 Peter J. Tschaplinski1*
Peter J. Tschaplinski1* David A. Reid
David A. Reid- 1Aquatic Conservation Science Section, Aquatic Ecosystems Branch, Ministry of Water, Land and Resource Stewardship, Victoria, BC, Canada
- 2Restoration Centre of Expertise, Salmon Enhancement Program, Department of Fisheries and Oceans Canada, Pacific Region, Nanaimo, BC, Canada
- 3Aquifer and Watershed Science Section, Watershed Stewardship and Security Branch, Ministry of Water, Land and Resource Stewardship, Victoria, BC, Canada
- 4BC Ministry of Forests, Victoria, BC, Canada
The Carnation Creek study is located in a 11.2 km2 watershed on southwestern Vancouver Island, British Columbia, in the territory of the Huu-ay-aht First Nations. It was initiated in 1970 as a case study of the effects of forestry practices on watershed biological and physical processes, stream channels, aquatic habitats, and fish populations. The project’s pre-harvest vs. post-harvest-impact observational design was intended to run for no more than 15 years, but continual and unanticipated changes in the responses of the biogeophysical aspects of the watershed to forest harvesting motivated the unprecedented long-term intensive study that continues. Now in year 55, the knowledge gained from this site is relevant not only for informing forestry legislation, regulations, and sustainable practices, but also (1) provides guidance on regenerating stand (second pass) forestry practices in coastal watersheds; (2) allows measurement of the contributions of climate variability to observed changes and the identification of mitigation options; (3) provides science-based guidance for watershed and stream channel restoration; and (4) serves as a logged but unrestored reference watershed for restoration effectiveness evaluations. Between 1976 and 1981, 41% of the watershed was harvested. A mixture of beneficial and harmful effects on juvenile coho salmon were initially observed. Between 1976 and 2004, a warmer thermal regime initially increased growth, freshwater survival, and boosted smolt production. Thermal benefits were partly counteracted by accelerated bank erosion, channel widening, loss of in-stream wood, formation of sediment wedges, and increased streambed scour and sediment transport downstream. Over the longer term, riparian-related impacts were greatly amplified and overwhelmed by the forestry-driven landslide effects that began in 1984. Landslide impacts occurring in the headwaters took more than 20 years to propagate downstream. In the absence of habitat restoration efforts, stream channel and fish habitat complexity and stability have become greatly reduced with accelerated bedload transport, deprived riparian wood input, and loss of large logjams. These delayed effects have reversed short-term, logging-related thermal benefits for coho salmon, with major declines in fry-to-smolt survival and smolt production. The results show that forestry-related impacts may take decades to fully develop and may continue to increase for decades after harvest.
1 Introduction
The Carnation Creek Watershed Experiment is a long-term, multi-disciplinary case study of the effects of historic and current forestry practices on a small salmon-bearing, coastal watershed. Located in southwestern Vancouver Island British Columbia (B.C.)., Carnation Creek is a small stream; approximately 7.8 km long that drains an area of approximately 11.2 km2 (Figure 1). Initiated in 1970, the investigation is now in its 55th year of continuous research and monitoring and has generated the longest time series of continuous data on fish-forestry interactions at the watershed scale available anywhere.
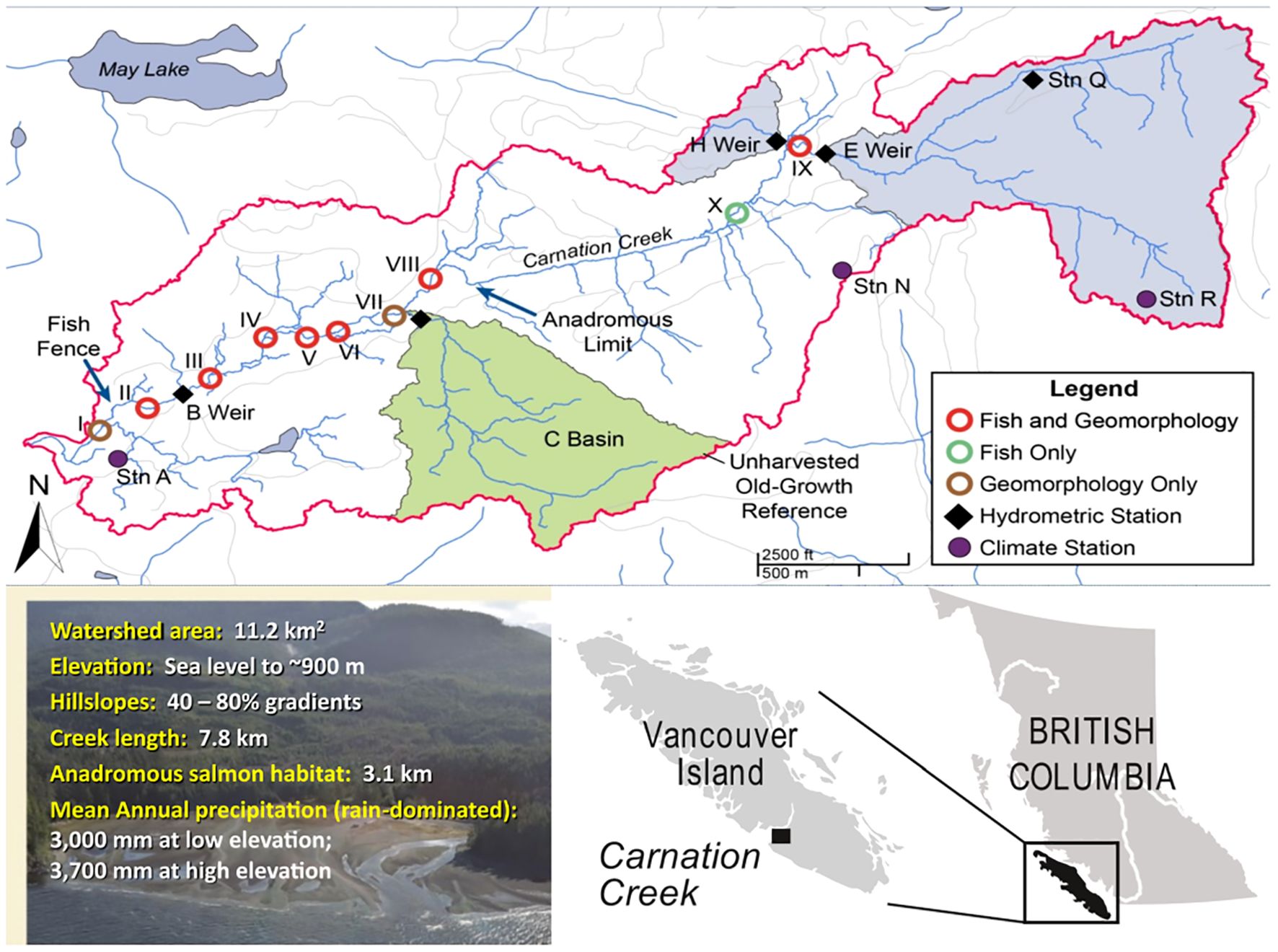
Figure 1. Location of the Carnation Creek watershed in southwestern Vancouver Island showing the watershed outline (red line), locations of permanent fish and channel morphology survey sections, hydrometric and climate (weather) monitoring stations, anadromous salmon distribution, and unharvested old-growth reference subbasin (shaded in green). Blue shaded areas corresponded to gauged sub-catchments.
In the 1960s, fisheries and forestry interests were more frequently coming into conflict in British Columbia. Natural resource managers in B.C. had to base judgments of forest practices effects on fish populations from studies conducted outside of the province; for example, in Oregon, Alaska, and as far away as New Hampshire, all in the United States of America (Tschaplinski and Pike, 2017). The forestry industry and government resource agencies all expressed concern that extrapolations might not lead to optimal planning decisions and practices for areas on the west coast of British Columbia. Therefore, the Carnation Creek study was initiated to provide fish–forestry interaction information on at least one type of drainage basin in coastal B.C.
The initial objectives of this study were to:
1. provide an understanding of the physical and biological processes operating within a coastal watershed;
2. reveal how the forest‐harvesting practices employed in the 1970s and early 1980s affected these processes; and,
3. apply the results to make reasonable and useful decisions concerning land‐use management, fish, and aquatic habitat protection.
The project has evolved with the knowledge gained to focus on quantifying the long‐term alterations and recovery of biological and physical watershed processes as the regenerating forest grows. Research continues to document the following processes and characteristics:
1. hydrologic/climatic regimes (precipitation, forest canopy interception, throughfall, stemflow, and streamflow);
2. hillslopes (mass wasting);
3. riparian forests (canopy closure and shade, streambank stability, and large wood supply to the channel);
4. stream channel morphology and responses;
5. in-stream wood dynamics;
6. aquatic habitats (e.g., fish habitats in the main channel, tributaries, and non-channel aquatic features in the valley bottom);
7. water temperatures; and
8. Pacific salmon and trout abundance, distribution, growth, age structure, survival, and smolt production.
Key questions include:
1. the length of time that observed physical and biological impacts persist;
2. whether all forestry‐related alterations are immediate (e.g., within the first 5 years post-harvest), or whether some are delayed (e.g., two decades or longer);
3. the timeline of post-harvest watershed and stream channel process recovery, and whether recovery resembles pre-harvest conditions or other states;
4. whether streams can recover their structural complexity and aquatic habitat diversity in the absence of restoration within the time prior to the second forest‐harvesting activities (potentially beginning between 2055 and 2065);
5. the potential implications for watershed and stream channel restoration prior to the next forest harvest based on the state and trajectory of biophysical recovery;
6. the contributions of climate change and variability to observed changes; and
7. the potential implications for second‐pass forestry practices in coastal watersheds with alterations or impacts potentially remaining from the first harvest.
The study has an intensive, pre-treatment vs. post-treatment-impact design consisting of five to six years of pre-harvest baseline data (1970-71 – 1975), six years of during-logging measurements when 41% of the basin was actively being harvested (1976 – 1981), and 44 years of post-harvest study which continues to the present (2025). The design incorporates clearcut and unharvested reference (control) sub-basins, and riparian forestry treatments along the mainstem that vary from intensive clearcutting to a variable-width riparian buffer. Main study components include the collection of data on climate, hydrology, channel geomorphology, fluvial processes, aquatic (salmon) habitat, and salmon populations. Quantifying long-term watershed physical and biological changes and salmonid responses to forest practices by species and life stage are central objectives of the Carnation Creek study.
The key findings and benefits of this long-term, intensive study approach will be illustrated by describing shifts in the dominant physical processes affecting the watershed and its salmonid species over four post-harvest decades.
The patterns in post-harvest watershed conditions, process alterations, and causes would not have been observed if the project been terminated in the mid-1980s as originally planned. Carnation Creek learnings highlight the benefits of long-term intensive watershed research and monitoring. Forestry-related impacts have been shown to potentially take decades to fully develop and at Carnation Creek persist four decades after the main period of forest harvesting. Results from this harvested but unrestored watershed are key for increasing restoration knowledge, informing priorities and actions, and evaluating restoration effectiveness that can be applied to other coastal watersheds. This paper will provide an overview of key findings from the Carnation Creek study, followed by a discussion of lessons learned for watershed management and habitat restoration.
2 Materials and methods
2.1 The study area
Carnation Creek is located approximately 20 km northeast of Bamfield on the south shore of Barkley Sound, in southwestern Vancouver Island (Figure 1; 48°55′24”N 124°57′47”W) in the traditional territory of the Huu-ay-aht First Nation. The watershed occurs within the Coastal Western Hemlock Biogeoclimatic Zone, which spans the west coast of North America from the Olympic peninsula in Washington State to Haida Gwaii and southeast Alaska (Krajina, 1969).
The stream drains an area of approximately 11.2 km2, which consists of rugged terrain from sea level to approximately 900‐m elevation. The valley walls have gradients up to 80%. The mainstream is about 7.8 km long, but only the lowermost 3.1 km extending from the stream mouth to the base of a steep‐gradient canyon is inhabited by anadromous salmon including coho salmon (Oncorhynchus kisutch), chum salmon (Oncorhynchus keta), rainbow (“steelhead”) trout (Oncorhynchus mykiss), and coastal cutthroat trout (Oncorhynchus clarki). Coastrange (Cottus aleuticus) and prickly (Cottus asper) sculpins also inhabit this lowest stream reach, which contains a valley bottom of about 55 ha that is 50–200 m wide. This area includes both perennial and seasonally flowing tributaries, ponds, and depressions used as winter habitat by juvenile coho salmon and coastal cutthroat trout (Brown and Hartman, 1988; Tschaplinski and Hartman, 1983; Tschaplinski and Pike, 2017). Populations of nonmigratory coastal cutthroat trout inhabit about 1.4 km of Carnation Creek upstream of the steep canyon, and about 1 km of Tributary C (C-Basin), an unlogged old-growth reference sub-basin of Carnation Creek (Figure 1).
Adult anadromous salmonids returning from the ocean to Carnation Creek to spawn have historically included 1–4,186 chum salmon, 74–426 coho salmon, ≤12 steelhead trout, and ≤9 cutthroat trout annually. Three other salmon species occasionally occur in small numbers as strays.
Average monthly air temperature at the lower elevations ranges from 4.5°C in December to 15.9°C in August (Figure 2c). Summer maximums are in the low 20’s°C and extremes can reach the high 30’s°C. Winter minimums range for 0 to 5°C with extremes below -10°C occurring only rarely. The upper region of the watershed is 4 to 6°C cooler than the lower elevations.
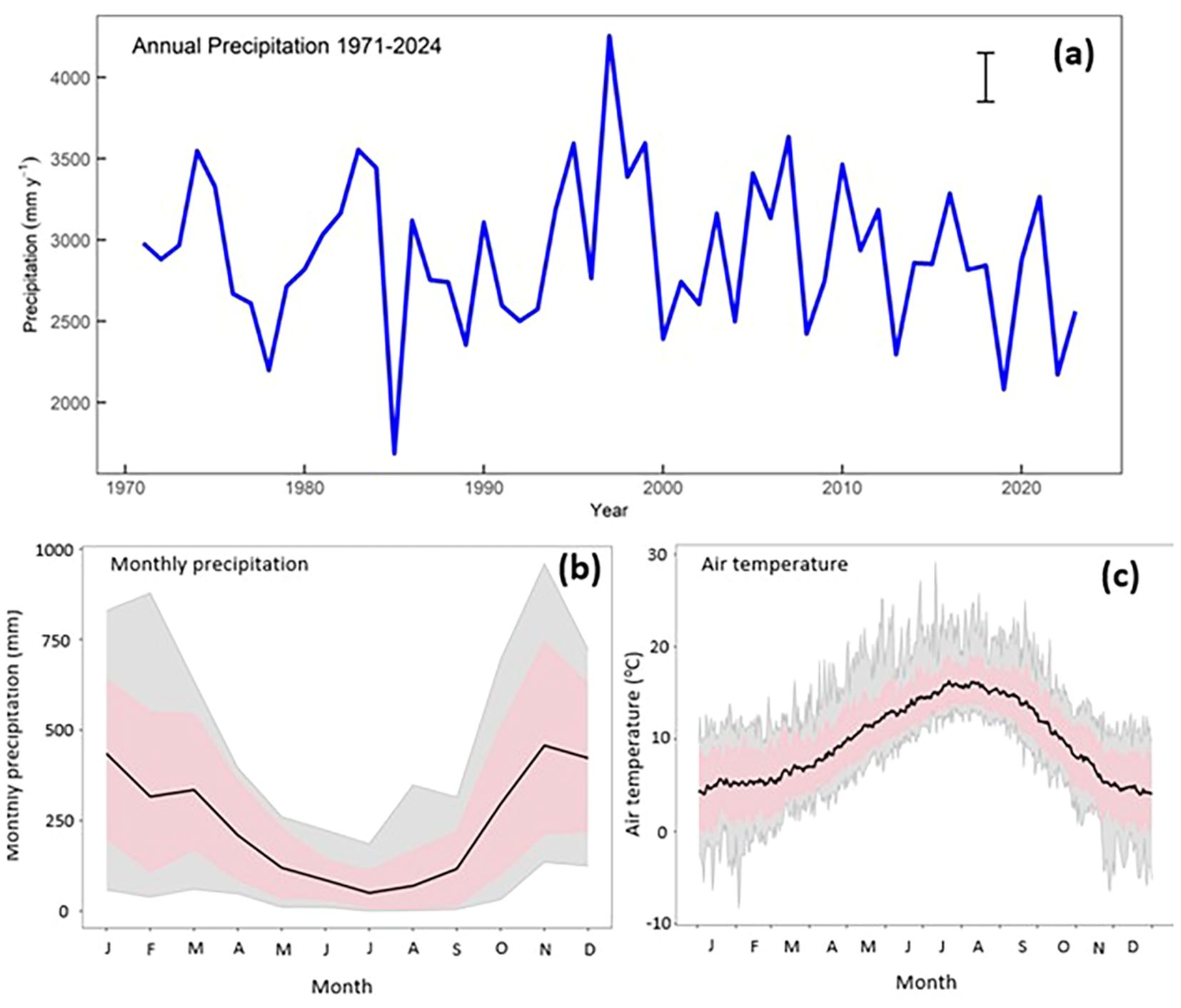
Figure 2. (a) Annual precipitation at station A 1971 to 2024. The bar indicates ±5% uncertainty. Similar inter-annual variability is seen at other locations in the watershed and at Tofino, about 70 km to the west. (b) Mean, maximum, minimum and the 90th and 10th percentiles of monthly precipitation (mm) at station A for 1971 to 2024. (c) Mean, maximum, minimum and the 90th and 10th percentiles of daily average air temperature (°C) at station A for 1971 to 2024. The dashed line corresponds to 0 degrees.
Carnation Creek is a rainfall-dominated watershed; high flows and low flows follow the seasonal cycle of rain (Figure 2a) with characteristic wet winters and dry summers. Mean annual precipitation in the lower part of the watershed is 3,000 mm (Figure 2b) with a range of 1,700 to 4,400 mm. The upper part of the watershed has a mean of 3,700 mm and a range of 2,300 to 5,200 mm. At station A, there is a wide range in monthly precipitation between months and between years ranging from 0 to over 900 mm (Figure 2b).
Streamflow in this system is dominated by rainfall, supplemented by groundwater inputs as well as episodic snowmelt (rain on snow). High flows occur in response to rainfall with some flood events in winter enhanced by melting snow from higher elevation areas. Snow is not a long-lasting feature at Carnation creek with accumulations typically lasting no more than a week or two, generally confined to upper elevations, during the coldest months.
The long-term average mean annual discharge at Carnation creek is 0.801 cubic meters/second (cms, Table 1). Stream discharge at Carnation Creek has varied from 5 to 23.2 cms (annual daily average) with maximum instantaneous flows ranging from 11 to 65 cms. Low flows occur primarily in the summer to early fall periods (Figure 3) but can also occur in the wintertime in absence of rainfall (Table 1). On an annual basis, low flows typically range from 0.001 to 0.091 cms (daily average). Table 1 summarizes the 1973 to 2023 long-term flow statistics for Water Survey Canada (WSC) Hydrometric station 08HB048 Carnation creek at the mouth.
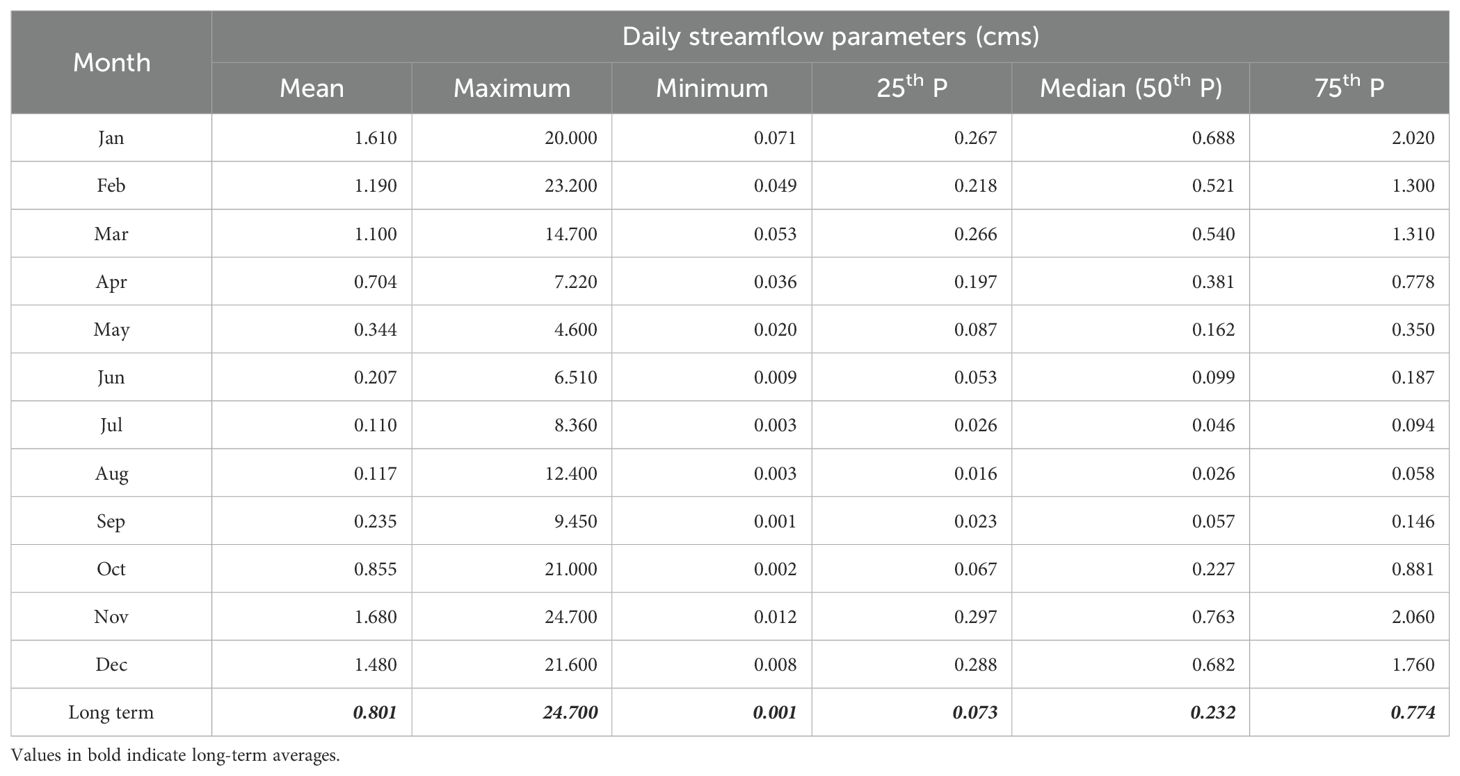
Table 1. Long term streamflow summary statistics, water survey Canada hydrometric station 08HB048 Carnation Creek at the mouth.
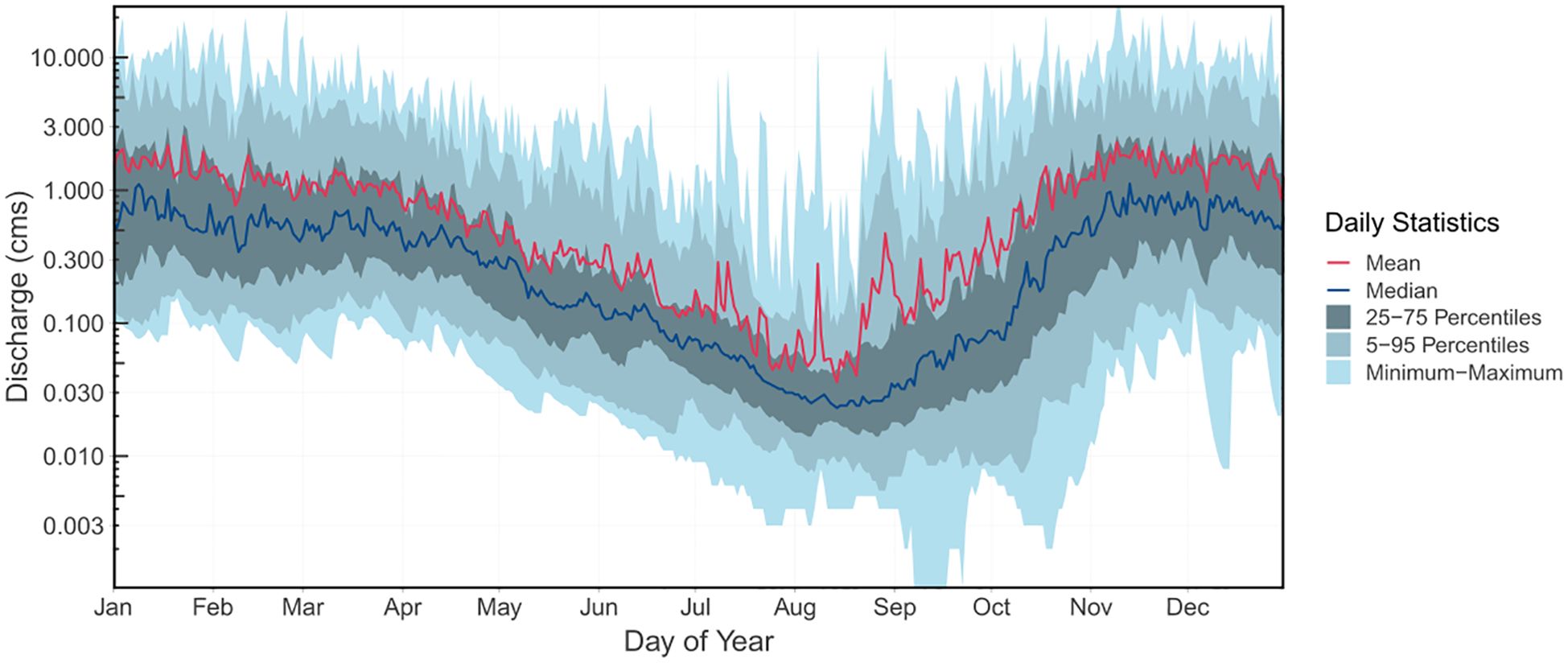
Figure 3. Long term streamflow (daily discharge percentiles) for water survey Canada hydrometric station 08HB048, Carnation Creek at the mouth, 1973-2023. Note the logarithmic y axis.
2.2 Project design and study components
Options originally considered for this fish-forestry investigation included a paired watershed study; however, the Carnation Creek Watershed Experiment was implemented as a single-watershed case study because a second watershed in the same region of comparable size and salmon populations that had not already been subject to forest harvesting could not be found.
There is no external control watershed, but spatial and temporal controls have been available for a variety of study components in Tributary C (C Basin), a sub‐basin more than 1 km2 in area that has remained unharvested for the duration of the investigation (Figure 1).
This study initially focused on the effects of progressive clearcutting (see Figure 4) and three different types of streamside forest‐harvest treatments on stream channels and fish populations. The three riparian treatments applied within the lowermost 3.1 km of the stream were as follows:

Figure 4. Pre-logging view of Carnation Creek in the headwater sub-basin upstream of the anadromous barrier (top left; coastal cutthroat trout habitat) and in the anadromous reach of the stream (top right). Bottom left: seaward view of the lower 3.5 km of Carnation Creek in 1981 after 41% of the watershed was logged at the end of the main phase of forest harvesting. Bottom right: seaward view from a similar vantage point in 2004 after 21–28 years of forest regrowth depending on the age of individual cutblocks.
1. A leave-strip riparian buffer 1–70 m wide from the estuary to 1,300 m upstream;
2. “Intensive” clearcutting along 900 m of stream channel immediately upstream from the leave‐strip treatment (no riparian trees were left, some trees were felled and yarded across the stream, and merchantable wind‐thrown trees were recovered from the stream channel); and,
3. “Careful” clearcutting along 900 m of stream immediately upstream from the intensive treatment area (no activity was permitted in the stream; vegetation on the stream bank such as salmonberry [Rubus spectabilis] was untouched, but red alders were removed).
The study has a pre-logging versus post‐logging observational design organized into three phases:
1. Pre-logging phase, 1970/1971–1975: 5–6 years of baseline data collection depending on salmon species and study component;
2. During‐logging phase, 1976–1981: main forest harvest period of 6 years in which 41% of the total watershed area was harvested; and
3. Post-logging phase, 1982–present: currently tracking 44 years of forestry‐related alterations and recovery processes.
Details of the study design and the methods employed for monitoring physical variables, fish populations, and biological processes before, during, and after forest harvesting have been described in Hartman and Scrivener (1990) and summarized by Tschaplinski (1998, 2000, 2010a), Tschaplinski et al. (2004), and Tschaplinski and Pike (2017).
Data collected historically have included variables on climate; stream temperatures and discharges; groundwater levels; water chemistry; stream channel morphology; large woody debris (LWD) abundance and distribution; streambed particle-size composition; suspended sediment generation and transport during high streamflows; streambed scour and deposition; forestry-related ground disturbance; post-harvest revegetation; biomass of aquatic algae (periphyton); abundance and distribution of benthic macroinvertebrates; and, fish habitats (see Hartman and Scrivener, 1990 for methods). Fish population studies have been focused primarily on salmonids and have included research on spawning and migration patterns; seasonal movements, rearing, growth, and survival of juvenile salmonids; chum egg incubation; egg survival; fry emergence; and, fecundity determinations for female chum and coho salmon for estimates of annual egg-to-fry survival (Tschaplinski, 2010a; Tschaplinski et al., 2004; Tschaplinski and Pike, 2017).
2.2.1 Climate
Continuous data on air temperature, precipitation, relative humidity, solar radiation, and wind speed and direction are currently collected electronically at three stations: Station A near sea level, Station N at 440 m elevation and a high elevation station (Station R at 820 m) installed to study higher elevation climate and monitor transient snow in the hyper-maritime environment (Figure 1). Air temperature and precipitation data are also collected at three streamflow weirs (Weirs C, Q and E – see Figure 1).
2.2.2 Hydrology and hydrologic recovery
Water levels, water temperatures, and supplementary climate data are collected continuously by data loggers at hydrometric stations on both the mainstream (two) and on each of three principal tributaries (Figures 1, 5). Main channel weirs are located at Station B (WSC 08HB048 - Carnation Creek at Mouth, about 450 m upstream from the estuary and near the headwaters at Station E (WSC 08HB069 Carnation Creek at 150m Contour, about 5.4 km upstream). Tributary weirs are established near the mouths of clearcut Tributary H, on the north fork of Carnation (Station Q) and near the outlet of the unharvested C Basin (Tschaplinski, 2010a). Station Q was installed recently to investigate perennial sources and timing of streamflow important to maintaining summer flows in Carnation Creek.

Figure 5. Streamflow weirs and climate stations. Streamflow and water temperature monitoring at main channel weir (Station B; upper panels) and Tributary C (old growth sub-basin, >1 km2; lower left). Climate monitoring at low‐elevation weather Station A near camp (lower center) and at Station N, a mid-elevation site above 300 m (lower right).
The effects of forest harvesting on precipitation interception by the regrowing, post-harvest forest canopy and subsequent changes in throughfall and stemflow have been compared between a second growth stand and reference old growth conditions as one component of hydrological recovery (see Spittlehouse and Maloney, 2023). Precipitation data falling through the forest canopy and down the tree trunks are used to track the progression toward hydrologic recovery as the stand grows and amounts of intercepted precipitation reaching the stream changes over time.
2.2.3 Hillslope processes
Landslide frequency, spatial extent, and connections to the stream channel network have been determined from low altitude aerial photographic surveys performed at 1–3 year intervals. Recently, a detailed aerial orthophotographic survey (2-cm digital imagery) as well as a high density survey by Light (laser) Detection and Ranging (LiDAR) technology (≥ 40 points/m2) has been performed for the entire watershed and surrounding area (approximately 20 km2). The purpose of all surveys is to describe watershed geomorphic features and changes through time, locate and delineate historic landslides, confirm old road networks, and confirm or re-map watershed boundaries. This information is useful to examine the structure of the tributaries to Carnation Creek in terms of channel slope, sediment supply (in-stream and hillslopes), and locations of terrestrial downed wood and in-stream large woody debris (LWD).
2.2.4 Stream channel morphology, erosion and sedimentation processes, and aquatic (fish) habitats
Changes in stream channel structure, physical processes, and fish habitats over time are determined from annual stream channel morphology and fish habitat surveys conducted in 8 permanent study sections that overlap with fish population assessment sites (Figure 1). Pool and riffle habitat areas, channel bed elevations, pool depths, LWD (numbered, measured, and mapped, Figure 6), streambed sediment size composition, gravel bars, overhanging banks, and annual bank erosion and channel scour and deposition processes are measured from geomorphic surveys which have used a range of methods over time (Reid et al., 2019). Channel characteristics are measured to ± 5-cm accuracy. In addition, four surveys have been conducted characterizing channel morphology and in-stream wood along the full length of anadromous fish habitat during the experimental period. These were undertaken in 1991, 1999, 2009, and 2017. Methodological details are outlined in Hartman and Scrivener (1990); Reid et al. (2019) and Reid and Hassan (2020).
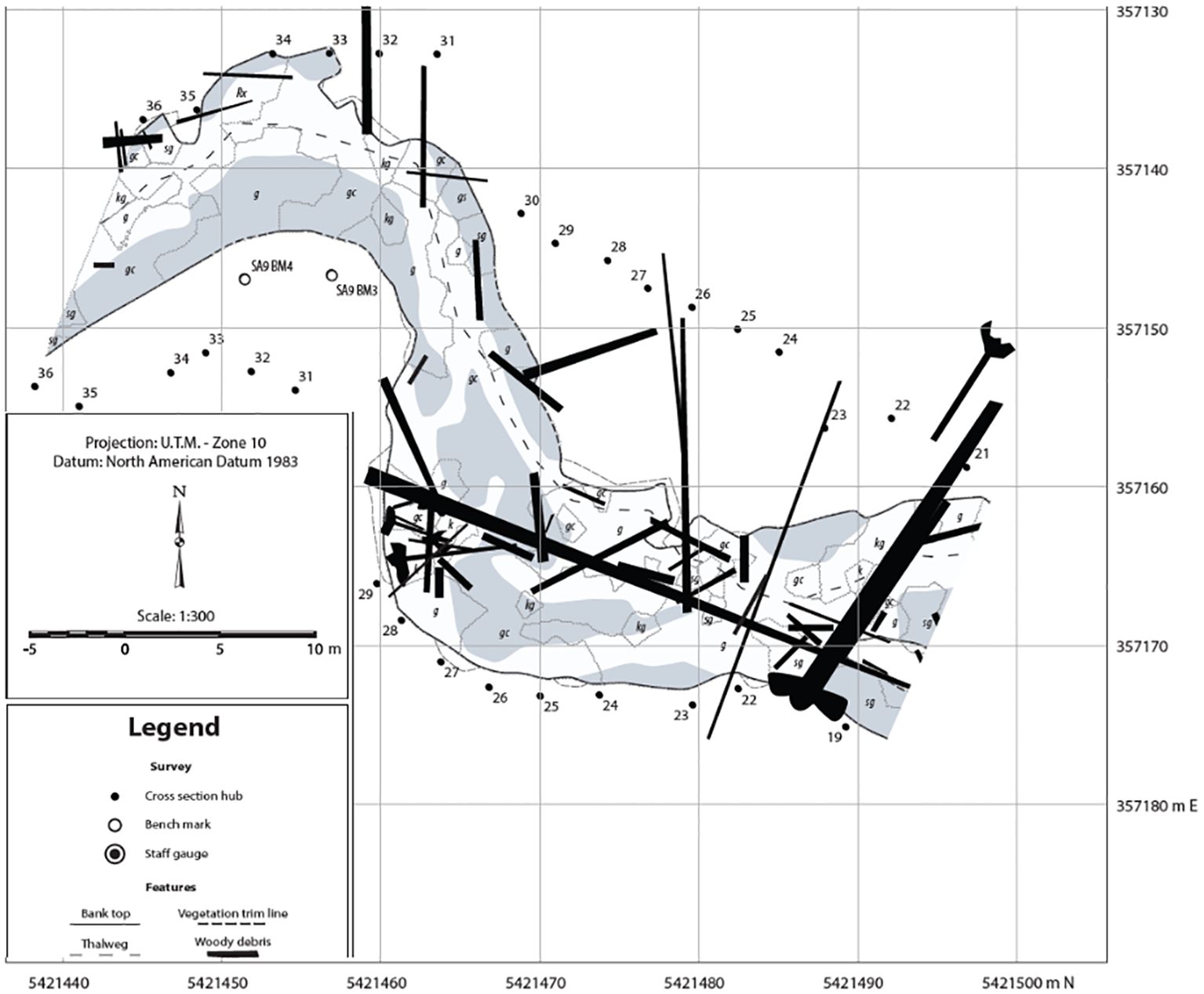
Figure 6. Annual stream channel morphology and aquatic physical habitat surveys are conducted in eight permanent study sections that overlap with fish population assessment sites. An example study area is shown here. Pool and riffle habitat areas, channel bed elevations, pool depths, LWD (logs numbered, measured, and mapped), streambed sediment size composition, gravel bars (shaded areas), and annual bank erosion and channel scour and deposition are measured from topographic surveys.
2.2.5 Fish populations, migrations, and dynamics
Interannual trends in fish populations in Carnation Creek are determined by quantifying migrations between the stream and marine environments at the main fish fence located at the head of tidal influence (see Figures 1, 7), and by assessing the abundance, growth, and survival of juveniles of species rearing in the watershed.

Figure 7. Adult and juvenile salmonid studies. Top left: fish fence configured to intercept seaward‐migrating salmonid smolts and fry (young of the year) in spring (looking upstream). Top right: fence configured to intercept adult spawners entering the stream from the ocean in the autumn (looking downstream). Bottom: summer assessments of populations rearing juvenile salmonids by a two‐catch removal method with electrofishing (left panel) and pole seining (right panel) in one of eight permanent survey sections.
Adult coho and chum salmon returning from the ocean in autumn to spawn in the creek are enumerated at the fish fence, identified to species and sex, and measured for length (± 1 mm) and weight (± 1 g). Scale samples (or alternately otoliths from dead salmon) are collected to determine age and time spent in freshwater and the ocean (Tschaplinski, 2000; Tschaplinski 2010a). During “spring” (mid-February to mid-June) seaward migrating coho salmon smolts, coho salmon fry, and the smolts of steelhead and cutthroat trout are enumerated, measured for length (± 1 mm), weighed (± 0.1 g). Scales are collected from samples of smolts to determine the age distribution of the migrants (see Hartman and Scrivener, 1990; Tschaplinski, 2000).
Populations of juvenile anadromous salmon rearing in fresh water are annually assessed from three replicate surveys performed each summer between mid-June and late September by two-catch removal methods (Seber and LeCren, 1967) employing both electrofishing gear and pole seines (Figure). Assessments are performed in eight standard study sections amounting to about 15% of the anadromous habitat in Carnation Creek. Abundance, age structure, growth and survival during summer are determined from these surveys for juvenile coho salmon, rainbow (steelhead) trout, and cutthroat trout. For each species, the abundance of juveniles in the last summer survey are compared to the numbers of smolts migrating seaward in spring to determine overwinter survival, a key population metric sensitive to habitat quantity and quality (Tschaplinski, 2000; Tschaplinski et al., 2004).
3 Results and discussion
3.1 Short and long-term changes in watershed characteristics and physical processes
While a broad suite of variables have been tracked through time at Carnation Creek, we will focus on changes to fish habitat and fish populations for this document. Carnation Creek research and monitoring has shown that fish population and habitat responses to harvest practices are complex. This complexity has made long‐term trends in anadromous salmon abundance, growth, and survival in fresh water difficult to interpret because direct effects caused by forestry may be confounded by and interwoven with changes in climate, ocean conditions, and fisheries management strategies. In addition, there may be interactions between freshwater and marine processes and corresponding salmon life history phases (Tschaplinski et al., 2004).
Carnation Creek research has identified three broad and interrelated categories of forestry-related effects upon fish and aquatic habitats: (1) physical habitat-structure alterations (Reid et al., 2019, 2020); (2) water temperature-related shifts; and, (3) trophic responses (Tschaplinski, 2000; Tschaplinski et al., 2004; Tschaplinski and Pike, 2017). These categories, separately and in combinations, have had different effects on fish depending upon the species, life stage, and distribution in the watershed. Dominant processes affecting aquatic habitats and fish species have changed over time (Tschaplinski and Pike, 2017). Corresponding to these temporal changes, fish population and habitat responses to harvest practices shifted over the long term. Carnation Creek fish population abundance, distribution, and survival trends over the past 43 post-harvest years were primarily driven by physical habitat and thermal environment changes. During logging and early in the post-harvest period (up to the mid- and late 1980s), both physical habitat impacts and changes in the aquatic thermal environment were largely the consequences of riparian clearcut harvesting. Longer-term stream channel and aquatic habitat changes were initiated and dominated by mass-wasting events (landslides) which are on-going in the present day. Discussion of fish population responses over both the short and long term is focused on coho and chum salmon which are the numerically dominant salmon species in the watershed.
3.1.1 Habitat changes and fish species response
Water temperatures at Carnation Creek increased immediately in response to clearcut riparian harvesting, which was initiated in 1976 and covered 1.8 km of stream length of the 3.1-km-long anadromous zone (58%; Holtby, 1988). Climate-related processes simultaneously contributed to an increase in mean monthly water temperatures in mid-summer (July and August) from approximately 10°C in the pre-harvest period to about 14°C during logging and after. Mean monthly temperatures in summer increased by as much as 5°C after streamside logging began. A mid-summer (August) increment of 3.2°C was attributed to riparian harvest (Hartman and Scrivener, 1990; Holtby, 1988; Holtby and Scrivener, 1989). The remaining portion of the seasonal increase was attributed to climate‐related processes including the Pacific Decadal Oscillation and the El Niño Southern Oscillation (ENSO). Winter temperature increased as well, but by a smaller amount of 0.7 °C in the decade post-harvest (Hartman and Scrivener, 1990).
Holtby (1988) and Holtby and Scrivener (1989) separated the logging‐related and climate variability contributions by (a) first establishing that air temperatures at nearby west coast Vancouver Island weather stations (e.g., Amphitrite Point, Port Alberni, and Estevan Point) were highly correlated with those measured at Carnation Creek, (b) building an air temperature versus water temperature relationship from Carnation Creek data (up to 9 years of pre-harvest data available from a headwater reach), and (c) using the two relationships to predict what water temperatures would have been at Carnation Creek if riparian forest harvesting had not occurred. The difference between the predicted water temperature regime and observed water temperatures in the during‐logging and early post-logging periods was identified as the logging effect.
Historic air temperature records dating back to 1923 for the west coast of Vancouver Island (Estevan Point) were used in similar multiple regression analyses based on Carnation Creek air and water temperature data to generate a back‐cast estimate of water temperatures for Carnation Creek dating back several decades (Holtby, 1988). These results indicated that water temperatures at Carnation Creek prior to the project were in a decade‐scale cycle slightly cooler than the predicted long‐term average (Holtby, 1988, 1988). A gradual return of water temperatures toward the long‐term mean in Carnation Creek was postulated to explain the climate‐related component of increased water temperatures recorded in the mid‐to‐late 1970s.
The effects on chum salmon, which use the freshwater environment primarily for spawning and egg incubation, were concluded to be detrimental to the survival of chum fry produced from the watershed (Holtby, 1988; Holtby and Scrivener, 1989). From a life-history-based multivariate analysis, this effect was estimated to be relatively small when compared to impacts on chum from increased streambed fine sediment accumulations after harvesting (see later discussion). However, warmer temperatures over autumn and winter accelerated the development of chum salmon eggs incubating in the streambed resulting in an earlier hatch, fry emergence from the gravel, and seaward migration of small-sized fry in spring. Smaller body size and early emergence were concluded to be survival disadvantages in the marine environment due to increased susceptibility to predators (Holtby, 1988; Holtby and Scrivener, 1989), as well as early-spring entry into a marine environment when ocean productivity is relatively low resulting in reduced growth and increased mortality early in their ocean life history (Holtby and Scrivener, 1989).
Juvenile coho salmon which spend up to two years rearing in freshwater benefited from increases in Carnation Creek water temperatures, responding with increased growth, freshwater survival, and smolt production (Hartman and Scrivener, 1990). However, Carnation Creek was a notably cold stream before logging. Riparian clearcut harvesting warmed the stream immediately and significantly in all seasons, though temperatures remained below 14°C (Hartman and Scrivener, 1990). The summer mean monthly increase were as much as 3.2°C, but the smaller winter increment of 0.7°C (December) was more ecologically important for juvenile coho salmon (Hartman and Scrivener, 1990). Such “small” thermal changes were nevertheless biologically significant. Fish egg development rates are highly sensitive and positively correlated to water temperatures (Holtby, 1988). Warmer water accelerated coho salmon egg development during autumn and winter such that coho fry emerged on average two weeks early (Holtby, 1988). Earlier emergence allowed for a longer period of summer growth for both fry and yearlings. Coho fry growth increased in response to elevated water temperatures such that fry were 11 mm longer (fork length) on average by the end of their first summer after logging compared with sizes during the pre-logging years (anadromous trout fry also increased in mean length by 18 mm after logging; Hartman and Scrivener, 1990). This larger body size was positively associated with improved overwinter survival after logging (r = 0.91, p <.001; Holtby, 1988). Larger coho were better able to survive winter conditions that included frequent scouring freshets (Tschaplinski and Hartman, 1983). Overwinter survival of juvenile coho increased from 19% in the pre-logging period to 55% in the early post-logging period between 1982 and 1995. The lower numbers of fry rearing in Carnation Creek early in the post-logging period also contributed to increased growth rates due to reductions in density‐dependent competition for food but thermal benefits predominated (Holtby, 1988; Scrivener and Andersen, 1984).
Increased overwinter survival ultimately caused by logging and climate-related increases in water temperatures promoted the production of seaward‐migrating smolts from Carnation Creek, increasing the number of smolts by 65% for nearly three decades (up to 2005) compared to pre-harvest smolt abundance (Figure 8) (Tschaplinski and Pike, 2017).
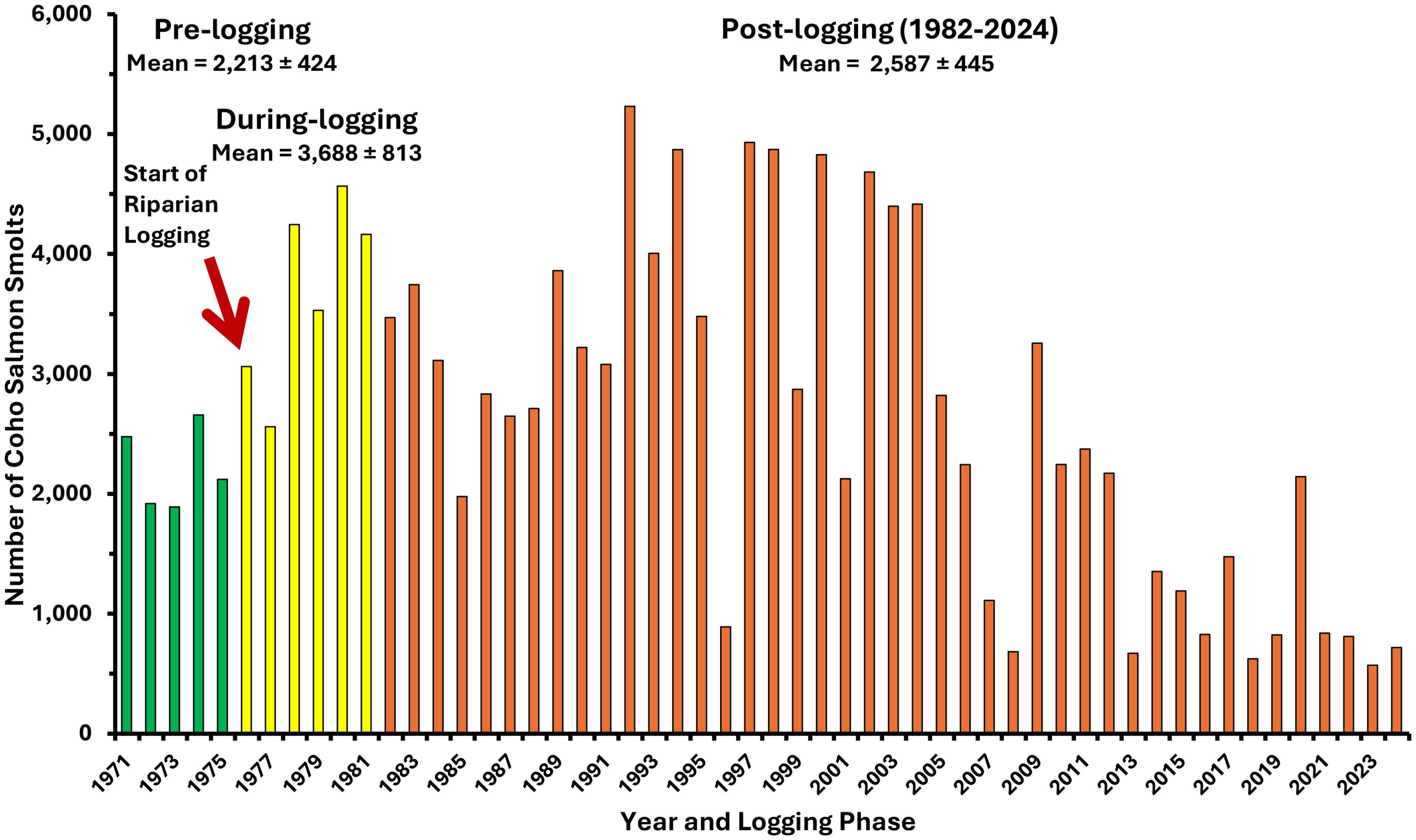
Figure 8. Numbers of coho salmon smolts produced from Carnation Creek from 1971 to 2024. Logging phases are color coded (pre-logging in green, during-logging in yellow, and post-logging in orange). Interannual variation is shown by means provided with ±95% confidence intervals.
In the during-logging period, and early in the post-logging phase (1982–1994), these increased smolt numbers were produced from rearing populations of juvenile coho fry and yearlings that had declined significantly on average to approximately one-half of the abundances observed in pre-logging and during‐logging years (Figure 9; Student’s t test, p <.05). Elevated smolt production from reduced rearing populations appear to have been driven by thermal boosts to overwinter survival due to increased summer growth and larger body size of juveniles entering the winter up to the late 1990s (Figure 10) (Holtby and Scrivener, 1989). Increased water temperatures were able to sustain greater coho salmon smolt production in the short‐to‐medium term until about 2005 (Figure 8).
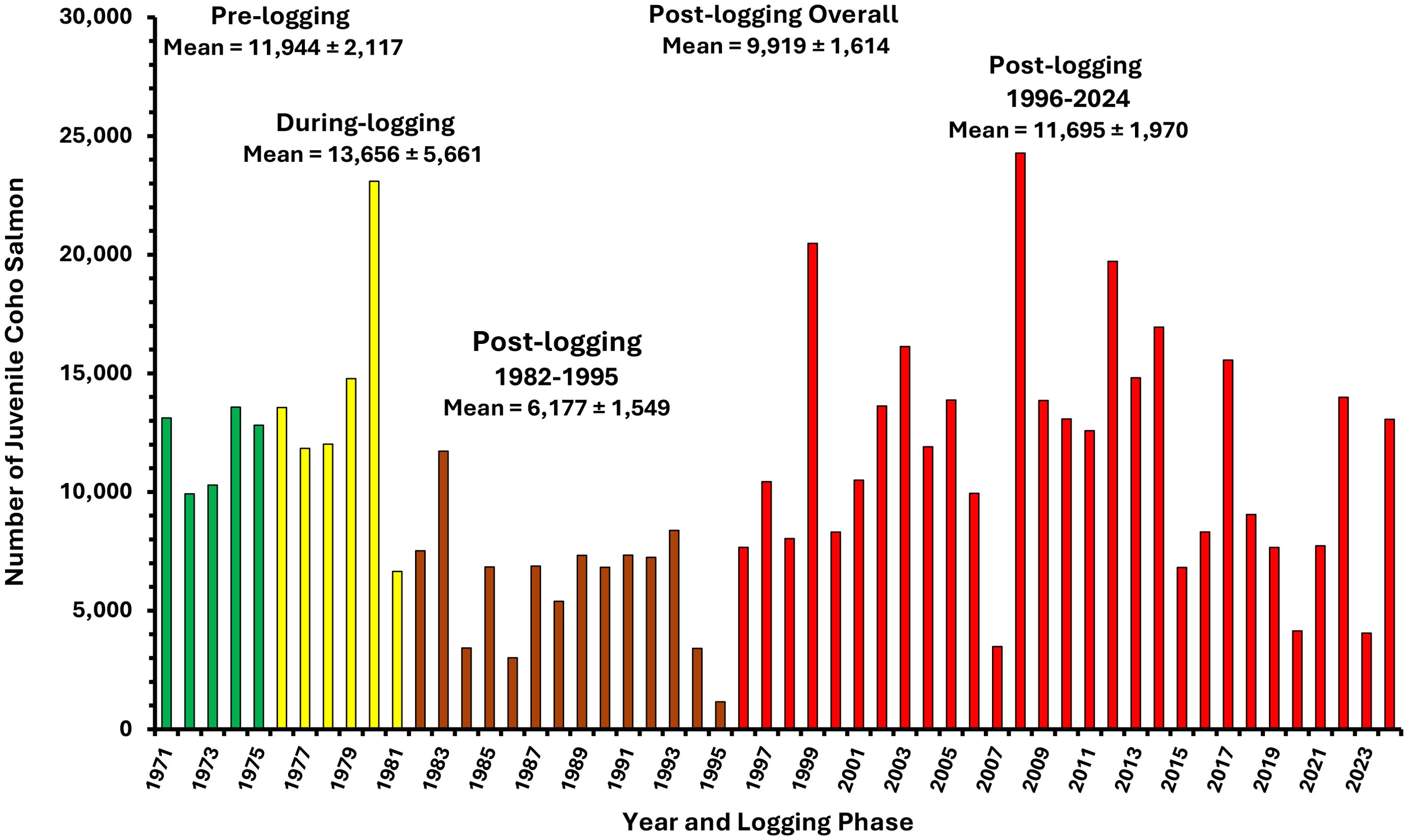
Figure 9. Numbers of juvenile coho salmon (fry and yearlings) rearing in Carnation Creek from 1971 to 2024. Logging phases are color coded as described in Figure 8 with the post-logging period after 1995 shown in red to identify years affecting juveniles after commercial and recreational fisheries were limited or closed. Means are provided with ±95% confidence intervals. Note: time series data are distributed normally (Shapiro–Wilk W = 0.96, p = .09) with no significant first‐order autocorrelation (Durbin Watson 1.54, p = .21).
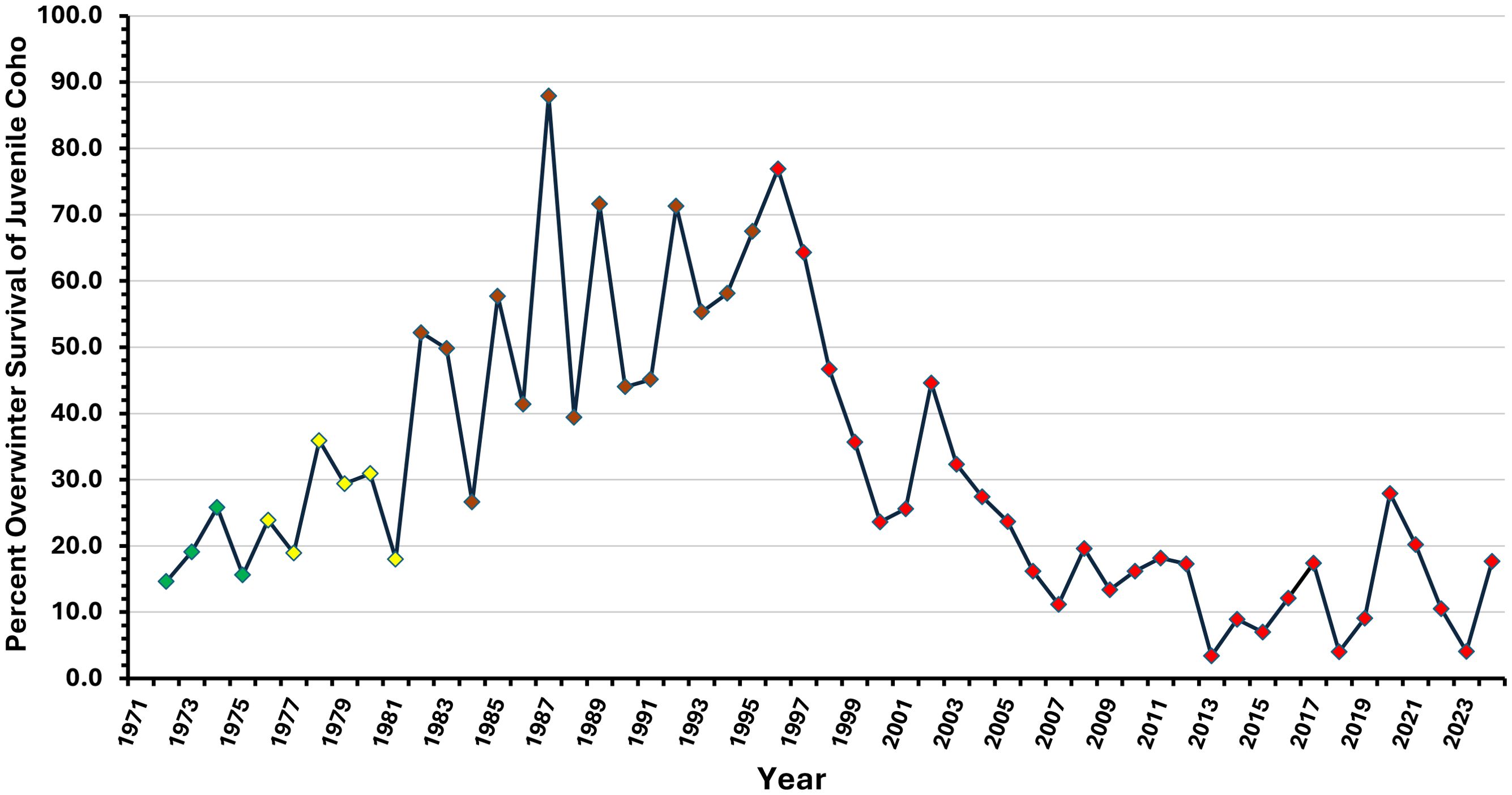
Figure 10. Changes in the percentage of juvenile coho salmon (fry and yearlings) surviving the winter at Carnation Creek from 1971 to 2024 to migrate seaward as smolts in the following spring. Data points are color coded as described in Figure 8 to indicate logging phases and to distinguish post-logging years affecting juveniles after fisheries commercial and recreational fisheries were restricted or closed.
3.1.2 Short-term sediment mobilization and transport in the post-harvest period
Within the first 10–12 years of the post-harvest period at Carnation Creek, changes in the morphology of the stream channel including the textural composition of streambed sediments were primarily caused by increased levels of streambank erosion and streambank collapse in both the “intensively” and “carefully” clearcut riparian treatments (Hartman et al., 1987; Hartman and Scrivener, 1990). During and after logging between 1978 and 1985, bank erosion accelerated in these areas clearcut to the streambanks when streamflows eroded weakened banks where tree stumps had lost rooting strength (Hartman et al., 1987; Scrivener, 1988a; Scrivener 1988b; Scrivener and Brownlee, 1989). Freshets transported eroded materials downstream into the leave-strip treatment including chum salmon spawning areas in the lowermost part of the stream (Hartman and Scrivener, 1990; Scrivener, 1988a).
Frozen core gravel samples taken in chum salmon spawning areas showed strong increases in the amounts of sand and “pea gravel” in the same early post-logging years where egg survival measured for capped chum salmon redds declined from 20% to 11% due to smothering (low oxygen supply) and burial by the excess fines (Scrivener and Brownlee, 1989). This 45% decline in chum egg survival was determined to be the primary cause of forestry-related impacts to chum early in the post-logging period from a life-history-based multivariate analysis by Holtby (1988) and Holtby and Scrivener (1989). Forestry-related impacts alone accounted for 26% of the interannual variation (decline) in adult chum salmon spawner returns (Figure 11).
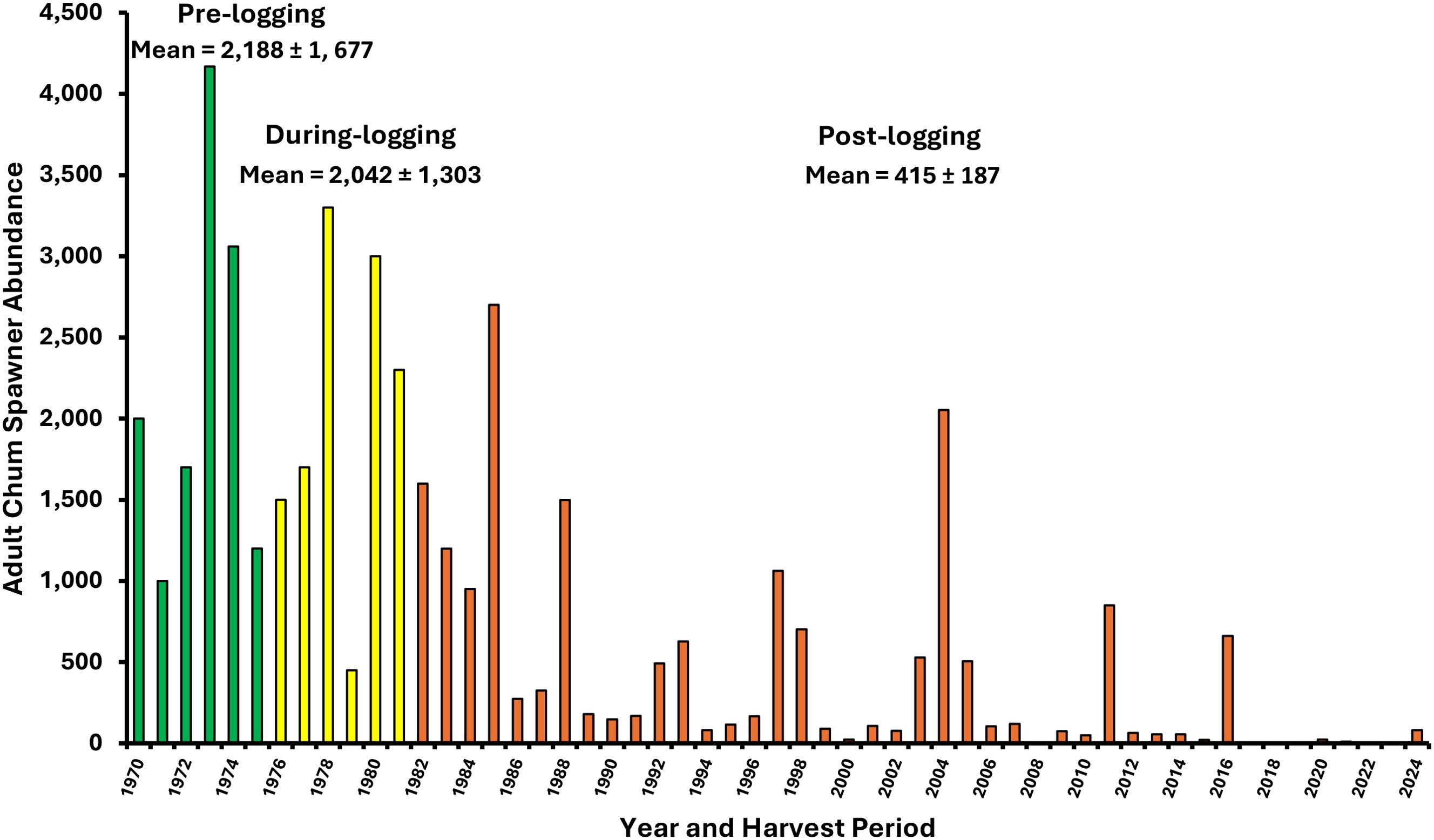
Figure 11. Numbers of adult chum salmon spawners entering Carnation Creek from 1971 to 2024. Logging phases are color coded (in green, during-logging in yellow, and post-logging in orange). Interannual variation is shown by means provided with ±95% confidence intervals.
Climate-related shifts which affected freshwater and ocean conditions (e.g., PDO, ENSO) explained most of the decline in chum salmon by the late 1980s (Holtby and Scrivener, 1989). However, the magnitude of changes in chum salmon abundance at Carnation Creek exceeded what was expected given this species uses freshwater habitats in a comparatively limited way both temporally and spatially; i.e., primarily for spawning and egg incubation. Therefore, these findings were conceptually counterintuitive with expectations at the time.
In contrast with findings for chum salmon, similar analysis for coho salmon showed that less than 10% of the decline in coho spawner returns (Figure 12) were explained by forestry-related effects within the first decade after logging despite coho requiring rearing periods in freshwater of up to two years. For coho salmon, thermal benefits in the short and medium term were counteracted with habitat-related impacts linked primarily to clearcutting which contributed to the reduction of rearing populations of fry and yearlings by nearly one-half (Figure 9). Initiated approximately two years after riparian clearcutting, rates of streambank erosion and streambank collapse increased, which consequently mobilized and transported sediments downstream, destabilized large, embedded in-stream wood, and created a wider, less stable and structurally diverse channel (Hartman and Scrivener, 1990). The life-history-based multivariate analysis for coho salmon trends also showed that the decline in abundance of spawners returning to Carnation Creek was dominated by impacts unrelated to forestry. Marine-related impacts due to climate variations (e.g., El Niño) predominated over both forestry and climate effects on the freshwater environment (Holtby and Scrivener, 1989). Carnation Creek findings have shown that researchers and natural resource managers need to be aware of the cumulative effects on biological and physical processes occurring within both watersheds and marine environments before interpreting observed patterns in anadromous salmonid production.
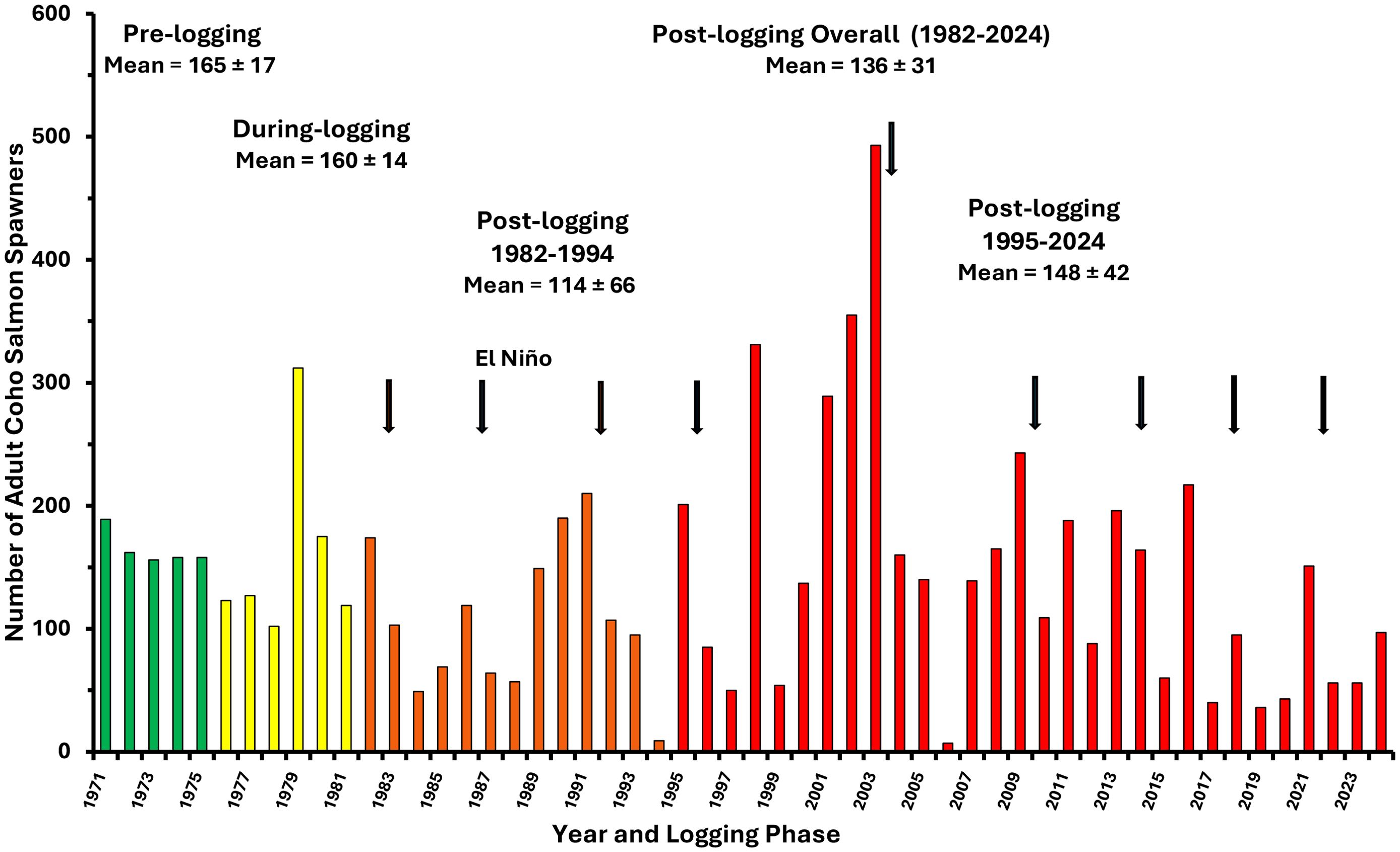
Figure 12. Numbers of adult coho salmon spawners entering Carnation Creek from 1971 to 2024. Logging phases are color coded as described previously with post-logging years when fisheries were restricted or closed shown in red. Interannual variation is shown by means provided with ±95% confidence intervals. Vertical arrows represent El Nino (warm water) conditions off coastal BC unfavorable for salmon growth and survival in the marine environment.
Compared with sediment-related impacts from riparian practices, the contribution of fine inorganic sediments including suspended particles originating from roads was relatively minor, episodic during early summer, and seasonally brief at Carnation Creek (Hartman and Scrivener, 1990). Roads are well known to be significant sources for fine sediments mobilized from their surfaces during rainfall events and delivered by way of ditches to streams at road-stream crossings. The relatively low road-related fine sediment signal at Carnation Creek has been suggested to be a function of operational forestry design. No road-stream crossings were constructed within the anadromous zone of the lower 3.1 km of stream. Only one short section of road entered the valley bottom in the portion of the watershed inhabited by anadromous salmon, and the sole crossing was over a major tributary near its mouth (Tschaplinski, 2000; Tschaplinski et al., 2004). All other roads were located on relatively stable hillslopes on both sides of the basin. Road-caused reductions in the quality of streambed substrates and harmful effects on salmonid egg-to-fry survival were likely minimized due to the lack of road-stream crossings and the short-term peak of fine sediments generated from the construction of valley-bottom roads and stream crossings. This atypical road network remains one of the limitations of the Carnation Creek study design. The chronic introduction of fines into streams from road surfaces and stream crossings is well known to be characteristic of many logged watersheds (Everest et al., 1987; Tschaplinski, 2010b; Tschaplinski and Pike, 2010; Reid et al., 2016).
3.1.3 Long-term changes to channel morphology and aquatic habitats
3.1.3.1 Channel morphology and sediment mobility
Channel and fish habitat changes in Carnation Creek initiated shortly after riparian logging were subsequently magnified from 1984 onward due to landslides that occurred in steep terrain about 1 km upstream of anadromous fish habitat (approximately near the midpoint of the stream). A snowfall followed by a heavy rain in the winter of 1984 resulted in landslides in three adjacent gullies filled with logging slash and crossed by a forestry road located above the gully headwalls (Hartman and Scrivener, 1990). Large volumes of logging debris and inorganic sediments (Figure 13) immediately slid into a confined section of channel. These materials were rapidly transported by high flows to the anadromous portion of Carnation Creek. The resulting partial blockage of the channel caused sediments to accumulate upstream of the logjam (more than 100 m long) and increased the rates of streambank erosion as the stream moved laterally to bypass the debris. Over time, lateral erosion processes widened the channel from twofold to threefold in many locations compared to pre-logging widths, and at the same time, mobilized and transported additional volumes of sediments downstream (Hartman and Scrivener, 1990; Tschaplinski and Pike, 2017; Reid et al., 2019). The progressive collapse of large logjams in the channel and the mobilization of sediment stored behind them has led to elevated sediment mobility and ongoing aggradation in lower reaches of the channel, especially in the last decade (Figure 14).

Figure 13. Landslides and long-term changes in channel characteristics. Clockwise from top left: (1) downstream face of the major logjam near the head of anadromous salmon habitat caused by the winter 1984 landslide that has driven the majority of long‐term channel and aquatic habitat impacts; (2) post-logging channel 1.6 km upstream from the mouth, 2–3 times wider than in pre-logging years with loss of LWD, little riparian LWD source remaining, and unstable sediment wedge and midchannel due to post-slide logjam (behind photographer); (3) and (4) Annual storm transported sediments (> 420 m3) and woody debris stopped at the fish fence near the stream mouth.
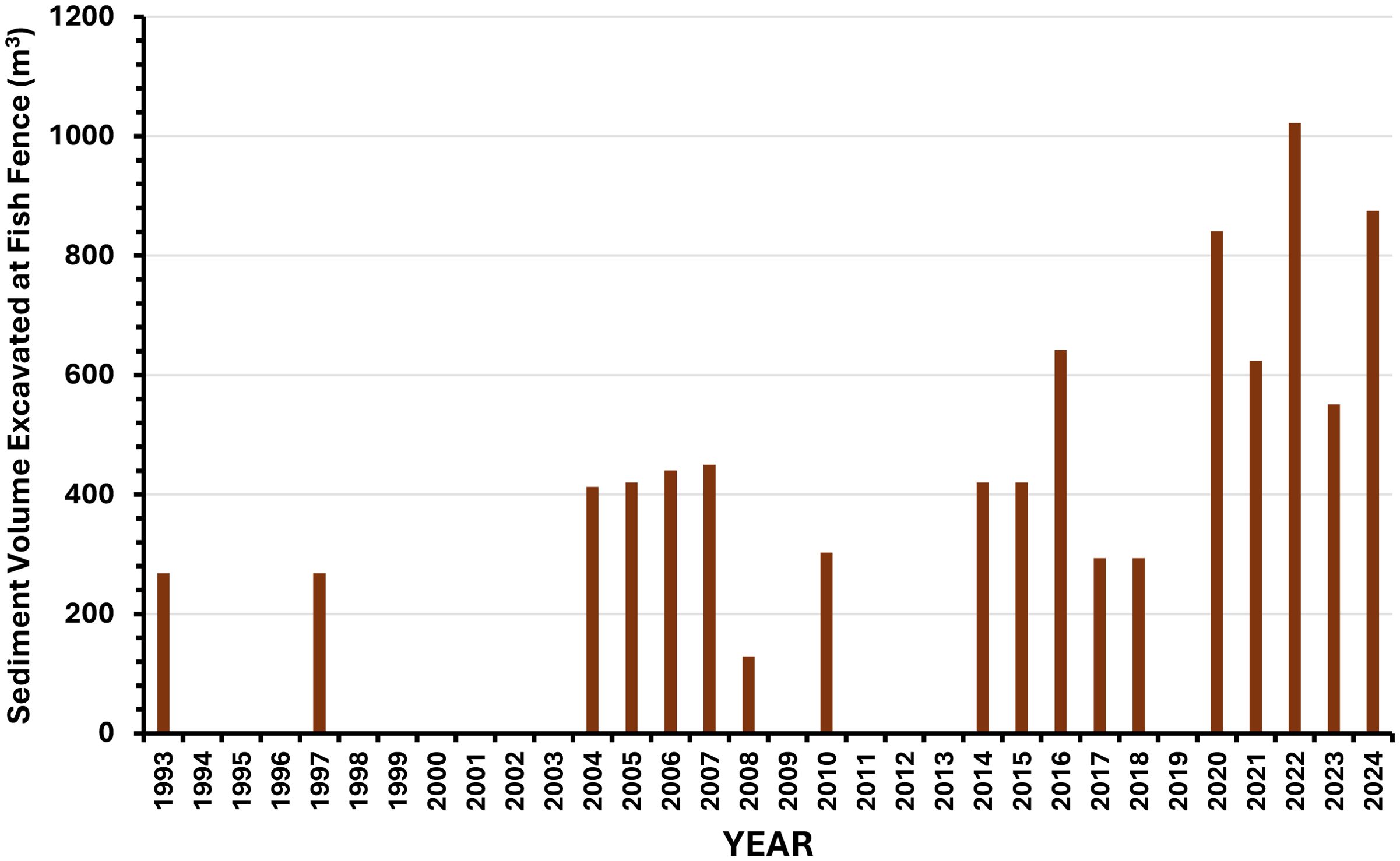
Figure 14. Excess storm-delivered sediments excavated from the fish fence. Since 1993, nearly 8,700 m3 of sediments have been excavated from the footings of the fence and nearby channel in order to operate the facility. The frequency of the excavations and volumes removed have increased over time. Nearly 70% of the total amounts excavated since 1993 have been removed between 2014 and 2024. Coincident with the loss of stable in-stream wood, this indicates on-going and increasing channel instability. No sediment was excavated during years with no bars shown in the figure.
Channel and fish habitat impacts have moved progressively downstream over a period exceeding three decades during the post-logging period. Periodic freshets initiated the breakup and bypass of the original logjams and transported some of the debris downstream to reform into new logjams with associated sediment wedges (Figure 13). Successive storm flows resulted in the destabilization and downstream displacement of embedded LWD. By the 1990s, these processes were augmenting the impacts of riparian clearcut harvesting to impact about one half the length of the stream occupied by coho salmon. After this time, the channel was characterized by shallower pools, long stretches of unstable midchannel gravel bars with elevations that sometimes exceeded those of the streambanks, little embedded LWD to stabilize the streambed and provide habitat structure, and consequently reduced coho salmon egg survival by 45% from 29% survival in the pre-logging period to 16% afterward (Hartman and Scrivener, 1990).
Presently, the entire 3.1-km anadromous portion of Carnation Creek has been morphologically altered by these landslide triggered processes in combination with a change in wood dynamics. Reduced structural integrity and aquatic habitat diversity has occurred due to a lack of functional, stable LWD (Figure 15).
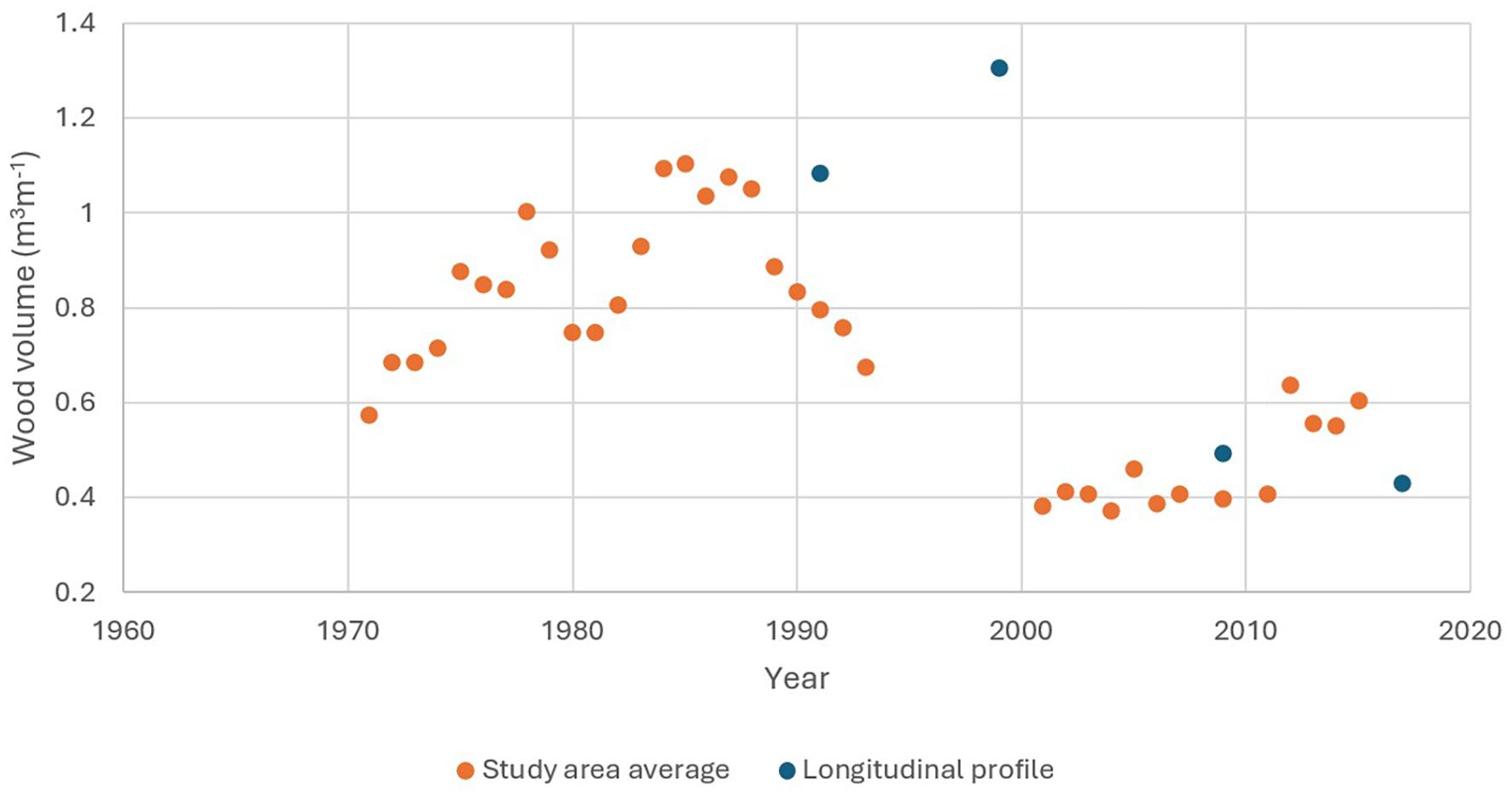
Figure 15. Wood volume scaled to channel length averaged across the eight study sections (orange) and for the spatially continuous surveys from river mouth to the upstream end of anadromous fish habitat (blue). Volume data includes individual wood pieces and logjams.
3.1.3.2 In-stream wood
In most forested streams, in-stream wood plays a critical role in providing habitat complexity and influencing channel morphology (e.g., Wohl and Scott, 2017). The harvesting along the Carnation Creek mainstem led to the elimination of most of the mature streamside trees, and consequently, lead to a reduction in the input of functional wood to the channel.
Data on in-stream wood collected in the eight geomorphic study sections and along the full 3.1 km of anadromous fish habitat reveal dramatic patterns in wood abundance through time (Figure 15). Additions of stable, functional wood to the channel were effectively eliminated following clearcut timber harvesting along 1.8 km of the stream in spite of riparian trees retained along the lowermost 1.3 km of channel within the variable-width riparian buffer treatment. Therefore, the available wood was limited to that already in the channel for most of the stream length. This wood has subsequently decayed, with a loss of large, channel spanning jams (Reid and Hassan, 2020).
At present, the majority of the anadromous zone of Carnation Creek has no supply of mature LWD and will not have a meaningful supply for many decades to come, suggesting a century-scale impact and recovery process (Reid and Hassan, 2020). Recent work integrating hydrological modeling data with wood abundances suggests that this loss of wood is a key driver of rearing habitat loss along Carnation Creek (Reid et al., 2020b). The lack of available future LWD likely signals sustained impacts to the channel, aquatic habitats, and fish production in the absence of mitigative actions through watershed restoration measures.
3.1.4 Cumulative interactions with marine conditions and fisheries management
In addition to the habitat alterations due to riparian practices and landslides, it was apparent that three stressors were reducing coho salmon abundance at Carnation Creek into the mid-1990s: (1) forestry related impacts in the watershed; (2) marine survival declines due to variation in ocean conditions; and (3) both commercial and recreational fisheries (Tschaplinski, 2000; Tschaplinski and Pike, 2017).
Declines in marine survival reduced the numbers of adult coho salmon that returned to freshwater to spawn from pre-logging and during-logging means of 165 and 160 respectively to 114 between 1982 and 1994 (Figure 12). Despite high interannual variation, the downward trend in spawner abundance contributed to reduced abundances of juveniles rearing in freshwater by approximately one-half (6,177) in the early post-logging years compared with means for the pre-logging (11,994) and during-logging (13,656) periods (Figure 9). Ocean survival reductions were marked by declines in spawner numbers for two years or longer after the initiation of El Niño conditions (Figure 12). Several moderate‐to‐strong El Niño events have affected the marine life history phase of Carnation Creek coho salmon throughout the post-logging period. During an El Niño event, warm ocean waters extend northward into BC coastal areas bringing conditions of relatively low biological productivity (impacting salmon food abundance) and abundant predators of young salmon; e.g., Pacific hake and mackerel (Tschaplinski, 2000; Tschaplinski and Pike, 2017).
In 1994, a persistent El Niño coupled together with commercial and recreational fishing pressure and several years of forestry‐related impacts in Carnation Creek reduced the number of adult coho spawners reaching the stream to an all-time observed low of nine salmon; including one female and 8 males (Figure 12). Other coho salmon populations along the west coast of Vancouver Island at the time were also experiencing strong reductions due to similar pressures. To conserve coho salmon stocks on the west coast of Vancouver Island, the federal Department of Fisheries and Oceans restricted the 1995 coho salmon fishery and implemented a full closure in 1998 that lasted several years based on the assumption that coho salmon production was spawner limited (Tschaplinski et al., 2004; Tschaplinski and Pike, 2017). These closures were supplanted in the 2000s with limited fishing opportunities. The purpose of these measures was to increase the numbers of adult coho reaching the spawning grounds to increase freshwater fry abundances and ultimately smolt production.
Fisheries restrictions and closures since 1995 resulted in five of the six highest spawner returns to freshwater over the history of the project; increasing the mean post-logging returns from 114 prior to 1995 to 148 afterward (Figure 12). The desired effect of increased spawner abundance appeared to be achieved at least in the short-term. More spawners increased the abundance of juveniles rearing in freshwater during summer to nearly 11,700 after 1996, approximating abundances prior to logging (Figure 9). However, more spawners and more juveniles in the stream in summer have not resulted in sustained increases in coho salmon smolt production from Carnation Creek through time. Although mean annual smolt production has remained above 2,500 for the post-logging period, strong reductions are apparent from 2005 to the present (Figure 8).
3.1.5 Long-term effects of habitat degradation on overwinter survival and coho salmon smolt production
Smolt production from Carnation Creek has trended downward since 2005 and has fallen below pre-logging smolt numbers in 13 of the past 20 years (Figure 8) although the mean smolt abundance of 1,448 since 2005 is not significantly different from either the overall post-logging mean or post‐1995 mean due to high interannual variability (Figure 8; Student’s t tests, p > 0.05). However, it is expected that declines in smolt production will continue as a consequence of declining amounts and quantity of freshwater habitats. Smolt numbers since 2013 have been less than one half the pre-logging numbers and coincide with the downstream progression of habitat impacts ultimately related to the 1984 landslides. Three decades after their initiation, these impacts now affect the entire 3.1 km of the anadromous salmon habitat at Carnation Creek. These delayed impacts to freshwater rearing habitats show that the short-to-mid-term thermal benefits that boosted smolt production in Carnation Creek have been surpassed by the gradual advances in habitat degradation. This suggests that Carnation Creek cannot return to nor maintain the increase in smolt production observed between 1976 and 2005 (Figure 8) despite spawner numbers and late summer rearing populations of fry and pre-smolt yearlings boosted as a consequence of fisheries management intervention.
Elevated numbers of coho salmon spawners since 1995 have increased the numbers of fry and pre-smolt yearlings entering the winter as shown by recent trends in late summer population abundances (Figure 9). However, recent analysis of the long‐term patterns in freshwater (overwinter) survival of juvenile coho salmon has shown that these enhanced populations have survived the winter in progressively lower proportions after 1995–1996, with historic low survivals being observed in one-half of the years since 2005 (Figure 10).
Trends in annual overwinter survival (survival proportions or “rates”) have been examined on the logit‐scale (logit = log(p/(1 − p) with data transformed to the natural log (Tschaplinski and Pike, 2017). Results of the transformed data showed no evidence of temporal autocorrelation (Durbin–Watson test, d = −0.11, p = 0.53). Segmented regressions (Oosterbaan, 1994) were fitted to the data separately for the pre-logging, during‐logging, early post-logging (1982–1995 with marine fisheries), and later post-logging phases (Tschaplinski and Pike, 2017). Increased overwinter survivals from the pre-logging to the early post-logging phases were observed; however, a strong and statistically significant decline in overwinter survival has occurred since 1996 when compared to the trends observed during the previous phases (Figure 10; p < 0.001). This depression in overwinter survival continues to the present day.
Overwinter mean survival increased to 55% during the early post-logging period compared to both pre-logging (19%) and during‐logging (26%) averages (Student’s t test, p < 0.05; Figure 10); however, the increase over the latter two periods cannot be solely attributed to the effects of logging on the aquatic thermal environment because of the interrelated effects of climatic variability and change simultaneously occurring with forest disturbances. Because both processes promoted better overwinter survival in juvenile coho salmon, the slopes of logit‐scale (segmented) regressions for overwinter survival spanning the pre-logging, during‐logging, and early post-logging segments (0.04 to 0.17) were not significantly different (all p > 0.05). Regardless of the mechanisms promoting increased overwinter survival and smolt production during the first two decades after riparian harvesting was initiated, the subsequent progressive reductions in both population metrics are a key long‐term finding of this research project and strongly show the value of long-term intensive monitoring and research at the watershed scale.
Short‐term, thermally driven survival benefits were eventually reversed by increased levels of physical habitat alterations due to logging-related impacts at Carnation Creek. By 2005, the downstream progression of sediment, wood, and altered channel compositions and morphology due to the landslides of 1984 had extended fully to the Carnation creek estuary. The effect of these alterations has been reflected in continual reductions in overwinter survival currently affecting coho salmon smolt production. The mean overwinter survival for the post-logging period after 1996 was approximately 23% (Figure 10), but survival has declined further to 14 ± 3% (95% CI) since 2005 and has been as low as 4% in recent years (Figure 10).
The simplification of stream habitat and destabilization of the stream channel due to loss of embedded in-stream wood is just one factor influencing the quantity and quality of overwinter shelter available for salmonid species. Both juvenile coho salmon and coastal cutthroat trout have been shown by research at Carnation Creek (and elsewhere) to seek the shelter of valley bottom tributaries and non-fluvial aquatic habitats in the floodplain in autumn to avoid the harmful effects of high seasonal flows in winter when water temperatures may fall to levels just above freezing and inhibit fish activity (Bustard and Narver, 1975; Brown and Hartman, 1988; Tschaplinski and Hartman, 1983). Upward of 23% of coho salmon smolts produced from Carnation Creek in the pre-logging period consisted of fish that used three primary “off-channel” tributaries to survive winter storm flows in winter (Hartman and Scrivener, 1990). During intervals throughout the post-logging period the connectivity of these tributaries with the main channel of Carnation Creek has been interrupted, often blocking fish access and use of these overwintering sites either by excess coarse sediment deposition in the main channel at the tributary junctions or by excess main channel scour at the junctions leaving the tributary mouth perched above the main stream. Since the mid-1990s, one of the three major valley bottom tributaries to Carnation Creek (Tributary 750 located 750 m upstream from tidal habitat) has not been accessible for juvenile coho salmon and coastal cutthroat trout seeking shelter in the winter due to main channel downcutting. Improvements to salmonid productivity through watershed restoration actions clearly need to address lateral drainage network connectivity.
3.1.6 Current state of the Carnation Creek stream channel
Carnation Creek trends show clearly that the impacts of historic forestry practices (i.e., practices implemented prior to the 1995 Forest Practices Code of BC) can be delayed and not fully developed for several decades. Short‐term benefits such as improvement in growth, survival, and smolt production might occur in circumstances where these population parameters may have been limited by pre-logging water temperature regimes. However, these benefits may be eclipsed by habitat degradation over the longer term. At Carnation Creek, these changes in habitat began to mature after three decades following forest harvest.
The entire 3.1 km portion of Carnation Creek occupied by anadromous fish has undergone substantial change post-harvest, with little structural and aquatic habitat diversity remaining due to a lack of LWD (Figure 15). The accelerated processes of lateral erosion responsible for channel widening have also impacted the riparian buffer treatments, resulting in toppling of streambank trees into the channel and negating the intended function of the buffer. In spite of this contribution of LWD to the channel from the riparian buffer, most of the anadromous zone of Carnation Creek has no supply of LWD at present and will not have a meaningful supply for many decades to come. The lack of future LWD has implications for sustained impacts to the channel, aquatic habitats, and fish production.
The channel bed is heavily aggraded with numerous midchannel bars and sediment wedges throughout the lower 3.1 km. This bedload material together with dislodged logging debris, landslide materials and in‐stream LWD is moving downstream in large volumes during major freshets (Figure 14). In most years since 2014, 300–450 m3 of inorganic sediments accumulated on the upstream side of the fish counting fence. To operate the fence, excess sediments delivered from upstream sources are removed periodically from the fence footings and streambed deck. Since 1993, nearly 8,700 m3 of sediments have been excavated. The number of excavations and volumes of materials removed have become more frequent over time. Nearly 70% of the total amounts excavated since 1993 have been removed between 2014 and 2024. Coincident with the loss of stable in-stream wood, this indicates on-going and increasing channel instability propagating downstream in Carnation creek.
These volumes of materials are only a fraction of the bedload transported annually. An unquantified amount passes through the fence to the estuary, which has become increasingly aggraded with bedload sediment, especially over the last two decades. Large volumes of LWD, formerly functioning as important in‐stream structure, also collect at the fish fence as the consequence of major freshets (Figure 13). All of these processes point to habitat alterations that currently reduce the capacity of the stream to support fish species such as coho and chum salmon. The forestry-related impacts continue to have a strong effect on chum salmon (Figure 11), but the causal mechanism has shifted. Impacts due to logging dominated by fine sediment accumulations in egg incubation gravels early in the post-logging period have been superseded by the near annual deposition of large volumes of coarse sediments directly on their spawning beds which include the upper intertidal portion of the stream, reducing the amount of suitable spawning substrate and burying eggs. This on-going process demonstrates that a continued and persistent outcome of an unstable stream channel at Carnation Creek is reduced salmonid production.
The shift in causal mechanisms affecting coho and chum salmon at Carnation Creek demonstrates the strength of the knowledge gained from long-term research projects, clearly demonstrating how conclusions may change through time. In the case of Carnation Creek, it was insufficient to limit the investigations at the discovery of salmonid population decline due to forestry-related impacts. It is essential to know what the specific causal mechanisms related to changes in the physical and biological watershed process have been in order to guide effective science informed watershed rehabilitation actions.
3.1.7 The value of long-term intensive watershed monitoring
Had the Carnation Creek study ended in 1985 as originally planned, conclusions on the effects of forest practices on watershed biological and physical processes, the stream channel network, aquatic habitats, and Pacific salmon would be very different from conclusions made presently after 55 years of research and monitoring. The study has shown that attention must be given not only to changes at the reach or stream habitat scale, but also to the physical and ecological process connections and linkages from the hillslopes to the channel network and how these may change over time and continue to cause further disturbances decades after forest harvest. Not all impacts are immediate, and some take decades to mature and persist for decades longer. Not all watersheds will have experienced what has occurred at Carnation Creek. However, detailed empirical study over the long-term can explain the proximal and ultimate causes of impacts on aquatic values including fish, and reveal that the dominant driving factors including forestry, climate, and fishery management can also change over time. Understanding processes (and disturbance effects and recovery timeline) is one of the most valuable outcomes from intensively monitored watersheds which is transferable broadly.
Changes to the physical environment and aquatic thermal characteristics of the Carnation Creek watershed continue and require on-going investigations focused on both forestry and climate change (i.e., trends in precipitation, air and water temperature). Climate change is playing an increasingly important role (PCIC, 2023). In 2023, the seaward smolt migration was blocked by low streamflows partway through the migration period, an unusual occurrence. Lack of precipitation together with a streambed heavily aggraded with excess sediments from forestry-related long-term changes reduced aquatic habitats to isolated pool refugia. Late summer coho salmon populations were reduced by nearly two-thirds to 4,200 compared with a post-1996 mean of nearly 11,700. Presently, effort continues on quantifying long‐term physical habitat changes from the annual channel morphology assessments and the flow-habitat modelling these assessments support. These analyses represent a rare opportunity to quantify fish species and life stage responses to long-term impacts of historic forestry practices in terms of freshwater population abundance (carrying capacity), distribution, and ultimately smolt production.
4 Concluding remarks
4.1 Implications of this study for watershed management and habitat restoration
Carnation Creek is, in many ways, a prototypical coastal stream in the Pacific Northwest. The history of landscape disturbance is similar to that experienced in a significant proportion of watersheds west of the Cascade Crest. To date, Carnation Creek has experienced no habitat restoration activities and, as a result, provides a valuable reference point against which restoration work in other systems can be compared.
The ongoing disturbance of habitat conditions and fish populations combined with the lack of notable improvement in watershed processes provides a strong case for the value of watershed-scale restoration work. In Carnation Creek, the primary mechanisms driving changes are related to sediment input and loss of both riparian and in-stream wood (Reid and Hassan, 2020). While restoration practitioners have sought to address wood and sediment challenges for decades (e.g., Slaney and Zaldokas, 1997), Carnation Creek findings reinforce the need for a more process-based focus on restoration approaches (e.g., Beechie et al., 2010) and help set expectations around recovery timelines. Similarly, the insight gained from pairing detailed fish population study with data collection on watershed processes helps clarify the mechanisms through which different restoration measures may assist recovery of ecological values directly. In particular, the timescale of responses in channel dynamics to watershed disturbance and responses in fish populations to channel dynamics provide approximate estimates of the timeline needed for large-scale restoration (e.g., Ogston et al., 2015).
The duration of the Carnation Creek study coupled with the lack of in-stream restoration interventions also provides a rare opportunity to assess longer-term environmental shifts tied to climate change or other modes of climate variability (i.e., PDO and ENSO). While the system continues to respond to the forest harvesting activities of the 1970s and 1980s, the concurrent collection of climatological and hydrological data serves to help parse out this signal from larger-scale patterns.
Very few watershed-scale studies have persisted beyond the decade timescale, and the Carnation Creek study represents the longest watershed-scale, multi-disciplinary fish-forestry interaction study undertaken. The multi-disciplinary value of long-term studies cannot be understated: the interconnectedness of critical watershed functions, processes, and changes between geophysical and ecological spheres require long-term, detailed analysis to parse out subtle but important effects through time. In mountainous coastal regions, the highly episodic nature of key formative processes (e.g., landslides, major rain events) also means that long records are often needed to capture infrequent but impactful events in the watershed.
The practical application of findings from Carnation Creek, and other intensively monitored watersheds, extends widely. From a habitat restoration perspective, this study has highlighted the need to undertake work capable of restoring sediment input rates and wood recruitment processes. As new issues and changes to the severity of existing watershed challenges continue in the face of a changing climate, long-term studies such as Carnation Creek will continue to be an essential component of applied research.
Data availability statement
The data analyzed in this study is subject to the following licenses/restrictions: The data used for this document is currently under analysis for additional research. Data is available upon request from the authors. Requests to access these datasets should be directed to Peter Tschaplinski, UGV0ZXIudHNjaGFwbGluc2tpQGdvdi5iYy5jYQ==.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
PT: Data curation, Formal Analysis, Funding acquisition, Writing – original draft, Writing – review & editing. DR: Data curation, Formal Analysis, Visualization, Writing – original draft, Writing – review & editing. RP: Data curation, Formal Analysis, Visualization, Writing – original draft, Writing – review & editing. DS: Data curation, Visualization, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Beechie T. J., Sear D. A., Olden J. D., Pess G. R., Buffington J. M., Moir H., et al. (2010). Process-based principles for restoring river ecosystems. BioScience 60, 209–222. doi: 10.1525/bio.2010.60.3.7
Brown T. G. and Hartman G. F. (1988). Contribution of seasonally flooded lands and minor tributaries to coho (Oncorhynchus kisutch) salmon smolt production in Carnation Creek, a small coastal stream in British Columbia. Trans. Am. Fish. Soc. 117, 546–551. doi: 10.1577/1548-8659(1988)117<0546:COSFLA>2.3.CO;2
Bustard D. R. and Narver D. W. (1975). Aspects of the winter ecology of juvenile coho salmon (Oncorhynchus kisutch) and steelhead trout (Salmo gairdneri). J. Fish. Res. Board Can. 32, 667–680. doi: 10.1139/f75-086
Everest F. H., Beschta R. L., Scrivener J. C., Sedell J. R., and Cederholm C. J. (1987). “Fine sediment and salmon production: A paradox,” in Streamside management: Forestry and fishery interactions. Ed. Salo and T.W. Cundy E. O. (Institute of Forest Resources, Contribution 57, University of Washington, AR-10Seattle, WA), 98–103.
Hartman G. F. and Scrivener J. C. (1990). Impacts of forestry practices on a coastal stream ecosystem, Carnation Creek, British Columbia. Can. Bull. Fish. Aquat. Sci., 223.
Hartman G. F., Scrivener J. C., Holtby L. B., and Powell L. (1987). “Some effects of different streamside treatments on physical conditions and fish population processes in Carnation Creek, a coastal rain forest stream in British Columbia,” in Streamside management: Forestry and fishery interactions. Ed. Salo and T.W. Cundy E. O. (Institute of Forest Resources, Contribution 57, University of Washington, AR-10 Seattle, WA), 330–372.
Holtby L. B. (1988). Effects of logging on stream temperatures in Carnation Creek, British Columbia, and associated impacts on the coho salmon (Oncorhynchus kisutch). Can. J. Fish. Aquat. Sci. 45, 502–515. doi: 10.1139/f88-060
Holtby L. B. and Scrivener J. C. (1989). “Observed and simulated effects of climatic variability, clear-cut logging, and fishing on the numbers of chum salmon (Oncorhynchus keta) and coho salmon (O. kisutch) returning to Carnation Creek, British Columbia,” in Proceedings of the national workshop on effects of habitat alteration on salmonid stocks, vol. 105. Eds. Levings C. D., L.B. Holtby L. B., and Henderson M. A. (Vancouver, BC: Can. Spec. Fish. Aquat. Sci), 62–81.
Krajina V. J. (1969). “Ecology of forest trees in British Columbia,” in Ecology of western north america. Eds. Krajina V. J. and Brooke. R. C. (Department of Botany, University of British Columbia, Vancouver, BC), 1–146.
Ogston L., Gidora S., Foy M., and Rosenfeld J. (2015). Watershed-scale effectiveness of floodplain habitat restoration for juvenile coho salmon in the Chilliwack River, British Columbia. Can. J. Fish. Aquat. Sci. 72, 479–490. doi: 10.1139/cjfas-2014-0189
Oosterbaan R. J. (1994). “Frequency and regression analysis of hydrologic data. Chapter 6,” in in Publication 16. Drainage principles and applications, Part II: Regression analysis (Second Revised Edition). Ed. Ritzema H. P. (International Institute for Land Reclamation and Improvement (ILRI, Wageningen, The Netherlands), 1–24.
Pacific Climate Impacts Consortium (PCIC) (2023). Climate projections for the Regional District of Nanaimo. Regional District of Nanaimo; Pacific Climate Impacts Consortium (PCIC). Available online at: https://www.rdn.bc.ca/sites/default/files/inlinefiles/Regional_District_of_Nanaimo_Report-Final_0.pdf (Accessed June 14, 2025).
Reid D. A. and Hassan M. A. (2020). Response of in-stream wood to riparian timber harvesting: Field observations and long-term projections. Water Resour. Res. 56, e2020WR027077. doi: 10.1029/2020WR027077
Reid D. A., Hassan M. A., Bird S., and Hogan D. (2019). Spatial and temporal patterns of sediment storage over 45 years in Carnation Creek, BC, a previously glaciated mountain catchment. Earth Surf. Pro Landforms 44, 1584–1601. doi: 10.1002/esp.4595
Reid D. A., Hassan M. A., Bird S., Pike R. G., and Tschaplinski P. (2020a). Does variable channel morphology lead to dynamic salmon habitat? Earth Surf. Pro Landforms 45, 295–311. doi: 10.1002/esp.4726
Reid D. A., Hassan M. A., and Floyd W. (2016). Reach-scale contributions of road-surface sediment to the Honna River, Haida Gwaii, BC. Hydrol. Pro 30, 3450–3465. doi: 10.1002/hyp.10874
Reid D. A., Pike R. G., Bird S., Tschaplinski P., and Wilford D. (2020b). Implications of legacy watershed disturbances for channel structure and salmon habitat availability under different low-flow levels: an analysis of 45 years of discharge–habitat relationships at Carnation Creek, British Columbia. Can. J. Fish. Aquat. Sci. 77, 1780–1793. doi: 10.1139/cjfas-2020-0120
Scrivener J. C. (1988a). “Changes in the composition of the streambed between 1973 and 1985 and the impacts on salmonids in Carnation Creek,” in Proceedings of the workshop: Applying 15 years of Carnation Creek results. Ed. Chamberlin T. W. (Carnation Creek Steering Committee, Pacific Biological Station, Nanaimo, British Columbia), 59–65.
Scrivener J. C. (1988b). “A summary of the population responses of chum salmon to logging in Carnation Creek, British Columbia, between 1971 and 1986,” in Proceedings of the workshop: Applying 15 years of Carnation Creek results. Ed. Chamberlin T. W. (Carnation Creek Steering Committee, Pacific Biological Station, Nanaimo, British Columbia) pp 150-158.
Scrivener J. C. and Andersen B. C. (1984). Logging impacts and some mechanisms that determine the size of spring and summer populations of coho salmon fry (Oncorhynchus kisutch) in Carnation Creek, British Columbia. Can. J. Fish. Aquat. Sci. 41, 1097–1105. doi: 10.1139/f84-129
Scrivener J. C. and Brownlee M. J. (1989). Effects of forest harvesting on spawning gravel and incubation survival of chum (Oncorhynchus keta) and coho salmon (O. kisutch) in Carnation Creek, British Columbia. Can. J. Fish. Aquat. Sci. 46, 681–696. doi: 10.1139/f89-087
Seber G. and LeCren E. (1967). Estimating population parameters from catches large relative to the total population. J. Anim. Ecol. 36, 631–643. doi: 10.2307/2818
Slaney P. and Zaldokas D. (1997). Fish habitat rehabilitation procedures [online] Vol. No. 9 (Vancouver BC: Watershed Restoration Technical Circular). Available online at: http://www.env.gov.bc.ca/wld/documents/wrp/wrtc_9.pdf (Accessed July 12, 2025).
Spittlehouse D. L. and Maloney D. (2023). Rainfall interception by mature coastal and interior forests in british columbia. Confl. J. Watershed Sci Manage. 6, 20. doi: 10.22230/jwsm.2023v6n1a55
Tschaplinski P. J. (1998). “A summary of the effects of forest harvesting, fishing, and environmental shifts on salmonid populations of Carnation Creek, Vancouver Island, British Columbia,” in Forest-fish conference: land management practices affecting aquatic ecosystems. Eds. Brewin M. K. and Monita D. M. K. (Calgary, Alberta: Canadian Forest Service).
Tschaplinski P. J. (2000). “The effects of forest harvesting, fishing, climate variation, and ocean conditions on salmonid populations of Carnation Creek, Vancouver Island, British Columbia,” in Sustainable fisheries management: Pacific salmon. Eds. Knudsen E. E., Steward C. R., MacDonald D. D., Williams J. E., and Reiser D. W. (CRC Press, Boca Raton, FL and New York, NY), 297–327.
Tschaplinski P. J. (2010a). Watershed research summary: Carnation Creek watershed experiment. Streaml. Watershed Manage. Bull. 14.
Tschaplinski P. J. (2010b). State of stream channels, fish habitats, and their adjacent riparian areas – Resource Stewardship Monitoring to evaluate the effectiveness of riparian management 2005 – 2008. FREP Rep. Ser. 27.
Tschaplinski P. J. and Hartman G. F. (1983). Winter distribution of juvenile coho salmon (Oncorhynchus kisutch) before and after logging in Carnation Creek, British Columbia, and some implications for overwinter survival. Canadian. J. Fish. Aquat. Sci. 40, 452–461. doi: 10.1139/f83-064
Tschaplinski P. J., Hogan D. L., and Hartman G. F. (2004). “Fish-forestry interaction research in coastal British Columbia: The Carnation Creek and Queen Charlotte Islands studies,” in Fishes and forestry: Worldwide watershed interactions and management (Blackwell Press, London, UK), 389–412.
Tschaplinski P. J. and Pike R. G. (2010). Chapter 15. Riparian management and effects on function, in Compendium of forest hydrology and geomorphology in British Columbia Vol. 66. Eds. Pike R. G., Redding T. E., Moore R. D., Winkler R. D., and Bladon K. D. (Victoria, BC: Land Management Handbook), 497–525.
Tschaplinski P. J. and Pike R. G. (2017). Carnation Creek watershed experiment – long-term responses of coho salmon populations to historic forest practices. Ecohydrology 10, e1812. doi: 10.1002/eco.1812
Keywords: watersheds, restoration effectiveness, forest practices, riparian practices, landslides, aquatic habitats, climate change, Pacific salmon
Citation: Tschaplinski PJ, Reid DA, Pike RG and Spittlehouse DL (2025) Long-term forestry and climate change effects on watershed processes and salmon populations at Carnation Creek, Vancouver Island, British Columbia. Front. Ecol. Evol. 13:1704400. doi: 10.3389/fevo.2025.1704400
Received: 12 September 2025; Accepted: 20 October 2025;
Published: 31 October 2025.
Edited by:
Robert Bilby, Retired, North Bend, WA, United StatesReviewed by:
Shannon Claeson, USDA Forest Service PNW Research Station, United StatesWarren Devine, Washington Department of Natural Resources, United States
Copyright © 2025 Tschaplinski, Reid, Pike and Spittlehouse. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter J. Tschaplinski, UGV0ZXIuVHNjaGFwbGluc2tpQGdvdi5iYy5jYQ==