 Hugues Terreaux de Félice
Hugues Terreaux de Félice Marc-Antoine Leclerc
Marc-Antoine Leclerc Hubert Morin
Hubert Morin- Department of Fundamental Sciences, Université du Québec à Chicoutimi, Saguenay, QC, Canada
Understanding the historical variability in the disturbance dynamics of the most important defoliator in eastern Canada, the spruce budworm (SBW), throughout its range will be paramount in anticipating potential changes associated with current climate change. There is growing concerns that black spruce-dominated forests may become increasingly affected by more frequent and severe outbreaks with warmer spring temperatures. Therefore, in this study, we used lepidopteran scales, a novel paleo-proxy found in lake sediments, to reconstruct periods of large spruce budworm populations (LSBP) along a latitudinal gradient and examined the relationship between large population event frequency relative to geographic location over the course of the Holocene in eastern Canada. The synchronous increase in LSBP event frequency in the four lakes observed around 5500 cal. yr BP culminating in maximum event frequency likely resulted from a combination of the northward migration of balsam fir, and low fire frequency. Although favorable Holocene climate conditions appear to have allowed for the expansion of the spruce budworm’s range into more northerly black spruce-dominated stands, event frequency remained low suggesting that the most suitable area for the SBW appears to be in more southern balsamfir-dominated stands.
1 Introduction
In Canada’s boreal and mixed-wood forests, fire and insect outbreaks are the two major natural disturbances (Blais, 1965; Morin and Laprise, 1990; Fleming, 2000). In eastern North America, the spruce budworm (Choristoneura fumiferana (Clem.)) (SBW) is the major defoliating insect undergoing episodic outbreaks causing significant growth reduction and mortality (Blais, 1958, 1981; Morin, 1994; Krause and Morin, 1995), in its primary host balsam fir (Abies balsamea (L.) Mill.) (MacLean, 1980, 1988; Bergeron et al., 1995). Balsam fir has historically been the spruce budworm’s preferred host tree because of the synchronous timing of bud burst and larval emergence (Blais, 1957; Lawrence et al., 1997; Nealis and Régnière, 2004; Fuentealba et al., 2017), along with more nutritious and softer needles (Mattson et al., 1983; Fuentealba and Bauce, 2012; Fuentealba et al., 2018), allowing for rapid population growth and maintaining very large populations via accelerated nutrient cycling (De Grandpré et al., 2022). Conversely, in secondary hosts such as black (Picea mariana (Mill.)), white (Picea glauca (Moench) Voss) and red (Picea rubens (Sarg.)) spruce, defoliation and mortality is less important (Hennigar et al., 2008), but large growth reductions are sometimes observed (MacLean, 1984; Nealis and Régnière, 2004). As such, areas of severe defoliation predominantly occur within balsam fir’s distribution and in proximity to balsam fir-dominated stands (Blais, 1981; Jardon et al., 2003; Hennigar et al., 2008). Over the past two centuries, major defoliation events have been episodic, and appear to occur every 30 to 40 years at the landscape-scale (Royama, 1984; Krause, 1997; Morin, 1998; Boulanger and Arseneault, 2004).
Although the SBW distribution overlaps with the distribution of black spruce, climate in recent history has limited the insect’s proliferation beyond the 50th parallel in Québec resulting in outbreaks that are less severe and frequent (Pureswaran et al., 2015), but the question remains whether these dynamics were also valid during the warmer periods of the Holocene. Natural disturbances, such as insect defoliators, are among the most climate-sensitive processes, and current climate change is expected to alter the extent, intensity, frequency, and severity of these disturbances (Gray, 2008; Lindner et al., 2010; Seidl et al., 2017, 2020). Recent work on the effect of warming suggests that secondary hosts, such as black spruce, may become more vulnerable with increasing temperature (Rossi et al., 2015; Pureswaran et al., 2018, 2019; Bellemin-Noël et al., 2021) potentially explaining the greater defoliation observed in black spruce stands during the current outbreak (Bognounou et al., 2017). Such a change in vulnerability would likely alter the dynamics of the black spruce forest, and the ability to observe and reconstruct past long-term variability in spruce budworm disturbance would be of great help in anticipating the future dynamics of the boreal forest ecosystem. Although paleoecological analyses cannot directly test the effects of non-mutually exclusive climate-mediated mechanisms, such as insect survival and development (Régnière et al., 2012; Pureswaran et al., 2015), and/or synchrony between black spruce bud burst and larval emergence (Nealis and Régnière, 2004; Antonucci et al., 2015; Fuentealba et al., 2017; Régnière and Nealis, 2018), these analyses allow for observance of long-term system behavior and trends. The objective of this study was to use lepidopteran scales, a novel spruce budworm paleo-proxy (Navarro et al., 2018a), that is easily identifiable, more abundant, and well-preserved in lake sediment (Montoro Girona et al., 2018; Navarro et al., 2018a) relative to larval head capsules, and frass (Simard et al., 2002, 2006), to reconstruct long-term variability of large spruce budworm population (LSBP) event frequencies during the Holocene within the insect’s current range (balsam fir-white birch domain) and at its northern limit (spruce-moss domain). If warm and humid periods were favorable to LBSP events in northern regions, then we expect greater SBW impacts in black spruce-dominated forests as registered by more frequent LSBP events because these favorable conditions would have enabled the spruce budworm to experience an expansion in host breadth by taking advantage of black spruce a once less preferred secondary host.
2 Materials and methods
2.1 Site description, selection and sediment coring
The four sampled lakes were located either in the balsam fir-white birch or the black spruce-moss bioclimatic domains (Saucier et al., 2009; Table 1). The earlier domain contains a diversity of tree species, generally composed of balsam fir, white spruce, white birch, and trembling aspen however, poorer sites tend to be dominated by black spruce, meanwhile, the latter domain is generally dominated by black spruce with the presence of jack pine, and balsam fir (Saucier et al., 2009; Table 1). The more southern balsam fir-white birch domain is typically warmer and wetter with a longer growing season relative to the more northern black spruce-moss domain (Saucier et al., 2009; Table 1). In both domains, there is an east-west precipitation gradient with the west being drier than the east resulting in shorter and longer fire return intervals respectively (Saucier et al., 2009). In addition to characterizing current site-level tree species composition (Table 1), long-term changes in forest composition were characterized using the closest available pollen profiles to our lakes from the Neotoma database (https://www.neotomadb.org; Table 1, Supplementary Figures S1-S3), along with accessing Holocene August summer temperature data based on chironomid reconstructions from Bajolle et al. (2018) made available in the NOAA Paleoclimatology database (NCEI; https://www.ncei.noaa.gov/access/paleo-search/study/27330; Kaufman et al., 2020). The long-term changes in forest composition and temperature were used to provide context for any observed changes in LSBP event frequency.

Table 1. Site-level characteristics of the cored lakes (from north to south) and associated regional climate.
The four lakes sampled in this study were selected due to their limited inflow and outflow, and a deep water column to avoid sediment mixing, and cored at each lake’s maximum depth (Table 1). The core of lake Flévy was collected in 2012 while lake Trouble and Totem were sampled in the summer of 2018, and lake Innu in the summer of 2019. The sediment surface was collected using a gravity corer (Renberg, 1991; Renberg and Hansson, 2008) and sampled at a 1 cm resolution (0.5 cm for lake Totem) in the field. The remainder of the sediment column was sampled using a Livingstone piston corer (Wright et al., 1984) from each lake’s deepest point and subsampled at a 1 cm resolution and stored in individual plastic bags to avoid desiccation. The lake Flévy core had already been analyzed in a previous study (Navarro et al., 2018b). The major difference in methodology between Navarro et al. (2018b) and the current study was the amount of sediment assessed at each depth with the earlier study using 5 cm3 as opposed to the 1 cm3 used in this study. This difference would not alter the observed trends but could possibly result in higher LSBP event frequencies detected at lake Flévy relative to the lakes analyzed in this study. All analyses in this section are for lakes Innu, Totem and Trouble. The lakes Innu and Totem are located in the spruce-moss domain while lakes Trouble and Flévy are found in the balsam fir-white birch domain (Figure 1).
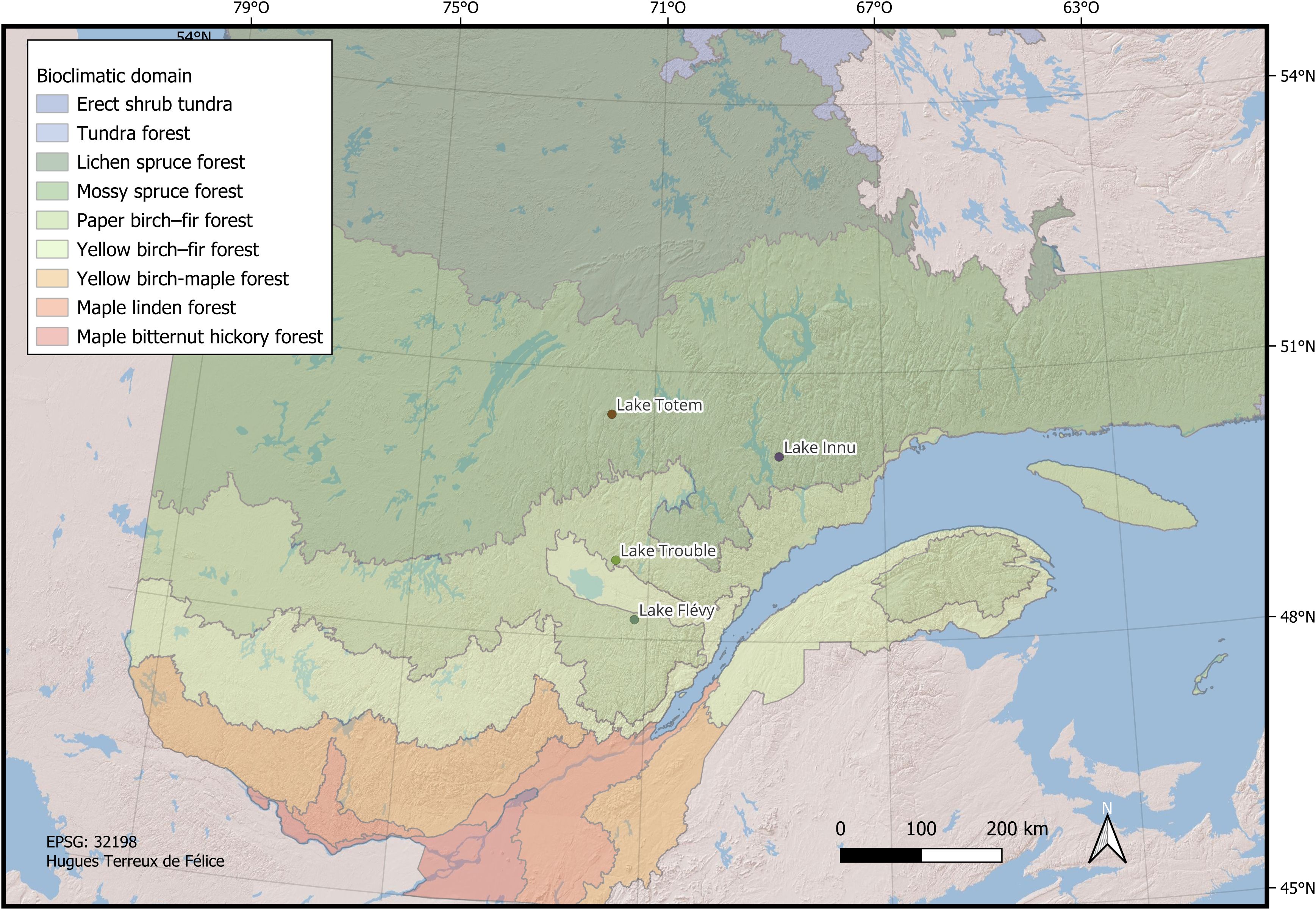
Figure 1. Location of the study sites within the bioclimatic domains of Québec (Canada).
2.2 Lead-210, and radiocarbon dating and sample preparation
The age-depth models for lakes Innu, Trouble, Totem were derived from a combination of Lead-210 (210Pb) to reconstruct recent history (approximately last 150 years), and deep history using radiocarbon (14C) dates (Table 2). Recent history was derived from bulk sediment samples that were sent and analyzed for 210Pb activity at Flett Research Ltd (Winnepeg, MN, Canada). A more detailed methodology is provided in Supplementary Material section 1.1, but in short, a dry mass was obtained from samples that were dried, homogenized, and then digested in nitric acid to help isolate the Polonium-210 (210Po) which provides an indirect measurement of the total 210Pb present in the sample (Eakins and Morrison, 1978). Also, samples were spiked with a tracer to measure and identify any potential losses (Eakins and Morrison, 1978), and the 210Po was isolated using spontaneous auto-deposition on silver planchets (Flynn, 1968). Supported 210Pb was derived from Radon-226 (226Ra; Mathieu et al., 1988) from which excess lead-210 could be calculated and Constant Rate of Supply models (Appleby and Oldfield, 1978; Appleby and Oldfieldz, 1983) could be applied to estimate the ages of the sediment in recent history.
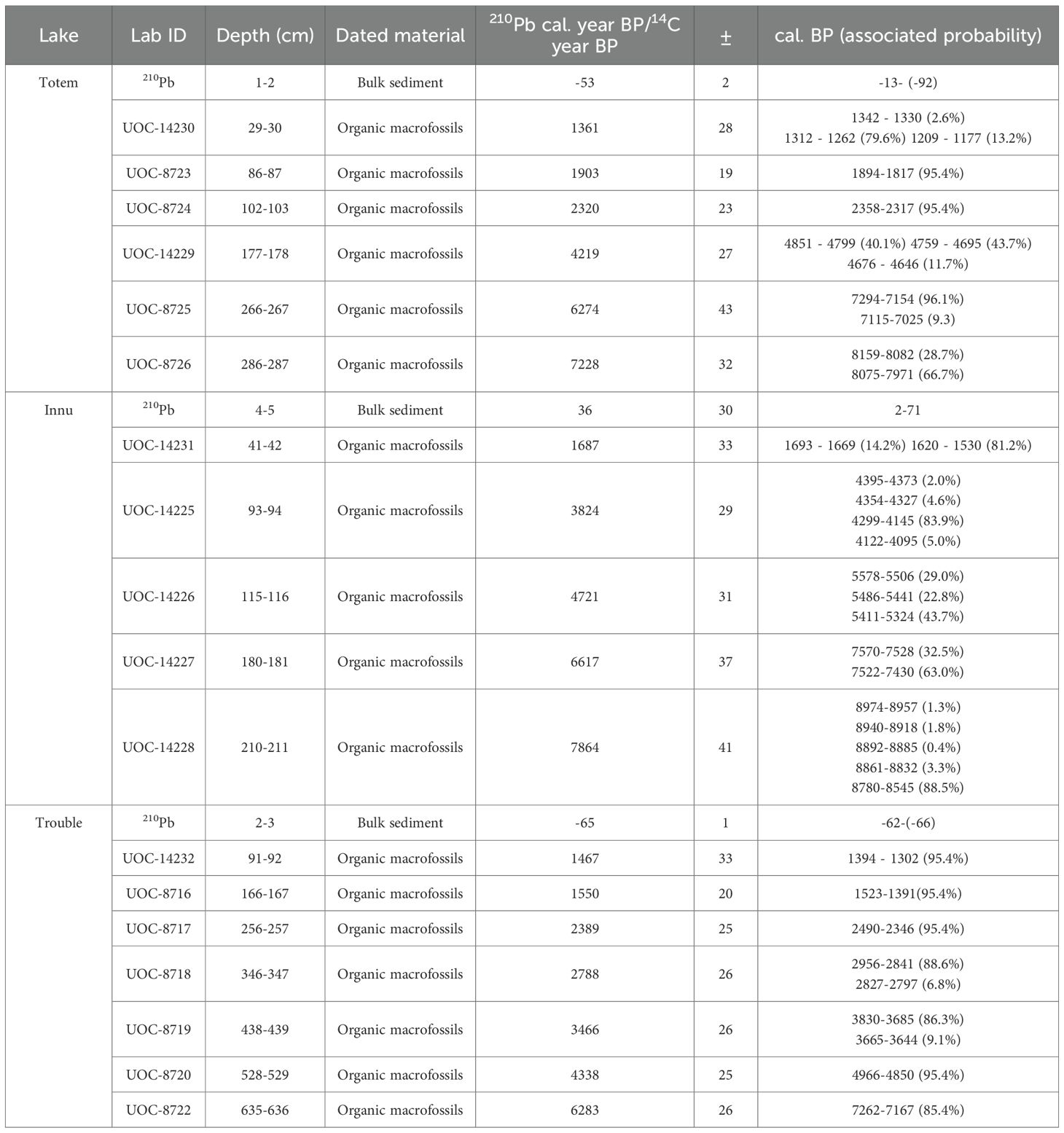
Table 2. Sampling depth, material, median (± standard deviation) calibrated ages of lakes Totem, Innu, and Trouble.
Deep history was reconstructed using radiocarbon dates derived from terrestrial plant macrofossils (leaves, needles, and seeds) sampled along the sediment cores, that were sent and analyzed at the André E. Lalonde AMS Laboratory at the University of Ottawa (Table 2). Briefly, macrofossils underwent pretreatment, combustion, graphitization, followed by accelerator mass spectrometry measurement (Crann et al., 2017; St-Jean et al., 2017). Plant macrofossils were inspected for contaminants, and sonicated in Milli-Q water when needed, underwent an acid-alkali-acid wash, and freeze-dried overnight (Brock et al., 2010; Crann et al., 2017). Samples were then combusted in a Thermoflash 1112 Elemental Analyzer in CN mode to trap pure carbon dioxide (CO2; Crann et al., 2017). The pure CO2 was converted to elemental carbon in semi-automated graphitization lines in the presence of iron and hydrogen then pressed into targets with a pneumatic press (Crann et al., 2017; St-Jean et al., 2017). Accelerator Mass Spectrometry measurements were done using a 3 MV Tandem Accelerator Mass Spectrometer built by High Voltage Engineering (The Netherlands; Kieser et al., 2015). A terminal voltage of 2.5 MV with Ar stripping was used to measure the 14C3+ ions (Crann et al., 2017). The fraction of modern carbon (F14C) was calculated following Reimer et al. (2004) by using the ratio of the sample 14C/12C to the standard 14C/12C in the same data block (Crann et al., 2017). Noting that all ratios were background corrected using corresponding process, oxalic acid-II, iron-only blanks along with secondary references (Crann et al., 2017). Results were also corrected for spectrometer and preparation and normalized to δ13C and finally 14C ages were calculated following Stuiver and Polach (1977). Further procedural and instrument details are provided in Supplementary Material section 1.2.
In the R software environment (R core team, 2020), Bayesian age-depth models for lake Innu, Trouble, and Totem were obtained using the IntCal20 calibration curve (Reimer et al., 2020) and the package Rbacon 2.5.8 with default settings (Blaauw et al., 2022) as these settings best reflected the sediment accumulation rate of the cores (see Results). The age-depth model for lake Flévy was derived by reanalyzing the radiocarbon dates in Navarro et al. (2018b) using Rbacon 2.5.8 and the IntCal20 calibration curve (Reimer et al., 2020; Blaauw et al., 2022). Broadly, this age-depth modelling package subdivided the respective sediment cores into sections of equal thickness within which linear accumulation rates, constrained by the provided 210Pb and radiocarbon dates, were estimated using Markov Chain Monte Carlo iterations (Blaauw et al., 2022).
For scale extraction we used a punch to collect exactly 1 cm3 of wet sediment and placed it in a 30 milliliter (mL) vial. Twenty mL of 20% KOH solution was added and the sample was placed in an autoclave (Amsco unit by Steris) at 121°C, 17 psi for 30 minutes (Leclerc et al., 2024) for complete deflocculation (Frey and Pamini, 1986). The slurry was then sieved through a mesh of 53 µm to retain most of the scales (Navarro et al., 2018a). Fifteen mL of sugary solution at 1.24 g/L was added to further separate sediment particles and isolate the scales, and then centrifuged at 4500 RPM for 20 minutes to concentrate sediment and reduce sample volume. The supernatant was removed with a Pasteur pipet and samples were stored at 4°C. The entire pellet of each sample was analyzed under a microscope (Leica DM 4000 B) with a digital camera (Leica DFC 425) at 100x magnification, and all scales were photographed.
2.3 Lepidopteran scale peak identification
Identification of scale peaks was done with CharAnalysis software (https://github.com/phiguera/CharAnalysis) (Higuera et al., 2009). This software was originally developed to identify, and extract charcoal peak accumulations that correspond to wildfire events, and the same methodology was applied to determine peak lepidopteran scale accumulations (Navarro et al., 2018a, b). These detected peak scale accumulations strongly agree with commonly used, reliable, and modern spruce budworm proxies (Leclerc et al., 2024, 2025) and are therefore assumed to accurately reflect spruce budworm populations around a lake. Indeed, a LSBP event would be related to an increase and a peak in the number of scales found in the sediment, noting however, that peak scale accumulations may encompass one or more outbreaks, and are interpreted as periods in time where SBW populations were elevated, and not as a count of individual outbreak events. To distinguish large population events from background scale accumulation rates, we obtained a constant time-step by interpolating each lake’s accumulations using its corresponding median temporal resolution (28, 40, 10, and 5 years for lakes Totem, Innu, Trouble, and Flévy respectively): Cint, and defined two components: Cback and Cpeak. The Cback component was identified using a LOWESS smoother robust to outliers in conjunction with the smallest possible smoothing window that yielded a Signal-to-Noise ratio (SNI) above 3. Cpeak was obtained by subtracting Cback from Cint. Further, a Gaussian mixture model was used to discriminate LSBPs within Cpeak from noise resulting from sediment mixing (Gavin et al., 2006; Higuera et al., 2010). We did not conduct the minimum count test; the final step used in CharAnalysis to avoid missing any LSBP events. Normally after Cpeak values are identified as being above a threshold value defined by the Gaussian mixture model, these potential LSBP events undergo the minimum count criterion test. This test determines if two detected events were generated from a single event by determining whether the two events belong to the same Poisson distribution and comparing the accumulation of scales in the event relative to the previous 75 years. Only peaks with a probability ∝<0.05 are considered LSBP events. However, during periods of large spruce budworm populations, the SBW numbers gradually increase to a maximum and then decreases, likely translating into gradual accumulations of scales in the sediment and therefore would more likely resemble a Normal and not a Poisson distribution.
2.4 Large spruce budworm population event frequency
LSBP event frequency was calculated from kernel rate occurrence estimation (Blarquez et al., 2014). This is a powerful approach because it is flexible, supports nonlinear trends, and produces a reliable statistical error (Mudelsee et al., 2004). The years of LSBP events identified by the Gaussian mixture model were used to run a bootstrap resampling analysis. LSBP event frequency was based on each lake’s median time-step and then estimated using a bandwidth of 1000 years and resampled 1000 times to ensure statistical robustness. To prevent over-representation of rare events, the bootstrap resampling was constrained to select between 1 and 10% of the total number of events, thereby reducing the likelihood of generating outlier reconstructions where a single event could be sampled multiple times (Blarquez et al., 2013). Finally, to compare LSBP event frequencies between northern (Totem and Innu) and southern (Flévy and Trouble) lakes we used an orthogonal contrast matrix coupled with an ANOVA. The event frequency values were logistically transformed to respect the assumption of normality. Three contrast pairs were tested: 1) southern vs. northern lakes, 2) Flévy vs. Trouble and 3) Innu vs. Totem.
3 Results
Innu and Totem lake cores were composed of homogenous gyttja, and relatively short, 202 and 283 cm respectively, likely explaining the particularly high sediment accumulation rates expressed in years/cm (Innu: mean = 41.63, median = 40, max = 57, min = 21; Totem: mean = 26.72, median = 28, max = 59, min = 6). Lake Trouble’s core was longer with a relative constant sediment accumulation rate (mean = 11.08, median = 10, max = 21, min = 3), and was composed of homogeneous gyttja until 5700 cal. yr BP (570 cm depth), where a large amount of sand was present limiting LSBP event data analysis beyond this point as microscopic observation of the scales was impossible. Finally, lake Flévy’s core was also long with a relatively constant sediment accumulation rate (mean = 19.02, median = 19, max = 42, min = 6) (Figure 2) and was composed of homogeneous gyttja (Navarro et al., 2018b). Markov Chain Monte Carlo simulations for all cores resemble white noise, and the posteriors for sediment accumulation rates, along with the variability in sediment accumulation rates overlap with the priors (Figure 2). Further, the age-depth models do not exhibit any strange elbows and generally resemble smooth curves (Figure 2). A total of 33 peaks were identified at Totem, 25 peaks at Innu, 43 at Trouble, and 93 at Flévy (Figures 3, 4).
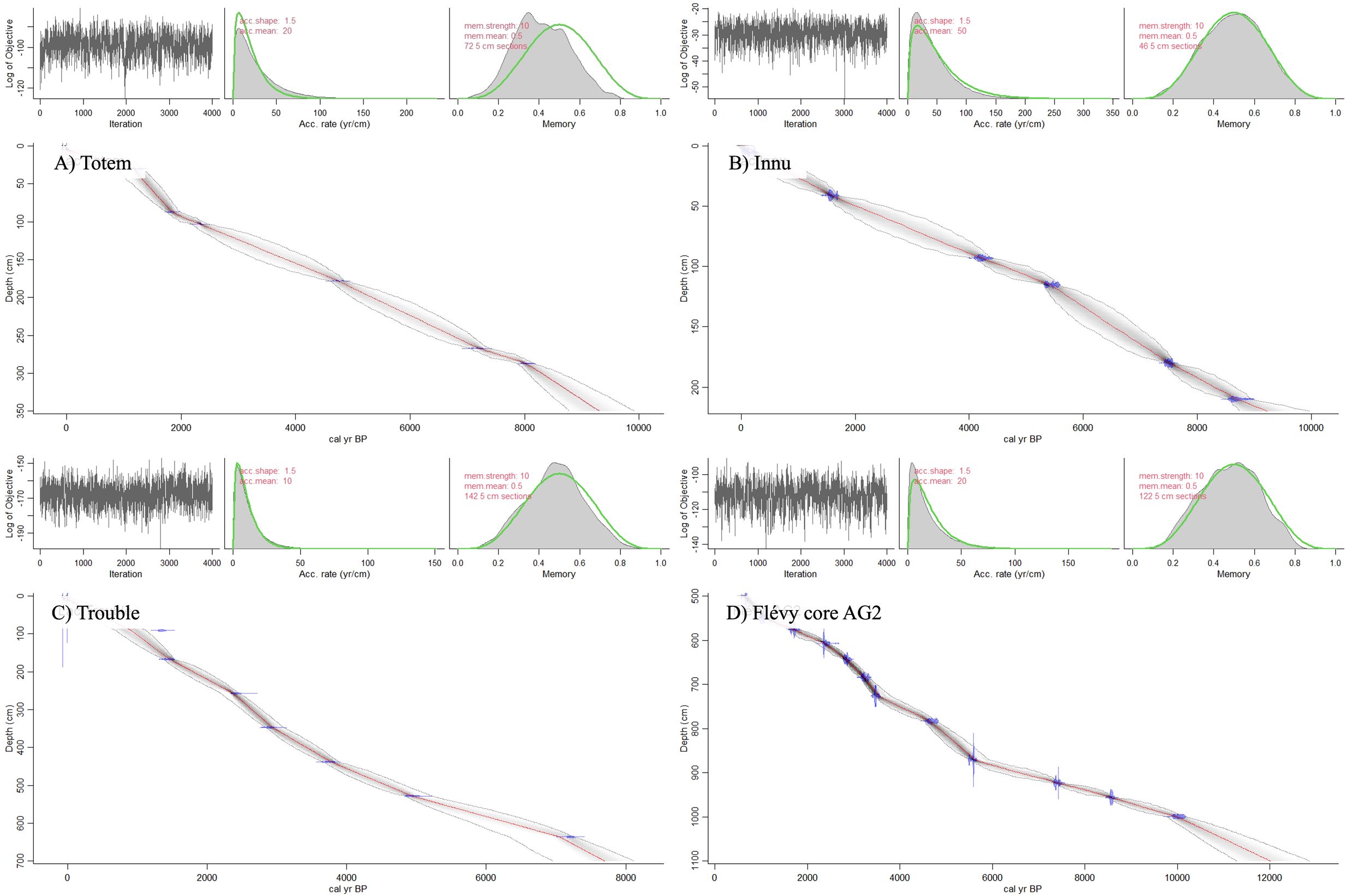
Figure 2. Lake sediment core age-depth models for lakes (A) Totem (B) Innu (C) Trouble (D) Flévy core AG2. Main panel: Red dashed line and dashed grey curves represent the mean model with 95% confidence interval, with calibrated 14C dates are shown in blue. Upper left: Markov Chain Monte Carlo simulations estimating accumulation rates of each core section. Upper center: Modelled accumulation rates compared to a gamma distribution (green line). Upper right: Variation in sediment accumulation through time (Memory) relative to a beta distribution (green line).

Figure 3. Large spruce budworm population event frequency (smooth curves) and associated uncertainty (shading) for each lake. Bars depict the retained lepidopteran scale peaks (Cfire) along with their respective magnitudes.
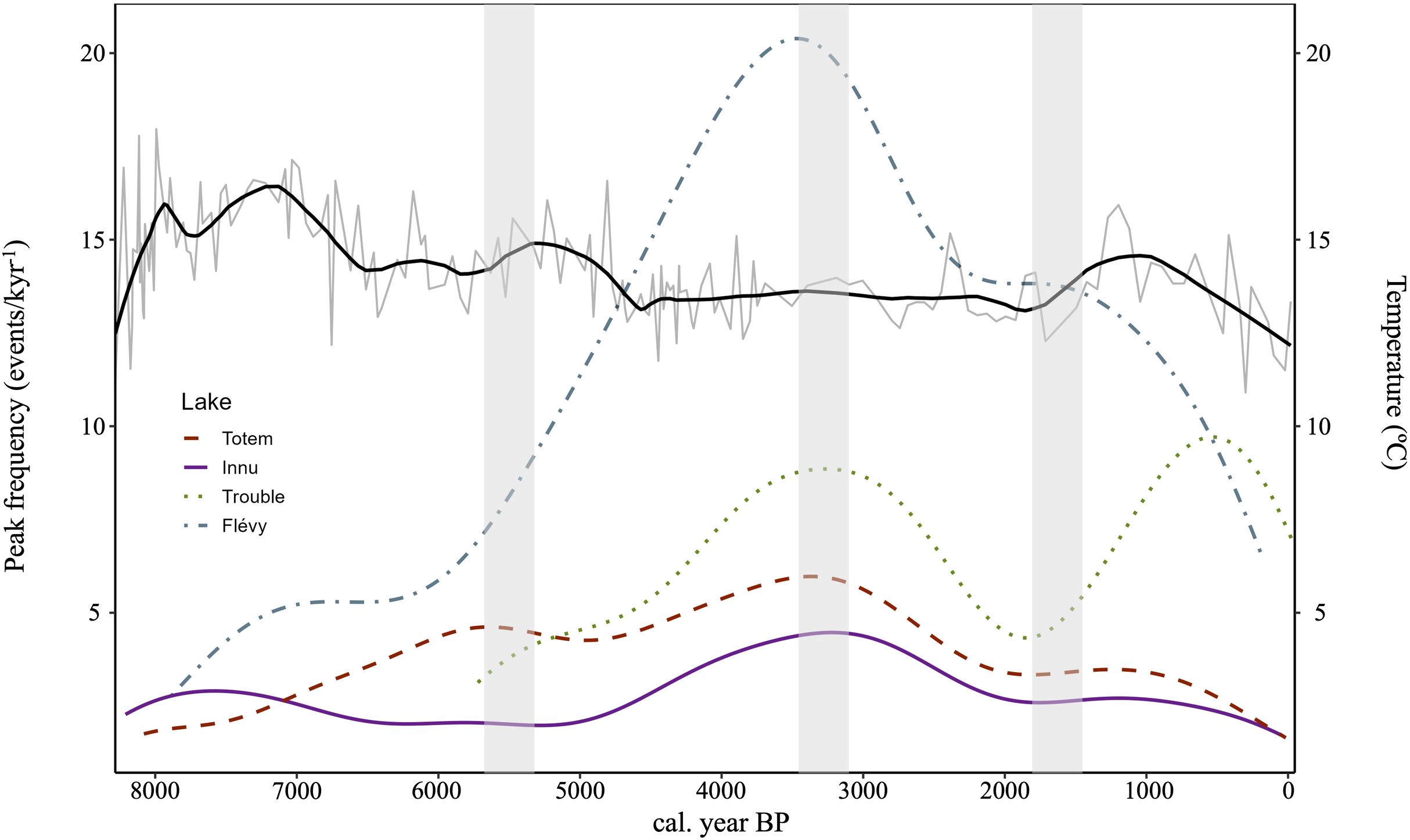
Figure 4. Large spruce budworm population event frequencies for the lakes after Kernel density bootstrap and chironomid-reconstructed August summer (grey) and smoothed temperatures (black). Vertical grey bands highlight the approximate timing of synchronous change in event frequency at 5500, 3100, and 1900 cal. year BP.
Variability in outbreak frequency was less important in northern lakes relative to southern lakes. Maximum peak frequency for lake Innu and Totem were respectively 4.471 and 5.974 kyr−1 and 9.712 and 20.391 kyr−1 for lake Trouble and Flévy (Figure 4). Large spruce budworm population event frequencies were somewhat synchronous prior to 5500 cal. yr BP, however, synchrony is most clearly observed and pronounced between lakes from 5500 cal. yr BP to the present. An increase and then a maximum was reached for all lakes around 3100 cal. yr BP followed by a particularly low period around 1900 cal. yr BP. Further, all lakes, except Innu, also showed a decrease in LSBP event frequency close to the present. The ANOVA indicated that means in event frequency differed between lakes (F(3,1677) = 1065.6, p≤ 0.001) (Figure 5). Specifically, event frequencies in the northern lakes differed from those in the southern lakes (F(1,1677) = 2726.8, p ≤ 0.001), between lakes Trouble and Flévy (F(1,1677) = 330.5, p≤ 0.001), and between lakes Innu and Totem (F(1,1677) = 139.7, p ≤ 0.001).
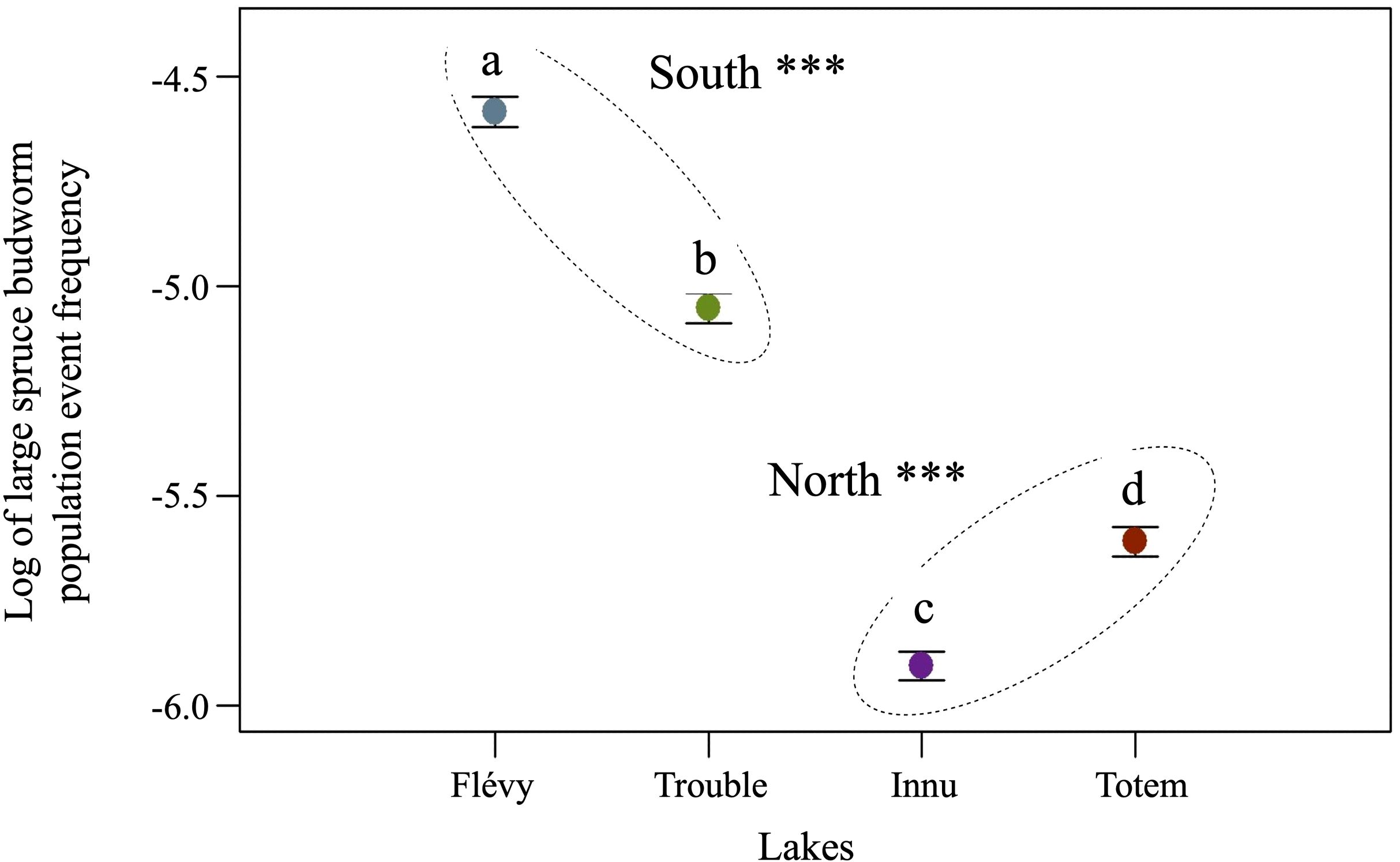
Figure 5. Orthogonal matrix contrast of mean large spruce budworm population event frequency after log transformation and ANOVA. Significant differences between localities are indicated by dashed circles with stars and significant differences between lakes are indicated by differing letters using a threshold of 0.05.
4 Discussion
4.1 Attribution of scale peaks to SBW
The regional consistency after 5500 cal. yr BP. suggests that the scales found in the sediment are mostly attributable to the SBW. The sampled sites are located in continental Québec, and in the boreal forest composed predominantly of coniferous tree species either at the core or at the northern periphery of the spruce budworm’s distribution (Pureswaran et al., 2015; Nealis, 2016). Moreover, historical spruce budworm outbreak epidemiology suggests that it is the only North American insect that can outbreak and simultaneously infest such large areas over several years (Morin and Laprise, 1990; Morin, 1998; Jardon, 2001; Jardon et al., 2003). Since 1975, 95% of the area defoliated in Québec has been affected by SBW, and among identified peak accumulations in the sediment using scale morphological features, more than 60% belonged to the SBW (Navarro et al., 2018a, b). Further, multi-proxy comparisons support the use of lepidopteran scales to reconstruct periods of large spruce budworm populations (Leclerc et al., 2024, 2025). It is therefore less likely that scales found in the sediment belong to other moths such as the hemlock looper (Lambdina fiscellaria (Guénée)), which tends to affect coastal areas, and have very patchy outbreaks that last only 1–2 years (Otvos et al., 1971, 1979) or the forest tent caterpillar (Malacosoma disstria Hübner) which outbreaks mainly in deciduous-dominated forests (Cooke and Lorenzetti, 2006; Cooke et al., 2012). Nevertheless, it is possible that these other species may contribute to the total of analyzed scales as scales from different species are likely equally-well preserved in the sediment (Shorohova et al., 2011; Navarro et al., 2018a), and broken and hidden scales along with somewhat similar scale morphology may lead to identification errors (Navarro et al., 2018a). However, in this study, the location of the lakes, the surrounding forest composition, past spruce budworm epidemiology, scale morphology, and multi-proxy comparisons strongly suggests that the analyzed scales do belong to the SBW.
The low LSBP event frequency observed in the northern lakes may reflect the low host-tree availability limiting spruce budworm population build-ups, and their extent relative to the southern lakes prior to 7000 cal yr. BP (Mott, 1976; Garralla, 1991; Garralla and Gajewski, 1992; Figures 3, 4; Supplementary Figures S1-S3). Lakes Innu and Totem were likely surrounded by tundra dominated by grasses and shrubs with patches of spruce, larch, and poplar (Saarnisto, 1974; Mcandrews, 1988; Heide, 1984; MacDonald et al., 1991) as the postglacial recolonization was slow during this period (Richard, 1973, 1975; Richard et al., 1992; Lavoie and Filion, 2001). Nevertheless, LSBP events would have likely occurred in spruce patches (Garralla, 1991).
4.2 Mid-Holocene increase and host availability
The warmest period of the Holocene (6000 to 3000 cal yr. BP) was characterized by a synchronous increase in LSBP event frequency (Figure 4) likely linked to the post-glacial recolonization by balsam fir (King, 1986; Remy et al., 2017) around 6000–5500 cal yr. BP as demonstrated by an increase in the proportion of the primary host in the pollen profiles (Supplementary Figures S1-S3). After the fragmentation of the Laurentian Ice Cap and deglaciation of the Hudson Bay (Dyke, 2005), the beginning of the warmest period of the Holocene generally exhibited low event frequencies despite mild climate and the boreal forest being continuous to 52°N (Viau and Gajewski, 2009). However, later on, the relatively warm, and humid mid-Holocene period likely resulted in a low fire frequency (Edwards et al., 1996; Ali et al., 2009; Blarquez et al., 2015). The low fire frequency during this period likely allowed late-successional species such as balsam fir to increase in density (Bergeron and Dubue, 1988, Supplementary Figure S1-S3). The greater density in primary host-tree species in conjunction with fewer fires in the northern parts of the SBW range would increase connectivity between trees and favor the establishment of periods of large populations over larger areas (James et al., 2011; Harper et al., 2002).
The combined effect of low fire frequency and increased host-tree density likely explains the maximum LSBP event frequency reached by all lakes around 3100 cal. yr BP (Figures 3, 4). The recurrent presence of the budworm in a stand is assumed to be dependent on temperature but especially on high balsam fir density (Morin et al., 2021). From around 5500 cal. yr BP, the scale signal appears to reflect regional variation (Figure 4). We believe that warm and humid periods would be most favorable to high LSBP event frequencies and the mid-Holocene period of the studied chronologies support this prediction via the possible mechanism of low fire frequencies. Synchronous variations in event frequency from 5500 cal. yr BP in several lakes located in different ecosystems is suggestive of the regional influence of large SBW populations along with the importance of the presence of balsam fir on the landscape.
4.3 Northward migration of the SBW
Recent studies on budworm phenology have shown that a reduction in asynchrony or an increase in synchrony between larval hatching and bud break of its secondary hosts (black, white spruce) and/or favorable larval development induced by climate could allow the insect to move north towards the edge of its historical range (Pureswaran et al., 2015; De Grandpré et al., 2019). If this is the case, black spruce stands at the edge of the spruce budworm’s range are expected to undergo greater defoliation, and should record more frequent LSBP events when conditions are favorable. However, in this study, LSBP event frequency over the course of the Holocene differed significantly between northern and southern lakes, and further, frequencies in the north never exceeded those in the south (except lake totem around 6000 cal. yr BP; Figure 5). Interestingly, a significant difference in LSBP event frequency between all lakes was detected. The difference between the southern lakes may be due to site-level productivity due to a slightly dissimilar current species composition, despite a similar past composition (Supplementary Figure S3). More likely however, is the influence of differences in lake basin shape (Lehman, 1975; Bennett and Buck, 2016) along with surrounding topography, and catchment basin size (e.g., Smith et al., 2010). It is also possible that the difference observed may stem from the fact that scale accumulations were not standardized between the lakes sampled in this study, and Navarro et al. (2018b). The difference observed between the northern lakes, Innu and Totem, is somewhat surprising given similar current (Table 1) and past vegetation composition (Richard et al., 2020; Fréchette et al., 2018, 2021, Supplementary Figures S1, S2). This difference may also most likely stem from site-level variability particularly lake basin shape (Lehman, 1975; Bennett and Buck, 2016) along with surrounding topography, and catchment basin size (e.g., Smith et al., 2010). Although the moth was able to establish itself homogeneously over the landscape as evidenced by the seemingly synchronous event peak frequency at 5500 cal. yr BP, the climatic and environmental conditions do not appear to have been sufficient to drastically alter northern event frequencies.
Understanding the relatively attenuated change in LSBP event frequencies observed in the northern lakes during favorable Holocene climate conditions may be elucidated by contrasting the northern and southern ecosystems by considering factors that may differ between the two systems. First, in the balsam fir-white birch domain host-insect phenological synchrony along with fir regeneration allows for the establishment of a feedback loop (Baskerville, 1975; Morin, 1994). Spruce budworm defoliation on balsam fir results in canopy tree mortality creating canopy gaps allowing for the release of the pre-established fir seedlings and saplings in the understory to replace the overstory (Morin and Laprise, 1997; Doucet and Blais, 2000; Lemay et al., 2018) predisposing the stand to subsequent outbreaks (Baskerville, 1975; Bouchard et al., 2006, 2007). This interaction is likely possible due to the low fire frequency in this domain (Couillard et al., 2012, 2013, 2021).
In contrast, aspects of nutrient cycling and foliage quality, regeneration, growth, and phenological synchrony in the black spruce system likely differ from those of the balsam fir-spruce budworm system. The higher nutrient quality of fir needles in the balsam fir system allows for the establishment of a positive feedback loop: defoliation by larvae results in greater nutrient availability to surviving trees through litterfall and frass, and these trees can then produce needles of higher nutritional quality allowing populations to increase favoring further defoliation (De Grandpré et al., 2022). This feedback ultimately allows for the build-up of spruce budworm populations permitting populations to move from endemic to epidemic levels. Conversely, defoliation by the insect did not appear to alter nutrient cycling and leaf nutritional quality in the black spruce system (De Grandpré et al., 2022). Further, and to a lesser extent, frequent periods of large budworm populations may be limited by the slower growth rates of black spruce at higher latitudes (Rossi et al., 2009, 2010), complex regeneration dynamics (Martin et al., 2019, 2020) affected by feeding on reproductive structures prior to bud flush (Schooley, 1980; Syme, 1981; Prévost et al., 1988; Wanner et al., 1997), and by previous disturbances (Paquin et al., 1999; Greene et al., 1999; Simard and Payette, 2005; Baltzer et al., 2021). As a result, the black spruce system may have sustained low endemic populations, and only during ‘spill-over’ events from more southern primary host-dominated regions could populations become large enough to affect northern sites.
4.4 Implications for future climate change
Alternatively, it is possible that black spruce remained a poor host even during favorable Holocene climate conditions as the warming was not enough to ameliorate synchrony between bud burst and larval emergence as has been observed in recent history (Pureswaran et al., 2015, 2019; Fuentealba et al., 2017). Indeed, recent warming in northern regions appears to exceed the temperatures experienced during the last 2000 years (Mayewski and Maasch, 2006; Viau et al., 2012; Pei et al., 2017) and even over the course of most of the Holocene (Viau et al., 2006; Viau and Gajewski, 2009; Marcott et al., 2013; Marsicek et al., 2018). Thus, it is likely that past Holocene warming may not serve as an analogue for the recent warming (Mitchell, 1990; Kaufman and Broadman, 2023) suggesting that past event frequencies observed in this study may not be indicative of future event behavior.
5 Conclusion
Our results are the first comparing SBW activity between the spruce-moss and balsam fir-white birch domains. Apparent synchronicity in LSBP event frequency was observed at a regional and supra-regional scale. The post-glacial recolonization of balsam fir around 5500 cal. yr BP likely allowed the SBW to spread more widely over the landscape. Black spruce does not appear to have been a preferred host of the SBW during the climatic variations of the last 8000 years. Nevertheless, current climate change is unmatched in terms of warming rates during the Holocene (Marsicek et al., 2018). Moreover, the average temperature of the Holocene is nearly 1.7 °C lower than that recorded in 2016, so our results are only suggestive of potential future LSBP event frequency behavior and not an accurate prediction.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
HT: Formal Analysis, Investigation, Visualization, Software, Validation, Conceptualization, Writing – review & editing, Data curation, Writing – original draft, Methodology. M-AL: Writing – review & editing. HM: Funding acquisition, Project administration, Supervision, Investigation, Writing – review & editing, Methodology, Conceptualization, Resources.
Funding
The author(s) declared financial support was received for this work and/or its publication. Funding was provided by the Natural Sciences and Engineering Research Council (NSERC) of Canada Industrial Research Chair on black spruce growth and the effect of the spruce budworm on landscape heterogeneity in the boreal forest grant number 499381-15. Funding was obtained by HM.
Acknowledgments
The authors would like to thank the 2 reviewers for their suggestions and feedback greatly improving the manuscript. Prior to an abrupt career change, Dr. Olivier Blarquez provided advice on methods, analysis, and interpretation. Thanks to Dr. Louis De Grandpré for providing comments on an earlier version of the manuscript and providing thoughts relating to our results. Thank you to Marika Tremblay and Guillaume Vigneault for help in the laboratory. Thanks to Claire Fournier, and Mireille Boulianne for preparing sucrose solution and providing lab equipment.
Conflict of interest
The author(s) declared that this work was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declared that generative AI was not used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1682907/full#supplementary-material
References
Ali A. A., Higuera P. E., Bergeron Y., and Carcaillet C. (2009). Comparing fire-history interpretations based on area, number and estimated volume of macroscopic charcoal in lake sediments. Quat Res. 72, 462–468. doi: 10.1016/j.yqres.2009.07.002
Antonucci S., Rossi S., Deslauriers A., Lombardi F., Marchetti M., and Tognetti R. (2015). Synchronisms and correlations of spring phenology between apical and lateral meristems in two boreal conifers. Tree Physiol. 35, 1086–1094. doi: 10.1093/treephys/tpv077
Appleby P. G. and Oldfield F. (1978). The calculation of lead-210 dates assuming a constant rate of supply of unsupported Pb-210 to the sediment. Catena 5, 1–8. doi: 10.1016/S0341-8162(78)80002-2
Appleby P. G. and Oldfieldz F. (1983). The assessment of 210Pb data from sites with varying sediment accumulation rates. Hydrobiologia 103, 29–35. doi: 10.1007/BF00028424
Bajolle L., Larocque-Tobler I., Gandouin E., Lavoie M., Bergeron Y., and Ali A. A. (2018). Major postglacial summer temperature changes in the central coniferous boreal forest of Quebec (Canada) inferred using chironomid assemblages. J. Quat Sci. 33, 409–420. doi: 10.1002/jqs.3022
Baltzer J. L., Day N. J., Walker X. J., Greene D., Mack M. C., Alexander H. D., et al. (2021). Increasing fire and the decline of fire adapted black spruce in the boreal forest. Proc. Natl. Acad. Sci. U.S.A. 118, e2024872118. doi: 10.1073/pnas.2024872118
Baskerville G. L. (1975). Spruce budworm- Super silviculturist. For Chron 51, 138–140. doi: 10.5558/tfc51138-4
Bellemin-Noël B., Bourassa S., Despland E., De Grandpré L., and Pureswaran D. S. (2021). Improved performance of the eastern spruce budworm on black spruce as warming temperatures disrupt phenological defences. Glob Chang Biol. 27, 3358–3366. doi: 10.1111/gcb.15643
Bennett K. and Buck C. E. (2016). Interpretation of lake sediment accumulation rates. Holocene 26, 1092–1102. doi: 10.1177/0959683616632880
Bergeron Y. and Dubue M. (1988). Succession in the southern part of the Canadian boreal forest. Vegetatio 79, 51–63. doi: 10.1007/BF00044848
Bergeron Y., Leduc A., Joyal C., and Morin H. (1995). Balsam fir mortality following the last spruce budworm outbreak in northwestern Québec. Can. J. For Res. 25, 1375–1384. doi: 10.1139/x95-150
Blaauw M., Christen J. A., and Aquino Lopez M. A. (2022). rbacon: Age-Depth Modelling using Bayesian Statistics. R package version 2.5.8. Available online at: https://cran.r-project.org/package=rbacon (Accessed October 5, 2025).
Blais J. R. (1957). Some relationships of the spruce budworm, Choristoneura fumiferana (Clem.) to black spruce, Picea mariana (Moench) Voss. For Chron 33, 364–372. doi: 10.5558/tfc33364-4
Blais J. R. (1958). Effects of defoliation by spruce budworm (Choristoneura fumiferana Clem.) on radial growth at breast height of balsam fir (Abies balsamea (L.) Mill.) and white spruce (Picea glauca (Moench) Voss.). For Chron 34, 39–47. doi: 10.5558/tfc34039-1
Blais J. R. (1965). Spruce budworm outbreaks in the past three centuries in the Laurentide Park, Québec. For. Sci. 11, 130–138. doi: 10.1093/forestscience/11.2.130
Blais J. R. (1981). Mortality of balsam fir and white spruce following a spruce budworm outbreak in the Ottawa River watershed in Québec. Can. J. For Res. 11, 620–629. doi: 10.1139/x81-085
Blarquez O., Ali A. A., Girardin M. P., Grondin P., Fréchette B., Bergeron Y., et al. (2015). Regional paleofire regimes affected by non-uniform climate, vegetation and human drivers. Sci. Rep. 5, 13356. doi: 10.1038/srep13356
Blarquez O., Girardin M. P., Leys B., Ali A. A., Aleman J. C., Bergeron Y., et al. (2013). Paleofire reconstruction based on an ensemble-member strategy applied to sedimentary charcoal. Geophys Res. Lett. 40, 2667–2672. doi: 10.1002/grl.50504
Blarquez O., Vannière B., Marlon J. R., Daniau A. L., Power M. J., Brewer S., et al. (2014). Paleofire: An R package to analyse sedimentary charcoal records from the Global Charcoal Database to reconstruct past biomass burning. Comput. Geosci 72, 255–261. doi: 10.1016/j.cageo.2014.07.020
Bognounou F., De Grandpré L., Pureswaran D. S., and Kneeshaw D. (2017). Temporal variation in plant neighborhood effects on the defoliation of primary and secondary hosts by an insect pest. Ecosphere 8, e01759. doi: 10.1002/ecs2.1759
Bouchard M., Kneeshaw D., and Bergeron Y. (2006). Forest dynamics after successive spruce budworm outbreaks in mixedwood forests. Ecology 87, 2319–2329. doi: 10.1890/0012-9658(2006)87[2319:FDASSB]2.0.CO;2
Bouchard M., Kneeshaw D., and Messier C. (2007). Forest dynamics following spruce budworm outbreaks in the northern and southern mixedwoods of central Québec. Can. J. For Res. 37, 763–772. doi: 10.1139/X06-278
Boulanger Y. and Arseneault D. (2004). Spruce budworm outbreaks in eastern Québec over the last 450 years. Can. J. For Res. 34, 1035–1043. doi: 10.1139/x03-269
Brock F., Higham T., Ditchfield P., and Bronk Ramsey C. (2010). Current pretreatment methods for AMS radiocarbon dating at the Oxford Radiocarbon Accelerator Unit (Orau). Radiocarbon 52, 103–112. doi: 10.1017/S0033822200045069
Cooke B. J. and Lorenzetti F. (2006). The dynamics of forest tent caterpillar outbreaks in Québec, Canada. For Ecol. Manage 226, 110–121. doi: 10.1016/j.foreco.2006.01.034
Cooke B. J., MacQuarrie C. J. K., and Lorenzetti F. (2012). The dynamics of forest tent caterpillar outbreaks across east-central Canada. Ecography 35, 422–435. doi: 10.1111/j.1600-0587.2011.07083.x
Couillard P.-L., Payette S., and Grondin P. (2012). Recent impact of fire on high-altitude balsam fir forests in south-central Quebec. Canadian Journal of Forest Research 42, 1289–1305. doi: 10.1139/x2012-081
Couillard P.-L., Payette S., and Grondin P. (2013).Long-term impact of fire on high-altitude balsam fir (Abies blasamea) forests in south-central Quebec deduced from soil charcoal. Canadian Journal of Forest Research 43, 188–199. doi: 10.1139/cjfr-2012-0414
Couillard P.-L., Payette S., Lavoie M., and Frégeau M. (2021).Precarious resilience of the boreal forest of eastern North America during the Holocene. Forest Ecology and Management485, 118954. doi: 10.1016/j.foreco.2021.118954
Crann C. A., Murseli S., St-Jean G., Zhao X., Clark I. D., and Kieser W. E. (2017). First status report on Radiocarbon sample preparation techniques at the A.E. Lalonde AMS Laboratory (Ottawa, Canada). Radiocarbon 59, 695–704. doi: 10.1017/RDC.2016.55
De Grandpré L., Kneeshaw D. D., Perigon S., Boucher D., Marchand M., Pureswaran D., et al. (2019). Adverse climatic periods precede and amplify defoliator-induced tree mortality in eastern boreal North America. J. Ecol. 107, 452–467. doi: 10.1111/1365-2745.13012
De Grandpré L., Marchand M., Kneeshaw D. D., Paré D., Boucher D., Bourassa S., et al. (2022). Defoliation-induced changes in foliage quality may trigger broad-scale insect outbreaks. Commun. Biol. 5, 463. doi: 10.1038/s42003-022-03407-8
Doucet R. and Blais L. (2000). “Comparative growth of balsam fir and black spruce advance regeneration after logging. J Sustainable Forestry 10, 235–239.
Dyke A. S. (2005). Late quaternary vegetation history of northern North America based on pollen, macrofossil and, faunal remains. Rev. Geogr. Montr 59, 211–262. doi: 10.7202/014755ar
Eakins J. D. and Morrison R. T. (1978). A new procedure for the determination of lead-210 in lake and marine sediments. Int. J. Appl. Radiat. Isot 29, 531–536. doi: 10.1016/0020-708X(78)90161-8
Edwards T. W. D., Wolfe B. B., and Macdonald G. M. (1996). Influence of changing atmospheric circulation on precipitation ∂18O-temperature relations in Canada during the Holocene. Quat Res. 46, 211–218. doi: 10.1006/qres.1996.0061
Fleming R. A. (2000). Climate change and insect disturbance regimes in Canada’s boreal forests. World Resource Rev. 12, 521–548.
Flynn W. W. (1968). The determination of low levels of polonium-210 in environmental materials. Anal. Chim. Acta 43, 221–227. doi: 10.1016/S0003-2670(00)89210-7
Fréchette B., Richard P. J. H., Grondin P., Lavoie M., and Larouche A. C. (2018). Histoire postglaciaire de la vegetation et du climat des pessières et des sapinières de l’ouest du Québec. Gouvernemnt du Québec, Ministère des forêts, de la faune et des parcs. Direction de la recherche forestière. Mémoire Recherche forestiére n 179, 165 pp.
Fréchette B., Richard P. J. H., Grondin P., Lavoie M., and Larouche A. C. (2021). Histoire postglaciaire de la vegetation et du climat des pessières et des sapinières de l’est du Québec et du Labrador méridional. Gouvernemnt du Québec, Ministère des forêts, de la faune et des parcs. Direction la Recherche forestière Mémoire Recherche forestière n 186, 170 pp.
Frey B. S. and Pamini P. (1986). “Cladocera analysis,” in Handbook of Holocene Palaeoecology and Palaeohydrology (Chichester: J. Wiley) 677–692.
Fuentealba A. and Bauce E. (2012). Carry-over effect of host nutritional quality on performance of spruce budworm progeny. Bull. Entomo Res. 102, 275–284. doi: 10.1017/S0007485311000617
Fuentealba A., Pureswaran D., Bauce E., and Despland E. (2017). How does synchrony with host plant affect the performance of an outbreaking insect defoliator? Oecologia 184, 847–857. doi: 10.1007/s00442-017-3914-4
Fuentealba A., Sagne S., Pureswaran D., Bauce E., and Despland E. (2018). Defining the window of opportunity for feeding initiation by second-instar spruce budworm larvae. Can. J. For Res. 48, 285–291. doi: 10.1139/cjfr-2017-0133
Garralla S. (1991). L’analyse pollinique des sédiments d’un lac de la région de Chibougamau, Québec (Sainte-Foy, QC: Université Laval).
Garralla S. and Gajewski K. (1992). Holocene vegetation history of the boreal forest near Chibougamau, central Québec. Can. J. Bot. 70, 1364–1368. doi: 10.1139/b92-171
Gavin D. G., Hu F. S., Lertzman K., and Corbett P. (2006). Weak climatic control of stand-scale fire history during the late Holocene. Ecology 87, 1722–1732. doi: 10.1890/0012-9658(2006)87[1722:WCCOSF]2.0.CO;2
Gray D. R. (2008). The relationship between climate and outbreak characteristics of the spruce budworm in eastern Canada. Clim Change 87, 361–383. doi: 10.1007/s10584-007-9317-5
Greene D. F., Zasada J. C., Sirois L., Kneeshaw D., Morin H., Charron I., et al. (1999). A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For Res. 29, 824–839. doi: 10.1139/x98-112
Harper K. A., Bergeron Y., Gauthier S., and Drapeau P. (2002). Post-fire development of canopy structure and composition in black spruce forests of Abitibi, Québec: A landscape scale study. Silva Fenn 36, 249–263. doi: 10.14214/sf.561
Heide K. (1984). Holocene pollen stratigraphy from a lake and small hollow in north-central Wisconsin, USA. Palynology 8, 3–19. doi: 10.1080/01916122.1984.9989268
Hennigar C. R., MacLean D. A., Quiring D. T., and Kershaw J. J. A. (2008). Differences in spruce budworm defoliation among balsam fir and white, red, and black spruce. For. Sci. 54, 158–166. doi: 10.1093/forestscience/54.2.158
Higuera P. E., Brubaker L. B., Anderson P. M., Hu F. S., and Brown T. A. (2009). Vegetation mediated the impacts of postglacial climate change on fire regimes in the south-central Brooks Range, Alaska. Ecol. Monogr. 79, 201–219. doi: 10.1890/07-2019.1
Higuera P. E., Gavin D. G., Bartlein P. J., and Hallet D. J. (2010). Peak detection in sediment-charcoal records: impacts of alternative data analysis methods on fire-history interpretations. Int. J. Wildland Fire 19, 996–1014. doi: 10.1071/WF09134
James P. M. A., Fortin M. J., Sturtevant B. R., Fall A., and Kneeshaw D. (2011). Modelling spatial interactions among fire, spruce budworm, and logging in the boreal forest. Ecosystems 14, 60–75. doi: 10.1007/s10021-010-9395-5
Jardon Y. (2001). Long term analysis of spruce budworm outbreak in a large scale area, a dendrochronological approach (Montréal, QC: Université du Québec à Montréal).
Jardon Y., Morin H., and Dutilleul P. (2003). Périodicité et synchronisme des épidémies de la tordeuse des bourgeons de l’épinette au Québec. Can. J. For Res. 33, 1947–1961. doi: 10.1139/x03-108
Kaufman D. S. and Broadman E. (2023). Revisiting the Holocene global temperature conundrum. Nature 614, 425–435. doi: 10.1038/s41586-022-05536-w
Kaufman D., McKay N., Routson C., Erb M., Davis B., Heiri O., et al. (2020). A global database of Holocene paleotemperature records. Sci. Data 7, 115. doi: 10.1038/s41597-020-0445-3
Kieser W. E., Zhao X.-L., Clark I. D., Cornett R. J., Litherland A. E., Klein M., et al. (2015). The André E. Lalonde AMS Laboratory- the new accelerator mass spectrometry facility at the University of Ottawa. Nucl. Instrum Methods Phys. Res. B 361, 110–114. doi: 10.1016/j.nimb.2015.03.014
King G. A. (1986). Deglaciation and vegetation history of western Labrador and adjacent Québec (Minneapolis, Saint Paul, Minnesota: University of Minnesota).
Krause C. (1997). The use of dendrochronological material from buildings to get information about past spruce budworm outbreaks. Can. J. For Res. 27, 69–75. doi: 10.1139/x96-168
Krause C. and Morin H. (1995). Impact of spruce budworm defoliation on the number of latewood tracheids in balsam fir and black spruce. Can. J. For Res. 25, 2029–2034. doi: 10.1139/x95-219
Lavoie M. and Filion L. (2001). Holocene vegetation dynamics of Anticosti island, Québec, and consequences of remoteness on ecological succession. Quat Res. 56, 112–127. doi: 10.1006/qres.2001.2239
Lawrence R. K., Mattson W. J., and Haack R. A. (1997). White spruce and the spruce budworm: defining the phenological window of susceptibility. Can. Entomo 129, 291–318. doi: 10.4039/Ent129291-2
Leclerc M.-A., Simard M., and Morin H. (2024). Lepidopteran scales in lakes sediments as a reliable proxy for spruce budworm outbreak events in the boreal forest of Eastern Canada. Holocene 34, 978–986. doi: 10.1177/09596836241236326
Leclerc M.-A., Simard M., and Morin H. (2025). Reconstructing spruce budworm outbreak severity: a comparison of paleoecological and tree-ring signals. PloS One 20, e0329406. doi: 10.1371/journal.pone.0329406
Lehman J. T. (1975). Reconstructing the rate of accumulation of lake sediment: the effect of sediment focusing. Quat Res. 5, 541–550. doi: 10.1016/0033-5894(75)90015-0
Lemay A., Krause C., Achim A., and Bégin J. (2018). Growth and wood quality of black spruce and balsam fir following careful logging around small merchantable stems (CLASS) in the boreal forest of Quebec, Canada. Forestry 91, 271–282. doi: 10.1093/forestry/cpw060
Lindner M., Maroschek M., Netherer S., Kremer A., Barbati A., Garcia-Gonzalo J., et al. (2010). Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For Ecol. Manage 259, 698–709. doi: 10.1016/j.foreco.2009.09.023
MacDonald G. M., Larsen C. P. S., Szeicz J. M., and Moser K. A. (1991). The reconstruction of boreal forest fire history from lake sediments: a comparison of charcoal, pollen, sedimentological, and geochemical indices. Quat Sci. Rev. 10, 53–71. doi: 10.1016/0277-3791(91)90030-X
MacLean D. A. (1980). Vulnerability of fir-spruce stands during uncontrolled spruce budworm outbreaks: A review and discussion. For Chron 56, 213–221. doi: 10.5558/tfc56213-5
MacLean D. A. (1984). Effects of spruce budworm outbreaks on the productivity and stability of balsam fir forests. For Chron 60, 273–279. doi: 10.5558/tfc60273-5
MacLean D. A. (1988). “Effects of spruce budworm outbreaks on vegetation, structure, and succession of balsam fir forests on Cape Breton island, Canada,” in Plant Form and Vegetation Structure (The Hague, The Netherlands: SPB Academic Pub), 253–261.
Marcott S. A., Shakun J. D., Clark P. U., and Mix A. C. (2013). A reconstruction of regional and global temperature for the past 11,300 years. Science 339, 1198–1201. doi: 10.1126/science.1228026
Marsicek J., Shuman B. N., Bartlein P. J., Shafer S. L., and Brewer S. (2018). Reconciling divergent trends and millennial variations in Holocene temperatures. Nature 554, 92–96. doi: 10.1038/nature25464
Martin M., Montoro Girona M., and Morin H. (2020). Driving factors of conifer regeneration dynamics in eastern Canadian boreal old-growth forests. PLoS One 15, e0230221. doi: 10.1371/journal.pone.0230221
Martin M., Morin H., and Fenton N. J. (2019). Secondary disturbances of low and moderate severity drive the dynamics of eastern Canadian boreal old-growth forests. Ann. For Sci. 76, 108. doi: 10.1007/s13595-019-0891-2
Mathieu G. G., Biscaye P. E., Lupton R. A., and Hammond D. E. (1988). System for measurement of 222Rn at low levels in natural waters. Health Phys. 55, 989–992. doi: 10.1097/00004032-198812000-00015
Mattson J., Slocum S. S., and Koller C. N. (1983). “Spruce budworm (Choristoneura fumiferana) performance in relation to foliar chemistry,” in Proceedings: Forest defoliator-host interactions: A comparison between gypsy moth and spruce budworm. Eds. Talerico R. L. and Montgomery M. (U.S Department of Agriculture, Forest Service, Northeeastern Forest Experiment Station, Broomall PA), 55–65.
Mayewski P. A. and Maasch K. A. (2006). Recent warming inconsistent with natural association between temperature and atmospheric circulation over the last 2000 years. Clim Past Discuss 2, 327–355. doi: 10.5194/cpd-2-327-2006
Mcandrews J. H. (1988). “Human disturbance of North American forests and grasslands: The fossil pollen recor,” in Vegetation history. Handbook of vegetation science. Eds. Huntley B. and Webb T. (Springer, Dordrecht), 673–697.
Mitchell J. F. B. (1990). Greenhouse warming: Is the mid-Holocene a good analogue? J. Clim 3, 1177–1192. doi: 10.1175/1520-0442(1990)003%3C1177:GWITMH%3E2.0.CO;2
Montoro Girona M., Navarro L., and Morin H. (2018). A secret hidden in the sediments: lepidoptera scales. Front. Ecol. Evol. 6. doi: 10.3389/fevo.2018.00002
Morin H. (1994). Dynamics of balsam fir forests in relation to spruce budworm outbreaks in the boreal zone of Québec. Can. J. For Res. 24, 730–741. doi: 10.1139/x94-097
Morin H. (1998). Importance et évolution des épidémies de la tordeuse des bourgeons de l’épinette dans l’est du Canada: l’apport de la dendrochronologie. Rev. Geogr. Montr 52, 237–244. doi: 10.7202/004856ar
Morin H., Gagnon R., Lemay A., and Navarro L. (2021). “Revisiting the relationship between spruce budworm outbreaks and forest dynamics over the Holocene in Eastern North America based on novel proxies,” in Plant Disturbance Ecology, 2nd edition (London, UK: Academic Press), 463–487. doi: 10.1016/B978-0-12-818813-2.00013-7
Morin H. and Laprise D. (1990). Histoire récente des épidémies de la Tordeuse des bourgeons de l’épinette au nord du lac Saint-Jean (Québec): une analyse dendrochronologique. Can. J. For Res. 20, 1–8. doi: 10.1139/x90-001
Morin H. and Laprise D. (1997). Seedling bank dynamics in boreal balsam fir forests. Can. J. For Res. 27, 1442–1451. doi: 10.1139/x97-113
Mudelsee M., Börngen M., Tetzlaff G., and Grünewald U. (2004). Extreme floods in central Europe over the past 500 years: Role of cyclone pathway “Zugstrasse vb. J. Geophys Res. Atmos 109, 1–21. doi: 10.1029/2004JD005034
Navarro L., Harvey A. E., Ali A., Bergeron Y., and Morin H. (2018b). A Holocene landscape dynamic multiproxy reconstruction: How do interactions between fire and insect outbreaks shape an ecosystem over long time scales? PLoS One 13, e0204316. doi: 10.1371/journal.pone.0204316
Navarro L., Harvey A. E., and Morin H. (2018a). Lepidoptera wing scales: a new paleoecological indicator for reconstructing spruce budworm abundance. Can. J. For Res. 48, 302–308. doi: 10.1139/cjfr-2017-0009
Nealis V. G. (2016). Comparative ecology of conifer-feeding spruce budworms (Lepidoptera: Tortricidae). Can. Ento 148, S33–S57. doi: 10.4039/tce.2015.15
Nealis V. G. and Régnière J. (2004). Insect-host relationships influencing disturbance by the spruce budworm in a boreal mixedwood forest. Can. J. For Res. 34, 1870–1882. doi: 10.1139/x04-061
Otvos I. S., Clark R. C., and Clarke L. J. (1971). “The hemlock looper in Newfoundland: the outbreak 1966 to 1971, and aerial spraying 1968 and 1969. Information Report N-X-68 (St John’s, Newfoundland: Environment Canada Forestry Service) 77 pp.
Otvos I. S., Clarke L., and Durling D. S. (1979). “A history of recorded eastern hemlock looper outbreaks in Newfoundland,” in Newfoundland Forest Research Centre No. N-X-179. (St John’s, Newfoundland: Environment Canada Forestry Service) 46 pp.
Paquin R., Margolis H. A., Doucet R., and Coyea M. R. (1999). Comparison of growth and physiology of layers and naturally established seedlings of black spruce in a boreal cutover in Québec. Can. J. For Res. 29, 1–8. doi: 10.1139/x98-171
Pei Q., Zhang D. D., Li J., and Lee H. F. (2017). Proxy-based Northern Hemisphere temperature reconstruction for the mid-to-late Holocene. Theor. Appl. Climatol 130, 1043–1053. doi: 10.1007/s00704-016-1932-5
Prévost Y. H., Laing J. E., and Haavisto V. F. (1988). Seasonal damage by insects and squirrels to female reproductive structures of black spruce, Picea mariana (Mill.) B.S.P. Can. Ent. 120, 1113–1121. doi: 10.4039/Ent1201113-12
Pureswaran D. S., De Grandpré L., Paré D., Taylor A., Barrette M., Morin H., et al. (2015). Climate-induced changes in host tree-insect phenology may drive ecological state-shift in boreal forests. Ecology 96, 1480–1491. doi: 10.1890/13-2366.1
Pureswaran D. S., Neau M., Marchand M., De Grandpré L., and Kneeshaw D. (2019). Phenological synchrony between eastern spruce budworm and its host trees increases with warmer temperatures in the boreal forest. Ecol. Evol. 9, 576–586. doi: 10.1002/ece3.4779
Pureswaran D. S., Roques A., and Battisti A. (2018). Forest insects and climate change. Curr. For Rep. 4, 35–50. doi: 10.1007/s40725-018-0075-6
Québec Ministry of Forests, Fauna, and Parks Data from: Fourth decadal forest inventory: Shapefiles of forest inventory data including forest composition and previous disturbance (Québec Ministry of Forests, Fauna and Parks). Available online at: https://www.donneesquebec.ca/recherche/dataset/carte-ecoforestiere-avec-perturbations (Accessed October 3, 2025).
R core team (2020). R: a language and environment for statistical computing (Vienna Austria: R foundation for statistical computing).
Régnière J. and Nealis V. G. (2018). Two sides of a coin: host-plant synchrony fitness trade-offs in the population dynamics of the western spruce budworm. Insect Sci. 25, 117–126. doi: 10.1111/1744-7917.12407
Régnière J., St-Amant R., and Duval P. (2012). Predicting insect distributions under climate change from physiological responses: Spruce budworm as an example. Biol. Invasions 14, 1571–1586. doi: 10.1007/s10530-010-9918-1
Reimer P. J., Austin W. E. N., Bard E., Bayliss A., Blackwell P. G., Ramsey C. B., et al. (2020). The IntCal20 Northern Hemisphere Radiocarbon Age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757. doi: 10.1017/RDC.2020.41
Reimer P. J., Brown T. A., and Reimer R. W. (2004). Discussion reporting and calibration of post-bomb 14C data. Radiocarbon 46, 1299–1304. doi: 10.1017/S0033822200033154
Remy C. C., Hély C., Blarquez O., Magnan G., Bergeron Y., Lavoie M., et al. (2017). Different regional climatic drivers of Holocene large wildfires in boreal forests of northeastern America. Environ. Res. Lett. 12, 0350035. doi: 10.1088/1748-9326/aa5aff
Renberg I. and Hansson H. (2008). The HTH sediment corer. J. Paleolimnol 40, 655–659. doi: 10.1007/s10933-007-9188-9
Richard P. (1973). Histoire postglaciaire comparée de la végétation dans deux localités au nord du Parc des Laurentides, Québec. Nat. Can. 100, 577–590.
Richard P. (1975). Histoire postglaciaire de la végétation dans la partie centrale du parc des Laurentides, Québec. Nat. Can. 102, 669–681.
Richard P. J. H. (1977). “Histoire post-wisconsinienne de la végétation du Québec méridional par l’analyse pollinique,” in Service de la recherche, Direction générale des forêts, Ministère des Terres et Forêts du Québec (Canada: Publications et rapports divers), 142 p. Tome 1, xxiv + 312 p.; tome 2.
Richard P. J. H., Fréchette B., Grondin P., and Lavoie M. (2020). Histoire postglaciaire de la végétation de la forêt boréale du Québec et du Labrador. Nat. Can. 144, 63–76. doi: 10.7202/1070086ar
Richard P. J. H., Larouche A. C., and Lortie G. (1992). Postglacial paleophytogeography and paleoclimates in the western part of the Lower Saint Lawrence River region, Québec. Rev. Geogr. Montr 46, 151–172. doi: 10.7202/032900ar
Rossi S., Cairo E., Krause C., and Deslauriers A. (2015). Growth and basic wood properties of black spruce along an alti-latitudinal gradient in Québec, Canada. Ann. For Sci. 72, 77–87. doi: 10.1007/s13595-014-0399-8
Rossi S., Morin H., and Tremblay M.-J. (2010). Growth and productivity of black spruce (Picea mariana) belonging to the first cohorts in stands within and north of the commercial forest in Quebec, Canada. Ann. For. Sci. 67, 807. doi: 10.1051/forest/2010043
Rossi S., Tremblay M.-J., Morin H., and Savard G. (2009). Growth and productivity of black spruce in even- and uneven-aged stands at the limit of the closed boreal forest. For Ecol. Manage 258, 2153–2161. doi: 10.1016/j.foreco.2009.08.023
Royama T. (1984). Population dynamics of the spruce budworm Choristoneura fumiferana. Ecol. Monogr. 54, 429–462. doi: 10.2307/1942595
Saarnisto M. (1974). The deglaciation history of the Lake Superior region and its climatic implications. Quat Res. 4, 316–339. doi: 10.1016/0033-5894(74)90019-2
Saucier J.-P., Robitaille A., and Grondin P. (2009). “Cadre bioclimatique du Québec,” in Manuel de Foresterie, 2nd Édition. Eds. Doucet R. and Côté M. (Éditions MultiMondes, Québec City, Québec), 186–205. Ordre des Ingénieurs du Québec.
Schooley H. O. (1980). Damage to black spruce cone crops by the spruce budworm. Can. For. Serv. Nfld. For. Res. Cent. Inf. Rep., N–X-187.
Seidl R., Honkaniemi J., Aakala T., Aleinikov A., Angelstam P., Bouchard M., et al. (2020). Globally consistent climate sensitivity of natural disturbances across boreal and temperate forest ecosystems. Ecography 43, 967–978. doi: 10.1111/ecog.04995
Seidl R., Thom D., Kautz M., Martin-Benito D., Peltoniemi M., Vacchiano G., et al. (2017). Forest disturbances under climate change. Nat. Clim Chang 7, 395–402. doi: 10.1038/nclimate3303
Shorohova E., Kneeshaw D., Kuuluvainen T., and Gauthier S. (2011). Variability and dynamics of old-growth forests in the circumboreal zone: Implications for conservation, restoration and management. Silva Fenn 45, 785–806. doi: 10.14214/sf.72
Simard I., Morin H., and Lavoie C. (2006). A millennial-scale reconstruction of spruce budworm abundance inSaguenay, Québec, Canada. Holocene 16, 31–37. doi: 10.1191/0959683606hl904rp
Simard I., Morin H., and Potelle B. (2002). A new paleoecological approach to reconstruct long-term history of spruce budworm outbreaks. Can. J. For Res. 32, 428–438. doi: 10.1139/x01-215
Simard M. and Payette S. (2005). Reduction of black spruce seed bank by spruce budworm infestation compromises postfire stand regeneration. Can. J. For Res. 35, 1686–1696. doi: 10.1139/x05-083
Smith D., Whitehouse N., Bunting M. J., and Chapman H. (2010). Can we characterise ‘openness’ in the Holocene palaeoenvironmental record? Modern analogue studies of insect faunas and pollen spectra from Dunham Massey deer park and Epping Forest, England. Holocene 20, 215–229. doi: 10.1177/0959683609350392
St-Jean G., Kieser W. E., Crann C. A., and Murseli S. (2017). Semi-automated equipment for CO2 purification and graphitization at the A.E. LaLonde AMS Laboratory (Ottawa, Canada). Radiocarbon 59, 941–956. doi: 10.1017/RDC.2016.57
Stuiver M. and Polach P. A. (1977). Discussion reporting of 14C data. Radiocarbon 19, 355–363. doi: 10.1017/S0033822200003672
Syme P. D. (1981). Black spruce cone and seed insects: a special study. Can. For. Serv. Gt. Lakes For. Cent. For. Res. Newsl. Summer 1981, 1–2.
Viau A. E. and Gajewski K. (2009). Reconstructing millennial-scale, regional paleoclimates of boreal Canada during the Holocene. J. Clim 22, 316–330. doi: 10.1175/2008JCLI2342.1
Viau A. E., Gajewski K., Sawada M. C., and Fines P. (2006). Millennial-scale temperature variations in North America during the Holocene. J. Geophys Res. Atmos 111, D09102. doi: 10.1029/2005JD006031
Viau A. E., Ladd M., and Gajewski K. (2012). The climate of North America during the past 2000 years reconstructed from pollen data. Glob Planet Change 84-85, 75–83. doi: 10.1016/j.gloplacha.2011.09.010
Wanner K. W., Helson B. V., and Kostyk B. C. (1997). Foliar and systemic applications of neem seed extract for control of spruce budworm, Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae), infesting black and white spruce seed orchards. Can. Entomol 129, 645–655. doi: 10.4039/Ent129645-4
Keywords: Holocene, outbreak, climate, Québec, lake sediments
Citation: Terreaux de Félice H, Leclerc M-A and Morin H (2025) Was black spruce a good host of the spruce budworm in warmer periods of the Holocene? a long-term reconstruction. Front. Ecol. Evol. 13:1682907. doi: 10.3389/fevo.2025.1682907
Received: 09 August 2025; Accepted: 25 November 2025; Revised: 25 November 2025;
Published: 17 December 2025.
Edited by:
Antonio Garcia-Alix, University of Granada, SpainReviewed by:
Felipe Dargent, University of Ottawa, CanadaLaura Jiménez Liébanas, Philosophisch-Naturwissenschaftliche Fakultät, Switzerland
Copyright © 2025 Terreaux de Félice, Leclerc and Morin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marc-Antoine Leclerc, bGVjbGVyY21hcmNhbnRvaW5lQGdtYWlsLmNvbQ==