 Maria Höhn1*†
Maria Höhn1*† Endre György Tóth2†Enikő Ibolya Major1†János Sisa1†Zsófia Kovács3†
Endre György Tóth2†Enikő Ibolya Major1†János Sisa1†Zsófia Kovács3† Zoltán Attila Köbölkuti4†
Zoltán Attila Köbölkuti4†- 1Department of Botany, Institute of Agronomy, University of Agriculture and Life Sciences (MATE), Budapest, Hungary
- 2National Coalition of Independent Scholars (NCIS), Brattleboro, VT, United States
- 3Department of Soil Biology, Institute for Soil Sciences, Centre for Agricultural Research, HUN-REN, Budapest, Hungary
- 4Forest Research Institute (UoS-FRI), University of Sopron, Sárvár, Hungary
In ecosystems dominated by acidic soils, species are forced to adapt to unfavorable and stressful substrate conditions and have evolved various strategies. These include a range of physiological and chemical mechanisms and signaling pathways that trigger molecular responses, which has led, in turn, to structural and functional changes in responsive genes. As a result of these adaptations, molecular approaches have enabled considerable progress in explaining the mechanisms and detection of genes responsible for acidic pH tolerance. The development of gene-specific molecular markers offers more options for the marker-assisted detection of substrate-specific populations. Scots pine (Pinus sylvestris L.) is a major forest forming conifer species of Eurasia capable of growing on substrates with different pH values. In this study, based on the annotation of pH-responsive genes, novel SNP markers were identified and tested using Scots pine samples originating from habitats with substrates of varying pH. By testing the primers, 33 markers from 27 gene regions were successfully amplified using PCR. Following the sequencing of the PCR products, gene regions were analyzed, focusing specifically on polymorphic sites and synonymous/non-synonymous SNPs. From a total of 15426 sites, it was possible to identify 8 synonymous and 5 non-synonymous SNPs. The genetic variation revealed by the newly designed markers, affecting species’ soil pH tolerance help to identify individuals adapted to divergent soil types. In such cases, the markers may serve as valuable genetic tools for studies of adaptive genetic variation in Scots pine, a species used in both reforestation and forest plantations.
1 Introduction
A high degree of genetic diversity in populations is essential to ecosystem stability and resilience to environmental change. Within ecosystems the distribution of plant populations is determined by habitat characteristics and biotope quality. Adaptive behavior of a population provided on the basis of high genetic diversity as a response to habitat characteristics is an important prerequisite for species survival. The morphological and phenological trait responses of individuals are often influenced by extreme environments, and this may induce population-level adaptations (Gray and Brady, 2016; Dussarrat et al., 2021; Everingham et al., 2023; Nacakci and Gülcü, 2025). The threat posed to natural ecosystems by climate change is the focus of many studies aiming to uncover the genetic background of the stress-response traits of various populations inhabiting divergent habitats. Genetic information is, moreover, crucial to the development of conservation strategies, and the sustainable use of genetic resources is therefore becoming ever more important (Hoffmann and Sgrò, 2011).
Forest tree species are key components of the forest ecosystems and as they have long life spans, the shift in the genetic background responsible for adaptation will require a much longer period of time (Davis and Shaw, 2001; Hamrick, 2004; Savolainen et al., 2004; Jump and Peñuelas, 2005). The accumulation of a greater degree of genetic diversity and population variability provides a good basis for the promotion of the selection of genes responsible to mitigate the effects of rapid environmental changes. SNPs in these genes represent an inexhaustible source of polymorphic markers. Since its advent, development in the use of DNA molecular markers has already provided important resources for the study of forest tree species. The identification of microsatellites and SNPs through population genetic analyses has already been widely used to demonstrate the degree of genetic polymorphism in populations. Moreover, the detection of SNPs within genes involved in adaptation has rapidly increased due to improvements of high-throughput DNA sequencing methods (Kuhner et al., 2000; Garvin et al., 2010; Rellstab et al., 2017; Price et al., 2020).
A frequent corollary of climate hazards, long-lasting droughts and extreme environmental events is the changes in soil properties, including shifts in pH. The reaction of forest-forming tree species to changes in soil pH and the resulting effects on their physiological properties are important in the modulation of their adaptive potential. Soil pH can be highly variable and one factor in this is changes in soil water content, which may substantially modify the pH while plants are then forced to react and respond, making changes in their physiological parameters to ensure their continued ability to acquire nutrients and regulate their water acquisition and exchange (Jing et al., 2023). Changes in soil pH may therefore have a strong impact on the plant rhizosphere, affecting respiration, the solubility of nutrients or even modifying a series of biotic interactions (Felle, 2001; Koyama et al., 2001; Gao et al., 2004). The impact of external pH on gene expression modulation has been noted in several previous studies. Gene discovery associated with tolerance to soil carbonate was detected in some genotypes of Arabidopsis thaliana having contrasting behavior. A bulk-segregant-analysis sequencing (BSA-Seq) was performed, and transcriptomics revealed 208 root and 2503 leaf differentially expressed genes under bicarbonate stress (Busoms et al., 2023). In the study altogether 69 genes were associated with carbonate tolerance and final set of 18 genes involved in bicarbonate stress responses that had relevant roles in soil carbonate tolerance and the modifications had an impact to the sugar, lipid and protein metabolism of the tolerant genotype. Changes in external pH act as signals. Plants should modify responses auxin signaling, or make changes in cell wall plasticity and extensibility, as well as in the salicylic acid signaling involved in pathogen responses (Lager et al., 2010; Bailey et al., 2023).
As one of the most important forest-forming tree species and due to its capacity to persist in different, even extreme ecological conditions, Scots pine is considered one of the most promising conifer species in the time of climate change. Among the conifers, Scots pine has one of the widest distributions, forming taiga forests across boreal Eurasia. With its broad ecological tolerance, Scots pine can inhabit many types of habitats (Kremenetski et al., 2000; Mátyás et al., 2004; Hallingbäck et al., 2021). It has been documented in Europe in ecologically marginal environments from its northernmost occurrence in Scandinavia and Russia to the southernmost regions of the Mediterranean, including Turkey, Spain or Central Europe (Tóth et al., 2017a, 2021; Wachowiak et al., 2022). Naturally, given this geographic spread, populations can be found on different substrates with great variation in their pH values. Scots pine is even known to grow on rocky limestone cliffs and volcanic outcrops, forming stable populations on both alkaline and acidic sandy soils, and it is also typical of low pH boreal peat bogs. Historical populations of Scots pine are known to have survived the glacial cycles of the Pleistocene, in which they faced large fluctuations of the climate (Birks and Willis, 2008; Tóth et al., 2019).
Despite the ecological and economic importance of various conifer species, achieving complete genome assemblies remains a challenge due to their large genome size and the highly repetitive content of the genomes. Technological advances have, however, enabled the sequencing of several conifer species (De La Torre et al., 2014; Neale and Wheeler, 2019; Scott et al., 2020; Xiong et al., 2021; Neale et al., 2022; Lo et al., 2024; Chen et al., 2025), allowing the study of genes potentially involved in ecological adaptation and the development of specific markers targeting SNP variation (Prunier et al., 2015). Given that Scots pine is adapted to multiple habitat types and grows on substrates with varying pH values, it can be considered a model species for identifying pH-responsive gene candidates within the genome. This, in turn, makes it possible to develop markers aimed at detecting variation among populations growing on substrates with different pH levels. In population genetic studies, such markers would enable the identification of substrate-specific populations and allow for directed and controlled forest management.
2 Materials and methods
2.1 Plant material
The plant material used was obtained from four Central European Scots pine populations previously sampled for a study by Tóth et al. (2017b), while one population was collected later. These populations represent different habitats, each with substrates of significantly divergent pH values, namely peat bogs with low pH (RPO, RMH), Ca-rich rocky outcrops (SKV, SLO), and an alkaline sandy substrate (HFE) (see Supplementary Table S1). Five specimens were selected randomly from each population.
2.2 Bioinformatics data mining
To develop markers for tracking pH-responsive genes or their respective functional sites, it was first necessary to perform a library preparation and annotation of functional regions. In the interests of clarity, we focused on genes displaying significant dissimilarities in gene expression patterns in response to changes in external pH levels. On the basis of work by Lager et al. (2010), a total of 1036 genes were selected and were denoted as pH-responsive if a significant difference (adjusted P < 0.05) was observed, plus at least a twofold change at pH 4.5, with the control value being pH 6.0. The “rentrez” package (Winter, 2017) was used, implemented in R (R Core Team, 2013), to select and download EST and protein repository records accurately for each gene from the NCBI database (https://www.ncbi.nlm.nih.gov/). The Pinus sylvestris draft genome (NCBI Accession ID: PRJEB1898) was employed which has a sequencing coverage of 12.5×, an assembly size of 6.795 Gbp, approximately 16.1 million contigs, and an NG50 of 447 bp (Nystedt et al., 2013). To map the unannotated Scots pine genome, which consists of 881,136 contigs > 1Kb in length, a total of 2,569 EST and 16,769 protein sequences were downloaded and used as queries. We used the NCBI BLAST+ ‘makeblastdb’ tool to create a BLAST database from the genome and then conducted BLASTN and TBLASTN searches to identify each locus on the open-source web-based platform Galaxy (https://usegalaxy.org/). Sequences with the highest BLAST hit were selected for further processing. Only sequences with an e-value ≤0.001, sequence identity >98% and alignment length >100 bp were retained. For each locus, redundant sequences were separately aligned using the ClustalW (Thompson et al., 2003) multiple sequence alignment tool in BioEdit (Hall et al., 2011) to create a consensus sequence capable of characterizing the maximal length of each locus. A total of 48 different partial gene sequences could be identified. Following alignment of the sequences in the P. sylvestris genome, the corresponding contigs were extracted and used for marker development. The complete workflow is shown in a flowchart, Supplementary Figure S1.
2.3 Primer design
The online software Primer 3.0 (http://primer3.ut.ee/) was used to Design primer pairs. The parameters for this were: (1) primer length, between 18 bp and 23 bp, with 20 bp as the optimum; (2) PCR product size from 90 to 800 bp for each gene; (3) a melting temperature of 57°–62°C, with 59°C as the optimum; and (4) a GC content of 30–70%, with 50% as the optimum. The PCR product size for each gene was kept below 800 bp to ensure efficient Sanger sequencing.
2.4 DNA extraction and PCR amplification
Total DNA was isolated from silica-dried needles using the E.Z.N.A.® SP plant DNA kit. (Omega Bio-tek, Inc., Norcross, GA, USA) following the manufacturer’s protocol. In the assessment of the DNA concentration and quality a NanoDrop spectrophotometer (BioScience, Budapest, Hungary) was employed, and the result visually checked on a 1% agarose gel.
PCR amplification was conducted in a total volume of 24.2 µl, of which 1 µl was genomic DNA (about 20 ng), 0.18 µl DreamTaq DNA polymerase (Fermentas, Szeged, Hungary), with, 4.5 µl (10×) Dream Taq Green PCR buffer (ThermoFisher, Waltham, MA, USA), 3 mM MgCl2, 200 μM of dNTPs, 1% BSA, 0.2 mM of each primer and Milli-Q ultrapure water (Merckmillipore, Billerica, MA, USA). The PCR protocol was as follow: (1) pre-denaturation at 94°C for 3 min; (2) 32 cycles at 94°C for 30s; (3) annealing at 58 or 60°C for 45s; and (4) extension at 72°C for 1.2 min, and a final elongation at (5) 72°C for 7 min. For markers with multiple bands, further optimization was achieved via an increase up to 60°C in the annealing temperature. The PCR products were analyzed on a 1.5% (w/v) ethidium bromide-stained agarose gel in 1xTBE buffer to verify the amplification.
2.5 Genotyping by SNP marker candidates
Products appearing as a single band of the expected size (in P. sylvestris) were used to genotype samples of different origins using Sanger sequencing. For this, the purification of the products after PCR was achieved in one step via the hydrolysis of the excess primers and dephosphorylated unincorporated dNTPs using ExoSAP-IT™ Express PCR Product Clean-up Reagent, (ThermoFisher Scientific, Carlsbad, CA, USA), according to the manufacturer’s protocol. The products of the ExoSAP-IT process were then sequenced on the basis of the forward and reverse primers, in both directions, at Biomi Ltd., Hungary. The amplified sequences were then edited, visualized and aligned using BioEdit Sequence Alignment Editor, software, version 7.0.9.0 (Hall et al., 2011). Following sequence alignment, the genotyping and final validation of all potential SNPs was performed visually on chromatograms with the aid of a CodonCode Aligner 8.0.2 (Richterich, 2004). The number of polymorphic sites and of insertions/deletions, plus the character of SNPs (i.e. synonymous or nonsynonymous) was calculated using DNA Sequence Polymorphism 6.10.01 (DNASP) software (Rosas and Rosas 1995) and validated with a visual check in CodonCode Aligner 8.0.2.
3 Results
3.1 Primer testing
A total of 40 primers were designed and PCR amplified, and of these 33 yielded high-quality PCR products. Of the total, six primers were not successful, while one was excluded as it yielded only a low-quality sequence. The size range of the amplified sequences fell between 238 and 633bp. The newly designed and successfully tested primers, together with their marker codes and expected PCR products lengths are found in Table 1.
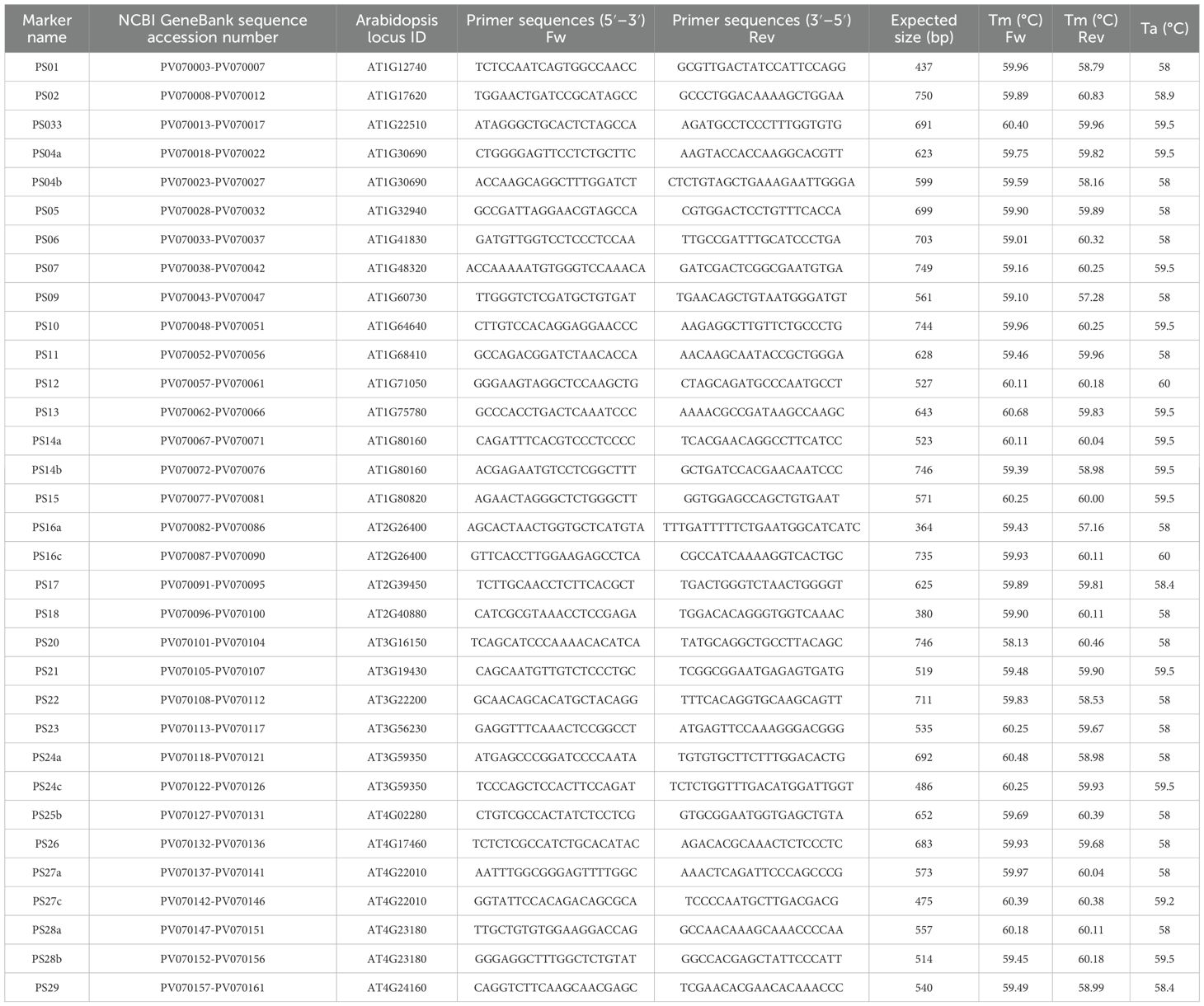
Table 1. List of selected genes sequences with NCBI GenBank sequence accession numbers, marker names, and forward and reverse primer sequences for pH responsive genes in P. sylvestris.
3.2 Evaluation of sequence variation
The newly designed molecular markers relate to 27 genes or partial gene regions that could be annotated as playing a role in trees’ adaptation to soil pH. The genes and their functions annotated in Arabidopsis are presented in Supplementary Table S2. Genetic characteristics, including the number of polymorphic sites (S), haplotypes (h), the degree of nucleotide diversity (Pi), and counts of synonymous and non-synonymous sites, were evaluated based on the sequence variation of 5 (or in some cases 4) Scots pine genotypes. Polymorphic site values ranged between 1 and 7, while the number of haplotypes of the newly developed markers ranged between 1 and 4. In nine of the cases, haplotype diversity was above zero. Further, eight synonymous and five non-synonymous sites were detected, and these are listed in Table 2, while the PCR products of the five non-synonymous sites are presented in Supplementary Figure S2.
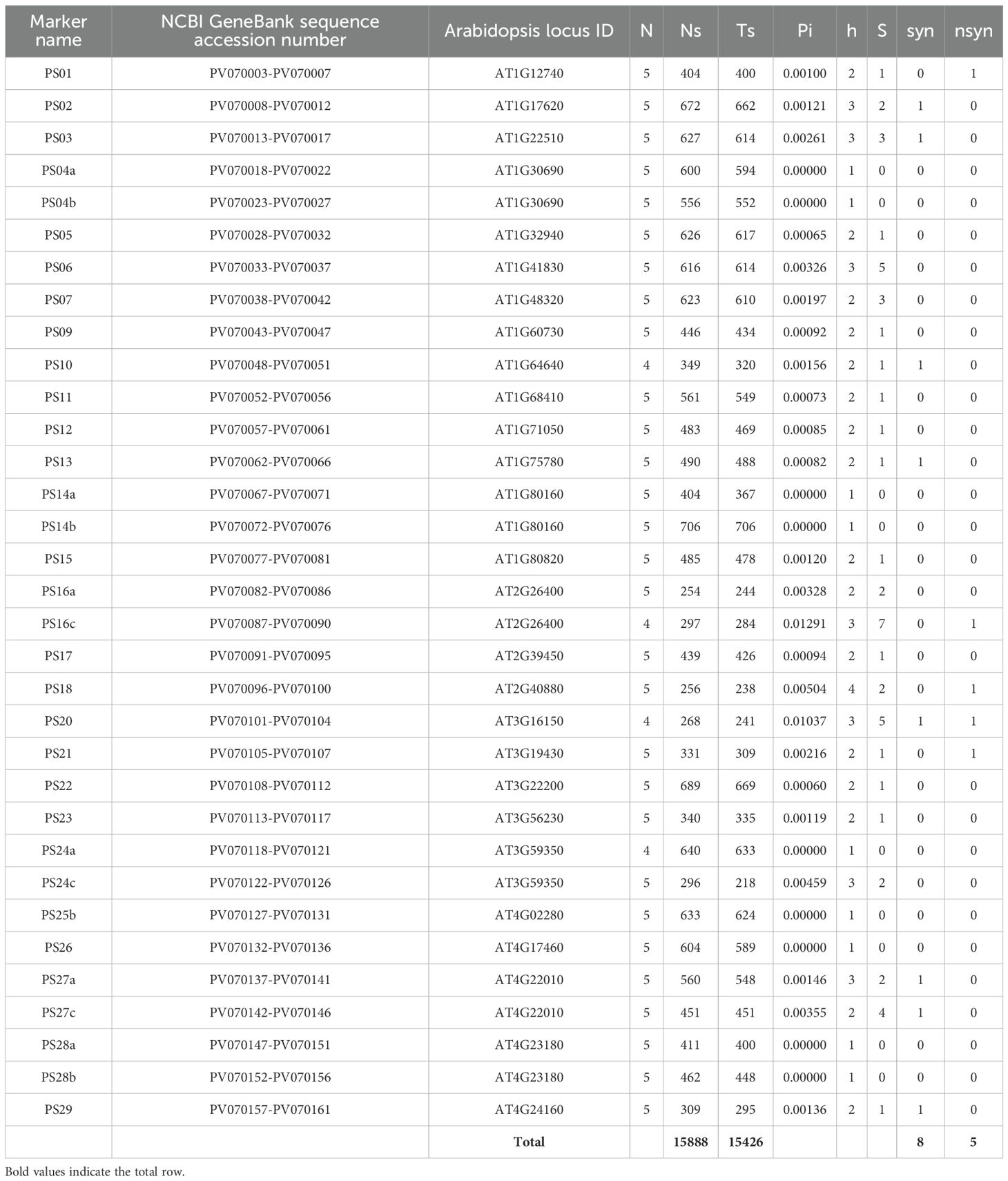
Table 2. Genetic characteristics of 33 newly developed markers of pH responsive genes in Scots pine genotypes, N: Number of sequences; Ns: Number of sites (alignment length); Ts: Total number of sites (excluding sites with gaps/missing data); Pi: Nucleotide diversity; h: Number of haplotypes; S: Number of polymorphic (segregating) sites; syn: synonymous mutations, nsyn: non-synonymous mutations.
4 Discussion
Soil pH is known to be a major environmental variable affecting plant life (Bloom et al., 2006; Neina, 2019). Soil pH plays a major role in environmental adaptation via its influence on a series of physiological-chemical reactions, acting as a signaling system that initiates responses at the molecular level, manifested firstly by the expression or alteration of responsive genes (Lager et al., 2010). Our genetic markers are based on DNA sequence variation of these genes. The newly designed primers yielded high-quality PCR products, and of the nine marker sites all turned out to be polymorphic. Our study also represents an SNP discovery test on genes presumed to be responsive to adaptation to soil pH, annotated by comparative genomics in the P. sylvestris genome. In the literature, cases are found in which SNP markers from sequences annotated by comparative genomics have been developed and successfully used on diverse plant species (Hall et al., 2002; Fulton et al., 2002; Köbölkuti et al., 2020; Vuruputoor et al., 2023). Translation of the annotated P. sylvestris genes followed by similarity searches in protein databases revealed that the corresponding DNA sequences or their encoded proteins have already been identified in other conifers or angiosperm tree species (Table), most likely because these genes occur in evolutionarily conserved genomic regions.
In the course of the sequence analysis, five non-synonymous SNPs were found in five different gene fragments. The biological functions of the proteins encoded by these genes are indicative: cytochrome P450 confers salt tolerance (Werck-Reichhart et al., 2002; Wang et al., 2016) acireductone dioxygenase has been demonstrated to have role in root growth behavior (Ramanathan et al., 2018) and tolerance to drought (Liang et al., 2019); and cystatins are universally present in plant taxa, suggesting a vital protease regulatory role for these proteins in plants in general (Benchabane et al., 2010) via the activation of protective metabolic cascades under abiotic stress conditions (Balbinott and Margis, 2022). Studies have reported an up-regulation of cystatin mRNA transcripts in leaves subjected to conditions adverse to growth, such as drought, salinity or low temperatures (Pernas et al., 1998); L-Asparaginase is a key enzyme for asparagine utilization by plants, and this has an important role in the nitrogen metabolism of developing plant tissues (Sieciechowicz et al., 1988); elevated levels of asparaginase activity are detected in developing tissues like leaves and roots (Michalska et al., 2006), and in late embryogenesis; abundant hydroxyproline-rich glycoprotein is involved in plant stress responses (Johnson et al., 2017).
The polymorphism observed in the sequences should be considered of particular importance, as the relevant genes seem to be essential to adaptation to substrate conditions. It may be presumed that the five non-synonymous SNPs detected influence the expression of some genes and modulate protein structures and function. The assignment of target sequences to genes and the coding sites responsible for the synthesis of the aforementioned proteins result in phenotypic changes of individuals in relation to adaptation.
The small number of samples used (4 and 5 per population) and the limited genotyped panels are methodological constraints that may have influenced our results, potentially leading to the omission of some relevant alleles. On the other hand, this allowed us to detect some putatively large effect loci. However, this sample size still affords the opportunity to further examine the connection between the selection and the soil/substrate properties of the species.
Having knowledge of the genetic background of the tree species in forest ecosystems is increasingly vital because climate change is expected to alter the species composition of forests globally. The decline of certain species opens up an ecological niche to others and leads to their expansion. Scots pine, with its tolerance of widely differing ecological conditions, and high adaptive capacity to extreme habitat sites, is a promising species in terms of its survival in forests and in parks and green spaces. Its presence on various, sometimes extreme substrates, and in conditions of widely varying soil pH suggests, however, that individuals have adapted differently over time. Knowledge of this adaptation is essential to the management and maintenance of future forests, landscapes and green spaces. Uncovering its genetic background, including the diversity of genes responsible for phenotypic variation, is an important prerequisite to our understanding of species’ ability to adapt to different or changing ecological conditions. The newly developed markers and the characterization of SNPs within the parts of the genome examined can contribute in an essential way to the investigation of the genetic basis of adaptive variation in Scots pine populations, making it possible to identify substrate-specific populations.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
MH: Conceptualization, Funding acquisition, Data curation, Investigation, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. EGT: Conceptualization, Data curation, Investigation, Methodology, Visualization, Validation, Writing – original draft, Writing – review & editing. EIM: Methodology, Investigation, Validation, Writing – review & editing. JS: Methodology, Validation, Writing – review & editing. ZK: Methodology, Investigation, Validation, Writing – review & editing. ZAK: Conceptualization, Data curation, Investigation, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The research was funded by a research excellence grant awarded to Maria Höhn from MATE, University of Agriculture and Life Sciences, Hungary.
Acknowledgments
The authors are grateful to MATE, the University of Agriculture and Life Sciences, for the Research Grant awarded to Mária Höhn.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1717368/full#supplementary-material
References
Bailey R. L., Zhu Y., and Shi M. (2023). Alkalinity modulates a unique suite of genes to recalibrate root responses to pH stress in barley. Plant Cell Environ. 46, 1812–1825. doi: 10.1111/pce.15567
Balbinott N. and Margis R. (2022). Unraveling the origin of the structural and functional diversity of plant cystatins. Plant Sci. 321, 111342. doi: 10.1016/j.plantsci.2022.111342
Benchabane M., Schlüter U., Vorster J., Goulet M. K., and Michaud D. (2010). Plant cystatins. Biochimie 92, 1657–1666. doi: 10.1016/j.biochi.2010.06.006
Birks H. J. B. and Willis K. J. (2008). Alpines, trees, and refugia in Europe. Plant Ecol. Divers. 1, 147–160. doi: 10.1080/17550870802349146
Bloom A., Frensch J., and Taylor A. R. (2006). Influence of inorganic nitrogen and pH on the elongation of maize seminal roots. Ann. Bot-London 97, 867–873. doi: 10.1093/aob/mcj605
Busoms S., Pérez-Martín L., Terés J., Huang X.-Y., Yant L., Tolrà R., et al. (2023). Combined genomics to discover genes associated with tolerance to soil carbonate. Plant Cell Environ. 46, 3986–3998. doi: 10.1111/pce.14691
Chen H., Qin X., Chen Y., Zhang H., Feng Y., Tan Y., et al. (2025). Chromosome-level genome assembly of Pinus massoniana provides insights into conifer adaptive evolution. GigaScience 14, giaf056. doi: 10.1093/gigascience/giaf056
Davis M. B. and Shaw R. G. (2001). Range shifts and adaptive responses to quaternary climate change. Science 292, 673–679. doi: 10.1126/science.292.5517.673
De La Torre A. R., Birol I., Bousquet J., Ingvarsson P. K., Jansson S., Jones S. J. M., et al. (2014). Insights into conifer giga-genomes. Plant Physiol. 166, 1724–1732. doi: 10.1104/pp.114.248708
Dussarrat T., Decros G., Díaz F. P., Gibon Y., Latorre C., Rolin D., et al. (2021). Another tale from the harsh world: how plants adapt to extreme environments. Annu. Plant Rev. 4(2), 551–603. doi: 10.1002/9781119312994.apr0758
Everingham S. E., Blick R. A. J., Sabot M. E. B., Slavich E., and Moles A. T. (2023). Southern hemisphere plants show more delays than advances in flowering phenology. J. Ecol. 111, 380–390. doi: 10.1111/1365-2745.13828
Felle H. H. (2001). pH: signal and messenger in plant cells. Plant Biol. (Stuttgart) 3, 577–591. doi: 10.1055/s-2001-19372
Fulton T. M., van der Hoeven R., Eannetta N. T., and Tanksley S. D. (2002). Identification, analysis, and utilization of conserved ortholog set markers for comparative genomics in higher plants. Plant Cell 14, 1457–1467. doi: 10.1105/tpc.010479
Gao D., Knight M. R., Trewavas A. J., Sattelmacher B., and Plieth C. (2004). Self-Reporting Arabidopsis Expressing pH and [Ca2]Indicators Unveil Ion Dynamics in the Cytoplasm and in the Apoplast under Abiotic Stress. Plant Physiol. 134, 898–908. doi: 10.1104/pp.103.032508
Garvin M. R., Saitoh K., and Gharrett A. J. (2010). Application of single nucleotide polymorphisms to non-model species: a technical review. Molec Ecol. Res. 10, 915–1108. doi: 10.1111/j.1755-0998.2010.02891.x
Gray S. B. and Brady S. M. (2016). Plant developmental responses to climate change. Dev. Biol. 419, 64–77. doi: 10.1016/j.ydbio.2016.07.023
Hall T., Biosciences I., and Carlsbad C. A. (2011). BioEdit: an important software for molecular biology. GERF Bull. Biosci. 2, 60–61.
Hall A. E., Fiebig A., and Preuss D. (2002). Beyond the Arabidopsis genome: opportunities for comparative genomics. Plant Physiol. 129, 1439–1447. doi: 10.1104/pp.004051
Hallingbäck H. R., Burton V., Vizcaíno-Palomar N., Trotter F., Liziniewicz M., Marchi M., et al. (2021). Managing uncertainty in scots pine range-wide adaptation under climate change. Front. Ecol. Evol. 9. doi: 10.3389/fevo.2021.724051
Hamrick J. L. (2004). Response of forest trees to global environmental changes. For. Ecol. Manage. 197, 323–335. doi: 10.1016/j.foreco.2004.05.023
Hoffmann A. and Sgrò C. (2011). Climate change and evolutionary adaptation. Nature 470, 479–485. doi: 10.1038/nature09670
Jing C. H. E., Zhao X. Q., and Shen R. F. (2023). Molecular mechanisms of plant adaptation to acid soils: A review. Pedosphere 33, 14–22. doi: 10.1016/j.pedsph.2022.10.001
Johnson K. L., Cassin A. M., Lonsdale A., Bacic A., Doblin M. S., and Schultz C. J. (2017). Pipeline to identify hydroxyproline-rich glycoproteins. Plant Physiol. 174, 886–903. doi: 10.1104/pp.17.00294
Jump A. S. and Peñuelas J. (2005). Running to stand still: adaptation and the response of plants to rapid climate change. Ecol. Lett. 8, 1010–1020. doi: 10.1111/j.1461-0248.2005.00796.x
Köbölkuti Z. A., Tóth E. G., Jahn D., Heinze B., and Höhn M. (2020). SNP marker development in Pinus sylvestris L. @ in stress-responsive genes characterized from Pinus cembra L. transcriptomes. Mol. Biol. Rep. 47, 4841–4847. doi: 10.1007/s11033-020-05527-y
Koyama H., Toda T., and Hara T. (2001). Brief exposure to low pH stress causes irreversible damage to the growing root in Arabodopsisthaliana. Pecti-Ca interactions may play an important role in proton rhizotoxicity. J. Exp. Bot. 52, 361–368. doi: 10.1093/jexbot/52.355.361
Kremenetski C. V., Liu K., and Macdonald M. (2000). “The late Quaternary dynamics of pines in northern Asia,” in Ecology and biogeography of pinus. Ed. Richardson D. M. (Cambridge UK: Cambridge University Press), p. 96–97.
Kuhner K. M., Beerli P., Yamato J., and Felsenstein J. (2000). Usefulness of single nucleotide polymorphism data for estimating population parameters. Genetics 156, 439–447. doi: 10.1093/genetics/156.1.439
Lager I. D. A., Andréasson O., Dunbar T. L., Andreasson E., Escobar M. A., and Rasmusson A. G. (2010). Changes in external pH rapidly alter plant gene expression and modulate auxin and elicitor responses. Plant Cell Environ. 33, 1513–1528. doi: 10.1111/j.1365-3040.2010.02161.x
Liang S., Xiong W., Yin C., Xie X., Jin Y. J., Zhang S., et al. (2019). Overexpression of OsARD1 improves submergence, drought, and salt tolerances of seedling through the enhancement of ethylene synthesis in rice. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01088
Lo T., Coombe L., Gagalova K. K., Marr A., Warren R. L., Kirk H., et al. (2024). Assembly and annotation of the black spruce genome provide insights on spruce phylogeny and evolution of stress response. G3: Genes Genomes Genet. 14, jkad247. doi: 10.1093/g3journal/jkad247
Mátyás Cs., Ackzell L., and Samuel C. J. A. (2004). EUFORGEN technical guidelines for genetic conservation and use for Scots pine (Pinus sylvestris) (Rome, Italy: International Plant Genetic Resources), ISBN: ISBN: 929043661-1.
Michalska K., Bujacz G., and Jaskolski M. (2006). Crystal structure of plant asparaginase. J. Mol. Biol. 360, 105–116. doi: 10.1016/j.jmb.2006.04.066
Nacakci F. M. and Gülcü S. (2025). Comparison of Scotch Pine (Pinus sylvestris L.) Origins in terms of Photosynthetic Gas Exchange and Chemical Properties. BioResources 20, 4681–4700. doi: 10.15376/biores.20.2.4681-4700
Neale D. B. and Wheeler N. C. (2019). The conifers: genomes, variation and evolution (Cham, Switzerland: Springer), 49–56. doi: 10.1007/978-3-319-46807-5
Neale D. B., Zimin A. V., Zaman S., Scott A. D., Shrestha B., Workman R. E., et al. (2022). Assembled and annotated 26.5 Gbp coast redwood genome. G3: Genes Genomes Genet. 12, jkab380. doi: 10.1093/g3journal/jkab380
Neina D. (2019). The Role of Soil pH in Plant Nutrition and Soil Remediation. Appl. Environ. Soil Sci. 5794869, 9. doi: 10.1155/2019/5794869
Nystedt B., Street N. R., Wetterbom A., Zuccolo A., Lin Y. C., Scofield D. G., et al. (2013). The Norway spruce genome sequence and conifer genome evolution. Nature 497, 579–584. doi: 10.1038/nature12211
Pernas M., Sánchez-Monge R., Gómez L., and Salcedo G. (1998). A chestnut seed cystatin differentially effective against cysteine proteinases from closely related pests. Plant Mol. Biol. 38, 1235–1242. doi: 10.1023/A:1006154829118
Price N., Lopez L., Platts A. E., and Lasky J. R. (2020). In the presence of population structure: From genomics to candidate genes underlying local adaptation. Ecol. Evol. 10, 1889–1904. doi: 10.1002/ece3.6002
Prunier J., Verta J.-P., and MacKay J. J. (2015). Conifer genomics and adaptation: at the crossroads of genetic diversity and genome function. New Phytol. 209, 44–62. doi: 10.1111/nph.13565
Ramanathan V., Rahman H., Subramanian S., Nallathambi J., Kaliyaperumal A., Manickam S., et al. (2018). OsARD4 encoding an acireductone dioxygenase improves root architecture in rice by promoting development of secondary roots. Sci. Rep. 8, 15713. doi: 10.1038/s41598-018-34053-y
R Core Team (2013). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing).
Rellstab C., Fischer M., Zolle R. S., Graf R., Tedder A., Shimizu K. K., et al. (2017). Local adaptation (mostly) remains local: reassessing environmental associations of climate-related candidate SNPs in Arabidopsis halleri. Heredity 118, 193–201. doi: 10.1038/hdy.2016.82
Richterich P. (2004). CodonCode aligner version 1.2 released. Genet. Med. 6, 162–163. doi: 10.1097/00125817-200405000-00012
Savolainen O., Bokma F., Garcıa-Gil R., Komulainen P., and Repo T. (2004). Genetic variation in cessation of growth and frost hardiness and consequences for adaptation of Pinus sylvestris to climatic changes. For. Ecol. Manag 197, 79–89. doi: 10.1016/j.foreco.2004.05.006
Scott A. D., Zimin A. V., Puiu D., Workman R., Britton M., Zaman S., et al. (2020). A reference genome sequence for giant sequoia. G3: Genes Genomes Genet. 10, 3907–3919. doi: 10.1534/g3.120.401612
Sieciechowicz K. A., Joy K. W., and Ireland R. J. (1988). The metabolism of asparagine in plants. Phytochemistry 27, 663–671. doi: 10.1016/0031-9422(88)84071-8
Thompson J. D., Gibson T. J., and Higgins D. G. (2003). Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinf. 1), 2–3. doi: 10.1002/0471250953.bi0203s00
Tóth E. G., Bagnoli F., Vendramin G. G., György Z., Spanu I., and Höhn M. (2021). Imprints of selection in peripheral and ecologically marginal central-eastern European Scots pine populations. Gene 779, 145509. doi: 10.1016/j.gene.2021.145509
Tóth E. G., Bede-Fazekas Á., Vendramin G. G., Bagnoli F., and Höhn M. (2019). Mid-Pleistocene and Holocene demographic fluctuation of Scots pine (Pinus sylvestris L.) in the Carpathian Mountains and the Pannonian Basin: Signs of historical expansions and contractions. Quatern Int. 504, 202–213. doi: 10.1016/j.quaint.2017.11.024
Tóth E. G., Köbölkuti Z. A., Pedryc A., and Höhn M. (2017a). Evolutionary history and phylogeography of Scots pine (Pinus sylvestris L.) in Europe based on molecular markers. J. Forestry Res. 28, 637–651. doi: 10.1007/s11676-017-0393-8
Tóth E. G., Vendramin G. G., Bagnoli F., Cseke K., and Höhn M. (2017b). High genetic diversity and distinct origin of recently fragmented Scots pine (Pinus sylvestris L.) populations along the Carpathians and the Pannonian Basin. Tree Genet. Genomes 13, 47. doi: 10.1007/s11295-017-1137-9
Vuruputoor V. S., Monyak D., Fetter K. C., Webster C., Bhattarai A., Shrestha B., et al. (2023). Welcome to the big leaves: Best practices for improving genome annotation in non-model plant genomes. Appl. Plant Sci. 11, e11533. doi: 10.1002/aps3.11533
Wachowiak W., Zukowska W. B., Perry A., Lewandowski A., Cavers S., and Łabiszak B. (2022). Phylogeography of Scots pine in Europe and Asia based on mtDNA polymorphisms. J. Syst. Evol. 61, 315–327. doi: 10.1111/jse.12907
Wang C., Yang Y., Wang H., Ran X., Li B., Zhang J., et al. (2016). Ectopic expression of a cytochrome P450 monooxygenase gene PtCYP714A3 from Populus trichocarpa reduces shoot growth and improves tolerance to salt stress in transgenic rice. Plant Biotechnol. J. 14, 1838–1851. doi: 10.1038/s41419-018-0948-4
Werck-Reichhart D., Bak S., and Paquette S. (2002). Cytochromes P450. Arabidopsis book Am. Soc. Plant Biologists U.S.A. 1, 28. doi: 10.1199/tab.0028
Winter D. J. (2017). rentrez: An R package for the NCBI eUtils API. PeerJ Preprints 5, e3179v2. doi: 10.7287/peerj.preprints.3179v2
Keywords: Scots pine, soil pH tolerance, primer design, SNP, adaptation
Citation: Höhn M, Tóth EG, Major EI, Sisa J, Kovács Z and Köbölkuti ZA (2025) Novel genetic markers for tracking pH-responsive genes in Pinus sylvestris (L.). Front. Ecol. Evol. 13:1717368. doi: 10.3389/fevo.2025.1717368
Received: 01 October 2025; Accepted: 13 November 2025; Revised: 05 November 2025;
Published: 05 December 2025.
Edited by:
Xiaoxue Ye, Chinese Academy of Tropical Agricultural Sciences, ChinaReviewed by:
Maximiliano Estravis-Barcala, Umeå University, SwedenFatma Merve Nacakci, Isparta University of Applied Sciences, Türkiye
Copyright © 2025 Höhn, Tóth, Major, Sisa, Kovács and Köbölkuti. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Höhn, aG9obi5tYXJpYUB1bmktbWF0ZS5odQ==
†ORCID: Maria Höhn, orcid.org/0000-0002-8587-8271
Endre György Tóth, orcid.org/0000-0001-7060-924X
Enikő Ibolya Major, orcid.org/0000-0003-4012-9856
János Sisa, orcid.org/0009-0009-1716-6801
Zsófia Kovács, orcid.org/0000-0001-5066-4911
Zoltán Attila Köbölkuti, orcid.org/0000-0002-6933-5648