 Bruna Fuga1,2,3,4*†
Bruna Fuga1,2,3,4*† Ingrith Neves1,2†
Ingrith Neves1,2† Herrison Fontana1,2Jessica Bispo2,3
Herrison Fontana1,2Jessica Bispo2,3 Elder Sano2,3
Elder Sano2,3 Adriana Cardenas-Arias2,3Fernanda Esposito1,2Brenda Cardoso2,3
Adriana Cardenas-Arias2,3Fernanda Esposito1,2Brenda Cardoso2,3 Susan Ienne5
Susan Ienne5 Fábio P. Sellera2,6,7
Fábio P. Sellera2,6,7 Nilton Lincopan1,2,3*
Nilton Lincopan1,2,3*- 1Department of Clinical Analysis, Faculty of Pharmaceutical Sciences, University of São Paulo, São Paulo, Brazil
- 2One Health Brazilian Resistance Project (OneBr), São Paulo, Brazil
- 3Department of Microbiology, Institute of Biomedical Sciences, University of São Paulo, São Paulo, Brazil
- 4Department of Cell Biology, Institute of Biological Sciences, University of Brasília, Brasília, Brazil
- 5Core Facility for Scientific Research (CEFAP), Institute of Biomedical Sciences, University of São Paulo, São Paulo, Brazil
- 6Department of Internal Medicine, School of Veterinary Medicine and Animal Science, University of São Paulo, São Paulo, Brazil
- 7School of Veterinary Medicine, Metropolitan University of Santos, Santos, Brazil
Background: Contamination of food by antimicrobial-resistant pathogens poses significant risk to consumers and environment, potentially leading to foodborne illnesses, silent colonization, and dissemination of antibiotic-resistant bacteria across geographic borders.
Methods: This study analyzed 32 retail meat samples (12 chicken, 10 beef, and 10 pork) from conventional (CN) and antibiotic-free (AF) production systems in Brazil, assessing WHO bacterial priority pathogens through whole genome sequencing and microbiological methods.
Results: Five broad-spectrum cephalosporin-resistant Escherichia coli strains were isolated from AF chicken meat, including four extended-spectrum β-lactamase (ESβL) producers belonging to sequence types (STs) ST117, ST443, ST1559 and ST3258, and one pAmpC producer carrying the blaCMY-2 gene and belonging to ST57. On the other hand, four E. coli strains resistant to 3rd generation cephalosporins were identified in CN chicken meat, being three ESβL producers of ST38, ST2179 and ST2040, and one pAmpC producer belonging to ST350. Genes conferring resistance to hazardous heavy metals, disinfectants, and pesticides were identified, whereas virulent potential of E. coli ST350 and ST2040 was predicted. Noteworthy, E. coli ST38 was genomically related to lineages previously identified in poultry (North America) and polluted environments (Europe), supporting an intercontinental dissemination within a One Health framework.
Conclusion: Our findings reinforce the need for continuous surveillance of WHO critical priority pathogens in the chicken meat supply chain from different production systems.
1 Introduction
Critical antimicrobial-resistant (AMR) bacteria exert hazardous effects on the environment or humans via contamination, causing serious economic losses and endangering human and environmental health (Antimicrobial Resistance Collaborators, 2022). Multisectoral aspects, including the overuse and misuse of antimicrobials in human and animal healthcare, agriculture, and livestock, have significantly contributed to the rapid emergence and spread of multidrug-resistant (MDR) bacterial strains (Aslam et al., 2021). Indeed, MDR bacteria have increasingly been documented beyond the confines of human hospital walls, adding an additional layer of complexity to this issue (Aslam et al., 2021; McEwen and Collignon, 2018). Given the interconnected nature of this problem, multidisciplinary approaches under the auspices of One Health have been encouraged to strengthen surveillance and mitigate the dissemination of these clinically important pathogens (Jesudason, 2023).
The 2021–2025 Action Plan on antimicrobial resistance by the Food and Agriculture Organization of the United Nations (FAO) has highlighted five key strategic priorities aimed at addressing this issue within the food and agriculture sectors, including: (i) promoting practices to prevent infections and mitigate the spread of antimicrobial resistance; and (ii) ensuring the prudent use of antimicrobials to preserve their efficacy. Among these priorities, surveillance is the basis for understanding the dynamics of antibiotic resistance in food, enabling the detection of emerging threats, and guiding targeted interventions to mitigate the spread of resistant microorganisms (Keck et al., 2023).
Particularly concerning is the widespread dissemination of third-generation cephalosporin-resistant Escherichia coli producing extended-spectrum (ESβLs) and/or plasmid-mediated AmpC (pAmpC) β-lactamases, rendering them clinically ineffective (Kaper et al., 2004; Foster-Nyarko and Pallen, 2022; Fuga et al., 2022). Due to its high mortality rates, healthcare burden, prevalence of resistance, and other significant impacts on public health, this sort of E. coli strains have been classified as a critical priority pathogen by the World Health Organization (2024). Therefore, contamination of food by critical priority E. coli represents serious a public health concern due to its potential transmission to humans through the food supply chain (Ramos et al., 2020; Alegría et al., 2020; Mughini-Gras et al., 2019). In this regard, chicken meat is a food highly susceptible to contamination by various microorganisms throughout the food chain, leading to its spoilage and risk to human health and the environment.
Currently, the production of antibiotic-free chicken meat has been encouraged as a response to growing concerns about AMR and the potential human health risks associated with the consumption of meat products containing antimicrobial residues (Haque et al., 2020; Mohammadi et al., 2023). AMR surveillance on antibiotic-free (AF) meat has been proposed to evaluate the effectiveness of antimicrobial stewardship practices in reducing the dissemination of antimicrobial-resistant bacteria in the food supply chain (Farooq et al., 2022). Although some studies have demonstrated that AF meat may present lower levels of antimicrobial residues compared to conventionally raised meat (Sarkar et al., 2023), it can still harbor bacteria of clinical interest, including critical priority E. coli strains (Rawat et al., 2024).
Since animal-derived foods can be contaminated with a wide variety of hazardous bacteria, such as E. coli, the identification and genetic context of pathogenicity and antibiotic resistance is very important for prevention against their widespread, especially MDR strains. In this study, as part of the Grand Challenges Explorations: New Approaches to Characterize the Global Burden of Antimicrobial Resistance Program, we report the occurrence of global clones of WHO critical priority E. coli in both conventional (CN) and AF retail meat, in Brazil, one of the world’s leading meat producers and exporters, and a significant player in the global meat industry (Klein and Vidal, 2022).
2 Materials and methods
2.1 Sample collection
Between August and February 2019–2020, 32 different samples of chicken, bovine and swine meat sold in supermarkets located at all regions of São Paulo, the most populous city in Latin America, were aseptically collected. The meat samples were obtained from CN or AF production systems, and include retail chicken (n = 12), beef (n = 10) and pork meat (n = 10) (Supplementary Table S1). The storage methods of meat samples, both fresh (FS) and frozen (FZ), were also assessed.
2.2 Isolation of broad-spectrum cephalosporin resistant Escherichia coli from conventional and antibiotic-free labeled commercial meat
Samples were purchased and immediately stored in thermic boxes and processed within 4 h. Microbiological analyses were carried out according to the Food and Drug Administration (FDA) protocol with modifications (U.S. Food and Drug Administration, 2021). In brief, 100 g of each sample were rinsed with 225 mL of Buffered Peptone Water (BPW) in sterile plastic bags (Whirl-Pak; Nasco, WI), and homogenized by hand-massage for 15 min. Subsequently, 25 mL was transferred to 25 mL of MacConkey broth flasks and cultured overnight at 37 ̊C. Then, 10 μL aliquot were transferred to MacConkey agar plates supplemented with ceftriaxone (2 μg/mL), as previously recommended for screening of potential ESβL-producing E. coli (Jacob et al., 2020).
After overnight incubation at 37°C, presumptive E. coli typical colonies were subculture into eosin methylene blue (EMB) agar and further confirmed by matrix-assisted laser desorption ionization–time of flight mass spectrometry (MALDI-TOF MS) analysis (Singhal et al., 2015).
2.3 Antimicrobial susceptibility testing and confirmation of extended Spectrum β-lactamases (ESβL) phenotype
Antimicrobial susceptibility profiles were accessed by disc diffusion method and results interpreted according to the Clinical and Laboratory Standards Institute (2024) breakpoints. A panel of 14 antimicrobials was tested, including amoxicillin/clavulanic acid (AMC), ceftriaxone (CRO), ceftazidime (CAZ), cefotaxime (CTX), cefoxitin (CFO), cefepime (CPM), ertapenem (ETP), imipenem (IMP), meropenem (MER), nalidixic acid (NAL), ciprofloxacin (CIP), aztreonam (ATM), gentamicin (GEN), and amikacin (AMI). Additionally, double disc synergy test (DDST) was used to detect ESβL production (Jarlier et al., 1998; Drieux et al., 2008). Isolates classified as non-susceptible to at least one agent in three or more antimicrobial categories were defined as MDR, according to the criteria established by Magiorakos et al. (2012).
2.4 Genome sequencing
Of the 14 strains recovered from the collected meat samples, nine E. coli isolates from chicken meat, representing both CN and AF systems, were selected for whole genome sequencing mainly based on their ESβL phenotype, as determined by the DDST test, with additional consideration of their MDR profiles (Table 1).
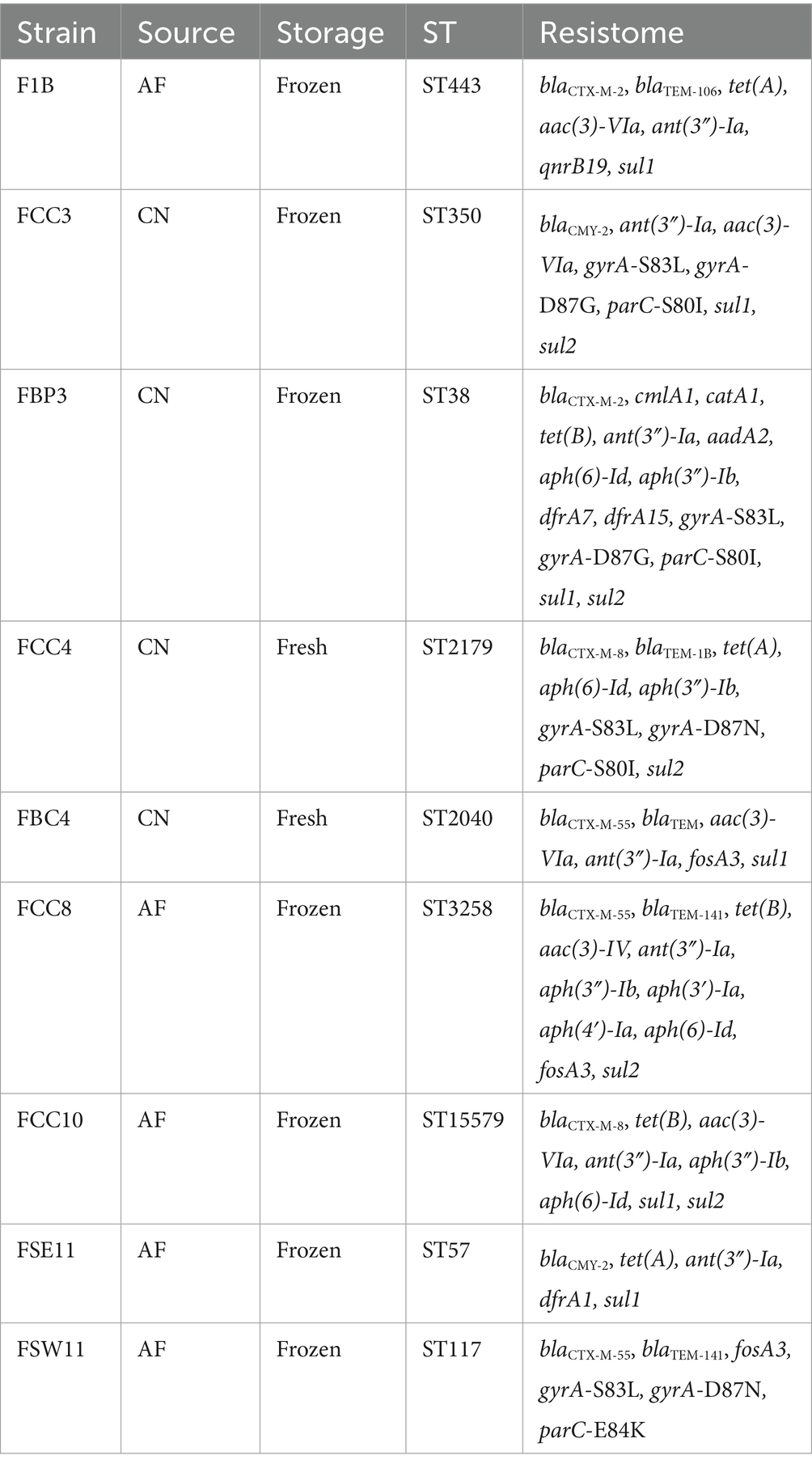
Table 1. Characteristics of sequenced hazardous Escherichia coli strains isolated from antibiotic-free (AF) and conventional (CN) chicken meat.
Total DNA of E. coli strains was extracted using a PureLink quick gel extraction kit (Life Technologies, CA), and subsequently utilized for constructing a Nextera DNA Flex Library Prep (Illumina Inc., San Diego, CA). Genomic sequencing was performed using the NextSeq platform (Illumina, San Diego, CA).
2.5 Bioinformatics analysis
The paired-end reads were quality checked and trimmed (PHRED quality score <20) using TrimGalore v.0.6.71 and assembled by Unicycler v.0.5.02. For the E. coli strain FCC4, we conducted both trimming and de novo assembly of sequences using CLC Genomics Workbench v.12.0.3 (Qiagen, Hilden, Germany). Complete genome annotation was carried out using NCBI Prokaryotic Genome Annotation Pipeline v.3.23.
Public databases for molecular typing and microbial genome diversity – PubMLST4 and Enterobase5 was used to determine sequence type of E. coli strains. To evaluate antimicrobial resistance genes, we employed the Resfinder v.4.4.2 tool available through the Center for Genomic Epidemiology (CGE) pipeline6, with default settings.
ABRicate v0.9.87 was used to predict virulence genes (VirulenceFinder v.2.0), plasmids (PlasmidFinder v.2.1), fimbriae (FimTyper v.1.0), and serotype (SerotypeFinder v.2.0) profiles through the CGE database. The Virulence Factor Database - VFDB8 was additionally employed for predicting the virulome. Heavy metal (HM), herbicide (glyphosate), and disinfectants (QACs) resistance genes were also identified using ABRicate through a database constructed from NCBI and BacMet29. Cutoff values of ≥90% identity and ≥ 80% coverage were used. Phylogroup stratification was performed using ClermonTypingv.1.4.010. The Mlplasmids v2.1.011 was used to predict plasmid and chromosome-derived sequences.
2.6 Phylogenetic analysis
For phylogenetic purposes, we downloaded all genome assemblies with data for country, year of collection and source of isolation for each of the STs from the Escherichia/Shigella database in Enterobase (see text footnote 5). For each ST, the average nucleotide identity (ANI) between E. coli strains and the downloaded dataset was obtained using FastANI v1.3212, and the 30 genomes with highest ANI were select for phylogenetic analysis. ST3258 had only 18 genome assemblies available, so all genomes were used. CSI Phylogeny v1.413 was used with default settings to generate maximum-likelihood trees based on SNP alignment. As reference genomes, chromosome sequences of E. coli ST38 strain 144 (accession number NZ_CP023364.1), ST57 strain NCTC10444 (NZ_LR134092.1), ST117 strain 14EC020 (NZ_CP024138.1), ST350 strain NCTC9112 (NZ_LR134079.1), ST443 strain 2014C-3307 (NZ_CP027368.1), and ST2179 strain BR03-DEC (NZ_CP035321.1) were used. On the other hand, for ST2040 and ST3258, which had no complete genome assemblies available on Enterobase, we used as reference genomes the chromosome sequences of ST191 (closely related to ST2040) strain 1,500 (NZ_CP040269.1) and ST117 (closely related to ST3258) strain 14EC020 (NZ_CP024138.1). Finally, for FCC10, we used the closely related ST641 strain 1916D18 (NZ_CP046000.1). All reference genomes were chosen based on Enterobase search results. All genome assemblies used on phylogenetic analysis were screened for resistance genes and plasmid replicons using ABRicate v1.0.1 (see text footnote 7) with ResFinder and PlasmidFinder databases. iTOL v614 was used to root the trees at midpoint and to annotate the trees with Enterobase and ABRicate data.
3 Results
A total of 14 E. coli isolates were recovered from 32 collected meat samples, mostly from chicken (12/14), including from both CN and AF production systems, followed by swine (2/14) samples obtained from CN alone (Supplementary Table S1). No E. coli strains were isolated from beef. Based on the sources of the meat samples, a higher frequency of E. coli isolates was obtained from chicken breast filet. A greater number of resistant E. coli strains were isolated from AF compared to the CN meat (6/9, and 6/23, respectively). Regardless of the storage method of meat samples, antimicrobial-resistant E. coli was mainly detected in FZ meat (6/8).
Antimicrobial susceptibility testing of the 14 E. coli isolates is shown in Figure 1. The highest resistance frequency was found for ceftriaxone and cefotaxime (100% of the tested strains), followed by cefepime, nalidixic acid and gentamicin (50%). The seven E. coli strains (F1B, FCC3, FBP3, FBC4, FCC10, FSE11, and FCC2) were classified as MDR according to the code previously established by Magiorakos et al. (2012). The MDR strains were more prevalent in chicken meat from antibiotic-free production systems (4/6) compared to conventional systems (3/8) (Figure 1). Among the tested strains, these nine strain, F1B, FCC3, FBP3, FCC4, FBC4, FCC8, FCC10, FSE11, and FSW11, were selected for whole genome sequencing based on their susceptibility profile and ESβL phenotype (Figure 1). We chose only chicken meat strains because they represented most of the isolates and allowed for a more consistent comparison between antibiotic-free and conventional production systems within the same meat type.
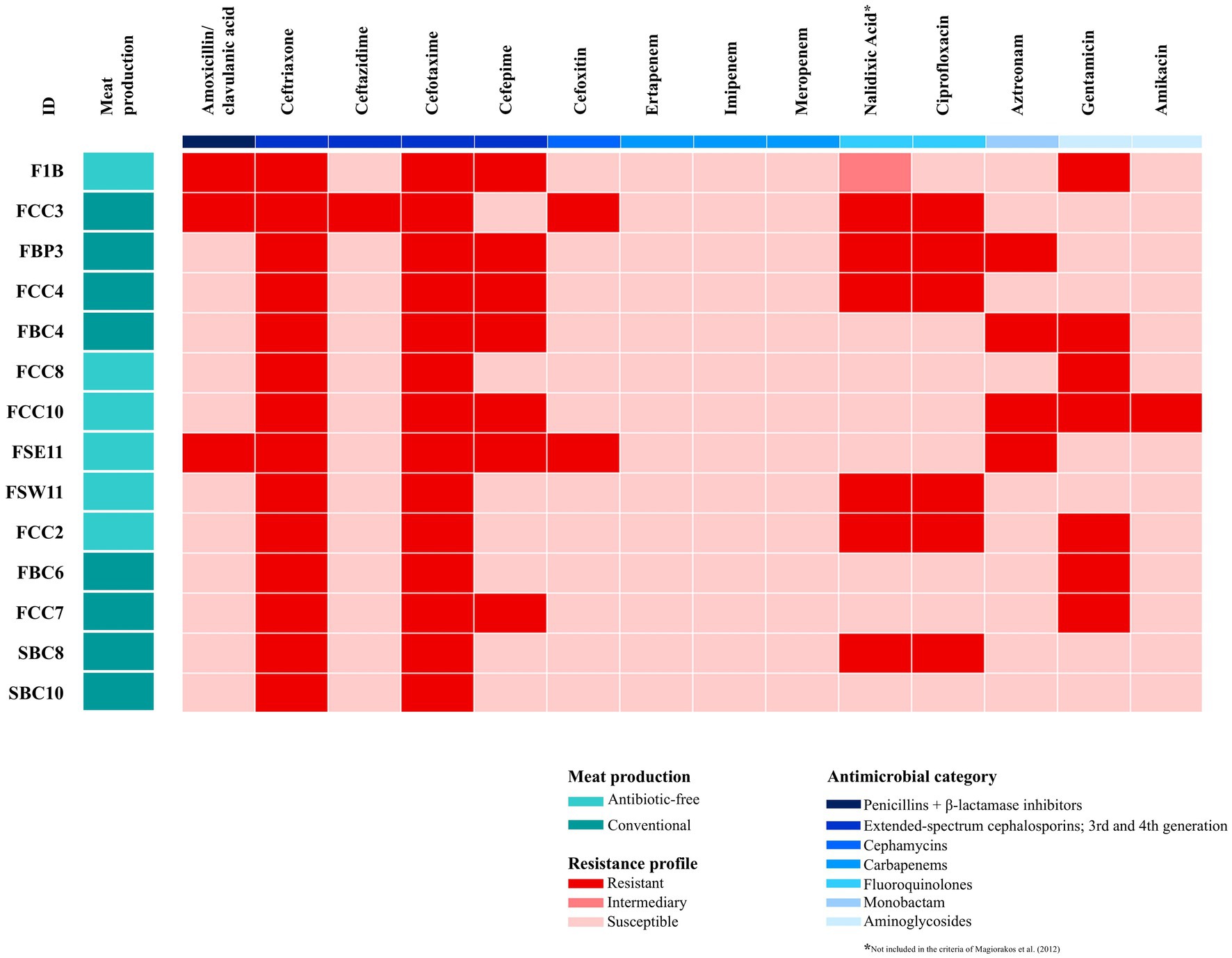
Figure 1. Heatmap showing the antimicrobial resistance profile of 14 Escherichia coli strains recovered from conventional and antibiotic-free retail meat in Brazil. Boxes highlighted in red indicate the antimicrobials for which the strains displayed resistance, while boxes in purple indicate susceptibility profiles. Aqua green color denotes antibiotic-free production systems, and dark green represents conventional systems.
Initial analysis of the sequenced strains was performed using an in silico MLST approach that revealed completely distinct sequence types (STs) among all strains (ST38, ST57, ST117, ST350, ST443, ST15579, ST2179, ST2040, and ST3258).
Overall, 63 antimicrobial resistance genes (ARGs) conferring resistance to eight antimicrobials categories, including β-lactams, phenicols, tetracyclines, aminoglycosides, fosfomycin, trimethoprim, quinolones, and sulfonamides were identified based on the WGS analysis (Figure 2A). Each genome of the nine strains harbored between three and 12 ARGs. The ß-lactam resistance genes blaCMY-2 or blaCTX-M (blaCTX-M-2, blaCTX-M-8, or blaCTX-M-55 variants) were detected in all sequenced E. coli strains, regardless of whether from CN or AF production systems (Table 1). Among these, the blaCTX-M-55 gene variant was the most frequently found in E. coli strains, being identified in three strains (FBC4/CN, FCC8/AF, and FSEW11/AF).

Figure 2. ESβL/pAmpC-producing Escherichia coli isolated from retail meat, considering the types of production (antibiotic-free or conventional) and storage (cold or frozen) system. (A) Antibiotic resistance, mutations and plasmidome diversity. (B) Metals, biocides and pesticide resistance. Highlighted boxes indicate the presence, while gray boxes indicate the absence of resistance determinants (to antimicrobials, metals, biocides, and pesticides), mutations, or plasmid incompatibility groups.
Resistance to fluoroquinolones was mainly associated with chromosomal mutations on gyrA [S83L, D87N, and D87G] and parC [S80I and E84K] genes. Only the F1B/AF E. coli strain presented gene associated with plasmid mobilization (qnrB19).
The sequenced strains showed a broad resistome related to heavy metals, herbicide, and disinfectants resistance genes, ranging between 28 and 39 resistance genes in each strain (Figure 2B). Overall, all of them carried resistance genes to heavy metal (arsenic: arsB, arsC, and arsR genes), herbicide (glyphosate: phnCDEFGHIJKLMNOP genes), and disinfectants (acrE, acrF, emrD, emrK, mdtE, mdtF, mdtK, mdtN, and tolC genes).
Our results also identify 12 distinct plasmid replicon types among the 9 genomes analyzed, including IncFIA, IncFIB, IncFIC, IncFII, IncI1-I, IncN, IncY, IncB/O/K/Z, IncX1, IncX4, p0111, and Col (Figure 2A). The most prevalent plasmid replicons were Col (88.9%, 8/9) and IncFIB (77.8%, 7/9). Interestingly, the E. coli strains harboring the highest number of resistance genes, FBP3/ST38/CN (12 ARGs) and FCC8/ST3258/AF (11 ARGs), commonly exhibited the plasmids IncFIB, IncFIC, and Col. Additionally, FCC8 strain also possessed IncFII and IncN plasmids. Although the short-read methodology employed did not allow for the circularization of plasmids, using the mlplasmid tool, we detected plasmid-derived sequences carrying the blaCTX-M-type or blaCMY-2 genes in F1B, FCC3, FBP3, and FSE11 strains. As a limitation, the study did not include plasmid electrophoretic analysis.
In total, 171 virulence genes representing different virulence pathogenicity mechanisms (adherence, bacteriocins, iron uptake, toxins, invasion, secretion systems, protectins/serum resistance, and other factors) were identified (Figure 3). The E. coli FBC4/ST2040 and FCC3/ST350 strains, from CN production system, harbored a higher number of virulence genes (124 and 120, respectively), followed by the FCC4/ST2179 (119 genes) and FBP3/ST38 (110 genes) strains. Despite this, those isolates from AF meat, FCC8/ST3258 and FSW11/ST117, also presented a broad virulome (104 and 100, respectively). All sequenced strains presented genes involved in processes of adherence (fimBCDEFGHI, ecpAR, cgsABCDEFG, and yehBCD genes), iron uptake (entABCDEFS, fepABCDEG, and fes genes), invasion (ibeBC genes), secretion system (espL1 gene), proctin/ serum resistance (ompT gene), among others (nlpI, and terC genes). Detailed genomic information of virulome is shown in Supplementary Table S1.
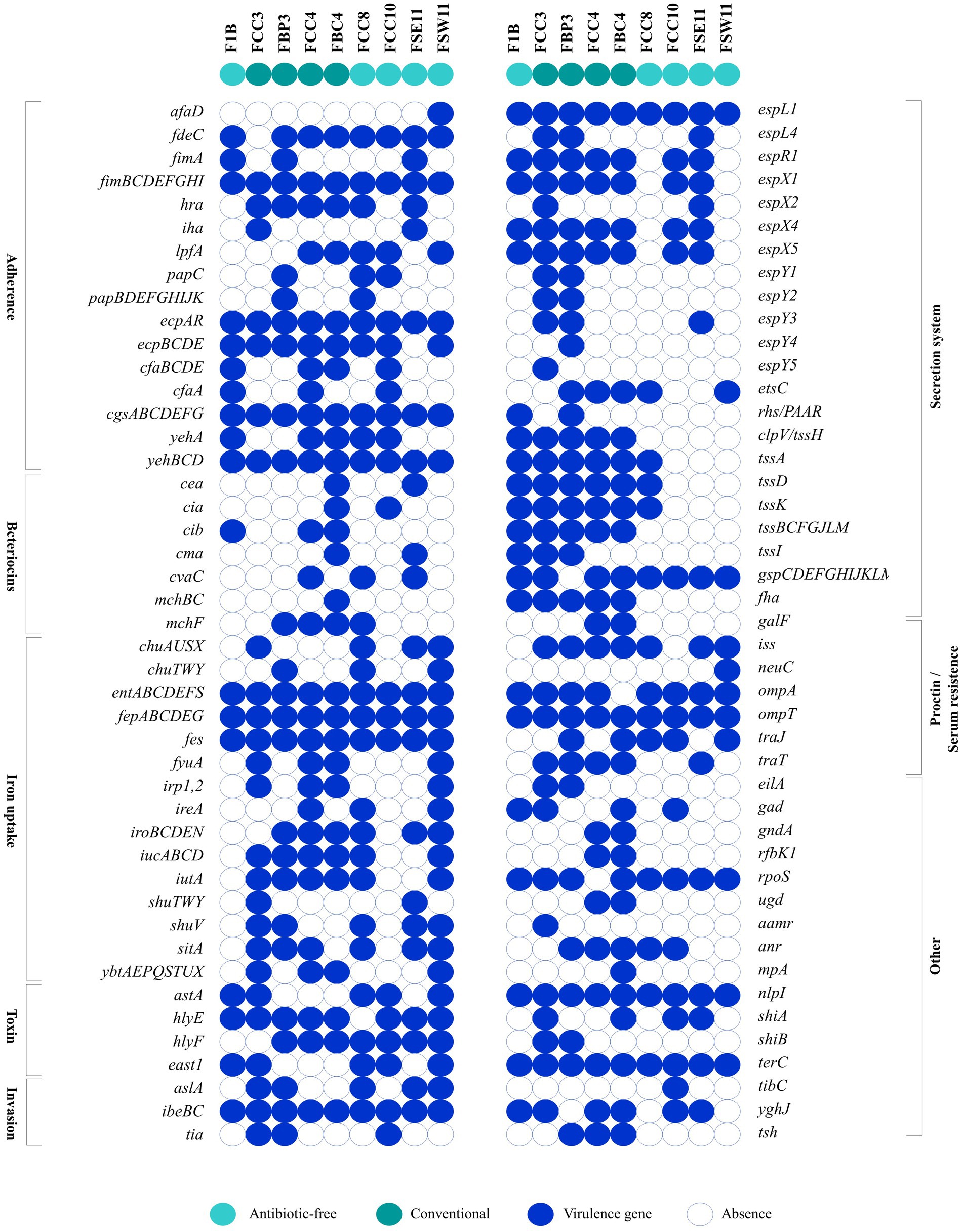
Figure 3. Comparative panel of virulence genes detected among antibiotic-free and conventional ESβL/pAmpC-producing Escherichia coli strains. The virulome scenario involves genes related to adherence, bacteriocins, iron uptake, toxins, invasion, secretion systems, protectins/serum resistance, and other factors of pathogenicity. Blue boxes represent the presence of virulence genes, while white boxes indicate their absence. Aqua green color denotes antibiotic-free production systems, and dark green represents conventional systems.
Fimbriae and serotyping analysis, accessed by FimTyper and SerotypeFinder, respectively, showed that all strains were distinct, although strains FCC3/CN, FCC8/AF and FSW11/AF presented the same H antigen (H4) (Supplementary Table S1). The most frequent Clermont phylogroup was B1 type (3/9), followed by E type (2/9) (Supplementary Table S1).
The phylogenetic analysis was conducted based on sequence typing (ST), so only one isolate from our study (FBP3) was included in the analysis of the pandemic clone ST38, together with other related isolates. Notably, the FBP3/CN/FZ strain was closely related to chicken meat isolates from Brazil in 2019, and to poultry isolates from USA in 2007, with SNP differences ranging from 128 to 142, respectively (Figure 4; Supplementary Table S1). All Brazilian strains carried blaCTX-M-2, sul1, sul2, aph(3″)-Ib, aph(6)-Id and drfA17 genes. The ST117 FSW11 (AF/FZ) strain showed close phylogenetic relatedness to other CTX-M-55-producing E. coli isolates from Brazil (2019), specifically those derived from swine meat (15 SNP differences) and poultry (17 SNP differences) (Figure 5; Supplementary Table S1). Additionally, the FBP3 and FSW11 strains were genomically linked (995–1843 SNPs) to the ST38 and ST117 lineages previously identified in environmental contamination in Europe (Figures 4, 5; Supplementary Table S1). From a plasmidome perspective, all ST117 strains carried the IncFIB, IncFIC, IncFII, and IncN plasmid groups (Figure 5). The presence of these plasmids in all strains suggests a high risk of antibiotic resistance spread, as these plasmid groups are known for efficient horizontal gene transfer and carriage of resistance genes (Liu et al., 2024; Felix et al., 2024; Yu et al., 2024).
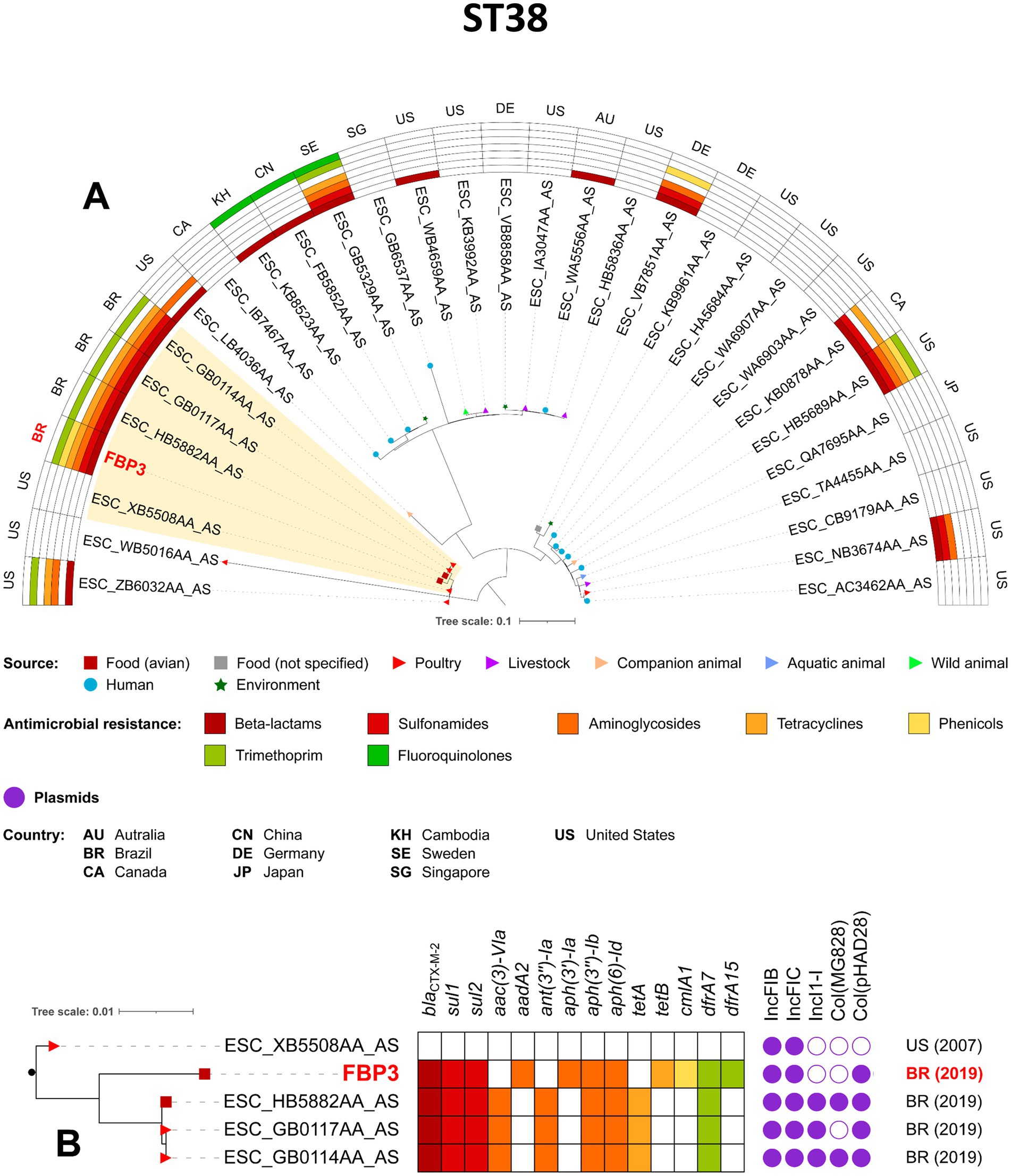
Figure 4. Phylogenomic analysis of FBP3 Escherichia coli strain belonging to ST38. In A, maximum-likelihood phylogenetic tree illustrating of 31 Escherichia coli ST38. Comparison of resistomes, isolation sources, and countries of origin of the strains. In B, zooming into the subtree comprising the clade A. The figure was generated with iTOL version 5.6.1 (https://itol.embl.de).
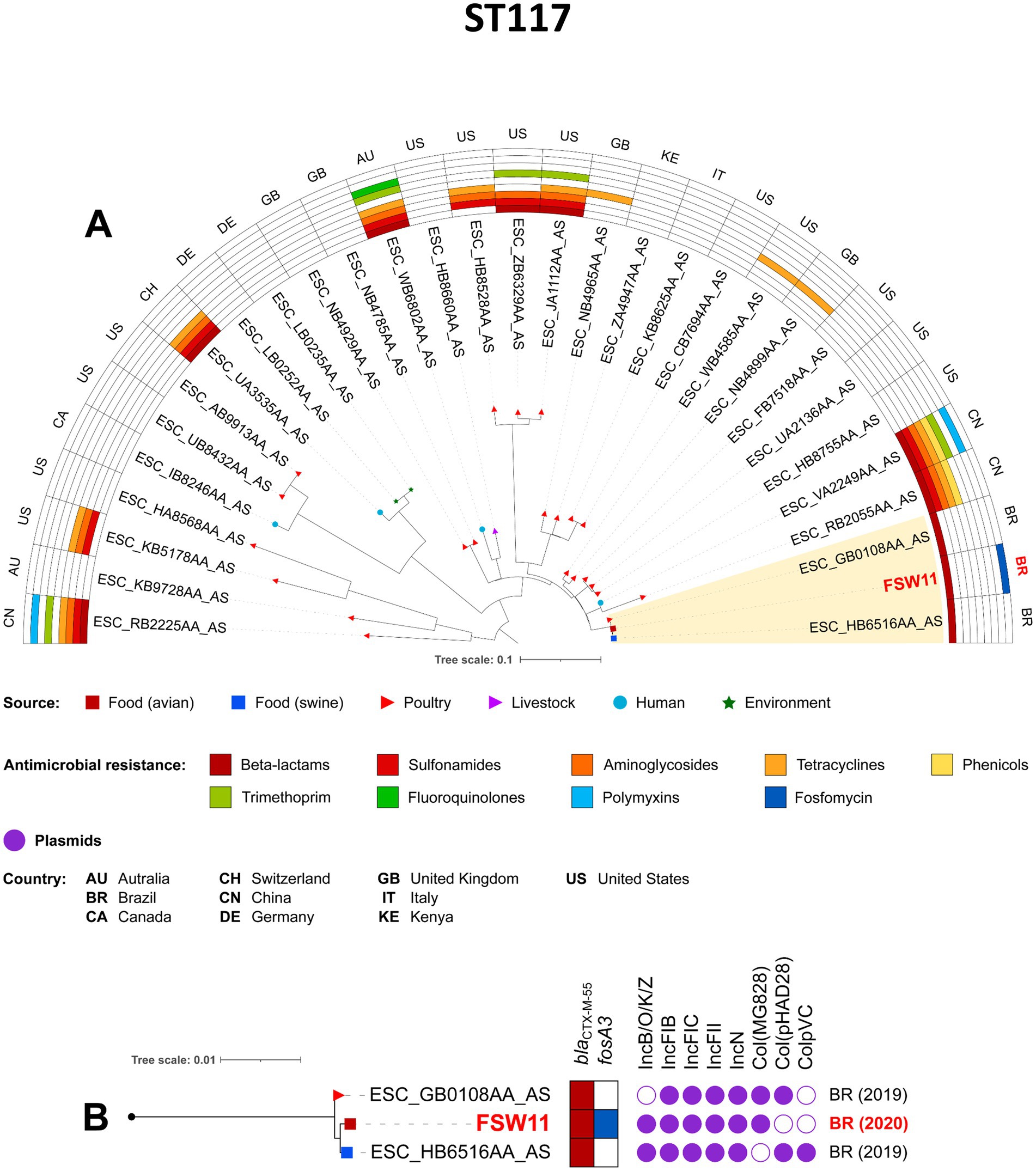
Figure 5. Phylogenomic analysis of FSW11 Escherichia coli strain belonging to ST117. In A, maximum-likelihood phylogenomic tree of 31 E. coli ST117. Comparison of resistomes, isolation sources, and countries of origin of the strains. In B, zooming into the subtree comprising the clade A. The figure was generated with iTOL version 5.6.1 (https://itol.embl.de).
In general, the F1B, FCC8, FBC4, FSE11 strains were nested within a food (avian), poultry and/or livestock clade (Supplementary Figure S1). On the other hand, the phylogenetic analysis of FCC4, FCC3, and FCC10 strains identified a clade comprising isolates from diverse sources, including avian food products, poultry, livestock, humans, and/or wild animals (Supplementary Figures S1B3,B5,B6). Furthermore, it was observed that some strains (FBP3, FCC4, and FBC4) share similar antibiotic resistance and plasmid content with their closest phylogenetic relatives, indicating the possible influence of horizontal gene transfer and environmental factors.
4 Discussion
Bidirectional transmission of antibiotic resistance is likely between humans, the food chain, and the environment. In this regard, industrial chicken farms represent an ideal environment for the spread of AMR bacteria, since the overuse of antimicrobials contributes to the selection of antibiotic-resistant bacteria in the gastrointestinal microbiota of animals, whereas the consumption of contaminated animal-source food, or environmental exposure can lead to the transmission of antimicrobial-resistant bacteria to humans and non-human hosts. In fact, antimicrobial resistance is regularly manifested in human clinical settings through food chain, with additional environmental spillovers.
This study prioritized the isolation and specific characterization of prevalent critical pathogens, following the FAO Action Plan and WHO’s global research guidelines on antimicrobial resistance (Keck et al., 2023; Bertagnolio et al., 2024). Therefore, as a limitation of this study, bacterial load was not measured before overnight enrichment, and virulence gene expression was not quantified. We report the occurrence of international clones of ESβL/pAmpC-positive E. coli isolated from both CN and AF retail meat sold at supermarkets across all regions of São Paulo, the most populous city in Latin America with over 11 million inhabitants (IBGE, 2025). This metropolitan area represents a diverse consumer population and a significant market for retail meat, making it a relevant setting for studying antimicrobial-resistant pathogens in food.
The dissemination of genes encoding ESβL in meat requires closer attention, particularly due to its association with the successful expansion of international high-risk clones (Fuga et al., 2022; Soncini et al., 2022). In this report, we highlight the occurrence of CTX-M-producing E. coli strains belonging to diverse lineages, including ST38, ST57, ST117, ST350, ST443, ST15579, ST2179, ST2040, and ST3258. While CTX-M-producing clones belonging to ST350 have already been widely reported in chicken samples from southwest China (Li et al., 2022), our phylogeographical analysis further confirmed the presence of ST350 clone in poultry samples from the USA, as well as its clonal relationship with the E. coli FCC3 strain identified in our study. Strikingly, E. coli ST350 has been reported since 1979 (accession number AASASW000000000.1), highlighting a long-standing circulation and persistence.
The E. coli ST3258 present in organic broiler farm has been detected in Netherlands (van Hoek et al., 2018) and ST2040 and has been mainly reported to harboring the blaCMY-2 gene (Castellanos et al., 2017; Che et al., 2023). The other STs (ST57, ST443, and ST2179) have been reported in Brazil associated with blaCTX-M-type or blaCMY-2 genes, being recovered from different hosts and sources (Fuga et al., 2022; Leigue et al., 2015; Palmeira et al., 2018; Dos Anjos Adur et al., 2022). To our knowledge, this is the first report of an E. coli ST15579 strain. The presence of closely related ST15579-FCC10, ST2179-FCC4, and ST350-FCC3 strains across avian, livestock, human, and/or wild animal hosts underscores potential transmission pathways at the human-animal-environment interface, emphasizing the importance of a One Health approach to antimicrobial resistance surveillance.
Worryingly, we identified the presence of high-risk lineages ST38 and ST117 in chicken meat. These E. coli clones have been increasingly identified in various sources including food, animals, humans, and the environment, emphasizing their high versatility at the One Health interface (Fuga et al., 2022; Soncini et al., 2022; Berg et al., 2017; Mo et al., 2023). Critically, these strains were recovered from chicken meat of AF production systems, which might suggest contamination during the food chain process or storage. In this regard, the AF meat production has been proposed to minimize the use of antimicrobials as prophylactic agents and growth promoters (World Health Organization, 2017; Food and Agriculture Organization of the United Nations, 2019; Tang et al., 2019; Murray et al., 2021). Indeed, there is global pressure to reduce the use of antimicrobials in chicken and swine production, as it raises concerns about the development of antimicrobial resistance (AMR) and its potential impact on public health (World Health Organization, 2017; Food and Agriculture Organization of the United Nations, 2019).
In the last years, the pandemic antimicrobial-resistant E. coli clone ST38 has been reported as a common environmental bacterial contaminant disseminated through hospital sewage in Norway, municipal wastewater in Croatia, wastewater treatment plants and rivers in China and Tunisia, hospital and community wastewater in Czech Republic, surface waters and sewage in Ireland, estuaries in Lebanon, and river water in Algeria (Grevskott et al., 2024; Puljko et al., 2024; Li et al., 2023; Davidova-Gerzova et al., 2023; Hooban et al., 2021; Hassen et al., 2021; Diab et al., 2018; Seni et al., 2018; Tafoukt et al., 2017; Oikarainen et al., 2019); serving, indeed, as an environmental sentinel for AMR. Strikingly, E. coli ST38, have been found to intersect variably across the human-animal-environment interface in Switzerland and Brazil (Fuga et al., 2022; Müller et al., 2016). On the other hand, E. coli ST117, one of the extended-spectrum β-lactamase (ESβL)-producing clone that we isolated in this study, has been previously found in chicken meat in Spain, imported Brazilian poultry meat, and human extraintestinal disease, presenting a risk to humans ingesting poultry products (Martínez-Álvarez et al., 2025; Saidenberg et al., 2024; da Silva et al., 2022; Casella et al., 2018).
Remarkably, our study revealed a higher number of antimicrobial-resistant E. coli strains recovered from meat sourced from AF farming systems.
The presence of antibiotic-resistant bacteria in antibiotic-free meat may seem contradictory and does not always mean antibiotics were used in poultry production (Davis et al., 2018; Rawat et al., 2024). In this regard, several factors could explain this phenomenon, including: (i) imported chicks or feed additives containing antibiotic-resistant bacteria; (ii) horizontal transmission of antibiotic-resistant bacteria from other animals, farm workers, or contaminated surfaces; (iii) soil, water, and/or feed contaminated with resistant bacteria, from neighboring farms or past practices (as resistant bacteria are known to persist in the environment and can colonize animals raised without antibiotics); and, (iv) unintentional cross-contamination during processing or transport (Adegbeye et al., 2024; Coppola et al., 2022; Tian et al., 2021; Sun et al., 2024; Thanner et al., 2016; Argudín et al., 2017; Park et al., 2017; De Cesare et al., 2022; Millman et al., 2013). In slaughterhouses that process animals from multiple sources, cross-contamination can occur if facilities are not properly cleaned, where tools, conveyor belts for poultry processing plant, and handlers can spread bacteria from one carcass to another (Warriner et al., 2002; Bergšpica et al., 2020; Park et al., 2017; De Cesare et al., 2022). While poor hygiene, inadequate disinfection, or substandard biosecurity practices on antibiotic-free farms can allow resistant bacteria to thrive or spread, resistance genes can persist in microbial communities for extended periods (Jaleta et al., 2024; Johnsen et al., 2009).
On the other hand, the meaning of antibiotic-free products can cause some confusion, mainly because there is no official or international accepted definition of what it is and how to classify different types of antibiotic-free products. Additionally, complications come with anticoccidials for prevention of coccidiosis in poultry, since in some countries they are classified as antimicrobials and as such they must be withdrawn from antibiotic-free production. In some countries, labels on retail poultry have been a source of misunderstanding to the consumer. Meat from chickens raised with sub-therapeutic doses of antibiotics may still contain labels claiming, “all natural” or “free range,” which imply a healthier product even though both statements are silent on antibiotics use. For retail poultry meat, the label “organic” [defined and certified by the United States Department of Agriculture (USDA)], implies not using antibiotics or hormones in poultry livestock after the first 24 h of life. Thus, injecting antibiotics into eggs or administering antibiotics to one-day-old chicks are practices that are often performed and do not violate the USDA organic standard (Sanchez et al., 2020). Moreover, various forms of “no antibiotic” labels have been used, such as raised without antibiotics (RWA), no antibiotics administered, no added antibiotics, or raised antibiotic free, which describe meat from chicken that has not been administered antibiotics during production. However, the “no antibiotics ever (NAE)” label seem to be slightly stricter than the others, as it also restricts the antibiotic use in the egg (Sanchez et al., 2020; Singer et al., 2019). Most likely, the antibiotic-free label should guarantee that the meat being sold does not carry detectable levels of antibiotics.
In brief, retail poultry products have been known sources of antibiotic-resistant E. coli, and although consumers have a range of choices for poultry meat, including conventional, organic, and antibiotic-free designations, which are used to indicate differences in quality and safety, the frequency of contamination with antibiotic-resistant E. coli in food sold in these categories is unknown.
Although a limitation of our study is that we did not determine the source of contamination of commercialized chicken meat, there is no doubt that the occurrence of ESβL-producing E. coli in food sold for human consumption should not occur. In this respect, the food chain has globally been recognized as a reservoir and critical pathway for the development and dissemination of AMR, involving farming, processing, transportation, distribution, storage, retail and consumption (Founou et al., 2021; Choy et al., 2024; De Cesare et al., 2022); whereas the dissemination of blaCTX-M-type ESβL genes in human health is one of the main problems related to broad-spectrum cephalosporin resistance, particularly when associated with the spread of successful pandemic clones (Chong et al., 2018). Therefore, the presence of CTX-M-producing E. coli in retail chicken poses risk to human health, and studies investigating human acquisition through food consumption are necessary. Although it has been suggested that the consumption of chicken meat could be related to the acquisition of ESβL-producing E. coli and urinary tract infections, in most cases, human infections with ESβL-/pAmpC-producing E. coli are preceded by asymptomatic carriage (Manges et al., 2007; Isendahl et al., 2019; Plaza-Rodríguez et al., 2021; Dantas et al., 2025).
Further studies must also be directed to understand the evolutionary changes of CTX-M-positive E. coli in poultry meat, evaluate biofilm-forming capacity on the food chain, investigate interactions with food microbiota, develop quantitative microbial risk assessment models to estimate the risk of human and animal exposure, and expand surveillance. On the other hand, to assess the presumptive food safety and microbiological quality of foods, in addition to estimating bacterial numbers (i.e., total coliform and E. coli counts) as indicators of unfavorable hygienic conditions and fecal contamination in food, the absence of WHO critical priority Enterobacterales (resistant to broad-spectrum cephalosporins and/or carbapenems) in chicken meat should be used as a microbiological standard. On the other hand, the occurrence of CTX-M-positive E. coli in food has significant environmental and public health implications, including: (i) potential for horizontal gene transfer; (ii) long-term persistence in food waste; (iii) colonization of companion animals and/or humans, via contaminated food, creating new reservoirs for AMR; and, (iv) risk of community-acquired infections, especially among immunocompromised individuals.
Finally, widespread dissemination of ESβL (CTX-M)-positive E. coli has been favored by globalization of food trade, and this could be contributing to the successful dissemination of international clones, reaching parts of the world where they were not previously present (Dhanji et al., 2010; Kawamura et al., 2014; Egervärn et al., 2014; Nahar et al., 2018; Kim et al., 2018; Eibach et al., 2018; Campos et al., 2018; Kelbert et al., 2025). Given the global dynamics of ESβL transmission, a multisectoral and multidisciplinary approach is critical to the success of the global action plan on AMR.
Tetracyclines, β-lactams (penicillins), aminoglycosides, quinolone and sulfonamides are among the most widely used classes of antimicrobials in food-producing animals worldwide (Centner, 2016; Caneschi et al., 2023). Interestingly, in this study, all E. coli strains carried genes conferring resistance to β-lactams, and the majority (8/9, 88.9%) exhibited one or more genes conferring resistance to aminoglycosides and sulfonamides. Additionally, several strains also harbored mechanisms conferring resistance to tetracycline and quinolones. In line with our findings, some studies have indicated that there is no significant disparity between meat from AF and CN production system related to AMR genes (Ferri et al., 2023; Farooq et al., 2022; Rawat et al., 2024).
The plasmids IncFIB and Col were the most frequently found in the strains analyzed, suggesting a high risk of antibiotic resistance spread, as these plasmid groups are known for efficient horizontal gene transfer and carriage of resistance genes (Liu et al., 2024; Felix et al., 2024).
Regarding the virulome, its context has been studied among lineages belonging to specific phylogroups of E. coli (Beghain et al., 2018). In this study, the FBC4 strain, belonging to phylogroup A* (with potential mutation), was the one that presented the largest set of virulence genes. Despite this, strains of phylogroup A are normally associated with commensal lineages (Mosquito et al., 2015).
Another interesting point is the presence of critical-priority E. coli in FZ meat. The ability of these bacteria to survive in low-temperature stress has already been demonstrated (Parvin et al., 2020), underscoring the significance of processors adopting and adhering to good slaughtering and processing practices.
5 Conclusion
In summary, we report the detection of global WHO critical priority clones of CTX-M-type/pAmpC-producing E. coli in commercially available FS and FZ chicken meat from both AF and CV production systems in Brazil, which is considered a major global chicken meat exporter. We highlight that meat could serve as potential reservoirs and vectors of medically important antimicrobial-resistant bacteria, posing a significant threat to consumers. Merely discontinuing the use of antimicrobials in food-producing animals without addressing other factors may not fully resolve the issue of AMR in the meat industry and food safety. Our findings raise questions about the efficacy of current agricultural practices, antimicrobial usage in animal husbandry, and potential routes of contamination during meat processing and distribution. Addressing these issues is crucial and requires collaborative efforts among stakeholders in the food industry, veterinary and medical sectors, and governmental agencies to implement strategies aimed at reducing the prevalence of these critical-priority bacteria in meat. Hence, it is imperative to adopt multi-faceted approaches across the meat production chain, including better farm practices and biosecurity, responsible use of antimicrobials, strict hygiene practices, proper storage and transportation, effective regulatory measures, and education – supplemented by continuous surveillance of AMR in these products, to effectively mitigate contamination routes.
Data availability statement
This Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accessions JAQQRG000000000 (F1B), JABEPS000000000 (FCC3), JABEPT000000000 (FBP3), JABEPU000000000 (FCC4), JAQQRI000000000 (FBC4), JAQQYK000000000 (FCC8), JAQQYJ000000000 (FCC10), JAQQYM000000000 (FSE11), and JAQQYL000000000 (FSW11). Additionally, genomic data are available at the OneBR platform under the number IDs ONE135 (F1B), ONE136 (FCC3), ONE137 (FBP3), ONE138 (FCC4), ONE139 (FBC4), ONE140 (FCC8), ONE141 (FCC10), ONE142 (FSE11), and ONE143 (FSW11) (http://onehealthbr.com/).
Author contributions
BF: Writing – original draft, Formal analysis, Data curation, Conceptualization. IN: Data curation, Conceptualization, Writing – original draft, Formal analysis. HF: Formal analysis, Data curation, Writing – review & editing. JB: Writing – review & editing, Methodology. ES: Data curation, Formal analysis, Writing – review & editing. AC-A: Methodology, Writing – review & editing. FE: Writing – review & editing, Methodology. BC: Writing – review & editing, Methodology. SI: Writing – review & editing, Methodology. FS: Formal analysis, Writing – original draft, Conceptualization. NL: Writing – review & editing, Resources, Project administration, Supervision, Funding acquisition, Conceptualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported, in whole or in part, by the Bill & Melinda Gates Foundation [Grand Challenges Explorations Brazil OPP1193112]. Additionally, this study was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (2020/08224-9 and 2019/15578-4), Conselho Nacional de Desenvolvimento Científico e Tecnológico (grants AMR 422984/2021-3, and 314336/2021-4), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior [grants 88887.358057/2019-00 and 1794306]. HF and BC are CAPES fellows [88887.506496/2020-00, 88882.333054/2019-01]. FE is FAPESP fellow [2019/15778-4].
Acknowledgments
We thank Cefar Diagnóstica Ltda (Brazil) for kindly supplying antibiotic discs for susceptibility testing and CEFAP-GENIAL for sequencing.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1593887/full#supplementary-material
SUPPLEMENTARY FIGURE S1 | Phylogenetic analysis. In A1–A7, maximum-likelihood phylogenetic tree of F1B (ST443), FCC8 (ST3258), FCC4 (ST2179), FBC4 (ST2040), FCC3 (ST350), FCC10 (ST15579), and FSE11 (ST57) Escherichia coli strains. Comparison of resistomes, isolation sources, and countries of origin of the strains. In B1–B7, zooming into the subtree comprising sequenced strains of Escherichia coli in this study. The figure was generated with iTOL version 5.6.1 (https://itol.embl.de).
SUPPLEMENTARY TABLE S1 | Epidemiological characteristics, genetic repertoire, and SNP matrix of ESβL/pAmpC-producing Escherichia coli isolated from conventional and antibiotic-free retail meat in Brazil.
Footnotes
1. ^https://github.com/FelixKrueger/TrimGalore
2. ^https://github.com/rrwick/Unicycler
3. ^http://www.ncbi.nlm.nih.gov/genome/annotation_prok/
5. ^https://enterobase.warwick.ac.uk
6. ^https://www.genomicepidemiology.org
7. ^https://github.com/tseemann/abricate
8. ^https://github.com/haruosuz/vfdb
9. ^http://bacmet.biomedicine.gu.se/
10. ^http://clermontyping.iame-research.center
11. ^https://sarredondo.shinyapps.io/mlplasmids/
12. ^https://github.com/ParBLiSS/FastANI
References
Adegbeye, M. J., Adetuyi, B. O., Igirigi, A. I., Adisa, A., Palangi, V., Aiyedun, S., et al. (2024). Comprehensive insights into antibiotic residues in livestock products: distribution, factors, challenges, opportunities, and implications for food safety and public health. Food Control 163:110545. doi: 10.1016/j.foodcont.2024.110545
Alegría, Á., Arias-Temprano, M., Fernández-Natal, I., Rodríguez-Calleja, J. M., García-López, M. L., and Santos, J. A. (2020). Molecular diversity of ESΒL-producing Escherichia coli from foods of animal origin and human patients. Int. J. Environ. Res. Public Health 17:1312. doi: 10.3390/ijerph17041312
Antimicrobial Resistance Collaborators (2022). Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399, 629–655. doi: 10.1016/S0140-6736(21)02724-0
Argudín, M. A., Deplano, A., Meghraoui, A., Dodémont, M., Heinrichs, A., Denis, O., et al. (2017). Bacteria from animals as a Pool of antimicrobial resistance genes. Antibiotics 6:12. doi: 10.3390/antibiotics6020012
Aslam, B., Khurshid, M., Arshad, M. I., Muzammil, S., Rasool, M., Yasmeen, N., et al. (2021). Antibiotic resistance: one health one world outlook. Front. Cell. Infect. Microbiol. 11:771510. doi: 10.3389/fcimb.2021.771510
Beghain, J., Bridier-Nahmias, A., Le Nagard, H., Denamur, E., and Clermont, O. (2018). Clermontyping: an easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb. Genom. 4:e000192. doi: 10.1099/mgen.0.000192
Berg, E. S., Wester, A. L., Ahrenfeldt, J., Mo, S. S., Slettemeås, J. S., Steinbakk, M., et al. (2017). Norwegian patients and retail chicken meat share cephalosporin-resistant Escherichia coli and Inc K/blaCMY-2 resistance plasmids. Clin. Microbiol. Infect. 23, 407.e9–407.e15. doi: 10.1016/j.cmi.2016.12.035
Bergšpica, I., Kaprou, G., Alexa, E. A., Prieto, M., and Alvarez-Ordóñez, A. (2020). Extended Spectrum β-lactamase (ESBL) producing Escherichia coli in pigs and pork meat in the European Union. Antibiotics 9:678. doi: 10.3390/antibiotics9100678
Bertagnolio, S., Dobreva, Z., Centner, C. M., Olaru, I. D., Donà, D., Burzo, S., et al. (2024). WHO global research priorities for antimicrobial resistance in human health. Lancet Microbe 5:100902. doi: 10.1016/S2666-5247(24)00134-4
Campos, J., Mourão, J., Silveira, L., Saraiva, M., Correia, C. B., Maçãs, A. P., et al. (2018). Imported poultry meat as a source of extended-spectrum cephalosporin-resistant CMY-2-producing Salmonella Heidelberg and Salmonella Minnesota in the European Union, 2014-2015. Int. J. Antimicrob. Agents 51, 151–154. doi: 10.1016/j.ijantimicag.2017.09.006
Caneschi, A., Bardhi, A., Barbarossa, A., and Zaghini, A. (2023). The use of antibiotics and antimicrobial resistance in veterinary medicine, a complex phenomenon: a narrative review. Antibiotics 12:487. doi: 10.3390/antibiotics12030487
Casella, T., Haenni, M., Madela, N. K., Andrade, L. K., Pradela, L. K., Andrade, L. N., et al. (2018). Extended-spectrum cephalosporin-resistant Escherichia coli isolated from chickens and chicken meat in Brazil is associated with rare and complex resistance plasmids and pandemic ST lineages. J. Antimicrob. Chemother. 73, 3293–3297. doi: 10.1093/jac/dky335
Castellanos, L. R., Donado-Godoy, P., León, M., Clavijo, V., Arevalo, A., Bernal, J. F., et al. (2017). High heterogeneity of Escherichia coli sequence types harbouring ESΒL/amp C genes on IncI1 plasmids in the Colombian poultry chain. PLoS One 12:e0170777. doi: 10.1371/journal.pone.0170777
Centner, T. J. (2016). Efforts to slacken antibiotic resistance: labeling meat products from animals raised without antibiotics in the United States. Sci. Total Environ. 563-564, 1088–1094. doi: 10.1016/j.scitotenv.2016.05.082
Che, M., Birk, T., and Hansen, L. T. (2023). Prevalence and transmission of extended-Spectrum cephalosporin (ESC) resistance genes in Escherichia coli isolated from poultry production systems and slaughterhouses in Denmark. Antibiotics 12:1602. doi: 10.3390/antibiotics12111602
Chong, Y., Shimoda, S., and Shimono, N. (2018). Current epidemiology, genetic evolution and clinical impact of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae. Infect. Genet. Evol. 61, 185–188. doi: 10.1016/j.meegid.2018.04.005
Choy, S. K., Neumann, E.-M., Romero-Barrios, P., and Tamber, S. (2024). Contribution of food to the human health burden of antimicrobial resistance. Foodborne Pathog. Dis. 21, 71–82. doi: 10.1089/fpd.2023.0099
Clinical and Laboratory Standards Institute (2024). Performance standards for antimicrobial susceptibility testing. Thirty-first informational supplement M100-S34. Wayne, PA: CLSI.
Coppola, N., Cordeiro, N. F., Trenchi, G., Esposito, F., Fuga, B., Fuentes-Castillo, D., et al. (2022). Imported one-day-old chicks as Trojan horses for multidrug-resistant priority pathogens harboring mcr-9, rmtG, and extended-Spectrum β-lactamase genes. Appl. Environ. Microbiol. 88:e0167521. doi: 10.1128/AEM.01675-21
Da Silva, C. R., do Valle, B. M., Gozi, K. S., Fontana, H., Nogueira, M. C. L., Lincopan, N., et al. (2022). Genomic analysis of Escherichia coli circulating in the Brazilian poultry sector. Braz. J. Microbiol. 53, 2121–2131. doi: 10.1007/s42770-022-00799-x
Dantas, K., Melocco, G., Esposito, F., Fontana, H., Cardoso, B., and Lincopan, N. (2025). Emergent Escherichia coli of the highly virulent B2-ST1193 clone producing KPC-2 carbapenemase in ready-to-eat vegetables. J. Glob. Antimicrob. Resist. 41, 105–110. doi: 10.1016/j.jgar.2024.11.020
Davidova-Gerzova, L., Lausova, J., Sukkar, I., Nesporova, K., Nechutna, L., Vlkova, K., et al. (2023). Hospital and community wastewater as a source of multidrug-resistant ESBL-producing Escherichia coli. Front. Cell. Infect. Microbiol. 15:1184081. doi: 10.3389/fcimb.2023.1184081
Davis, G. S., Waits, K., Nordstrom, L., Grande, H., Weaver, B., Papp, K., et al. (2018). Antibiotic-resistant Escherichia coli from retail poultry meat with different antibiotic use claims. BMC Microbiol. 18:174. doi: 10.1186/s12866-018-1322-5
De Cesare, A., Oliveri, C., Lucchi, A., Savini, F., Manfreda, G., and Sala, C. (2022). Pilot study on poultry meat from antibiotic free and conventional farms: can metagenomics detect any difference? Food Secur. 11:249. doi: 10.3390/foods11030249
Dhanji, H., Murphy, N. M., Doumith, M., Durmus, S., Lee, S. S., Hope, R., et al. (2010). Cephalosporin resistance mechanisms in Escherichia coli isolated from raw chicken imported into the UK. J. Antimicrob. Chemother. 65, 2534–2537. doi: 10.1093/jac/dkq376
Diab, M., Hamze, M., Bonnet, R., Saras, E., Madec, J. Y., and Haenni, M. (2018). Extended-spectrum beta-lactamase (ESBL)- and carbapenemase-producing Enterobacteriaceae in water sources in Lebanon. Vet. Microbiol. 217, 97–103. doi: 10.1016/j.vetmic.2018.03.007
Dos Anjos Adur, M., Châtre, P., Métayer, V., Drapeau, A., Pillonetto, M., Penkal, M. L., et al. (2022). Escherichia coli ST224 and Inc F/blaCTX-M-55 plasmids drive resistance to extended-spectrum cephalosporins in poultry flocks in Parana, Brazil. Int J Food Microbiol 380:109885. doi: 10.1016/j.ijfoodmicro.2022.109885
Drieux, L., Brossier, F., Sougakoff, W., and Jarlier, V. (2008). Phenotypic detection of extended-spectrum beta-lactamase production in Enterobacteriaceae: review and bench guide. Clin. Microbiol. Infect. 14, 90–103. doi: 10.1111/j.1469-0691.2007.01846.x
Egervärn, M., Börjesson, S., Byfors, S., Finn, M., Kaipe, C., Englund, S., et al. (2014). Escherichia coli with extended-spectrum beta-lactamases or transferable AmpC beta-lactamases and Salmonella on meat imported into Sweden. Int. J. Food Microbiol. 171, 8–14. doi: 10.1016/j.ijfoodmicro.2013.11.005
Eibach, D., Dekker, D., Gyau Boahen, K., Wiafe Akenten, C., Sarpong, N., Belmar Campos, C., et al. (2018). Extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae in local and imported poultry meat in Ghana. Vet. Microbiol. 217, 7–12. doi: 10.1016/j.vetmic.2018.02.023
Farooq, M., Smoglica, C., Ruffini, F., Soldati, L., Marsilio, F., and Di Francesco, C. E. (2022). Antibiotic resistance genes occurrence in conventional and antibiotic-free poultry farming, Italy. Animals 12:2310. doi: 10.3390/ani12182310
Felix, M. A., Sopovski, D., Commichaux, S., Yoskowitz, N., Aljahdali, N. H., Grim, C. J., et al. (2024). Genetic relatedness and virulence potential of Salmonella Schwarzengrund strains with or without an IncFIB-IncFIC (FII) fusion plasmid isolated from food and clinical sources. Front. Microbiol. 15:1397068. doi: 10.3389/fmicb.2024.1397068
Ferri, G., Buonavoglia, A., Farooq, M., Festino, A. R., Ruffini, F., Paludi, D., et al. (2023). Antibiotic resistance in Italian poultry meat production chain: a one-health perspective comparing antibiotic free and conventional systems from the farming to the slaughterhouse. Front. Food Sci. Technol. 3:1168896. doi: 10.3389/frfst.2023.1168896
Food and Agriculture Organization of the United Nations. (2019). Prudent and efficient use of antimicrobials in pigs and poultry. Available online at: https://www.fao.org/documents/card/fr/c/CA6729EN (Accessed January 15, 2025).
Foster-Nyarko, E., and Pallen, M. J. (2022). The microbial ecology of Escherichia coli in the vertebrate gut. FEMS Microbiol. Rev. 46:fuac008. doi: 10.1093/femsre/fuac008
Founou, L. L., Founou, R. C., and Essack, S. Y. (2021). Antimicrobial resistance in the farm-to-plate continuum: more than a food safety issue. Future Sci. OA 7:FSO692. doi: 10.2144/fsoa-2020-0189
Fuga, B., Sellera, F. P., Cerdeira, L., Esposito, F., Cardoso, B., Fontana, H., et al. (2022). WHO critical priority Escherichia coli as one health challenge for a post-pandemic scenario: genomic surveillance and analysis of current trends in Brazil. Microbiol. Spectr. 10:e0125621. doi: 10.1128/spectrum.01256-21
Grevskott, D. H., Radisic, V., Salvà-Serra, F., Moore, E. R. B., Akervold, K. S., Victor, M. P., et al. (2024). Emergence and dissemination of epidemic-causing OXA-244 carbapenemase-producing Escherichia coli ST38 through hospital sewage in Norway, 2020-2022. J. Hosp. Infect. 145, 165–173. doi: 10.1016/j.jhin.2023.12.020
Haque, M. H., Sarker, S., Islam, M. S., Islam, M. A., Karim, M. R., Kayesh, M. E. H., et al. (2020). Sustainable antibiotic-free broiler meat production: current trends, challenges, and possibilities in a developing country perspective. Biology 9:411. doi: 10.3390/biology9110411
Hassen, B., Abbassi, M. S., Ruiz-Ripa, L., Mama, O. M., Ibrahim, C., Benlabidi, S., et al. (2021). Genetic characterization of extended-spectrum β-lactamase-producing Enterobacteriaceae from a biological industrial wastewater treatment plant in Tunisia with detection of the colistin-resistance mcr-1 gene. FEMS Microbiol. Ecol. 97:fiaa231. doi: 10.1093/femsec/fiaa231
Hooban, B., Fitzhenry, K., Cahill, N., Joyce, A., O' Connor, L., Bray, J. E., et al. (2021). A point prevalence survey of antibiotic resistance in the Irish environment, 2018-2019. Environ. Int. 152:106466. doi: 10.1016/j.envint.2021.106466
IBGE. (2025) Brazilian institute of geography and statistics. Available online at: https://cidades.ibge.gov.br/brasil/sp/sao-paulo/panorama (Accessed May 04, 2025).
Isendahl, J., Giske, C. G., Hammar, U., Sparen, P., Tegmark Wisell, K., Ternhag, A., et al. (2019). Temporal dynamics and risk factors for bloodstream infection with extended-spectrum β-lactamase-producing bacteria in previously-colonized individuals: national population-based cohort study. Clin. Infect. Dis. 68, 641–649. doi: 10.1093/cid/ciy539
Jacob, M. E., Keelara, S., Aidara-Kane, A., Matheu Alvarez, J. R., and Fedorka-Cray, P. J. (2020). Optimizing a screening protocol for potential extended-spectrum β-lactamase Escherichia coli on MacConkey agar for use in a global surveillance program. J. Clin. Microbiol. 58:e01039-19. doi: 10.1128/JCM.01039-19
Jaleta, M., Junker, V., Kolte, B., Börger, M., Werner, D., Dolsdorf, C., et al. (2024). Improvements of weaned pigs barn hygiene to reduce the spread of antimicrobial resistance. Front. Microbiol. 14:1393923. doi: 10.3389/fmicb.2024.1393923
Jarlier, V., Nicolas, M. H., Fournier, G., and Philippon, A. (1998). Extended broad-spectrum beta-lactamases conferring transferable resistance to newer beta-lactam agents in Enterobacteriaceae: hospital prevalence and susceptibility patterns. Rev. Infect. Dis. 10, 867–878. doi: 10.1093/clinids/10.4.867
Jesudason, T. (2023). A new research agenda to combat antimicrobial resistance. Lancet Infect. Dis. 23:e281. doi: 10.1016/S1473-3099(23)00446-2
Johnsen, P. J., Townsend, J. P., Bøhn, T., Simonsen, G. S., Sundsfjord, A., and Nielsen, K. M. (2009). Factors affecting the reversal of antimicrobial-drug resistance. Lancet Infect. Dis. 9, 357–364. doi: 10.1016/S1473-3099(09)70105-7
Kaper, J. B., Nataro, J. P., and Mobley, H. L. (2004). Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2, 123–140. doi: 10.1038/nrmicro818
Kawamura, K., Goto, K., Nakane, K., and Arakawa, Y. (2014). Molecular epidemiology of extended-spectrum β-lactamases and Escherichia coli isolated from retail foods including chicken meat in Japan. Foodborne Pathog. Dis. 11, 104–110. doi: 10.1089/fpd.2013.1608
Keck, N., Treilles, M., Gordoncillo, M., Ivette, O. L. I., Dauphin, G., Dorado-Garcia, A., et al. (2023). A systematic approach toward progressive improvement of national antimicrobial resistance surveillance systems in food and agriculture sectors. Front. Vet. Sci. 9:1057040. doi: 10.3389/fvets.2022.1057040
Kelbert, L., Barmettler, K., Biggel, M., Stephan, R., and Nüesch-Inderbinen, M. (2025). Occurrence and characteristics of extended-spectrum ß-lactamase-producing Escherichia coli in Swiss and imported retail chicken meat. J. Glob. Antimicrob. Resist. 43, 285–292. doi: 10.1016/j.jgar.2025.05.013
Kim, Y. J., Moon, J. S., Oh, D. H., Chon, J. W., Song, B. R., Lim, J. S., et al. (2018). Genotypic characterization of ESBL-producing E. coli from imported meat in South Korea. Food Res. Int. 107, 158–164. doi: 10.1016/j.foodres.2017.12.023
Klein, H., and Vidal, F. (2022). The emergence of Brazil as the leading world exporter of chicken meat. Hist. Agrar. Am. Lat. 3, 75–99. doi: 10.53077/haal.v3i02.127
Leigue, L., Warth, J. F., Melo, L. C., Silva, K. C., Moura, R. A., Barbato, L., et al. (2015). MDR ST2179-CTX-M-15 Escherichia coli co-producing RmtD and AAC(6′)-Ib-cr in a horse with extraintestinal infection, Brazil. J. Antimicrob. Chemother. 70, 1263–1265. doi: 10.1093/jac/dku520
Li, Q., Li, Z., Wang, Y., Chen, Y., Sun, J., Yang, Y., et al. (2022). Antimicrobial resistance and transconjugants characteristics of sul3 positive Escherichia coli isolated from animals in Nanning, Guangxi Province. Animals 12:976. doi: 10.3390/ani12080976
Li, Q., Zou, H., Wang, D., Zhao, L., Meng, M., Wang, Z., et al. (2023). Tracking spatio-temporal distribution and transmission of antibiotic resistance in aquatic environments by using ESBL-producing Escherichia coli as an indicator. J. Environ. Manag. 344:118534. doi: 10.1016/j.jenvman.2023.118534
Liu, H., Fan, S., Zhang, X., Yuan, Y., Zhong, W., Wang, L., et al. (2024). Antibiotic-resistant characteristics and horizontal gene transfer ability analysis of extended-spectrum β-lactamase-producing Escherichia coli isolated from giant pandas. Front. Vet. Sci. 11:1394814. doi: 10.3389/fvets.2024.1394814
Magiorakos, A. P., Srinivasan, A., Carey, R. B., Carmeli, Y., Falagas, M. E., Giske, C. G., et al. (2012). Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 18, 268–281. doi: 10.1111/j.1469-0691.2011.03570.x
Manges, A. R., Smith, S. P., Lau, B. J., Nuval, C. J., Eisenberg, J. N., Dietrich, P. S., et al. (2007). Retail meat consumption and the acquisition of antimicrobial resistant Escherichia coli causing urinary tract infections: a case-control study. Foodborne Pathog. Dis. 4, 419–431. doi: 10.1089/fpd.2007.0026
Martínez-Álvarez, S., Châtre, P., François, P., Zarazaga, M., Madec, J. Y., Haenni, M., et al. (2025). Comparative phylogenomics of extended-spectrum beta-lactamase-producing Escherichia coli revealed a wide diversity of clones and plasmids in Spanish chicken meat. Int. J. Food Microbiol. 426:110900. doi: 10.1016/j.ijfoodmicro.2024.110900
McEwen, S. A., and Collignon, P. J. (2018). Antimicrobial resistance: a one health perspective. Microbiol. Spectr. 6. doi: 10.1128/microbiolspec.arba-0009-2017
Millman, J. M., Waits, K., Grande, H., Marks, A. R., Marks, J. C., Price, L. B., et al. (2013). Prevalence of antibiotic-resistant E. coli in retail chicken: comparing conventional, organic, kosher, and raised without antibiotics. F1000Res 2:155. doi: 10.12688/f1000research.2-155.v2
Mo, S. S., Fiskebeck, E. Z., Slettemeås, J. S., Lagesen, K., Nilsson, O., Naseer, U., et al. (2023). Escherichia coli multilocus sequence type 38 from humans and broiler production represent distinct monophyletic groups. Front. Microbiol. 14:1173287. doi: 10.3389/fmicb.2023.1173287
Mohammadi, H., Saghaian, S., and Boccia, F. (2023). Antibiotic-free poultry meat consumption and its determinants. Food Secur. 12:1776. doi: 10.3390/foods12091776
Mosquito, S., Pons, M. J., Riveros, M., Ruiz, J., and Ochoa, T. J. (2015). Diarrheagenic Escherichia coli phylogroups are associated with antibiotic resistance and duration of diarrheal episode. Sci. World J. 2015. doi: 10.1155/2015/610403
Mughini-Gras, L., Dorado-García, A., van Duijkeren, E., van den Bunt, G., Dierikx, C. M., Bonten, M. J. M., et al. (2019). Attributable sources of community-acquired carriage of Escherichia coli containing β-lactam antibiotic resistance genes: a population-based modelling study. Lancet Planet Health 3, e357–e369. doi: 10.1016/S2542-5196(19)30130-5
Müller, A., Stephan, R., and Nüesch-Inderbinen, M. (2016). Distribution of virulence factors in ESBL-producing Escherichia coli isolated from the environment, livestock, food and humans. Sci. Total Environ. 541, 667–672. doi: 10.1016/j.scitotenv.2015.09.135
Murray, M., Salvatierra, G., Dávila-Barclay, A., Ayzanoa, B., Castillo-Vilcahuaman, C., Huang, M., et al. (2021). Market chickens as a source of antibiotic-resistant Escherichia coli in a Peri-Urban Community in Lima, Peru. Front. Microbiol. 12:635871. doi: 10.3389/fmicb.2021.635871
Nahar, A., Awasthi, S. P., Hatanaka, N., Okuno, K., Hoang, P. H., Hassan, J., et al. (2018). Prevalence and characteristics of extended-spectrum β-lactamase-producing Escherichia coli in domestic and imported chicken meats in Japan. J. Vet. Med. Sci. 80, 510–517. doi: 10.1292/jvms.17-0708
Oikarainen, P. E., Pohjola, L. K., Pietola, E. S., and Heikinheimo, A. (2019). Direct vertical transmission of ESBL/pAmpC-producing Escherichia coli limited in poultry production pyramid. Vet. Microbiol. 231, 100–106. doi: 10.1016/j.vetmic.2019.03.001
Palmeira, J. D., Ferreira, H., Madec, J. Y., and Haenni, M. (2018). Draft genome of a ST443 mcr-1- and blaCTX-M-2-carrying Escherichia coli from cattle in Brazil. J. Glob. Antimicrob. Resist. 13, 269–270. doi: 10.1016/j.jgar.2018.05.010
Park, J. H., Kim, H. S., Yim, J. H., Kim, Y. J., Kim, D. H., Chon, J. W., et al. (2017). Comparison of the isolation rates and characteristics of Salmonella isolated from antibiotic-free and conventional chicken meat samples. Poult. Sci. 96, 2831–2838. doi: 10.3382/ps/pex055
Parvin, M. S., Talukder, S., Ali, M. Y., Chowdhury, E. H., Rahman, M. T., and Islam, M. T. (2020). Antimicrobial resistance pattern of Escherichia coli isolated from frozen chicken meat in Bangladesh. Pathogens 9:420. doi: 10.3390/pathogens9060420
Plaza-Rodríguez, C., Mesa-Varona, O., Alt, K., Grobbel, M., Tenhagen, B. A., and Kaesbohrer, A. (2021). Comparative analysis of consumer exposure to resistant Bacteria through chicken meat consumption in Germany. Microorganisms 9:1045. doi: 10.3390/microorganisms9051045
Puljko, A., Babić, I., Rozman, S. D., Barišić, I., Jelić, M., Maravić, A., et al. (2024). Treated municipal wastewater as a source of high-risk and emerging multidrug-resistant clones of E. Coli and other Enterobacterales producing extended-spectrum β-lactamases. Environ. Res. 243:117792. doi: 10.1016/j.envres.2023.117792
Ramos, S., Silva, V., Dapkevicius, M. L. E., Caniça, M., Tejedor-Junco, M. T., Igrejas, G., et al. (2020). Escherichia coli as commensal and pathogenic bacteria among food-producing animals: health implications of extended spectrum β-lactamase (ESΒL) production. Animals 10:2239. doi: 10.3390/ani10122239
Rawat, N., Anjali, S., Sabu, B., Bandyopadhyay, A., and Rajagopal, R. (2024). Assessment of antibiotic resistance in chicken meat labelled as antibiotic-free: a focus on Escherichia coli and horizontally transmissible antibiotic resistance genes. LWT 194:115751. doi: 10.1016/j.lwt.2024.115751
Saidenberg, A. B. S., Edslev, S. M., Hallstrøm, S., Rasmussen, A., Park, D. E., Aziz, M., et al. (2024). Escherichia coli ST117: exploring the zoonotic hypothesis. Microbiol. Spectr. 3:e0046624. doi: 10.1128/spectrum.00466-24
Sanchez, H. M., Whitener, V. A., Thulsiraj, V., Amundson, A., Collins, C., Duran-Gonzalez, M., et al. (2020). Antibiotic Resistance of Escherichia coli Isolated from Conventional, No Antibiotics, and Humane Family Owned Retail Broiler Chicken Meat. Animals (Basel). 26:2217. doi: 10.3390/ani10122217
Sarkar, S., Souza, M. J., Martin-Jimenez, T., Abouelkhair, M. A., Kania, S. A., and Okafor, C. C. (2023). Tetracycline, sulfonamide, and erythromycin residues in beef, eggs, and honey sold as "antibiotic-free" products in East Tennes(see USA) Farmers' Markets. Vet. Sci. 10:243. doi: 10.3390/vetsci10040243
Seni, J., Moremi, N., Matee, M., van der Meer, F., DeVinney, R., Mshana, S. E., et al. (2018). Preliminary insights into the occurrence of similar clones of extended-spectrum beta-lactamase-producing bacteria in humans, animals and the environment in Tanzania: a systematic review and meta-analysis between 2005 and 2016. Zoonoses Public Health 65, 1–10. doi: 10.1111/zph.12387
Singer, R. S., Porter, L. J., Thomson, D. U., Gage, M., Beaudoin, A., and Wishnie, J. K. (2019). Raising Animals Without Antibiotics: U.S. Producer and Veterinarian Experiences and Opinions. Front Vet Sci. 6:452. doi: 10.3389/fvets.2019.00452
Singhal, N., Kumar, M., Kanaujia, P. K., and Virdi, J. S. (2015). MALDI-TOF mass spectrometry: an emerging technology for microbial identification and diagnosis. Front. Microbiol. 6:791. doi: 10.3389/fmicb.2015.00791
Soncini, J. G. M., Cerdeira, L., Sano, E., Koga, V. L., Tizura, A. T., Tano, Z. N., et al. (2022). Genomic insights of high-risk clones of ESΒL-producing Escherichia coli isolated from community infections and commercial meat in southern Brazil. Sci. Rep. 12:9354. doi: 10.1038/s41598-022-13197-y
Sun, Z., Hong, W., Xue, C., and Dong, N. (2024). A comprehensive review of antibiotic resistance gene contamination in agriculture: challenges and AI-driven solutions. Sci. Total Environ. 953:175971. doi: 10.1016/j.scitotenv.2024.175971
Tafoukt, R., Touati, A., Leangapichart, T., Bakour, S., and Rolain, J. M. (2017). Characterization of OXA-48-like-producing Enterobacteriaceae isolated from river water in Algeria. Water Res. 120, 185–189. doi: 10.1016/j.watres.2017.04.073
Tang, K. L., Caffrey, N. P., Nóbrega, D. B., Cork, S. C., Ronksley, P. E., Barkema, H. W., et al. (2019). Comparison of different approaches to antibiotic restriction in food-producing animals: stratified results from a systematic review and meta-analysis. BMJ Glob. Health 4:e001710. doi: 10.1136/bmjgh-2019-001710
Thanner, S., Drissner, D., and Walsh, F. (2016). Antimicrobial resistance in agriculture. MBio 7:e02227. doi: 10.1128/mBio.02227-15
Tian, M., He, X., Feng, Y., Wang, W., Chen, H., Gong, M., et al. (2021). Pollution by antibiotics and antimicrobial resistance in LiveStock and poultry manure in China, and countermeasures. Antibiotics 10:539. doi: 10.3390/antibiotics10050539
U.S. Food and Drug Administration (2021). National antimicrobial resistance monitoring system (NARMS): retail meat surveillance laboratory protocol. Maryland: FDA.
van Hoek, A. H. A. M., Veenman, C., Florijn, A., Huijbers, P. M. C., Graat, E. A. M., de Greeff, S., et al. (2018). Longitudinal study of ESΒL Escherichia coli carriage on an organic broiler farm. J. Antimicrob. Chemother. 73, 3298–3304. doi: 10.1093/jac/dky362
Warriner, K., Aldsworth, T. G., Kaur, S., and Dodd, C. E. (2002). Cross-contamination of carcasses and equipment during pork processing. J. Appl. Microbiol. 93, 169–177. doi: 10.1046/j.1365-2672.2002.01678.x
World Health Organization (2017). Stop using antibiotics in healthy animals to prevent the spread of antibiotic resistance. Geneva: World Health Organization.
World Health Organization (2024). WHO bacterial priority pathogens list, 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance. Geneva: World Health Organization.
Keywords: foodborne pathogens, retail meat, chicken meat, Enterobacterales, genomic surveillance
Citation: Fuga B, Neves I, Fontana H, Bispo J, Sano E, Cardenas-Arias A, Esposito F, Cardoso B, Ienne S, Sellera FP and Lincopan N (2025) Extended-spectrum β-lactamase (ESβL)-producing Escherichia coli in antibiotic-free and conventional chicken meat, Brazil. Front. Microbiol. 16:1593887. doi: 10.3389/fmicb.2025.1593887
Edited by:
Monica Alejandra Delgado, CONICET Higher Institute of Biological Research (INSIBIO), ArgentinaReviewed by:
Marta Laranjo, University of Evora, PortugalDiana Claudia Marcela Alvarez Espejo, Pontificia Universidad Católica de Chile, Chile
Copyright © 2025 Fuga, Neves, Fontana, Bispo, Sano, Cardenas-Arias, Esposito, Cardoso, Ienne, Sellera and Lincopan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bruna Fuga, YnJ1bmEuZnVnYUBob3RtYWlsLmNvbQ==; Nilton Lincopan, bGluY29wYW5AdXNwLmJy
†These authors have contributed equally to this work