 Borja González-Rabanal1,2*
Borja González-Rabanal1,2* Ana B. Marín-Arroyo2*Eduardo Carmona-Ballestero3,4
Ana B. Marín-Arroyo2*Eduardo Carmona-Ballestero3,4 David Cuenca-Solana5Igor Gutiérrez-Zugasti5Miguel Ángel Martín-Merino6
David Cuenca-Solana5Igor Gutiérrez-Zugasti5Miguel Ángel Martín-Merino6 Ana Isabel Ortega-Martínez7Lawrence G. Straus2,8Cristina Vega-Maeso9Manuel R. González Morales5Emanuela Cristiani10*
Ana Isabel Ortega-Martínez7Lawrence G. Straus2,8Cristina Vega-Maeso9Manuel R. González Morales5Emanuela Cristiani10*- 1Departamento de Prehistoria, Arqueología, Antropología Social y Ciencias y Técnicas Historiográficas, Universidad de Valladolid, Valladolid, Spain
- 2Grupo I+D+i EvoAdapta, (Evolución Humana y Adaptaciones durante la Prehistoria), Departamento de Ciencias Históricas, Universidad de Cantabria, Santander, Spain
- 3Servicio Territorial de Cultura, Turismo y Deporte de Valladolid, Junta de Castilla y León, Valladolid, Spain
- 4Departamento de Historia, Geografía y Comunicación, Universidad de Burgos, Burgos, Spain
- 5Instituto Internacional de Investigaciones Prehistóricas de Cantabria,Universidad de Cantabria, Banco Santander, Gobierno de Cantabria, Santander, Spain
- 6Sociedad Española de Espeleología y Ciencias del Karst, Fundación Gómez Pardo, Madrid, Spain
- 7Real Academia Burgense de Historia y Bellas Artes, Institución Fernán González, Burgos, Spain
- 8Department of Anthropology, University of New Mexico, Albuquerque, NM, United States
- 9Servicio Territorial de Cultura, Turismo y Deporte de Segovia, Junta de Castilla y León, Segovia, Spain
- 10DANTE−Diet and ANcient TEchnology Laboratory, Department of Oral and Maxillo−Facial Sciences, Sapienza University of Rome, Rome, Italy
Introduction: Research on the origin and spread of agriculture in northern Atlantic Iberia remains partially limited because archaeobotanical data are scarce due to old excavations or preservation biases.
Methods: In this paper, we present starch grain analyses of dental calculus of 18 individuals from 10 sites dated to the 4th/2nd millennium cal BC
Results: This research supports the development of extensive and stable agriculture, based on the cultivation of wheat and barley species, from the Neolithic to the Bronze Age, when millets were introduced, marking the primary shift of the Cantabrian farming economy and revealing a more diversified and mixed agriculture thereafter. In parallel, legumes were less cultivated since the Neolithic and probably used as a secondary plant resource by the Cantabrian communities.
Discussion: Our results have also allowed us to corroborate previous regional archaeobotanical and isotopic data and enhance the archaeological evidence of plant consumption during Late Prehistory, establishing a diachronic multiapproach to the development of agricultural practices in this area and providing a methodological framework for future studies.
1 Introduction
Plant domestication constituted one of the most radical changes of the “Neolithic Revolution”, leading to biological, technological, socioeconomic, and cultural transformations that began in the Levant approximately during the 9th-millennium cal BC (Fuller et al., 2010; Abbo and Gopher, 2017). Iberia was the last Mediterranean region to adopt farming, three thousand years later than the eastern Mediterranean and after the marine colonization of early Near East farmers (Martins et al., 2015; García-Puchol et al., 2018), bringing pottery, ground stone tools, and domesticated plants and animals (Guilaine, 2017). Although the contributions of Mesolithic indigenous groups in the neolithization process must not be underestimated (Arias, 2007; Guilaine and Manen, 2007), genetic evidence available for Iberia shows a greater genetic input of Anatolian farmers (Olalde et al., 2019; Villalba-Mouco et al., 2019), as observed across Europe (Haak et al., 2010; Mathieson et al., 2015).
The neolithization process of the Cantabrian Region (Atlantic northern Iberia) was belated compared to other Iberian regions (Cubas et al., 2016). The chronological evidence places the arrival of the “Neolithic package” at the beginning of the 5th-millennium cal BC (Fano et al., 2015). In this way, the Ebro Valley constitutes the main geographical corridor for the spread of the Neolithic from the Mediterranean coast to northern Iberia (Arias, 2007), although the Atlantic corridor between the Bay of Biscay and the western end of the Pyrenees cannot be ruled out as a potential route for the introduction of domestics (Zapata and Pena-Chocarro, 2005; González Morales, 2012). Archaeobotanical studies have revealed that a wide variety of crops were cultivated during the Early Neolithic across Iberia, including cereals (wheat and barley species), pulses (pea, lentil, fava bean) and other non-edible plants such as flax or poppy (Zapata et al., 2004). However, the early stages and development of agriculture in northern Iberia are still little understood, mainly due to the scarcity of carpological data because of old excavations or preservation biases. Even so, the limited data available point to the cultivation of naked and hulled varieties of wheat and barley, such as emmer wheat, einkorn wheat, common wheat, and barley (Peña-Chocarro et al., 2018). Other early Neolithic crops, such as lentil, bean, pea, flax or poppy, are absent in the carpological record (Zapata, 2002; López-Dóriga, 2016). During the Chalcolithic and the Bronze Age, a few seeds from new crops such as oats, peas, and beans appeared in the record, but they remain undated, and the date of their arrival is unknown (Zapata, 2002; López-Dóriga, 2016; Tereso et al., 2016). Finally, other crops, such as millets and rye, would not be cultivated until the Late Bronze Age and Iron Age, respectively (Moreno-Larrazabal et al., 2015; Seabra et al., 2023).
Dental calculus is the mineralized layer of dental plaque adhering to the tooth enamel that comprises calcium phosphate, mineral salts, bacteria, carbohydrates, lipids and proteins (Dobney and Brothwell, 1986). This biofilm accumulates on the tooth surface during life due to the continuous production of saliva in the mouth; hence its formation continues until the individual’s death (Lieverse, 1999). Due to its structure, dental calculus favors biomolecular preservation as molecules are entrapped rapidly by mineralization and protected from post-mortem environmental alteration (Dobney, 1994). In this sense, plant microremains (starch granules, phytoliths, fibers, pollens, spores, and other plant and animal microresidues) can survive in the ancient dental calculus matrix, being considered a repository of biographic information related to the hygiene, dietary and non-dietary habits of ancient past human communities and paleoenvironments (Radini et al., 2017). In the last 20 years, developments in dental calculus analysis and extraction protocols have made this bioarcheological matrix a key source of information in human evolution studies (Warinner, 2016; Radini et al., 2017; Hardy et al., 2018). As starch granules within human dental calculus are capable of taxonomic identification to the species level (Torrence and Barton, 2016), this bioarcheological mineralized matrix has become a relevant proxy specifically for understanding the role of plant foods in ancient human communities (Hardy et al., 2009).
The Cantabrian region has yielded an extraordinary funerary record in which many caves were selected as burial locations between the 5th-2nd millennia cal BC. However, until now, dental calculus analyses had never been applied to remains from this spatio-temporal range for inferring plant consumption during Late Prehistory. With the aim of contributing to knowledge of the origin and development of agricultural practices during Late Prehistory in northern Iberia, a dental calculus approach was achieved to ascertain what types of plants were involved in the diet of these farming communities.
2 Materials and methods
Dental calculus was available on teeth belonging to 21 of the 39 individuals anthropologically identified in 10 burial caves located in coastal and inland areas of the Cantabrian Region (Figure 1), a Eurosiberian biogeographical area located in the northern Atlantic Iberia, separated from the Spanish central plateau and the Ebro valley by steep, high mountains, running parallel to the coast and reaching their highest summits 2500m above sea level. It has an oceanic climate, with year-round rainfall and relatively limited seasonal temperature variation. The archaeological sites studied here are located in three administrative Spanish provinces: Asturias, Cantabria and Burgos. In terms of the chronocultural framework, the individuals have been directly dated by C14-AMS between 3700–1500 cal BC (González-Rabanal, 2022) and ranging from the Late Neolithic to the Middle Bronze Age. In total, we have analyzed six individuals dated in the Late Neolithic, six individuals dated in the Chalcolithic, six individuals dated in the Early Bronze Age and three individuals dated in the Middle Bronze Age (Table 1). Detailed information about the archaeological and osteological evidence of the sites is included in Supplementary Text 1.
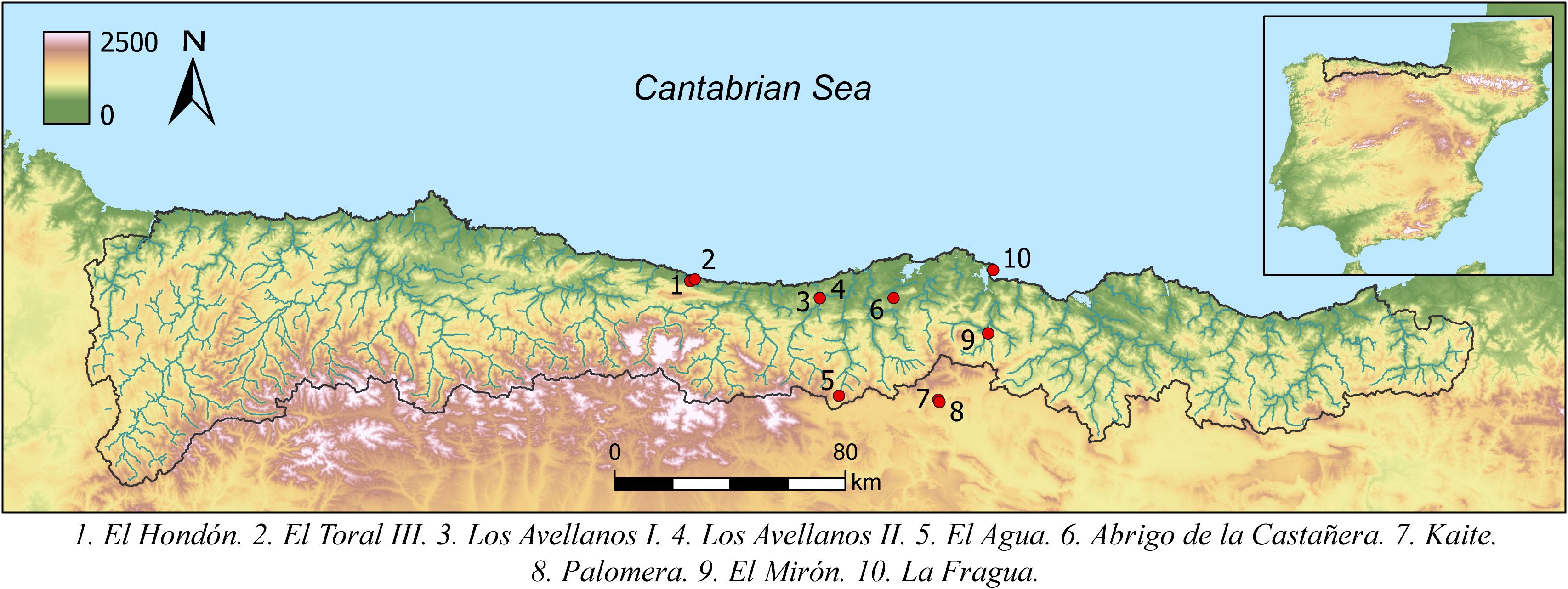
Figure 1. Geographical location of the archaeological sites studied in this work.
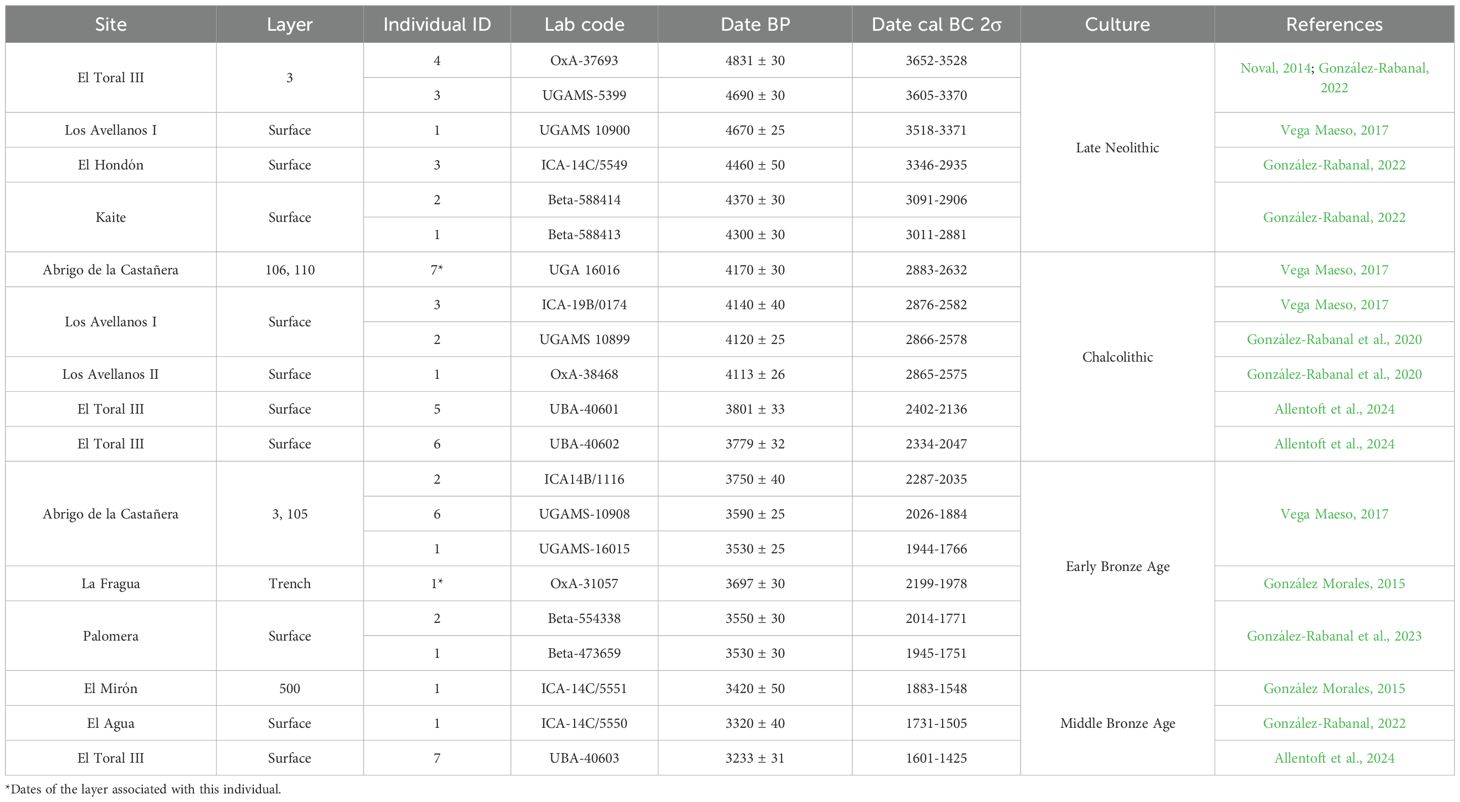
Table 1. Summary of the archaeological and chronological information of the sites.
Sampling was conducted under a Leica S8APO stereomicroscope with a magnification up to 80x. The dental calculus matrix was removed from the teeth following the protocol of Sabin and Fellow (Sabin and James, 2020) using a disposable blade and wearing powder-free gloves to prevent contamination. Disposable blades were changed after each sample extraction. Calculus samples were stored in sterile Eppendorf tubes after being left remnant of dental calculus on the teeth for future research. Decontamination and extraction procedures for micro-debris were carried out according to standard protocols as described by (Cristiani et al., 2016; 2018) and (Fiorin et al., 2021), and they were conducted in dedicated clean spaces under strict environmental monitoring of the DANTE – Diet and Ancient Technology – laboratory of Sapienza University of Rome. In this facility, strict anticontamination rules were followed, including the daily room cleaning and the prohibition of food. Bench space surfaces were cleaned prior to the analysis of each sample, using soap and ethanol, followed by covering of the surfaces by aluminum foil, and using of clean starch-free nitrile gloves at all times. Dental calculus decontamination was carried out on a Petri dish previously washed and immersed in hot ultrapure water, under the stereomicroscope with magnifications up to 100x. Soil particles adhered to the plaque’s surface were cleaned using tweezers to hold the sample and a fine sterile acupuncture needle to scratch the external layer of the calculus. The procedure was performed using drops of 0.05 M hydrochloric (HCl) acid to dissolve the mineralized flecks of soil and ultrapure water to block the demineralization. Once clean, the contaminated soil was checked for possible cross-contamination and calculus samples were washed in ultrapure water up to three times in order to remove any trace of sediment. Later, calculus was demineralized in a weak solution of 0.05 M HCl with the aim to extract the microfossils entrapped in the calculus matrix. The dissolved calculus was mounted on slides using a solution of 50:50 glycerol and ultrapure water. Bench working areas and dust traps placed in the laboratory were also regularly checked as control samples for comparative purposes in order to prevent any type of modern contamination. The examination of the microfossils was carried out using a Zeiss Imager2 polarized microscope (100x–630x) at the DANTE laboratory and a Leica DVM6 M digital microscope at the EvoAdapta laboratory from the University of Cantabria.
For the identification of archaeological starch granules, a reference collection of organic residues created from more than 300 modern wild and domestic plants natives to the Mediterranean area and housed at DANTE laboratory, was used for morphological comparison along with published literature through the observation and record of morphological criteria habitually established: shape, hilum, lamellae, fissures, Maltese cross, and size. In addition, a second botanical collection of more than 100 modern species from northern Spain was made ex professo for a more local approach considering the carpological and palynological record of the Holocene in the Cantabrian Region (Zapata, 2002; Peña-Chocarro et al., 2005; López-Merino et al., 2010; López-Dóriga, 2016; Tereso et al., 2016; Núñez de la Fuente, 2018). It is housed at the EvoAdapta laboratory. In order to search associations of qualitative and quantitative variables among sites, cultures and starch morphotypes, we implemented a Factor Analysis of Mixed Data (FAMD), a multivariate statistical technique used to reduce the dimensionality of data by transforming qualitative and quantitative variables into a smaller, uncorrelated set of new variables called dimensions.
An experimental analysis of 22 modern plant species from these collections, considering the most commonly gathered wild fruits and cultivated crops in this area, was undertaken in order to compare morphologically and statistically the archaeological starch grains with the experimental ones and provide a potential identification to species level. Six species belong to the Triticeae tribe: Hordeum vulgare L., Secale cereale L., Triticum aestivum L., Triticum dicoccoides (Körn. ex Asch. & Graebn.) Schweinf., Triticum dicoccum L., and Triticum monococcum L.; four species belong to the Paniceae tribe: Panicum miliaceum L., Setaria italica (L.) P.Beauv., Setaria verticillata (L.) P.Beauv., and Setaria viridis (L.) P.Beauv.; one species belonging to the Andropogoneae tribe: Sorghum halepense (L.) Pers.; four species to the Fabeae tribe: Vicia sylvatica L., Vicia faba L., Lens culinaris Medik. (also known as Vicia lens (L.) Coss. & Germ.), and Pisum sativum L. (also known as Lathyrus oleraceus Lam.); and two species to Aveneae/Poeae tribe: Avena fatua L. and Avena sativa L. Finally, six wild plants species were also sampled including: Quercus robur L., Quercus ilex L., Corylus avellana L., Arbutus unedo L., Pinus pinea L., Cornus sanguinea L. and Prunus spinosa L. Morphological traits of each experimental plant species were recorded following the ICSN 2011, The International Code for Starch Nomenclature (https://www.fossilfarm.org/ICSN/Code.html) and can be consulted in Supplementary Text 2. In order to extract the starch grains for statistical analysis, one experimental seed of each species was grounded using pestle and mortar. Starch powder (0.5 mg) was re-suspended in 100 μL of sterile distilled water and vortexed for 5 minutes. Later, the sample was observed by optical light microscopy. One hundred starch granules were randomly selected, and their length was measured. Minimum and maximum lengths, mean, and median values with relative standard deviations and their IQR ranges were recorded for each species. To investigate the existence of significant differences, the length distribution of starch grains of each experimental species was statistically compared with that of its tribe, and later with the length of archaeological starch grains from each site. This statistical analysis was carried out through a Pairwise Wilcoxon test. Results were considered significant for p-values <0.05 (*<0.05; **<0.01; ***<0.001) and not significant (n.s.) for measurements >0.05.
3 Results
3.1 Control sampling
We retrieved 90 microremains during the 12 environmental control samples carried out during the dental calculus analyses (Supplementary Table 1). Our results based on this procedure summarize as follows: synthetic and plant fibers and hairs, fungal spores and hyphae, conifer and palm pollens, insect debris, a very low number of phytoliths and some unidentified small starch granules (Supplementary Figure 1). These tests control have allowed us to characterize the flow of contaminations through seasons, suggesting the possible relation with environmental causes. The residues more commonly found were fibers, fungal remains and pollens. Starch granules were very rare in the control samples, and the morphotypes described below were never recovered, supporting the archaeological nature of the microfossils identified within dental calculus.
3.2 State of preservation and general results
The state of preservation of dental calculus was moderate, with most of the samples showing weak bands of tartar or small patches on the buccal and lingual surfaces of the teeth. The weight of the dental calculus samples ranged between 0.1–34 mg (mean= 5.1; median= 2.8) (Supplementary Table 2). A total of 194 starch grains were retrieved within dental calculus samples. This number can be considered quantitatively low given the number of individuals analyzed. The relatively low presence of starch grains could be explained by the small size of the calculus samples, because only small amounts of dental calculus could be sampled from most individuals. In fact, there is a significant positive correlation between the size of the samples and the number of starch grains identified by individual (p = 0.01, rho = 0.55), which suggests that the preservation conditions of dental calculus determine the number of starch grains found in our archaeological samples. However, our results can be considered satisfactory since it has allowed the recovery of microresidues in such small quantities.
Of the total number of 21 individuals who preserved dental calculus on their teeth and, therefore, were analyzed, the vast majority (n = 18) provided starch grains in greater or lesser quantities (Table 2). Only the Chalcolithic/Middle Bronze Age individuals from El Toral III failed to discover starch grains. We have identified 47 starch grains in Late Neolithic individuals, 20 starch grains in Chalcolithic individuals, 81 starch grains in Early Bronze Age individuals and 46 starch grains in Middle Bronze Age individuals (Table 2).
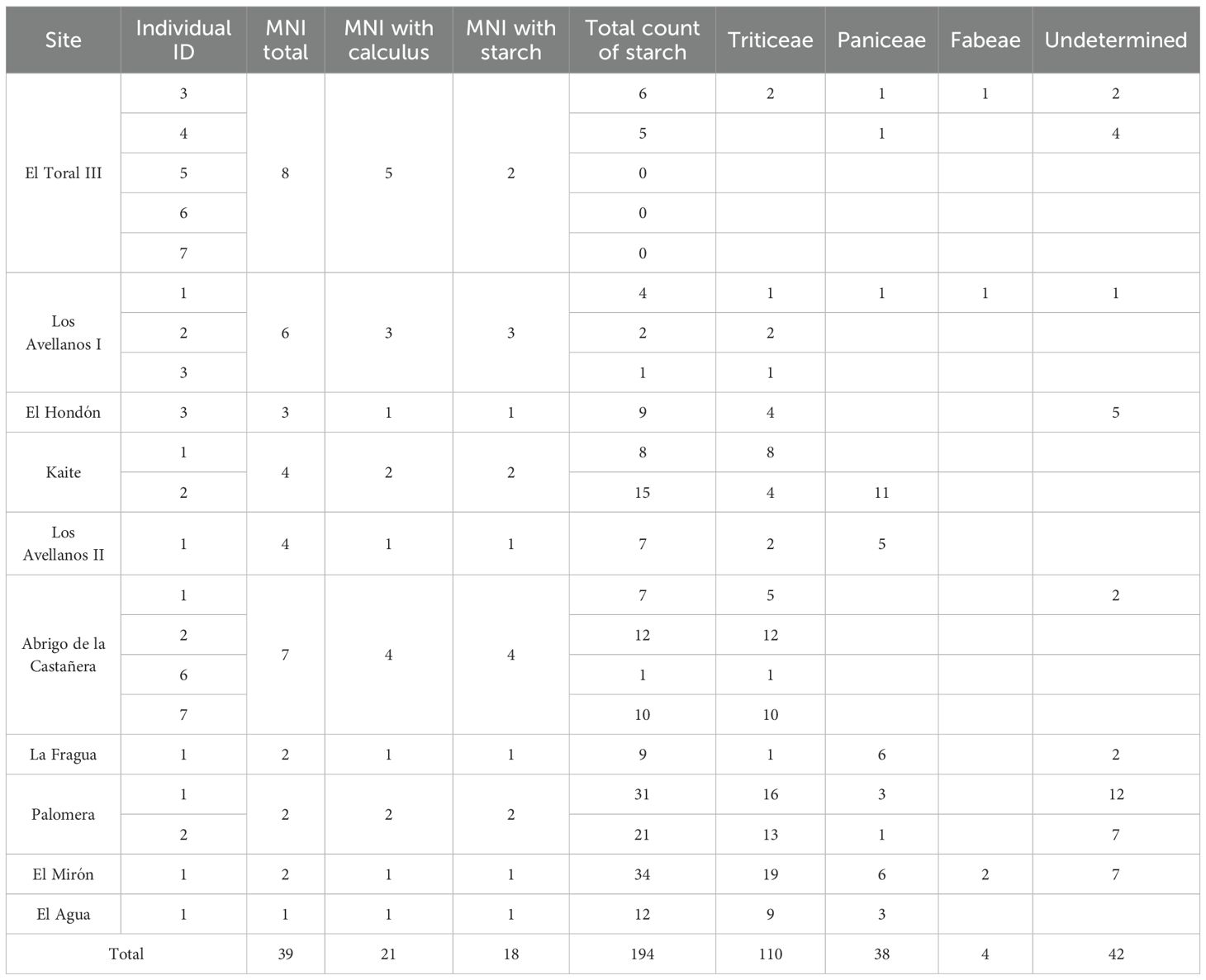
Table 2. Summary table of the quantitative results of starch grains identified by individual and site.
3.3 Morphological and statistical features of the archaeological starch grains
From the 194 starch grains identified, 152 granules were taxonomically addressed to a tribe level, and they correspond to Triticeae, Paniceae and Fabeae tribes. Triticeae grains were the most found in the sample and all periods, followed by Paniceae grains and the testimonial presence of Fabeae grains. The other 42 starch grains were assigned as undetermined because their small size prevents a more precise taxonomic identification. Based on their morphometric characteristics, three different morphotypes of starch grains were identified among calculus samples (Supplementary Table 2; Figure 2). Morphotype I represented 72.4% of the identified grains (n = 110), and it is characterized by a bimodal distribution typical of most grasses of the Triticeae tribe (Figure 3). Large grains (≤ 15 μm), defined as type A, are round to oval in 2D shape, lenticular in 3D, with a central or slightly sunken hilum and few lamellae. Smaller grains (< 10 μm), defined as type B, are almost spherical and have a central hilum. The mean size of these starch grains was 21.1 μm. The length of the Triticeae grains ranges between 4.7 and 63.4 μm. The size by sites was: El Hondón (14.6-35.6 μm), El Toral III (23.3-30.8 μm), Los Avellanos I (13.1-52.9 μm), Los Avellanos II (16.1-24.2 μm), La Castañera (4.7-35.3 μm), La Fragua (21.3 μm), El Mirón (6.4-56 μm), El Agua (9.5-49.5 μm), Kaite (8.3-48.4 μm), and Palomera (9.3-63.4 μm) (Supplementary Table 3). Morphotype II described 25% of the identified starch grains (n = 38). They have a 3D polyhedral shape, with a central hilum and fissures radiating from it. The extinction cross is visible, with straight arms, while lamellae are not identified. This type of grain is consistent with those of the Paniceae tribe (Figure 4). The mean size for all sites was 19.8 μm, while the length by site was: El Toral III (11.9-20 μm), Los Avellanos I (25.4 μm), Los Avellanos II (17.1-27.3 μm), La Fragua (13.9-16.9 μm), Palomera (23.8-33 μm), El Mirón (17.9-30.8 μm), and El Agua (8.2-12.5 μm) (Supplementary Table 3). Morphotype III represented 2.6% of the identified grains (n = 4) and has an oval/reniform morphology, with a Maltese cross -x- shaped, whose lamellae are visible and have fissures inside the hilum. These characteristics are usually common in legume species of the Fabeae family (Figure 5). The mean size of them was 31.8 μm. El Toral III Fabeae grain, whose length is 17.7 μm, is fractured and may have been processed. Los Avellanos I grain was greater and measured 52.9 μm, similar to the El Mirón grains, whose dimensions were 34 μm and 44.2 μm, respectively (Supplementary Table 3). The length of this starch grain was 9.3 μm (Supplementary Table 3). No other diagnostic morphotype of starch grain characteristic of domesticated plants or modern wild species of nuts or fruits was identified within the dental calculus samples (Figure 6).
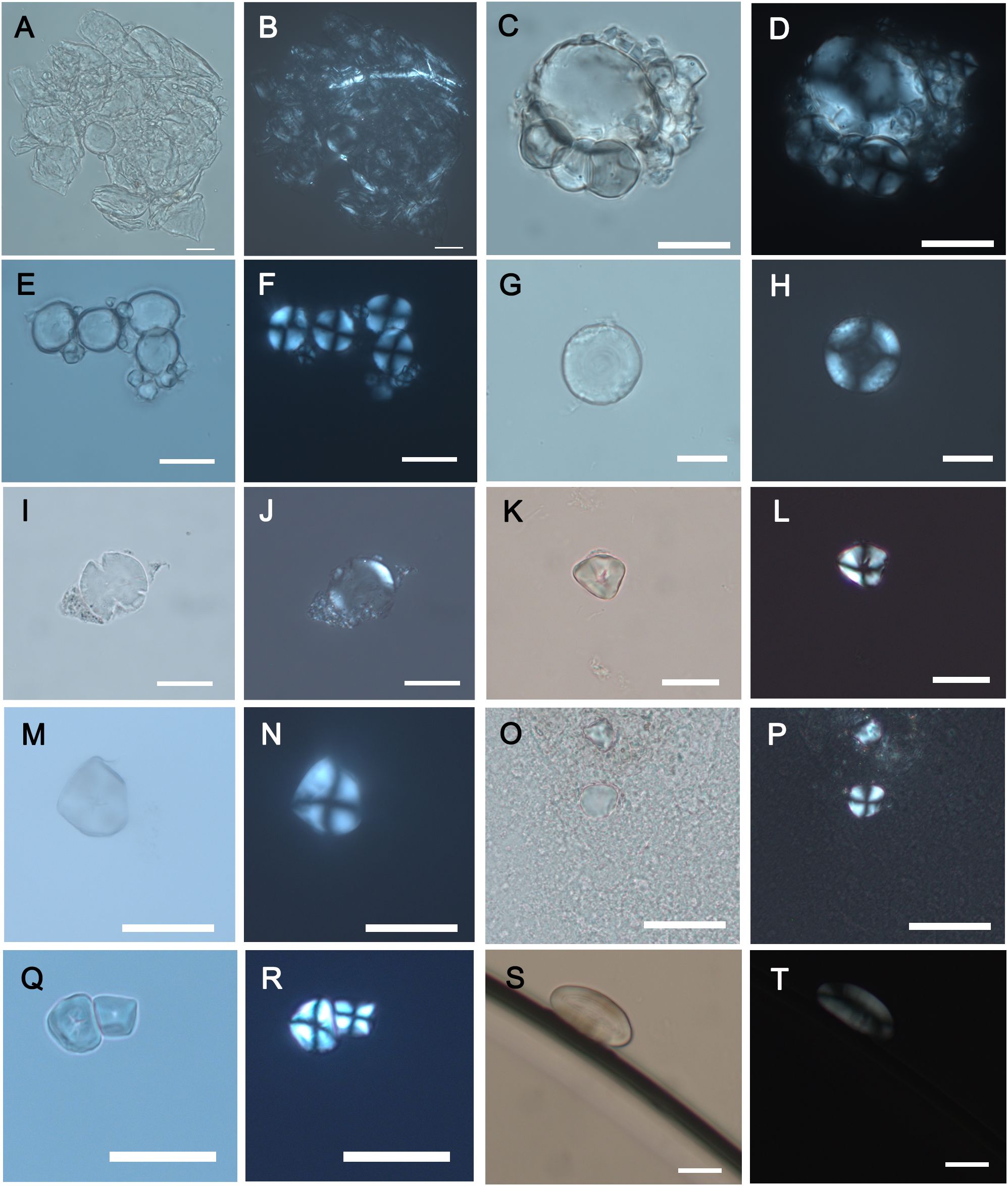
Figure 2. Archaeological starch grains embedded within the dental calculus of individuals under study, as viewed under a light microscope and a polarised light microscope. (A–J) Morphotype I consistent with the Triticeae tribe: (A–D) El Abrigo de la Castañera individuals 2 and 7; (E, F) El Mirón individual 1; (G, H) El Hondón individual 3; and (I, J) El Toral III individual 3. (K–R) Morphotype II associated with the Paniceae tribe: (K, L) Palomera individual 2; (M, N) Kaite individual 2; (O, P) El Toral III individual 3; and (Q, R) El Agua individual 1. (S, T) Morphotype III assigned to the Fabeae tribe from Los Avellanos I individual 1. (Scale bar, 20 μm).
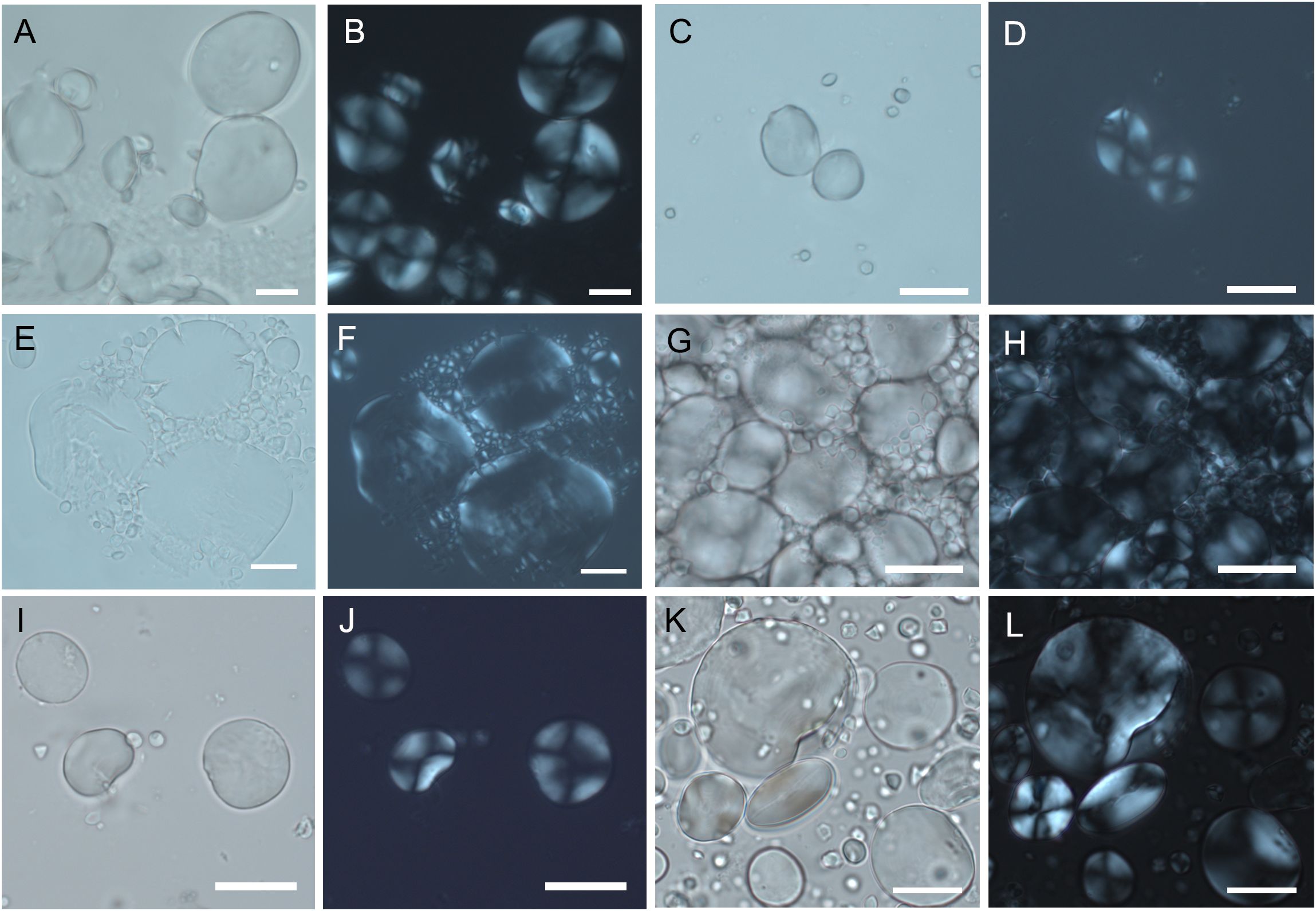
Figure 3. Experimental starch grains from different species of the Triticeae tribe. (A, B) T. aestivum. (C, D) T. monococcum. (E, F) T. dicoccum. (G, H) T. dicoccoides. (I, J) H. vulgare. (K, L) S. cereale. Starch granules in light microscope and polarized light microscope. (Scale bar, 20 μm).
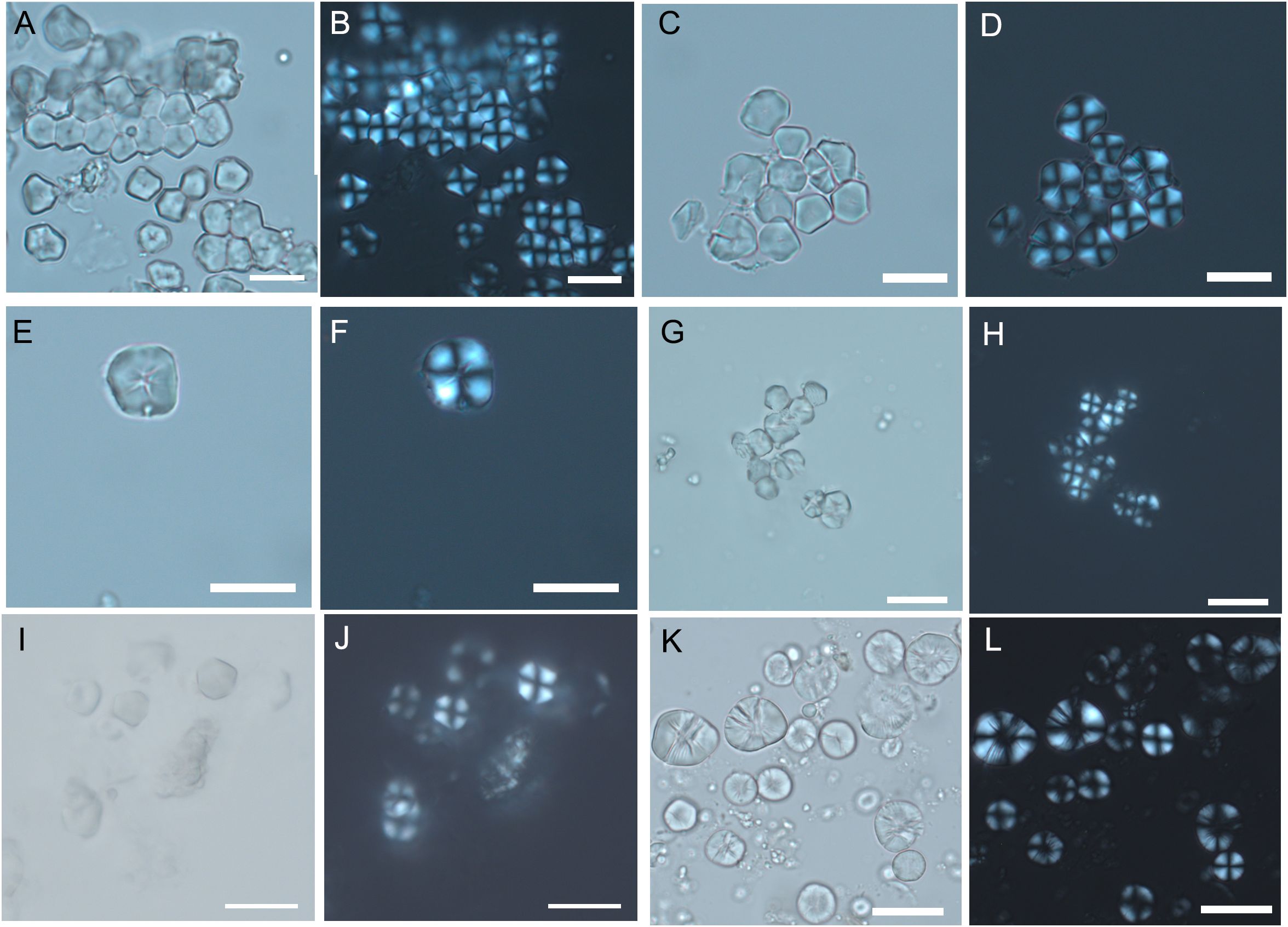
Figure 4. Experimental starch grains from different species of Paniceae and Andropogoneae tribes. (A, B) P. miliaceum. (C–F) S. italica. (G, H) S. viridis. (I, J) S. verticillata. (K, L) S. halepense. (Scale bar, 20 μm).
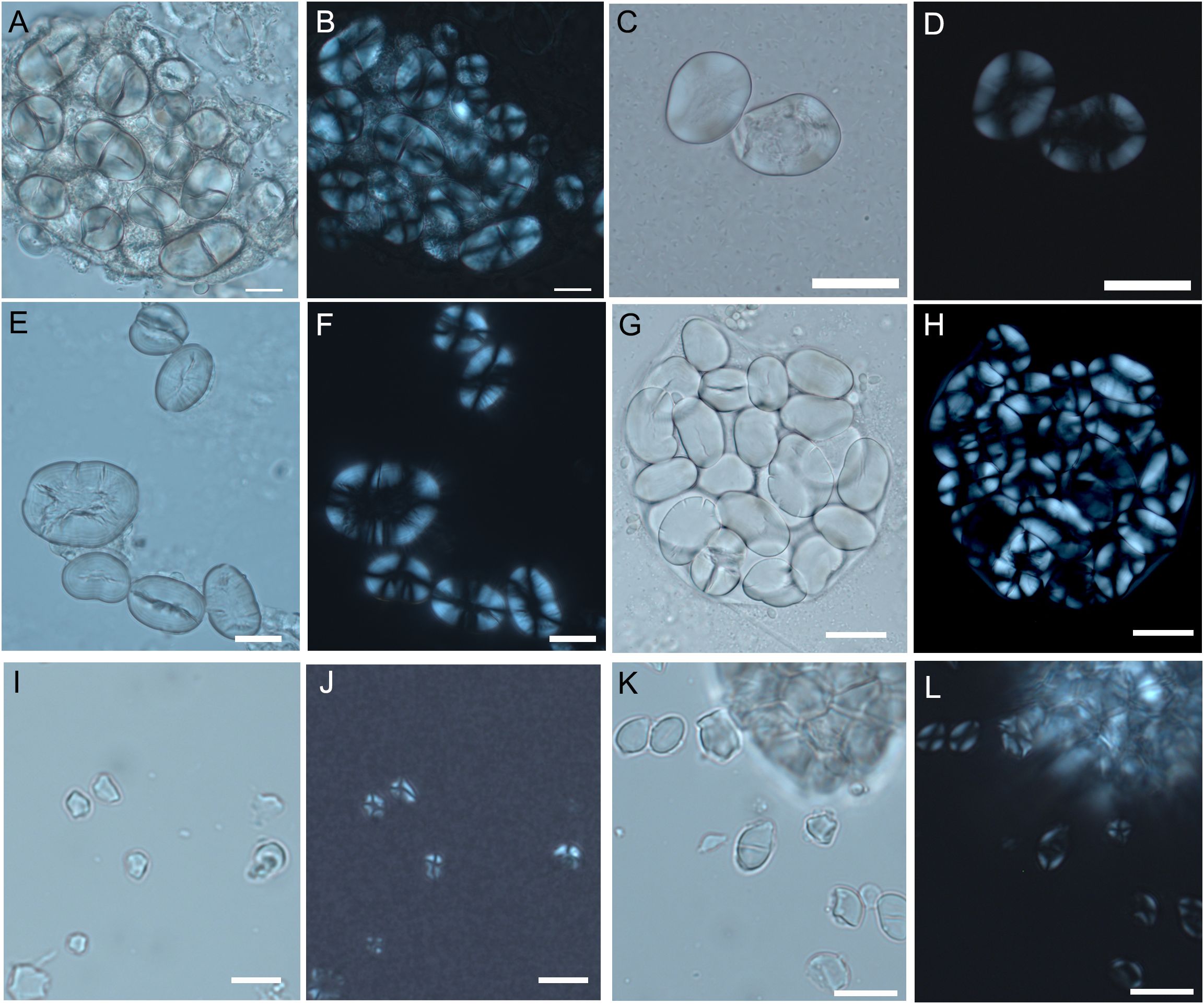
Figure 5. Experimental starch grains from different species of Fabeae and Aveneae/Poeae tribes. (A, B) V. sylvatica. (C, D) V. faba. (E, F) L. culinaris. (G, H) P. sativum. (I, J) A. fatua. (K, L) A. sativa. (Scale bar, 20 μm).
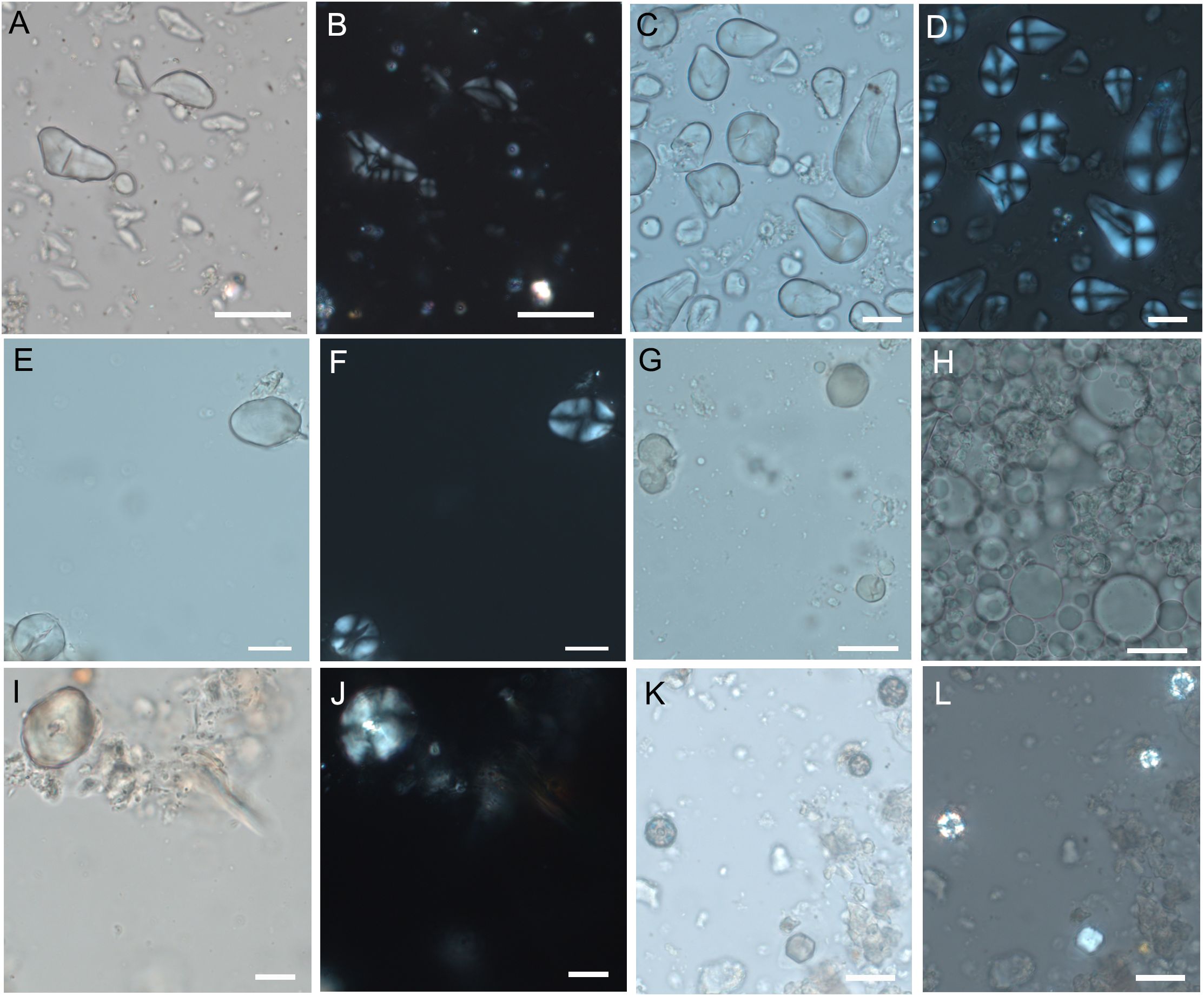
Figure 6. Experimental starch grains from different wild plants species (A, B) Q. robur. (C, D) Q. ilex. (E, F) C. avellana. (G) A. unedo. (H) P. pinea. (I, J) C. sanguinea. (K, L) P. spinosa. (Scale bar, 20 μm).
3.4 Measurement comparisons of experimental starch grains
Triticeae experimental starch grains were the tribe that provided the most homogeneous results between species (Supplementary Table 5). Although all of them showed great variability and were relatively common in terms of mean sizes, T. aestivum experimental grains were larger than others, ranging between 4.9-45.6 μm (mean size of 16.7 μm). This evidence is supported by the Pairwise Wilcoxon test, which highlights that T. aestivum starch measurements are significantly different from those obtained for all Triticeae sampled species (Supplementary Table 6). T. dicoccum size was lower than T. aestivum, ranging between 3.4-72 μm (mean size of 11.4 μm), but some starch grains reported the highest maximum values of the taxa analyzed. The rest of the Triticeae experimental species were very similar in size and did not report (or with very little significance) statistically significant differences (Supplementary Table 6). T. monococcum between 3.3-31.5 μm (mean size of 9.6 μm). H. vulgare between 3.6-34.7 μm (mean size of 9 μm). T. dicoccoides between 4.1-32.5 μm (mean size of 8.8 μm). And S. cereale between 2.9-50.3 μm (mean size of 8.5 μm) (Supplementary Table 5; Figure 7A).
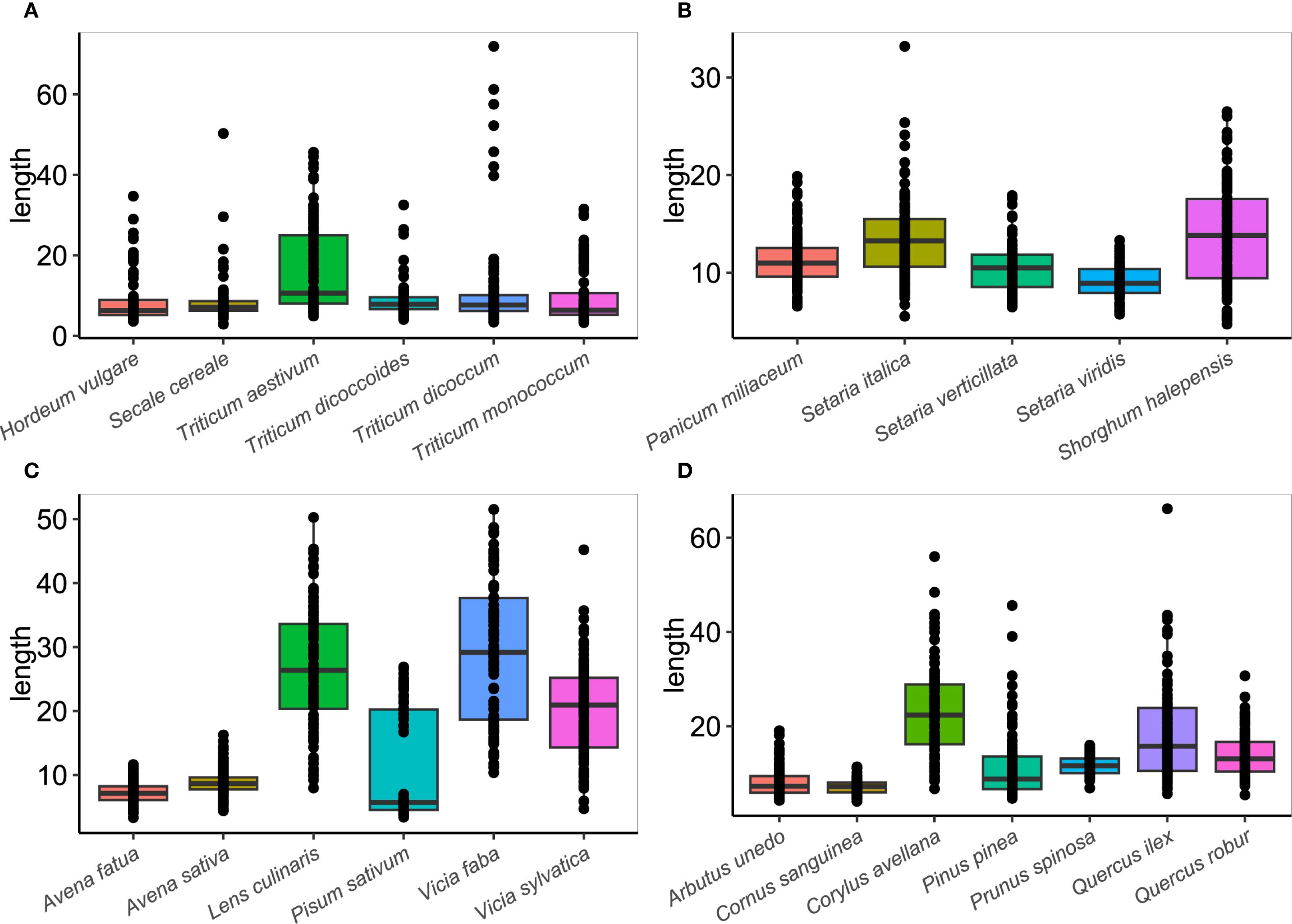
Figure 7. Boxplots of comparing the length of starch granules in 20 modern plant species (Length given in μm). (A) Triticeae tribe. (B) Paniceae and Andropogoneae tribes. (C) Aveneae/Poeae and Fabeae tribes. (D) Wild fruits species.
With regard to Paniceae/Andropogoneae experimental starch grains, the size of wild species was smaller than that of domestic ones. S. verticillata ranged between 6.5-17.9 μm (mean size of 10.4 μm), and S. viridis ranged between 5.7-13.3 μm (mean size of 9.1 μm). Among domestic millets, S. halepense was slightly greater than S. italica and P. miliaceum, ranging between 4.7-26.5 μm (mean size of 13.9 μm). S. italica ranged between 5.5-33.2 μm (mean size of 13.6 μm), being larger than P. miliaceum, which ranged between 6.6-19.9 μm (mean size of 11.4 μm) (Supplementary Table 5; Figure 7B). These results were confirmed through statistical comparison, with their lengths being significantly different, except for S. italica and S. halepense (Supplementary Table 6).
Fabeae experimental species showed the greatest variation among tribes. L. culinaris ranged lengths between 8-50.2 μm (mean size of 26.5 μm), V. faba ranged between 13.1-51.5 μm (mean size of 29.1 μm), V. sylvatica ranged between 4.7-45.2 μm (mean size of 20 μm), and P. sativum ranged between 3.4-26.9 μm (mean size 11 μm). L. culinaris and V. faba experimental grains were similar in size, and they had lengths larger than those of the wild native V. sylvatica grains and domestic P. sativum grains, especially in their minimum values (Supplementary Table 5; Figure 7C). In fact, L. culinaris and V. faba were the only species which did not show significant differences between them (Supplementary Table 6). Concerning the two Aveneae/Poeae species experimentally analyzed, both taxa had small sizes. A. sativa starch grains ranged between 4.4-16.3 μm (mean size of 8.9 μm), while A. fatua ranged between 3.3-11.6 μm (mean size of 7.3 μm). Thus, A. sativa reported slightly greater lengths than A. fatua (Supplementary Table 5; Figure 7C), being statistically different (Supplementary Table 6).
Finally, some wild fruit species were also statistically analyzed. C. avellana experimental starch grains were the greatest and most variable species, ranging between 6.7-56 μm (mean size of 23.3 μm). Q. ilex ranged between 5.7-66.2 μm (mean size of 18.4 μm), Q. robur ranged between 5.4-30.7 μm (mean size of 14 μm), P. pinea ranged between 4.7-45.6 μm (mean size of 11.5 μm), C. sanguinea ranged between 4.1-11.4 μm (mean size of 7.1 μm), P. spinosa ranged between 6.9-16 μm (mean size of 11.8 μm) and A. unedo ranged between 4.3-19 μm (mean size of 7.3 μm) (Supplementary Table 5; Figure 7D). All of them reported statistically significant differences (Supplementary Table 6).
3.5 Other plant microremains
A variety of other plant structures of probable non-dietary origin were also found entrapped with the tartar, including, from largest to smallest quantity, fibers, fungal spores, phytoliths, charcoals, pollen granules, and wooden tissues (Supplementary Table 4; Supplementary Figure 2). They may have been related to a wide range of non-dietary activities (Radini et al., 2017). The presence of plant fibers can be associated with different craft activities such as cordage, textile, basketry or net-making in which the mouth is used as a third hand (Cristiani et al., 2018). Although fungal spores may become useful food items, such as mushrooms and fungi, the ubiquity of fungal spores in the environment suggests that they may well result from accidental ingestion or inhalation (Hardy et al., 2016). On the other hand, phytoliths might have also been naturally present in the environment, water, and soil, suggesting the inclusion of phytoliths in dental calculus for non-dietary reasons (Norström et al., 2019). Charcoals may also have reached the mouth by accidental ingestion through food and/or breathing due to exposure to hearths (Hardy et al., 2016). And pollen granules are most likely the result of inhalation, reflecting the environmental conditions in which this individual lived (Hardy et al., 2018). Finally, the wood inclusion in calculus can also vary from the use of a toothpick to crafting activities (Cristiani et al., 2021).
4 Discussion
4.1 Starch grain variability between individuals, sites, geographic environments and chronological phases
Triticeae grains were recovered at all sites, individuals and cultures, being quite frequent in all the samples. This trend suggests a broader cultural pattern in which Triticeae species were cultivated across regions, both in coastal and inland landscapes, and all individuals consumed these cereals. The individuals with more Triticeae starch grains were dated in the Early and Middle Bronze Age, while Late Neolithic and Chalcolithic individuals provided fewer quantities of granules. These differences in the number of starch grains recovered in each individual could be explained by different consumption among the members of these communities, or instead, by the state of preservation of the dental calculus of the oldest individuals. Concerning Paniceae grains, they are abundant in the samples, but quantitatively lower than Triticeae grains, suggesting that these crops were of lesser importance in the diet of these groups. They were also documented in all cultures and geographic locations, but only in eight of the ten sites and in ten of the 18 individuals, indicating a greater variability among individuals and sites. There are sites where these grains have not been preserved in all individuals (i.e. Los Avellanos I), and even sites without any evidence of their consumption (i.e. Abrigo de La Castañera). At the individual level, most of the individuals show an equal, low proportion of Paniceae grains, except for Individual 2 from Kaite, an adolescent female with signs of hypoplasia and caries. On the other hand, Fabeae starch grains were identified in three individuals and sites: El Toral III and Los Avellanos I (Late Neolithic) and El Mirón (Middle Bronze Age). However, they were recovered in a very small proportion, which prevents us from interpreting why they appear indistinctly in some individuals and specific chronological periods.
The FAMD distinguished two groupings of the different archaeological starch grains (Figure 8). The first one is represented by the sites analyzed in this work and dated from the Late Neolithic to the Middle Bronze Age. Triticeae starch grains are more commonly grouped with these sites and periods, although they are located in the center of both dimensions, suggesting that these cereals were widely cultivated across cultures. The second one is characterized by an assemblage studied previously by us and dated to the Late Bronze Age (González-Rabanal et al., 2022). Paniceae starch grains are predominantly associated with this site, indicating that these species are more highly associated with the Late Bronze Age and supporting the turning point in the Late Prehistory of the Cantabrian Region. Instead, Fabeae is located at the upper right side of the FAMD but is more associated with the first grouping. However, their small sample size in the record can constitute a comparative bias.
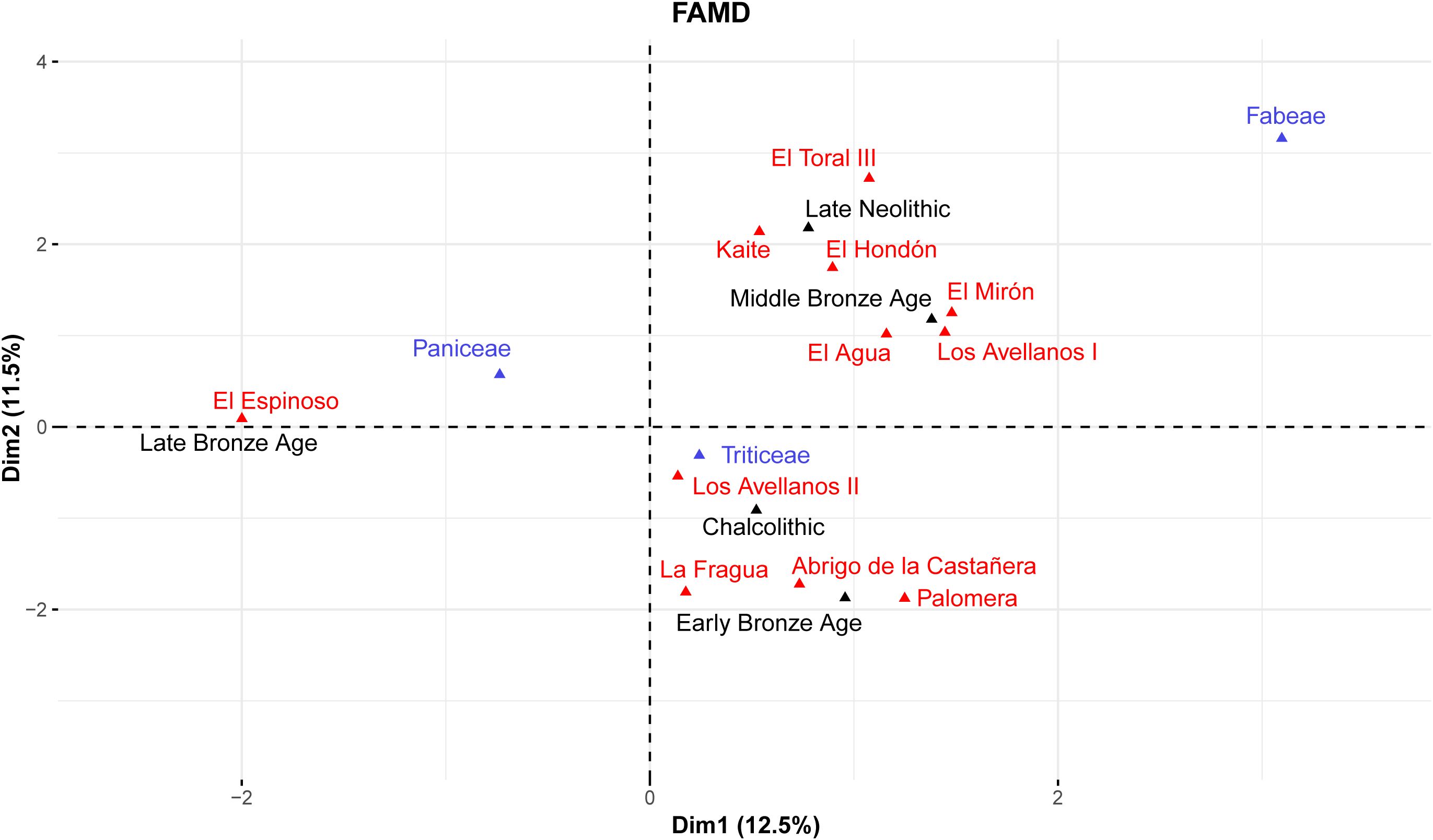
Figure 8. Factor analysis of mixed data showing the association of the archaeological samples (this study and González-Rabanal et al., 2022) grouped by site, culture and starch grain morphotype, considering quantitative and qualitative variables.
4.2 The cultivation of wheat and barley species during late prehistory
The starch grains most documented in this study have been morphologically assigned to the Triticeae tribe (Henry and Piperno, 2008) (Figure 2). This morphotype was already common among hunter-gatherers of Balkans and Italy (Cristiani et al., 2016; 2018; 2021; Nava et al., 2021; Oxilia et al., 2021; Carra et al., 2023). Likewise, this tribe is the most identifiable starch morphotype in the dental calculus analyses conducted in Iberian farmers up to now (Bucchi et al., 2019; González-Rabanal et al., 2022; Salazar-García et al., 2022; Coutinho et al., 2024), suggesting that the cultivation of different species of wheat and barley was key to the economy of these farming societies from the Neolithic onwards. Previous experimental analyses in species of the Triticeae tribe have revealed that wild starch grains (Aegilops genus) show a larger size distribution than domestic ones (Triticum and Hordeum genera) (Cristiani et al., 2021). Our experimental measurements support the morphological features of the archaeological starch grains and have revealed a wide range of sizes for the different Triticeae domestic species, avoiding their taxonomic identification to species level (Supplementary Table 5; Figure 3). This finding aligns with previous experimental analyses (Bocanegra and Sáez, 2012; Cristiani et al., 2016). In fact, there is no homogeneity in the existence of statistically significant differences between the length of the archaeological and experimental starch grains of the Triticeae tribe, varying the presence and/or absence of significance between species and sites (Supplementary Table 7). However, the morphology and size obtained for archaeological starch grains type A are compatible with those of larger starch grains of wheat and barley species (Figure 7A; Figure 9A).
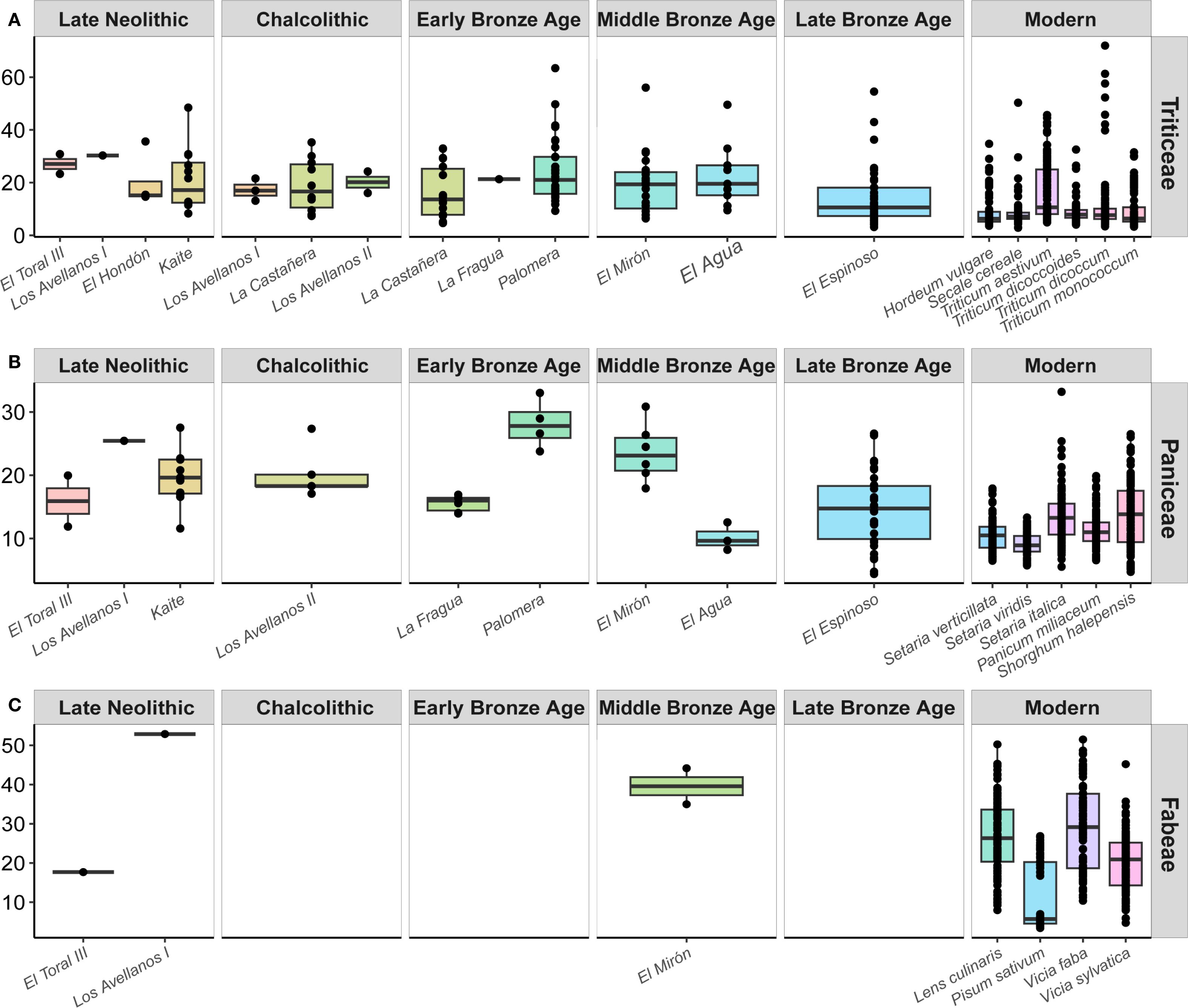
Figure 9. Boxplots comparing the length of archaeological (this study and González-Rabanal et al., 2022) and experimental starch granules by site and species (Length given in μm). (A) Triticeae tribe. (B) Paniceae and Andropogoneae tribes. (C) Fabeae tribe.
Wheat and barley were the most widely cultivated cereals in Iberia during Late Prehistory (Buxó and Piqué, 2008). Carpological evidence for the Cantabrian region supports the presence of H. vulgare (naked and hulled varieties), T. dicoccum, T. monococcum, T. aestivum and T. durum in several Cantabrian sites such as Arangas, El Mirón, Los Gitanos, Arenaza, Kobaederra, Lumentxa or Pico Ramos (Zapata, 2002; López-Dóriga, 2016) (Table 3). In fact, direct dating of cereal seeds from El Mirón, Kobaederra and Pico Ramos demonstrates the early cultivation of these cereals since the second half of the fifth millennium cal BC (Zapata et al., 2004; Peña-Chocarro et al., 2005; Zapata et al., 2007). Such an early date is less likely for such species as S. cereale, which did not reach the peninsula until the Iron Age (Seabra et al., 2023). Thus, our methodological approach to the identification of starch grains in dental calculus corroborates the archaeobotanical data previously obtained for this region. Similarly, stable isotope evidence supports this attribution, so all the individuals analyzed here show a C3 terrestrial diet based on the consumption of cereals, meat and dairy products (Jones et al., 2019; González-Rabanal et al., 2020; González-Rabanal, 2022).
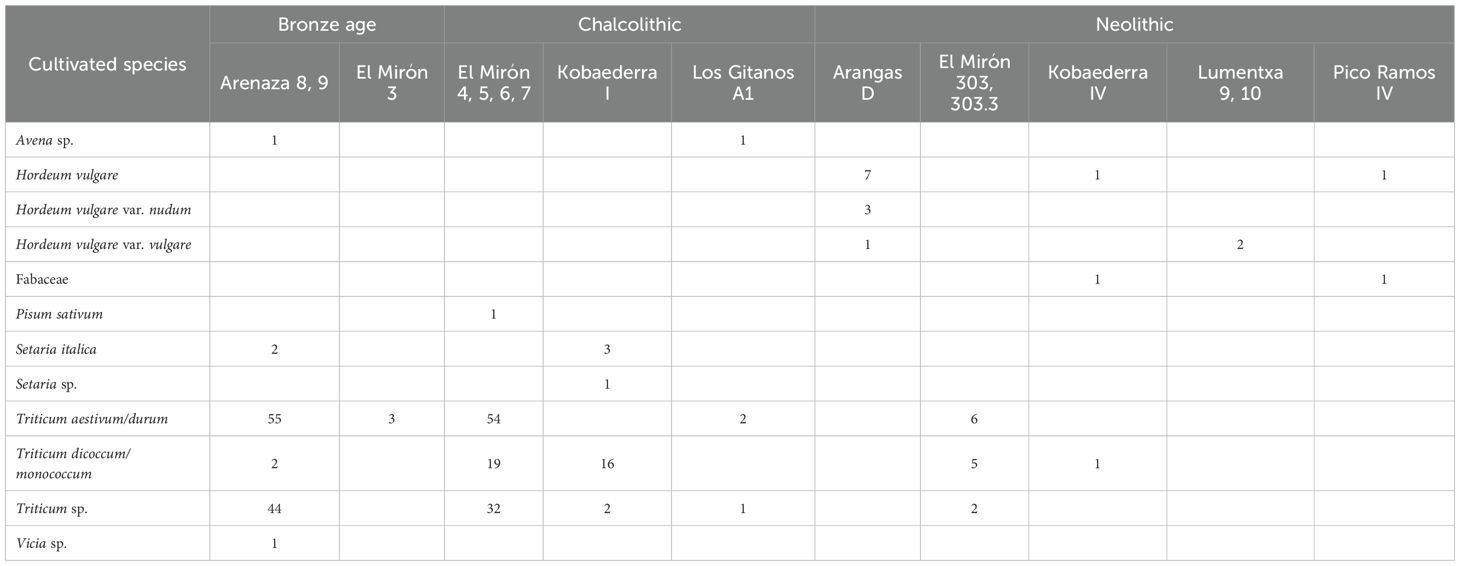
Table 3. Carpological data from Cantabrian sites/layers by cultural period.
The combination of naked and hulled wheat varieties such as emmer (T. dicoccum), einkorn (T. monococcum) or hard wheat (T. durum) and bread wheat (T. aestivum), and hulled and naked barley (H. vulgare var. vulgare and H. vulgare var. nudum) was possible in some periods of the Late Prehistory of the Cantabrian Region (Zapata and Pena-Chocarro, 2005). However, hulled species were more common during the Early Neolithic, while free-threshing wheats were almost absent, reflecting peculiar features for agricultural practices, suggesting that hulled species were primarily cultivated due to ecological factors. Hulled wheats are resistant to poor soil conditions and fungal diseases, and barley also tolerates poorer soils (Zapata et al., 2004). Similarly, they demonstrate a good adaptation to wet and mountainous conditions. As part of the Eurosiberian region, the Cantabrian Region is a mountainous area with an oceanic climate with generally mild temperatures and high precipitation, and no notable summer droughts. Only in this particular region of Iberia is emmer still cultivated using traditional methods due to its favorable yields under these conditions (Peña-Chocarro and Zapata, 1998). The late adoption (5th millennium cal BC) of a farming economy in this region (Fano et al., 2015) would be in line with this hypothesis. The first cereals cultivated in this geographical area would have taken some time to acclimate to the new ecological conditions, influenced by the geographic characteristics of the short, narrow valleys that descend from the Cantabrian Cordillera to the Atlantic shore (Peña-Chocarro et al., 2005), a landscape more suitable for pastoralism in the initial stage of the neolithization process (López-Merino et al., 2010).
4.3 Dental calculus evidence of millet consumption, but when?
Polyhedral starch grains (morphotype II) found in the sample may be morphologically attributed to different Paniceae/Andropogoneae plants, such as millet species (Figure 2). The identification of starch grains from wild grasses of the Paniceae and Andropogoneae tribes was found among foragers (Cristiani et al., 2016; 2021; Nava et al., 2021; Oxilia et al., 2021). Based on their average size and morphological features this morphotype was also identified within the dental calculus of farmers in the Chalcolithic-age farmers from El Mirador cave in the Sierra de Atapuerca (Burgos, Spain) (Bucchi et al., 2019), in the Chalcolithic/Bronze Age site of Grotta dello Scoglietto (Tuscany, Italy) (Mariotti Lippi et al., 2017), in the Bronze Age site of Laderas del Castillo (Alicante, Spain) (Romero et al., 2024) and in the Late Bronze Age site of El Espinoso (Asturias, Spain) (González-Rabanal et al., 2022). The experimental starch grains from Paniceae species studied in this work exhibit similar morphological features as the archaeological starch grains, excepting S. halepense (Andropogoneae tribe), which showed different morphological traits than the archaeological ones (Supplementary Table 5; Figures 4, 7B). However, their length suggests that there are notable differences in grain size distributions between experimental and most of the oldest archaeological samples such as the Late Neolithic individuals from El Toral III, Los Avellanos I and Kaite, the Chalcolithic individual from Los Avellanos II, the Early Bronze Age individuals from Palomera and the Middle Bronze Age individual from El Mirón (Figure 9B). These differences in size are corroborated by the Pairwise Wilcoxon test, finding significant differences with all the experimental Paniceae species analyzed, except in cases in which the archaeological sample size bias the results (Supplementary Table 7). For these reasons, we can consider as aff. Paniceae to this set of starch grains. Instead, the Paniceae starch grains from the Early/Bronze Age individuals from La Fragua and El Agua are similar in size to the large starch grains of S. italica and P. miliaceum. In this case, the statistical comparisons showed no significant differences between the length of the Paniceae grains of these sites, and the S. italica grains, similar to what happens with the El Espinoso Late Bronze Age site (Supplementary Table 7). With respect to the experimental species analyzed, S. italica can have occasional oval grains and a centric hilum traversed by fissures which vary in form. In contrast, starch grains from P. miliaceum have a mostly polyhedral shape with a centric hilum where fissures are less common (Yang et al., 2012). Experimental starch grain size differs slightly between both species, being marginally smaller and statistically different the grains of P. miliaceum (Supplementary Table 6). Wild millet species, such as S. viridis or S. verticillata are even smaller, with a more characteristic spherical morphology and short fissures (Madella et al., 2016; González-Rabanal et al., 2022), which allows us to discard them as the source of the archaeological starch grains. The duality mentioned above could indicate that the larger starch grains of the aff. Paniceae tribe of the oldest individuals would belong to different, still underexplored species than those smaller Paniceae grains documented in some Bronze Age individuals. A great variety of Paniceae and Andropogoneae species grow in water environments (Cristiani et al., 2021), and a mixture of species from such genera might likely have been used by Cantabrian human groups for different purposes, while the domestic millets could be the source of the smallest Paniceae archaeological starch grains. This hypothesis could explain the differences in size between some archaeological Paniceae starch grains and the experimental ones.
The first carpological evidence of millet exploitation in Iberia dates to the Middle Bronze Age (Buxó and Piqué, 2008). However, millets were not systematically exploited until the Late Bronze Age and Iron Age (Tereso et al., 2016). Millets are poorly represented in the archaeological record of the Cantabrian Region. Macroremains of S. italica were identified in Kobaederra (Level 1) and Arenaza (Layer 9), chronologically assigned to the Chalcolithic and Early Bronze Age, respectively (Zapata, 2002) (Table 3). However, those seeds have not been directly dated; thus, we cannot exclude the possibility that they might be intrusive. Moreover, there is a notable absence of P. milliaceum seeds in the carpological records of the Cantabrian Region. By contrast, the carpological record of Northwest Iberia (Galicia and Northern Portugal) is richer, with both species being abundant in Middle and Late Bronze Age settlements (Bettencourt, 2000; Tereso et al., 2016; Figueiral et al., 2017; Jesus et al., 2020). Anyway, both species might have been consumed in a mixed way since millet poly-cropping has been suggested ethnographically (Moreno-Larrazabal et al., 2015; Teira-Brión, 2022). On the other hand, S. halepense can be discounted as a potentially consumed species since the morphology of its starch grains is quite different from those of S. italica and P. miliaceum and sorghum did not reach Iberia until the Middle Ages (Pérez-Jordà et al., 2024). No carpological evidence of the wild native species (Panicum repens L., S. viridis, or S. verticillata) has been found in the Holocene sites of northern Iberia, although today, they are widely distributed across central and southern Europe, but are almost unknown in Atlantic Europe (Pyankov et al., 2010).
The Late Neolithic/Chalcolithic individuals of El Toral III, Kaite and Los Avellanos I and II have reported some sparse Paniceae grains, but their belonging to domestic millets is discounted since they are larger than experimental millet species and a prior arrival of these species is not supported by carpological research, and they probably belong to the group widely known as “forgotten millets” or other Paniceae/Andropogoneae taxa (Lucarini et al., 2016). More likely is this association in the Early Bronze Age individuals of La Fragua and Palomera, although the archaeobotanical evidence for this period is very doubtful, and both sites provided differences in starch grain size. Finally, the starch grains from the Middle Bronze Age individuals, at least those from El Agua, offer a higher probability of belonging to domestic millets because in this period there is carpological evidence of millets. If we pay attention to the stable isotope evidence of Late Neolithic/Chalcolithic and Early/Middle Bronze Age individuals, the carbon isotope values do not support the consumption of C4 plants in the Cantabrian Region (Arias, 2005; Sarasketa-Gartzia et al., 2018; Jones et al., 2019; González-Rabanal et al., 2020). In any case, if millets were already cultivated at this time, their consumption may have been occasional because the long-term bone collagen record does not register the carbon enrichment typical of eating millets. Thus, the agriculture of millet species was not fully developed until the Late Bronze Age, as supported by stable isotopes and dental calculus from El Espinoso cave (González-Rabanal et al., 2022). In that research, in addition to the high carbon isotope values, we identified 74 starch grains, of which 43 belong to the Triticeae tribe and 29 to the Paniceae tribe. As millets allow for two growing seasons per year, other crops such as those of the Triticeae tribe could also have been cultivated alongside them annually. These findings reveal a more diversified and mixed agriculture during this time, marking the primary shift in the farming economy of the Cantabrian Region during Late Prehistory.
4.4 The consumption of other plant resources
The existence of starch grains within dental calculus identified as morphotype III suggests that some legumes were also part of the diet of human groups during the Holocene (Figure 2). However, the sparse evidence of their starch grains prevents calculating the real weight in their diet. All Fabeae grains are large granules that fall within the margins observed in experimental species of L. culinaris and V. faba (Supplementary Table 5; Figures 5, 7C, 9C), whose maximum lengths also coincide with this size. In fact, there are no significant differences between the archaeological Fabeae starch grains and the experimental ones (Supplementary Table 7). V. sylvatica and P. sativum experimental starch grains were statistically smaller; therefore, the archaeological grains likely belong to a domestic legume species. Wild species of the family Fabaceae are well-represented in the Early Mesolithic and Late Mesolithic individuals from the Danube Gorges area of the Balkan peninsula (Cristiani et al., 2016; 2021). In Iberia, a substantial quantity of starch grains was recovered from the Early Neolithic individuals at La Dehesilla cave (Cádiz) (Coutinho et al., 2024).
Legumes are traditionally poorly represented in archaeobotanical records due to taphonomic problems related to their processing. Although the existence of legumes in the Cantabrian archaeobotanical record is unknown until the Neolithic, some wild legumes could have been present in this region since the beginning of the Holocene. In fact, preliminary anthracological analysis of El Toral III has documented charcoals containing burnt legume remains in several stratigraphic units of the Mesolithic shell midden (Gutiérrez-Zugasti, personal communication). Later, several legumes such as lentil (L. culinaris), pea (P. sativum) broad bean (V. faba), grass and red pea (Lathyrus sativus L. and Lathyrus cicera L.), bitter betch (Vicia ervilia L. Willd.) and common vetch (Vicia sativa L.) were common in the Early Neolithic carpological record of numerous regions of Iberia (Zapata et al., 2004; Peña-Chocarro et al., 2018). In contrast, seeds of these species are absent in the Cantabrian Region, although some Fabaceae vegetal remains have been found in the Neolithic levels of El Mirón, Kobaederra, Lumentxa and Pico Ramos (Table 3). The first taxonomically legume remains are a Vicia sp. seed documented in the Arenaza Bronze Age layer 9 and a P. sativum seed discovered in El Mirón Chalcolithic level 4. Likewise, V. faba and P. sativum were widely identified in several sites of northwest Iberia during the Bronze Age (Tereso et al., 2016); therefore, it would not be surprising if these crops were also cultivated in other northern areas of the peninsula. Therefore, it is possible that some domestic legumes, such as peas, lentils, or beans, were used as food supplements by the Cantabrian communities from the Neolithic onwards.
Our results have not shown any starch grains with key morphological characteristics to be assigned to the Aveneae/Poeae tribe. However, a single, dubious angular starch was discovered in an Early Bronze Age individual from La Fragua. This grain has an irregular morphology, a central hilum and a visible but asymmetric Maltese cross. The lamellae are not observable. This morphology can be compatible with that documented in modern species of the Aveneae/Poeae tribe (Figure 5), but the absence of other morphological traits and its poor preservation suggest caution. For these reasons, it has been considered undetermined. The length of this starch grain was 9.3 μm (Supplementary Table 3), being consistent with the averages obtained in our experimental oat species (Supplementary Table 5, Figure 5, Figure 7C). Several starch granules of the Aveneae/Poeae tribe have already been identified in dental calculus from foragers, especially during the Late Mesolithic in the Balkans (Cristiani et al., 2018; 2021) and within Chalcolithic/Bronze Age individuals from El Mirador (Spain) and Grotta dello Scoglieto (Italy) (Mariotti Lippi et al., 2017; Bucchi et al., 2019). For the Cantabrian Region, carpological references to Aveneae/Poeae species are very scarce, with possible grains of Avena sp. in the A1 level of Los Gitanos (López-Dóriga, 2016) and in layer 9 of Arenaza (Zapata, 2002) (Table 3). The Iberian archaeobotanical record indicates that Avena species were not cultivated until a late stage of Late Prehistory (Buxó and Piqué, 2008). Thus, both archaeobotanical and dental calculus Iberian evidence suggest that, if consumed, these crops, typical of temperate ecosystems, would be cultivated sporadically and in addition to other cereals.
So far, no other starch grains from flax (Linum usitatissimum L.), poppy (Papaver somniferum L.) or additional crops have been identified within the dental calculus of late prehistoric humans in Cantabrian Spain. Similarly, starch grains associated with wild fruits have not been identified (Figure 6), although the smallest starch grains without taxonomical identification could belong to them. Wild nuts and fruits such as hazelnuts, acorns and Rosaceae fruits must have played a major role in the diet of the farming societies, as they were abundant, predictable, and storable food resources in this region (Zapata, 2000). In fact, some species are abundant in the Cantabrian carpological record, mainly hazelnuts and acorns (Zapata, 2002; López-Dóriga, 2016). Moreover, the anthracological analysis of charcoal remains has indicated the exploitation of several woody taxa in the archaeological sites of the region, and their wood was probably used as fuel (Uzquiano, 1995).
5 Conclusions
This research has allowed us to demonstrate, for the first time, that dental calculus analysis constitutes a valuable proxy for reconstructing agricultural practices during Late Prehistory in northern Iberia, providing visibility to the farming activities limited by the scarce carpological record. The combination of archaeobotanical and isotopic data and the starch grains entrapped in human dental calculus highlights a deep, diachronic approach to the farming diet of Cantabrian human groups. Our results support an extensive and stable agriculture based on the cultivation of wheat and barley species from the Early Neolithic to the Bronze Age, when millets were introduced and became major crops, integrated into a mixed farming economy. New sampling of Early Neolithic individuals will help to refine our knowledge of the origin and development of the first crops in this area.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author contributions
BG-R: Data curation, Writing – review & editing, Conceptualization, Methodology, Software, Writing – original draft, Investigation, Visualization, Validation, Formal Analysis. ABM-A: Funding acquisition, Supervision, Resources, Project administration, Writing – review & editing. EC-B: Writing – review & editing, Resources. DC-S: Funding acquisition, Resources, Writing – review & editing. IG-Z: Funding acquisition, Resources, Writing – review & editing. MAM-M: Writing – review & editing, Resources. AIO-M: Resources, Writing – review & editing, Funding acquisition. LGS: Writing – review & editing, Funding acquisition, Resources. CV-M: Resources, Writing – review & editing. MRGM: Supervision, Resources, Funding acquisition, Writing – review & editing, Project administration. EC: Resources, Funding acquisition, Writing – review & editing, Formal Analysis, Visualization, Methodology, Data curation, Validation, Supervision, Conceptualization, Project administration.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. BG-R benefited from a Juan de la Cierva Grant (JDC2022-048798-I), funded by MCIN/AEI/10.13039/501100011033 and the European Union «NextGeneration EU»/PRTR. This study was part of BG-R’s Doctoral dissertation, supervised by ABM-A and MRGM. This research was funded by the research projects of the Spanish Economy, Industry and Competitiveness Ministry HAR2016-75605-R to MRGM and HAR2017- 84997-P to ABM-A The analysis of dental calculus was carried out under the facilities of the ERC Starting Grants (HIDDEN FOODS ref. 639286 and SUBSILIENCE ref. 818299), awarded to EC and ABM-A. IG-Z, DC-S, MRGM and LGS were supported by a grant from the Consejería de Cultura, Deporte y Turismo of the Gobierno de Cantabria. We also thank the research teams that recovered the human remains involved in this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1644052/full#supplementary-material
References
Abbo S. and Gopher A. (2017). Near eastern plant domestication: A history of thought. Trends Plant Sci. 22, 491–511. doi: 10.1016/j.tplants.2017.03.010
Allentoft M. E., Sikora M., Refoyo-Martínez A., Irving-Pease E. K., Fischer A., Barrie W., et al. (2024). Population genomics of post-glacial western Eurasia. Nature 625(7994), 301–311. doi: 10.1038/s41586-023-06865-0
Arias P. (2005). “Determinaciones de isótopos estables en restos humanos de la región Cantábrica,” in Aportación al estudio la dieta las poblaciones del Mesolı́tico y el Neolı́tico. Munibe Antropologia-Arkeologia, (Cambridge: Cambridge University Press), 57, 359–374.
Arias P. (2007). “Neighbours but diverse: social change in north-west Iberia during the transition from the Mesolithic to the Neolithic, (5500–4000 cal BC),” in Going over: The Mesolithic-Neolithic Transition in Northwest Europe. Eds. Whittle A. and Cummings V., 53–71.
Bettencourt A. M. S. (2000). O povoado da Idade do Bronze da Sola, Braga, norte de Portugal. Cadernos Arqueologia 9, 29–44.
Bocanegra F. J. A. and Sáez J. A. L. (2012). Caracterización morfológica de almidones de los géneros Triticum y Hordeum en la Península Ibérica. Trabajos Prehistoria 69, 332–348.doi: 10.3989/tp.2012.12095
Bucchi A., Burguet-Coca A., Expósito I., Aceituno Bocanegra F. J., and Lozano M. (2019). Comparisons between methods for analyzing dental calculus samples from El Mirador cave (Sierra de Atapuerca, Spain). Archaeol. Anthropol. Sci. 11, 6305–6314. doi: 10.1007/s12520-019-00919-z
Buxó R. and Piqué R. (2008). Arqueobotánica: los usos de las plantas en la Península Ibérica (Barcelona: Grupo Planeta (GBS)).
Carra M., Zupancich A., Fiorin E., Sarti L., Lo Vetro D., Martini F., et al. (2023). Plant foods in the Late Palaeolithic of Southern Italy and Sicily: Integrating carpological and dental calculus evidence. Quaternary international: J. Int. Union Quaternary Res. 653–654, 53–68. doi: 10.1016/j.quaint.2022.06.007
Coutinho A. P., Moreira M., Silva E., García-Rivero D., and Umbelino C. (2024). Identification of neolithic diet by the morphology of the starch grains of dental calculus found in the Dehesilla Cave (Cadiz – south of the Iberian peninsula). Archaeol. Anthropol. Sci. 16, 62. doi: 10.1007/s12520-024-01972-z
Cristiani E., Radini A., Borić D., Robson H. K., Caricola I., Carra M., et al. (2018). Dental calculus and isotopes provide direct evidence of fish and plant consumption in Mesolithic Mediterranean. Sci. Rep. 8, 8147. doi: 10.1038/s41598-018-26045-9
Cristiani E., Radini A., Edinborough M., and Borić D. (2016). Dental calculus reveals Mesolithic foragers in the Balkans consumed domesticated plant foods. Proc. Natl. Acad. Sci. USA 113, 10298–10303. doi: 10.1073/pnas.1603477113
Cristiani E., Radini A., Zupancich A., Gismondi A., D’Agostino A., Ottoni C., et al. (2021). Wild cereal grain consumption among Early Holocene foragers of the Balkans predates the arrival of agriculture. eLife 10, 1–37. doi: 10.7554/eLife.72976
Cubas M., Altuna J., Álvarez-Fernández E., Armendariz A., Fano M.Á., López-Dóriga I. L., et al. (2016). Re-evaluating the neolithic: the impact and the consolidation of farming practices in the Cantabrian region (Northern Spain). J. World Prehistory 29, 79–116. doi: 10.1007/s10963-016-9091-2
Dobney K. (1994). “Study of the dental calculus,” in The Jewish Burial Ground at Jewbury. Eds. Addyman P. and Kinsler V. (Council for British Archaeology, York), 507–521.
Dobney K. and Brothwell D. (1986). Dental calculus: its relevance to ancient diet and oral ecology. Teeth Anthropology 291, 55–81.
Fano M.Á., Cubas M., and Wood R. (2015). The first farmers in Cantabrian Spain: Contribution of numerical chronology to understand an historical process. Quaternary Int. 364, 153–161. doi: 10.1016/j.quaint.2014.09.026
Figueiral I., Jesus Sanches M., and Cardoso J. L. (2017). Crasto de Palheiros (Murça, NE Portugal, 3rd - 1st millennium BC): from archaeological remains to ordinary life. Estudos do Quaternário 17, 13–28. doi: 10.30893/eq.v0i17.158
Fiorin E., Moore J., Montgomery J., Lippi M. M., Nowell G., and Forlin P. (2021). Combining dental calculus with isotope analysis in the Alps: New evidence from the Roman and medieval cemeteries of Lamon, Italy. Quaternary Int. 653-654, 89–102. doi: 10.1016/j.quaint.2021.11.022
Fuller D. Q., Allaby R. G., and Stevens C. (2010). Domestication as innovation: the entanglement of techniques, technology and chance in the domestication of cereal crops. World Archaeol. 42, 13–28. doi: 10.1080/00438240903429680
García-Puchol O., Bernabeu-Aubán J., Michael Barton C., Pardo-Gordó S., McClure S. B., and Diez-Castillo A. (2018). A bayesian approach for timing the neolithization in mediterranean iberia. Radiocarbon 60, 181–205. doi: 10.1017/RDC.2017.61
González Morales M. R. (2012). “Cantabria,” in El Neolítico En La Península Ibérica y Su Contexto Europeo. Eds. Rojo-Guerra M. A., Garrido Pena R., and García-Martínez de Lagrán I. (Cátedra, Barcelona), 255–290.
González Morales M. R. (2015). La Prehistoria del Valle del Asón: un resumen de 30 años de investigaciones. Sautuola, XX, 53–72.
González Morales M. R., González-Rabanal B., Gutiérrez-Zugasti I., Cuenca-Solana D., and Straus L. G (2024). Chalcolithic/Early Bronze Age and additional Magdalenian radiocarbon dates for El Mirón cave. Date List VII. Radiocarbon, (Spain: Ramales de la Victoria, Cantabria). 65, 1372–1385. doi: 10.1017/RDC.2023.123
González-Rabanal B. (2022). Dinámicas de población, dieta y prácticas funerarias de los últimos cazadores-recolectores y primeras sociedades campesinas de la Región Cantábrica. Santander: Universidad de Cantabria.
González-Rabanal B., Marín-Arroyo A. B., Cristiani E., Zupancich A., and González Morales M. R. (2022). The arrival of millets to the Atlantic coast of northern Iberia. Sci. Rep. 12, 18589. doi: 10.1038/s41598-022-23227-4
González-Rabanal B., Marín-Arroyo A. B., Vidal-Cordasco M., Martín M. Á., and Ortega A. I. (2023). Early Bronze Age violence in Ojo Guareña (Merindad de Sotoscueva, Burgos, Spain). Perimortem modifications in two male individuals. Quat. Sci. Adv. 12, 100120. doi: 10.1016/j.qsa.2023.100120
González-Rabanal B., Marín-Arroyo A. B., Jones J. R., Agudo Pérez L., Vega-Maeso C., and González Morales M. R. (2020). Diet, mobility and death of Late Neolithic and Chalcolithic groups of the Cantabrian Region (northern Spain). A multidisciplinary approach towards studying the Los Avellanos I and II burial caves. J. Archaeol. Sci. Rep. 34, 1–13. doi: 10.1016/j.jasrep.2020.102644
Guilaine J. (2017). “The neolithic transition: from the eastern to the western mediterranean,” in Times of Neolithic Transition along the Western Mediterranean. Eds. García-Puchol O. and Salazar-García D. C. (Springer International Publishing, Cham), 15–32.
Guilaine J. and Manen C. (2007). From mesolithic to early neolithic in the western mediterranean. Proc. Br. Acad. 144, 21–51. doi: 10.5871/bacad/9780197264140.003.0003
Haak W., Balanovsky O., Sanchez J. J., Koshel S., Zaporozhchenko V., Adler C. J., et al. (2010). Ancient DNA from European early neolithic farmers reveals their near eastern affinities. PLoS Biol. 8, e1000536. doi: 10.1371/journal.pbio.1000536
Hardy K., Blakeney T., Copeland L., Kirkham J., Wrangham R., and Collins M. (2009). Starch granules, dental calculus and new perspectives on ancient diet. J. Archaeol. Sci. 36, 248–255. doi: 10.1016/j.jas.2008.09.015
Hardy K., Buckley S., and Copeland L. (2018). Pleistocene dental calculus: Recovering information on Paleolithic food items, medicines, paleoenvironment and microbes. Evol. Anthropol. 27, 234–246. doi: 10.1002/evan.21718
Hardy K., Radini A., Buckley S., Sarig R., Copeland L., Gopher A., et al. (2016). Dental calculus reveals potential respiratory irritants and ingestion of essential plant-based nutrients at Lower Palaeolithic Qesem Cave Israel. Quaternary Int. 398, 129–135. doi: 10.1016/j.quaint.2015.04.033
Henry A. G. and Piperno D. R. (2008). Using plant microfossils from dental calculus to recover human diet: a case study from Tell al-Raqā’i, Syria. J. Archaeol. Sci. 35, 1943–1950. doi: 10.1016/j.jas.2007.12.005
Jesus A., Tereso J. P., and Gaspar R. (2020). Interpretative trajectories towards the understanding of negative features using Terraço das Laranjeiras Bronze Age site as a case study. J. Archaeol. Sci. Rep. 30, 1–14. doi: 10.1016/j.jasrep.2020.102222
Jones J. R., Maeso C. V., Ballestero E. C., Martín L. V., Arceo M. E. D., and Marín-Arroyo A. B. (2019). Investigating prehistoric diet and lifeways of early farmers in central northern Spain, (3000–1500 CAL BC) using stable isotope techniques. Archaeol. Anthropol. Sci. 11, 3979–3994. doi: 10.1007/s12520-019-00841-4
Lieverse A. R. (1999). Diet and the aetiology of dental calculus. Int. J. Osteoarchaeology 9, 219–232. doi: 10.1002/(SICI)1099-1212(199907/08)9:4<219::AID-OA475>3.0.CO;2-V
López-Dóriga I. (2016). The use of plants during the Mesolithic and the Neolithic in the Atlantic coast of the Iberian peninsula. Santander: Universidad de Cantabria.
López-Merino L., Cortizas A. M., and López-Sáez J. A. (2010). Early agriculture and palaeoenvironmental history in the North of the Iberian Peninsula: a multi-proxy analysis of the Monte Areo mire (Asturias, Spain). J. Archaeol. Sci. 37, 1978–1988. doi: 10.1016/j.jas.2010.03.003
Lucarini G., Radini A., Barton H., and Barker G. (2016). The exploitation of wild plants in Neolithic North Africa. Use-wear and residue analysis on ground stone tools from the Farafra Oasis, Egypt. Quaternary Int. 410, 77–92. doi: 10.1016/j.quaint.2015.11.109
Madella M., Lancelotti C., and García-Granero J. J. (2016). Millet microremains—an alternative approach to understand cultivation and use of critical crops in Prehistory. Archaeol. Anthropol. Sci. 8, 17–28. doi: 10.1007/s12520-013-0130-y
Mariotti Lippi M., Pisaneschi L., Sarti L., Lari M., and Moggi-Cecchi J. (2017). Insights into the Copper-Bronze Age diet in Central Italy: Plant microremains in dental calculus from Grotta dello Scoglietto (Southern Tuscany, Italy). J. Archaeol. Sci. Rep. 15, 30–39. doi: 10.1016/j.jasrep.2017.07.005
Martins H., Oms F. X., Pereira L., Pike A. W. G., Rowsell K., and Zilhão J. (2015). Radiocarbon dating the beginning of the Neolithic in Iberia: new results, new problems. J. Mediterr. Archaeology 28, 105–131. doi: 10.1558/jmea.v28i1.27503
Mathieson I., Lazaridis I., Rohland N., Mallick S., Patterson N., Roodenberg S. A., et al. (2015). Genome-wide patterns of selection in 230 ancient Eurasians. Nature 528, 499–503. doi: 10.1038/nature16152
Moreno-Larrazabal A., Teira-Brión A., Sopelana-Salcedo I., Arranz-Otaegui A., and Zapata L. (2015). Ethnobotany of millet cultivation in the north of the Iberian Peninsula. Vegetation History Archaeobotany 24, 541–554. doi: 10.1007/s00334-015-0518-y
Nava A., Fiorin E., Zupancich A., Carra M., Ottoni C., Di Carlo G., et al. (2021). Multipronged dental analyses reveal dietary differences in last foragers and first farmers at Grotta Continenza, central Italy (15,500–7000 BP). Sci. Rep. 11, 1–14. doi: 10.1038/s41598-021-82401-2
Norström E., Gustavsson R., Molin F., and Gummesson S. (2019). Micro-fossil analysis of Mesolithic human dental calculus, Motala, Sweden - Indications of health status and paleo-diet. J. Archaeol. Sci. Rep. 26, 101866. doi: 10.1016/j.jasrep.2019.05.031
Noval M. (2014). Excavación arqueológica en la cueva de El Toral III (Andrín, Llanes). In Excavaciones Arqueológicas en Asturias, 2007-2012. 381–384. Oviedo: Gobierno del Principado de Asturias.
Núñez de la Fuente S. (2018). Dinámicas socio-ecológicas, resiliencia y vulnerabilidad en un paisaje atlántico montañoso: la Región Cantábrica durante el Holoceno. Santander: Universidad de Cantabria.
Olalde I., Mallick S., Patterson N., Rohland N., Villalba-Mouco V., Silva M., et al. (2019). The genomic history of the Iberian Peninsula over the past 8000 years. Science 363, 1230–1234. doi: 10.1126/science.aav4040
Oxilia G., Bortolini E., Badino F., Bernardini F., Gazzoni V., Lugli F., et al. (2021). Exploring late Paleolithic and Mesolithic diet in the Eastern Alpine region of Italy through multiple proxies. Am. J. Phys. Anthropol. 174, 232–253. doi: 10.1002/ajpa.24128
Peña-Chocarro L., Pérez-Jordà G., and Morales J. (2018). Crops of the first farming communities in the Iberian Peninsula. Quaternary Int. 470, 369–382. doi: 10.1016/j.quaint.2017.06.002
Peña-Chocarro L. and Zapata L. (1998). “Hulled wheats in Spain: history of minor cereals,” in Triticeae III. Ed. Jaradat A. A. (Enfield: Oxford & NBH), 45–52.
Peña-Chocarro L., Zapata L., Iriarte M. J., González Morales M., and Straus L. G. (2005). The oldest agriculture in northern Atlantic Spain: new evidence from El Mirón Cave (Ramales de la Victoria, Cantabria). J. Archaeol. Sci. 32, 579–587. doi: 10.1016/j.jas.2004.12.001
Pérez-Jordà G., Peña-Chocarro L., Sabato D., Peralta Gómez A., Ribera A., García Borja P., et al. (2024). The Path of African Millets (Pennisetum glaucum and Sorghum bicolor) to Iberia. Agronomy 14 (10), 2375. doi: 10.3390/agronomy14102375
Pyankov V. I., Ziegler H., Akhani H., Deigele C., and Lüttge U. (2010). European plants with C4 photosynthesis: geographical and taxonomic distribution and relations to climate parameters. Botanical J. Linn. Soc. 163, 283–304. doi: 10.1111/j.1095-8339.2010.01062.x
Radini A., Nikita E., Buckley S., Copeland L., and Hardy K. (2017). Beyond food: The multiple pathways for inclusion of materials into ancient dental calculus. Am. J. Phys. Anthropol. 162 Suppl 63, 71–83. doi: 10.1002/ajpa.23147
Romero A., Basso Rial R. E., Javier Jover Maestre F., Herrero-Otal M., Cuesta-Aguirre D. R., Fiorin E., et al. (2024). Dental evidence of textile-related task activities in the Bronze Age site of Laderas del Castillo (Alicante, Spain). J. Archaeol. Sci. Rep. 60, 104818. doi: 10.1016/j.jasrep.2024.104818
Sabin S. and James A. (2020). Dental Calculus Field-Sampling Protocol (Sabin version) v2 [WWW Document]. (protocols.io.bqecmtaw). doi: 10.17504/protocols.io.bqecmtaw
Salazar-García D. C., Power R. C., Daura J., and Sanz M. (2022). Diet at the onset of the Neolithic in northeastern Iberia: An isotope–plant microremain combined study from Cova Bonica (Vallirana, Catalonia). Front. Earth Sci. 10, 957344. doi: 10.3389/feart.2022.957344
Sarasketa-Gartzia I., Villalba-Mouco V., Le Roux P., Arrizabalaga Á., and Salazar-García D. C. (2018). Late Neolithic-Chalcolithic socio-economical dynamics in northern Iberia. A multi-isotope study on diet and provenance from Santimamiñe and Pico Ramos archaeological sites (Basque Country, Spain). Quaternary Int. 481, 14–27. doi: 10.1016/j.quaint.2017.05.049
Seabra L., Teira-Brión A., López-Dóriga I., Martín-Seijo M., Almeida R., and Tereso J. P. (2023). The introduction and spread of rye (Secale cereale) in the Iberian Peninsula. PLoS One 18, e0284222. doi: 10.1371/journal.pone.0284222
Teira-Brión A. (2022). “Traditional millet cultivation in the Iberian Peninsula: Ethnoarchaeological reflections through the lens of social relations and economic concerns,” in Millet and What Else?: The Wider Context of the Adoption of Millet Cultivation in Europe, Scales of Transformation in Prehistoric and Archaic Societies. Eds. Kirleis W., Dal Corso M., and Filipović D. (Sidestone Press, Leiden), 263–278.
Tereso J. P., Bettencourt A. M. S., Ramil-Rego P., Teira-Brión A., López-Dóriga I., Lima A., et al. (2016). Agriculture in NW Iberia during the Bronze Age: A review of archaeobotanical data. J. Archaeol. Sci. Rep. 10, 44–58. doi: 10.1016/j.jasrep.2016.07.011
Uzquiano P. (1995). L’évolution de la végétation à l’Holocène initial dans le nord de l’Espagne à partir de l’étude anthracologique de trois sites archéologiques. Quaternaire 6, 77–83. doi: 10.3406/quate.1995.2040
Vega Maeso C. (2017). La cerámica inciso-impresa en el tránsito del III al II milenio cal. B.C. en la región Cantábrica. Nadir Ediciones, Santander.
Villalba-Mouco V., van de Loosdrecht M. S., Posth C., Mora R., Martínez-Moreno J., Rojo-Guerra M., et al. (2019). Survival of late pleistocene hunter-gatherer ancestry in the Iberian Peninsula. Curr. Biol. 29, 1169–1177. doi: 10.1016/j.cub.2019.02.006
Warinner C. (2016). Dental calculus and the evolution of the human oral microbiome. J. - California Dental Assoc. 44, 411–420. doi: 10.1080/19424396.2016.12221034
Yang X., Zhang J., Perry L., Ma Z., Wan Z., Li M., et al. (2012). From the modern to the archaeological: starch grains from millets and their wild relatives in China. J. Archaeol. Sci. 39, 247–254. doi: 10.1016/j.jas.2011.09.001
Zapata L. (2000). La recolección de plantas silvestres en la subsistencia mesolítica y neolítica: datos arqueobotánicos del País Vasco. Complutum 11, 157–170.
Zapata L. (2002). La explotación de los recursos vegetales y el origen de la agricultura en el País Vasco. Vitoria: Universidad del País Vasco.
Zapata L., Milner N., and Rosello E. (2007). “Pico Ramos cave shell midden: the Mesolithic-Neolithic transition by the Bay of Biscay,” in Shell Middens in Atlantic Europe. Eds. Milner N., Craig O. E., and Bailey G. N. (Oxford: Oxbow Books), 150–157.
Zapata L. and Pena-Chocarro L. (2005). L’agriculture néolithique de la façade atlantique européenne. Bulletin de la Société Préhistorique Française Memoire XXXVI: 189–199.
Keywords: farming, Cantabrian region, dental calculus, starch grains, late prehistory
Citation: González-Rabanal B, Marín-Arroyo AB, Carmona-Ballestero E, Cuenca-Solana D, Gutiérrez-Zugasti I, Martín-Merino MÁ, Ortega-Martínez AI, Straus LG, Vega-Maeso C, González Morales MR and Cristiani E (2025) New insights into prehistoric agriculture of northern Iberia from the analysis of starch grains embedded in dental calculus. Front. Ecol. Evol. 13:1644052. doi: 10.3389/fevo.2025.1644052
Received: 09 June 2025; Accepted: 10 September 2025;
Published: 26 September 2025.
Edited by:
Antonella Pedergnana, University of Zurich, SwitzerlandReviewed by:
Robert C. Power, University College Dublin, IrelandAngelo Gismondi, University of Rome Tor Vergata, Italy
Fiona Pichon, Agencia Estatal Consejo Superior de Investigaciones Cientificas, Spain
Copyright © 2025 González-Rabanal, Marín-Arroyo, Carmona-Ballestero, Cuenca-Solana, Gutiérrez-Zugasti, Martín-Merino, Ortega-Martínez, Straus, Vega-Maeso, González Morales and Cristiani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Borja González-Rabanal, Ym9yamEuZ29uemFsZXpAdXZhLmVz; Ana B. Marín-Arroyo, YW5hYmVsZW4ubWFyaW5AdW5pY2FuLmVz; Emanuela Cristiani, ZW1hbnVlbGEuY3Jpc3RpYW5pQHVuaXJvbWExLml0