 Karla Cáceres-Mago
Karla Cáceres-Mago Alicia Cáceres1
Alicia Cáceres1 Eleinis Ávila-Lovera
Eleinis Ávila-Lovera Wilmer Tezara
Wilmer Tezara- 1Centro de Botánica Tropical, Instituto de Biología Experimental, Universidad Central de Venezuela, Caracas, Venezuela
- 2Instituto Multidisciplinario de Biología Vegetal, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Universidad Nacional de Córdoba, Córdoba, Argentina
- 3School of Biological Sciences, University of Utah, Salt Lake City, UT, United States
- 4Facultad de Ciencias Agropecuarias, Universidad Técnica Luis Vargas Torres, Esmeraldas, Ecuador
The tropical dry forests (TDFs) of the Macanao Peninsula (Margarita Island, Venezuela) have been severely degraded by open-cast sand mining for over four decades, reducing vegetation cover and disrupting soil biological processes, including those mediated by arbuscular mycorrhizal fungi (AMF). To assess whether enriching native AMF communities can improve plant performance, we evaluated the physiological traits and growth of three woody species (Bulnesia arborea, Caesalpinia mollis and Piptadenia flava) grown for eight months under greenhouse conditions in noninoculated (NI) and inoculated (I) treatments, both using the same non-sterile forest soil to ensure that inoculation represented enrichment of the native AMF community, which reflects realistic restoration scenarios. Species differed markedly in their responses: in B. arborea, AMF enrichment increased net photosynthetic rate and stomatal conductance by 50% and 31%, respectively, and significantly enhanced the maximum rate of RuBisCO carboxylation and biomass accumulation; in C. mollis, inoculation increased CO2-saturated photosynthetic rate but did not affect growth; and in P. flava, inoculated plants showed greater shoot biomass and root length despite no detectable changes in photosynthetic parameters. These species-specific responses suggest that B. arborea may hold potential for future restoration testing in the Macanao Peninsula, and further ecological evaluation is needed before selecting species for restoration.
1 Introduction
Tropical dry forests (TDFs) constitute over 40% of the world’s tropical forests (Murphy and Lugo, 1995) and comprise tree communities adapted to warm climates and seasonal droughts (Santos et al., 2022; Oyedeji, 2023). These ecosystems are among the most threatened due to anthropogenic activities, which degrade soil properties, vegetation structure, and environmental conditions (Fajardo et al., 2013; Ávila-Lovera et al., 2023; Mesa-Sierra et al., 2025). Recognized for their high endemism and timber species richness, TDFs are rapidly disappearing, emphasizing the need for research and restoration efforts in this biome (Banda et al., 2016; Shahzad et al., 2024). Restoring degraded areas is crucial for conserving biodiversity and improving ecosystem services, particularly when native species are used (Bertolazi et al., 2025).
Soil disturbance during deforestation, such as that caused by opencast mining, disrupts essential biological processes, including those mediated by arbuscular mycorrhizal fungi (AMF), key mutualistic symbionts in terrestrial ecosystems (Xu et al., 2023a). AMF form associations with most tropical and dryland plant species, enhancing the uptake of water and nutrients (particularly P and N) while providing protection against root pathogens (Wahab et al., 2023). Through their extensive extraradical mycelium and the production of glomalin, AMF improve soil aggregation and water retention, thereby enhancing plant stomatal conductance, transpiration, and photosynthetic performance (Shukla et al., 2025; Zaman et al., 2025). These functions are particularly critical in arid and semi-arid ecosystems such as TDFs, where limited water and nutrient availability constrain plant establishment and survival, and AMF enhance plant drought tolerance by improving root absorptive capacity, hydraulic continuity, and water-use efficiency during dry periods (Shukla et al., 2025). Beyond individual plant benefits, AMF promote long-term soil recovery and ecosystem resilience through improved soil structure and organic matter stabilization (Li et al., 2024). This ecological facilitation underlies their growing use in restoration programs of degraded TDFs, where AMF enhance seedling establishment, water relations, and overall soil functionality, offering a sustainable bio-alternative to chemical fertilization.
In the Macanao Peninsula, Margarita Island, Venezuela, four decades of open-cast sand mining have caused severe deforestation and habitat destruction (González, 2007). Mining is considered one of the main causes of deforestation in the region, involving excavation and a transport system, with negative impacts that lead to soil erosion, damage to wildlife habitat, and changes in soil hydrology (Chambi-Legoas et al., 2021; Moulatlet et al., 2023). Studies in TDFs impacted by mining suggest that native species from mature forests can thrive in disturbed sites, supporting their use in ecological restoration plans (Ávila-Lovera et al., 2023; Bertolazi et al., 2025).
Research in Macanao has demonstrated that AMF inoculation enhances growth, physiological performance, and survival of native plants under both drought and well-watered conditions (Cáceres et al., 2014; Tezara et al., 2014). For instance, Piscidia carthagenensis (Fabaceae) inoculated with native AMF exhibited improved gas exchange and water-use efficiency compared to control plants planted in a disturbed area (Kalinhoff, 2012). Eight years postrevegetation, soil quality improvement, including enhanced chemical and microbiological properties, was observed in restored sites (Sierraalta et al., 2021).
Despite evidence that AMF enhance physiological parameters such as photosynthetic rate, stomatal conductance, and water potential under drought stress (Frosi et al., 2016; Chen et al., 2020; Rani et al., 2023), their role remains underexplored in restoration plans for Macanao’s semi-arid ecosystems. To address this knowledge gap and support restoration planning, three native woody species with contrasting functional traits were selected to test their responses to AMF enrichment using an inoculum obtained from soils of the Macanao TDF (Table 1). Bulnesia arborea, Caesalpinia mollis, and Piptadenia flava were chosen based on the following criteria: (a) they are all native species, (b) they present rapid growth, and (c) they belong to families of high ecological importance in these TDFs and associated xerophytic scrubs. B. arborea is a dominant species in the riparian deciduous forest of the Macanao Peninsula. Both C. mollis and P. flava belong to the Fabaceae family, the most species-rich in Venezuelan TDFs and the largest group present in the study area (Fajardo, 2007). Furthermore, C. mollis is classified as vulnerable (VU) according to the Red Book of Venezuelan Flora (Huérfano et al., 2020), warranting special conservation attention. These species are also ecologically important for the yellow-shouldered parrot (Amazona barbadensis), an endangered species endemic to the region. B. arborea and C. mollis are the primary trees used for nesting during the reproductive period, while the flowers and fruits of B. arborea and the seeds of P. flava are a significant part of the parrot’s diet (González, 2007; Rojas-Suárez, 2014).

Table 1. Tree plant species studied in a tropical dry forest of the Macanao Peninsula, including family, leaf habit, and means ± SE of height and diameter at breast height (DBH) (modified from Ávila-Lovera et al., 2019).
This study evaluated how AMF enrichment, through the inoculation of native communities, influences the physiological performance and growth of these three woody species, focusing on gas exchange, photochemical activity, and biomass production. We hypothesized that AMF enrichment would promote species-specific responses reflecting their contrasting adaptations to drought, water-use efficiency, and nutrient acquisition strategies. The outcomes of this study aim to inform restoration strategies for degraded areas in Macanao, contributing to global efforts to recover TDFs—an ecosystem of high conservation priority, yet often overlooked compared to tropical wet forests.
2 Materials and methods
2.1 Study area
The field component of this study was conducted in the Macanao Peninsula, a mountainous region in the western part of Margarita Island (10°56′–11°06′N; 64°10′–64°25′W), approximately 38 km off the coast of mainland Venezuela. The climate of Macanao is semi-arid, with a mean annual temperature of 27° ± 0.3°C and an average annual precipitation of 524 mm ± 87 mm, based on climate data recorded from 1986 to 2006 at the San Francisco de Macanao meteorological station (11°01′30″N, 64°17′26″W; Ávila-Lovera et al., 2019). Precipitation peaks occur in August, with a secondary, smaller peak in December, while the dry season extends from January to June (Fajardo et al., 2013).
Sand extraction in the region has been ongoing for over 46 years, and following the abandonment of mining sites, vegetation recovery is notably slow due to the removal of the soil seed bank and the region’s characteristic aridity (González, 2007).
The TDF of the Macanao Peninsula represents a mature plant community dominated by Spondias mombin (Anacardiaceae), Lonchocarpus punctatus (Fabaceae), Handroanthus chrysanthus (Bignoniaceae), Handroanthus serratifolius (Bignoniaceae), Bulnesia arborea (Zygophyllaceae), and P. carthagenensis (Fabaceae) (González, 2007). This forest serves as the reference ecosystem prior to disturbance, with its lowland areas among the plant communities most threatened by sand mining. Excavated areas are typically abandoned after mining activities, allowing for natural regeneration of vegetation in and around the mine sites, although this process is extremely slow and is often interrupted by the resumption of mining activities after 20–25 years of abandonment.
2.2 Field measurements
To compare the physiological performance of greenhouse-grown plants with that of adult trees in the study area, gas exchange and photochemical activity traits were assessed in adult trees from the TDF located in the Macanao Peninsula during the rainy season of the year (see details below).
2.3 Greenhouse experiment
The greenhouse experiment was carried out at the Institute of Experimental Biology, Central University of Venezuela, located in Caracas (10°24′N, 67°36′W; 1200 masl). The greenhouse was covered with transparent polyethylene and maintained under natural light conditions. During the experiment, air temperature ranged from 20°C to 26°C, relative humidity varied from 83% to 65%, maximum photosynthetic photon flux density reached 550 µmol m−2 s−1, and vapor pressure deficit ranged between 0.4 and 1.6 kPa. Measurements were taken at 08:00 and 13:00 hours, indicating relatively stable microclimatic conditions with moderate daily fluctuations (see Supplementary Table S1 for microclimatic values in both the greenhouse and the field).
Seeds were collected from relatively undisturbed xerophytic scrubs and deciduous forests in the surroundings of mining activity areas. They were stored in plastic bags in a cool environment until transferred to the greenhouse. In the greenhouse, seeds were planted in germination trays, and after the emergence of the first true leaves, 60 seedlings of each species were transplanted into 3 kg polyethylene bags and assigned to two experimental treatments: (1) nonsterile control, with 30 seedlings of each species planted in unsterilized forest soil without AMF inoculum (not inoculated (NI)), and (2) inoculated treatment, with 30 seedlings of each species planted in unsterilized forest soil supplemented with 50 g of AMF inoculum (inoculated (I)). Plants were watered regularly throughout the experiment, and after 8 months of growth (240 days), gas exchange, photochemical activity, response curves of photosynthesis (A) to intercellular CO2 concentration (Ci) (A/Ci curves), and growth variables were measured.
It is important to note that both treatments were established using the same unsterilized forest soil to preserve the native microbial and AMF community naturally present in the substrate. This approach provided ecologically realistic conditions that simulate field restoration scenarios, where soils are never sterile. Therefore, the I treatment represents an enrichment of the native AMF community compared with the NI treatment.
2.4 Soils and AMF inoculum
The soil used in the greenhouse experiment was collected from the TDF of the study area and was characterized as moderately alkaline, with a pH of 7.87 ± 0.08, low P concentration (7.37 ppm ± 1.02 ppm), high N concentration (0.74% ± 0.04%), and elevated soil organic matter content (13% ± 6.3%), according to the classification criteria of the Ministerio de Obras Públicas (MOP) (1972).
To characterize the native AMF community present in these soils, AMF spores were extracted from rhizospheric soil samples collected from adult individuals of the studied plant species. Spores were isolated from 5 g of soil (in triplicate) using the wet-sieving, decanting, and sucrose centrifugation method described by Sieverding et al. (1991). Only intact spores with visible lipid reserves were counted under a stereomicroscope (× 40), and results were expressed as the number of spores per 100 g of dry soil. Morphological identification of AMF morphospecies was based on spore characteristics such as size, wall type and number, color in water and in polyvinyl alcohol mounting medium, reaction to Melzer’s reagent, and other diagnostic features such as ornamentation, subtending hyphae, or scars when present. Morphospecies were identified using the descriptions and identification keys of Schenck and Pérez (1990) and the International Culture Collection of Arbuscular Mycorrhizal Fungi (INVAM, https://invam.ku.edu/species-descriptions). The AMF community identified in these soils (Table 2) represents the natural assemblage used as the basis for preparing the mixed inoculum.

Table 2. Morpho-species of arbuscular mycorrhizal fungi (AMF) isolated from tropical dry forest soils in the Macanao Peninsula.
The AMF inoculum was then prepared using the trap pot method, which consists of preparing a suitable substrate for fungal development and using a highly mycotrophic plant species to promote rapid root colonization and sporulation, generating infective AMF propagules in the soil (Sieverding et al., 1991). For this purpose, plants of Vigna luteola, a highly mycotrophic herbaceous legume (Hernández et al., 2000), were grown in pots containing fresh soil from the TDF to obtain a mixed AMF inoculum (Bever, 1994). After 8 months of cultivation, the aerial parts of the plants were harvested, and the roots were preserved as part of the inoculum. This method allowed an increase in the AMF propagule density in the substrate, reaching 860.67 spores per 100 g of soil ± 120.81 spores per 100 g of soil, between 12 and 30 times higher than the spore density found in the rhizospheric soil of the studied species (Table 2).
Additionally, root samples from the experimental greenhouse plants were collected at harvest and examined microscopically to verify the presence of mycorrhizal structures, confirming that the seedlings had established AMF colonization during the experiment. Fine root fragments were cleared in 10% KOH, acidified with 1% HCl, stained with trypan blue (Phillips and Hayman, 1970), and examined under a light microscope (Nikon Eclipse E200, Tokyo, Japan) for the presence of hyphae, vesicles, and arbuscules.
2.5 Instantaneous gas exchange
Net photosynthetic rate (A), transpiration rate (E), and stomatal conductance (gs) were measured in leaves of five trees per species in the field and 10 individuals per species per treatment in the greenhouse between 08:00 and 11:00 hours using an open-system portable infrared gas analyzer (CIRAS-II, PP Systems Inc., Amesbury, MA, USA) connected to a leaf chamber PLC(B) at a CO2 concentration of 400 µmol mol−1 ± 10 µmol mol−1, a leaf chamber temperature of 29.0°C ± 0.3°C, leaf-to-air vapor pressure deficit (ΔW) of 1.6 kPa ± 0.01 kPa, and photosynthetic photon flux density (PPFD) of 1,500 µmol m−2 s−1 ± 50 µmol m−2 s−1 (light was provided by a LED-based light unit from the same manufacturer). Instantaneous measurements were made on fully expanded healthy leaves. Intercellular CO2 concentration (Ci) was calculated by the program in the CIRAS, after Farquhar et al. (1980). Instantaneous water-use efficiency was estimated as WUE=A/E.
2.6 A/Ci curves
For each species, A/Ci curves (n=5) were determined in well-irrigated plants in both treatments (NI and I) by decreasing atmospheric CO2 concentration (Ca) from 400 µmol mol−1 to zero, and then progressively increasing Ca from zero to 1,800 µmol mol−1. The CO2 was provided by a cylinder filled with pure gas connected to the IRGA. Conditions during A/Ci measurements were the same as in the instantaneous measurements. The A/Ci curves were fitted to an empirical equation where A=b + d ekCi (Tezara et al., 2003). Carboxylation efficiency (CE) was calculated from the initial slope of the curve, and the CO2 compensation point was calculated as Γ=Ln (− b/d)/k. Relative stomatal limitation (Ls) was calculated as Ls=100 × (Ao − A)/Ao, where Ao is A at Ci=Ca and A is the net photosynthesis rate measured at Ci=280 µmol mol−1 (Farquhar and Sharkey, 1982). Relative mesophyll limitation (Lm) was calculated as Lm=100 × (AI − ANI)/AI, where AI is A of leaves in inoculated plants at Ci=800 µmol mol−1, and ANI is A of leaves in noninoculated plants at the same Ci (Jacob and Lawlor, 1991). Thus, Lm is a measure of the capacity of the mesophyll to fix CO2 at saturating Ci, and its value in inoculated plants was assumed to be minimum. Maximum rate of RuBisCO carboxylation (Vcmax), maximum rate of electron transport (Jmax), and triose phosphate utilization rate (TPU) were calculated from the A/Ci curves using the model proposed by Sharkey (2015).
2.7 Chlorophyll a fluorescence of PSII
Maximum quantum yield of PSII (Fv/Fm) was measured on attached dark-adapted leaves at predawn in well-irrigated plants using a fluorometer (PAM 2100, WALZ, Effeltrich, Germany) following the protocol described by Genty et al. (1989). Variable fluorescence (Fv) is Fm − F0, where F0 and Fm are dark-adapted minimum and maximum fluorescence, respectively. Relative quantum yield of PSII at steady-state photosynthesis was calculated as ΦPSII=(F′m − Fs)/F′m, where Fs and F′m are fluorescence at steady-state photosynthesis and maximum fluorescence in light, respectively. Electron transport rate through PSII (J) was estimated as J=ΦPSII × PPFD × α × 0.5, where α is the fraction of incident PPFD absorbed by the leaf, assumed to be 0.84 (Krall and Edwards, 1992). The photochemical (qP=(F′m − Fs)/(F′m − F′0)) and nonphotochemical (qN=1 − (F′m − F′0)/(F′m − F′0)) quenching coefficients were also calculated. Fluorescence parameters were measured simultaneously on the same leaves where gas exchange was measured in five trees per species in the field and 10 plants per species per treatment in the greenhouse.
2.8 Growth variables
After physiological measurements in the greenhouse, 10 plants per species per treatment were harvested to carry out a growth analysis. The number of leaves of each plant was counted, and the shoots and roots were separated. Roots were washed, and soil particles were removed. Total root length was measured using the WinRHIZO Pro program (version 2003b, Regent Instrument, Quebec, Canada). Shoots and roots were dried in an oven at 60°C for 48 h and weighed to determine shoot, root, and total biomass. The shoot–root ratio (S:R) was determined by dividing the shoot dry mass by the root dry mass. Based on the total plant biomass, the mycorrhizal response index (MRI) was calculated as MRI=[(DW(I) − DW(NI))/DW(I)] × 100, where DW(I) is the dry weight of the inoculated plant and DW(NI) is the dry weight of the noninoculated plant. The MRI is considered an indirect indicator of the plant’s response to AMF inoculation and of the functional effectiveness of the mycorrhizal symbiosis, as it does not directly quantify root colonization by fungi but rather the functional outcome of that colonization on plant performance, expressed here in terms of growth (biomass).
2.9 Statistical analysis
Data analysis was performed using Statistica v10 (StatSoft Inc., Tulsa, OK, USA). A one-way analysis of variance (ANOVA) was conducted to determine the significant differences in the variables evaluated among species in the field. In the greenhouse experiment, a two-way ANOVA was applied to all response variables, using species and inoculation treatment as fixed factors. Prior to the analysis, the assumptions of normality (Shapiro–Wilk test) and homogeneity of variances (Levene’s test) were verified, and data were transformed when necessary to meet these criteria. When significant effects were detected, Tukey’s post-hoc test was used for mean comparisons at a significance level of p=0.05. All results are presented as means ± standard error (SE). Graphical representations were produced in SigmaPlot 14.0 (Systat Software, San Jose, CA, USA).
3 Results
3.1 Field measurements
The three tree species studied differed significantly in height, with B. arborea and C. mollis being deciduous and P. flava evergreen (Table 1). Significant differences were found in almost all the physiological traits related to gas exchange (E, gs, and WUE) and in qN among the species in the field (Table 3). Values of A ranged from 4.9 to 6.4 μmol m−2 s−1, showing no significant differences among species. In contrast, E (1.9–3.2 mmol m−2 s−1), gs (162.3–423.1 mmol m−2 s−1), and WUE (1.8–2.9 mmol mol−1) differed significantly (p < 0.001) among species. P. flava differed significantly from the other two species, with notably higher gs values, whereas B. arborea showed the highest WUE. Photochemical activity parameters showed no signs of photoinhibition, as Fv/Fm values remained between 0.81 and 0.83 during the study period. Values of J (74.7–75.8 μmol e− m−2 s−1), ΦPSII (0.11–0.14), and qP (0.19–0.22) did not differ among species (Table 3).

Table 3. Physiological traits measured in the field during the wet season for adult trees from TDF.
3.2 Greenhouse experiment
A total of 18 AMF morphospecies were identified in the soils of the TDF of the Macanao Peninsula, which were used to produce the native inoculum applied in the greenhouse experiment. The AMF community was dominated by species of the genus Glomus (nine species), followed by Sclerocystis (three species), and Diversispora, Gigaspora, and Rhizophagus (two species each). The remaining AMF genera were represented by a single species. This composition reflects the naturally occurring AMF community in the study area and was used to inoculate the seedlings of B. arborea, C. mollis, and P. flava.
3.2.1 Effects of AMF inoculation on physiological traits
The effects of AMF inoculation on physiological traits were species-dependent (Supplementary Table S2). The AMF inoculation in B. arborea plants caused a significant increase in A (p=0.0001) and gs (p=0.0409) of 50% and 31%, respectively, compared to the NI plants, with no significant differences in E, Ci, and WUE (Figure 1). A significant species × inoculation interaction was observed for A (p =0.0177), but not for gs, E, Ci, or WUE. B. arborea showed higher A, gs, and E than C. mollis and P. flava. Significant species effects were observed in leaf photochemical activity parameters such as Fv/Fm (p=0.0002), J (p=0.0038), ΦPSII (p=0.0018), and qN among species, while qP was not affected. However, no significant effects of inoculation or species × inoculation interactions were detected for the photochemical variables.

Figure 1. Instantaneous gas exchange parameters in the greenhouse: (A) net photosynthetic rate [A], (B) stomatal conductance [gs], (C) transpiration rate [E], (D) intercellular CO2 concentration [Ci], (E) water use efficiency (WUE), and photochemical parameters: (F) maximum quantum yield [Fv/Fm], (G) electron transport rate [J], (H) relative quantum yield of PSII [ΦPSII], (I) photochemical quenching coefficient [qP], and (J) nonphotochemical quenching coefficient [qN] measured in leaves of B. arborea, C. mollis, and P. flava, in noninoculated (black bars) and AMF-inoculated (white bars) plants. Values are means ± standard error (n=10). Different letters indicate statistical differences at p ≤ 0.05.
Analyzing the A/Ci curves, significant increases in Asat were found in B. arborea and C. mollis plants inoculated with AMF, whereas in P. flava, inoculation had a negative effect on the photosynthetic capacity, decreasing Asat (Figure 2). Inoculation significantly increased Vcmax in B. arborea (p =0.0560, Figure 3), and a significant species × inoculation interaction was observed for Jmax (p=0.0107) and TPU (p=0.0094). For all species, no changes in Γ, Ls (58 µmol mol−1 and 18.33%, respectively), or Lm were observed between treatments, species, or due to the species × inoculation interaction (Figure 3).

Figure 2. Responses of leaf photosynthetic rate (A) to intercellular CO2 concentration (Ci) in leaves of (A) B. arborea, (B) C. mollis, and (C) P. flava, in noninoculated (black circles) and AMF-inoculated (white circles) plants. Values are means ± standard error (n=5), and standard errors are shown when larger than the symbol size.

Figure 3. Effect of inoculation with AMF on (A) Ci-saturated photosynthetic rate [Asat], (B) carboxylation efficiency (CE), (C) CO2 compensation point [Γ], (D) relative stomatal limitation [Ls], (E) maximum carboxylation rate [VCmax], (F) maximum rate of electron transport [Jmax], (G) rate of triose phosphate utilization (TPU), and (H) relative mesophyll limitation [Lm] in the three tree species studied: B. arborea, C. mollis, and P. flava. Noninoculated (black bars) and AMF-inoculated (white bars) plants. Values are means (n=4) ± standard error. Different letters indicate statistical differences at p ≤ 0.05.
Overall, AMF enrichment enhanced photosynthetic performance only in B. arborea, while C. mollis and P. flava showed no consistent improvements, underscoring the species-specific nature of the physiological responses to the symbiosis.
3.2.2 Effects of AMF inoculation on plant growth
AMF inoculation and species significantly influenced most growth variables, while species × inoculation interactions were generally not significant (Supplementary Table S3). Inoculation did not significantly affect the number of leaves, although this trait differed among species (p < 0.0001).
Inoculation with AMF resulted in larger plants and a greater root system overall (Figure 4). AMF inoculation significantly increased root and total biomass in B. arborea (p=0.0217 and p=0.0126, respectively) and shoot biomass and root length in P. flava (p=0.0254 and p=0.0061, respectively). Inoculation did not significantly affect S:R, although species differences were significant (p < 0.0001). The values obtained for the S:R ratio indicate that, in all treatments, the plants showed a greater allocation of biomass to the aboveground part (Figures 4, 5). Species effects were significant for all biomass and root traits, and no significant species × inoculation interactions were detected. B. arborea, C. mollis, and P. flava responded to AMF inoculation, albeit to different degrees, with MRI of 31%, 3%, and 23%, respectively.

Figure 4. Growth of Bulnesia arborea, Caesalpinia mollis, and Piptadenia flava plants after 240 days in the greenhouse with AMF inoculation and without inoculation (control).
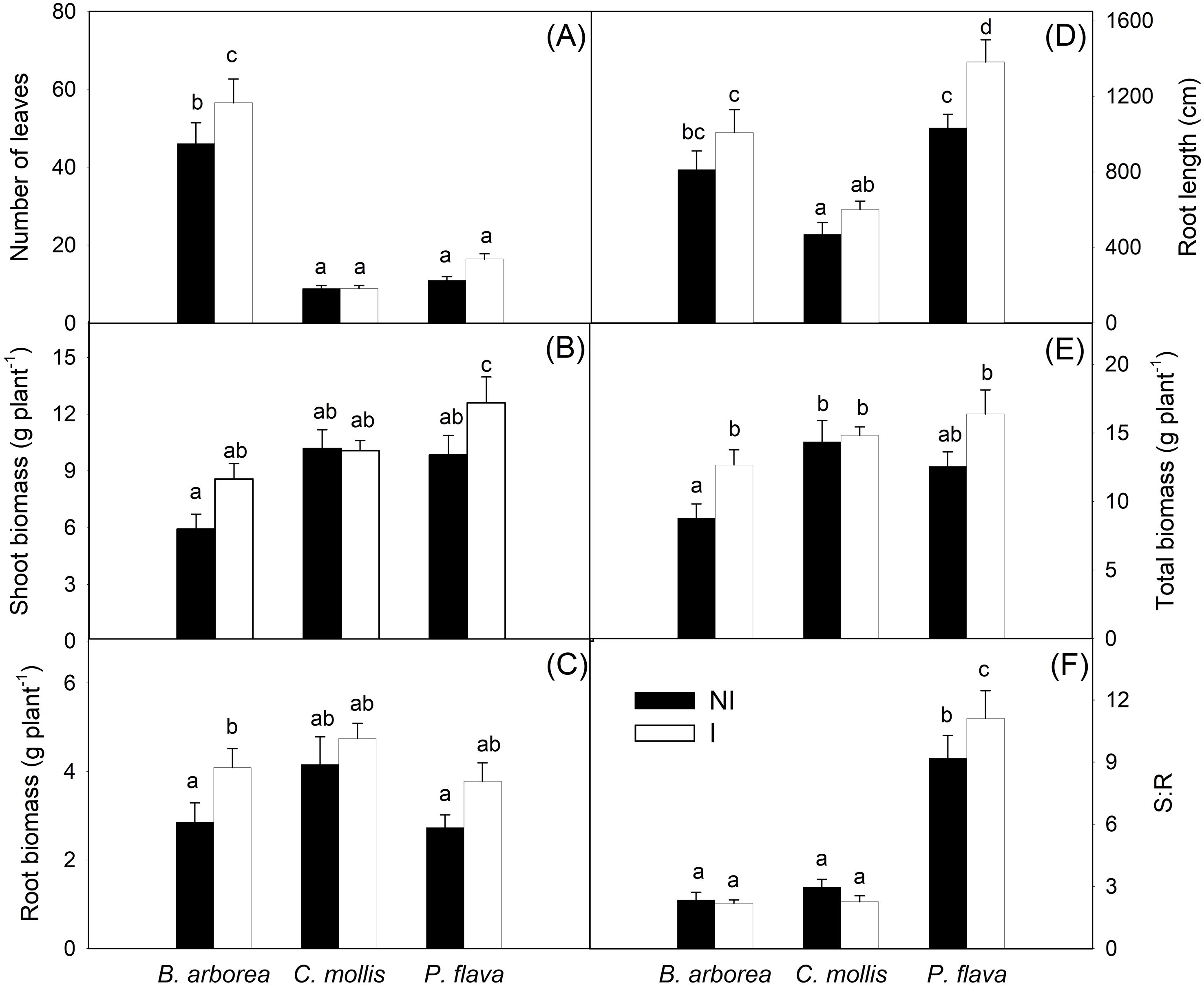
Figure 5. Growth variables measured in B. arborea, C. mollis, and P. flava, in noninoculated (black bars) and AMF-inoculated (white bars) plants, including (A) number of leaves, (B) shoot biomass, (C) root biomass, (D) root length, (E) total biomass, and (F) shoot–root ratio (S:R) (n=10). Values are means ± standard error. Different letters indicate statistical differences at p ≤ 0.05.
Growth responses to AMF enrichment were also species-dependent, with marked biomass increases in B. arborea, partial enhancement in P. flava, and no significant effects in C. mollis, confirming that the magnitude of AMF-induced growth stimulation varied among host species.
4 Discussion
Our results showed that enrichment of the native AMF community via inoculation enhanced certain physiological traits and promoted growth in native plant species, with the most pronounced effects observed in B. arborea. In P. flava, plant growth also increased following AMF inoculation, although no significant differences were detected in its physiological traits, suggesting species-specific functional responses to AMF. These findings highlight the importance of evaluating the compatibility between native AMF consortia and target plant species when designing restoration strategies for disturbed areas. In this context, specific AMF strains may be more effective with certain hosts, depending on their functional traits and ecological adaptations (Wu et al., 2024). Therefore, large-scale restoration programs should consider the use of compatible or locally adapted AMF inoculum, rather than generalist strains, to optimize plant establishment, growth, and ecosystem recovery.
Adult trees evaluated in the field showed significant differences in gas exchange parameters (E, gs, WUE), with the physiological performance of B. arborea, C. mollis, and P. flava being consistent with values previously reported in the same forest during the wet season (Ávila-Lovera et al., 2019). Compared with greenhouse plants, trees in the field likely experienced greater environmental stress, as indicated by the lower A values in B. arborea and C. mollis. The higher light intensity, temperature, and vapor pressure deficit in the field likely imposed stronger evaporative demands, leading to reduced photosynthetic performance compared with the more stable and moderate greenhouse conditions.
The AMF community identified in soils of the Macanao Peninsula TDF shared similarities with those reported for the Caatinga dry forests in Brazil, where Glomus and Gigaspora were likewise common; however, Acaulospora was the predominant genus in those ecosystems (Teixeira-Rios et al., 2018; da Nóbrega Veras et al., 2024). AMF are widely distributed symbiotic fungi, yet many taxa possess physiological and ecological adaptations to specific environments, including semi-arid ecosystems, where factors such as rainfall and soil properties influence AMF community structure (Sousa et al., 2018; da Nóbrega Veras et al., 2024). Interestingly, B. arborea exhibited the highest response to mycorrhization (MRI of 31%), despite belonging to the Zygophyllaceae family, which has previously been described as mycorrhizal-free (Varma, 1999). The high MRI value suggests a functional response to the inoculum, consistent with effective AMF symbiosis. Our findings are consistent with reports of AMF colonization in other Zygophyllaceae species (Ramos-Zapata et al., 2013; Alrajhei et al., 2022; Xu et al., 2023b).
Fabaceae species are known to associate simultaneously with AMF and rhizobia, and previous studies have documented reciprocal influences between these symbionts: AMF infection can increase nodule number and biomass, whereas rhizobia inoculation may reduce AMF hyphal colonization of roots (Larimer et al., 2014). In addition, the mixed inoculum used in this study was likely enriched not only in AMF propagules but also in rhizobia, as V. luteola formed root nodules during trap culture. These combined factors may help explain the comparatively lower mycorrhizal response observed in C. mollis and P. flava, not necessarily reflecting limited AMF colonization capacity, but rather the concurrent contribution of nitrogen-fixing symbiosis or nutrient enrichment from the inoculum to plant growth—aspects that should be considered in future studies aimed at selecting species for restoration.
4.1 Effects of AMF inoculation on physiological traits
Enrichment of the native AMF community through inoculation improved the physiological performance of B. arborea, increasing A and gs, with minor effects on photosynthetic capacity except for Vcmax. Similar responses have been reported in other woody species from semi-arid ecosystems, which exhibited enhanced A, gs, E, and/or WUE following AMF inoculation (Barros et al., 2018; Chen et al., 2020). These improvements likely reflect enhanced water uptake, carboxylation efficiency, and RuBP regeneration, supported by the observed increases in gs, Vcmax, and Jmax. Since all plants were grown under optimal irrigation in the greenhouse, the benefits of AMF enrichment could be even more pronounced under water-limited conditions (Püschel et al., 2021).
The effects of the AMF inoculum were not evident in fluorescence parameters for any of the plant species studied, suggesting that photochemical activity, light use rate, or energy dissipation as heat were unaffected by AMF enrichment. B. arborea and C. mollis maintained Fv/Fm values near the optimal range for healthy plants (~ 0.83; sensu Maxwell and Johnson, 2000), while P. flava showed lower values, suggesting partial PSII impairment. These results are consistent with reports for other semi-arid shrub species associated with AMF (Martínez-García et al., 2011; Yang et al., 2022; Bertolazi et al., 2025).
Photosynthetic limitations often arise from constraints in RubisCO carboxylation (Vcmax) and electron transport (Jmax; Farquhar et al., 1980; Amthor, 1995). In this study, B. arborea plants subjected to AMF enrichment through inoculation showed increased A and Asat, as well as Vcmax, indicating that AMF promoted photosynthesis by enhancing RuBisCO carboxylation capacity. Similar effects have been reported in other species, where AMF inoculation improved RubisCO activity and photosynthetic efficiency (Tezara et al., 2014; Chen et al., 2017). Increases in Asat have also been reported in plants inoculated with AMF, where the symbiosis alleviated photosynthetic limitations caused by biochar-induced reductions in N and P availability through enhanced Vcmax and Jmax (Yang et al., 2022).
4.2 Effects of AMF inoculation on plant growth
The significant increase in B. arborea growth after AMF enrichment through inoculation could be associated with improved nutrient uptake and assimilation through enhanced root biomass and photosynthetic activity (Khan et al., 2022). This increase in root biomass, combined with the extensive extra-root mycelium of AMF, allows the inoculated plants to extract more water and nutrients (mainly P) from the soil for subsequent translocation to the aboveground parts (White, 2019). In P. flava, some growth variables also increased following AMF enrichment, but these changes cannot be explained by the physiological traits measured here, as no changes in photosynthetic performance were detected. The enhancement in shoot biomass and root length, without changes in leaf number, root biomass, or total biomass, suggests that the symbiosis influenced specific growth components rather than overall carbon gain. This apparent decoupling between photosynthesis and growth could reflect early resource allocation to structural development or the carbon costs of sustaining the fungal partner, which can offset potential benefits to photosynthetic activity. Overall, these species-specific responses highlight the complexity of AMF–plant interactions and their dependence on the functional traits of both partners.
Beyond improving plant physiological performance and growth, enrichment of the native AMF community can exert long-term effects on soil recovery and microbial community dynamics. Through glomalin production and hyphal network formation, AMF contribute to soil aggregation, carbon sequestration, and nutrient cycling, enhancing soil stability in degraded dry ecosystems (Wu and Zou, 2017; Li et al., 2024). These processes favor the establishment of other beneficial soil microorganisms, promoting a more complex and resilient rhizosphere community (Boyno et al., 2025). Although this study was limited to 8 months under greenhouse conditions, the sustained physiological and growth responses observed suggest that AMF enrichment could have lasting benefits if maintained under field conditions. The use of native AMF inoculum, as applied here, may thus accelerate the reestablishment of soil biological functions disrupted by mining activities, ultimately improving soil health and supporting ecosystem resilience. Future research should include long-term monitoring of soil structure and fertility parameters, such as glomalin content, aggregate stability, and nutrient availability, together with microbial and biochemical analyses, to evaluate the persistence of AMF effects and their contribution to ecosystem restoration in the dry forests of Macanao and other tropical dry ecosystems worldwide.
5 Conclusion
Bulnesia arborea, C. mollis, and P. flava showed different responses to inoculation both in growth variables and in physiological traits, and it is well documented that these responses vary depending on the species, the inoculum, and the environmental conditions. AMF enrichment improved both growth and physiological performance in B. arborea during the first months of growth, corresponding to the most critical stage of plant establishment and survival, especially in arid and semi-arid ecosystems. Therefore, B. arborea may hold potential for future restoration efforts in the TDFs of the Macanao Peninsula, and further ecological evaluation is required before its selection for restoration programs. In contrast, the AMF enrichment could represent a carbon cost for P. flava, potentially affecting its photosynthetic capacity; however, it would be advisable to conduct experiments under water stress conditions to assess whether inoculation with AMF elicits a positive response when water is limiting. Overall, our results advance the understanding of plant responses to AMF and support the development of restoration strategies through the enrichment of native AMF communities. Our findings, together with evidence from other studies, suggest that the improvement in photosynthetic capacity and growth is not general across species, and species-specific responses and interactions with AMF inoculum are necessary to study to determine with certainty whether this symbiotic relationship is beneficial for plants.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
KC-M: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing, Resources. AC: Conceptualization, Investigation, Methodology, Resources, Supervision, Writing – review & editing. EÁ-L: Investigation, Writing – review & editing, Resources. WT: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial suppor was received for the research, and/or publication of this article.
Acknowledgments
We would like to thank Jenny De Almeida for the help provided in the greenhouse measurements and Carolina Kalinhoff for providing data on the AMF morpho-species present in the rhizosphere of the studied species.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1702917/full#supplementary-material
References
Alrajhei K., Saleh I., and Abu-Dieyeh. M. H. (2022). Biodiversity of arbuscular mycorrhizal fungi in plant roots and rhizosphere soil from different arid land environment of Qatar. Plant Direct 6, e369. doi: 10.1002/pld3.369
Amthor J. S. (1995). Terrestrial higher-plant response to increasing atmospheric [CO2] in relation to the global carbon cycle. Glob. Chang. Biol. 1, 243–274. doi: 10.1111/j.1365-2486.1995.tb00025.x
Ávila-Lovera E., Urich R., Coronel I., and Tezara W. (2019). Seasonal gas exchange and resource-use efficiency in evergreen versus deciduous species from a tropical dry forest. Tree Physiol. 39, 1561–1571. doi: 10.1093/treephys/tpz060
Ávila-Lovera E., Urich R., Coronel I., and Tezara W. (2023). Ecophysiological traits change little along a successional gradient in a tropical dry deciduous woodland from Margarita Island, Venezuela. Front. For. Glob. Change. 6. doi: 10.3389/ffgc.2023.1043574
Banda K., Delgado-Salinas A., Dexter K. G., Linares-Palomino R., Oliveira-Filho A., Prado D., et al. (2016). Plant diversity patterns in neotropical dry forests and their conservation implications. Science 353, 1383–1387. doi: 10.1126/science.aaf5080
Barros V., Frosi G., Santos M., Ramos D. G., Falcão H. M., and Santos M. G. (2018). Arbuscular mycorrhizal fungi improve photosynthetic energy use efficiency and decrease foliar construction cost under recurrent water deficit in woody evergreen species. Plant Physiol. Biochem. 127, 469–477. doi: 10.1016/j.plaphy.2018.04.016
Bertolazi A. A., Passamani L. Z., de Souza S. B., Rodrigues W. P., Campostrini E., Pinto V. B., et al. (2025). Comparative effects of Serendipita indica and a mix of arbuscular mycorrhizal fungi on the growth, photosynthetic capacity, and proteomics of Schinus terebinthifolius Raddi. Planta 261, 34. doi: 10.1007/s00425-025-04608-1
Bever J. (1994). Feedback between plants and their soil communities in an old field community. Ecology 75, 1965–1977. doi: 10.2307/1941601
Boyno G., Rezaee Danesh Y., Çevik R., Teniz N., Demir S., Durak E. D., et al. (2025). Synergistic benefits of AMF: development of sustainable plant defense system. Front. Microbiol. 16. doi: 10.3389/fmicb.2025.1551956
Cáceres A., Kalinhoff C., and Cáceres-Mago K. (2014). Experiencias de inoculación con micorrizas arbusculares (MA) nativas sobre el crecimiento y sobrevivencia de algunas especies arbóreas del bosque seco tropical y matorral xerófito. Experimentia 4, 47–53.
Chambi-Legoas R., Ortega Rodriguez D. R., Figueiredo de Marques F., Peña Valdeiglesias J., Zevallos Pollito P. A., Marcelo-Peña J. L., et al. (2021). Natural regeneration after gold mining in the Peruvian amazon: implications for restoration of tropical forests. Front. For. Glob. Change. 4. doi: 10.3389/ffgc.2021.594627
Chen W., Meng P., Feng H., and Wang C. (2020). Effects of arbuscular mycorrhizal fungi on growth and physiological performance of Catalpa bungei C.A.Mey. under drought stress. Forests 11, 1117. doi: 10.3390/f11101117
Chen S., Zhao H., Zou C., Li Y., Chen Y., Wang Z., et al. (2017). Combined inoculation with multiple arbuscular mycorrhizal fungi improves growth, nutrient uptake and photosynthesis in cucumber seedlings. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.02516
da Nóbrega Veras J. S., Escobar Costa I. E., Mendes-Alvarenga R. L., dos Santos V. M., Alves da Silva D. K., Alves da Silva J., et al. (2024). Rainfall and soil properties driver the temporal dynamics of arbuscular mycorrhizal fungal assemblages in a seasonally dry tropical forest. Acta Oecol. 123, 104000. doi: 10.1016/j.actao.2024.104000
Fajardo L. (2007). Bases ecológicas para la restauración de bosques secos tropicales de la Península de Macanao. Isla de Margarita. Instituto Venezolano de Investigaciones Científicas (IVIC, San Antonio de los Altos, Miranda (VE.
Fajardo L., Rodríguez J. P., González V., and Briceño-Linares J. M. (2013). Restoration of a degraded tropical dry forest in Macanao, Venezuela. J. Arid Environ. 88, 236–243. doi: 10.1016/j.jaridenv.2012.08.009
Farquhar G. D. and Sharkey T. D. (1982). Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 33, 317–345. doi: 10.1146/annurev.pp.33.060182.001533
Farquhar G. D., von Caemmerer S., and Berry J. A. (1980). A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 149, 78–90. doi: 10.1007/BF00386231
Frosi G., Barros V. A., Oliveira M. T., Santos M., Ramos D. G., Maia L. C., et al. (2016). Symbiosis with AMF and leaf Pi supply increases water deficit tolerance of woody species from seasonal dry tropical forest. J. Plant Physiol. 207, 84–93. doi: 10.1016/j.jplph.2016.11.002
Genty B., Briantais J. M., and Baker N. R. (1989). The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 990, 87–92. doi: 10.1016/S0304-4165(89)80016-9
González V. (2007). La vegetación de la Isla de Margarita y sus interrelaciones con el ambiente físico. Memoria la Fundación La Salle Cienc. Naturales 167, 131–161.
Hernández G., Cuenca G., and García A. (2000). Behavior of arbuscular-mycorrhizal fungi on Vigna luteola growth and its effect on the exchangeable (³²P) phosphorus of soil. Biol. Fertil Soils. 31, 232–236. doi: 10.1007/s003740050650
Huérfano A., Fedón I., and Mostacero J. (2020). Libro Rojo de la Flora Venezolana. Segunda edición (Caracas: Instituto Experimental Jardín Botánico Dr. Tobías Lasser, Universidad Central de Venezuela).
Jacob J. and Lawlor D. W. (1991). Stomatal and mesophyll limitations of photosynthesis in phosphate deficient sunflower, maize and wheat plants. J. Exp. Bot. 42, 1003–1011. doi: 10.1093/jxb/42.8.1003
Kalinhoff C. (2012). Influencia de las micorrizas arbusculares sobre el crecimiento y respuesta a la sequía de Piscidia cartaguenensis Jacq.: Implicaciones en la recuperación de un bosque seco de la península de Macanao, Isla de Margarita. Universidad Central de Venezuela, Caracas (VE.
Khan Y., Shah S., and Tian H. (2022). The roles of arbuscular mycorrhizal fungi in influencing plant nutrients, photosynthesis, and metabolites of cereal crops-A review. Agronomy 12, 2191. doi: 10.3390/agronomy12092191
Krall J. P. and Edwards G. E. (1992). Relationship between photosystem II activity and CO2 fixation in leaves. Physiol. Plant 86, 180–187. doi: 10.1111/j.1399-3054.1992.tb01328.x
Larimer A. L., Clay K., and Bever J. D. (2014). Synergism and context dependency of interactions between arbuscular mycorrhizal fungi and rhizobia with a prairie legume. Ecology 95, 1045–1054. doi: 10.1890/13-0025.1
Li M. Y., Wang W., Mo F., Ren A. T., Wang Z. Y., Zhu Y., et al. (2024). Seven-year long-term inoculation with Funneliformis mosseae increases maize yield and soil carbon storage evidenced by in situ13C-labeling in a dryland. Sci. Total Environ. 944, 173975. doi: 10.1016/j.scitotenv.2024.173975
Martínez-García L. B., Armas C., de Dios Miranda J., Padilla F. M., and Pugnaire F. I. (2011). Shrubs influence arbuscular mycorrhizal fungi communities in a semi-arid environment. Soil Biol. Biochem. 43, 682–689. doi: 10.1016/j.soilbio.2010.12.006
Maxwell K. and Johnson G. N. (2000). Chlorophyll fluorescence—a practical guide. J. Exp. Bot. 51, 659–668. doi: 10.1093/jexbot/51.345.659
Mesa-Sierra N., de la Peña-Domene M., Campo J., and Giardina C. P. (2025). Restoration of tropical dry forest: an analysis of constraints and successes across a highly threatened biome. Front. Environ. Sci. 12. doi: 10.3389/fenvs.2024.1458613
Ministerio de Obras Públicas (MOP) (1972). Normas y especificaciones para los estudios de suelos en la división de edafología. Primera edición (Caracas: Dirección de Recursos Hidráulicos).
Moulatlet G. M., Yacelga N., Rico A., Mora A., Hauser-Davis R. A., Cabrera M., et al. (2023). A systematic review on metal contamination due to mining activities in the Amazon basin and associated environmental hazards. Chemosphere 339, 139700. doi: 10.1016/j.chemosphere.2023.139700
Murphy P. G. and Lugo A. E. (1995). “Dry forests of Central America and the Caribbean,” in Seasonally dry tropical forests. Eds. Bullock S. H., Mooney H. A., and Medina E. (Cambridge University Press, Cambridge), 9–34.
Oyedeji S. (2023). “Plant adaptations in dry tropical biomes: an ecophysiological perspective,” in Ecophysiology of Tropical Plants. Eds. Tripathi S., Bhadouria R., Srivastava P., Singh R., and Devi R.S. (CRC Press, Boca Raton, FL), 3–14. doi: 10.1201/9781003335054
Phillips J. M. and Hayman D. S. (1970). Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc 55, 158–163. doi: 10.1016/S0007-1536(70)80110-3
Püschel D., Bitterlich M., Rydlová J., and Jansa J. (2021). Drought accentuates the role of mycorrhiza in phosphorus uptake. Soil Biol. Biochem. 157, 108243. doi: 10.1016/j.soilbio.2021.108243
Ramos-Zapata J., Marrufo-Zapata D., Guadarrama-Chávez P., Solís-Rodríguez U., and Salinas-Peba L. (2013). Ruderal plants: temporary hosts of arbuscular mycorrhizal fungi in traditional agricultural systems? Trop. Subtrop Agroecosystems 16, 399–406. doi: 10.56369/tsaes.1635
Rani B., Jatttan M., Dhansu P., Madan S., Kumari N., Sharma K. D., et al. (2023). Mycorrhizal symbiosis improved drought resistance in wheat using physiological traits. Cereal Res. Commun. 51, 115–124. doi: 10.1007/s42976-022-00281-2
Rojas-Suárez F. (2014). Conservación de la cotorra cabeciamarilla (Caracas: Fundación Empresas Polar-PROVITA).
Santos E. A., Haro-Carrión X., and Oshun J. (2022). Age-specific and species-specific tree response to seasonal drought in tropical dry forests. Sci. Total Environ. 850, 157908. doi: 10.1016/j.scitotenv.2022.157908
Schenck N. C. and Pérez Y. (1990). Manual for the Identification of VA Mycorrhizal Fungi (Gainesville: Synergistic Publications).
Shahzad K., Ali W. S., Muhammad S., Dai J., Zeb U., and Zhu M. (2024). Assessment of plant biodiversity in tropical dry forests of Sialkot, Pakistan; insight into environmental, anthropogenic influence and conservation strategies. Front. For. Glob. Change. 7. doi: 10.3389/ffgc.2024.1362117
Sharkey T. D. (2015). What gas exchange data can tell us about photosynthesis. Plant Cell Environ. 39, 1161–1163. doi: 10.1111/pce.12641
Shukla S., Didwania N., and Choudhary R. (2025). Arbuscular mycorrhizal fungi (AMF): a pathway to sustainable soil health, carbon sequestration, and greenhouse gas mitigation. J. Saudi Soc Agric. Sci. 24, 22. doi: 10.1007/s44447-025-00023-w
Sierraalta D., Cáceres A., Hernández Valencia I., Gajardo R., Cáceres-Mago K., and Rodríguez A. (2021). Efecto de la revegetación con la especie nativa Piscidia carthagenensis Jacq. sobre la calidad del suelo en un matorral xerófito intervenido para la extracción de arena. Multequina 30, 143–156.
Sieverding E., Friedrichsen J., and Suden W. (1991). Vesicular-arbuscular mycorrhiza management in tropical agrosystems (Eschborn: TZ-Verlagsgesellschaft).
Sousa N. M., Veresoglou S. D., Oehl F., Rillig M. C., and Maia L. C. (2018). Predictors of arbuscular mycorrhizal fungal communities in the Brazilian tropical dry forest. Microb. Ecol. 75, 447–458. doi: 10.1007/s00248-017-1042-7
Teixeira-Rios T., Alves da Silva D. K., Tomio Goto B., and Yano-Melo A. M. (2018). Seasonal differences in arbuscular mycorrhizal fungal communities in two woody species dominating semiarid caatinga forests. Folia Geobot. 53, 191–200. doi: 10.1007/s12224-018-9314-7
Tezara W., Kalinhoff C., Marín O., and Cáceres A. (2014). Efecto de las micorrizas arbusculares sobre las limitaciones estomáticas y no-estomáticas de la fotosíntesis de Piscidia carthagenensis creciendo en un suelo degradado de un matorral xerofítico tropical. Experimentia 4, 69–74.
Tezara W., Martínez D., Rengifo E., and Herrera A. (2003). Photosynthetic responses of the tropical spiny shrub Lycium nodosum (Solanaceae) to drought, soil salinity and saline spray. Ann. Bot. 92, 757–765. doi: 10.1093/aob/mcg199
Varma A. (1999). “Functions and Application of Arbuscular Mycorrhizal Fungi in Arid and Semi-Arid Soils,” in Mycorrhiza. Eds. Varma A. and Hock B. (Springer, Berlin, Heidelberg), 521–556. doi: 10.1007/978-3-662-03779-9_22
Wahab A., Muhammad M., Munir A., Abdi G., Zaman W., Ayaz A., et al. (2023). Role of arbuscular mycorrhizal fungi in regulating growth, enhancing productivity, and potentially influencing ecosystems under abiotic and biotic stresses. Plants 12, 3102. doi: 10.3390/plants12173102
White P. J. (2019). Root traits benefitting crop production in environments with limited water and nutrient availability. Ann. Bot. 124, 883–890. doi: 10.1093/aob/mcz162
Wu Y., Chen C., and Wang G. (2024). Inoculation with arbuscular mycorrhizal fungi improves plant biomass and nitrogen and phosphorus nutrients: a meta-analysis. BMC Plant Biol. 24, 960. doi: 10.1186/s12870-024-05638-9
Wu Q. S. and Zou Y. N. (2017). “Arbuscular Mycorrhizal Fungi and Tolerance of Drought Stress in Plants,” in Arbuscular Mycorrhizas and Stress Tolerance of Plants. Ed. Wu Q. S. (Springer, Singapore), 25–41. doi: 10.1007/978-981-10-4115-0_2
Xu D., Li X., Chen J., and Li J. (2023). Research progress of soil and vegetation restoration technology in open-pit coal mine: a review. Agriculture 13, 226. doi: 10.3390/agriculture13020226
Xu D., Yu X., Chen J., Liu H., Zheng Y., Qu H., et al. (2023b). Arbuscular mycorrhizae fungi diversity in the root–rhizosphere–soil of Tetraena mongolica, Sarcozygium xanthoxylon, and Nitraria tangutorum Bobr in Western Ordos, China. Agronomy 13, 1485. doi: 10.3390/agronomy13061485
Yang Q., Ravnskov S., Pullens J. W. M., and Andersen M. N. (2022). Interactions between biochar, arbuscular mycorrhizal fungi and photosynthetic processes in potato (Solanum tuberosum L.). Sci. Total Environ. 816, 151649. doi: 10.1016/j.scitotenv.2021.151649
Keywords: gas exchange, chlorophyll fluorescence, fungal inoculation, photosynthesis, restoration
Citation: Cáceres-Mago K, Cáceres A, Ávila-Lovera E and Tezara W (2025) Effect of arbuscular mycorrhizal fungi on the physiological traits and growth of three woody species from a tropical dry forest. Front. Ecol. Evol. 13:1702917. doi: 10.3389/fevo.2025.1702917
Received: 10 September 2025; Accepted: 24 November 2025; Revised: 18 November 2025;
Published: 10 December 2025.
Edited by:
Giovanbattista Domenico de Dato, Council for Agricultural Research and Agricultural Economy Analysis | CREA, ItalyReviewed by:
Karin E. Groten, Max Planck Institute for Chemical Ecology, GermanyMohammed Bouskout, Cadi Ayyad University, Morocco
Copyright © 2025 Cáceres-Mago, Cáceres, Ávila-Lovera and Tezara. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karla Cáceres-Mago, a2NhY2VyZXNtYWdvQGltYml2LnVuYy5lZHUuYXI=; Wilmer Tezara, d2lsbWVyLnRlemFyYUBjaWVucy51Y3YudmU=; d2lsbWVyLnRlemFyYUB1dGVsdnQuZWR1LmVj