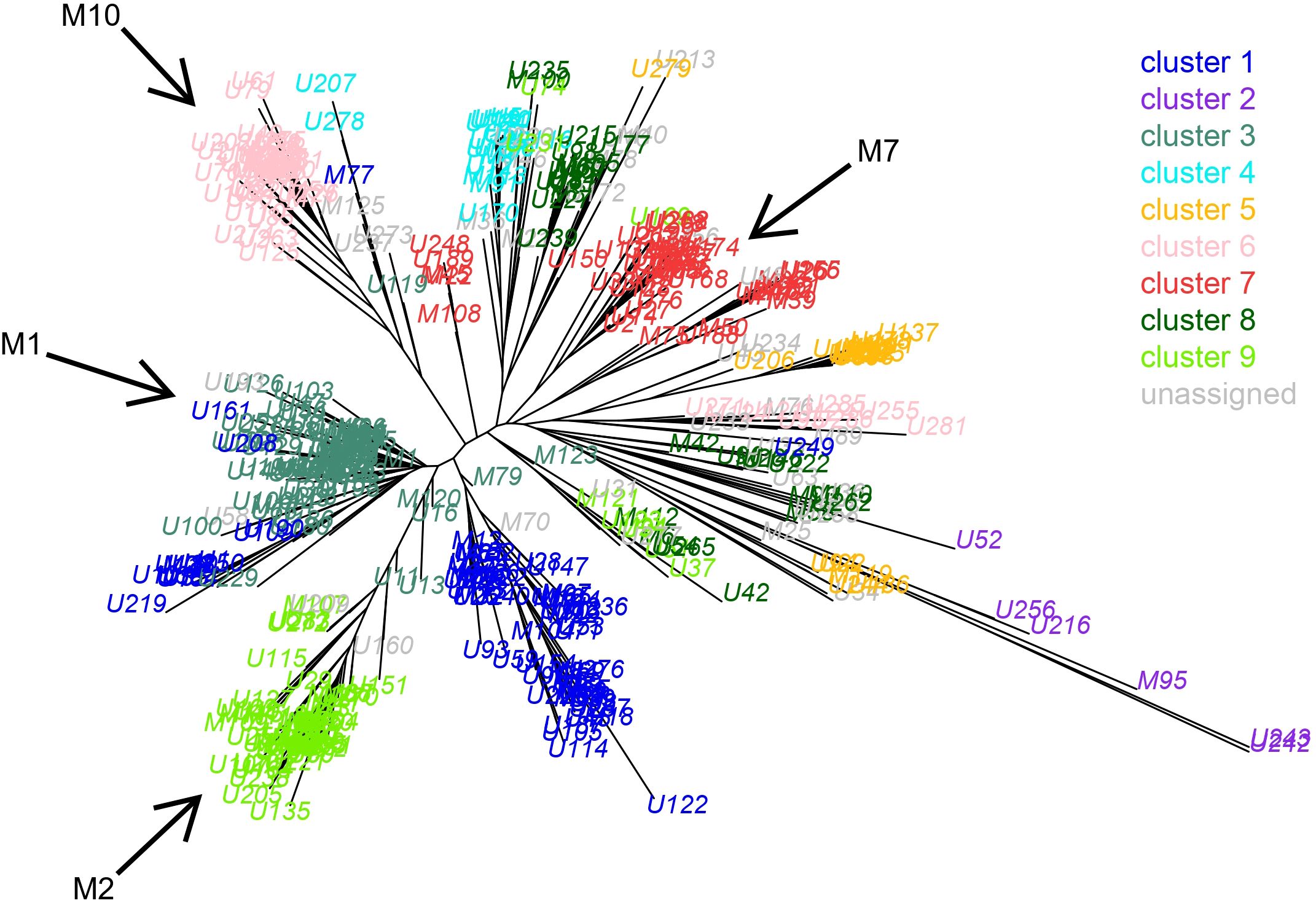
Figure 4. Neighbor-Joining tree based on the Cavalli-Sforza distance measured between the 412 different multilocus genotypes (MLGs) of M. persicae. Each MLG is colored according to its cluster membership as inferred by the STRUCTURE analysis for K=9. The position in the tree of the four most common superclones is indicated by black arrows.
F-statistics analyses further revealed considerable genetic differentiation between the different clusters, with FST ranging from 0.12 to 0.38. FIS revealed that all clusters (except Cluster 2) had a large excess of heterozygotes (indicated by strongly negative FIS, ranging from -0.24 to -0.59), while cluster 2 has a positive FIS (0.25) indicating homozygote excess (Supplementary Table S3). Lastly, we observed that the number of allelic differences between MLGs belonging to the same STRUCTURE cluster (at K=9) was very low (Supplementary Figure S3), whereas allelic differences between MLGs from different clusters were high, averaging 8.5. Cluster 3 and Cluster 6 showed average differences of only 2.7 and 3.1 alleles between MLG pairs, respectively. Clusters 1, 4, 5, 7, and 9 also exhibited relatively low values, ranging from 3.5 to 5.3 alleles. In contrast, cluster 2 showed outlier values, with the average number of allelic differences among MLGs reaching 11.5. Therefore, it appears that STRUCTURE has grouped together MLGs that are genetically very similar, often differing by only a few alleles. This likely reflects clonal diversification within M. persicae populations, where mutations accumulate successively and independently from an ancestral clonal lineage, resulting in sets of closely related MLGs (e.g., those in Cluster 6) that differ markedly from other clusters. We propose that clusters 1 to 9 (with the exception of Cluster 2) are the result of this clonal diversification process. This interpretation is further supported by the high among-cluster FST values and the observed excess of heterozygotes. In contrast, we hypothesize that MLGs in Cluster 2 originated from sexual reproduction, consistent with the high within-group allelic differences, longer branch lengths in the tree, and a contrasting FIS signature.
No clear association between host plants and the distribution of superclones or genetic clusters
Superclones M1 and M2 were found on 15 different host-plants while M7 and M10 were found on six and nine hosts, respectively (Supplementary Table S1). Comparison of the four superclones across the five most common host plants revealed no clear differences in distribution or evidence of host specialization (Figure 5). The annotation of collection plants on the Neighbor-Joining tree of the 412 MLGs further supports the absence of genetically based host-plant specialization, as no association was observed between genetic clusters and host plants (Figure 6, see also Supplementary Figure S4).
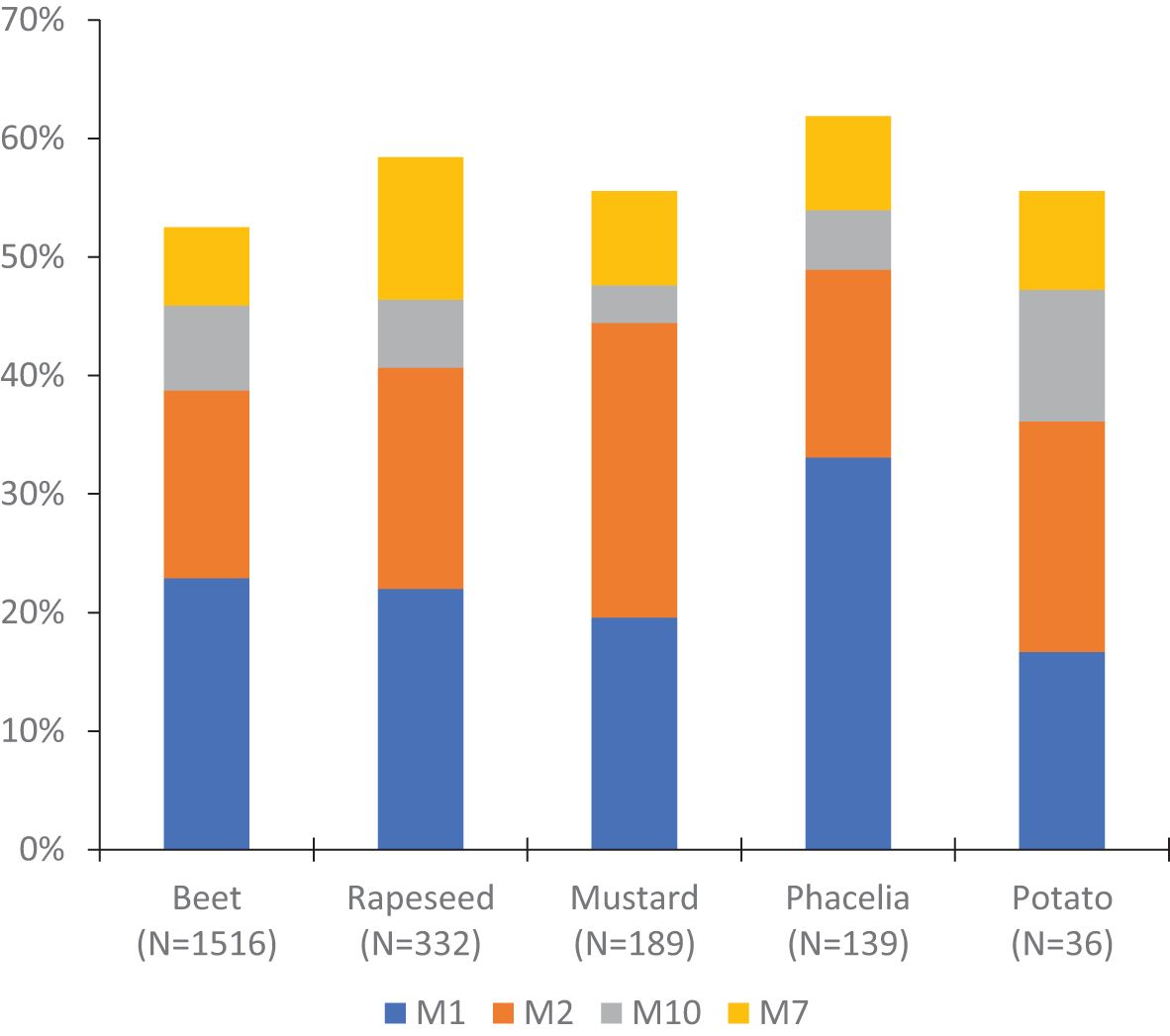
Figure 5. Distribution of the four superclones of M. persicae in five host-plant species. N represents the number of individuals sampled on the different plants.
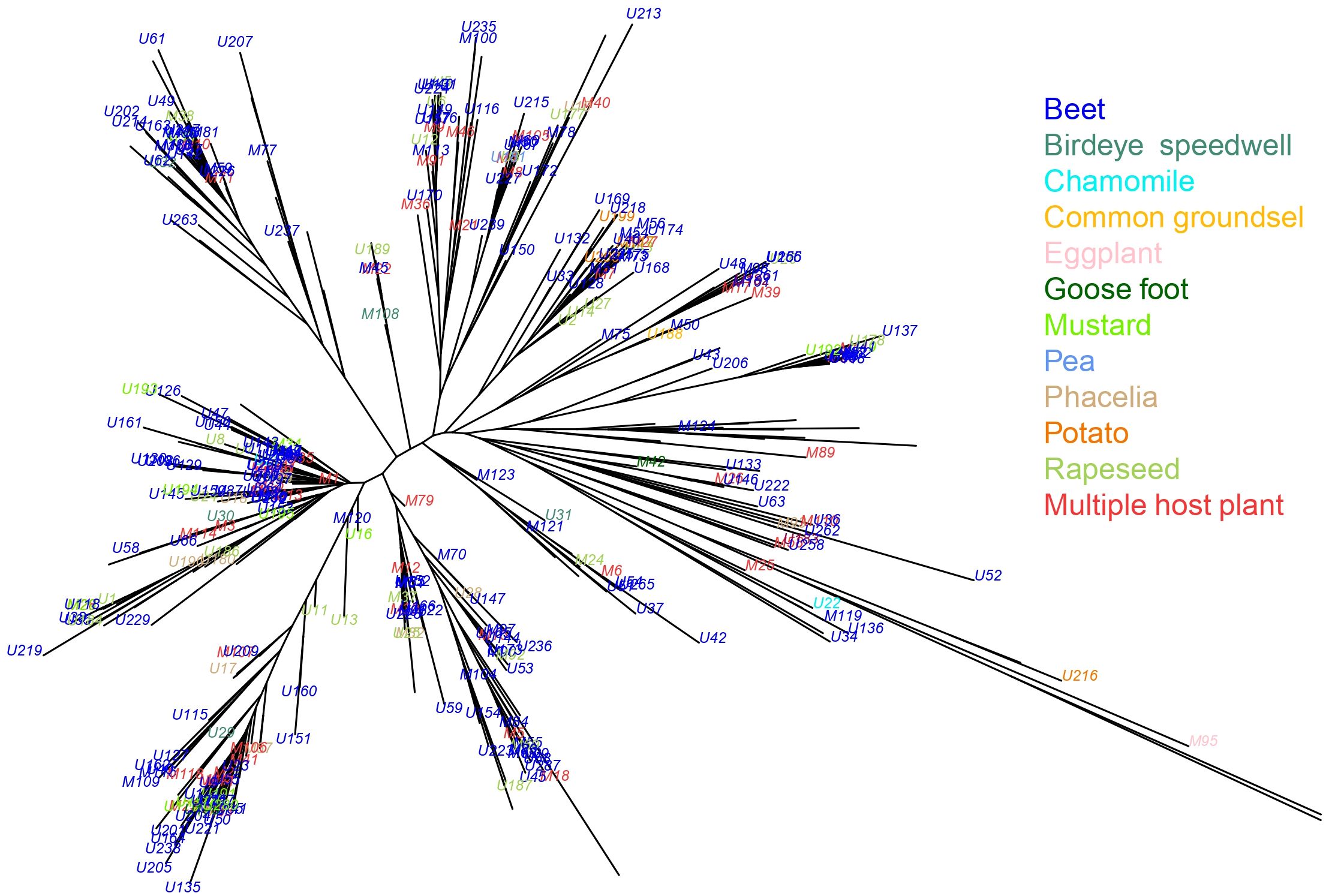
Figure 6. Neighbor-Joining tree based on the Cavalli-Sforza distance measured between the 412 different multilocus genotypes (MLGs) of M. persicae. Each MLG is colored according to the host plant on which it was collected. MLGs represented by multiple copies found on different host plants are not shown, as they cannot be assigned to a single host.
Discussion
Our study provides robust evidence that Myzus persicae populations in northern France are predominantly composed of a few highly successful superclones that exhibit remarkable ecological generalism. Contrary to the hypothesis that M. persicae might be composed of cryptic host-specialized lineages, we found no clear association between multilocus genotypes (MLGs) or genetic clusters and specific host plants, even across distantly related botanical families. These results reinforce the status of M. persicae as a true polyphagous (=generalist) species and underscore the ecological dominance of a small number of genotypes in natural populations of this aphid.
The high clonal structure observed, reflected by a low G:N ratio (0.14), the overwhelming representation of four superclones, and signs of a clonal diversification provides evidence that asexual reproduction plays a dominant role in shaping the population structure of M. persicae in the northern half of France. The predominance of asexual reproduction in this region is likely driven by the relatively mild winter climate, which permits parthenogenetic overwintering and consequently favours aphid genotypes that do not invest in the production of energetically costly sexual morphs (Rispe et al., 1998; Simon et al., 2002).
One particularly intriguing aspect of our results is the identification of Cluster 2, which differed markedly from other clusters. This group exhibited higher genetic diversity, longer branch lengths, and a positive inbreeding coefficient, all consistent with a history of sexual reproduction (Halkett et al., 2005). These genotypes were genetically distant from others, possibly representing rare recombinants produced during sexual cycles. Similar patterns of rare sexual recombination within largely asexual populations have been reported in previous works on aphids including M. persicae, Rhopalosiphum padi and Sitobion avenae (Haack et al., 2000; Delmotte et al., 2002; Vorburger et al., 2003a; Gilabert et al., 2009). This pattern is rare among the predominantly parthenogenetic populations of M. persicae in temperate zones and could reflect either recent loss of sex or immigration from regions where cyclical parthenogenesis – i.e. the alternation of asexual and sexual generations within a year – is more common because of harsher winters and more widespread primary hosts (Guillemaud et al., 2003; Roy et al., 2022).
Interestingly, the MLGs in Cluster 2 were not dominant in the population, suggesting that although sexual reproduction introduces novel genetic variation, these genotypes may not be as competitively successful under current field conditions as the well-established superclones. The fact that the four superclones were found consistently across multiple years, over large geographic areas, and on diverse host plants suggests they possess broad ecological plasticity. Their prevalence in suction and yellow traps further suggests that, in addition to their ability to exploit a broad spectrum of host plant species, they are also efficient dispersers, likely enhancing their capacity to dominate landscapes with mixed or shifting host-plant communities. Similar patterns have been observed in other aphid species where parthenogenesis allows for the rapid expansion of fit genotypes (Vorburger et al., 2003a; Gilabert et al., 2009; Figueroa et al., 2018).
While the dominance of a few clones might suggest potential bottlenecks or founder effects, our STRUCTURE and Neighbor-Joining analyses show that on the contrary the major superclones are genetically distinct and belong to separate clusters. This argues for the independent evolutionary origin of the MLGs concerned rather than clonal amplification from a single recent ancestor. The presence of multiple closely related MLGs within clusters, especially those with low allelic divergence and connected by short branches, supports the view of clonal diversification through stepwise mutations, a hallmark of long-term asexuality (Barraclough et al., 2003; Halkett et al., 2005; Tang et al., 2014). These clonal radiations likely represent microevolutionary processes driven by mutation accumulation in the absence of recombination (Delmotte et al., 2002; Barraclough et al., 2003). Clustering and genetic distance analyses identified an optimal partitioning into nine distinct clusters. Notably, if Cluster 2 is predominantly composed of cyclically parthenogenetic lineages (a hypothesis we could not firmly demonstrate, as we were unable to directly assess their reproductive phenotype), this suggests that the populations of M. persicae in northern France are primarily represented by eight main clonal lineages, each derived from independent ancestral origins followed by subsequent diversification. The precise origins, both temporal and geographical, of these independent clonal diversification events remain unclear and warrant further investigation through the integration of our dataset into a broader survey encompassing a wider geographical range.
Our findings provide important insights into the ecology of generalism in aphids. One might expect that exploitation of multiple host species would be achieved through a mosaic of specialized genotypes, but our results do not support this view. The absence of host-associated differentiation in the STRUCTURE analysis, and the lack of phylogenetic clustering by host plant in the genetic tree, provide strong evidence against the existence of cryptic host races within our sampled populations. This contrasts with previous studies on M. persicae that reported host-based population structure (Zamoum et al., 2005; Singh et al., 2021; Roy et al., 2022). However, much of the host-associated differentiation in those studies was driven by individuals collected from tobacco and peach, hosts that were not included in the present survey. Our results also contrast with findings in other aphids such as the pea aphid (Acyrthosiphon pisum) or the cotton-melon aphid (Aphis gossypii), where host-associated lineages show clear genetic structuring (Vanlerberghe-Masutti and Chavigny, 1998; Peccoud et al., 2009). Our results further demonstrate that broad host use is a property of individual clones of M. persicae, particularly the superclones, rather than a result of population-level polymorphism for host specialization. In herbivorous insects, generalist feeding strategies are often thought to be evolutionarily unstable due to trade-offs in host use and specialization (Futuyma and Moreno, 1988; Joshi and Thompson, 1995). However, M. persicae stands out as an exception, as do some other species that are partially or exclusively sexual, such as whiteflies, fall armyworms and desert locusts. Despite being a generalist species with over 400 recorded host plant species (Blackman and Eastop, 2000), our study confirms that M. persicae generalism is not merely a species-level trait but a property of individual superclones. These lineages, notably M1, M2, M7, and M10, were found on diverse hosts spanning ten botanical families and constituted more than half of all genotyped individuals. Two main hypotheses explain the persistence of parthenogenetic lineages. The frozen niche variation (FNV) hypothesis suggests these lineages consist of specialized genotypes, each adapted to a narrow niche, potentially outcompeting their more generalist sexual ancestors (Vrijenhoek, 1979; Vrijenhoek and Parker, 2009). In contrast, the general-purpose genotype (GPG) hypothesis proposes that parthenogenetic lineages have broadly adapted genotypes resulting from long term selection by environmental heterogeneity (Baker, 1965; Lynch, 1984). While both hypotheses have received empirical support (Vorburger et al., 2003b; Maraun et al., 2022; Godefroid et al., 2024), our results favor the GPG hypothesis, suggesting that generalist clones persist longer than specialist ones over evolutionary timescales.
Several traits may contribute to the broad host range observed in the M. persicae superclones. First, these clones may exhibit greater behavioral plasticity in host choice and acceptance, enabling flexible foraging strategies and enhancing their survival and reproduction. Second, they may possess specific metabolic or physiological adaptations that enable them to adjust to a wide variety of host plants. An experimental study involving a generalist M. persicae clone exposed to unrelated host plant species under controlled laboratory conditions revealed coordinated expression of gene clusters linked to host shift, notably including cathepsin B and cuticular protein genes (Mathers et al., 2017). These findings mean that transcriptional plasticity and epigenetic regulation play a key role in facilitating host plant acclimation in generalist (i.e. phytophagous) clones (Shih et al., 2023). Third, detoxification capacity plays a critical role in shaping host range by enabling herbivorous insects to neutralize or tolerate harmful plant compounds encountered during feeding (Simon et al., 2015; Shih et al., 2023). Interestingly, insecticide-resistant M. persicae genotypes have been shown to exhibit elevated expression of detoxifying enzymes such as cytochrome P450s, esterases, and glutathione S-transferases (Silva et al., 2012; Troczka et al., 2021; Bass and Nauen, 2023). In particular, a study on the tobacco-adapted race, which is tolerant to nicotine, revealed overexpression of a cytochrome P450 gene, CYP6CY3, driven by gene amplification and promoter alterations (Puinean et al., 2010; Bass et al., 2013). Subsequent research confirmed that these regulatory changes in CYP6CY3 expression were responsible for adaptation to tobacco, potentially contributing to race formation (Singh et al., 2020). Such genetic innovations likely involved transposable elements, which often accumulate near loci encoding xenobiotic metabolism genes (Baril et al., 2023). The four superclones identified in this study, along with many other multilocus genotypes (MLGs), are likely to carry multiple genes conferring insecticide resistance, consistent with recent findings in southern French populations of M. persicae (Roy et al., 2022). Characterizing the resistance mechanisms in these superclones and investigating their potential role in broad tolerance to plant defenses will doubtless provide valuable insights into the molecular basis of generalism and xenobiotic resistance. Together, these traits may explain the nutritional plasticity associated with M. persicae superclones.
Our work, which reveals the dominance of a few M. persicae clones with broad ecological tolerance, raises important concerns from a pest management perspective. While ecological theory suggests that specialization can confer fitness advantages under stable environmental conditions (Colles et al., 2009; Wang et al., 2019), our results indicate that in the highly dynamic context of conventional agriculture, generalist genotypes may have a selective advantage. Agricultural landscapes are generally characterized by frequent crop rotation, the use of mixed cultivars, and regular chemical applications, factors that create a constantly changing environment in which flexible and resilient genotypes are likely to thrive (Gilabert et al., 2009; Hardy et al., 2020). The M. persicae superclones capable of exploiting multiple host plants and dispersing over long distances are also likely to be those best equipped to resist insecticides and to transmit a broad array of phytopathogenic viruses (Bass et al., 2014; Nio et al., 2025). Continuous monitoring of the clonal composition of M. persicae in these landscapes is therefore strongly advocated, not only to understand ecological and evolutionary dynamics, but also in order to anticipate the spread of aphid-borne viruses and the emergence and dissemination of insecticide resistance (van Emden and Harrington, 2017).
In conclusion, our study supports the hypothesis that M. persicae is a true ecological generalist, with a population structure dominated by a small number of widespread and highly plastic superclones. These genotypes show no evidence of host-plant specialization and appear to thrive in diverse agricultural environments. Future research should focus on the molecular mechanisms underlying their ecological success and explore whether their dominance is maintained under different environmental pressures or pest control strategies.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
J-CS: Formal analysis, Writing – original draft, Resources, Methodology, Funding acquisition, Conceptualization, Investigation, Project administration, Writing – review & editing, Supervision. YN: Formal analysis, Writing – review & editing, Data curation, Investigation, Methodology. CB: Investigation, Resources, Writing – review & editing, Methodology. FD: Resources, Methodology, Writing – review & editing, Investigation. FM: Formal analysis, Writing – review & editing, Methodology. SB: Resources, Writing – review & editing. AL: Resources, Writing – review & editing. QC: Methodology, Writing – review & editing, Resources. GAM: Writing – review & editing, Investigation, Resources. VB: Writing – review & editing, Resources, Methodology. TC: Methodology, Resources, Investigation, Writing – review & editing. JJ: Writing – review & editing, Investigation, Formal analysis, Methodology, Visualization. AM: Resources, Writing – review & editing, Methodology, Investigation. GHM: Writing – review & editing, Resources, Methodology.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was funded by FranceAgriMer through the Resaphid, BeetRes, and Redivibe projects as part of Plan National de Recherche et Innovation to control beet yellows with alternative strategies.
Acknowledgments
We thank our colleagues from INRAE, the University of Amiens (UMR EDYSAN), and the Institut Technique de la Betterave (ITB) for their valuable assistance with aphid sampling, with special thanks to Fabienne Maupas (ITB) for her support. We are also grateful to Alex Greenslade (Rothamsted Insect Survey, UK) for his help with species identification. This study was funded by FranceAgriMer through the Resaphid, BeetRes, and Redivibe projects.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1635527/full#supplementary-material
Supplementary Table 1 | Description of the 3,112 individuals of M. persicae sampled during the survey, including their code, origin, date and site of collection, genotypes at 14 microsatellite markers and assigned multilocus genotype (MLG).
Supplementary Table 2 | Host-plants from which individuals of M. persicae were collected during the survey. We were unable to obtain complete multilocus genotypes for individuals sampled on Camelina sativa, Fumaria officinalis and Leucanthemum vulgare.
Supplementary Table 3 | Pairwise FST (above diagonal) between MLGs from the different clusters inferred from the STRUCTURE analysis (at K=9). The 95% CI obtained by bootstrapping over loci is shown below the diagonal. Also shown is the FIS for each cluster and it 95% CI. FST analyses revealed an outstanding genetic differentiation between some of the clusters (reaching more than 30%). The FIS revealed that the clusters that supposedly group MLGs that evolved via clonal diversification (all except Cluster 2) show very large excesses of heterozygotes (negative FIS), whereas the cluster containing MLGs supposedly resulting from sexual reproduction (Cluster 2) shows a positive FIS.
Supplementary Figure 1 | Diagnostic plots to infer the most likely number of clusters from the STRUCTURE analyses based on the Delta-K method (Evanno et al., 2005) implemented in the R package ‘pophelper’ (https://github.com/royfrancis/pophelper).
Supplementary Figure 2 | Neighbor-Joining tree based on the Cavalli-Sforza distance measured between the 412 different multilocus genotypes (MLGs) of M. persicae. Each MLG is colored according to its cluster membership as inferred by the STRUCTURE analysis for K=4.
Supplementary Figure 3 | Number of different alleles between each pair of MLGs (N=412) over the 14 microsatellite loci. This value ranges from 0 (on the diagonal, when each MLG is compared to itself) to a theoretical maximum value of 28 (2 × the number of loci). MLGs are sorted according to the STRUCTURE analysis, which inferred that the most likely number of clusters was K=9. The identity of the inferred STRUCTURE cluster is shown in the first row and column, and cells are colored using the same colour code as that used in Figure 4. The matrix of the pairwise number of allelic differences between samples is colored according to the values (green: few differences, red: many differences). It appears that STRUCTURE has grouped together MLGs that are genetically very close, often differing by only a few alleles. For example, Clusters 3 and 6 show an average difference of only 2.7 and 3.1 alleles between pairs of MLG, and Clusters 1, 4, 5, 7 and 9 remain at very low values (3.5 to 5.3). This probably reflects clonal diversification, with successive and independent accumulation of mutations from an ancestral clone, leading to a set of MLGs that are genetically very close (e.g. those belonging to Cluster 6) but differ considerably from the other clusters.
Supplementary Figure 4 | Radial representation of the Neighbor-Joining tree (based on the Cavalli-Sforza distance) shown in Figure 6 to better see the distribution of the collection plants. Each MLG is colored according to the host plant on which it was collected. MLGs represented by multiple copies found on different host plants are not shown, as they cannot be assigned to a single host.
References
Bailleul D., Stoeckel S., and Arnaud-Haond S. (2016). RClone: a package to identify multilocus clonal lineages and handle clonal data sets in R. Methods Ecol. Evol. 7, 966–970. doi: 10.1111/2041-210X.12550
Baker H. (1965). “Characteristics and modes of origin of weeds,” in The genetics of colonizing species: Proc. 1st Internat. Union biol Sci. (Asilomar, California: Academic Press), 147–172.
Baril T., Pym A., Bass C., and Hayward A. (2023). Transposon accumulation at xenobiotic gene family loci in aphids. Genome Res. 33, 1549–5469. doi: 10.1101/gr.277820.123
Barraclough T. G., Birky C. W. Jr., and Burt A. (2003). Diversification in sexual and asexual organisms. Evolution 57, 2166–2172. doi: 10.1111/j.0014-3820.2003.tb00394.x
Bass C. and Nauen R. (2023). The molecular mechanisms of insecticide resistance in aphid crop pests. Insect Biochem. Mol. Biol. 156, 103937. doi: 10.1016/j.ibmb.2023.103937
Bass C., Puinean A. M., Zimmer C. T., Denholm I., Field L. M., Foster S. P., et al. (2014). The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 51, 41–51. doi: 10.1016/j.ibmb.2014.05.003
Bass C., Zimmer C. T., Riveron J. M., Wilding C. S., Wondji C. S., Kaussmann M., et al. (2013). Gene amplification and microsatellite polymorphism underlie a recent insect host shift. Proc. Natl. Acad. Sci. 110, 19460–19465. doi: 10.1073/pnas.1314122110
Blackman R. L. (1987). Morphological discrimination of a tobacco-feeding form from Myzus persicae (Sulzer) (Hemiptera: Aphididae), and a key to New World Myzus (Nectarosiphon) species. Bull. Entomological Res. 77, 713–730. doi: 10.1017/S0007485300012219
Blackman R. L. and Eastop V. F. (2000). Aphids on the World’s Crops: an Identification and Information Guide (New York: Willey and sons).
Cavalli-Sforza L. L. and Edwards A. W. (1967). Phylogenetic analysis. Models and estimation procedures. Am. J. Hum. Genet. 19, 233–257.
Coeur d’acier A., Cruaud A., Artige E., Genson G., Clamens A.-L., Pierre E., et al. (2014). DNA barcoding and the associated phylAphidB@se website for the identification of European aphids (Insecta: Hemiptera: Aphididae). PloS One 9, e97620. doi: 10.1371/journal.pone.0097620
Colles A., Liow L. H., and Prinzing A. (2009). Are specialists at risk under environmental change? Neoecological, paleoecological and phylogenetic approaches. Ecol. Lett. 12, 849–863. doi: 10.1111/j.1461-0248.2009.01336.x
De Barro P. J., Liu S.-S., Boykin L. M., and Dinsdale A. B. (2011). Bemisia tabaci: A statement of species status. Annu. Rev. Entomology 56, 1–19. doi: 10.1146/annurev-ento-112408-085504
Delmotte F., Leterme N., Gauthier J.-P., Rispe C., and Simon J.-C. (2002). Genetic architecture of sexual and asexual populations of the aphid Rhopalosiphum padi based on allozyme and microsatellite markers. Mol. Ecol. 11, 711–723. doi: 10.1046/j.1365-294X.2002.01478.x
Drès M. and Mallet J. (2002). Host races in plant-feeding insects and their importance in sympatric speciation. Philos. Trans. R. Soc. Lond B Biol. Sci. 29, 471–492. doi: 10.1098/rstb.2002.1059
Durand K., Yainna S., and Nam K. (2022). Incipient speciation between host-plant strains in the fall armyworm. BMC Ecol. Evol. 22, 52. doi: 10.1186/s12862-022-02008-7
Edger P. P., Heidel-Fischer H. M., Bekaert M., Rota J., Glöckner G., Platts A. E., et al. (2015). The butterfly plant arms-race escalated by gene and genome duplications. Proc. Natl. Acad. Sci. 112, 8362–8366. doi: 10.1073/pnas.1503926112
Ehrlich P. R. and Raven P. H. (1964). Butterflies and plants: a study in coevolution. Evolution 18, 586–608. doi: 10.2307/2406212
Evanno G., Regnaut S., and Goudet J. (2005). Detecting the number of clusters of individuals using the software structure: a simulation study. Mol. Ecol. 14, 2611–2620. doi: 10.1111/j.1365-294X.2005.02553.x
Falush D., Stephens M., and Pritchard J. K. (2003). Inference of population structure using multilocus genotype data: linked loci and correlated allele frequencies. Genetics 164, 1567–1587. doi: 10.1093/genetics/164.4.1567
Figueroa C. C., Fuentes-Contreras E., Molina-Montenegro M. A., and Ramírez C. C. (2018). Biological and genetic features of introduced aphid populations in agroecosystems. Curr. Opin. Insect Sci. 26, 63–68. doi: 10.1016/j.cois.2018.01.004
Finlay-Doney M. and Walter G. H. (2012). The conceptual and practical implications of interpreting diet breadth mechanistically in generalist predatory insects. Biol. J. Linn. Soc. 107, 737–763. doi: 10.1111/j.1095-8312.2012.01991.x
Forister M. L., Novotny V., Panorska A. K., Baje L., Basset Y., Butterill P. T., et al. (2015). The global distribution of diet breadth in insect herbivores. Proc. Natl. Acad. Sci. 112, 442–447. doi: 10.1073/pnas.1423042112
Futuyma D. J. and Agrawal A. A. (2009). Macroevolution and the biological diversity of plants and herbivores. Proc. Natl. Acad. Sci. United States America 106, 18054–18061. doi: 10.1073/pnas.0904106106
Futuyma D. J. and Moreno G. (1988). The evolution of ecological specialization. Annu. Rev. Ecology Evolution Systematics 19, 207–233. doi: 10.1146/annurev.es.19.110188.001231
Gilabert A., Simon J. C., Mieuzet L., Halkett F., Stoeckel S., Plantegenest M., et al. (2009). Climate and agricultural context shape reproductive mode variation in an aphid crop pest. Mol. Ecol. 18, 3050–3061. doi: 10.1111/j.1365-294X.2009.04250.x
Godefroid M., Meynard C. N., Clamens A.-L., Popkin M., and Jousselin E. (2024). On the role of niche specialization in the geographic distribution of aphid asexual lineages: a case study using the leaf-curl plum aphid superclones. Oikos 2024, e10481. doi: 10.1111/oik.10481
Goudet J. (2005). HIERFSTAT, a package for R to compute and test hierarchical F-statistics. Mol. Ecol. Notes 5, 184–186. doi: 10.1111/j.1471-8286.2004.00828.x
Govind G., Mittapalli O., Griebel T., Allmann S., Böcker S., and Baldwin I. T. (2010). Unbiased transcriptional comparisons of generalist and specialist herbivores feeding on progressively defenseless Nicotiana attenuata plants. PLoS One 5, e8735. doi: 10.1371/journal.pone.0008735
Guillemaud T., Mieuzet L., and Simon J. C. (2003). Spatial and temporal genetic variability in French populations of the peach–potato aphid, Myzus persicae. Heredity 91, 143–152. doi: 10.1038/sj.hdy.6800292
Haack L., Simon J.-C., Gauthier J.-P., Plantegenest M., and Dedryver C.-A. (2000). Evidence for predominant clones in a cyclically parthenogenetic organism provided by combined demographic and genetic analyses. Mol. Ecol. 9, 2055–2066. doi: 10.1046/j.1365-294X.2000.01108.x
Halkett F., Simon J.-C., and Balloux F. (2005). Tackling the population genetics of clonal and partially clonal organisms. Trends Ecol. Evol. 20, 194–201. doi: 10.1016/j.tree.2005.01.001
Hardy N. B., Kaczvinsky C., Bird G., and Normark B. B. (2020). What we don’t know about diet-breadth evolution in herbivorous insects. Annu. Rev. Ecology Evolution Systematics 51, 103–122. doi: 10.1146/annurev-ecolsys-011720-023322
Joshi A. and Thompson J. N. (1995). Trade-offs and the evolution of host specialization. Evolutionary Ecol. 9, 82–92. doi: 10.1007/BF01237699
Lynch M. (1984). Destabilizing hybridization, general-purpose genotypes and geographic parthenogenesis. Q. Rev. Biol. 59, 257–290. doi: 10.1086/413902
Maraun M., Bischof P. S. P., Klemp F. L., Pollack J., Raab L., Schmerbach J., et al. (2022). Jack-of-all-trades” is parthenogenetic. Ecol. Evol. 12, e9036. doi: 10.1002/ece3.9036
Margaritopoulos J. T., Malarky G., Tsitsipis J. A., and Blackman R. L. (2007). Microsatellite DNA and behavioural studies provide evidence of host-mediated speciation in Myzus persicae (Hemiptera: Aphididae). Biol. J. Linn. Soc. 91, 687–702. doi: 10.1111/j.1095-8312.2007.00828.x
Mathers T. C., Chen Y., Kaithakottil G., Legeai F., Mugford S. T., Baa-Puyoulet P., et al. (2017). Rapid transcriptional plasticity of duplicated gene clusters enables a clonally reproducing aphid to colonise diverse plant species. Genome Biol. 18, 27. doi: 10.1186/s13059-016-1145-3
Miller S. A., Dykes D. D., and Polesky H. F. (1988). A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 16, 1215. doi: 10.1093/nar/16.3.1215
Nio Y., Buchard C., Duval F., Faure C., Mahéo F., Malatesta G., et al. (2025). Identification des réservoirs des vecteurs et des virus responsables des jaunisses de la betterave. Innov. Agronomiques. 103, 1–9. doi: 10.17180/ciag-2025-vol103-art01
Paradis E. and Schliep K. (2018). ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35, 526–528. doi: 10.1093/bioinformatics/bty633
Peccoud J., Mahéo F., de la Huerta M., Laurence C., and Simon J.-C. (2015). Genetic characterisation of new host-specialised biotypes and novel associations with bacterial symbionts in the pea aphid complex. Insect Conserv. Diversity 8, 484–492. doi: 10.1111/icad.12131
Peccoud J., Ollivier A., Plantegenest M., and Simon J. C. (2009). A continuum of genetic divergence from sympatric host races to species in the pea aphid complex. Proc. Natl. Acad. Sci. U.S.A 106, 7495–7500. doi: 10.1073/pnas.0811117106
Peccoud J., Simon J.-C., von Dohlen C., Coeur d’acier A., Plantegenest M., Vanlerberghe-Masutti F., et al. (2010). Evolutionary history of aphid-plant associations and their role in aphid diversification. Comptes Rendus Biologies 333, 474–487. doi: 10.1016/j.crvi.2010.03.004
Puinean A. M., Foster S. P., Oliphant L., Denholm I., Field L. M., Millar N. S., et al. (2010). Amplification of a cytochrome P450 gene is associated with resistance to neonicotinoid insecticides in the aphid Myzus persicae. PLoS Genet. 6, e1000999. doi: 10.1371/journal.pgen.1000999
Rispe C., Pierre J. S., Simon J. C., and Gouyon P. H. (1998). Models of sexual and asexual coexistence in aphids based on constraints. J. Evolutionary Biol. 11, 685–701. doi: 10.1046/j.1420-9101.1998.11060685.x
Roy L., Barrès B., Capderrey C., Mahéo F., Micoud A., Hullé M., et al. (2022). Host plants and insecticides shape the evolution of genetic and clonal diversity in a major aphid crop pest. Evolutionary Appl. 15, 1653–1669. doi: 10.1111/eva.13417
Shih P.-Y., Sugio A., and Simon J.-C. (2023). Molecular mechanisms underlying host plant specificity in aphids. Annu. Rev. Entomology 68, 431–450. doi: 10.1146/annurev-ento-120220-020526
Showler A. T. (2002). A summary of control strategies for the desert locust, Schistocerca gregaria (Forskål). Agriculture Ecosyst. Environ. 90, 97–103. doi: 10.1016/S0167-8809(01)00167-0
Silva A. X., Jander G., Samaniego H., Ramsey J. S., and Figueroa C. C. (2012). Insecticide resistance mechanisms in the green peach aphid Myzus persicae (Hemiptera: Aphididae) I: A transcriptomic survey. PLoS One 7, e36366. doi: 10.1371/journal.pone.0036366
Simon J.-C., d’Alençon E., Guy E., Jacquin-Joly E., Jaquiéry J., Nouhaud P., et al. (2015). Genomics of adaptation to host-plants in herbivorous insects. Briefings Funct. Genomics 14, 413–423. doi: 10.1093/bfgp/elv015
Simon J.-C., Rispe C., and Sunnucks P. (2002). Ecology and evolution of sex in aphids. Trends Ecol. Evol. 17, 34–39. doi: 10.1016/S0169-5347(01)02331-X
Singh K. S., Cordeiro E. M. G., Troczka B. J., Pym A., Mackisack J., Mathers T. C., et al. (2021). Global patterns in genomic diversity underpinning the evolution of insecticide resistance in the aphid crop pest Myzus persicae. Commun. Biol. 4, 847. doi: 10.1038/s42003-021-02373-x
Singh K. S., Troczka B. J., Duarte A., Balabanidou V., Trissi N., Carabajal Paladino L. Z., et al. (2020). The genetic architecture of a host shift: An adaptive walk protected an aphid and its endosymbiont from plant chemical defenses. Sci. Adv. 6, eaba1070. doi: 10.1126/sciadv.aba1070
Tang C. Q., Obertegger U., Fontaneto D., and Barraclough T. G. (2014). Sexual species are separated by larger genetic gaps than asexual species in rotifers. Evolution 68, 2901–2916. doi: 10.1111/evo.12483
Tay W. T., Meagher R. L., Czepak C., and Groot A. T. (2023). Spodoptera frugiperda: ecology, evolution, and management options of an invasive species. Annu. Rev. Entomology 68, 299–317. doi: 10.1146/annurev-ento-120220-102548
Troczka B. J., Singh K. S., Zimmer C. T., Vontas J., Nauen R., Hayward A., et al. (2021). Molecular innovations underlying resistance to nicotine and neonicotinoids in the aphid Myzus persicae. Pest Manage. Sci. 77, 5311–5320. doi: 10.1002/ps.6558
Vanlerberghe-Masutti F. and Chavigny P. (1998). Host-based genetic differentiation in the aphid Aphis gossypii Glover, evidenced from RAPD fingerprints. Mol. Ecol. 7, 905–914. doi: 10.1046/j.1365-294x.1998.00421.x
Vorburger C., Lancaster M., and Sunnucks P. (2003a). Environmentally related patterns of reproductive modes in the aphid Myzus persicae and the predominance of two ‘superclones’ in Victoria, Australia. Mol. Ecol. 12, 3493–3504. doi: 10.1046/j.1365-294X.2003.01998.x
Vorburger C., Sunnucks P., and Ward S. A. (2003b). Explaining the coexistence of asexuals with their sexual progenitors: no evidence for general-purpose genotypes in obligate parthenogens of the peach-potato aphid, Myzus persicae. Ecol. Lett. 6, 1091–1098. doi: 10.1046/j.1461-0248.2003.00536.x
Vrijenhoek R. C. (1979). Factors affecting clonal diversity and coexistence. Am. Zoologist 19, 787–797. doi: 10.1093/icb/19.3.787
Vrijenhoek R. C. and Parker E. D. (2009). “Geographical parthenogenesis: general purpose genotypes and frozen niche variation,” in Lost Sex: The Evolutionary Biology of Parthenogenesis. Eds. Schön I., Martens K., and Dijk P. (Springer Netherlands, Dordrecht), 99–131.
Wang A., Singh A., Huang Y., and Agrawal A. F. (2019). Ecological specialization in populations adapted to constant versus heterogeneous environments. Evolution 73, 1309–1317. doi: 10.1111/evo.13725
Zamoum T., Simon J. C., Crochard D., Ballanger Y., Lapchin L., Vanlerberghe-Masutti F., et al. (2005). Does insecticide resistance alone account for the low genetic variability of asexually reproducing populations of the peach-potato aphid Myzus persicae? Heredity 94, 630–639. doi: 10.1038/sj.hdy.6800673
Keywords: herbivorous insects, host range, clonal reproduction, ecological tolerance, specialization, generalism, genetic structure
Citation: Nio Y, Buchard C, Duval F, Mahéo F, Buzy S, Le Ralec A, Chesnais Q, Mardoc G, Brault V, Candresse T, Malatesta G, Monteiro A, Jaquiéry J and Simon J-C (2025) Host specialisation or generalism? Population genetics of the aphid Myzus persicae reveals dominance of superclones across diverse host plants. Front. Ecol. Evol. 13:1635527. doi: 10.3389/fevo.2025.1635527
Received: 26 May 2025; Accepted: 30 June 2025;
Published: 17 July 2025.
Edited by:
Hugh David Loxdale, Cardiff University, United KingdomReviewed by:
Sarah Mansfield, AgResearch Ltd., New ZealandAstrid Forneck, University of Natural Resources and Life Sciences Vienna, Austria
Copyright © 2025 Nio, Buchard, Duval, Mahéo, Buzy, Le Ralec, Chesnais, Mardoc, Brault, Candresse, Malatesta, Monteiro, Jaquiéry and Simon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jean-Christophe Simon, amVhbi1jaHJpc3RvcGhlLnNpbW9uQGlucmFlLmZy