 Pilar Truchado1
Pilar Truchado1 Márcia Oliveira2
Márcia Oliveira2 Rebeca Cordero-García2Manuel Abellán Soler3Amador Rancaño4Francisca García4
Rebeca Cordero-García2Manuel Abellán Soler3Amador Rancaño4Francisca García4 Avelino Álvarez-Ordóñez2
Avelino Álvarez-Ordóñez2 Ana Allende1*
Ana Allende1*- 1Research Group on Microbiology and Quality of Fruit and Vegetables, Food Science and Technology Department, CEBAS-CSIC, Murcia, Spain
- 2Department of Food Hygiene and Technology and Institute of Food Science and Technology, Universidad de León, León, Spain
- 3Entidad Regional de Saneamiento y Depuración de Murcia (ESAMUR), Murcia, Spain
- 4Acciona Agua, S.A.U., Madrid, Spain
Wastewater reuse for agricultural irrigation is increasingly essential, but it carries potential public health risks due to the dissemination of antimicrobial resistance (AMR). This study evaluates the effectiveness of four tertiary wastewater treatment technologies—peracetic acid (PAA), PAA combined with low-intensity ultraviolet-C (PAA/UV Low), high-intensity UV-C (UV High), and ultrafiltration (UF)—in reducing extended-spectrum β-lactamase-producing Escherichia coli (ESBL-E. coli) and antimicrobial resistance genes (ARGs) in reclaimed water used for irrigation. The relative abundance of the genes, normalized to the 16S RNA gene present in the water samples, was then estimated to assess whether there is an amplification of these genes during the reuse process in the wastewater treatment plant (WWTP). The results indicate that while all treatments significantly reduced ESBL-E. coli (≥3 logs cfu/100 mL) and ARGs (≥ 1.5 logs gc/100 mL), complete elimination was not achieved in any WWTP. Among the treatments, UF demonstrated the highest removal efficiency (≈4 log gc ARG/100 mL), against ARGs, followed by UV High (≈3 log gc ARG/100 mL), whereas PAA and PAA/UV Low were less effective (≈2 log gc ARG/100 mL). The study also found that while absolute ARG levels were reduced, their relative abundance remained stable or showed minimal decline, suggesting a persistent environmental reservoir of resistance genes. Among the ARGs analyzed, the most frequently detected were associated with tetracyclines (tetW, tetA), quinolones (qnrB, qnrS), and sulfonamides (sul1, sul2), highlighting potential public health concerns. Moreover, multidrug-resistant (MDR) ESBL-E. coli isolates were present across all WWTPs, exhibiting resistance to β-lactams, quinolones, tetracyclines, and sulfonamides. Nevertheless, notably low levels of resistance to last-resort antibiotics (tigecycline, colistin, and meropenem) were observed. These findings underscore the critical role of tertiary treatments in mitigating antimicrobial resistance (AMR) risks in water reuse systems. However, the persistence of ARGs in effluents suggests that current WWTP processes require further optimization.
Highlights
• Multidrug-resistant ESBL-E. coli persisted in WWTP effluents.
• ARGs were reduced but not completely eliminated in treated wastewater.
• Ultrafiltration (UF) was the most effective in reducing ESBL-E. coli and ARGs.
• High-dose UV was more effective than PAA and PAA/UV Low for ARG removal.
• Detection of ARBs and ARGs emphasizes the need for improved water reuse strategies.
1 Introduction
Antimicrobial resistance (AMR) is a significant global public health concern, as bacteria carrying antimicrobial resistance genes (ARGs) and deployment of antibiotic residues are the main contributing agent causing thousands of deaths each month (GBD 2021 Antimicrobial Resistance Collaborators, 2024). The WHO considers this threat a priority and has endorsed a global action plan on AMR, which includes the development of a global public health research agenda to address major knowledge gaps [(World Health Organization (WHO), 2018)]. One contributing factor to this crisis is the excessive and inappropriate use of antibiotics in medical, livestock, and agricultural contexts (McConnell et al., 2018). This has led to the continuous deployment of antibiotic residues into the environment, facilitating the emergence of antimicrobial resistant bacteria (ARB) and ARGs. Europe, and particularly, the European Commission places significant importance on addressing AMR due to its critical impact on public health, healthcare systems, and economies.
The support that the European Commission gives to the One Health approach recognizes that human health, animal health, and the environment are all interconnected. AMR is a prime example of this interdependence, as resistant microbes and resistance genes can be transmitted between humans, animals, crops, and the environment. This recognition is embedded in several key policy and legislative instruments, including Regulation (EU) 2019/6 on veterinary medicinal products, which introduces stricter measures to reduce the use of antimicrobials in animals, and Regulation (EU) 2019/4 on medicated feed. Moreover, the European Commission's 2019 Communication on the Strategic Approach to Pharmaceuticals in the Environment highlights the environmental dimension of AMR, promoting actions to address the release of pharmaceuticals, including antibiotics, throughout their life cycle. These efforts are further supported by the European One Health Action Plan against AMR (2017) and integrated into the broader Farm to Fork Strategy and the Zero Pollution Action Plan, which aim to reduce pollution from antimicrobial substances in water, soil, and food systems (European Commission, 2017). Wastewater treatment plants (WWTPs) receive wastewater from various sources, including municipalities, hospitals, and industries, and have been underscored as potential hotspots for the transmission of ARGs and, consequently, ARB (Hazra et al., 2024; Li et al., 2022). The main hypothesis is that wastewater contains antibiotics, high bacterial loads, and nutrient-rich substances, creating ideal conditions for the selection and spread of ARGs and ARBs, which confer resistance and survival advantages to microbial communities (Buriánková et al., 2021; Haberecht et al., 2019; Oliveira et al., 2018). While WWTPs design allows a successful removal of dissolved nutrients and solids, including bacteria, there is currently no specific technology in place for the targeted reduction of ARGs (Gao et al., 2022; Rizzo et al., 2013). Several studies have focused on evaluating the presence of ARB and ARGs in the influent and effluent of various WWTPs (Macrı̀ et al., 2024; Sanz et al., 2024). Overall, most of these studies have reported a decrease in absolute concentrations after treatment. Generally, wastewater treatment reduces the overall abundance of ARBs and ARGs by ~2–3 logs, thereby lowering the risk of ARG dissemination into the environment (Ben et al., 2017; Hultman et al., 2018; Rodriguez-Mozaz et al., 2014). However, some studies have suggested that WWTPs may contribute to the proliferation of ARGs in the environment (Kumar et al., 2020; Stachurova et al., 2021; Wang et al., 2020).
The current European regulatory framework includes regulations on the use of antimicrobials in human and veterinary medicine, as well as measures to prevent the spread of resistant bacteria in healthcare settings and the environment, including water reuse systems [European Commission, 2020a; World Health Organization (WHO), 2018]. However, there is growing awareness of its connection to climate change and how climate change can influence AMR in agriculture (van Bavel et al., 2024). The EFSA Scientific Opinion [(EFSA (European Food Safety Authority, Panel on Biological Hazards), 2021)] further underscores the major role of the environment in the emergence and spread of AMR through food chains, identifying key transmission routes such as fecal-origin fertilizers, irrigation water, and animal feed. The Opinion also highlights the presence of critical ARB and ARGs [e.g., blaCTX − M, mcr, van(A), qnr] across plant-based and animal production systems, emphasizing the importance of mitigation strategies across sectors.
In Mediterranean countries, which are severely affected by water scarcity, Europe is promoting the use of reclaimed water for irrigation as part of a sustainable water management strategy (Echaide et al., 2021; European Commission, 2020b). Nonetheless, ensuring the microbiological quality of reclaimed water is crucial by monitoring not only pathogenic bacteria but also ARB and ARGs, as they could potentially be transferred to crops through irrigation (Macrı̀ et al., 2024; Rodriguez-Mozaz et al., 2014; Sanz et al., 2024). However, the effectiveness of different water treatment technologies in reducing or eliminating ARBs and ARGs in WWTPs remains uncertain due to insufficient data (Ben et al., 2017; Osińska et al., 2020; Wang et al., 2020).
This study aims to investigate and compare various water treatments implemented in four distinct WWTPs for the reclamation of urban wastewater used in agriculture. The methods include peracetic acid (PAA), PAA combined with low-intensity ultraviolet-C (UV-C; PAA/UV Low), high-intensity UV-C (UV High), and ultrafiltration (UF). The research will evaluate the efficacy of these water treatments in reducing ARB carrying ARGs, with a particular focus on ESBL-producing Escherichia coli and 10 different ARGs. The assessment will be conducted using quantitative real-time PCR (qPCR).
2 Material and methods
2.1 Urban wastewater treatment plants
Four WWTPs located in the Region of Murcia, Spain, were selected for this study, each employing different tertiary treatment technologies. The primary and secondary treatment processes were previously described by Oliveira et al. (2023). Briefly, primary treatment typically includes aeration, solids and suspended solids separation, grit removal/degreasing, and a primary clarifier of varying size. Secondary treatment involves an aerobic or anaerobic biological process in a secondary clarifier, with coagulation/flocculation and supplementary lamella clarification. For tertiary treatment, the four WWTPs utilize different water treatment technologies: peracetic acid (PAA), PAA combined with low-intensity ultraviolet-C (UV-C; PAA/UV Low), high-intensity UV-C (UV High), and ultrafiltration (UF). The doses of PAA and UV are summarized in Table 1. Each WWTP receives both domestic and agricultural wastewater. Additionally, the WWTP using PAA also treats a significant amount of pre-treated hospital wastewater.
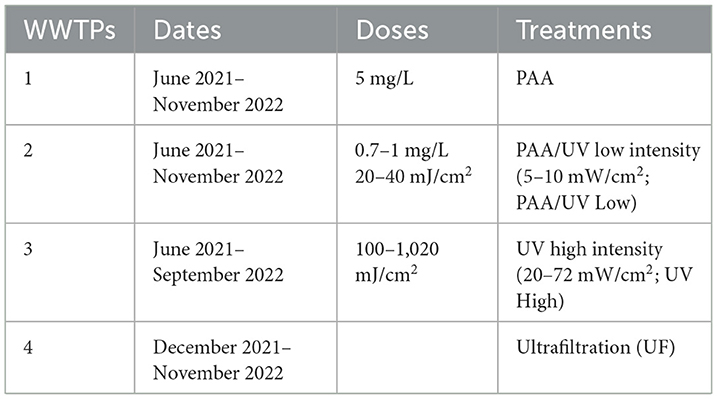
Table 1. Water treatments used in the WWTPs included in this study.
2.2 Sampling
From June 2021 to December 2022, monthly influent and effluent samples were collected from each WWTP. However, UF samples were only collected from December 2021 to December 2022. A total of 132 samples were included in this study: 18 influent and 18 effluent samples were collected from each WWTPs, except for the UF system, from which 12 influent and 12 effluent samples were taken. One liter of each sample was collected in sterile polypropylene plastic bottles (Labbox Labware S.L., Barcelona, Spain), stored under refrigerated conditions, transported to the laboratory within 2 h, and kept at 4°C until analysis.
2.3 Presence of ESBL-producing E. coli in wastewater samples
The levels of extended-spectrum β-lactamase (ESBL)-producing E. coli (cfu/100 mL) in each influent water sample were determined by plating. Serial 10-fold dilutions were prepared in buffered peptone water (BPW, 2 g/L; Oxoid) and then spread-plated onto CHROMagar ESBL (CHROMagar, Paris, France). For effluent water samples, aliquots of 1, 10, and 100 mL were filtered through sterile cellulose nitrate filters (0.45 μm, Sartorius, Madrid, Spain) using a vacuum filtration system (Sartorius). Dark pink to reddish colonies were counted after 24 h of incubation at 37°C. All analyses were performed in duplicate, and results were expressed as log cfu/100 mL.
2.4 Antibiotic susceptibility testing
A total of 366 E. coli isolates producing ESBL were selected for antibiotic susceptibility testing (AST). The isolates were obtained from both influent and effluent samples across the four wastewater treatment processes (PAA, PAA/UV Low, UV High, and UF). The number of isolates tested per sample was proportionally selected based on the initial recovery rates of presumptive ESBL-producing E. coli, ensuring representative coverage of all water matrices. Susceptibility to a panel of antibiotics was assessed using Sensititre EUVSEC3 plates (Thermo Scientific, TREK Diagnostic Systems Ltd., East Grinstead, UK) and the broth microdilution method, following the manufacturer's instructions, as previously described by Oliveira et al. (2023). Briefly, isolates were cultured in BHI broth at 37°C for 24 h. The resulting bacterial suspension was adjusted to a 0.5 McFarland turbidity standard and transferred to Mueller-Hinton broth. Subsequently, 50 μL of the suspension were dispensed into each well of the AST plate using the Sensititre AIM Automated Inoculation Delivery System (Thermo Scientific, TREK Diagnostic Systems Ltd., East Grinstead, UK). After 24 h of incubation at 37°C, growth was visually assessed to determine the minimum inhibitory concentration (MIC) for each antibiotic. Interpretation of susceptibility or resistance was based on the epidemiological cut-off values (ECOFF) provided by the European Committee on Antimicrobial Susceptibility Testing (EUCAST). The antibiotics tested included sulfamethoxazole, trimethoprim, ciprofloxacin, tetracycline, meropenem, azithromycin, nalidixic acid, cefotaxime, chloramphenicol, tigecycline, ceftazidime, colistin, ampicillin, and gentamicin.
2.5 Wastewater DNA extraction
Ten milliliters of influent samples were concentrated by centrifugation at 3,000 × g for 10 min. The supernatant was removed, and the pellet was resuspended in 1 mL of phosphate-buffered saline (PBS, Sigma-Aldrich, LS, USA). The resuspended pellet was then centrifuged at 9,000 × g for 10 min at 4°C, and the supernatant was discarded. For effluent samples, 100 mL of water was vacuum-filtered through sterile cellulose nitrate filters (0.45 μm). The filters were placed in 50 mL Falcon tubes containing 20 mL of PBS supplemented with Tween 80 (1 mL/L; Sigma-Aldrich). The tubes were vortexed for 7 min, after which the filters were removed. The sample was then centrifuged at 3,000 × g for 10 min, the supernatant was discarded, and the pellet was resuspended in 1 mL of PBS. The resuspended pellet was further concentrated by centrifugation at 9,000 × g for 10 min at 4°C. Both influent and effluent pellets were stored at −20°C until genomic DNA extraction. Genomic DNA extraction from wastewater concentrates was performed using the Maxwell® RSC Instrument (Promega) and the Maxwell RSC Pure Food GMO and Authentication Kit (Promega), following the Maxwell RSC Viral Total Nucleic Acid running program. DNA concentration and purity were determined using an Implen NanoPhotometer N60/50 (Implen, Munich, Germany). The DNA samples were then stored at −20°C.
2.6 Antimicrobial resistance genes qPCR assays
Quantitative PCR (qPCR) was used to quantify ARGs encoding resistance to beta-lactams (blaCTX − M and blaTEM), chloramphenicol (cmlA and catI), quinolones (qnrB and qnrS), sulfonamides (sul1 and sul2), and tetracyclines (tetA and tetW). Additionally, the 16S rRNA gene was analyzed to quantify total bacterial populations and normalize ARG abundance in the collected samples. All qPCR assays were performed using a QuantStudio 5 system (Applied Biosystems, USA) in 96-well plates with KAPA SYBR FAST and KAPA PROBE FAST Universal qPCR Master Mix kits (KapaBiosystems, Massachusetts, USA). The selection of primers and probes used to quantify ARGs as well as cycling parameters, are detailed in Table 2. Each qPCR analysis was conducted in triplicate wells containing both undiluted and diluted DNA samples. Wastewater samples were diluted 50-fold, while reclaimed water samples were diluted by a factor of 5. Each qPCR assay included three negative control wells (nuclease-free water). When SYBR Green was used for detection, a melting curve analysis was performed after each assay to confirm that the fluorescence signal originated from a specific PCR product. The standard curve for each ARG was established following the method described by Truchado et al. (2016). The limit of detection (LOD) and limit of quantification (LOQ) were determined for each target gene through a series of dilutions, with 4–10 replicates per dilution. ARG quantification results were expressed as absolute and relative abundance. Absolute abundance was reported as the number of copies per 100 mL of sample, while relative abundance was calculated by normalizing the absolute copy number of each gene to 16S rRNA gene copy numbers (i.e., ARG copies/16S rRNA copies). All values were log-transformed.
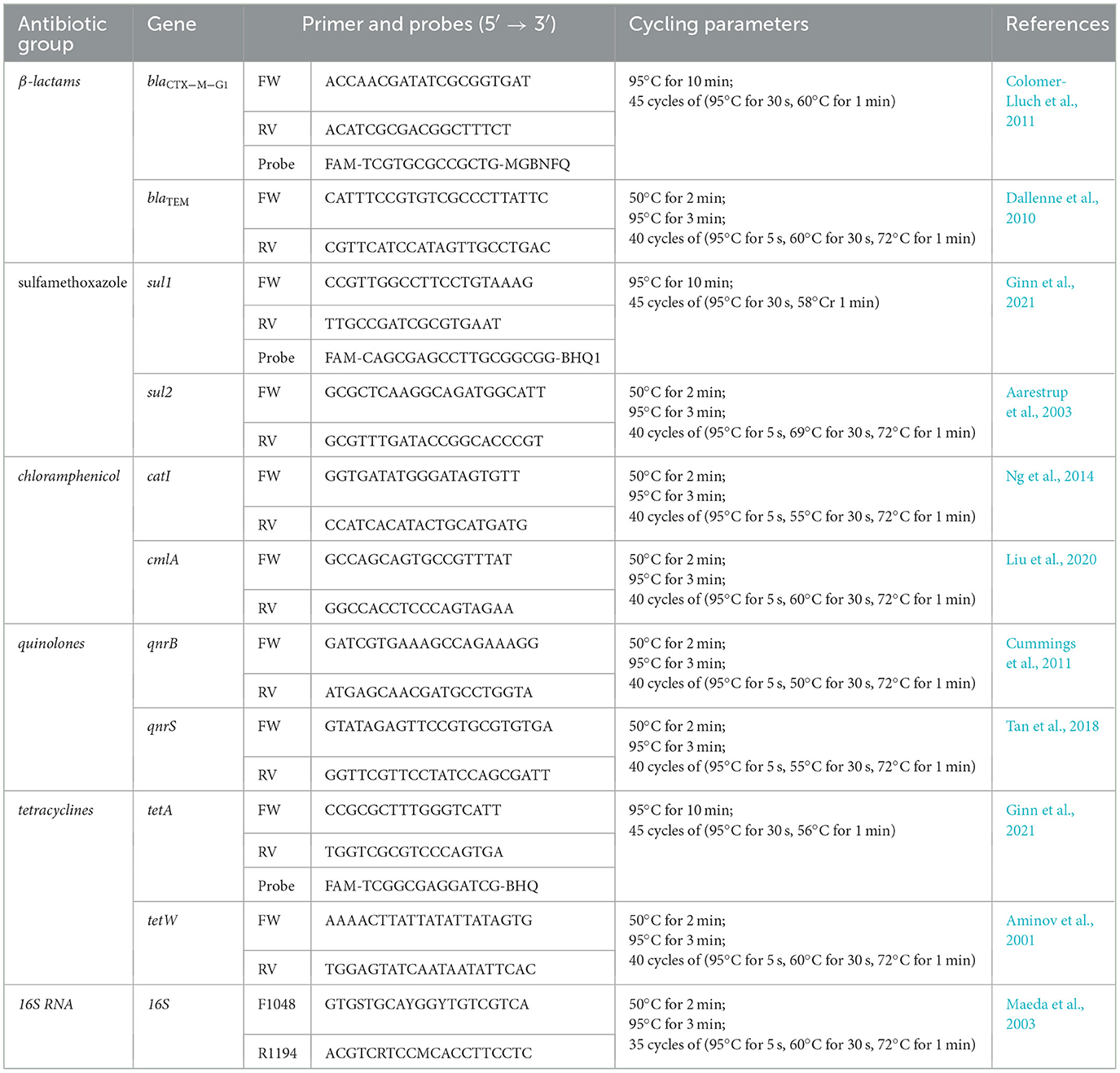
Table 2. Selected primers and probes used to quantify ARGs and the cycling parameters us in the PCR.
2.7 Statistical analysis
Treatment efficacy was calculated as follows: Log10 influent (cfu or gene copies/100 mL) – Log10 effluent (cfu or gene copies/100 mL). Graphical representations were created using R software (R Core Team, 2021) and SigmaPlot 14 (Systat Software, Inc., Addilink Software Scientific, S.L., Barcelona, Spain).
The Shapiro-Wilk test was performed to assess the normality of the data (P > 0.05). For normally distributed data, an ANOVA test was conducted, and when significant differences were detected, Tukey's HSD (Honestly Significant Difference) test was applied. The Student's t-test was used to compare the relative abundance of ARGs between influent and effluent samples.
3 Results and discussion
3.1 Effectiveness of reclamation treatments in removing ESBL-producing E. coli from WWTPs
ESBL-producing E. coli was detected in all influent samples from the four WWTPs (Figure 1). The median counts in influent samples were 6.1 ± 0.0, 5.7 ± 0.1, 5.2 ± 0.0, and 5.4 ± 0.0 log cfu/100 mL for WWTPs PAA, PAA/UV Low, UV High, and UF, respectively (Figure 1). These concentrations are in line with those reported in previous studies on untreated urban wastewater (Haberecht et al., 2019; Schmiege et al., 2021; Xie et al., 2023), indicating that our findings are consistent with values observed in similar settings.
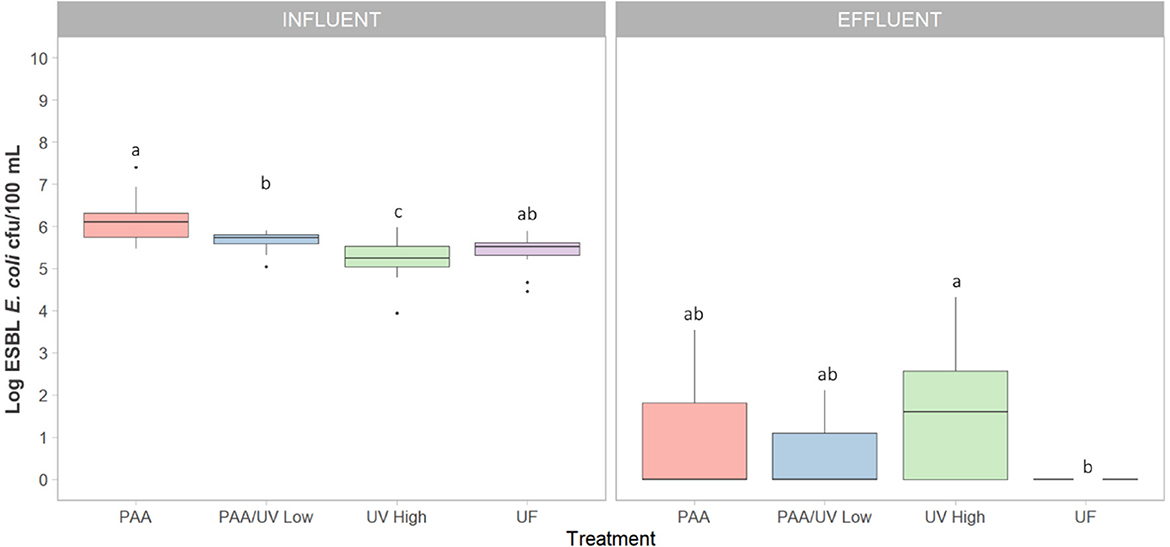
Figure 1. Level of ESBL-producing E. coli detected in the four WWTPs using different treatments. Box plots shows the median with the 25th and 75th percentile values of reduction. Box plots labeled with different letters indicate significant differences among treatments at P < 0.05.
On the other hand, differences were observed among wastewater samples, which may be influenced by various factors such as antibiotic usage patterns, population density, industrial and agricultural activities, proximity to hospitals, waste management practices, and mobility patterns. While we do not have direct evidence to determine the specific contribution of each factor, these represent plausible hypotheses that warrant further investigation. Notably, the highest levels of ESBL-producing E. coli were detected in the PAA WWTP, which is located near a hospital. This raises the possibility that hospital effluents may contribute to an increased presence of ESBL-producing E. coli in municipal WWTPs, as previously suggested by Lépesová et al. (2020).
After treatment, the ESBL-producing E. coli levels in effluent samples showed a notable reduction (Figure 1), with significant differences observed among treatments. In most WWTPs (PAA, PAA/UV Low, and UF), average counts were below 1.0 log cfu/100 mL, with several samples falling below the detection limit (0 log cfu/100 mL or 1 cfu/100 mL), particularly in the case of UF treatment. UF's effectiveness can be attributed to its ability to retain bacteria, given that their size exceeds the pore size of UF membranes (Michael et al., 2022). However, UV High treatment showed median counts exceeding 1.4 log cfu/100 mL (Figure 1), which is concerning as it sometimes surpasses the limits set by Regulation (EU) 2020/741 for water quality intended for edible agricultural crops (10 cfu/100 mL for E. coli). It is important to note that this regulation focuses on microbial indicators such as E. coli but does not include controls for ARB or ARGs. The observed high counts in UV High-treated effluent may be due to incomplete treatment during the early phase of sampling (June 2021–March 2022), before the full implementation of UV and PAA treatments. Raven et al. (2019) also reported ESBL-producing E. coli levels ranging from 2 to 21 cfu/mL in reclaimed water after UV treatment.
The detection of samples below the limit of detection at several sampling points demonstrates the potential effectiveness of certain water treatments in significantly reducing ESBL-E. coli loads. This finding is consistent with previous studies that reported substantial reductions in E. coli concentrations by up to 6 log units, through biological and tertiary treatments, including chlorination and UV-based systems (Xie et al., 2023). However, the persistence of ESBL-E. coli in final effluents, even at low levels, remains a concern. Similarly, Bréchet et al. (2014) found that urban wastewater contained high concentrations of ESBL-E. coli, with WWTP treatment reducing the load by ~95%. The authors also highlighted the genotypic overlap between clinical and environmental isolates, suggesting potential dissemination from hospitals into the environment. In the Spanish context, Oliveira et al. (2023) observed that although tertiary treatments such as UV and PAA-UV combinations resulted in significant reductions, ESBL-E. coli was still detected in several effluent samples, particularly where treatment was incomplete or inconsistent. Notably, the resistance profiles of isolates remained largely unchanged before and after treatment, indicating that current WWTP processes may reduce bacterial counts without fully eliminating resistance determinants. However, high levels of ESBL-producing E. coli have also been reported in both influent (4.5 ± 2.9 cfu/mL) and effluent (61–49 cfu/mL) samples treated with chlorine in a WWTP in Japan (Azuma et al., 2022). Similarly, Nzima et al. (2020) reported influent counts ranging from 4.1 to 4.2 log cfu/mL and effluent counts between 2.5 and 3.3 log cfu/mL. These findings collectively underscore the importance of ongoing surveillance and the optimization of advanced treatment technologies to prevent the environmental dissemination of ARB and ARGs.
3.2 Antibiotic resistance profile of ESBL-producing E. coli isolates
A total of 366 ESBL-producing E. coli isolates from influent and effluent samples of the four WWTPs were analyzed. The antibiotic resistance profiles for 210, 126, 20, and 10 isolates from PAA, PAA/UV Low, UV High, and UF WWTPs, respectively, are shown in Figures 2–5. All isolates exhibited resistance to multiple antibiotics, classifying them as MDR since they were resistant to at least three different antimicrobial classes.
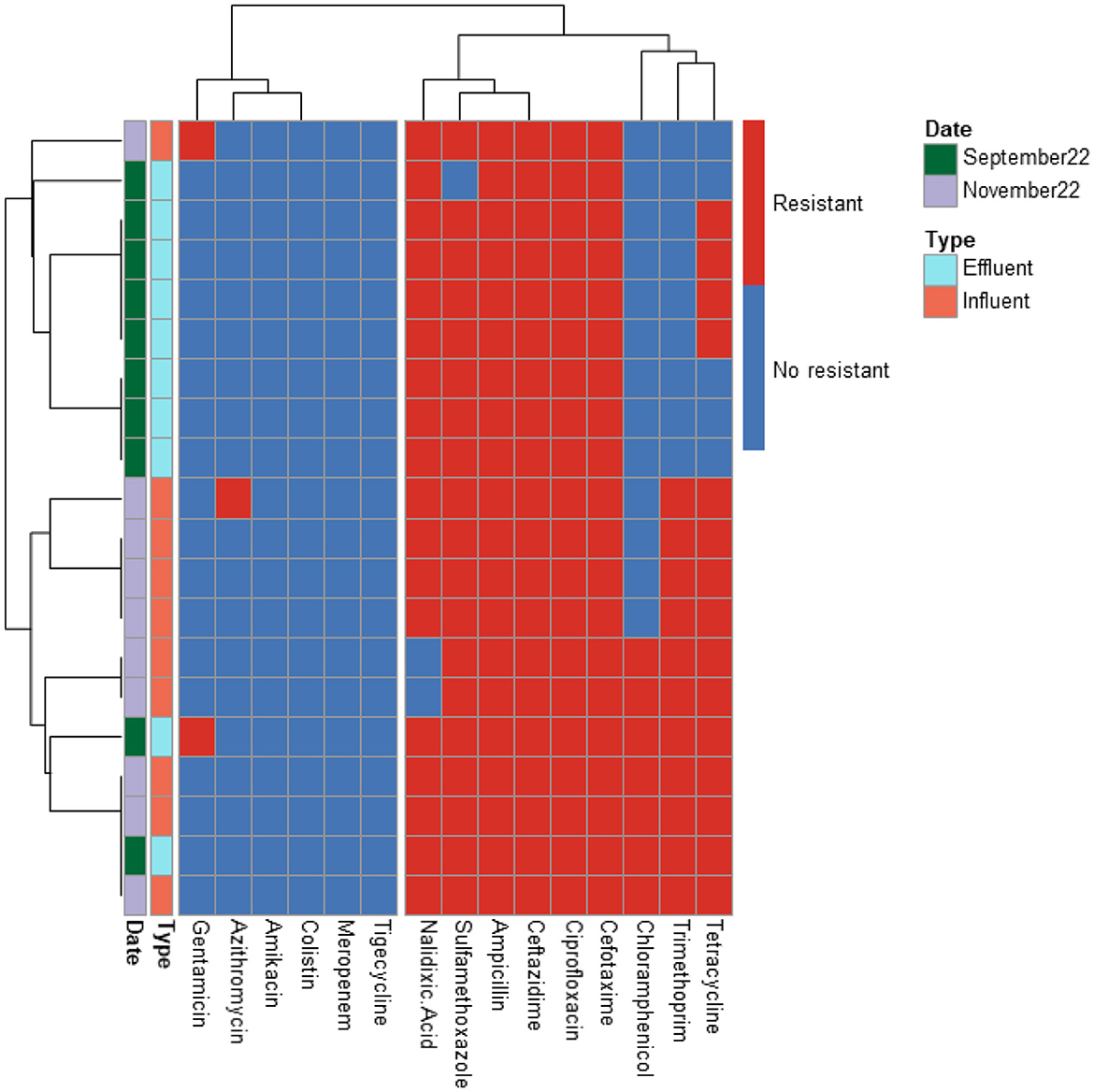
Figure 2. Heatmap displaying the antibiotic resistance profile of ESBL-producing E. coli isolates retrieved from PAA WWTP. The isolates (n = 20) were classified as antibiotic resistant (in red) or susceptible (in blue) based on ECOFF values from EUCAST, which served as the threshold.
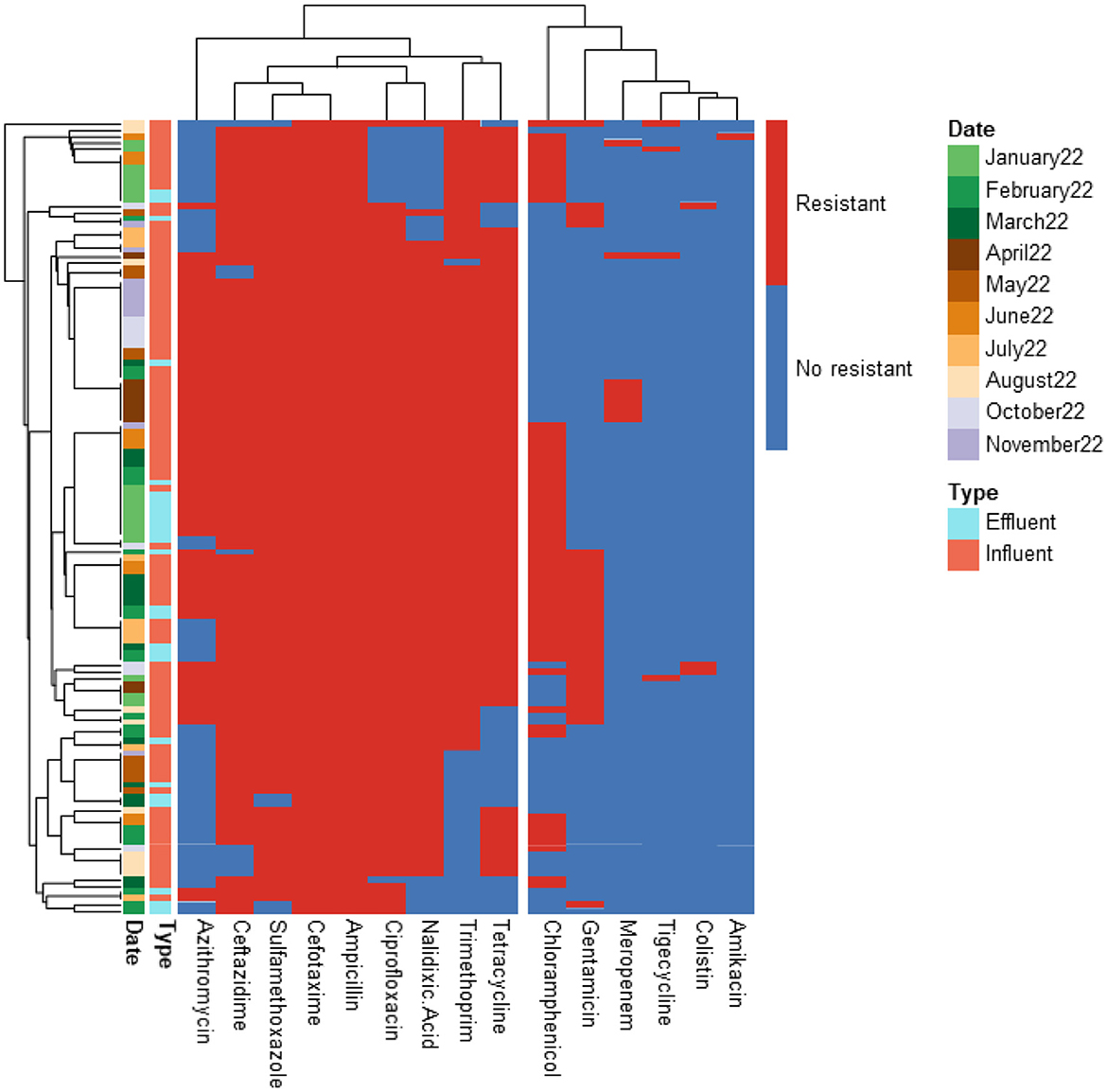
Figure 3. Heatmap displaying the antibiotic resistance profile of ESBL-producing E. coli isolates retrieved from PAA/UV Low WWTP. The isolates (n = 126) were classified as antibiotic resistant (in red) or susceptible (in blue) based on ECOFF values from EUCAST, which served as the threshold.
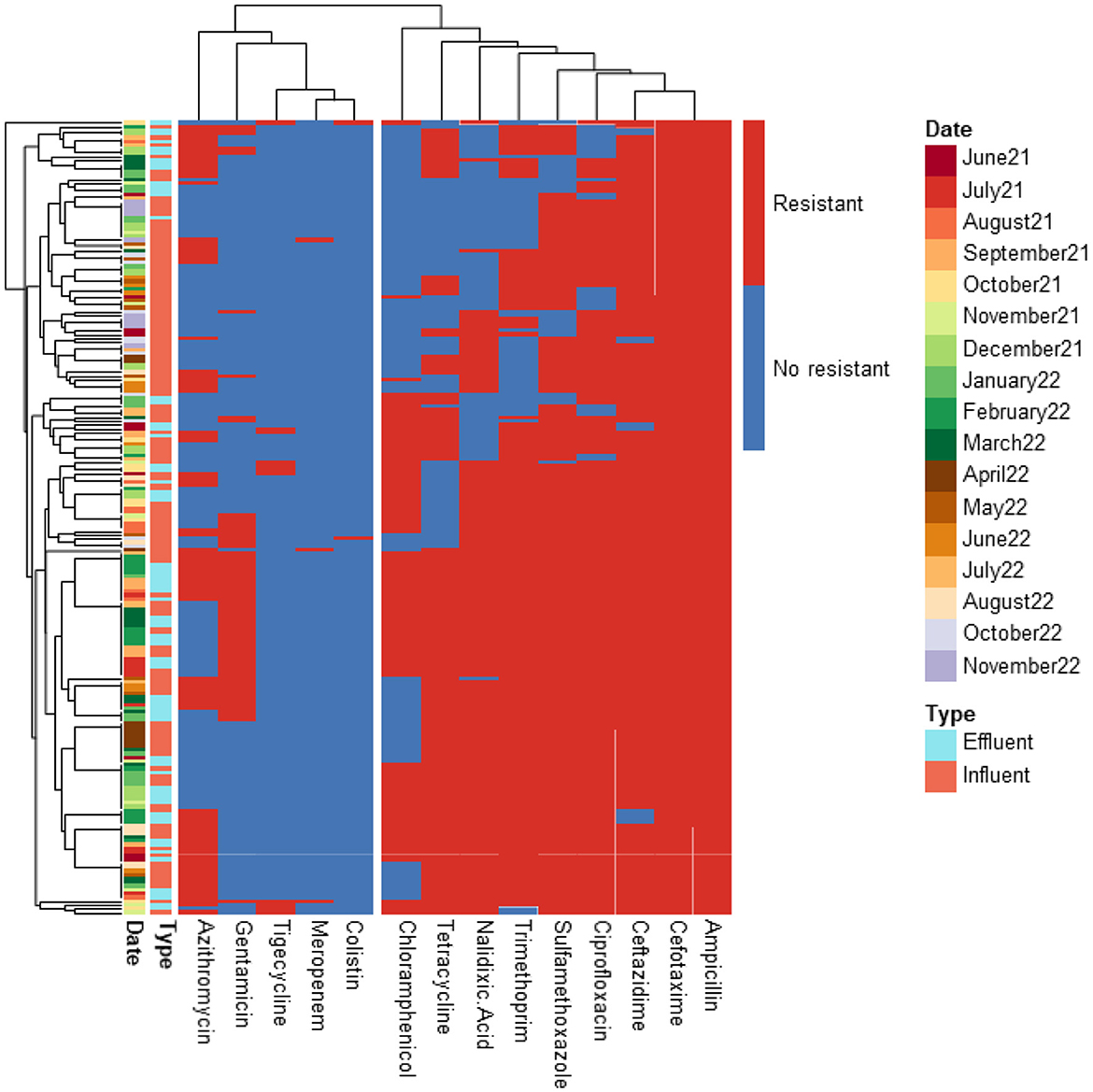
Figure 4. Heatmap displaying the antibiotic resistance profile of ESBL-producing E. coli isolates retrieved from UV High WWTP. The isolates (n = 210) were classified as antibiotic resistant (in red) or susceptible (in blue) based on ECOFF values from EUCAST, which served as the threshold.
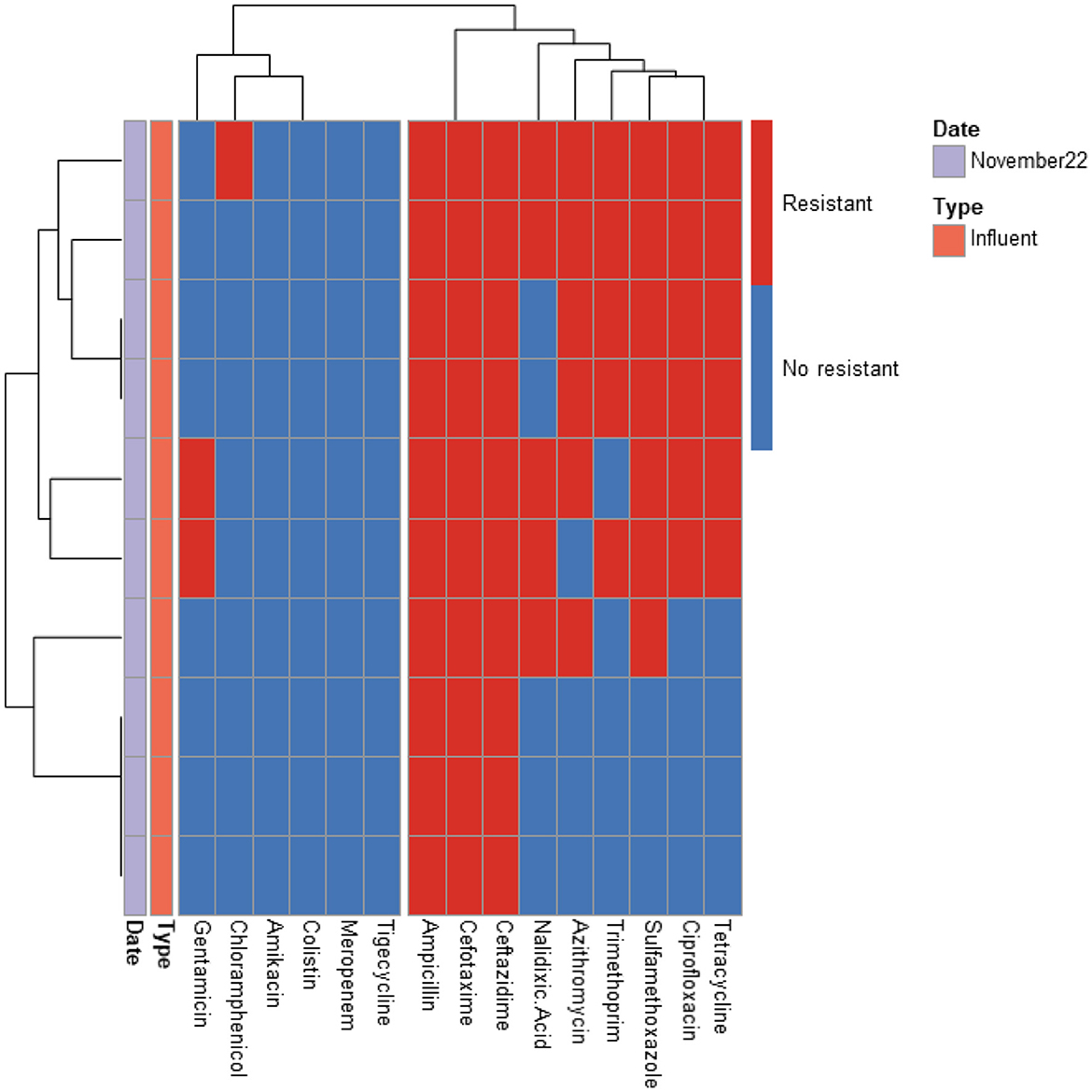
Figure 5. Heatmap displaying the antibiotic resistance profile of ESBL-producing E. coli isolates retrieved from UF WWTP. The isolates (n = 10) were classified as antibiotic resistant (in red) or susceptible (in blue) based on ECOFF values from EUCAST, which served as the threshold.
The predominant resistance phenotypes included: β-lactams (cefotaxime, ceftazidime, ampicillin); Quinolones (ciprofloxacin, nalidixic acid); Tetracyclines (tetracycline); Sulfonamides (sulfamethoxazole); Dihydrofolate reductase inhibitors (trimethoprim). However, meropenem, tigecycline, colistin, and amikacin retained antibacterial activity against most isolates. A greater number of isolates (15 isolates) exhibited resistance to tigecycline compared to meropenem (12 isolates), colistin (5 isolates), and amikacin (2 isolates). The tigecycline-resistant isolates were found in the influent of the UV High and PAA/UV Low WWTPs, as well as in the effluent of the UV High WWTP. Most of the meropenem-resistant isolates were detected in the influent samples from the PAA/UV Low WWTP. Colistin-resistant isolates were identified in an effluent sample from the UV High WWTP and in influent samples from the UV High and PAA/UV Low WWTPs. The two isolates resistant to amikacin were originated from the influent samples from the UV High and PAA/UV Low WWTPs. Notably, two isolates from the influent of UV High and PAA/UV Low showed combined resistance to tigecycline and meropenem, while one isolate from an effluent sample from the UV High WWTP showed combined resistance to tigecycline and colistin. The presence of these resistant phenotypes in extended-spectrum β-lactamase (ESBL)-producing isolates, although not predominant, is noteworthy. Glycylcyclines (tigecycline), polymyxins (colistin), and carbapenems (meropenem) are considered last-resort antibiotics, primarily recommended for treating infections caused by ESBL-producing Gram-negative bacteria. Resistance to these antibiotics was not detected in isolates from PAA and UF WWTPs. However, it is important to note that the number of isolates analyzed from these WWTPs was considerably lower, which may have influenced the results. Similarly, Amador et al. (2015) reported a high prevalence of antibiotic resistance in Enterobacteriaceae from WWTPs, including resistance to β-lactam antibiotics, tetracycline, ciprofloxacin, and trimethoprim/sulfamethoxazole. Other studies have also characterized AMR profiles of E. coli isolates from WWTPs, frequently detecting resistance to sulfonamides, tetracyclines, and aminopenicillins, while resistance to quinolones was reported at lower rates (Ferreira da Silva et al., 2006; Łuczkiewicz et al., 2010).
Our findings suggest that ESBL-producing E. coli isolates recovered from influent and effluent samples commonly exhibit multidrug resistance phenotypes, including resistance to β-lactams, quinolones, tetracyclines, and sulfonamides. While descriptive data did not show major shifts in resistance profiles between sample types or sampling dates, the limited number of isolates from some WWTPs prevent definitive conclusions. The detection of isolates resistant to last-resort antibiotics, particularly in effluent samples from certain WWTPs, warrants further investigation to clarify whether wastewater treatment processes may contribute to the selection or persistence of specific resistance phenotypes
Nonetheless, the presence of MDR in effluent samples remains a public health concern, as reclaimed water is frequently reused for irrigation in agricultural fields. This highlights the need for continuous monitoring of AMR in WWTPs to mitigate potential risks associated with environmental and human exposure.
3.3 Monitoring of absolute ARG abundance in reclamation treatments at WWTPs
In this study, the 16S rRNA gene and 10 ARGs were detected in all influent and effluent water samples from the four WWTPs (Table 3), except for the sul1 gene in the effluent of the UV High WWTP and both sul1 and catl genes in the effluent of the UF WWTP. These genes have been frequently reported in wastewater across various geographical regions, including Europe, America, Asia, and Africa (Oliveira et al., 2023; Pazda et al., 2019; Wang et al., 2020). In all WWTP influents, the most prevalent ARG was tetW (11.1–11.7 gc/100 mL), followed by tetA (10.2–10.9 gc/100 mL), cmlA (10.8–11.3 gc/100 mL), and qnrS (8.9–9.3 gc/100 mL; Figure 6).
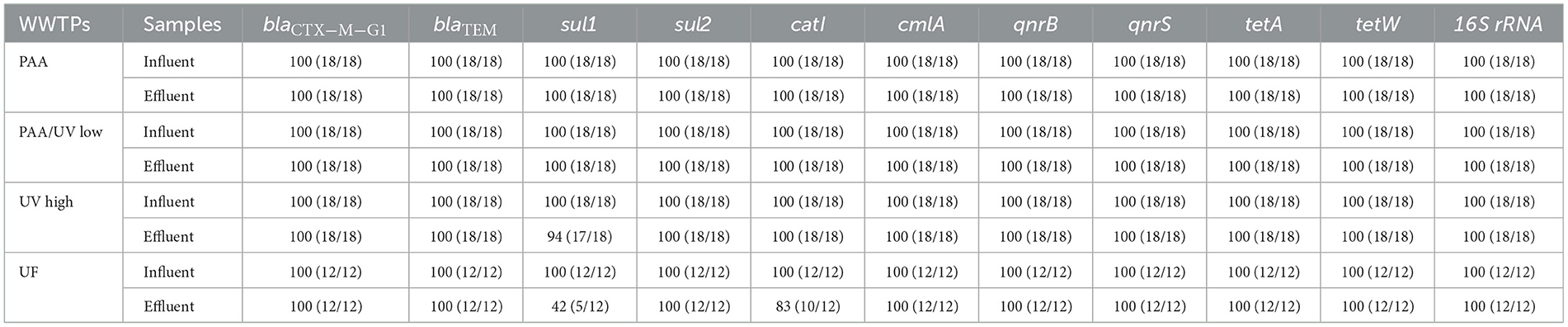
Table 3. Prevalence of ARGs analyzed in the current study.
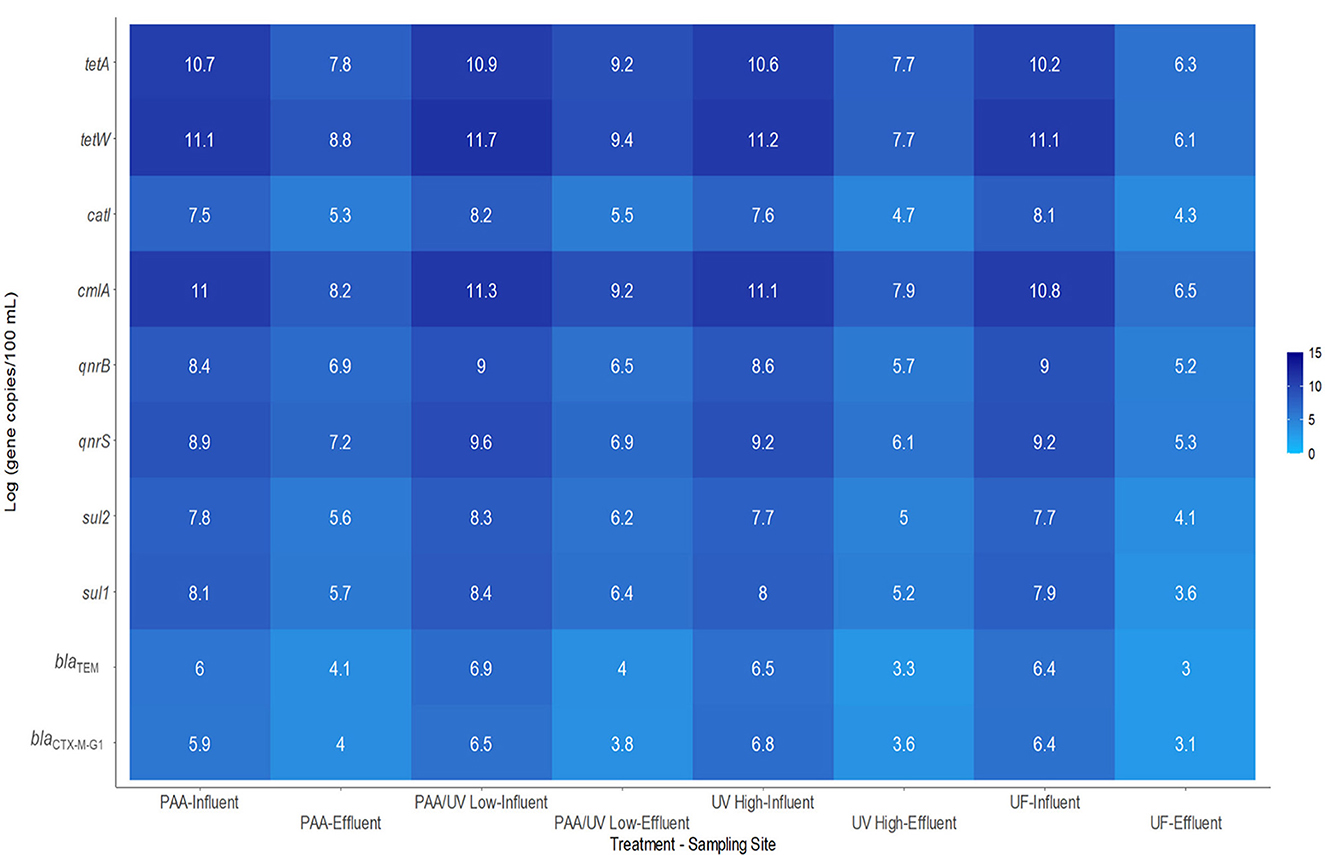
Figure 6. Heatmap representing the absolute levels of ARGs measured by qPCR in wastewater samples upon arrival at the WWTP studied and in its effluents after applying the tertiary treatment corresponding to each plant over an 18-month sampling period. Values are presented as Log (gene copies/100 mL).
The absolute abundance of ARGs in influent samples ranged as follows: (i) PAA: 5.9–11.1 gc/100 mL; (ii) PAA/UV Low: 6.5–11.7 gc/100 mL; (iii) UV High: 6.5–11.2 gc/100 mL; and (iv) UF: 6.4–11.1 gc/100 mL (Figure 6). On the other hand, effluent samples exhibited significantly lower ARG levels (P ≤ 0.05): (i) PAA: 4.0–8.3 gc/100 mL; (ii) PAA/UV Low: 3.8–9.4 gc/100 mL; (iii) UV High: 3.6–7.7 gc/100 mL; (iv) WWTP UF: 3.0–6.1 gc/100 mL (Figure 6). Consistent with influent samples, tetW, tetA, cmlA, and qnrS remained the most abundant genes in effluents (Figure 6). The high prevalence of tetracycline and quinolone resistance genes in wastewater and effluent is likely due to the widespread use of these broad-spectrum antibiotics. These findings align with previous studies demonstrating that tetracycline, quinolone, sulfonamide, and macrolide resistance genes are frequently detected in WWTP effluents (Pazda et al., 2019; Wang et al., 2020). While wastewater treatment reduced ARG abundance, significant levels were still detected in effluents, consistent with other reports (Luo et al., 2024; Munir et al., 2011; Pallares-Vega et al., 2019; Raza et al., 2022).
Significant differences were observed in ARG levels among the WWTPs (Figure 7). UF treatment achieved the most substantial reduction in ARGs, consistent with studies suggesting that membrane filtration is an effective approach for removing emerging contaminants (Guo et al., 2021; Zhang et al., 2023). This high effectiveness is likely due to the physical retention of bacterial cells and extracellular DNA fragments by the membrane pores, which are small enough to block ARG carriers. However, studies on ARG removal in reclaimed water systems using UF remain limited.

Figure 7. Absolute concentration of the 10 ARGs in the influent and effluent samples collected from each WWTPs. Box plots show median with the 25th and 75th percentile values of reduction. Box plots labeled with different lower-case letters indicate significant differences among influents at P < 0.05. Box plots labeled with different upper-case letters indicate significant differences among treatments at P < 0.05.
Conversely, PAA/UV Low WWTP exhibited the highest absolute ARG levels in effluent, despite the fact that PAA/UV is often proposed as a promising disinfection strategy. However, a previous laboratory-scale study reported that PAA/UV Low treatment resulted in lower ARG levels than PAA or UV High alone (Ping et al., 2022). Several factors could explain these discrepancies. First, operational conditions such as variations in PAA dose, UV intensity, and contact time could have impacted the overall efficiency. For instance, if UV exposure or PAA concentration is insufficient or poorly synchronized, incomplete inactivation of microorganisms or ARG degradation may occur. In this particular case, the UV High doses used in that study (216 mJ/cm2) were lower than those applied in this study. All tertiary treatments reduced 16S rRNA gene levels, indicating a decrease in total bacterial load (Figure 8). UF showed the most significant reduction, suggesting its superior effectiveness in removing bacteria from wastewater. These results align with previous research evaluating 16S rRNA levels in WWTP influent and effluent (Pallares-Vega et al., 2019; Wang et al., 2021). The observed differences across WWTPs may therefore be influenced by a combination of treatment-specific parameters and site-specific factors, such as influent microbial load, composition of the wastewater, or presence of resistance-selection agents (e.g., residual antibiotics or heavy metals). These aspects highlight the complexity of ARG removal and suggest that further optimization and standardization of treatment parameters are essential to improve performance across different settings.

Figure 8. Absolute concentration of the 16S rRNA gene in the influent and effluent samples collected from each WWTPs. Box plots show the median with the 25th and 75th percentile values of reduction. Box plots labeled with different letters indicate significant differences among treatments at P < 0.05.
3.4 Monitoring of the relative abundance of ARGS in the water treatments applied at the WWTPs
To evaluate the impact of the four water treatments on ARG occurrence in WWTPs, the relative abundance of 10 ARGs was quantified as copies of ARGs per 16S rRNA gene copies using quantitative polymerase chain reaction (qPCR). The results showed that all four WWTPs either maintained or reduced the relative abundance of the analyzed ARGs, except for blaCTX − M−G1 and blaTEM in the UF-treated effluent and qnrB and qnrS in the PAA-treated effluent (Figure 9). These findings align with previous studies, where some ARGs exhibited slight reductions or remained unchanged after wastewater treatment (Bengtson-Palme et al., 2016; Di Cesare et al., 2016; Laht et al., 2014). Similarly, McConnell et al. (2018) found no increase in ARG abundance following tertiary UV treatment at two WWTPs in Canada. Pallares-Vega et al. (2019) also reported no increase in sul1, sul2, tetM, qnrS, or blaCTX − M relative abundance in 62 Belgian WWTPs. Conversely, other studies have reported increases in ARG levels following wastewater treatment, including Lee et al. (2017), Makowska et al. (2016), Marti et al. (2013), and Rodriguez-Mozaz et al. (2014). These conflicting results may stem from variations in microbial community composition post-treatment, the presence of AMR-selective agents in effluents, and differences in sampling design (Pallares-Vega et al., 2019).
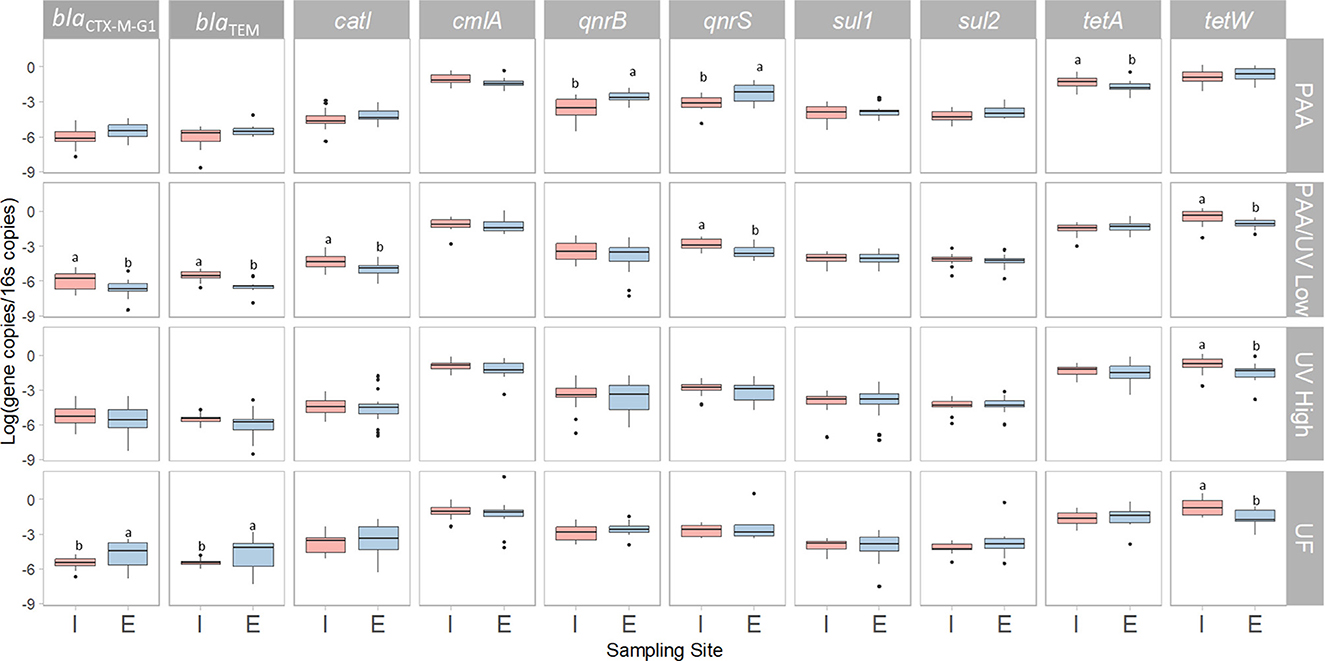
Figure 9. Relative abundance of ARGs in the influent (I) and effluent (E) samples collected from each WWTP. Box plots show the median with the 25th and 75th percentile values of reduction. Box plots labeled with different letters indicate significant differences between influent and effluent at P < 0.05.
3.5 Efficacy of tertiary treatments in WWTPs
Significant differences were observed between UV High and the other treatments (PAA, PAA/UV Low, and UF). The obtained results are consistent with studies evaluating the efficacy of UF, PAA/UV, and UV in removing ARBs (Balachandran et al., 2021; Chhetri et al., 2022; McKinney and Pruden, 2012; Michael et al., 2022; Oliveira et al., 2023; Raven et al., 2019). For instance, when UF is applied to activated sludge effluent achieved a 3–4 log reduction of enteric opportunistic pathogens and significant ARB removal (Michael et al., 2022). The application of a PAA dose of 4 mg/L for 7 min led to a 4-log reduction in multidrug-resistant E. coli (Balachandran et al., 2021). In fact, PAA treatment was effective in reducing ciprofloxacin-resistant bacteria in municipal and hospital wastewater (Chhetri et al., 2022). In the case of UV-C, this treatment alone or in combination with PAA or chlorine efficiently removed ESBL-producing E. coli from influent water (Oliveira et al., 2023). UV doses of 10–20 mJ/cm2 led to 4–5 log reductions in ARBs (McKinney and Pruden, 2012). In this study, UV High showed a lower reduction than the other treatments (Figure 10), suggesting lower efficacy in removing ESBL-producing E. coli. However, the median log reduction for UV High was ~3.5 logs, similar to the 3.2 log reduction observed in UV-treated effluents from a WWTP in southern England (Raven et al., 2019). Regarding ARG removal, the different WWTPs achieved an average reduction of 2.2–3.9 log/100 mL (Figure 11). These reductions are consistent with those observed in previous studies evaluating UV, PAA, UF, and PAA/UV combinations as tertiary treatments (Manoharan et al., 2022; Wang et al., 2020).

Figure 10. Box-plot representing the reduction of ESBL-producing E. coli after tertiary water treatments applied in each WWTP studied. The 0 log reduction (ESBL_I/ESBL_E) indicates no reduction of ARGs in the water. Box plots show the median with the 25th and 75th percentile values of reduction. Box plots labeled with different letters indicate significant differences among treatments at P < 0.05.
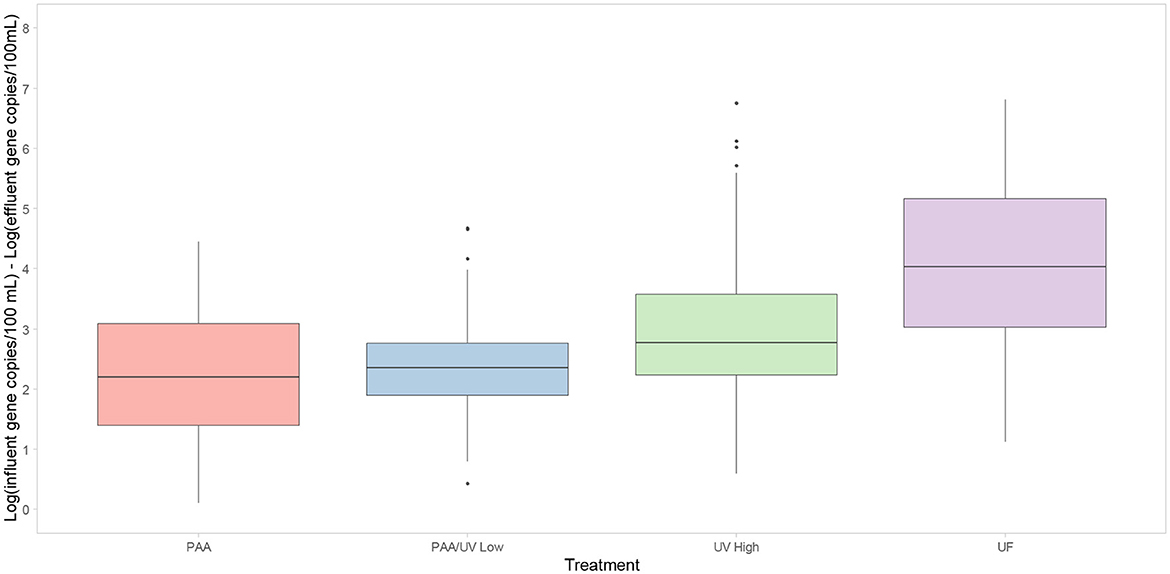
Figure 11. Box-plot representing the reduction of 10 ARGs evaluated after tertiary water treatments applied in each WWTP studied. The 0 log indicates no reduction of ARGs in the water. Box plots show median with the 25th and 75th percentile values of reduction. Box plots labeled with different letters indicate significant differences among treatments at P < 0.05.
In the case of ARG, significant differences were observed among WWTPs. UF was the most effective treatment for reducing ARG concentrations. UV High followed UF in effectiveness, with better performance than PAA/UV Low and PAA alone. On the other hand, PAA/UV Low and PAA exhibited the lowest ARG removal efficiencies (Figure 12). These findings align with Liang et al. (2021), who reported that integrated membrane filtration reduced ARGs by nearly 3 logs (99.79%), from 3.02 × 108 to 6.45 × 105 cg/mL.
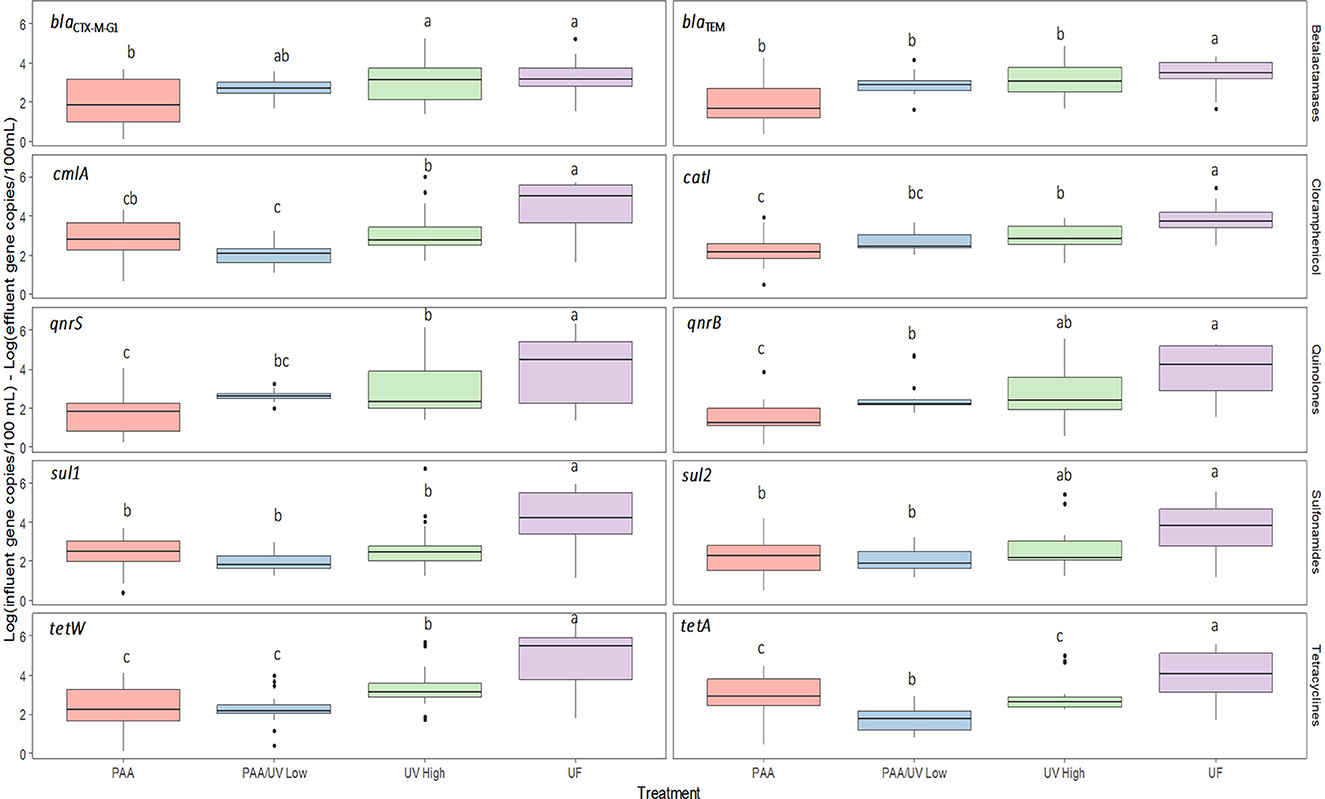
Figure 12. Reduction of each ARGS after tertiary water treatments applied in each WWTP studied. The 0 log indicates no reduction of ARGs in the water. Box plots show median with the 25th and 75th percentile values of reduction. Box plots labeled with different letters indicate significant differences among treatments at P < 0.05.
Several studies have demonstrated the ability of UV light to reduce ARGs in wastewater (Jia et al., 2021; Zhuang et al., 2014). However, other studies have reported minimal or no ARG reductions following UV treatment (Auerbach et al., 2007; Lee et al., 2017; Munir et al., 2011; Yang et al., 2019). These discrepancies are likely due to variations in UV dose. For instance, McConnell et al. (2018) found that WWTPs using 250 mJ/cm2 UV doses achieved greater ARG removal than those using lower doses (50 mJ/cm2). Studies suggest that combining PAA with low UV doses enhances ARG removal, but in this study, UV High alone was more effective than PAA/UV Low or PAA alone. This is likely due to higher UV doses applied in that WWTP. Ping et al. (2022) recently demonstrated that a high UV dose (108 mJ/cm2) and PAA (4 mg/L) effectively reduced ARGs, whereas lower UV doses (18 mJ/cm2) with the same PAA concentration were less effective. These findings highlight the role of tertiary treatments in mitigating ARG prevalence, emphasizing the importance of effective water treatments before discharge to reduce antibiotic resistance risks.
4 Conclusions
This study assesses the efficacy of different tertiary wastewater treatment technologies in reducing ESBL-producing E. coli and ARGs in reclaimed water intended for irrigating horticultural crops. The findings highlight the role of these treatments in limiting AMR dissemination and their implications for water reuse safety. Tertiary treatments significantly reduced ESBL-producing E. coli and ARGs, though complete elimination was not achieved. Ultrafiltration (UF) was the most effective, outperforming peracetic acid (PAA), PAA/UV Low, and UV High treatments. While absolute ARG abundance decreased across all WWTPs, relative abundance remained stable or only slightly reduced, with tetracycline (tetW, tetA), quinolone (qnrS, qnrB), and sulfonamide (sul1, sul2) resistance genes being the most persistent. UV efficacy was dose-dependent, with UV High performing better than PAA/UV Low and PAA alone, emphasizing the need to optimize UV intensity for ARG inactivation. Antibiotic resistance profiles revealed MDR E. coli in all WWTPs, primarily resistant to β-lactams, quinolones, tetracyclines, and sulfonamides. Additionally, resistance to last-resort antibiotics (tigecycline, colistin, meropenem) was detected at low frequencies, raising concerns about the potential dissemination of highly resistant bacterial strains through wastewater effluents. These findings underscore the need for improved wastewater treatment strategies and stricter monitoring to mitigate AMR risks in reclaimed water intended for agricultural use.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
PT: Data curation, Formal analysis, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing. MO: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. RC-G: Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. MA: Conceptualization, Resources, Writing – review & editing. AR: Conceptualization, Funding acquisition, Resources, Supervision, Writing – review & editing. FG: Conceptualization, Funding acquisition, Resources, Writing – review & editing. AÁ-O: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing. AA: Conceptualization, Formal analysis, Funding acquisition, Investigation, Resources, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors are thankful for the financial support from the Spanish Ministry of Science and Innovation (Project TED2021-131427B-C21 and TED2021-131427B-C22), as well as the a Fundación Séneca – Agencia de Ciencia y Tecnología de la Región de Murcia, FSRM/10.13039/100007801 (22713/PI/24), Spain, and AGROALNEXT programme supported by MCIN with funding from European Union NextGenerationEU (PRTR-C17.I1).
Acknowledgments
The technical assistance of Macarena Moreno (CEBAS-CSIC) and Pedro J. Simón-Andreu (ESAMUR) was very much appreciated.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Gen AI was used in the creation of this manuscript. Revise the English of the manuscript. To elaborate the first draft of the summary.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aarestrup, F. M., Lertworapreecha, M., Evans, M. C., Bangtrakulnonth, A., Chalermchaikit, T., Hendriksen, R. S., et al. (2003). Antimicrobial susceptibility and occurrence of resistance genes among Salmonella enterica serovar Weltevreden from different countries. J. Antimicrob. Chemother. 52, 715–718. doi: 10.1093/jac/dkg426
Amador, P. P., Fernandes, R. M., Prudencio, M. C., Barreto, M. P., and Duarte, I. M. (2015). Antibiotic resistance in wastewater: occurrence and fate of Enterobacteriaceae producers of class A and class C β-lactamases. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 50, 26–39. doi: 10.1080/10934529.2015.964602
Aminov, R. I., Garrigues-Jeanjean, N., and Mackie, R. I. (2001). Molecular ecology of tetracycline resistance: development and validation of primers for detection of tetracycline resistance genes encoding ribosomal protection proteins. Appl. Environ. Microbiol. 67, 22–32. doi: 10.1128/AEM.67.1.22-32.2001
Auerbach, E. A., Seyfried, E. E., and McMahon, K. D. (2007). Tetracycline resistance genes in activated sludge wastewater treatment plants. Water Res. 41, 1143–1151. doi: 10.1016/j.watres.2006.11.045
Azuma, T., Usui, M., and Hayashi, T. (2022). Inactivation of antibiotic-resistant bacteria in wastewater by ozone-based advanced water treatment processes. Antibiotics 11:210. doi: 10.3390/antibiotics11020210
Balachandran, S., Charamba, L. V. C., Manoli, K., Karaolia, P., Caucci, S., and Fatta-Kassinos, D. (2021). Simultaneous inactivation of multidrug-resistant Escherichia coli and Enterococci by peracetic acid in urban wastewater: exposure-based kinetics and comparison with chlorine. Water Res. 202:117403. doi: 10.1016/j.watres.2021.117403
Ben, W., Wang, J., Cao, R., Yang, M., Zhang, Y., and Qiang, Z. (2017). Distribution of antibiotic resistance in the effluents of ten municipal wastewater treatment plants in China and the effect of treatment processes. Chemosphere 172, 392–398. doi: 10.1016/j.chemosphere.2017.01.041
Bengtson-Palme, J., Hammerén, R., Pal, C., Östman, M., Björnlenius, B., Flach, C.-F., et al. (2016). Elucidating selection processes for antibiotic resistance in sewage treatment plants using metagenomics. Sci. Total Environ. 572, 697–712. doi: 10.1016/j.scitotenv.2016.06.228
Bréchet, C., Plantin, J., Sauget, M., Thouverez, M., Talon, D., Cholley, P., et al. (2014). Wastewater treatment plants release large amounts of extended-spectrum β-lactamase-producing Escherichia coli into the environment. Clin. Infect. Dis. 58, 1658–1665. doi: 10.1093/cid/ciu190
Buriánková, I., Kuchta, P., Molíková, A., Sovová, K., Výravský, D., Rulík, M., et al. (2021). Antibiotic resistance in wastewater and its impact on a receiving river: a case study of WWTP Brno-Modrice, Czech Republic. Water 13:2309. doi: 10.3390/w13162309
Chhetri, R. K., Sanchez, D. F., Lindholst, S., Hansen, A. V., Sanderbo, J., Løppenthien, B. K., et al. (2022). Disinfection of hospital-derived antibiotic-resistant bacteria at source using peracetic acid. J. Water Process Eng. 45:102507. doi: 10.1016/j.jwpe.2021.102507
Colomer-Lluch, M., Jofre, J., and Muniesa, M. (2011). Antibiotic resistance genes in the bacteriophage DNA fraction of environmental samples. PLoS ONE 6:e17549. doi: 10.1371/journal.pone.0017549
Cummings, D. E., Archer, K. F., Arriola, D. J., Baker, P. A., Faucett, K. G., Laroya, J. B., et al. (2011). Broad dissemination of plasmid-mediated quinolone resistance genes in sediments of two urban coastal wetlands. Environ. Sci. Technol. 45, 447–454. doi: 10.1021/es1029206
Dallenne, C., Da Costa, A., Decré, D., Favier, C., and Arlet, G. (2010). Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 65, 490–495. doi: 10.1093/jac/dkp498
Di Cesare, A., Eckert, E. M., D'Urso, S., Bertoni, R., Gillan, D., Wattiez, R., et al. (2016). Co-occurrence of integrase 1, antibiotic, and heavy metal resistance genes in a municipal wastewater treatment plant. Water Res. 94, 208–214. doi: 10.1016/j.watres.2016.02.049
Echaide, M., Rivas, A., Díaz-Cruz, M. S., and Barceló, D. (2021). Antibiotic resistance in reclaimed water used for irrigation: management strategies to ensure its safe use. Curr. Opin. Environ. Sci. Health 20:100234. doi: 10.1007/978-3-319-78000-9_178
EFSA (European Food Safety Authority, Panel on Biological Hazards). (2021). Scientific Opinion on the role played by the environment in the emergence and spread of antimicrobial resistance (AMR) through the food chain. EFSA J. 19:6651, 188 pp.
European Commission (2017). European One Health Action Plan against Antimicrobial Resistance (AMR). Brussels: European Commission. Available online at: https://health.ec.europa.eu/system/files/2020-01/amr_2017_action-plan_0.pdf
European Commission (2020a). A Farm to Fork Strategy for a Fair, Healthy and Environmentally-Friendly Food System. Available online at: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A52020DC0381
European Commission (2020b). Regulation (EU) 2020/741 of the European Parliament and of the Council of 25 May 2020 on minimum requirements for water reuse. Off. J. Eur. Union L 177/32, 32–55.
Ferreira da Silva, M., Tiago, I., Veríssimo, A., Boaventura, R. A. R., Nunes, O. C., and Manaia, C. M. (2006). Antibiotic resistance of enterococci and related bacteria in an urban wastewater treatment plant. FEMS Microbiol. Ecol. 55, 322–329. doi: 10.1111/j.1574-6941.2005.00032.x
Gao, Y. X., Li, X., Fan, X. Y., Zhao, J. R., and Zhang, Z. X. (2022). Wastewater treatment plants as reservoirs and sources for antibiotic resistance genes: a review on occurrence, transmission, and removal. J. Water Process Eng. 188:116482. doi: 10.1016/j.jwpe.2021.102539
GBD 2021 Antimicrobial Resistance Collaborators (2024). Global burden of bacterial antimicrobial resistance 1990–2021: a systematic analysis with forecasts to 2050. Lancet 404, 1199–1226. doi: 10.1016/S0140-6736(24)01867-1
Ginn, O., Nichols, D., Rocha-Melogno, L., Bivins, A., Berendes, D., Soria, F., et al. (2021). Antimicrobial resistance genes are enriched in aerosols near impacted urban surface waters in La Paz, Bolivia. Environ. Res. 194:110730. doi: 10.1016/j.envres.2021.110730
Guo, Y., Liang, H., Li, G., Xu, D., Yan, Z., Chen, R., et al. (2021). A solar photo-thermochemical hybrid system using peroxydisulfate for organic matter removal and improving ultrafiltration membrane performance in surface water treatment. Water Res. 188:116482. doi: 10.1016/j.watres.2020.116482
Haberecht, H. B., Nealon, N. J., Gilliland, J. R., Holder, A. V., Runyan, C., Oppel, R. C., et al. (2019). Antimicrobial-resistant Escherichia coli from environmental waters in Northern Colorado. J. Environ. Public Health 2019:3862949. doi: 10.1155/2019/3862949
Hazra, M., Watts, J. E. M., Williams, J. B., and Joshi, H. (2024). An evaluation of conventional and nature-based technologies for controlling antibiotic-resistant bacteria and antibiotic resistance genes in wastewater treatment plants. Sci. Total Environ. 917:170433. doi: 10.1016/j.scitotenv.2024.170433
Hultman, J., Tamminen, M., Pärnänen, K., Cairns, J., Karkman, A., and Virta, M. (2018). Host range of antibiotic resistance genes in wastewater treatment plant influent and effluent. FEMS Microbiol. Ecol. 94:fiy038. doi: 10.1093/femsec/fiy038
Jia, S., Li, T., and Zhang, X. X. (2021). Integrated metagenomic and metatranscriptomic analyses of ultraviolet disinfection effects on antibiotic resistance genes and bacterial communities during wastewater treatment. Ecotoxicology 30, 1610–1619. doi: 10.1007/s10646-020-02313-1
Kumar, M., Ram, B., Sewwandi, H., Sulfikar, H., Honda, R., and Chaminda, T. (2020). Treatment enhances the prevalence of antibiotic-resistant bacteria and antibiotic resistance genes in the wastewater of Sri Lanka and India. Environ. Res. 183:109179. doi: 10.1016/j.envres.2020.109179
Laht, M., Karkman, A., Voolaid, V., Ritz, C., Tenson, T., Virta, M., et al. (2014). Abundances of tetracycline, sulphonamide, and beta-lactam antibiotic resistance genes in conventional wastewater treatment plants (WWTPs) with different waste load. PLoS ONE 9:e88882. doi: 10.1371/journal.pone.0103705
Lee, J., Jeon, J. H., Shin, J., Jang, H. M., Kim, S., Son, M. S., et al. (2017). Quantitative and qualitative changes in antibiotic resistance genes after passing through treatment processes in municipal wastewater treatment plants. Sci. Total Environ. 605–606, 906–914. doi: 10.1016/j.scitotenv.2017.06.250
Lépesová, K., Olejníková, P., Mackulak, T., Cverenkárová, K., Krahulcová, M., and Bírošová, L. (2020). Hospital wastewater – important source of multidrug-resistant coliform bacteria with ESBL production. Int. J. Environ. Res. Public Health 17:7827. doi: 10.3390/ijerph17217827
Li, S., Ondon, B. S., Ho, S. H., Jiang, J., and Li, F. (2022). Antibiotic-resistant bacteria and genes in wastewater treatment plants: from occurrence to treatment strategies. Sci. Total Environ. 838:156544. doi: 10.1016/j.scitotenv.2022.156544
Liang, C., Wei, D., Zhang, S., Ren, Q., Shi, J., and Liu, L. (2021). Removal of antibiotic resistance genes from swine wastewater by membrane filtration treatment. Ecotoxicol. Environ. Saf. 210:111885. doi: 10.1016/j.ecoenv.2020.111885
Liu, X., Wang, H., and Zhao, H. (2020). Propagation of antibiotic resistance genes in an industrial recirculating aquaculture system located in northern China. Environ. Pollut. 261:114155. doi: 10.1016/j.envpol.2020.114155
Łuczkiewicz, A., Jankowska, K., Fudala-Ksiażek, S., and Olańczuk-Neyman, K. (2010). Antimicrobial resistance of fecal indicators in municipal wastewater treatment plant. Water Res. 44, 5089–5097. doi: 10.1016/j.watres.2010.08.007
Luo, L., Wang, Z., Huang, X., Gu, J. D., Yu, C., and Deng, O. (2024). The fate of antibiotic resistance genes in wastewater containing microalgae treated by chlorination, ultraviolet, and Fenton reaction. Water Res. 254:121392. doi: 10.1016/j.watres.2024.121392
Macrì, M., Bonetta, S., Di Cesare, A., Sabatino, R., Corno, G., Catozzo, M., et al. (2024). Antibiotic resistance and pathogen spreading in a wastewater treatment plant designed for wastewater reuse. Environ. Pollut. 363:125051. doi: 10.1016/j.envpol.2024.125051
Maeda, H., Fujimoto, C., Haruki, Y., Maeda, T., Kokeguchi, S., Petelin, M., et al. (2003). Quantitative real-time PCR using TaqMan and SYBR Green for Actinobacillus actinomycetemcomitans, Porphyromonas gingivalis, Prevotella intermedia, tetQ gene, and total bacteria. FEMS Immunol. Med. Microbiol. 39, 81–86. doi: 10.1016/S0928-8244(03)00224-4
Makowska, N., Koczura, R., and Mokracka, J. (2016). Class 1 integrase, sulfonamide, and tetracycline resistance genes in wastewater treatment plant and surface water. Chemosphere 144, 1665–1673. doi: 10.1016/j.chemosphere.2015.10.044
Manoharan, R. K., Ishaque, F., and Ahn, Y. H. (2022). Fate of antibiotic-resistant genes in wastewater environments and treatment strategies – a review. Chemosphere 298:134671. doi: 10.1016/j.chemosphere.2022.134671
Marti, E., Jofre, J., and Balcazar, J. L. (2013). Prevalence of antibiotic resistance genes and bacterial community composition in a river influenced by a wastewater treatment plant. PLoS ONE 8:e78906. doi: 10.1371/journal.pone.0078906
McConnell, M. M., Truelstrup Hansen, L., Jamieson, R. C., Neudorf, K. D., Yost, C. K., and Tong, A. (2018). Removal of antibiotic resistance genes in two tertiary level municipal wastewater treatment plants. Sci. Total Environ. 643, 292–300. doi: 10.1016/j.scitotenv.2018.06.212
McKinney, C. W., and Pruden, A. (2012). Ultraviolet disinfection of antibiotic-resistant bacteria and their antibiotic resistance genes in water and wastewater. Environ. Sci. Technol. 46, 13393–13400. doi: 10.1021/es303652q
Michael, S. G., Drigo, B., Michael-Kordatou, I., Michael, C., Jäger, T., Aleer, S. C., et al. (2022). The effect of ultrafiltration process on the fate of antibiotic-related microcontaminants, pathogenic microbes, and toxicity in urban wastewater. J. Hazard. Mater. 435:128943. doi: 10.1016/j.jhazmat.2022.128943
Munir, M., Wong, K., and Xagoraraki, I. (2011). Release of antibiotic resistant bacteria and genes in the effluent and biosolids of five wastewater utilities in Michigan. Water Res. 45, 681–693. doi: 10.1016/j.watres.2010.08.033
Ng, K. H., Samuel, L., Kathleen, M. M., Leong, S. S., and Felecia, C. (2014). Distribution and prevalence of chloramphenicol-resistance gene in Escherichia coli isolated from aquaculture and other environments. Int. Food Res. J. 21, 1321–1325.
Nzima, B., Adegoke, A. A., Ofon, U. A., Al-Dahmoshi, H. O. M., Saki, M., Ndubuisi-Nnaji, U. U., et al. (2020). Resistotyping and extended-spectrum beta-lactamase genes among Escherichia coli from wastewater treatment plants and recipient surface water for reuse in South Africa. New Microbes New Infect. 38:100803. doi: 10.1016/j.nmni.2020.100803
Oliveira, L. G., Ferreira, L. G. R., Nascimento, A. M. A., Reis, M. D. P., Dias, M. F., Lima, W. G., et al. (2018). Antibiotic resistance profile and occurrence of AmpC among Pseudomonas aeruginosa isolated from a domestic full-scale WWTP in southeast Brazil. Water Sci. Technol. 1, 108–114. doi: 10.2166/wst.2018.091
Oliveira, M., Truchado, P., Cordero-García, R., Gil, M. I., Soler, M. A., Rancaño, A., et al. (2023). Surveillance on ESBL-Escherichia coli and indicator ARG in wastewater and reclaimed water of four regions of Spain: impact of different disinfection treatments. Antibiotics. 12:400. doi: 10.3390/antibiotics12020400
Osińska, A., Korzeniewska, E., Harnisz, M., and Niestepski, S. (2020). Impact of type of wastewater treatment process on the antibiotic resistance of bacterial populations. E3S Web Conf. 17:00070. doi: 10.1051/e3sconf/20171700070
Pallares-Vega, R., Blaak, H., van der Plaats, R., de Roda Husman, A. M., Hernández-Leal, L., van Loosdrecht, M. C. M., et al. (2019). Determinants of presence and removal of antibiotic resistance genes during WWTP treatment: a cross-sectional study. Water Res. 161, 319–328. doi: 10.1016/j.watres.2019.05.100
Pazda, M., Kumirska, J., Stepnowski, P., and Mulkiewicz, E. (2019). Antibiotic resistance genes identified in wastewater treatment plant systems – a review. Sci. Total Environ. 697:134023. doi: 10.1016/j.scitotenv.2019.134023
Ping, Q., Yan, T., Wang, L., Li, Y., and Lin, Y. (2022). Insight into using a novel ultraviolet/peracetic acid combination disinfection process to simultaneously remove antibiotics and antibiotic resistance genes in wastewater: mechanism and comparison with conventional processes. Water Res. 210:118019. doi: 10.1016/j.watres.2021.118019
R Core Team (2021). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online at: https://www.R-project.org/
Raven, K. E., Ludden, C., Gouliouris, T., Blane, B., Naydenova, P., Brown, N. M., et al. (2019). Genomic surveillance of Escherichia coli in municipal wastewater treatment plants as an indicator of clinically relevant pathogens and their resistance genes. Microb. Genom. 5:e000267. doi: 10.1099/mgen.0.000267
Raza, S., Shin, H., Hur, H. G., and Unno, T. (2022). Higher abundance of core antimicrobial resistant genes in effluent from wastewater treatment plants. Water Res. 208:117882. doi: 10.1016/j.watres.2021.117882
Rizzo, L., Manaia, C., Merlin, C., Schwartz, T., Dagot, C., Ploy, M., et al. (2013). Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: a review. Sci. Total Environ. 447, 345–360. doi: 10.1016/j.scitotenv.2013.01.032
Rodriguez-Mozaz, S., Chamorro, S., Marti, E., Huerta, B., Gros, M., Sanchez-Melsio, A., et al. (2014). Occurrence of antibiotics and antibiotic resistance genes in hospital and urban wastewaters and their impact on the receiving river. Water Res. 69, 234–244. doi: 10.1016/j.watres.2014.11.021
Sanz, C., Casado, M., Martinez-Landa, L., Valhondo, C., Amalfitano, S., Di Pippo, F., et al. (2024). Efficient removal of antibiotic resistance genes and enteric bacteria from reclaimed wastewater by enhanced soil aquifer treatments. Sci. Total Environ. 953:176078. doi: 10.1016/j.scitotenv.2024.176078
Schmiege, D., Zacharias, N., Sib, E., Falkenberg, T., Moebus, S., Evers, M., et al. (2021). Prevalence of multidrug-resistant and extended-spectrum beta-lactamase-producing Escherichia coli in urban community wastewater. Sci. Total Environ. 785:147269. doi: 10.1016/j.scitotenv.2021.147269
Stachurova, T., Pikovay, A., Bartas, M., Semerad, J., Svobodova, K., and Malachova, K. (2021). Beta-lactam resistance development during the treatment processes of municipal wastewater treatment plants. Chemosphere 280:130749. doi: 10.1016/j.chemosphere.2021.130749
Tan, L., Li, L., Ashbolt, N., Wang, X., Cui, Y., Zhu, X., et al. (2018). Arctic antibiotic resistance gene contamination, a result of anthropogenic activities and natural origin. Sci. Total Environ. 621, 1176–1184. doi: 10.1016/j.scitotenv.2017.10.110
Truchado, P., Gil, M. I., Kostic, T., and Allende, A. (2016). Optimization and validation of a PMA qPCR method for Escherichia coli quantification in primary production. Food Control 62, 150–156. doi: 10.1016/j.foodcont.2015.10.014
van Bavel, B., Berrang-Ford, L., Moon, K., Gudda, F., Thornton, A. J., Robinson, R. F. S., et al. (2024). Intersections between climate change and antimicrobial resistance: a systematic scoping review. Lancet Planet Health. 8, e1118–e1128. doi: 10.1016/S2542-5196(24)00273-0
Wang, J., Chu, L., Wojnárovits, L., and Takács, E. (2020). Occurrence and fate of antibiotics, antibiotic-resistant genes (ARGs) and antibiotic-resistant bacteria (ARB) in municipal wastewater treatment plant: an overview. Sci. Total Environ. 744:140997. doi: 10.1016/j.scitotenv.2020.140997
Wang, R., Ji, M., Zhai, H., Guo, Y., and Liu, Y. (2021). Occurrence of antibiotics and antibiotic resistance genes in WWTP effluent-receiving water bodies and reclaimed wastewater treatment plants. Sci. Total Environ. 796:148919. doi: 10.1016/j.scitotenv.2021.148919
World Health Organization (WHO) (2018). WHO REPORT on Surveillance of Antibiotic Consumption: 2016–2018 Early Implementation. Geneva: WHO.
Xie, H., Yamada, K., Tamai, S., Shimamoto, H., Nukazawa, K., and Suzuki, Y. (2023). Disappearance and prevalence of extended-spectrum β-lactamase-producing Escherichia coli and other coliforms in the wastewater treatment process. Environ. Sci. Pollut. Res. Int. 30, 83950–83960. doi: 10.1007/s11356-023-28382-3
Yang, L., Wen, Q., Chen, Z., Duan, R., and Yang, P. (2019). Impacts of advanced treatment processes on elimination of antibiotic resistance genes in a municipal wastewater treatment plant. Front. Environ. Sci. Eng. 13:32. doi: 10.1007/s11783-019-1116-5
Zhang, J., Li, G., Yuan, X., Li, P., Yu, Y., Yang, W., et al. (2023). Reduction of ultrafiltration membrane fouling by the pretreatment removal of emerging pollutants: a review. Membranes 13:77. doi: 10.3390/membranes13010077
Keywords: wastewater treatment, sustainable agriculture, ESBL-E. coli, antimicrobial resistance (AMR), irrigation water
Citation: Truchado P, Oliveira M, Cordero-García R, Abellán Soler M, Rancaño A, García F, Álvarez-Ordóñez A and Allende A (2025) Sustainable water reuse in food production: risks of extended-spectrum β-lactamase (ESBL)-producing Escherichia coli and antimicrobial resistance gene release from tertiary-treated reclaimed water. Front. Microbiol. 16:1591202. doi: 10.3389/fmicb.2025.1591202
Received: 10 March 2025; Accepted: 15 May 2025;
Published: 19 June 2025.
Edited by:
Alberto Garre, Polytechnic University of Cartagena, SpainReviewed by:
Pedro Rodríguez-López, Centre de Recerca en Sanitat Animal (CReSA), SpainManuela Macrì, University of Turin, Italy
Copyright © 2025 Truchado, Oliveira, Cordero-García, Abellán Soler, Rancaño, García, Álvarez-Ordóñez and Allende. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Allende, YWFsbGVuZGVAY2ViYXMuY3NpYy5lcw==