 Thomas Eltz
Thomas Eltz Hannah Luisa Kimm1
Hannah Luisa Kimm1 Maximilian Sigl
Maximilian Sigl- 1Department of Animal Ecology, Evolution and Biodiversity, Ruhr-Universität Bochum, Bochum, Germany
- 2Colegio de Ciencias Biológicas y Ambientales, Universidad San Francisco de Quito, Quito, Ecuador
- 3Center for Integrative Biodiversity Discovery, Leibniz Institute for Evolution and Biodiversity Science, Museum für Naturkunde, Berlin, Germany
- 4Institute for Biology, Humboldt University, Berlin, Germany
Frog-biting midges (Corethrellidae) are tropical, blood-sucking micro-predators of frogs that locate their prey by eavesdropping on anuran mating calls. In the present study we used acoustic traps broadcasting the call of Savage’s Thin-toed Frog (Leptodactylus savagei) to elucidate frog-biting midge stratification in the rainforest as well as the directionality and spatial scale of their phonotactic approach. In an Amazonian rainforest in Ecuador, midges were attracted to acoustic traps in all forest strata, including the canopy of an emergent tree 38 m above the forest floor, with a significant effect of stratum on midge species composition. Of the eight putative species (MOTUs) of Corethrella delimited using COI barcoding (N = 158) four were also found in the highest forest stratum (28.5–38 m), with one of them showing a preference for the middle and top canopy. In both Ecuador and Costa Rica catch numbers of ground-based traps differed strongly depending on the orientation of an attached mesh tunnel (65 cm) that was used to filter approach directions. Very few midges entered traps when mesh tunnels allowed horizontal (0°) or inclined (45°) approach vectors, while the highest catch numbers were recorded when tunnels were oriented vertically (90°). Consistent results were obtained when the length of the attached tunnel was extended to 250 cm, confirming that the primary approach to a calling host follows a vertical trajectory, and demonstrating that frog-biting midges can perceive acoustic stimuli in the acoustical far field. When stimulus intensity was varied, midges showed a significant phonotactic response at sound pressure levels as low as 56 dBA at tunnel entrance (65 cm), equivalent to a male Leptodactylus savagei calling from ~30 m distance. Our results emphasize that, despite their minute size, frog-biting midges are highly sensitive to acoustic stimuli. They are also highly mobile, occupy all strata of the neotropical rainforests and preferentially home in on their prey from above.
Introduction
Frog-biting midges (Corethrellidae), the sister group of mosquitoes (Culicidae) and phantom-midges (Chaoboridae), are mostly tropical, tiny (1–2 mm), blood-sucking midges that have an unusual way of locating their blood hosts, frogs. Female midges eavesdrop on the nocturnal mating calls of male frogs, being attracted maximally by certain combinations of frequency and pulse duration/rate (Bernal et al., 2006; Camp, 2006; Grafe et al., 2008; Virgo et al., 2019). Dozens of midges can sometimes be seen imbibing blood from an individual male frog at a tropical pond, and several hundreds of midges may feed on blood from a frog in one night (Bernal et al., 2006). While there are other dipteran species using acoustic stimuli for prey location, e.g. certain frog-biting culicid mosquitoes (Bartlett-Healy et al., 2008) or tachinid parasitoids of crickets (Müller and Robert, 2001), in corethrellids phonotactic predation is characteristic for the entire pan-tropical group (Borkent, 2008). Corethrellid activity is strictly nocturnal and frogs appear to be the only natural blood hosts (Borkent, 2008; Cutajar and Rowley, 2020). While specialized on locating frog calls they seem to have a relatively generalized acoustic template (Legett et al., 2018), with at least the more common species in the neotropics being attracted to a broad range of frog calls and even pure sinusoidal tones (Legett et al., 2018; Virgo et al., 2019; Geisler et al., 2024).
Although frog-biting midges have gained increasing interest for their phonotactic behavior in recent years, many aspects remain unknown. One major area of uncertainty is the spatial range within which frog-biting midges detect and react to auditory cues, which has important implication for the ecology and evolution of interactions with their hosts. Hearing in Nematocera (“midges” within the insect order Diptera) is mediated by flagellar ears, i.e. the plumose antennae that are vibrated in response to particle-velocity changes resulting from sound in air (Göpfert et al., 1999; Pantoja-Sánchez et al., 2023). Traditionally such ears are considered to function only in the near field, i.e. over distances of centimeters to decimeters (Albert and Kozlov, 2016). However, the effective range of a hearing organ depends ultimately on its sensitivity, and more recent research suggests that mosquitoes can detect sounds from distances of > 5 m if these sounds were of sufficient intensity (Bartlett-Healy et al., 2008). In behavioral and physiological experiments sounds broadcast from distances up to 10 m (perceived at a minimum sound level of 31 dB) evoked stimulus-related responses in Aedes aegypti (Menda et al., 2019). The numbers of female Corethrella attracted to acoustic lures, and the immediacy of their approach, would suggest that they possess comparable capacities, but this has not been investigated in a systematic way.
In the present study we investigated spatial aspects of corethrellid foraging behavior in the field using acoustic traps. Initially, we investigated the directionality of the phonotactic approach to increase our understanding on how these tiny insects locate their blood hosts in a complex three-dimensional environment. These experiments were stimulated by visual observations made in the light cone of a strong LED torch in La Gamba (R. Böddicker & T. Eltz, pers. obs.). When a torch was pointed to the dark night sky above a sound trap in a forest clearing, midges could be observed “falling down” towards the speaker from >3 m height (the distance at which the torch produced visible reflections from midges) within seconds after playback commenced. In our experiments we therefore tested the hypothesis that midges prefer a vertical approach to an acoustical stimulus over a horizontal or inclined one. Additionally, we measured the minimum stimulus intensity that is sufficient to elicit a phonotactic approach from a certain distance and estimated the distance from which natural frog calls might be approached in the natural habitat. Finally, having obtained evidence for a strong vertical component in the phonotactic approach, we assessed the presence of frog-biting midges in the different vertical strata of the rain forest and examined whether there is stratum-specificity among different species/molecular taxonomic units (MOTUs) of midges. Although vertical stratification has been investigated in several insect taxa, including also hematophagous Diptera in tropical forests (Basset et al., 2008), this aspect has not been investigated in detail in frog-biting midges (Corethrellidae; but see de Souza Amorim et al., 2022).
Methods
The experiments were done from 2022 to 2024 at two lowland neotropical forest sites: at the La Gamba research station in southern pacific Costa Rica (www.lagamba.at), and at Tiputini Biodiversity Station (TBS) located in Amazonian Ecuador adjacent to the Yasuní National Park (www.tiputini.com). Both localities harbor species-rich frog communities (Cisneros-Heredia, 2006; Franzen and Kollarits, 2018). Whereas La Gamba is relatively well studied with regard to frog-biting midges (Sigl et al. 2025, Virgo et al., 2019, 2021, Virgo and Eltz, 2022) very little is known about Corethrella in the western Amazon.
In all experiments midges were trapped with custom made, compact (height 17 cm, diameter 13 cm), fan-driven (Biogents 5V) traps broadcasting a call of the widespread Savage’s Thin-toed Frog, Leptodactylus savagei, consisting of series of ‘whorups’ of 0.25 s pulse duration and a dominant frequency of 379 Hz (see https://homepage.ruhr-uni-bochum.de/thomas.eltz/Frogs/Leptodactylidae.html). Series of L. savagei-calls have proven to be highly attractive for Corethrella spp. at several neotropical localities (Virgo et al., 2019, Virgo & Eltz, pers. obs.) and for a wide range of Corethrella species (Virgo et al., 2021). For example, in La Gamba, traps broadcasting L. savagei calls attracted all of the five species of Corethrella that are regularly collected directly from frogs, including one species that feeds predominantly on tree frogs (Virgo et al., 2021). While L. savagei only ever calls from ground level, or even from holes in the ground, the broad attractiveness of its call led us to use it as standardized attractant also in the vertical stratification assay.
In the present study we played repeated series of 24 L. savagei-’whorups’ (1 per 1.8 s) followed by 15 s intervals of silence for variable times (see below). If not indicated otherwise these series were played at a standardized intensity using Jay‐Tech K10/A100 speakers placed 10 cm above the opening of the actual suction trap where midges were caught in a fine nylon mesh bag (Biogents). At the used speaker level the L. savagei call produced an average peak sound pressure level (SPL) of 78.06 dB(flat) at 65 cm from the source (measured in the lab with an Acoustilizer AL1, NTi Audio AG, Liechtenstein), corresponding to 74.3 dB at 1 m. Midge samples were immobilized in a freezer and counted under a stereomicroscope. Excluded from counts were the occasional culicid and other non-corethrellid midges.
If not indicated otherwise all statistical tests were done with GraphPad Prism 10.
Vertical stratification assay
In November 2022 we used the Canopy Tower at TBS to assess midge stratification. This tower has been erected next to a massive emergent tree (Ceiba pentandra, height ca 50 m Figure 1) without substantial alterations of the natural forest surroundings, i.e. the tree and tower is embedded in the continuous natural forest matrix with other herbaceous and woody vegetation nearby (see Figure 1). We trapped midges with three separate trap units at nine evenly spaced heights (platforms) along the stairs of the tower, from 0 m (forest floor) to 38 m (top platform). Traps were operated for ten minutes to obtain samples, with speakers placed horizontally above the trap (0°) and always aligned in the same compass direction.

Figure 1. Frog-biting midges (Corethrella spp.) in acoustic traps broadcasting Leptodactylus savagei calls (right) on the canopy tower at Tiputini Biodiversity Station in Amazonian Ecuador (left). Grey shades in the right panel show an approximation of the vegetation structure around the tower.
Trap units were moved to other heights following a scheme that left at least one platform empty between traps at all times (i.e., maintained > 9.50 m distance in height between them) and avoided an association of night and height. Four rotations were done per night (from 20:30 to 21:30 h), repeated in four nights, resulting in 3 (traps) x 4 (rotations) x 4 (nights) = 48 midge samples, with 5 to 6 replicates per height. These samples were stored in 2 ml Eppendorf tubes filled with EtOH (abs.). Midges were counted per sample and Spearman’s rank correlation was used to test for an effect of height on midge number. From 31 samples that contained midges a total of 264 individual midges were selected randomly for DNA barcoding, without prior knowledge of species affiliation and aiming at covering all heights equally. Genomic DNA was isolated following the GeneReleaser protocol (BioVentures Inc.), adapted by (Weigand, 2013) and optimized for Corethrella by Virgo et al. (2021). For barcoding, a 658 bp fragment of the cytochrome oxidase subunit I (COI) gene was amplified using primers HCO1490/LCO02198 (Folmer et al., 1994), each tagged with a unique 9 bp sequence as described by Srivathsan et al. (2023). Pooled amplicons were sequenced on an Oxford Nanopore MinION at the Center for Integrative Biodiversity Discovery (Museum für Naturkunde Berlin). All barcodes were queried against the NCBI (BLASTN with minimum e-value of 1e-10) and BOLD databases to check for membership in Corethrella. For species delimitation, barcodes confirmed as likely belonging to Corethrella spp. were analyzed using ASAP (Assemble Species by Automatic Partitioning; Puillandre et al., 2021). For partitioning we included Corethrella sequences of 17 putative species obtained from Genbank, that had been corroborated with a second marker (ITS2; Virgo et al., 2019).
For statistical analysis of MOTU composition the 31 samples from which the sequenced midges originated were grouped in three strata, combining three heights each (low=0-9.5 m, middle =14.25-23.75 m, and high=28.50–38 m). To test for differences in midge MOTU composition a one-way PERMANOVA was calculated using the R package vegan (Oksanen et al., 2015), using Bray-Curtis distances as a distance metric, with ‘stratum’ as a grouping variable and 999 permutations.
Homing-angle experiment
To assess the vertical (meridional) angle of the approach to acoustic stimuli we put trap units in mesh tunnels that were midge-proof and had only one opening. This allowed us to “point” the whole arrangement at variable angles away from the sound source. Two different setups were used, short and long: 90x60x60 cm nylon mesh Life Unearthed Pop-Up-Insect Cages (90 x 60 x 60 cm) were cut open at one small side (60x60 cm) to provide the short setup, and fully collapsible, 250x50 cm cylindrical nylon Mesh Tunnels (Spordas, for children) were closed at one end with a plastic bag, providing the long set-up. In both setups a trap unit was placed at the closed end, resulting in distances of 65 cm (short setup) or 250 cm (long setup) from the opening to the sound source/trap. These distances represent the minimal distances at which trapped midges must have perceived the stimulus to make the decision to orient towards it. We used setups of different length to vary the minimal distance of perception/orientation.
The short setup was operated at horizontal (0°), inclined (45°) and vertical (90°) orientation in two separate experiments at TBS (November 2022) and La Gamba (March 2023). The traps themselves remained upright regardless of angular treatment to allow equal suction efficiency, but the trap speakers were oriented in alignment with the tunnel to maintain sound pressure levels at the tunnel opening. All experiments were conducted within the first 3 hours of the night. The trap setups were always placed at the edge of forest clearings, just outside of overhanging vegetation, at sites that were at least 15 meters apart from each other. Each site was only used once for a given angular treatment and traps were run for ten minutes per replicate. Three (TBS) or six (La Gamba) trap units were rotated systematically through sites and angle treatments. We collected 9 replicate samples per angle treatment in TBS, resulting in 27 samples, and 6 replicate samples in La Gamba, resulting in 18 samples. In TBS we also collected control samples without attached tunnel and with the speaker in vertical orientation (90°; 8 samples).
The long setup was only used in La Gamba (March 2023) with four different angle treatments (0°, 45°, 72,5°, 90°). Only two tunnels were available, so we rotated two trap units/tunnels through treatments and sites. The general procedure was similar, but in this case we used SoundCore Anker Mini 3 loudspeakers, and trapping duration was 30 min per replicate. At the used sound level, the L. savagei call produced an average sound pressure level (SPL) of 68.5 dBA at the opening 250 cm from the source (measured in the field with a Benetech Digital Sound Level Meter GM1351), corresponding to 76,5 dBA at 1 m.
Behavioral response threshold
To determine the minimal sound pressure level that would trigger phonotactic behavior to L. savagei calls in midges we used the short setup (see homing-angle experiment) with vertical orientation of the tunnel and variable speaker level setting (level 5 to 10, Jay-Tech speakers, see Results). For every treatment trial we ran a silent control that except for the lack of the frog-call play-back was identical to the treatment. Individual pairs of treatment and control traps were placed at a distance of ten meters in similar situations and operated for 20 minutes simultaneously. A total of 72 paired trials were run during the first three hours of nine nights in March 2024 in the station garden or near the Laguna in La Gamba, Costa Rica (see Results for number of replicates per speaker-level setting). During each trial we measured SPL of the played L. savagei call at the tunnel entrance of the treatment trap using a Benetech Digital Sound Level Meter GM1351. The device was regularly checked for deviations before the trials using a Voltcraft SLC-100 calibrator. For comparison we took a series of controlled measurements in the lab, thereby excluding the partly substantial nocturnal background noise in the field. To test for differences in midges captured between treatment and silent control traps we used Wilcoxon matched-pairs test.
To determine at what distance the SPL of natural calls of Leptodactylus savagei would fall below the behavioral response threshold we measured SPL at individual males calling in Costa Rica and calculated threshold distances using the inverse square law. A total of four frog individuals were measured from distances between 3.5 to 8 m, with 1 to 4 replicate measurements each. The frogs were calling from variable sites: one from underneath a pile of fallen leaves, one from a hole in a road embankment, one from within a bush and one openly from the edge of a pond. Distances to frogs (or their hideouts) were measured using a RockSeed laser distance meter. Repeated SPL measurements were taken from different angles. Mean threshold distances as well as SPL at 1 m were calculated for each measurement using the inverse square law, means calculated for individual frogs, and grand means across frog individuals. Although the field conditions certainly violated some of the assumption of the calculations (spherical sound emission, no reflections), we are confident that they allow a crude estimate of threshold distance.
Results
Vertical stratification assay
A total of 439 frog-biting midges were trapped at different heights on the TBS canopy tower, with an overall decrease in numbers from bottom to top (Figure 1; Spearman Rank Correlation: N = 45; Rs=0.52; p<0.001). However, there were numerous midges active also at higher strata, with the third and fourth highest midge counts at 33.25 m, in the lower canopy of the emergent support tree.
We obtained 158 COI barcodes that matched the genus Corethrella in BLAST analyses (average of 97.6% percent pairwise identity; range 86.1-100%).
Based on these sequences, the best-supported ASAP partition, defined by the minimum ASAP score (4.50), was selected for further analysis and yielded 24 MOTUs, with the next two best-ranked partitions suggesting 42 (score = 5.0) or 23 (score = 7.50) MOTUs, respectively. Of these 24 MOTUs, nine derived from the stratification assay, of which one could be assigned to the widely distributed Corethrella amazonica (96.5-97.7% pairwise identity; N = 31; Borkent, 2008; Geisler et al., 2024). The remaining eight could only be identified to the genus level as Corethrella sp. (Figure 2). However, four of these putative species corresponded to Corethrella MOTUs delimited in previous studies (96.5-100.0% pairwise identity; see Supplementary Material Table 1). The delimitation of the 17 included reference MOTUs obtained from Genbank was largely recovered, except for two MOTUs (Corethrella peruviana and Corethrella sp. peruviana 1), which were combined into a single MOTU (Supplementary Material 2).
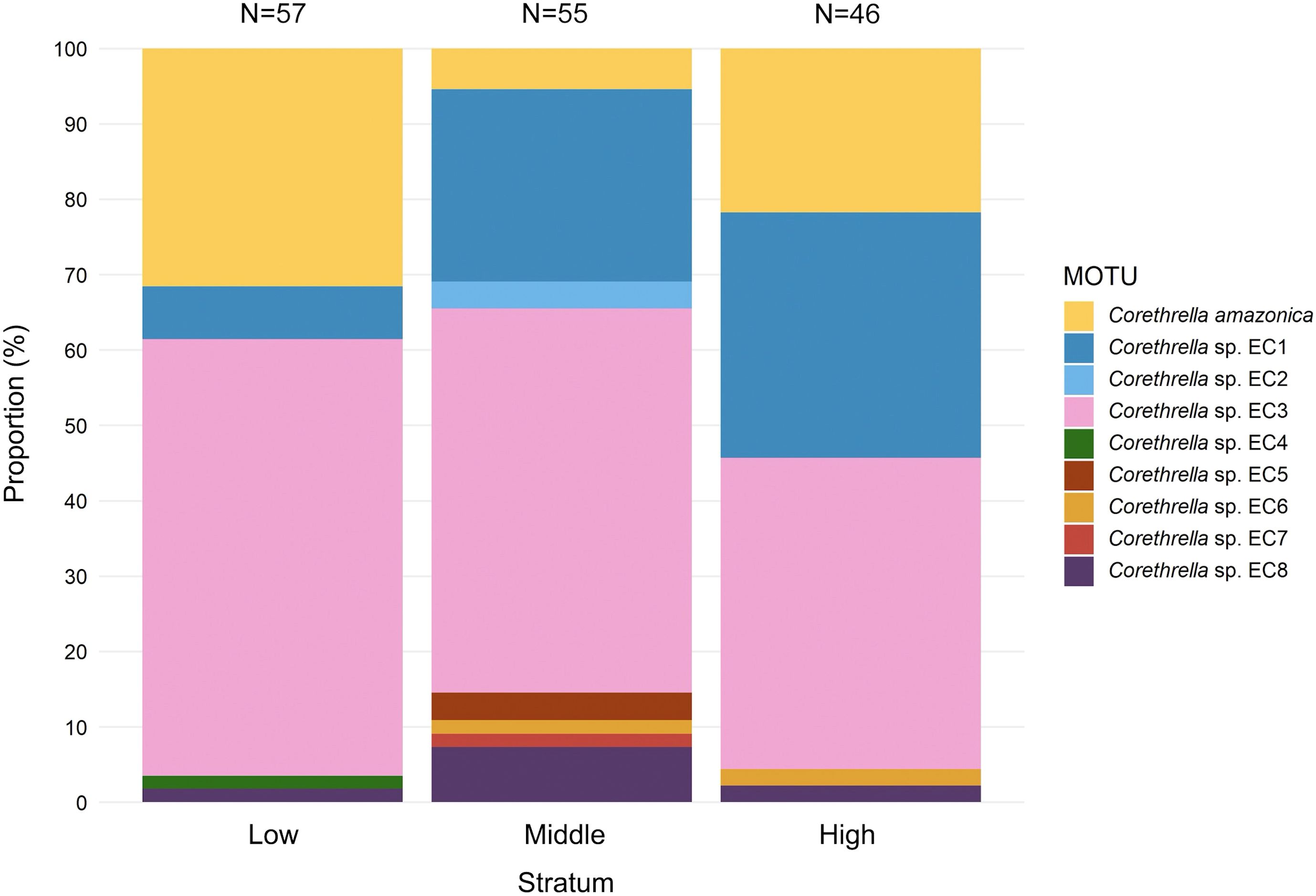
Figure 2. MOTU composition of frog-biting midges (Corethrella spp.) in acoustic traps broadcasting Leptodactylus savagei calls on the canopy tower at Tiputini Biodiversity Station in Amazonian Ecuador. Sequenced midges (N = 158) were drawn from samples taken at nine different heights (see Figure 1), which were grouped into three strata for analysis and visualization.
There were 5 (low), 8 (middle) and 5 (high) different MOTUs in the three different strata, with the four most common ones having been trapped in all three strata. Based on PERMANOVA there was a significant effect of stratum on midge MOTU composition (N = 31 samples; F = 2.72; p=0.0105). This effect seems to have been driven mostly by one MOTU, Corethrella sp. EC1, which increased in both absolute and relative numbers from low to middle to high stratum.
Homing-angle experiment
There was a significant effect of angular treatment on midge numbers, a result that was qualitatively identical in TBS and La Gamba (Kruskal-Wallis Test: TBS: N = 27; p<0.01; La Gamba: N = 18: p<0.001; see Figure 3 for post-hoc comparisons). Very few midges entered traps when mesh tunnels were oriented horizontally (0°) or were inclined by 45°, while maximal catch numbers were recorded when tunnels were oriented vertically (90°; Figure 3). Absolute catch numbers were approximately three times as high in La Gamba compared to TBS, which is in accordance with our impression of outstanding frog-biting midge abundance around the La Gamba research station (Virgo et al., 2019).
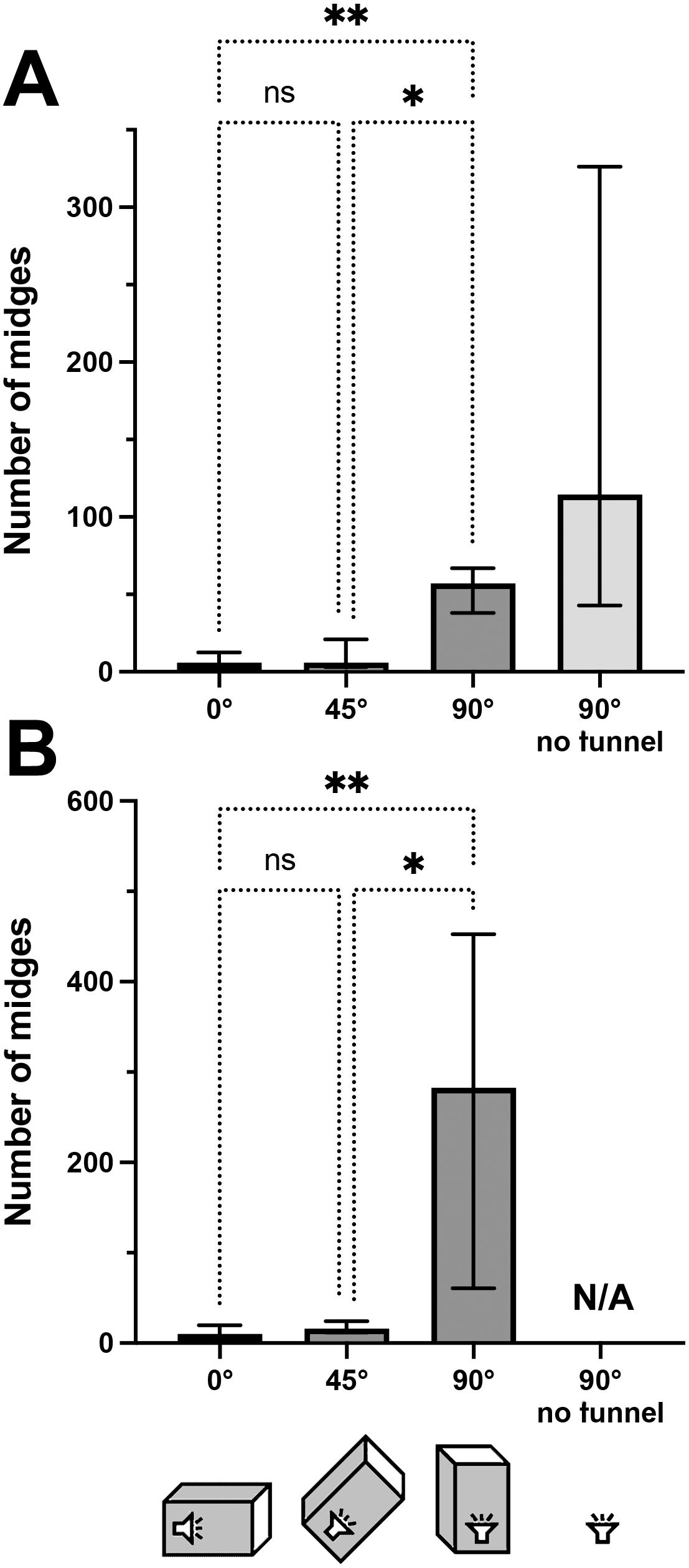
Figure 3. Number of midges trapped per replicate in the homing-angle experiments using the short tunnel setup (65 cm) at TBS (A) and La Gamba (B). Median and interquartile range are plotted. Brackets above columns represent results of Kruskal-Wallis post-hoc tests for multiple comparisons (*p<0.05, **p<0.01).
Absolute midge numbers were substantially lower with the long setup in La Gamba, reflecting lower sound pressure levels at the tunnel entrance. However, relative proportions of catches corresponded closely to the short setup, with the 67.5° treatment being intermediate between 45° and 90° (Kruskal-Wallis Test: N = 48; p<0.001; see Figure 4 for post-hoc comparisons).
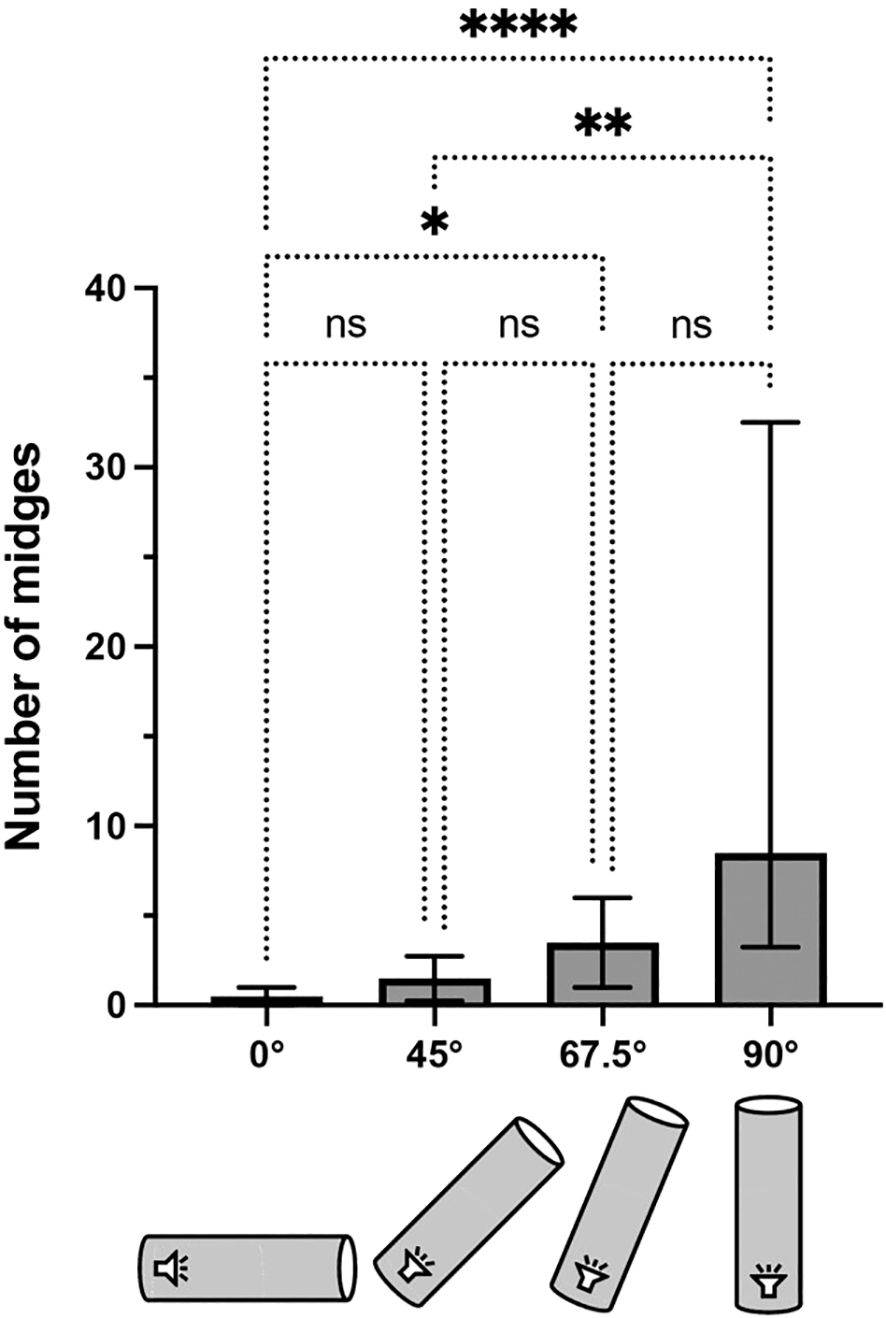
Figure 4. Number of midges trapped per replicate in the homing-angle experiments using the long tunnel setup (250 cm) at La Gamba. Median and interquartile range are plotted. Brackets above columns represent results of Kruskal-Wallis post-hoc tests for multiple comparisons (*p<0.05,**p<0.01,****p<0.0001).
The species composition of midge samples was not investigated in the Homing-angle experiment (see Discussion).
Behavioral response threshold
In the experiment to determine the behavioral response threshold the silent control traps were all empty except for a single trap that had captured 1 midge. Treatment traps broadcasting L. savagei calls captured 0 to 369 midges. The percentage of traps with more midges than in the paired control increased from lower to higher speaker levels/sound pressure levels, with a steep increase in trapping success between level 7 and 8 (Figure 5). However, already at level 7 there were significantly more midges in the treatment traps than in silent controls (Wilcoxon matched-pairs test: N = 19; p<0.05). This level corresponded to a measured mean peak SPL of the played-back L. savagei calls at the tunnel entrance of 56.5 dBA (field) or 55.8 dBA (lab). Calls of wild male L. savagei measured in Costa Rica produced an average SPL at 1m of 84.5 (+/-4.5) dBA (N = 4). Assuming unobscured (free field) sound propagation their call SPL would fall below the Corethrella behavioral threshold at a distance of 28.7 (+/-11.9) m.
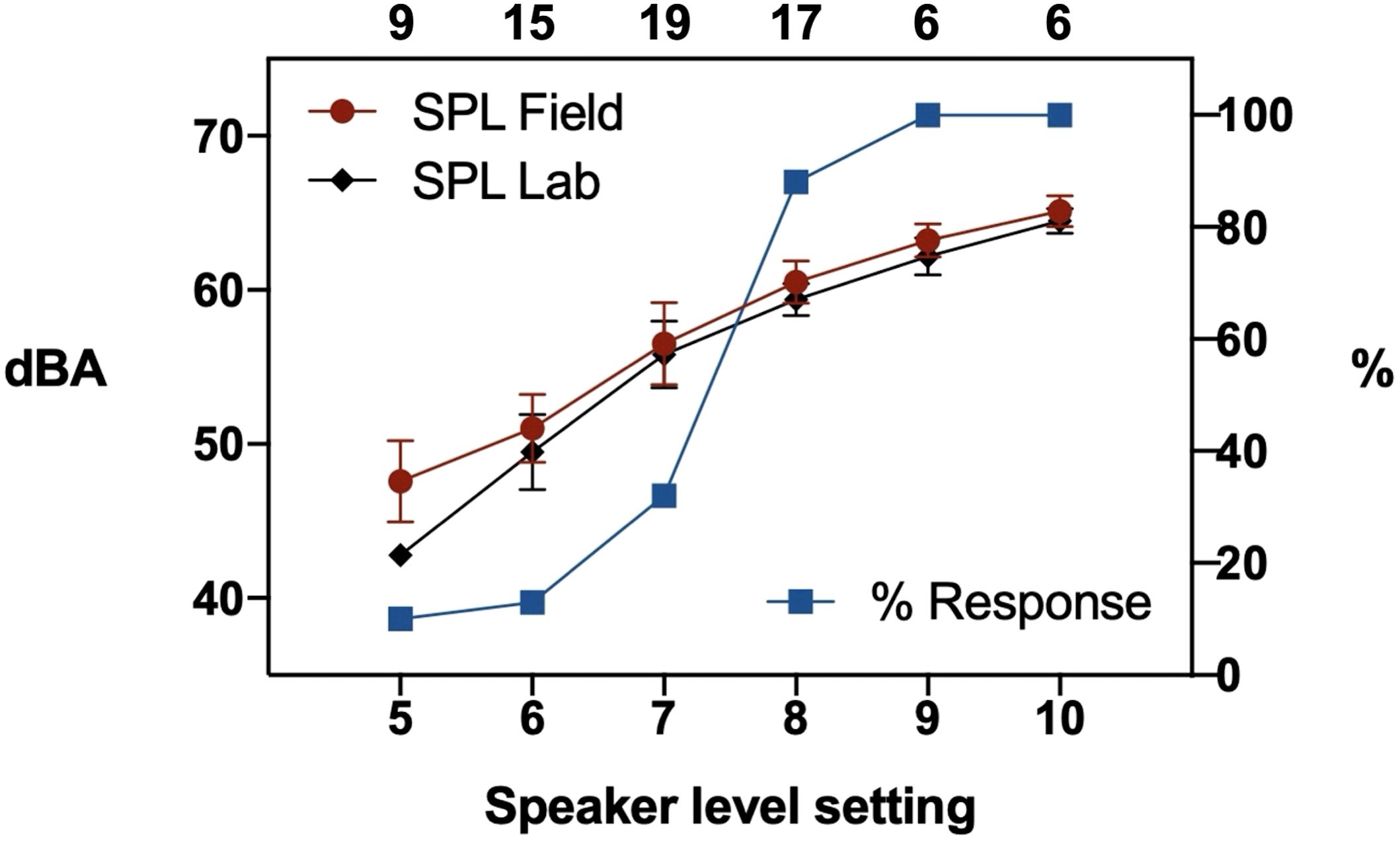
Figure 5. Trapping success (% traps with midges = % response) of acoustic midge traps broadcasting recorded Leptodactylus savagei calls at variable speaker level settings. The resulting sound pressure level was recorded at the tunnel entrance (65 cm from speaker, means across used individual speakers with standard deviation) during the field trial (red circles) and in the lab (black diamonds). The number of replicate trap assays (treatment plus silent control trap) is given by numbers above the diagram. Starting with speaker level 7 there was a significant difference of trapping success compared to silent control traps (see Text for details).
Discussion
This study expands our understanding of spatial aspects of frog-biting midge foraging and is the first to explore Corethrella stratification in a rainforest with complex vertical structure. Our results demonstrate that frog-biting midges are highly mobile and present in all strata of the rainforest, including the top canopy. This corroborates the findings of a previous study using Malaise traps in the Brazilian Amazon, which had found a small number of Corethrella sp. at a height of 24 m in the forest canopy (de Souza Amorim et al., 2022). McKeever and French (1991) sound-trapped corethrellids at 0.5, 1.0, 2.0, and 3.0 m above ground in swampland in Georgia, USA, and found that midge numbers declined sharply with height. They attributed this to humidity being highest near the ground, resting sites most abundant, and air movement slowest, an environment deemed suitable for Corethrella. In Amazonian Ecuador rainforest we also found declining numbers of midges with height above ground, but there were still substantial numbers of midges in the canopy. Presumably, low humidity and wind are less of a problem for small midges in a tropical rainforest, and, clearly, resting sites were present in all strata around the canopy tower.
We found a moderate stratification among Corethrella MOTUs, with the four most common MOTUs being present in all three strata. For the rarer MOTUs sample size was too small to evaluate their vertical distribution. Of the more common ones, Corethrella sp. EC1 showed perhaps the clearest trend towards a canopy preference, but future studies with larger sample sizes will have to confirm this trend. From the literature, the evidence for hematophagous dipterans to stratify vertically in forest environments is mixed, with some species/groups having a predictable ground or canopy preference (Swanson and Adler, 2010), whereas others show no, weak or seasonally dependent preferences (McGregor et al., 2018). Generally, it is unclear whether stratum preferences are caused by host preferences or if travelling height is determined by other factors (e.g. search for breeding sites, avoidance of predation) and preferred hosts are simply the ones most commonly encountered at that height. It should be emphasized that most studies on stratification of hematophagous dipterans were conducted in temperate forests and/or on only moderate vertical scales (e.g., ground vs. 10 m for canopy in (Swanson and Adler, 2010)). In tropical forests the scale and complexity of stratification is more substantial as are the challenges of canopy access and species identification. Here, limited data exists for medically important groups such as Ceratopogonidae and Phlebotominae, suggesting patterns of moderate stratification (Dias-Lima et al., 2002; Basset et al., 2008 and references therein) similar to what we found for Corethrellidae in the present study.
It could be argued that in our acoustical trap-based stratification assay frog-biting midges were lured to canopy traps from lower strata. Based on our own calculations, the SPL of the used calls could have attracted midges from distances of up to 7 m (see also discussion below). While we cannot exclude that inter-stratum attraction has influenced the stratum-specific community composition, frog-biting midges were attracted to canopy traps quickly, i.e. already during the first of a given days’ short (10 min) broadcast intervals. Thus, it seems likely that they were not lured to those heights by the traps but were already present in the canopy when traps started to broadcast. Tree frogs (Hylidae), of which at least two species were regularly heard calling from the epiphyte-covered branches at the site, are common targets of Corethrella in the neotropics (Virgo and Eltz, 2022; Virgo et al., 2021). Indeed, one Corethrella sp. EC8, captured at a height of 23.75 m during our assay, contained a blood meal later identified by DNA barcoding (Sigl & Eltz, unpublished). It was derived from Osteocephalus fuscifacies, a medium-sized tree frog previously recorded in Tiputini (Cisneros-Heredia, 2006) and known to breed in bromeliads (Jungfer et al., 2000). Corethrellids may have also been drawn to the canopy by water-filled bromeliad leave axils, which we have found to be important breeding habitats in La Gamba (Virgo, Sigl & Eltz, unpublished data). The upper canopy of the canopy tower support tree in Tiputini is densely populated with epiphytes, including many sizable bromeliads from the genus Aechmea.
Judging from the homing angle experiments, the primary approach to auditory cues includes a phase of vertical descent. Catch numbers of traps were maximal when mesh tunnels were open to the vertical but very low when mesh tunnels were oriented horizontally. These results were obtained at both rainforest localities (Amazonian Ecuador and Costa Rica), suggesting that a vertical approach may be common among species of Corethrella. Although we have not identified the midges in the homing angle experiment to species level, the species composition in Ecuador was probably similar to that of the vertical stratification assay (Figure 3; the two assays were done during the same 10-days visit and at sites only 500 m distant from each other). In La Gamba, Costa Rica, we can refer to previous studies that have found a different set of species dominated by the super-abundant C. ranapungens, which represented 65 to 98% of trap catches depending on season (Virgo et al., 2019, 2021). Thus, it seems safe to assume that the vertical approach pattern is not a quirk of just one species.
Vertical descent on acoustic stimuli has also been demonstrated for females of the cricket-parasitizing tachinid fly, Ormia ochracea (Müller and Robert, 2001). In Ormia the descent is remarkably accurate in complete darkness: flies are able to land only centimeters away from the loudspeaker after a flight distance of approximately 4 m (Müller and Robert, 2001). While Ormia ochracea uses tympanal ears for directional hearing, the vertical approach of Corethrella may be related to the mechanism that enables directional hearing with flagellar ears. Behavioral and electrophysiological experiments with Culex pipiens suggest that the in-flight position of antennae is optimal for binaural hearing focused in front of, above and below a mosquito (Lapshin and Vorontsov, 2023), resulting in an auditory space of female mosquitoes that has sufficient resolution to estimate the direction of a sound source. Possibly, an even more narrow directional tuning of flagellar responsiveness improves phonotactic targeting in even smaller insects such as frog-biting midges.
Alternatively, the vertical approach to acoustic stimuli may represent an optimal foraging strategy for Corethrella. In a dense rainforest environment, where flight paths are obstructed by branches, leaves, and other vegetation, a vertical descent may be energetically more efficient. By leveraging gravity to assist in their downward movement, host-seeking midges can cover greater distances more quickly and with less energy expenditure than in the horizontal plane. Additionally, this approach would minimize the risk of overshooting the target as the frog’s call can only originate from as far as ground level, providing a clear and defined location for the midge to target. Finally, sound scattering in forests results mostly from tree trunks which are oriented vertically, resulting in significant reduction of sound intensity along the horizontal plane (Bullen and Fricke, 1982). Propagation in the vertical may be largely unobscured and, using a vertical descent, Corethrella may be able to achieve more precise targeting of sound sources.
Our results also have implications regarding the distance that sounds may travel to be perceived by flagellar ears. Traditionally, such ears were considered to function only in the near field, i.e. over distances of centimeters to decimeters (Albert and Kozlov, 2016), in accordance with a function of detecting flight tones produced by conspecifics in mate-seeking mosquitoes (Pantoja-Sánchez et al., 2023). However, in behavioral experiments female Aedes aegypti mosquitoes became airborne in response to acoustic stimuli originating 3 m away from their mesh cage while their antennal sensilla were stimulated from even larger distances (10 m), at a sound pressure level of only 31 dB (in an anechoic chamber, Menda et al., 2019). Our field experiments support these results, demonstrating that female Corethrella respond to calls played from a distance of at least 2.5 m, the length of the long-tunnel setup, clearly in the far field. It seems that even tiny insects with flagellar ears are capable of exploring an acoustic space of surprisingly large dimensions. Just how large is this space in a 1 mm frog-biting midge encountering natural frog call intensities? We have determined that behavioral responses to L. savagei playback calls are elicited at sound pressure levels as low as 56 dBA. Based on averaged SPL measurements of four bull frogs calling in their natural environment in Costa Rica, and assuming zero reflectance from vegetation, we estimated that calls could be detected from a distance of up to 28.7 m. Clearly, there are many potential errors involved in these calculations. However, an perceptual space with a diameter of several dozen meters would fit well with observations of hundreds of frog-biting midges being attracted quickly and reliably to acoustic traps at neotropical forest sites.
Data availability statement
Datasets are deposited at Figshare and will publicly available as of the date of publication (https://doi.org/10.6084/m9.figshare.30539921). The sequences used in the stratification assay are available in BOLD at dx.doi.org/10.5883/DS-TIPEC22 and in GenBank under the accession numbers PX525671–PX525828.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
TE: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Supervision, Visualization, Writing – original draft, Writing – review & editing. HK: Investigation, Methodology, Writing – review & editing. RB: Investigation, Methodology, Writing – review & editing. MS: Conceptualization, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. RM: Investigation, Methodology, Resources, Supervision, Writing – original draft, Writing – review & editing. JV: Investigation, Methodology, Writing – original draft, Writing – review & editing.
Funding
The author(s) declared that financial support was received for this work and/or its publication. This work was supported by Ruhr-University Bochum and Deutsche Forschungsgemeinschaft (EL 249/13). Additional molecular expenses were covered by the core funding for the Center for Integrative Biodiversity Discovery. Open Access funding was provided by the Open Access Publication Fund of the Ruhr-Universität Bochum.
Acknowledgments
We thank Werner Huber and the staff of the La Gamba field station for their continuous support and Thomas Riedinger for help with sequencing. Tomi Sugahara, David Romo and Catalina Ulloa made our visits to Tiputini Biodiversity Station possible. We further thank the Ministerio de Ambiente y Energía (MINAE) of Costa Rica and the Ministerio del Ambiente, Agua y Transicíon Ecológica of Ecuador for granting permission to conduct this research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1705905/full#supplementary-material
References
Albert J. T. and Kozlov A. S. (2016). Comparative aspects of hearing in vertebrates and insects with antennal ears. Curr. Biol. 26, R1050–R1061. doi: 10.1016/j.cub.2016.09.017
Bartlett-Healy K., Crans W., and Gaugler R. (2008). Phonotaxis to amphibian vocalizations in Culex territans (Diptera: Culicidae). Ann. Entomol. Soc Am. 101, 95–103. doi: 10.1603/0013-8746(2008)101[95:Ptavic]2.0.Co;2
Basset Y., Kitching R., Miller S., and Novotny V. (2008). Arthropods of tropical forests: spatio-temporal dynamics and resource use in the canopy (Cambridge, United Kingdom: Cambridge University Press), 492.
Bernal X. E., Rand A. S., and Ryan M. J. (2006). Acoustic preferences and localization performance of blood-sucking flies (Corethrella Coquillett) to tungara frog calls. Behav. Ecol. 17, 709–715. doi: 10.1093/beheco/arl003
Borkent A. (2008). The frog-biting midges of the world (Corethrellidae: Diptera). Zootaxa 1804, 1–456. doi: 10.11646/zootaxa.1804.1.1
Bullen R. and Fricke F. (1982). Sound propagation through vegetation. J. Sound Vib. 80, 11–23. doi: 10.1016/0022-460X(82)90387-X
Camp J. V. (2006). Host attraction and host selection in the family Corethrellidae (Wood and Borkent)(Diptera). Dissertation (Statesboro, Georgia, USA: Georgia Southern University).
Cisneros-Heredia D. F. (2006). La Herpetofauna de la Estación de Biodiversidad Tiputini, Ecuador (Quito, Ecuador: Universidad San Francisco de Quito).
Cutajar T. P. and Rowley J. J. L. (2020). Surveying frogs from the bellies of their parasites: invertebrate-derived DNA as a novel survey method for frogs. Glob. Ecol. Conserv. 22. doi: 10.1016/j.gecco.2020.e00978
de Souza Amorim D., Brown B. V., Boscolo D., Ale-Rocha R., Alvarez-Garcia D. M., Balbi M. I. P. A., et al. (2022). Vertical stratification of insect abundance and species richness in an Amazonian tropical forest. Sci. Rep. 12, 1734. doi: 10.1038/s41598-022-05677-y
Dias-Lima A., Bermúdez E. C., Medeiros J. F., and Sherlock I. (2002). Vertical stratification of phlebotomine sandfly fauna (Diptera, Psychodidae) in a primary non-flooded forest of the Central Amazon, Amazonas State, Brazil. Cadernos Saúde Pública 18, 823–832. doi: 10.1590/S0102-311X2002000300026
Folmer O., Black M., Hoeh W., Lutz R., and Vrijenhoek R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 3, 294–299.
Franzen M. and Kollarits D. (2018). Pocket guide to the amphibians and reptiles of La Gamba, Costa Rica (Laurenti-Verlag, Bielefeld, Germany).
Geisler E., de F., de Campos L. L. F., Carvalho-Rocha V., Ferreira A. S., Dayrell J. S., et al. (2024). Specialisation in frog-biting midges (Diptera: Corethrellidae): A landscape perspective. Ecol. Entomol. 49, 530–543. doi: 10.1111/een.13325
Göpfert M. C., Briegel H., and Robert D. (1999). Mosquito hearing: sound-induced antennal vibrations in male and female Aedes aEgypti. J. Exp. Biol. 202, 2727–2738. doi: 10.1242/jeb.202.20.2727
Grafe T. U., Saat H. B. M., Hagen N., Kaluza B., Berudin Z. B. H., and Wahab M. A. B. A. (2008). Acoustic localisation of frog hosts by blood-sucking flies Corethrella Coquillet (Diptera: Corethrellidae) in Borneo. Aust. J. Entomol. 47, 350–354. doi: 10.1111/j.1440-6055.2008.00667.x
Jungfer K.-H., Ron S., Seipp R., and Almendáriz A. (2000). Two new species of hylid frogs, genus Osteocephalus, from Amazonian Ecuador. Amphibia-Reptilia 21, 327–340. doi: 10.1163/156853800507525
Lapshin D. N. and Vorontsov D. D. (2023). Mapping the auditory space of Culex pipiens female mosquitoes in 3D. Insects 14, 743. doi: 10.3390/insects14090743
Legett H. D., Baranov V. A., and Bernal X. E. (2018). Seasonal variation in abundance and diversity of eavesdropping frog-biting midges (Diptera, Corethrellidae) in a neotropical rainforest. Ecol. Entomol. 43, 226–233. doi: 10.1111/een.12492
McGregor B. L., Runkel I. V. A. E., Wisely S. M., and Burkett-Cadena N. D. (2018). Vertical stratification of Culicoides biting midges at a Florida big game preserve. Parasites Vectors 11, 505. doi: 10.1186/s13071-018-3080-5
McKeever S. and French F. E. (1991). Corethrella (Diptera, Corethrellidae) of eastern North America - laboratory life-history and field responses to Anuran calls. Ann. Entomol. Soc Am. 84, 493–497. doi: 10.1093/aesa/84.5.493
Menda G., Nitzany E. I., Shamble P. S., Wells A., Harrington L. C., Miles R. N., et al. (2019). The long and short of hearing in the mosquito Aedes aEgypti. Curr. Biol. 29, 709–714. doi: 10.1016/j.cub.2019.01.026
Müller P. and Robert D. (2001). A shot in the dark: the silent quest of a free-flying phonotactic fly. J. Exp. Biol. 204, 1039–1052. doi: 10.1242/jeb.204.6.1039
Oksanen J., Blanchet F., Kindt R., Legendre P., Minchin P., and O’Hara R. (2015). “vegan: community ecology package,” in R package version 2.3-0.
Pantoja-Sánchez H., Leavell B. C., Rendon B., de-Silva W. A. P. P., Singh R., Zhou J., et al. (2023). Tiny spies: mosquito antennae are sensitive sensors for eavesdropping on frog calls. J. Exp. Biol. 226. doi: 10.1242/jeb.245359
Puillandre N., Brouillet S., and Achaz G. (2021). ASAP: assemble species by automatic partitioning. Mol. Ecol. Resour. 21, 609–620. doi: 10.1111/1755-0998.13281
Sigl M., Virgo J., Grafe U., Henske J., Schweinsberg M., and Eltz T. (2025). Uninvited guests: diversity and specificity of Trypanosoma infections in frog-biting midges (Corethrella spp.). Parasites Vectors 18, 348. doi: 10.1186/s13071-025-06993-8
Srivathsan A., Feng V., Suárez D., Emerson B., and Meier R. (2023). ONTbarcoder 2.0: rapid species discovery and identification with real-time barcoding facilitated by Oxford Nanopore R10.4. Cladistics 40, 192–203. doi: 10.1111/cla.12566
Swanson D. A. and Adler P. H. (2010). Vertical distribution of haematophagous Diptera in temperate forests of the southeastern U.S.A. Med. Vet. Entomol. 24, 182–188. doi: 10.1111/j.1365-2915.2010.00862.x
Virgo J. and Eltz T. (2022). Feeding site specificity in frog-biting midges (Corethrellidae). J. Insect Behav. 35, 155–159. doi: 10.1007/s10905-022-09807-2
Virgo J., Ruppert A., Lampert K. P., Grafe T. U., and Eltz T. (2019). The sound of a blood meal: acoustic ecology of frog-biting midges (Corethrella) in lowland Pacific Costa Rica. Ethology 125, 465–475. doi: 10.1111/eth.12871
Virgo J., Ufermann L., Lampert K. P., and Eltz T. (2021). More than meets the eye: decrypting diversity reveals hidden interaction specificity between frogs and frog-biting midges. Ecol. Entomol. 47, 95–108. doi: 10.1111/een.13095
Keywords: acoustic ecology, ectoparasites, insect hearing, Nematocera, Culicomorpha
Citation: Eltz T, Kimm HL, Böddicker R, Sigl M, Meier R and Virgo J (2025) Look up! Stratification and vertical approach to auditory prey cues in frog-biting midges (Corethrellidae). Front. Ecol. Evol. 13:1705905. doi: 10.3389/fevo.2025.1705905
Received: 15 September 2025; Accepted: 29 October 2025;
Published: 09 December 2025.
Edited by:
Fabio Santos Nascimento, University of São Paulo, BrazilReviewed by:
Eduardo De Farias Geisler, National Institute of Amazonian Research (INPA), BrazilLuiz Carmo, Federal University of Rio de Janeiro, Brazil
Copyright © 2025 Eltz, Kimm, Böddicker, Sigl, Meier and Virgo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thomas Eltz, dGhvbWFzLmVsdHpAcnViLmRl
‡ORCID: Thomas Eltz, orcid.org/0000-0002-8465-7944
Maximilian Sjgl, orcid.org/0009-0004-3484-0021
Rudolf Meier, orcid.org/0000-0002-4452-2885
Jonas Virgo, orcid.org/0000-0003-4395-7737