 Cecilia Ferreira de Mello1
Cecilia Ferreira de Mello1 Letícia de Mattos Pavão1,2
Letícia de Mattos Pavão1,2 Shayenne Olsson Freitas Silva1Amanda Bastos Queiroz1
Shayenne Olsson Freitas Silva1Amanda Bastos Queiroz1 Jeronimo Alencar1*
Jeronimo Alencar1*- 1Laboratório Diptera, Instituto Oswaldo Cruz (FIOCRUZ), Rio de Janeiro, Brazil
- 2Programa de Pós-Graduação em Biologia Animal, Instituto de Biologia, Universidade Federal Rural do Rio de Janeiro, Seropédica, Brazil
The structural complexity of host plants plays a significant role in shaping the heterogeneity of phytotelmata, influencing the taxonomic composition and ecological dynamics of associated communities, including medically important organisms such as vector mosquitoes. These microhabitats perform essential ecological functions, such as facilitating nutrient cycling and regulating insect populations, thereby contributing to the stability and conservation of biodiversity in forest environments. In this context, the present study investigated the population fluctuation and species composition of culicids using seven Lecythis pisonis (sapucaia) fruits as natural traps in an Atlantic Forest fragment located in the municipality of Silva Jardim, state of Rio de Janeiro, Brazil, between January 2023 and December 2024. A total of 1,235 specimens were collected. Statistical analyses included the Kruskal-Wallis test followed by Dunn’s post-hoc test. Significant differences in species abundance and richness were observed among the different fruit-traps, which were associated with microenvironmental factors such as light exposure, location, and water accumulation. Aedes albopictus (Skuse, 1895) exhibited clear dominance in a specific microhabitat, while Haemagogus leucocelaenus (Dyar & Shannon, 1924) showed higher prevalence in certain traps, reflecting an aggregated distribution in sites with distinct environmental characteristics and highlighting the importance of entomological surveillance in arbovirus risk areas. Trap 3 showed the highest richness and abundance, making it the most attractive microhabitat, whereas trap 5 demonstrated greater ecological balance among species. Notably, Ae. albopictus and Hg. leucocelaenus, both known vectors of human pathogens, were found among the species collected. The sampling method using sapucaia fruits proved effective, providing a practical and sustainable alternative for monitoring culicid populations in natural environments. The results contribute to our understanding of vector ecology and may inform surveillance and control strategies for mosquito-borne pathogens, as well as support integrated public health and biodiversity conservation actions.
Introduction
Knowledge of mosquito diversity in Atlantic Forest remnants is essential to under-standing changes in the activity patterns and behavior of vector populations, particularly in response to environmental changes caused by human activity. Faunistic studies of culicids have direct implications for public health, as they support the monitoring of etiological agents of epidemiological relevance, such as the viruses responsible for dengue, Zika, chikungunya, and yellow fever (YF), as well as the protozoa that cause malaria (Forattini, 2002; WHO, 2019).
Identifying the presence and estimating the population density of vector mosquitoes allows for the anticipation of outbreaks and the implementation of preventive measures before pathogens begin to spread. Early detection of vector species and assessment of their population abundance provide critical information to predict potential outbreaks and guide timely intervention strategies. Habitat fragmentation tends to favor opportunistic species, which are more adaptable to altered environments, while specialist species that depend on specific ecological conditions become more vulnerable to population declines and local extinction (Fahrig, 2003).
Microclimatic changes along the edges of forest fragments, known as edge effects, can significantly alter environmental conditions such as temperature, humidity, and sunlight exposure, directly impacting mosquito breeding habitats (Reisen, 2010). These changes make culicid populations more vulnerable to environmental stressors like dehydration and predation, which can compromise their survival and dynamics (Laurance et al., 2007).
Fahrig (2003) states that habitat fragmentation not only reduces the total available area but also alters the spatial configuration of remnants, which directly influences species composition and richness. These changes are especially critical for biological groups sensitive to environmental disturbances, such as culicids, whose populations may respond differently to landscape structure changes, often resulting in impoverished communities or dominance by opportunistic species in more degraded areas (Patz et al., 2000).
These variations are important for identifying potential behavioral shifts in species originally native to the wild that, faced with human pressures like urbanization and land-use changes, may adapt to altered environments such as peri-urban areas and re-generating forests (Mckinney, 2006). This pattern is especially evident in tropical and subtropical regions where mosquitoes, due to their ecological plasticity, thrive in a wide variety of larval habitats. Among these, phytotelmata, small water bodies formed by plant structures, play a key role in the proliferation of medically important species like Aedes and Haemagogus (Forattini, 2002; Frank and Lounibos, 2009; Marcondes and Alencar, 2010).
These microhabitats, found in plant structures such as leaf axils, tree holes, fruits, flowers, pods, and bamboo stems, offer ideal microclimatic conditions for the development of immature culicids (Kitching, 2000). Ngai and Srivastava (Ngai and Srivastava, 2006) showed that bromeliad eco-systems not only sustain aquatic communities but also fulfill essential ecological roles.
The structural diversity of host plants directly influences the heterogeneity of phytotelmata (Frank and Lounibos, 2009), which in turn affects the composition and dynamics of the associated communities, including mosquito species of epidemiological importance (Forattini, 2002). In addition to serving as breeding sites, these environments perform essential ecological functions, such as nutrient cycling and the trophic regulation of insect populations (Frank and Lounibos, 1983), contributing significantly to the maintenance of biodiversity in tropical forest ecosystems (Ngai and Srivastava, 2006).
Culicid species that breed in phytotelmata exhibit selectivity and specific adaptations for oviposition and the development of their immature stages. The structure of the larval habitat influences key factors, including development time, survival, and feeding behavior. With the increasing need to improve mosquito population monitoring in natural environments, particularly in light of the ecological diversity of these ecosystems and the presence of pathogen-vector species, it is essential to develop and implement collection methods that are both environmentally compatible and effective in sampling immature forms.
Using Lecythis pisonis Cambess. (sapucaia) fruits as natural traps is a promising alternative for entomological studies. Their hollow and durable structure provides an ideal microhabitat for the oviposition of various culicid species, closely simulating the natural conditions of phytotelm breeding sites. However, a central issue that arises is the extent to which sapucaia fruits can function as an effective and innovative tool for studying the diversity of immature culicids in Atlantic Forest remnants. In this regard, and considering the high ecological and epidemiological relevance, the present study aimed to compare and identify the diversity of immature culicids collected with sapucaia traps in a remnant of the Atlantic Forest in the municipality of Silva Jardim, state of Rio de Janeiro, Brazil (Lima Bersot et al., 2023).
The use of L. pisonis capsules as an alternative method for collecting immature mosquito stages may improve the understanding of culicid community structure in forested areas, particularly regarding medically significant species, and contribute to strengthening entomological surveillance efforts.
Materials and methods
Study area
The study was conducted in an Atlantic Forest fragment located at Renascente Farm (22°36’49.1”S, 42°27’34.1”W), in the central-northern region of the state of Rio de Janeiro, within the municipality of Silva Jardim (Figure 1). The study area lies within the critical habitat of the golden lion tamarin, Leontopithecus rosalia (Linnaeus, 1766), an endemic and threatened primate of the Atlantic Forest that was severely affected during the 2016–2018 YFV outbreak (Abreu et al., 2019). In this epidemiological context, the region is particularly important for investigating the dynamics of culicid vectors across different forest habitats. According to updated data from the Fundação SOS Mata Atlântica and INPE (Fundação SOS Mata Atlântica & Instituto Nacional de Pesquisas Espaciais, 2021), approximately 18.6% of the state’s original Atlantic Forest vegetation cover remains in forest remnants, representing an important natural heritage and one of the last refuges of biodiversity within this global hotspot.
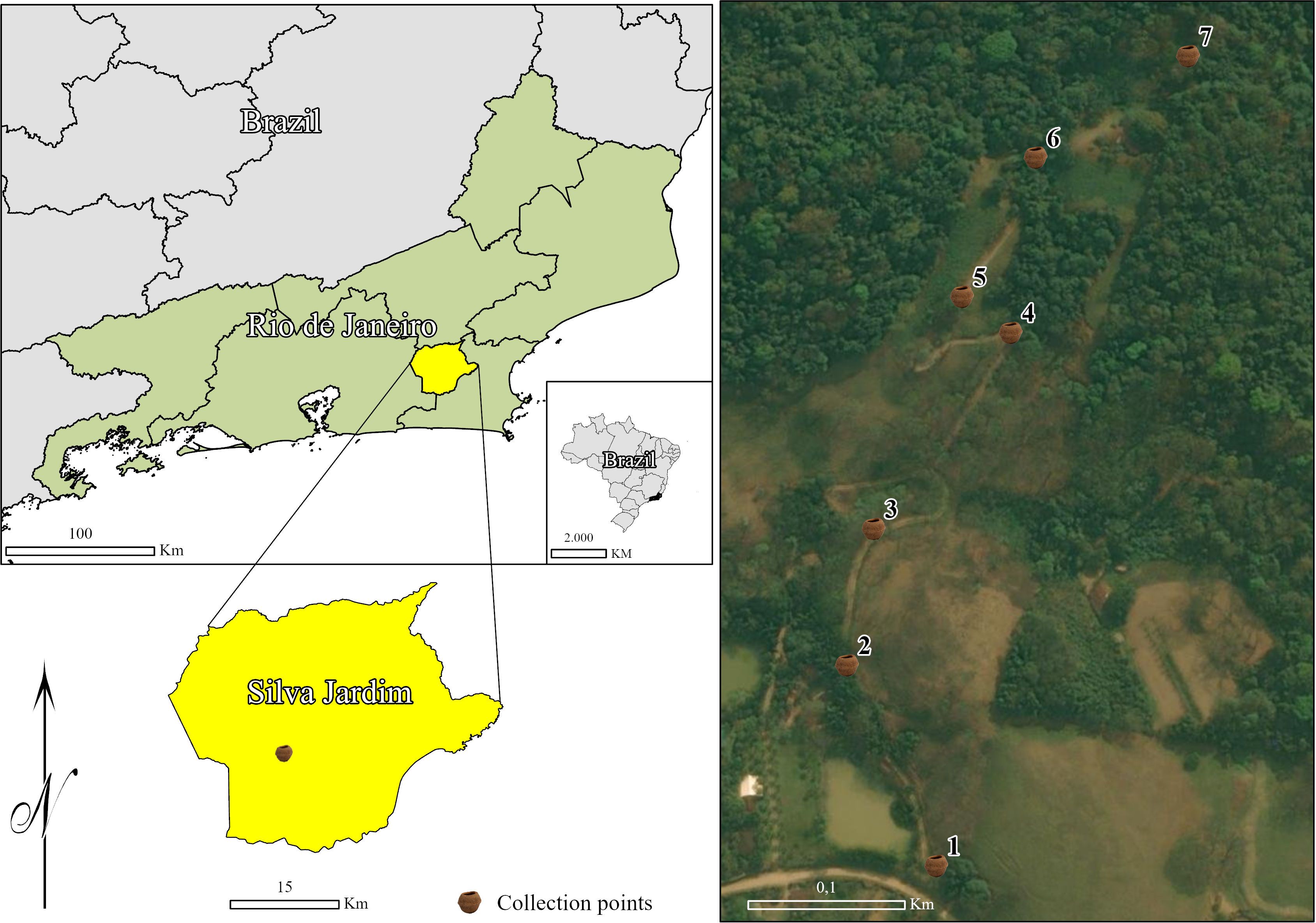
Figure 1. Sampling site at Renascente Farm, located in the central-northern region of the state of Rio de Janeiro, within the municipality of Silva Jardim, Brazil.
The predominant vegetation in the Poço das Antas Biological Reserve, also located in the municipality of Silva Jardim, includes characteristic vegetation formations of the Atlantic Dense Ombrophilous Forest, such as submontane, alluvial, and pioneer types, as well as secondary growth at various successional stages (Guedes-Bruni et al., 2006). These vegetation types provide diverse microhabitats that favor faunal diversity, including vector insects such as culicids.
The local climate, according to the Köppen-Geiger classification, is tropical with a dry winter season (Aw), characterized by a rainy summer and a dry winter period. The average annual rainfall is approximately 1,400 mm, and average temperatures range from 18°C to 23°C (INMET, 2025). These climatic conditions, combined with the environmental heterogeneity of forest fragments, create a favorable setting for the reproduction and proliferation of vector mosquito populations, especially during the warmer and more humid months of the year.
This ecological setting highlights the importance of the study area for exploring culicid population dynamics and their role in pathogen transmission. It also helps deepen our understanding of the ecological processes driving the emergence and reemergence of arboviruses in forested habitats and transition zones between natural and human-impacted environments.
Collections were conducted monthly on a single day between 9:00 a.m. and 4:00 p.m., from January 2023 to December 2024. This sampling design enabled continuous monitoring of the mosquito fauna across seasonal variations.
Natural traps (sapucaia)
To sample the culicid fauna, natural traps were prepared using capsules of the tree L. pisonis, commonly known as “sapucaia” or “monkey-pot.” Lecythis pisonis is a large tree that can reach 20 to 30 m in height (Figure 2) (Santos et al., 2020; Vieira et al., 2020). Its leaves are leathery and glabrous, showing a reddish-pink hue during the flowering period. The flowers are violet, and the urn-shaped capsules contain between 10 and 30 seeds (nuts) (Lima Bersot et al., 2023). The sapucaia trap is designed as an artificial analogue of natural phytotelmata, specifically tree holes. It intentionally mimics the preferred ecosystem for oviposition and larval development of species in the tribe Aedini (e.g., Aedes spp. and Haemagogus spp.) in wild or peri-urban settings. Its effectiveness is therefore evaluated based on its dual capacity to recreate attractive conditions, since fruit decomposition releases specific volatiles that lure gravid females adapted to these natural microhabitats, and to support complete development by providing a viable environment that allows egg hatching, larval development, pupation, and adult emergence in situ. This functional design distinguishes the sapucaia trap from standard ovitraps.

Figure 2. Natural trap of the tree Lecythis pisonis, commonly known as “sapucaia.” (A) Metal wire; (B) original operculum of the fruit; (C) capsule surface with an asphalt membrane.
The capsules used were globose, with diameters ranging from 12 to 18 cm, based on morphological descriptions of the species reported in previous botanical studies (Prance and Mori, 1979; ITTO, 2025; NPARKS, 2025). Each trap was prepared by covering the outer surface of the capsule with an asphalt membrane to prevent permeability through the natural walls and minimize water loss by infiltration. The original operculum of the fruit was used as a lid, positioned to remain slightly open, allowing access for females to oviposit while simultaneously reducing the evaporation rate of the water accumulated inside. For field deployment, a metal wire was attached to opposite sides of the capsule’s edge near the opening, forming a small hanging basket and allowing the traps to be easily secured to branches or tree trunks (Figure 2).
A total of seven traps were installed. Although the internal volume varied among the capsules used, each was standardized by filling it with 500 mL of clean water. The traps were spaced at least 100 m apart and hung vertically on the trunks of seven trees, approximately 2 m above the ground, aiming to minimize environmental interference and ensure consistent sampling.
The use of natural traps stems from the ecological relevance of phytotelmata, which are aquatic microhabitats formed by cavities in plants that serve as natural breeding sites for various mosquito species. This approach offers greater ecological relevance to the methodology by simulating real oviposition conditions found in forest environments, thereby enhancing attractiveness to wild species and improving the effectiveness of data collection.
Collection, maintenance, and identification of immature stages
The immature mosquito forms (larvae and pupae) found in the traps were collected using sterile plastic pipettes and transferred to 1-L round plastic containers with screw caps (Plasvale, Santa Catarina, Brazil), properly labeled with information on the date and collection site. These containers were used for transporting and temporarily maintaining the specimens until sorting at the laboratory. During transport, the containers were kept in insulated coolers to preserve sample integrity and minimize thermal stress.
In the laboratory, the immature specimens were kept in plastic containers filled with the same water collected from the traps, with periodic replacement using dechlorinated water as needed to preserve the original physicochemical characteristics of the micro-habitat. The immature mosquitoes developed to the adult stage in a Biochemical Oxygen Demand (B.O.D.) climate chamber (Eletrolab, São Paulo, Brazil) under controlled conditions of temperature (28 ± 1°C), relative humidity between 75% and 90%, and a 12-hour light/12-hour dark photoperiod.
The larvae were fed twice a week with commercial fish food (TetraMin®, Tetra GmbH, Niedersachsen, Germany), which was previously ground and diluted in 10 mL of water to ensure adequate nutrient supply for proper larval development.
After emergence, the adult specimens were euthanized using chloroform and mounted on entomological pins for subsequent morphological identification. Species were identified through direct observation of morphological characteristics under a stereoscopic microscope (Leica DMD108®), based on consultation of specific descriptions and diagnoses using classical and updated dichotomous keys (Lane, 1953; Arnell, 1973; Consoli and Oliveira, 1994; Forattini, 2002; Marcondes and Alencar, 2010). Generic and subgeneric name abbreviations followed the standardization proposed by Reinert (Reinert, 2009). All specimens have been cataloged and are awaiting the assignment of voucher numbers by the Oswaldo Cruz Institute Entomological Collection (Fiocruz). Once confirmed, these numbers will be made publicly available and included in the supplementary material, ensuring long-term preservation, traceability, and accessibility for future research.
Statistical analyses
Data were analyzed using the Kruskal-Wallis test followed by Dunn’s post-hoc test, with a significance level of 5% (p < 0.05), to identify significant differences in species composition and abundance between collection months and among traps. To evaluate the diversity and structure of the culicid community, several ecological indices were calculated: the Shannon-Wiener Index (H′) to estimate diversity; Pielou’s Evenness Index (J′) to assess the uniformity of species distribution; Simpson’s Index (D) to measure ecological dominance; Species Richness (S) to count the total number of taxa identified per trap and month; and Relative Abundance to indicate the proportion of individuals of each species relative to the total catch. These metrics allowed us to compare trap effectiveness and changes in the mosquito fauna over time, considering seasonal and microenvironmental variations. All analyses were performed using PAST software (version 4.05).
Results
Statistical analyses revealed significant differences in culicid diversity and abundance among the sapucaia traps. The Kruskal-Wallis test followed by Dunn’s post-hoc test showed that trap 3 differed significantly from traps 1 and 6 (p = 0.04 for both). These differences are likely due to the higher species richness observed in trap 3 (S = 12), which was notably greater than in traps 1 and 6 (S = 5 each). Additionally, trap 3 had the highest abundance, with 273 specimens collected.
Trap 5 showed the second highest species richness (S = 10) along with the highest diversity (H = 2.06) and evenness (J = 0.89) indices, suggesting a more uniform distribution of the collected species. In contrast, trap 1 exhibited the highest dominance index (D = 0.74), with Aedes albopictus predominating, representing 86% of the specimens collected in this trap.
Traps 2, 5, and 7 showed similar species richness, abundance, and diversity, suggesting homogeneity in environmental conditions or trap structure. The most abundant species collected in the traps were Sabethes cyaneus (Fabricius, 1805) (n = 269), Ae. albopictus (n = 175), Aedes terrens (Walker, 1856) (n = 116), and Culex pleuristriatus Theobald, 1903 (n = 100). Notably, Haemagogus leucocelaenus, a known vector of the yellow fever virus (YFV), was also present, with 22 individuals collected in trap 6 and 48 in trap 7, highlighting the importance of natural traps for monitoring species of epidemiological interest. Furthermore, Sabethes albiprivus Theobald, 1903, recognized as a secondary vector of YFV and confirmed to be susceptible to infection, was also registered in the collections (trap 1: n = 1; trap 3: n = 4), as detailed in Table 1. These findings highlight the effectiveness of sapucaia traps for collecting immature mosquitoes and their value in monitoring species diversity and abundance in Atlantic Forest remnants. Differences observed between traps may be influenced by factors such as host plant structure, water availability, and closeness to ecotones between natural and disturbed habitats.
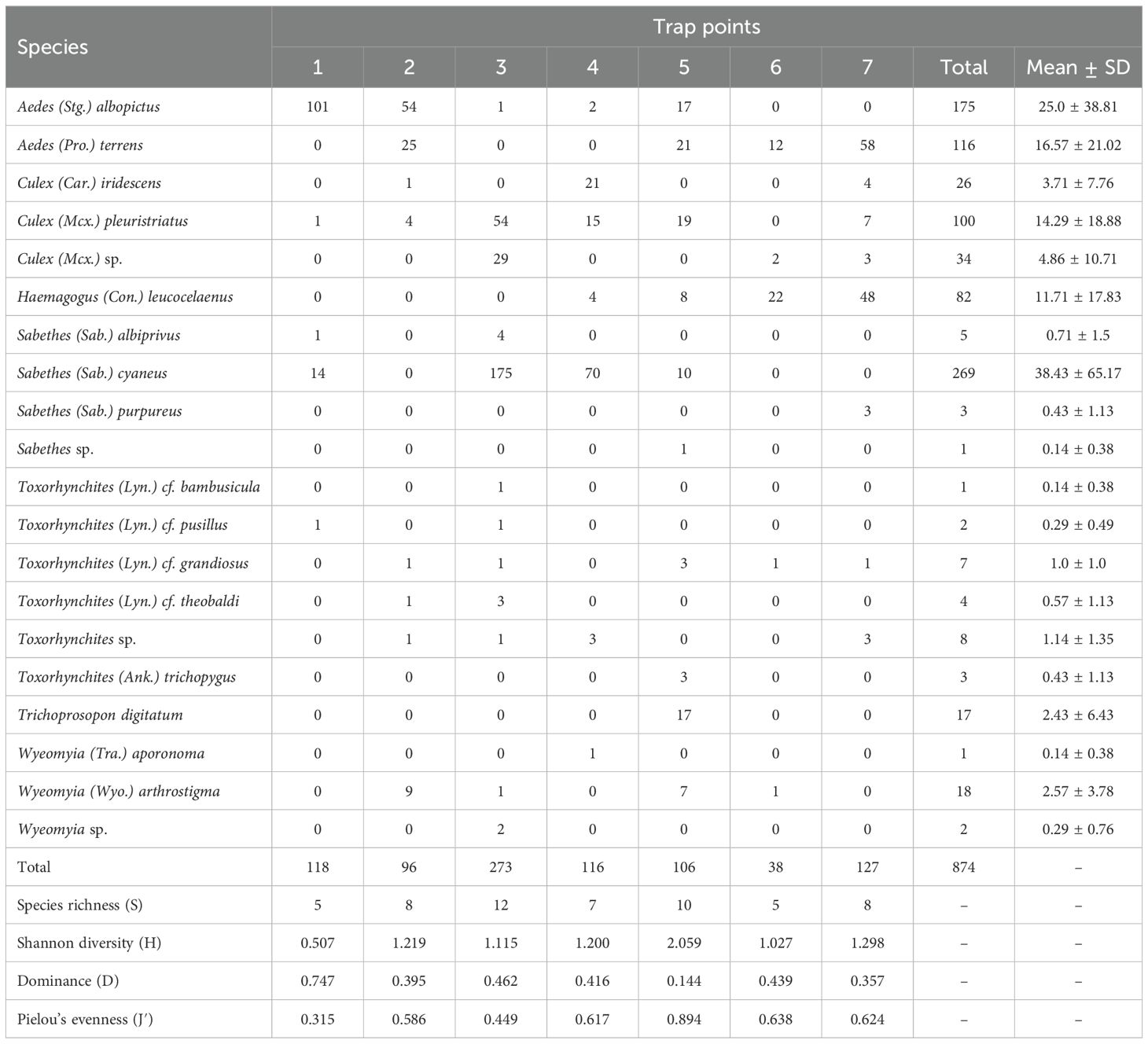
Table 1. Abundance of culicid species collected at Renascente Farm, municipality of Silva Jardim, state of Rio de Janeiro, Brazil, from January 2023 to December 2024.
Trap 1 was dominated by Ae. albopictus (n = 101) and Sa. cyaneus (n = 14), accounting for 86% and 12% of all culicids collected in this trap, respectively. Similarly, trap 2 showed a predominance of Ae. albopictus (n = 54; 56%), followed by Ae. terrens (n = 25; 26%), and Wyeomyia arthrostigma (Lutz, 1905) (n = 9; 9%). In contrast, traps 3 and 4 were characterized by higher abundances of Sa. cyaneus, with 175 individuals (64%) and 70 individuals (60%), respectively. The second most abundant species in trap 3 was Cx. pleuristriatus (n = 54; 20%), whereas in trap 4, the most representative species were Culex iridescens (Lutz, 1905) (n = 21; 18%) and Cx. pleuristriatus (n = 15; 13%).
Additionally, trap 3 exhibited the highest richness of Toxorhynchites species (Tx. cf. (Lyn.) bambusicula, Tx. cf. (Lyn.) pusillus, Tx. cf. grandiosus, and Tx. cf. theobaldi), all predatory larvae. Trap 5 exhibited a high evenness index, with a relatively balanced distribution of species. Notable representatives included Ae. albopictus (n = 17; 16%), Ae. terrens (n = 21; 20%), Cx. pleuristriatus (n = 19; 18%), Hg. leucocelaenus (n = 8; 8%), Sa. cyaneus (n = 10; 9%), Trichoprosopon digitatum (Rondani, 1848) (n = 17; 16%), and Wy. arthrostigma (n = 7; 7%). Traps 6 and 7 were noteworthy for their high abundance of Hg. leucocelaenus, an important YFV vector, with 22 individuals (58%) collected in trap 6 and 48 individuals (38%) in trap 7. In trap 6, Ae. terrens was the second most abundant species (n = 12; 32%), whereas in trap 7 it was the most abundant overall, with 58 individuals (46%) (Figure 3).
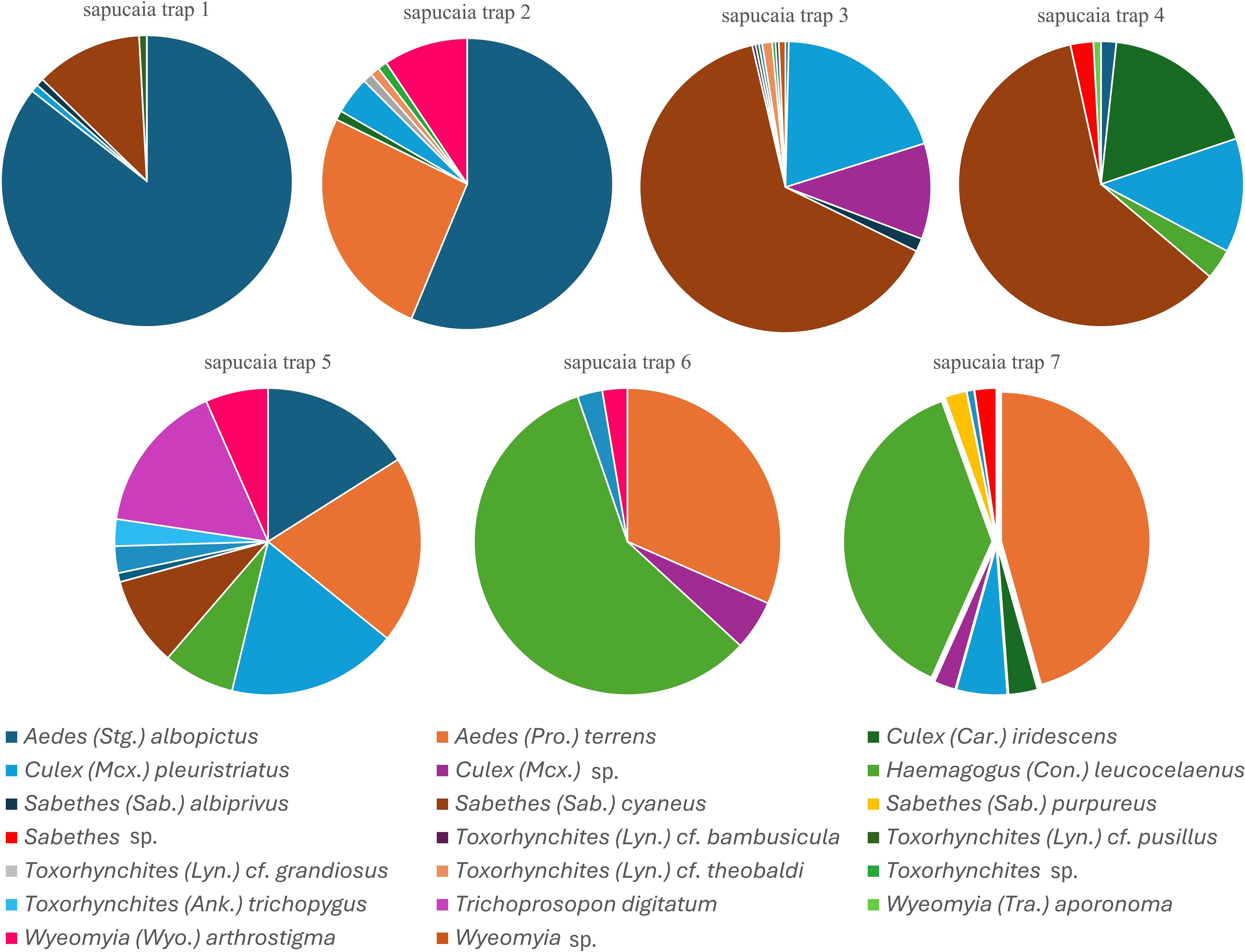
Figure 3. Abundance of culicid species per sapucaia trap (1–7) at Renascente Farm, municipality of Silva Jardim, state of Rio de Janeiro, Brazil, from January 2023 to December 2024.
Seasonal Fluctuation of Mosquito Populations During the Collection Period (2023–2024).
The mosquito population showed marked seasonal fluctuations throughout the collection period, from January 2023 to December 2024. In both years, the numbers recorded in January were relatively low, a pattern likely associated with reduced re-productive activity during the drier and cooler months at the beginning of the year. From February onward, however, there was a consistent increase in abundance, reflecting the influence of more favorable climatic conditions such as rising temperatures and intensified rainfall, which are typical of the summer season in the study region.
In March 2023, a decline in mosquito abundance was recorded, whereas the same month in 2024 marked the highest population peak of the entire study period, with 256 individuals collected. This year-to-year difference may be attributed to interannual climatic variation, changes in the availability of breeding sites, or temporary microenvironmental shifts within the sampled habitat.
In 2023, the main population peaks occurred in February (n = 110) and April (n = 49), with secondary increases observed in June (n = 28) and September (n = 22). In 2024, in addition to the pronounced peak recorded in March (n = 256), moderate increases were observed in February (n = 65), April (n = 37), and May (n = 34), indicating that seasonal factors influenced population dynamics differently across the two consecutive years.
There was a widespread decline in mosquito population density during the dry season (May to October), a pattern expected due to the reduced availability of natural breeding sites and lower relative humidity. However, in November 2024, a notable increase in abundance was recorded (n = 98), suggesting the start of a new reproductive cycle triggered by the return of rainfall at the end of spring.
Overall, considering the combined data from both years, the months with the highest mosquito abundance were February (n = 175), March (n = 281), and November (n = 108). Conversely, the lowest abundances were recorded in June (n = 14), July (n = 7), and October (n = 10). Statistical analysis revealed significant differences in population abundance between certain months. February differed significantly from July (p = 0.03), August (p = 0.01), and October (p = 0.02). March also showed significant differences compared to August (p = 0.02) and October (p = 0.03). Additionally, April differed significantly from August (p = 0.03) (Figure 4).
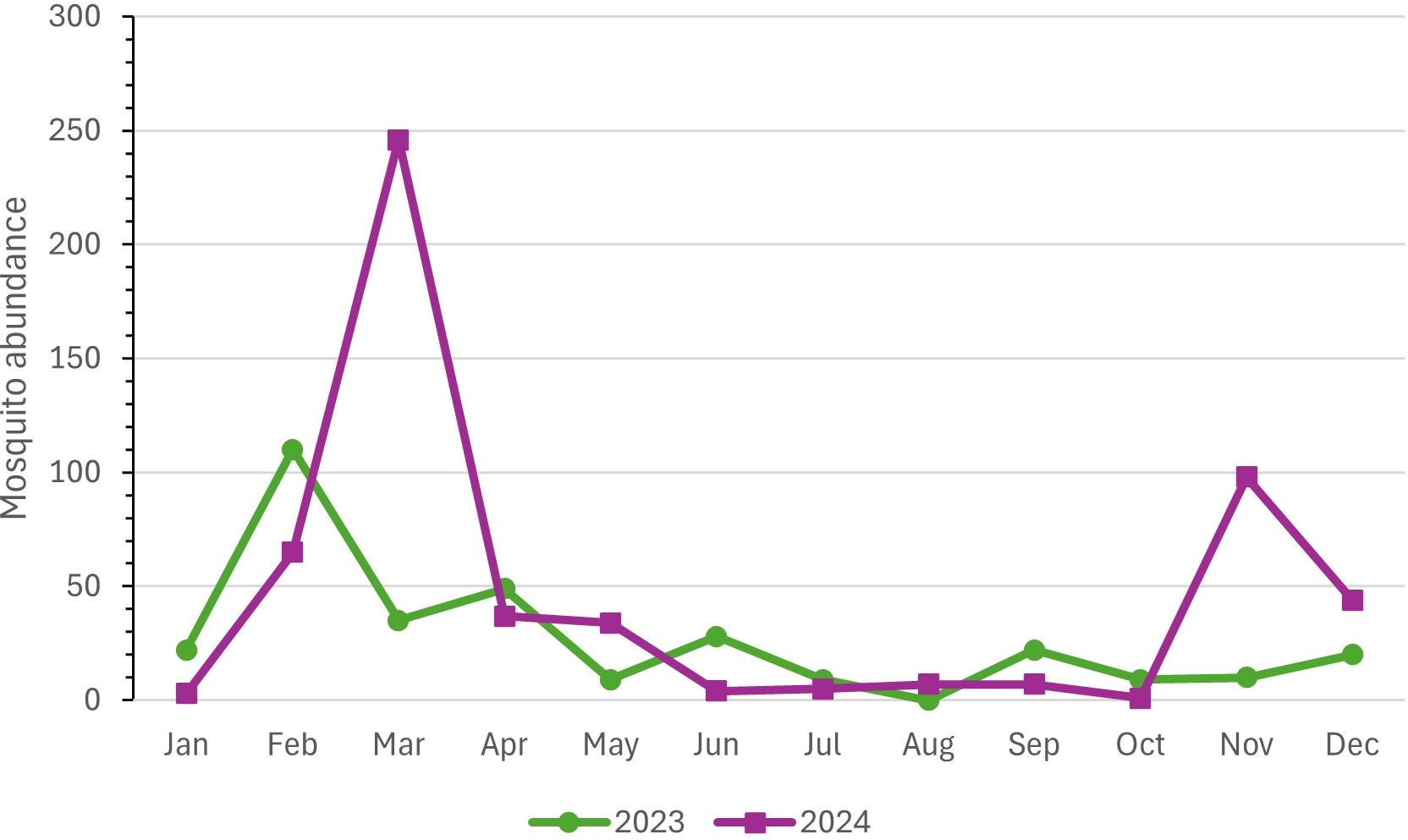
Figure 4. Abundance of mosquitoes collected from all traps at Renascente Farm, municipality of Silva Jardim, state of Rio de Janeiro, from January 2023 to December 2024.
These results indicate that the mosquito population exhibits marked seasonality, with population peaks mainly concentrated at the end of summer and the beginning of autumn. This pattern suggests an influence of climatic factors such as temperature and rainfall on the population dynamics of these insects (Table 2).
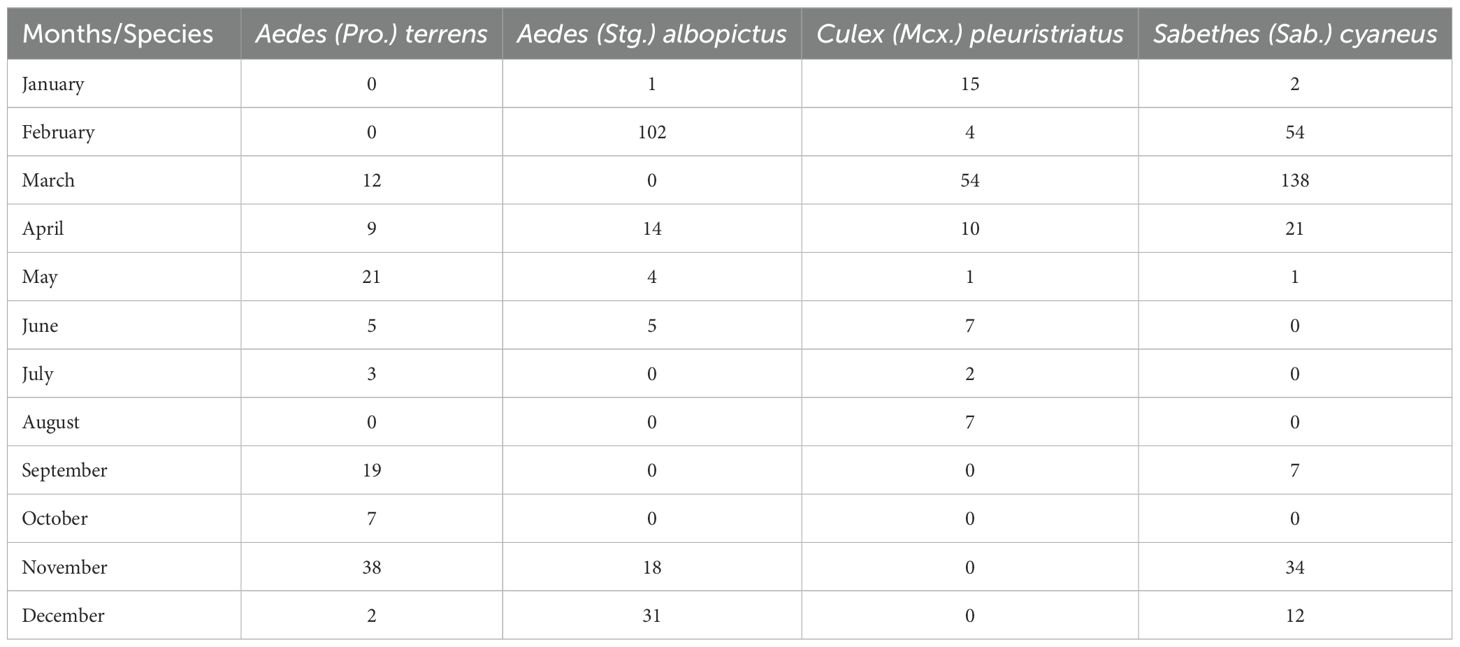
Table 2. Most abundant culicid species collected per month at Renascente Farm, municipality of Silva Jardim, state of Rio de Janeiro, Brazil, from January 2023 to December 2024.
Discussion
The present results reveal statistically significant differences in the composition and structure of mosquito communities sampled using sapucaia traps, underscoring the role of microenvironmental factors in shaping mosquito diversity and abundance in Atlantic Forest remnants.
Furthermore, the configuration of the operculum opening emerged as a crucial factor influencing the composition of the sampled mosquito community. The observation that Aedini species such as Hg. leucocelaenus, Ae. albopictus, and Ae. terrens, which are known for laying eggs on the inner surface of breeding sites, colonized the traps when the lid was dislocated or dropped corroborates previous findings. Vieira et al. (2020) reported that the fauna differs markedly when the operculum is “closing,” leaving only a small perforated opening. Our results reinforce that the degree of operculum opening acts as a fine-scale environmental filter, actively selecting for species with distinct oviposition behaviors and, consequently, directly influencing the species composition observed compared to other studies employing different operational protocols.
Our findings provide new insights into mosquito ecology in the Atlantic Forest by demonstrating the critical influence of fine-scale environmental heterogeneity, a factor often unmeasured in broader surveys. The significant variation in community composition between closely placed traps, revealed by our method, challenges conventional habitat classifications and underscores the patchy distribution of mosquitoes even within preserved forest fragments. Achieving this level of ecological resolution with a low-cost, naturalistic sampling tool represents a key contribution of our study, highlighting the utility or our method in identifying micro-environmental preferences that are crucial for targeted surveillance.
Trap 3 showed the highest species richness (S = 12) and abundance (n = 273) among those evaluated, contrasting significantly with traps 1 and 6 (both with S = 5). Although no specific environmental characteristics were identified in the immediate surroundings of this trap, such as different patterns of water accumulation, vegetation structure, or geographic position, it was more exposed to sunlight than the others. This suggests that even subtle and not fully characterized environmental heterogeneity can directly influence the composition and structure of the mosquito community. Greater sun exposure may be associated with microvariations in temperature, humidity, and water evaporation, which in turn may affect the trap’s attractiveness or suitability for different species (Chaves et al., 2011; Ferraguti et al., 2016).
This heterogeneity is recognized in the literature as a key factor for maintaining diversity in insect communities, including mosquito vectors (Chaves et al., 2011). Therefore, even without a detailed characterization of environmental variables, the data reinforce the importance of considering microenvironmental variation when interpreting mosquito distribution and diversity patterns, emphasizing the need to incorporate more precise ecological measurements in future research.
The high Pielou’s evenness index (J = 0.89) observed in trap 5, along with the highest Shannon diversity index (H = 2.06), indicates a more ecologically stable environment where multiple species coexist without marked dominance. This pattern is commonly associated with habitats that have a balanced distribution of resources and favorable environmental conditions (Magurran, 2021). In contrast, trap 1 showed dominance of Ae. albopictus (86%), a highly adaptable species often linked to anthropized areas (Juliano and Philip Lounibos, 2005). The abundance of this species may reflect local environmental changes, such as increased sunlight exposure, reduced vegetation cover, and/or greater human influence.
While Ae. albopictus predominated in traps 1 and 2, typically sylvatic species such as Sa. cyaneus and Hg. leucocelaenus were more abundant in traps 3, 4, 6, and 7, suggesting the coexistence of environments with varying degrees of ecological conservation. The notable presence of Hg. leucocelaenus, a recognized vector of the sylvatic YFV in forested areas, serves as a warning for the potential risk of reemergence of this arbovirus, as observed in recent outbreaks in Brazil (Cardoso et al., 2010; Possas et al., 2018).
The present analysis revealed a diverse Toxorhynchites community, with five species recorded (Tx. cf. bambusicula, Tx. cf. pusillus, Tx. cf. grandiosus, Tx. cf. theobaldi, and Tx. trichopygus). However, this diversity contrasts with the low abundance of specimens observed, a pattern previously reported in specific microhabitats (Lozovei, 1998). This low density is characteristic of the genus and is attributed to its specialized ecology and population regulation mechanisms, with larval intraspecific cannibalism being the predominant factor (Campos, 2013; Campos, 2016). As Toxorhynchites larvae are voracious predators, the absence of alternative prey often leads to conspecific predation, resulting in the occurrence of a single larva per larval habitat. This behavior acts as a self-regulating mechanism that severely limits the total number of adults. Thus, population dynamics are influenced by the availability of larvae from other mosquito species, where fluctuations in target populations producing even more pronounced oscillations in Toxorhynchites counts (Zequi and Lopes, 2001). Despite this density limitation, species diversity remains ecologically significant, as these mosquitoes play an important role in biological control (Medeiros-Sousa et al., 2013). The coexistence of multiple species suggests niche complementarity, enabling the community to regulate vector mosquito populations across a wider range of microhabitats and highlighting the need for a deeper understanding of the biology of these species for effective use in biocontrol programs (Collins and Blackwell, 2000). In direct counterpoint to the presence of these natural predators and their regulatory function, the presence of vector species in the area demonstrates the epidemiological complexity of the studied ecosystem.
The detection of Sa. albiprivus and Sa. purpureus underscores the importance of comprehensive entomological surveillance, which should not be limited to primary vectors (Haemagogus spp.) (Gomes et al., 2010; Conceição, 2024). The diversity of vectors indicates that the ecosystem is capable of supporting the sylvatic YF viral cycle, with Sa. albiprivus likely serving as an important secondary vector, corroborating studies reporting the role of this species in virus maintenance (Gomes et al., 2010; de Oliveira et al., 2023). For Sa. purpureus, although records of natural infection are less common, its detection highlights the need for future investigations (such as viral infection assays and laboratory studies on vector competence) to clarify its role in YF epidemiology in the area, particularly in comparison with other Sabethes species (Carvalho et al., 1997; Mascheretti et al., 2013). The co-occurrence of these vector species, combined with the presence of natural predators, increases the risk of accidental transmission to humans entering the forest, especially during epizootics in non-human primates (NHPs) (Vasconcelos, 2003).
The temporal variation in mosquito abundance highlights a seasonal population pattern, with peaks occurring mainly at the end of summer and the beginning of autumn, particularly in February, March, and November. This fluctuation is likely associated with climatic conditions favorable to mosquito reproduction, such as increased rainfall and higher temperatures (Honório et al., 2009; Morin and Comrie, 2013). Statistical analysis detected significant differences between months, suggesting that these peaks are not random but follow predictable patterns.
The use of sapucaia capsules as natural traps proved to be an effective and environmentally compatible tool for sampling immature mosquitoes in forest environments. This method offers several advantages, including simplicity, low cost, and the ability to simulate natural breeding sites, making it particularly useful for ecological studies and entomological surveillance programs.
Therefore, this study provides valuable insights into the ecology of pathogen vector species, their seasonal patterns, and interactions with the environment, contributing to more effective epidemiological and entomological surveillance strategies in tropical regions.
Our data show that environmental temperature and precipitation patterns between January 2023 and December 2024 significantly altered mosquito population dynamics, primarily by favoring the development of immature stages through the formation of optimal breeding habitats. Consequently, abundance peaks were observed February, March, and November. These findings highlight the strong influence of seasonal climatic factors and further confirm the established role of temperature and rainfall as key drivers of mosquito population fluctuations (Reinhold et al., 2018).
The increase in mosquito abundance in February and March, particularly in 2024 when the highest peak was recorded (n = 256 in March), coincides with the late summer period in the study region. During this time, high temperatures and frequent rainfall create ideal conditions for mosquito larval development (Mordecai et al., 2019). Previous studies have shown that these factors directly affect the availability of breeding sites and accelerate the mosquito life cycle (Christophers, 1960; Consoli and Oliveira, 1994). The sharp decline in populations during the winter months, especially from June to August, with July showing the lowest count (n = 7), reflects the unfavorable climatic conditions, such as lower temperatures and reduced humidity, which limit reproductive activity below 18 °C.
The population increase in November 2024 (n = 98) suggests the onset of a new reproductive cycle driven by rising temperatures and the return of rainfall, both characteristic of spring in the region. A study on the vertical behavior and temporal distribution of mosquitoes in an Atlantic Forest fragment in the state of Rio de Janeiro, using ovitraps for egg collection, presented results similar to those of the present study. The highest abundances of Hg. leucocelaenus and Hg. janthinomys were recorded in November and December 2020, December 2022, January and November 2023, and January and February 2024 (Silva et al., 2025). These findings corroborate our observations, in which population peaks occurred during warmer months. This pattern aligns with other studies that identified November as a critical transition period for the growth of arbovirus vector populations in the state of Rio de Janeiro, including those responsible for YF transmission (Silva et al., 2025).
The statistical analysis indicated significant seasonality, highlighted by differences between rainy/warm months and dry/cold months (García-Suárez et al., 2024). The differences observed between February and July, August, and October, as well as between March and the drier months, support the influence of abiotic factors on mosquito density. Increased rainfall promotes the reproduction of mosquito vectors capable of transmitting pathogens. When combined with other favorable epidemiological factors, this scenario can trigger epidemics (Vasconcelos et al., 2001). In this regard, the seasonality observed in our study highlights the relevance of entomological monitoring to anticipate pathogen transmission risks and support more effective vector control interventions.
The use of sapucaia fruits as natural traps proved to be an efficient, low-cost, and environmentally compatible strategy for sampling culicids in natural environments. This methodology significantly contributes to understanding the ecology of vector species, their distribution patterns, and their interactions with the environment. The present findings provide valuable support for developing entomological surveillance strategies, vector control, and biodiversity conservation in tropical areas. Additionally, they can guide integrated public health actions, especially in regions with a history or risk of arbovirus circulation.
This study reveals significant variations in mosquito species richness and abundance among the different sapucaia traps, which possibly reflect microenvironmental differences such as spatial location, light intensity, water accumulation and retention, as well as factors related to the surrounding vegetation structure. Trap 3 stood out as the most favorable environment for colonization, showing the highest indices of species richness and abundance. The high dominance of Ae. albopictus in trap 1 indicates a possible preference of this species for specific microenvironmental conditions, such as greater exposure to sunlight or increased interspecific competition in that trap. In contrast, trap 5 exhibited a more balanced species composition, suggesting greater ecological diversity and stability in the use of the microhabitat by different mosquito species.
Traps 6 and 7, which showed a notable presence of Hg. leucocelaenus, deserve special attention, as this species is associated with the transmission of arboviruses such as the sylvatic YFV. Its occurrence underscores the importance of continuous monitoring in forested areas with potential epidemiological risk. Its occurrence under-scores the importance of continuous monitoring in forested areas with potential epidemiological risk, where the collection of immature stages of mosquitoes specialized in these microhabitats represents a low-cost alternative that utilizes locally available materials.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
CM: Conceptualization, Methodology, Writing – original draft, Writing – review & editing. LP: Methodology, Writing – original draft, Writing – review & editing. SO: Formal analysis, Writing – original draft, Writing – review & editing. AQ: Conceptualization, Writing – original draft, Writing – review & editing. JA: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing.
Ethics statement
The permanent license for the collection, capture, and transport of zoological material was granted by the Chico Mendes Institute for Biodiversity Conservation (ICMBio) and the Authorization and Information System on Biodiversity (SISBIO) under license No. 84318-4.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; Grant No. 303286/2021-0, J.A.), the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ; Grants No. E-26/200.956/2022, Fellowship No. 2008.03683.9, J.A.; and E-26/205.649/2022, Fellowship No. 2012.04774.0, C.F.M.), and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). Author C.F.M. acknowledges FAPERJ for granting the Postdoctoral Fellowship Nota 10 (Process No. E-26/205.649/2022, Fellowship No. 2012.04774.0).
Acknowledgments
We thank Hercio Gomes and Tatiana Gomes for work support at the Renascente site in the central–north region of Rio de Janeiro, municipality of Silva Jardim, Rio de Janeiro State, Brazil.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abreu F. V. S. D., Ribeiro I. P., Ferreira-De-Brito A., Santos A. A. C. D., Miranda R. M. D., Bonelly I. D. S., et al. (2019). Haemagogus leucocelaenus and Haemagogus janthinomys are the primary vectors in the major yellow fever outbreak in Brazil 2016–2018. Emerging Microbes infections 8, 218–231. doi: 10.1673/031.013.930110.1080/22221751.2019.1568180
Arnell J. H. (1973). “Mosquito studies (Diptera, culicidae),” in XXXII. A revision of the genus Haemagogus (Contributions of the American Entomological Institute). 10, 1–174. Available online at: https://www.medigraphic.com/pdfs/revbio/bio-2010/bio103j.pdf
Campos R. E. (2013). The aquatic communities inhabiting internodes of two sympatric bamboos in Argentinean subtropical forest. J. Insect Sci. 13(1), 1–17. doi: 10.1673/031.013.9301
Campos R. E. (2016). Phytotelmata colonization in bamboo (Guadua sp.) culms in northeast Argentina. J. Natural History 50(15-16), 923–941. doi: 10.1080/00222933.2015.1091096
Cardoso J. D. C., De Almeida M. A. B., Dos Santos E., Da Fonseca D. F., Sallum M. A. M., Noll C. A., et al. (2010). Yellow fever virus in Haemagogus leucocelaenus and Aedes serratus mosquitoes, southern Brazil 2008. Emerging Infect. Dis. 16, 1918. doi: 10.3201/eid1612.100608
Carvalho M. E. S. D., Naves H. A. M., Carneiro E., and De Miranda M. F. (1997). Distribuição vertical de mosquitos dos géneros Haemagogus e Sabethes, em zona urbana de Goiânia-Goiás-Brasil. Rev. Patologia Tropical/Journal Trop. Pathol. 26(1), 1–3. doi: 10.5216/rpt.v26i1.17371
Chaves L. F., Hamer G. L., Walker E. D., Brown W. M., Ruiz M. O., and Kitron U. D. (2011). Climatic variability and landscape heterogeneity impact urban mosquito diversity and vector abundance and infection. Ecosphere 2, art70. doi: 10.1890/ES11-00088.1
Christophers S. (1960). Aëdes aEgypti (L.) the Yellow Fever Mosquito: its Life History, Bionomics and Structure (London: The Syndics of the Cambridge University Press, Bentley House, 200, Euston Road, N.W.I).
Collins L. E. and Blackwell A. (2000). “The biology of Toxorhynchites mosquitoes and their potential as biocontrol agents,” in Biocontrol news and information. Wallingford, Oxfordshire, UK. 21, 105N–116N. Available online at: https://www.cabi.org/bni/FullTextPDF/2000/20003030873.pdf
Conceição J. P. A. D. (2024). Risco de exposição a vetores da febre amarela silvestre (Diptera: Culicidae) no Parque Nacional de Brasília, Brasil. Parque Nacional de Brasília, Brasil: University of Brasília, Brasília. Available online at: http://repositorio2.unb.br/handle/10482/51083.
Consoli R. and Oliveira R. L. (1994). Principais mosquitos de importância sanitária no Brasil. Rio de janeiro (Brasil. Rio de janeiro: Editora da Fundação Oswaldo Cruz).
de Oliveira C. H., Andrade M. S., Campos F. S., Da C. Cardoso J., Gonçalves-Dos-Santos M. E., Oliveira R. S., et al. (2023). Yellow fever virus maintained by Sabethes mosquitoes during the dry season in Cerrado, a semiarid region of Brazil, in 2021. Viruses 15, 757. doi: 10.3390/v15030757
Fahrig L. (2003). Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecology Evolution Systematics 34, 487–515. doi: 10.1146/annurev.ecolsys.34.011802.132419
Ferraguti M., Martínez-De La Puente J., Roiz D., Ruiz S., Soriguer R., and Figuerola J. (2016). Effects of landscape anthropization on mosquito community composition and abundance. Sci. Rep. 6, 29002. doi: 10.1038/srep29002
Forattini O. P. (2002). Culicidologia médica: identificação, biologia, epidemiologia (Sâo Paulo: EDUSP).
Frank J. H. and Lounibos L. P. (1983). Phytotelmata: terrestrial plants as hosts for aquatic insect communities (Plexus Pub).
Frank J. H. and Lounibos L. P. (2009). Insects and allies associated with bromeliads: a review. Terrestrial Arthropod Rev. 1, 125–153. doi: 10.1163/187498308X414742
Fundação Sos Mata Atlântica, & Instituto Nacional de Pesquisas Espaciais (2021). Atlas dos remanescentes florestais da Mata Atlântica: período 2019/2020. (São Paulo: Fundação SOS Mata Atlântica/Instituto Nacional de Pesquisas Espaciais). Available online at: http://urlib.net/ibi/8JMKD3MGP3W34T/4C2J9PM.
García-Suárez O., Tolsá-García M. J., Arana-Guardia R., Rodríguez-Valencia V., Talaga S., Pontifes P. A., et al. (2024). Seasonal mosquito (Diptera: Culicidae) dynamics and the influence of environmental variables in a land use gradient from Yucatan, Mexico. Acta Tropica 257, 107275. doi: 10.1016/j.actatropica.2024.107275
Gomes A. D. C., Torres M. A. N., Paula M. B. D., Fernandes A., Marassá A. M., Consales C. A., et al. (2010). “Ecologia de Haemagogus e Sabethes (Diptera: Culicidae) em áreas epizoóticas do vírus da febre amarela,” in Epidemiologia e Serviços de Saúde, vol. 19, 101–113. (Rio Grande do Sul, Brasil: Epidemiologia e Serviços de Saúde). doi: 10.5123/S1679-49742010000200003
Guedes-Bruni R. R., Silva S. J. D., Morim M. P., and Mantovani W. (2006). Composição florística e estrutura de dossel em trecho de floresta ombrófila densa atlântica sobre morrote mamelonar na Reserva Biológica de Poço das Antas, Silva Jardim, Rio de Janeiro, Brasil. Rodriguésia 57, 429–442. doi: 10.1590/2175-7860200657304
Honório N. A., Codeço C. T., Alves F. C., Magalhães M. A., and Lourenço-De-Oliveira R. (2009). Temporal distribution of Aedes aEgypti in different districts of Rio de Janeiro, Brazil, measured by two types of traps. J. Med. Entomol 46, 1001–1014. doi: 10.1603/033.046.0505
INMET (2025). Banco de dados meteorológicos para ensino e pesquisa (Pecuária e Abastecimento: Instituto Nacional de Meteorologia - Ministério da Agricultura). Available online at: https://portal.inmet.gov.br/ (Accessed Jun 13, 2025).
ITTO (2025). Lecythis pisonis. Timber Species Database. Available online at: http://www.tropicaltimber.info/pt-br/specie/sapucaia-lecythis-pisonis/ (Accessed Jun 13, 2025).
Juliano S. A. and Philip Lounibos L. (2005). Ecology of invasive mosquitoes: effects on resident species and on human health. Ecol. Lett. 8, 558–574. doi: 10.1111/j.1461-0248.2005.00755
Kitching R. L. (2000). Food webs and container habitats: the natural history and ecology of phytotelmata (Cambridge University Press). doi: 10.1017/CBO9780511542107
Laurance W. F., Nascimento H. E. M., Laurance S. G., Andrade A., Ewers R. M., Harms K. E., et al. (2007). Habitat fragmentation, variable edge effects, and the landscape-divergence hypothesis. PloS One 2, e1017. doi: 10.1371/journal.pone.0001017
Lima Bersot M. I., Vieira G., De Moraes J. R., Rocha Pereira G., Albuquerque Motta M., and Lourenço-De-Oliveira R. (2023). Biological and behavioral features and colonization of the sylvatic mosquito Sabethes identicus (Diptera: Culicidae). PloS One 18, e0296289. doi: 10.1371/journal.pone.0296289
Lozovei A. L. (1998). Mosquitos dendrícolas (Diptera, Culicidae) em internódios de taquara da floresta atlântica, serra do mar e do primeiro planalto, Paraná, Brasil. Braz. Arch. Biol. Technol. 41, 501–510. doi: 10.1590/S1516-89131998000400016
Magurran A. E. (2021). Measuring biological diversity. Curr. Biol. 31, R1174–r1177. doi: 10.1016/j.cub.2021.07.049
Marcondes C. B. and Alencar J. (2010). Revisión de los mosquitos del género Haemagogus Williston (Diptera: Culicidae) de Brasil. Revista Biomédica. 21, 221–238.
Mascheretti M., Tengan C. H., Sato H. K., Suzuki A., Souza R. P. D., Maeda M., et al. (2013). Yellow fever: reemerging in the state of Sao Paulo, Brazi. Rev. Saude Publica 47, 881–889. doi: 10.1590/s0034-8910.2013047004341
Mckinney M. L. (2006). Urbanization as a major cause of biotic homogenization. Biol. Conserv. 127, 247–260. doi: 10.1016/j.biocon.2005.09.005
Medeiros-Sousa A. R., Ceretti-Junior W., Urbinatti P. R., Natal D., Carvalho G. C. D., Paula M. B. D., et al. (2013). Biodiversidade de mosquitos (Diptera: Culicidae) nos parques da cidade de São Paulo I. Biota Neotropica 13(1), 317–321. doi: 10.1590/S1676-06032013000100030
Mordecai E. A., Caldwell J. M., Grossman M. K., Lippi C. A., Johnson L. R., Neira M., et al. (2019). Thermal biology of mosquito-borne disease. Ecol. Lett. 22, 1690–1708. doi: 10.1111/ele.13335
Morin C. W. and Comrie A. C. (2013). Regional and seasonal response of a West Nile virus vector to climate change. Proc. Natl. Acad. Sci. 110, 15620–15625. doi: 10.1073/pnas.1307135110
Ngai J. T. and Srivastava D. S. (2006). Predators accelerate nutrient cycling in a bromeliad ecosystem. Science 314, 963–963. doi: 10.1126/science.1132598
NPARKS (2025). “Lecythis pisonis (sapucaia).,” in Flora Fauna web. Singapore: Flora Fauna web. Available online at: https://www.nparks.gov.sg/florafaunaweb/flora/7/5/7553.
Patz J. A., Graczyk T. K., Geller N., and Vittor A. Y. (2000). Effects of environmental change on emerging parasitic diseases. Int. J. Parasitol. 30, 1395–1405. doi: 10.1016/s0020-7519(00)00141-7
Possas C., Lourenço-De-Oliveira R., Tauil P. L., Pinheiro F. D. P., Pissinatti A., Cunha R. V. D., et al. (2018). Yellow fever outbreak in Brazil: the puzzle of rapid viral spread and challenges for immunisation. Memórias do Instituto Oswaldo Cruz 113, e180278. doi: 10.1590/0074-02760180278
Prance G. T. and Mori S. A. (1979). Lecythidaceae: part I: the actinomorphic-flowered new world lecythidaceae (Asteranthos, gustavia, grias, allantoma, & Cariniana). New York Botanical Garden Press 21, 1–270.
Reinert J. F. (2009). List of abbreviations for currently valid generic-level taxa in family Culicidae (Diptera). Eur. Mosq. Bull. 27, 68–76. Available online at: https://e-m-b.myspecies.info/sites/e-m-b.org/files/EMB%2827%2968-76.pdf.
Reinhold J. M., Lazzari C. R., and Lahondère C. (2018). Effects of the Environmental Temperature on Aedes aEgypti and Aedes albopictus Mosquitoes: A Review. Insects 9, 158. doi: 10.3390/insects9040158
Reisen W. K. (2010). Landscape epidemiology of vector-borne diseases. Annu. Rev. entomology 55, 461–483. doi: 10.1146/annurev-ento-112408-085419
Santos M., Mariscal L. C., Henríquez B., Garzón J., González P., Carrera J. P., et al. (2020). Implementação de armadilhas de bambu e macacos para amostragem de mosquitos reprodutores de cavidades em Darién, Panamá. Acta Trópica 205, 105352. doi: 10.1016/j.actatropica.2020.105352
Silva S. O. F., De Mello C. F., Maia D., and Alencar J. (2025). Vertical and temporal distribution of medically important mosquito species in an atlantic forest fragment of southeastern Brazil: ecological insights and public health implications. J. Am. Mosq. Control Assoc. 41, 61–68. doi: 10.2987/25-7217
Vasconcelos P. F. d. C. (2003). Febre amarela. Rev. da Sociedade Bras. Med. Trop. 36, 275–293. doi: 10.1590/S0037-86822003000200012
Vasconcelos P. F. C., Travassos Da Rosa A., Rodrigues S. G., Travassos Da Rosa E. S., Dégallier N., and Travassos Da Rosa J. F. S. (2001). Inadequate management of natural ecosystem in the Brazilian Amazon region results in the emergence and reemergence of arboviruses (Cadernos de Saúde Pública), 17, S155–S164. doi: 10.1590/S0102-311X2001000700025
Vieira G., Bersot M. I. L., Pereira G. R., de Abreu F. V. S., Nascimento-Pereira A. C., Neves M. S. A. S., et al. (2020). Documentação em vídeo de alta velocidade do comportamento de oviposição catapultadora de ovos do mosquito Sabethes albiprivus (Diptera: Culicidae). Entomologia Neotropical 49 , 662–667. doi: 10.1007/s13744-020-00782-x
WHO (2019). World health statistics 2019: monitoring health for the SDGs, sustainable development goals (World Health Organization).
Keywords: arboviruses, insect ecology, Culicidae, natural trap, Lecythis pisonis, vectors, entomo-logical surveillance
Citation: Mello CFd, Pavão LdM, Olsson Freitas Silva S, Queiroz AB and Alencar J (2025) Interaction between mosquito (Diptera: Culicidae) diversity and phytotelmata used as traps in an Atlantic forest ecosystem, state of Rio de Janeiro, Brazil. Front. Ecol. Evol. 13:1709756. doi: 10.3389/fevo.2025.1709756
Received: 20 September 2025; Accepted: 27 October 2025;
Published: 13 November 2025.
Edited by:
Paulino Siqueira Ribeiro, Harvard University, United StatesReviewed by:
Agostinho Nascimento Pereira, Federal University of Sergipe, BrazilAnthony Kiszewski, Bentley University, United States
Copyright © 2025 Mello, Pavão, Olsson Freitas Silva, Queiroz and Alencar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jeronimo Alencar, amFsZW5jYXJAaW9jLmZpb2NydXouYnI=