 Juan Vicente Farizano1
Juan Vicente Farizano1 Emilia Castagnaro1
Emilia Castagnaro1 Julián Thomas Arroyo-Egea1
Julián Thomas Arroyo-Egea1 Juan Daniel Aparicio2Alicia Cecilia Vallejos1,3
Juan Daniel Aparicio2Alicia Cecilia Vallejos1,3 Elvira María Hebert4
Elvira María Hebert4 Lucila Saavedra4
Lucila Saavedra4 Viviana Andrea Rapisarda1
Viviana Andrea Rapisarda1 Josefina María Villegas1
Josefina María Villegas1 Mariana Grillo-Puertas1*
Mariana Grillo-Puertas1*- 1Instituto Superior de Investigaciones Biológicas (INSIBIO), CONICET-UNT, and Instituto de Química Biológica “Dr. Bernabé Bloj”, Facultad de Bioquímica, Química y Farmacia, UNT, San Miguel de Tucumán, Argentina
- 2Planta Piloto de Procesos Industriales Microbiológicos (PROIMI-CONICET), Tucumán, Argentina
- 3Hospital Eva Perón, Ministerio de Salud Pública de Tucumán, Tucumán, Argentina
- 4Centro de Referencia para Lactobacilos (CERELA-CONICET), Tucumán, Argentina
Indwelling devices, such as double-J stents, are commonly used in urological surgery and are often associated with a high risk of urinary tract infections (UTIs) due to biofilm-related complications. In this study, we characterized 27 clinical bacterial isolates from double-J catheters, including Staphylococcus spp., Enterococcus faecalis, Klebsiella pneumoniae, Escherichia coli, and Bacillus spp., to investigate their pathogenic potential. Our findings revealed that strong biofilm producers (E. coli, S. aureus, and B. subtilis) exhibited robust extracellular matrix synthesis. Notably, multidrug resistance was observed in E. coli, K. pneumoniae, and E. faecalis. Mixed-culture experiments demonstrated that Bacillus spp. enhanced the biofilm formation of uropathogens, suggesting a potential impact on clinical outcomes. The characterization of the polymicrobial population colonizing double-J catheters, as conducted in this study, is essential for understanding the complexity and clinical behavior of biofilm-related infections associated with medical devices. Additionally, our results highlight the clinical relevance of underreported genera, such as Bacillus, which are often overlooked in routine clinical diagnostics. Gaining insights into the interaction mechanisms and survival strategies of several bacterial species colonizing double-J catheters may help shift current paradigms in understanding catheter-related infections.
Introduction
Urinary tract infections (UTIs) affect a large number of people, with prevalence varying by age group, sex, and the presence of risk factors (Echverría-Zarate et al., 2006; Pigrau, 2013; Delcaru et al., 2016; Klein and Hultgren, 2020). The high incidence of UTIs, along with increased recurrence rates and the potential for complications, contributes to the cost of medical treatment, amounting to billions of dollars in healthcare expenditures (Cepas et al., 2019). The primary cause of UTIs is the invasion of the urinary tract by microorganisms known as uropathogens (UP). The leading etiologic agent in human UTIs is uropathogenic E. coli (UPEC), responsible for up to 80% of all cases (Ronald, 2002). Other microorganisms, such as Staphylococcus spp., Proteus mirabilis, Proteus vulgaris, Klebsiella spp., Enterococcus faecalis, and Pseudomonas aeruginosa, account for the remaining 20% of infections (Nicolle, 2000; Flores-Mireles et al., 2015).
The risk of developing an UTI significantly increases with the use of indwelling devices, as they disrupt host defense mechanisms and facilitate the access of UP to the bladder or the upper urinary tract (Wu et al., 2015). The double-J stent is a commonly used device in urological surgery, serving dual functions by providing both support and drainage (Ozturk, 2015; Gauhar et al., 2022; Lin et al., 2022). However, ureteral stents are often associated with severe complications, primarily due to biofilm formation on the stent surface, which can lead to infections. These infections may result in bacteremia, renal function deterioration, pyelonephritis, or even mortality due to sepsis (Ranganathan et al., 2009). Biofilms developing on ureteral stents can originate from the urinary tract microbiota or from contamination during stent insertion (Bossa et al., 2017). The internal environment of ureteral stents provides an ideal medium for bacterial adhesion, colonization, and biofilm formation (Zumstein et al., 2017). An additional challenge with these stents lies in diagnosis, as colonizing bacteria may not be detected in urine or prostatic secretion samples (Klis et al., 2009). Therefore, identifying the pathogenic bacteria involved in double-J catheter colonization is crucial for effective treatment of these infections.
Bacterial populations have the capacity to rapidly adapt to changes in their environmental surroundings (O'Toole and Kolter, 1998; Monds and O'Toole, 2009). The ability of UP strains to cause various types and severities of diseases depends on the expression of multiple virulence traits, including biofilm formation, motility, hemolytic activity, expression of several virulence genes related to adhesin production, toxins, siderophores, and secretion systems, among others (Blanco et al., 2002; Johnson, 2002). UP are often found in a sessile state on the intra- and/or extraluminal surfaces of urinary catheters removed from patients with UTIs (Li et al., 2023).
Over the years, several reports have shown that the majority of biofilms formed on long-term catheters are polymicrobial, with an average of 2–5 microbial species isolated per catheter, predominantly comprising Gram-negative bacteria (Nicolle, 2012; Azevedo et al., 2017; Chua et al., 2017; Flores-Mireles et al., 2019; Zboromyrska et al., 2019). UP isolates from polymicrobial biofilms in catheterized patients have exhibited significant resistance to commonly used antibiotics, thereby increasing the risk of developing severe infections (Croxall et al., 2011; Kazi et al., 2015). In the context of the urinary tract, the presence of multiple microorganisms in a midstream urine sample is often attributed to contamination from periurethral or vaginal microbiota (Hooton, 2012; Kline and Lewis, 2016). Polymicrobial samples may also be classified as negative cultures if they lack a single dominant species, or they may be categorized as containing secondary pathogens or doubtful pathogens, depending on the level of colonization. As a result, the virulence and complexity of the often underestimated polymicrobial colonization of double-J catheters remain poorly characterized. In this context, this study aimed to investigate the virulence-associated phenotypic traits and antimicrobial resistance of bacteria isolated from double-J catheters, as well as to explore the interactions within mixed biofilms formed by co-isolated bacterial pairs. Through the comprehensive characterization of 27 isolates from this complex niche, we analyzed the virulent potential and interactions between known UP and understudied species such as Bacillus spp. Such knowledge is crucial for better managing catheter-associated infections, which significantly impact patient outcomes and healthcare systems.
Materials and methods
Catheter processing and UP isolation
Samples were taken from eight double-J-type urinary catheters provided by a specialized private urological center in San Miguel de Tucumán, Argentina. In all cases, 30–60 days urinary catheters were collected from adult male and female patients without UTI symptoms and with a negative urine culture. Each catheter was processed under sterile conditions to remove adherent and non-adherent bacteria, as reported by Mandakhalikar et al. (2018). Briefly, to extract unattached cells, four washes with physiological saline (PS) solution were performed. To extract adhered bacteria from catheters, the vortexing–sonication–vortexing (V-S-V) method was carried out (Mandakhalikar et al., 2018). Both unattached and attached bacteria were concentrated by centrifugation for 10 min at 7012 × g. Then, cells were resuspended in PS and seeded with a Digralsky spatula onto Cystine Lactose Electrolyte Deficient Agar (CLED-agar, Britania) plates. The plates were incubated for 24–48 h at 37°C to evaluate microbial growth. Colonies with different morphologies were purified using the quadrant streak method on CLED-agar medium. Pure isolates of potential UP were preserved for subsequent identification.
Identification of UP isolates
To identify the obtained UP isolates, genomic DNA was extracted using the organic solvent extraction method reported by Sambrook et al. (1989). The extracted genomic DNA was quantified using a NanoDrop spectrophotometer (UV/Vis nano spectrophotometer, Nabi).
Then, amplification of the 16S rRNA gene was conducted and sequenced to identify the genus of the obtained UP isolates (Sanger sequencing, Macrogen Inc.). Moreover, the identification of UP at the species level was carried out according to the standard methods and recommendations of the International Committee of Clinical Laboratories (CLSI, 2020). Standard biochemical tests and confirmation by MALDI-TOF mass spectrometry techniques were also used for identification (Bruker Daltonics; Perilla et al., 2009; Murray et al., 2021).
Culture conditions
Isolates were routinely grown under aerobic conditions in BHI medium at 37°C with shaking (180 rpm) or in static growth at 37°C on CLED-agar plates. Bacterial growth was monitored by measuring the absorbance at 600 nm (A600 nm) to determine turbidity in a liquid medium or by observing the appearance of isolated colonies in a solid medium. MacConkey (Britania), Luria Broth (LB, Sigma-Aldrich), M63 (in g/L: 2.0 ammonium sulfate, 13.6 monopotassium phosphate, 0.005 ferrous sulfate heptahydrate, 0.12 magnesium sulfate, 2.0 glycerol), Tryptic Soy Broth (TSB, Britania), Brain Heart Infusion (BHI, Britania) and urine media were used, depending on the assay. Human urine media was prepared according to Eberly et al. (2020). Briefly, urine was collected in equal volumes from male and female healthy volunteers and filtered through a 0.22 μm filter prior to use. Healthy volunteers were classified as individuals who are urologically asymptomatic, not menstruating, and those who have not taken antibiotics in the last 90 days.
Biofilm formation and quantification
Biofilm formation was assessed by measuring the ability of cells to adhere and grow in 96-well polystyrene plates using various culture media (O'Toole and Kolter, 1998). Briefly, 24-h cultures of all isolates were washed in PS, adjusted to an A600 nm of 1.0, and then diluted to an A600 nm of 0.1 in the corresponding media. Suspensions were loaded into multiwell plates and incubated under static conditions at 30°C for the specified times in each assay. Then, unattached (planktonic) cells were removed, and wells were washed three times with distilled water. The quantification of adherent cells or biofilm biomass was performed as described by O'Toole and Kolter (1998). Each condition was performed in quadruplicate, and each experiment was repeated at least five times.
UP isolates were classified according to their biofilm formation capacity after 72 h, following the Stepanovic criteria (Stepanovic et al., 2007). The A595nm cut-off value (A595nmC) was defined as the average value of the blank absorbance plus three standard deviations. The blank was the non-inoculated medium. Consequently, the following classification criteria were established: A595nm ≤ A595nmC, non-biofilm producers (–); A595nmC < A595nm ≤ 2 fold A595nmC, weak biofilm producers (+); 2 fold A595nmC < A595nm ≤ 4 fold A595nmC, moderate biofilm producers (++); 4 fold A595nmC < A595nm ≤ 8 fold A595nmC, strong biofilm producers (+++); and A595nm > 8 fold A595nmC, robust biofilm producers (++++).
Microscopy biofilm visualization
Selected isolates were grown under biofilm-forming conditions in 6-well polystyrene plates containing M63 medium. Each well was previously fitted with a sterile glass coverslip (provided by the microscopy facilities). After 72 h at 30°C, non-adherent cells were gently removed, and the coverslips were washed with distilled water and air-dried for 10 min. For confocal laser scanning microscopy (CLSM), biofilms were stained with 20 μM DAPI (Sigma) in 0.1 M Tris-HCl buffer (pH 8) for 10 min (in dark conditions), followed by distilled water washes. Samples were fixed with 4% paraformaldehyde (for 20 min), rinsed with PBS, and imaged using a Zeiss LSM800 microscope. For scanning electron microscopy (SEM), biofilms on coverslips were fixed with a 2.5% glutaraldehyde and 2.5% paraformaldehyde (v/v) solution, dehydrated through an acetone/ethanol series, and sputter-coated with gold using a JEOL JFC-1100 ion coater. Samples were then mounted on aluminum stubs and imaged with a Carl Zeiss SUPRA-55 SEM at resolutions of 1.0 nm (15 kV) and 1.7 nm (1 kV) in high-vacuum mode or 2 nm (30 kV) in variable-pressure mode.
Colony morphotypes: amyloid-like fiber and cellulose production
Colony morphology and dye-binding may serve as indicators of several physiological and metabolic states in microbes (Martin-Rodriguez et al., 2021). To analyze colony morphology, culture plates supplemented with Congo Red (CR, Cicarelli) and Brilliant Blue (BB, Sigma-Aldrich) were used, as described by Da Re and Ghigo (2006). Briefly, BHI cultures of all isolates were grown overnight (ON) at 37°C, washed, and diluted to an A600 nm of 0.1 in PS. Subsequently, 5 μL of these suspensions were spotted on LB-agar plates (low salt) supplemented with 40 μg/mL CR and 20 μg/mL BB. Plates were incubated at 30°C for 96 h and monitored every 24 h. Colony phenotype analysis was performed for each bacterial genus, as per previous reports. For E. coli and K. pneumoniae, the analysis was according to Bokranz et al. (2005): ras (red and smooth colonies) and pas (pink and smooth colonies) morphotypes. For Staphylococcus spp. isolates, the results were interpreted as reported by Arciola et al. (2002): reddish-black colonies with a rough and dry consistency were considered to be biofilm-related extracellular matrix-producing strains. The morphology of E. faecalis strains was analyzed as reported by Torres-Rodríguez et al. (2020), where strains with black and rough colonies are considered biofilm producers, and strains with red or white colonies are considered non-producers. For Bacillus spp., bacteria that bind CR dye have previously been classified as producers of functional bacterial amyloid fibers (Romero et al., 2010). The presence of amyloid-like fiber was denoted as “+.”
Cellulose production was assessed as described by White et al. (2006). For this, BHI cultures of each isolate were grown ON at 37°C, washed, and diluted to an A600 nm of 0.1 in PS. Subsequently, 5 μL of the isolates were spotted on LB-agar plates supplemented with 50 μg/mL Calcofluor White (CW, Sigma-Aldrich) dye. The plates were incubated at 30°C for 96 h and monitored every 24 h. Cellulose production was qualitatively assessed by observing the fluorescence of colonies when irradiated with UV light. Fluorescent colonies were denoted as +.
Motility assay
Bacterial motility was assessed according to Ulett et al. (2006), with minor modifications. Briefly, ON BHI cultures were washed and diluted to an A600 nm of 0.1 in PS. Suspensions were then seeded onto semi-solid LB-agar plates (0.3%) using a sterile toothpick. The plates were incubated at 30°C, and the colony diameter was evaluated over time. Isolates with colony diameters between 0.5 and 1 cm were denoted as “+” and those with diameters >1 cm as “++.”
Hemolytic capacity
The isolates' hemolytic activity was assessed on blood agar plates using the method described by Gerhardt et al. (1994). Briefly, isolates were grown ON in BHI medium, and then the cells were washed and diluted to an A600 nm of 0.1 in PS. Aliquots of the bacterial suspensions were plated on blood agar medium and incubated for 24 h at 30°C. A β-hemolytic reaction involves the complete lysis of red blood cells, resulting in a clear area on the agar surrounding the colony, known as total hemolysis. In contrast, an α-hemolytic reaction occurs when the hemoglobin in red blood cells is converted to methemoglobin, resulting in a greenish tint in the agar surrounding the colonies. Finally, the absence of hemolysis or discoloration is referred to as γ-hemolysis (Buxton, 2005).
Siderophore production assay
The siderophore production was determined using a qualitative technique adapted in our laboratory, based on the color change of Cromoazurol S (CAS, Sigma-Aldrich). Strains were cultured ON in M9 minimal medium (Sigma-Aldrich) supplemented with 0.2% glucose, with shaking at 37°C. Subsequently, 5 μL aliquots of each cell culture were seeded onto M9 minimal medium supplemented with 0.2% glucose and incubated at 37°C for 48 h. Then, an overlay of semisolid CAS medium (Cromoazurol S, 60.5 mg; piperazine acid, 72.9 mg; FeCl3, 1 mM; dissolved in HCl at 10 mM; 10 mL of this solution per 1 L) was poured onto the grown plates. Isolates capable of producing siderophores exhibited a color change in the CAS medium from blue to yellowish around the colony.
Hydrogen peroxide, acid, and human serum tolerance assays
Stressor susceptibility was carried out according to Shea et al. (2022). Briefly, bacterial cultures were incubated ON in BHI medium at 37°C. Cultures were normalized to 0.5 McFarland (~1.5 × 108 CFU mL−1) in 1 mL of either BHI, BHI containing fresh 0.2% H2O2, or BHI buffered to pH 7, 5, or 2.5. Samples were immediately vortexed and incubated either for 15 or 60 min at room temperature for the H2O2 tolerance assay or for 1 h at 37°C with aeration for the acid tolerance assay. At each time point, suspensions were serially diluted in PS to further determine CFU mL−1 in CLED-agar medium. For human serum tolerance, ON BHI cultures (1 mL) were pelleted by centrifugation, and 0.5 McFarland suspensions were prepared in sterile PS. The suspensions were diluted 1:200 in either 100% human serum or 100% heat-inactivated human serum (Sigma-Aldrich). The mixture was incubated for 1 h at 37°C, and then the number of CFU mL−1 was calculated by serially diluting the bacterial-serum suspension and plating it on CLED-agar medium. The CFU mL−1 of the bacterial inoculum was calculated by serially diluting the bacterial suspension and plating it on CLED-agar medium.
Antibiotic susceptibility
Antibiograms were performed for each UP using the disc diffusion method on Müller-Hinton agar medium (MH-agar, Britania; CLSI, 2020). The choice of antimicrobial agents is based on national and international recommendations for urinary infections in adults, covering both inpatient and outpatient settings (WHO, 2022). The antibiotics (ATB) used for Gram-negative and Gram-positive bacteria in this study were 10 μg norfloxacin (NOR), 10 μg gentamicin (GEN), 300 μg nitrofurantoín (NIT), 10/10 μg ampicillin/sulbactam (AMS), 1.25/23.75 μg trimethoprim/sulfamethoxazole (SXT), 30 μg nalidixic acid (NA), 5 μg ciprofloxacin (CIP),10 μg ampicillin (AMN), 30 μg amikacin (AKN), 30 μg ceftazidime (CAZ), 30 μg cefuroxime (CXM), and 10 μg imipenem (IMP). In addition, ATB specifically targeting Gram-positive cocci were also tested: 1 μg oxacillin (OXA), 15 μg erythromycin (ERY), clindamycin (CLI) 2 μg, 30 μg vancomycin (VAN), and 30 μg cefoxitin (FOX). Inhibition halos were measured and interpreted according to the Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2020). Multi-drug resistance (MDR) was defined as acquired non-susceptibility to at least one agent in three or more antimicrobial categories (Siegel et al., 2006).
Detection of genes encoding virulence factors
The presence or absence of virulence-associated genes was screened using PCR. For each bacterial genus, the following genes were tested: for E. coli, hemolysin (hlyA), cytotoxic necrotizing factor (cnf1), fimbria type I regulator (fimB), type 1 fimbria (fimA), fimbria P (papA) and iron uptake related genes (iroN and iutA); for K. pneumoniae, hemolysin (hlyA), type 1 fimbrial subunit (fimA), the type 3 fimbrial adhesin (mrkD), enterobactin (entB), and an outer membrane porin (ompK36); for E. faecalis, gelatinase (gelE), sortase-type enzyme (srt), bacterial adherence protein (efaA), collagen adhesin protein (ace), cytolicin production activator (cylA); for Staphylococcus spp., staphylococcal enterotoxin A (sea), slime production related protein (ica), arginine metabolism related protein (argB, argC), lipase (gehC), fibrinogen binding protein (sdrG) and for Bacillus spp., bacillus enterotoxins (entB, entA), pore-forming toxin (cytK), hemolysin (hlyIII), sporulation related -ATPase subunit (clpC), capsule synthesis protein (capA). PCR reactions were performed using the primers and parameters described in Supplementary Table S1. The PCR reaction mixture (25 μL total) consisted of 12.5 μL of 2 × SYBR Green PCR Master Mix (Bio-Rad), 300 nmol of each forward and reverse primer, and 10 ng of genomic DNA as the template. The amplification protocol consisted of an initial denaturation step at 95°C for 4 min, followed by 35 cycles of denaturation at 95°C for 15 s and annealing/extension at 56°C for 30 s. Fluorescence signals were recorded at the end of each extension step, and a melt curve analysis was performed to verify the specificity of the product. To confirm amplification, 5 μL of the PCR product was analyzed on a 2% agarose gel, with a 100 bp DNA ladder (Promega) included for size reference. Positive and negative controls were included, consisting of a 16S gene and a free-template tube, respectively.
Bacterial interaction assays: colony interaction, liquid medium co-culture, and mixed biofilm formation
Bacterial interaction was evaluated in co-isolated pairs obtained from the same catheter. Four co-isolated pairs were selected, consisting of Bacillus strains with other UP: E. faecalis Ef5/B. pumilus Bp1, S. epidermidis Se3/B. subtilis Bs1, S. epidermidis Se4/B. subtilis Bs2, and K. pneumoniae Kp2/B. megaterium Bm2. For colony interaction assays, a working protocol developed in the laboratory was followed. Briefly, bacteria were grown overnight (ON) at 37°C in a BHI liquid medium with shaking (150 rpm). Then, 5 μL aliquots of the cultures were taken and placed on the surface of a solid BHI-agar medium plate, following the scheme of one central bacterium and four concentric bacteria at different distances. Bacteria (central or peripheral) were seeded simultaneously or in a deferred manner (24 h after the growth of the central colony). The interaction was analyzed qualitatively over a period of 2 days at 30°C, and changes in colony aspects, such as growth, size, and morphology, were documented. For liquid media co-culture assays, a previously described protocol was carried out (Juarez and Galvan, 2018; Learman et al., 2019; Gaston et al., 2020). Briefly, isolates were grown ON at 37°C in BHI, harvested by centrifugation, and adequately diluted in BHI medium to an A600 nm of 0.1. Monospecies or mixed cultures were grown under shaking conditions at 37°C for varying times, and viability was measured using appropriate media and ATB to facilitate single-species identification in the interaction. For mono- or multispecies biofilm assays, isolates were grown at 37°C in BHI and diluted to an A600 nm of 0.1. Cells of different bacteria (alone or in co-culture) were grown in multiwell plates during 24 h at 30°C. After incubation under static conditions, the non-adherent fraction (planktonic cells) was removed, and total biofilm biomass was quantified using the crystal violet technique (O'Toole and Kolter, 1998). The non-adherent fraction containing a mixed culture was quantified by CFU mL−1 using the appropriate media, as mentioned earlier. To quantify microorganisms in the biofilm biomass, adhered cells were washed three times with sterile PS and incubated with 0.1% Triton X-100 for 10 min. Biofilm cells were extracted by scraping vigorously with a sterile tip. These bacteria were resuspended in sterile PS and quantified by determining CFU mL−1 using appropriate media, as mentioned before (Juarez and Galvan, 2018).
Statistical analysis
Statistical analyses were performed using R (version 4.4.1) and RStudio (version 2024.09.0+375). A significance level of α = 0.05 was set for all inferential tests. Prior to applying parametric tests, the assumptions of normality and homogeneity of variances (homoscedasticity) were assessed using the Shapiro–Wilk test and Levene's test, respectively. When these assumptions were violated, such as with microbial counts spanning several orders of magnitude and exhibiting heteroscedasticity, a logarithmic transformation [log(x+1)] was applied to stabilize variances and improve normality. A two-way analysis of variance (ANOVA) was conducted to evaluate the effects of the independent variables and their interaction with the response variable. Model residuals were examined to verify the assumptions of normality and homoscedasticity required for the analysis of variance (ANOVA). When significant effects were detected, Fisher's Least Significant Difference (LSD) post-hoc test was used for pairwise group comparisons. To explore multivariate patterns and identify potential groupings among bacterial genera, a principal component analysis (PCA) was performed. The phenotypic traits of the bacterial genera were included as variables. All variables were standardized, and PCA was conducted using a correlation matrix to account for differences in measurement scales. A biplot based on the first two principal components was generated to visualize the distribution of observations and the contribution of variables to the principal components, represented by the PCA loadings. Clusters were visually identified and characterized using ellipses, and the within-cluster sum of squares (WSS) was calculated to assess the compactness of each cluster.
Results and discussion
Clinical isolates from double-J catheters reveal polymicrobial colonization
A total of 27 bacteria were isolated from 8 catheters (Table 1). Monobacterial isolates were obtained from three catheters (catheters 1, 4, and 5), whereas multiple bacterial species were recovered from the remaining catheters, confirming the polymicrobial etiology of the double-J stents (Chatterjee et al., 2014; Klis et al., 2014; Wang et al., 2021). The predominant bacterial species were Bacillus spp., including B. pumilus, B. subtilis, and B. megaterium (7 isolates, 27%), followed by Staphylococcus spp., including S. aureus and S. epidermidis (1 and 6 isolates, respectively, 27%). E. faecalis (5 isolates, 19%), K. pneumoniae (4 isolates, 15%), and E. coli (3 isolates, 12%) were also identified (Table 1). All patients had their catheters in place for a period of 1–3 months and received ATB therapy prior to extraction. Despite negative urine cultures (UC), both adhered and non-adhered bacteria were isolated from all devices. This finding aligns with several reports indicating that colonization of double-J catheters does not correlate linearly with urine culture (UC) or prostatic secretion samples (Klis et al., 2009). The identified species (except for Bacillus spp.) are in agreement with prior literature concerning bacterial species in catheters, which describes them as common urinary tract infection (UTI) pathogens (Gould et al., 2010; Kart et al., 2017; Pérez et al., 2017; Al-Qahtani et al., 2019). Although only 12% of the isolates were UPEC, this bacterium is the most common pathogen in both community-acquired and nosocomial urinary tract infections, accounting for approximately 24%−39% of catheter-associated urinary tract infections (CAUTIs; Jacobsen et al., 2008). It is widely known that K. pneumoniae is an opportunistic pathogen commonly distributed in the perineum, which can easily colonize the urinary tract and medical devices (Pérez et al., 2017). E. faecalis establishes a symbiotic relationship with other UP and is frequently found in the context of CAUTIs (Salm et al., 2023). Additionally, S. aureus and S. epidermidis, which are normally present on the skin surface, can enter the urinary tract during catheter placement (Klis et al., 2014; Badhan et al., 2016; Oliveira et al., 2018). Often, the cause of this colonization may be that catheterization and its manipulation promote ascending infections due to the presence of commensal microorganisms in the surrounding skin of the urethra or potential contamination resulting from the manipulation of healthcare personnel (Kalsi et al., 2003; Chatterjee et al., 2014). Notably, several bacterial isolates adhered to different catheters belonging to the Bacillus genus. It is well-known that human infections caused by Bacillus spp., except for B. anthracis or B. cereus, are rarely reported in the literature. The primary reason for this can be attributed to the fact that most of these bacteria are often considered laboratory contaminants with little to no clinical relevance (Drobniewski, 1993; Fekete, 2015). However, some reports described that Bacillus spp. has been isolated from bacteremia, endocarditis, wounds, respiratory, urinary, and gastrointestinal tract infections, food poisoning, and meningitis (Drobniewski, 1993; Fekete, 2015). Therefore, the identification of aerobic Gram-positive spore-forming bacilli at the species level, the evaluation of their pathogenic potential, and the interpretation of susceptibility ATB tests for these bacilli would be relevant in elucidating their potential role in the mentioned infections (Klis et al., 2014; Kandi et al., 2016).
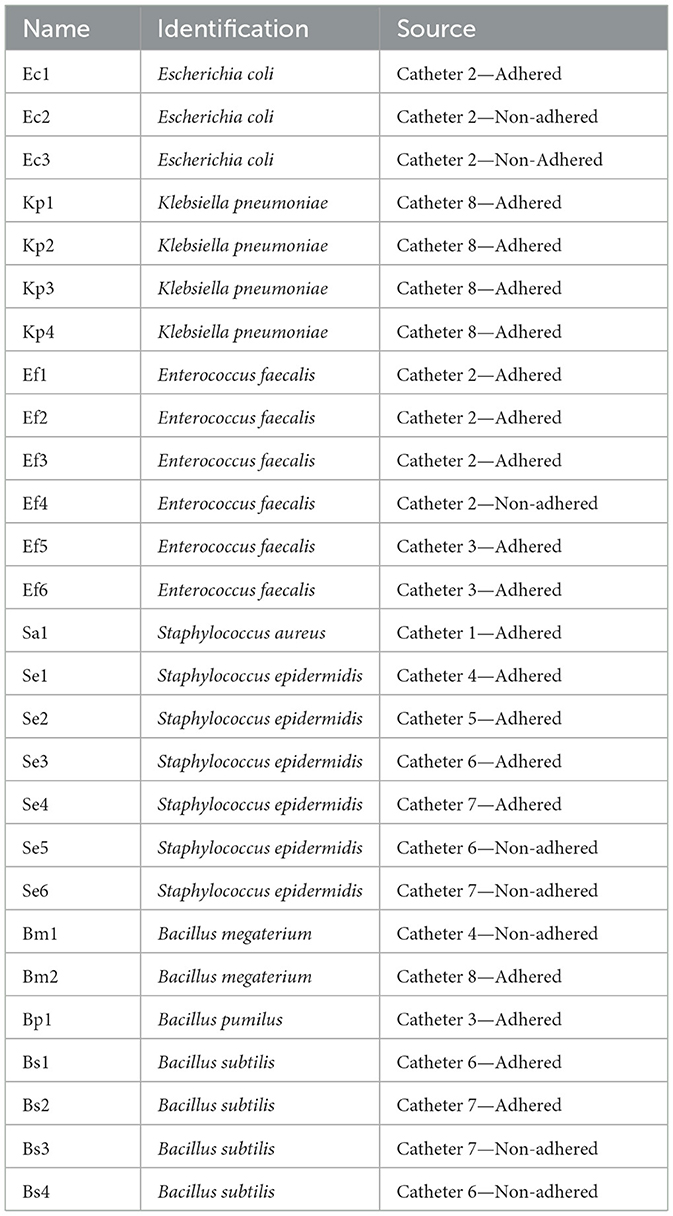
Table 1. Clinical isolates obtained from double-J catheters.
Our data showed a high percentage of bacterial catheter colonization. Several authors have reported that the catheter colonization rate exceeds the urinary infection rate, indicating a significant inconsistency between urinary infections and catheter colonization. This discrepancy complicates the estimation of stent colonization (Kehinde et al., 2004; Klis et al., 2009). Therefore, a thorough study should be conducted regarding the prevalence of etiological agents and their antimicrobial susceptibility in double-J catheters to improve treatments for long-term catheterized patients.
Clinical isolates exhibit multiple virulence-associated phenotypes that support potential catheter colonization
Measuring experimental phenotypic outcomes could predict the UP potential infectivity. To understand the virulence strategies of the isolated strains, we performed an extensive set of phenotypic microbiological assays, which included evaluating biofilm formation capacity, amyloid-type fiber production, cellulose, and mucoid substance production as components of the extracellular matrix, motility, siderophore production, and hemolytic capacity, along with their ATB resistance profile. These phenotypes are summarized in Table 2 and described below.
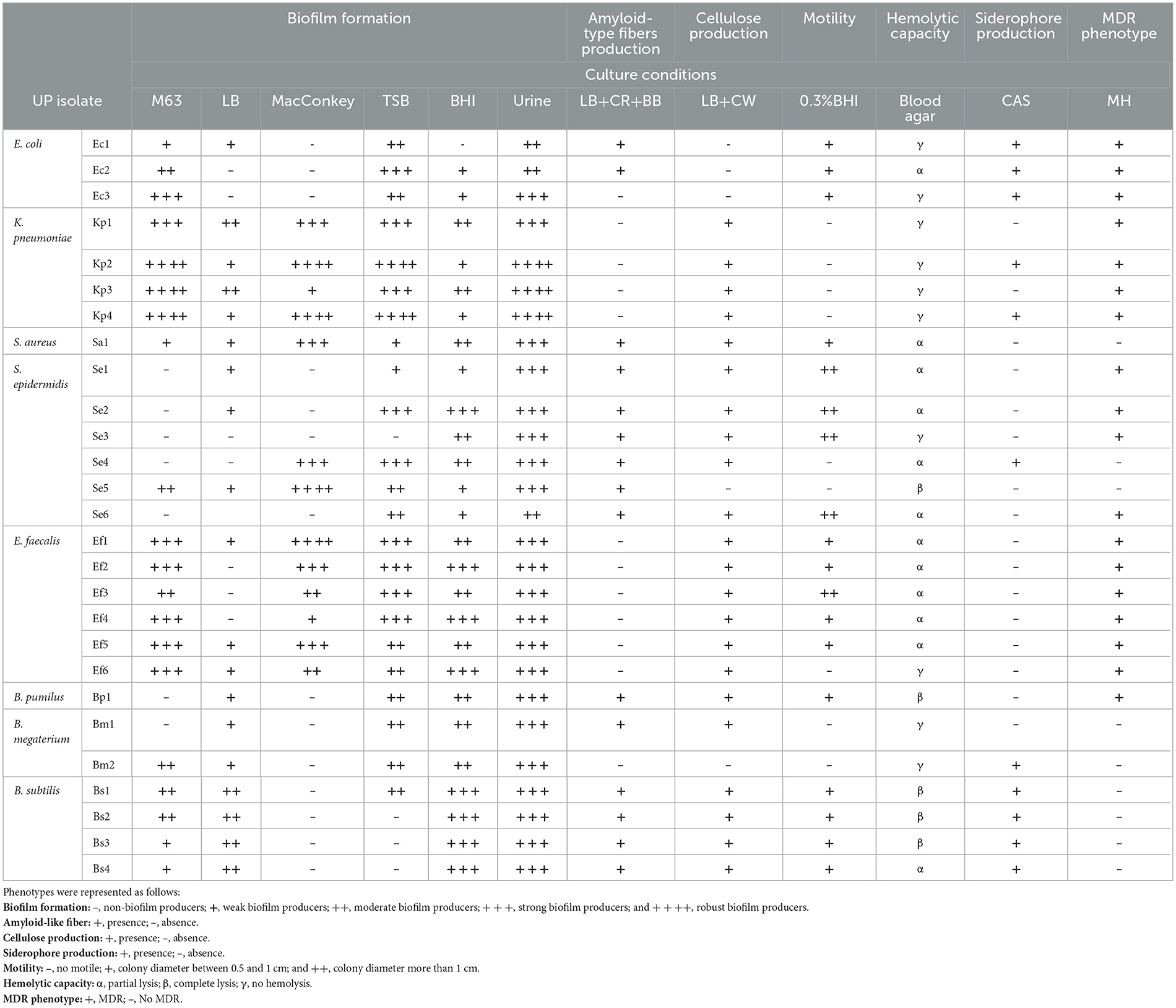
Table 2. Virulence associated-phenotypes.
Biofilm formation
Since biofilm development is influenced by environmental factors, such as nutrient availability, surface type, and shear forces, assessing this trait in multiple growth media will allow us to infer the versatility of bacterial populations and their potential to form biofilms in various clinical contexts. Biofilm formation was tested in M63, MacConkey, BHI, TSB, and urine culture media at various time points. Results displayed in Table 2 show that the biofilm formation ability of E. coli, S. epidermidis, and Bacillus spp. strains varied depending on the culture medium used, being higher in TSB, BHI, and urine medium. S. aureus, E. faecalis, and the majority of the K. peumoniae isolates were considered strong or robust biofilm formers in all tested media. Differences in the ability to form biofilm among genera and conditions are typical in clinical isolates (Grillo-Puertas et al., 2015; Del Pozo, 2018; Uruen et al., 2020). It is well-known that biofilm formation is one of the most relevant virulence mechanisms used by UP during UTIs (Del Pozo, 2018; Klein and Hultgren, 2020; Lenchenko et al., 2020). Indeed, it has been reported that microorganisms living in these structures are even more virulent and resistant to ATB. Therefore, the ability of UP to form biofilms in urinary catheters and the urothelium is a crucial factor for the persistence and recurrence of UTI (Vuotto et al., 2017). Additionally, the release of bacteria from biofilms into the bloodstream can lead to widespread infections, especially in immunocompromised patients (Saint and Chenoweth, 2003; Flemming and Wingender, 2010; Lazar and Chifiriuc, 2010). The description of the isolates' biofilm-forming ability is insufficient to classify them as non-biofilm formers, as in other circumstances, such as certain clinical settings, they may have a greater capacity for adhesion and biofilm formation. Studying biofilm formation under different in vitro conditions provides valuable insights into the adaptability and persistence strategies of clinical isolates. Such variability may reflect the ability of these organisms to persist on medical devices or within host niches, even under fluctuating conditions. In addition to visualizing the biofilm structure, qualitative analysis was performed using confocal laser scanning microscopy (CLSM) and scanning electron microscopy (SEM) on four selected UP isolates in BHI medium (Supplementary Figures S1, S2). Results showed that S. aureus Sa1, E. coli Ec1, K. pneumoniae Kp2, and E. faecalis Ef5 strains were able to adhere to the glass surface. Although the surface coverage of Sa1 and Ec1 was relatively lower compared to Kp2, all these strains demonstrated a strong adhesion, presenting characteristic agglomerates. The biofilm surface of the Sa1, Ec1, and Kp2 strains showed a “Christmas tree forest” appearance with internal water channels. These structures, characterized by their towering growth and interconnected channels, allow for efficient exchange of nutrients and waste, enabling the biofilm to sustain itself over time (Quan et al., 2022). It is worth mentioning that the Ef5 isolate was not able to form a structured biofilm. Thus, it was not possible to reconstruct the 3D structure. In SEM assays (Supplementary Figure S2), the Ec1 strain showed a significant coverage of the glass surface. In addition, a delicate layer encapsulating groups of cells interspersed by channels was observed (indicated by an arrow in the insert), probably indicating the presence of an extracellular matrix, as previously described for this genus (Cui et al., 2020). In the Ef5 isolate, a large number of cell clusters surrounded by abundant extracellular material were found. Finally, the Kp2 isolate formed a strong biofilm with protrusions that rose from the surface, as observed by CLSM. Pili, curli, or nanotube-like structures were observed, with abundant extracellular matrix surrounding the bacteria (arrows in inserts of Supplementary Figure S2). These appendages have been reported to play essential roles during cell adhesion and biofilm formation (Busscher et al., 2008; Rodrigues and Elimelech, 2009). Regarding the Sa1 isolate, it was unable to form a structured biofilm in this assay, as dispersed cells were observed (Supplementary Figure S2). The discrepancies observed in the biofilm formation ability among the different isolates on polystyrene and glass surfaces were not surprising since surface topography is a parameter that significantly influences microbial adhesion (El Abed et al., 2012).
Colony morphotypes
The assessment of colonies' morphology and dye-binding capacity can be used to describe, identify, and characterize microorganisms, serving as an indicator of microbial physiology and metabolism. Binding to CR and BB dyes, as well as the interpretation of the obtained results, is specific to each bacterial genus. Table 2, Supplementary Figure S3 show the phenotypes of bacterial colonies obtained on LB plates supplemented with CR and Brilliant Blue. When curli or other amyloid-like fibers were studied, it was observed that only Escherichia, Staphylococcus, and Bacillus exhibited a colony morphotype characteristic of fiber or adhesion-type component production. Based on the morphotype classification, E. coli isolates showed a ras phenotype (Supplementary Figure S3), indicating moderate production of fimbriae curli and cellulose. S. epidermidis isolates Se1, Se2, Se3, and Se6 exhibited clear, flat, circular colonies with rough edges at 24 h, which turned red, wrinkled, and irregularly shaped by 96 h (Supplementary Figure S3). According to the colony morphotype, Staphylococcus spp. isolates were considered as matrix producers (Arciola et al., 2002). In contrast, Bacillus isolates Bp1, Bs1, Bs2, Bs3, and Bs4 exhibited dark or red colonies with central black halos and rough, irregular, and lobed margins (Supplementary Figure S3). This morphotype indicates the production of TasA proteins (Romero et al., 2010), which are the major amyloid-type proteins of the biofilm matrix in Bacillus spp. (Chapman et al., 2002; Branda et al., 2006; Bohning et al., 2022). K. pneumoniae or E. faecalis strains were characterized by white colonies with pink or reddish centers and a mucoid appearance (Supplementary Figure S3). The distinctive mucoid morphology of K. pneumoniae isolates indicated a high capsule production (Table 2). This phenotype was reported as one of the main virulence factors of K. pneumoniae involved in adhesion to host cells and biofilm formation (Vernet et al., 1995; Cavalcanti et al., 2019). The presence of a pigmented ring that stands out in the Ef5 isolate was consistent with biofilm formation capacity (Torres-Rodríguez et al., 2020).
Cellulose production is another relevant extracellular matrix component for biofilms and several virulence traits, as it provides an important physical barrier to antimicrobials (Matz et al., 2008; Mulcahy et al., 2008; Hansen and Vogel, 2011). In addition to the observed RC colony morphotypes, cellulose production was also visualized on LB-agar plates supplemented with CW, as described in the Materials and Methods section. As shown in Table 2, except for E. coli, all bacterial genera produced cellulose, as evidenced by the formation of fluorescent colonies when they were irradiated with UV light (Supplementary Figure S3). No fluorescence was observed in E. coli colonies after irradiation; only the Ec3 isolate showed slight fluorescence in the center of its colony, as expected for its ras morphotype.
Colony morphology depends on numerous environmental factors, such as media composition, temperature, and humidity, among others, which lead to the expression of specific genes or intercellular communication processes that control this process (Bokranz et al., 2005; Da Re and Ghigo, 2006; Kaiser et al., 2013; Sydow et al., 2022). Analysis of these components under various conditions could determine the potential high capacity of the isolates to form a biofilm in UTIs. Although the correlation between morphology, expression of extracellular components, and biofilm formation capacity has been noted in some bacterial isolates (i.e., B. subtilis and S. epidermidis strains), it has not been generalized across all bacterial genera. Although K. pneumoniae and E. faecalis strains exhibited a strong biofilm-forming ability, they did not produce amyloid-type fibers under the tested conditions (Table 2).
Bacterial motility
In some pathogenic bacteria, motility plays a crucial role in the initial phase of infection (Josenhans and Suerbaum, 2002), likely to overcome the electrostatic repulsion between cells and surfaces (Pratt and Kolter, 1998; Walker et al., 2007). In addition, motility contributes to the colonization of different environmental niches by facilitating the spread of the infectious agent, which is crucial in the context of UTI (Josenhans and Suerbaum, 2002). As observed in Table 2, all isolates exhibited significant motility capacity, except for K. pneumoniae, S. epidermidis strains Se4 and Se5, B. megaterium, and E. faecalis strain Ef6. Although both S. aureus and S. epidermidis lack flagella and, therefore, are considered non-motile bacteria, it was observed that Sa1, Se1, Se2, and Se3 isolates exhibited significant motility. Recent studies have reported flagellum-independent forms of motility in microorganisms of this genus (Pollitt and Diggle, 2017). For instance, spreading involves a sliding movement, where bacteria spread radially outward from an inoculation site, forming multiple layers of densely packed cells (Henrichsen, 1997; Kaito and Sekimizu, 2007). E. faecalis is also regarded as a non-motile bacterium due to the absence of flagella. However, in this study, most E. faecalis isolates showed motility, with the exception of the Ef6 strain. It demonstrated an E. faecalis migration mechanism, where the synthesis and secretion of extracellular polysaccharides were required (Ramos and Morales, 2019). Therefore, when cells are grown on semisolid agar, they penetrate and invade the medium, creating a “colony impression” (Ramos and Morales, 2019). This mechanism is relevant for translocation through monolayers of human epithelial cells, conferring adaptive advantages during infection, as it enables them to translocate from the urinary tract to the bloodstream and colonize distant anatomical sites (Muscholl-Silberhorn et al., 2000; Ubeda et al., 2010; van der Heijden et al., 2014). In general, it has been described that environmental isolates of B. subtilis exhibit robust motility (Kearns and Losick, 2003), in agreement with the exacerbated motility shown by the strains used in this study. There is a complex relationship between motility and biofilm formation, which depends on environmental conditions or bacterial requirements. A study conducted on B. cereus strains reported that bacterial motility influenced biofilm formation through three mechanisms: (1) motility is necessary for the bacteria to reach adequate surfaces to form biofilm; (2) motility promotes the recruitment of planktonic cells to invade the preformed biofilm; and (3) motility is involved in biofilm spreading and propagation (Houry et al., 2010). Based on the observed motility of B. subtilis, it could be assumed that motility is necessary for the bacteria to reach adequate surfaces to form biofilm (Dunne, 2002; Grillo-Puertas et al., 2015).
Siderophore production
Iron, a vital nutrient required for bacterial growth, is highly restricted within human hosts (Hood and Skaar, 2012; Subashchandrabose and Mobley, 2015). Most UP strains encode several iron acquisition systems, such as siderophores, to acquire iron sequestered by the host (Garcia et al., 2011). Here, we tested siderophore production by the studied clinical isolates using the CAS assay described previously. All E. coli and B. subtilis strains, including Kp2 and Kp4, Se4, and Bm1, were able to produce siderophores (Table 2, Supplementary Figure S4A). It was reported that siderophore production by different UP is an important characteristic that contributes to the potential virulence of these isolates, relevant in the catheter context (Danese et al., 2000; Yue et al., 2003; Zou et al., 2023).
Hemolytic capacity
Hemolysins are proteins secreted by bacteria that constitute another important virulence factor in UTIs. These toxins can disrupt host cell signaling cascades, altering the inflammatory response and inducing cell death (Guyer et al., 2002; Wiles et al., 2008). It is well-known that α-hemolysin, HlyA, can stimulate epithelial barrier rupture, leading to bacterial translocation from the urinary tract to the bloodstream (Whelan et al., 2023). Table 2 shows that 19.2% of the isolates exhibited total hemolysis (β hemolysis) with defined lysis zones (Supplementary Figure S4B). Partial hemolysis (α hemolysis) was carried out by 42.3% of the isolates, while the remaining 38.5% did not present hemolytic capacity. As observed in Table 2, Staphylococcus strains exhibited partial or total hemolysis capacity. It is well-known that hemolytic capacity is one of the most common virulence factors of coagulase-positive (S. aureus) and coagulase-negative (S. epidermidis) staphylococci (Moraveji et al., 2014). In E. faecalis isolates, only partial hemolysis was observed, in agreement with several studies that describe this genus as not being able to produce total hemolysis (Stepien-Pysniak et al., 2019; Torres-Rodríguez et al., 2020). It is worth mentioning that four out of the five isolates exhibiting total hemolysis belong to the Bacillus spp. genus. Similar results were obtained with Bacillus strains isolated from river water samples with fecal contamination, where all isolates presented total hemolytic capacity (Ostensvik et al., 2004). The pathogenicity of Bacillus spp. has been poorly investigated, except for B. cereus, a known pathogen whose pathogenic potential has been related to the secretion of several virulence proteins, such as hemolysins phospholipases, cytotoxin K (CytK) and proteases (Ramarao and Sanchis, 2013; Jessberger et al., 2015), and diverse motility factors, such as those involve in swimming and swarming (Senesi et al., 2010).
Taken together, the ability to form biofilms and produce extracellular components, such as capsules and cellulose, along with virulence factors like hemolysins and siderophores, are critical elements in the pathogenicity of UP associated with colonization of double-J catheters. These factors not only facilitate the adhesion and colonization of devices but also confer resistance to antimicrobial treatments, complicating the resolution of infections.
Double-J catheter-associated isolates exhibit prevalence of MDR phenotype
Knowledge of the ATB susceptibility (AS) pattern of clinical isolates is necessary to characterize the microorganism, as it provides valuable context for understanding their clinical relevance and potential treatment challenges. Therefore, antibiograms using conventional ATB were performed for each isolate (Supplementary Table S2). Results in Table 2 show that E. coli and K. pneumoniae isolates exhibited a multidrug-resistant (MDR) phenotype. E. coli strains were resistant to 6 out of 12 antibiotics tested, with K. pneumoniae isolates susceptible only to imipenem and amikacin (Supplementary Table S2). Both species have frequently been reported to carry plasmids encoding extended-spectrum β-lactamases (ESBLs), which confer resistance to third-generation cephalosporins and other antibiotics of the β-lactam class (Paterson, 2006; Garau, 2008; Chen et al., 2013; Pendleton et al., 2013; Mazzariol et al., 2017). Although ESBL production was not specifically tested in our study, resistance to second- and third-generation cephalosporins supports this possibility. The preserved susceptibility to imipenem suggests that carbapenems may remain effective despite increasing reports of carbapenem-resistant strains (Logan and Weinstein, 2017). Regarding the Staphylococcus genus, only Se1, Se2, Se3, and Se6 showed an MDR phenotype (Table 2). Multi-drug-resistant S. epidermidis is increasingly recognized as a cause of opportunistic infections, particularly in patients with indwelling medical devices (Otto, 2009). These infections can be difficult to treat due to biofilm formation and the ability of S. epidermidis to acquire resistance genes through horizontal gene transfer. Our results are in agreement with those reported by Socohou et al. (2020), who found that S. epidermidis strains isolated from the surfaces of medical materials exhibited high susceptibility to gentamicin and ciprofloxacin. Regarding E. faecalis strains, all isolates exhibited a multidrug-resistant (MDR) profile (Supplementary Table S2). The observed resistance of E. faecalis isolates to ERY, CLI, OXA, and FOX is clinically concerning, particularly in hospital settings. Notably, two strains (Ef5 and Ef6) exhibited resistance to vancomycin (VAN) and erythromycin (ERY), indicating a vancomycin-resistant enterococci (VRE) phenotype. VRE infections are increasingly reported in catheterized patients and raise a serious therapeutic challenge due to limited treatment options (Rivera and Boucher, 2011; Arias and Murray, 2012; Dalhoff, 2012; Sievert et al., 2013). Resistance to macrolides and lincosamides is particularly problematic in patients allergic to β-lactams, as it restricts alternative therapeutic options. Moreover, the persistence and spread of such resistant strains increase the risk of hospital outbreaks that are difficult to control.
Results show that Bacillus spp. presented limited resistance to the tested ATB. Particularly, only the Bp1 strain exhibited resistance to several antimicrobials, including CAZ, ERY, CLI, OXA, and FOX, while the Bm1 and Bs3 strains were resistant only to AMP. Similar results were found in B. pumilus and B. subtilis by Adamski et al. (2023). Additionally, since most Bacillus species are not considered human pathogens but rather saprophytic skin microorganisms, antimicrobial susceptibility studies are typically not performed for this genus (Kalsi et al., 2003; Chatterjee et al., 2014). As environmental organisms, Bacillus are not frequently subjected to the selective pressures found in hospital settings, where the continuous use of ATB often leads to the development of resistance (Stenfors Arnesen et al., 2008). This limited exposure may contribute to the observed lower rates of resistance compared to other pathogens commonly encountered in healthcare environments.
Genotypic analysis evidences the presence of key virulence determinants among clinical isolates
The ability of UP isolates to cause different diseases is mediated by multiple virulence factors, including the expression of genes associated with the production of adhesins, toxins, siderophores, and secretion systems, among others (Blanco et al., 2002; Johnson, 2002). These factors are involved in the colonization of specific host surfaces, evading immune defenses, or causing direct damage to cells and tissues (Johnson, 2002). A genotypic analysis of some virulence-associated genes facilitates the identification of different phenotypic traits that may contribute to the persistence and severity of infections. E. coli isolates harbored genes encoding iron acquisition systems (iutA and iroN) and the hemolysin-encoding gene hlyA. Additionally, the Pap fimbriae (papA) and cytotoxic necrotizing factor (cnf-1) genes were detected in all isolates, suggesting a highly virulent phenotype. The iutA and iroN genes are known to enhance bacterial survival by facilitating iron acquisition in iron-limited environments, such as the human body (Johnson et al., 2005). Kanamaru et al. (2003) reported the prevalence of putative uropathogenic virulence factors in 427 E. coli strains, finding a significant prevalence rate of the iroN gene in the UTI isolates. The presence of hlyA and cnf-1 further suggests that these isolates have the potential to cause severe infections, as both genes are associated with tissue damage and inflammatory responses, as typically observed in UPEC (Rendon et al., 2007; Totsika et al., 2012). Soto et al. (2007) reported that the papA and hlyA genes were present in 42% and 36% of the UPEC strains analyzed, respectively. It is worth mentioning that 50% of UPEC produce α-hemolysin, and its expression is strongly associated with symptomatic UTIs (Pigrau, 2013).
All K. pneumoniae isolates were positive for all virulence genes tested (Table 3), indicating their potential to cause infections by adhering to host tissues and evading the immune system. OmpK36 expression has been linked to ATB resistance (Wyres et al., 2020), consistent with the MDR profile previously described for this genus (Table 2, Supplementary Table S2). Additionally, the combination of adhesion factors and hemolysin suggests that these strains may be particularly adept at establishing infections, especially in immunocompromised patients (Paczosa and Mecsas, 2016).
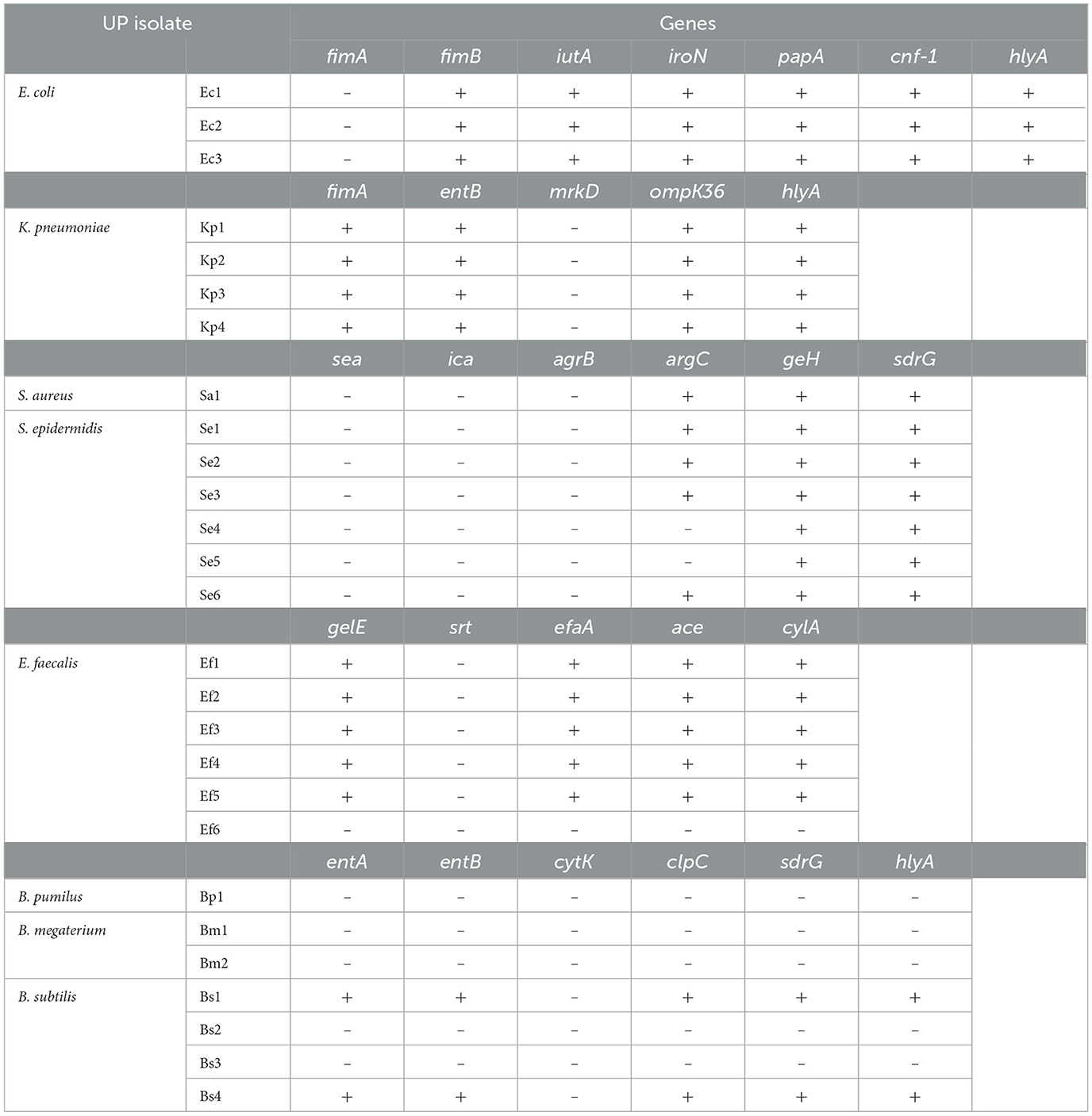
Table 3. Presence of virulence factor coding genes.
The S. aureus isolates exhibited virulence genes associated with the accessory gene regulator (agr) system, including argC and geH. This system is involved in the expression of toxins and surface proteins that aid in immune evasion and tissue invasion (Novick and Geisinger, 2008). The agr system is a well-documented key virulence regulator in S. aureus, controlling the production of toxins, such as hemolysins and proteases (Novick and Geisinger, 2008). Additionally, the sdrG gene, associated with fibrinogen-binding proteins that contribute to biofilm formation and immune evasion, was observed in this isolate. The presence of this gene indicates an enhanced biofilm-forming capacity, exhibiting strong resistance to immune clearance and ATB treatment (Kong et al., 2016). S. epidermidis isolates also contained the agrC, geH, and sdrG genes, all of which are associated with biofilm formation (Kavanaugh and Horswill, 2016). This ability enhances the survival of S. epidermidis on indwelling medical devices, such as catheters and prosthetic joints (Otto, 2009). The presence of these virulence factors suggests that these S. epidermidis isolates may be well-adapted and capable of causing persistent infections in healthcare settings.
Except for Ef6, all E. faecalis isolates were positive for gelE, efaA, ace, and cylA genes. The presence of these virulence factors highlights the pathogenic potential of E. faecalis. Gelatinase, encoded by the gelE gene, is a protease that degrades host tissues and promotes biofilm development by facilitating bacterial adhesion (Hancock and Perego, 2004). The Ace protein is involved in adhesion to extracellular matrix proteins and plays a crucial role in host tissue colonization and the formation of biofilms on medical devices (Singh et al., 2010). EfaA is also involved in bacterial adherence and is considered essential for biofilm formation during infection (Nallapareddy et al., 2005). The expression of these genes is consistent with the considerable biofilm formation observed in these isolates. Finally, cytolysin (cylA) is involved in the lysis of red and white blood cells, contributing to immune evasion and tissue destruction (Hancock and Gilmore, 2006). This result is in agreement with the partial hemolytic capacity exhibited by these isolates.
Among the analyzed Bacillus species, only B. subtilis (Bs1 and Bs4) exhibited hlyA and sdrG genes. This was unexpected since Bacillus spp. are generally regarded as environmental bacteria with limited clinical relevance (Turnbull, 1996). However, recent studies by Bianco et al. (2020) and Fayanju et al. (2024) identified specific virulence genes in B. subtilis strains that contribute to their pathogenicity. In addition to virulence factors, the antimicrobial resistance profiles of Bacillus strains play a critical role in determining their pathogenicity and potential impact on indwelling device contexts.
Clinical isolates display differential survival under host-mimicking stress conditions
In the urinary tract, bacteria are exposed to various host-mediated stress responses, including osmotic stress, pH changes, reactive oxygen species (ROS) generated by the immune system, and nutrient limitation (Agace et al., 1995). Therefore, pathogens develop adaptive advantages to cope with these environmental stressors.
All isolates were exposed to pH levels of 7, 5, and 2.5 for 2 h to simulate the expected pH ranges that bacteria may encounter in the gut, urine, or during neutrophil attack, respectively (Figure 1). The results showed that both Gram-positive and Gram-negative isolates exhibited significant decreases in viability when exposed to pH 2.5. Only Ec2, Sa1, Bs1, and Bs4 strains were tolerant to all tested pH conditions (Figure 1A). The decreased viability when exposed to pH 2.5 is consistent with previous findings that show that acidic conditions can severely compromise bacterial membrane integrity and metabolic function in both bacterial genera (Cotter and Hill, 2003; Foster, 2004).
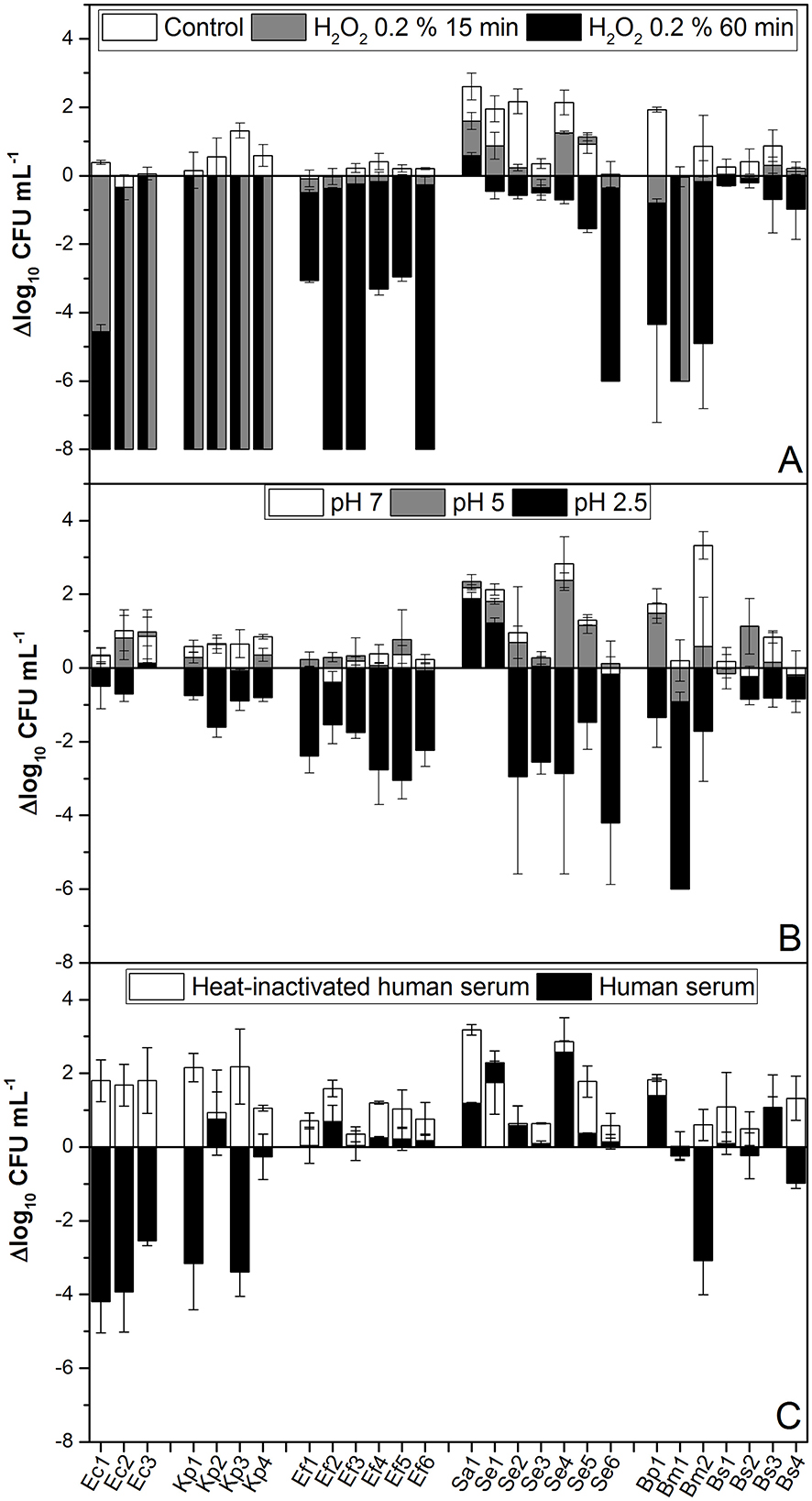
Figure 1. pH, H2O2, and human serum tolerance assays. (A) A total of 108 CFU mL−1 of each isolate was inoculated into BHI containing 0.2% H2O2 and maintained statically at room temperature. Samples were taken and serially diluted at 0, 15, and 60 min. (B) From an overnight culture, the isolates were diluted to 108 CFU mL−1 in BHI buffered to pH 7 (control), 5, or 2.5, as indicated. Cultures were incubated for 2 h at 37°C with aeration. (C) Isolates were cultured overnight in BHI, washed, and then resuspended in PBS. Then, 108 CFU mL−1 was added to 100% complete human serum and heat-inactivated human serum. Samples were incubated statically at 37°C for 60 min. After incubation, the CFU mL−1 was determined. Data represent the mean ± SD of at least four independent experiments. A one-way ANOVA was performed with the Fisher test, yielding a p-value of 0.05.
To study oxidative stress, the survival of the tested isolates exposed to H2O2 was evaluated. E. coli and K. pneumoniae strains (both Gram-negative bacteria) were highly sensitive to 0.2% H2O2 even after 15 min (Figure 1B). However, Gram-positive bacteria were more tolerant to the mentioned stress, displaying variable susceptibility when exposed for up to 1 h (Figure 1B). The higher resistance in Gram-positive isolates could be due to differences in their cell wall structures and antioxidant defense systems (Imlay, 2013).
Complement-mediated killing is also a crucial innate immune defense that can be assessed by measuring bacterial survival in human serum. Only E. coli isolates Kp1, Kp3, and Se5 strains were partially susceptible to serum (Figure 1C). This is in agreement with the fact that, although the immune pressure of serum complement, all virulent pathogens capable of inducing active infections have evolved immune evasive strategies that primarily target the complement system (Sharma et al., 2020).
The significant resistance to exogenous stressors observed in many isolates implies that these bacteria may have a high survival rate during infections, potentially leading to chronic or recurrent infection.
Principal component analysis of phenotypic diversity
The Principal Components Analysis (PCA) plot provides a visual summary of the relationships between five different bacterial genera (color- and shape-coded by taxonomic group) based on eleven phenotypic traits: antimicrobial resistance, biofilm formation, hemolytic capacity, motility, siderophore production, virulence factor coding genes, curli production, cellulose production and H2O2, pH 2.5, and human serum tolerances (Figure 2). The variable “pH 5 resistance” was excluded from the PCA since all bacteria were tolerant (constant value). The X-axis (PC1) explained 27% of the variance, while the Y-axis (PC2) explained 23%, together accounting for 50% of the total variance, which allowed an adequate interpretation of the characteristics associated with virulence (Figure 2).
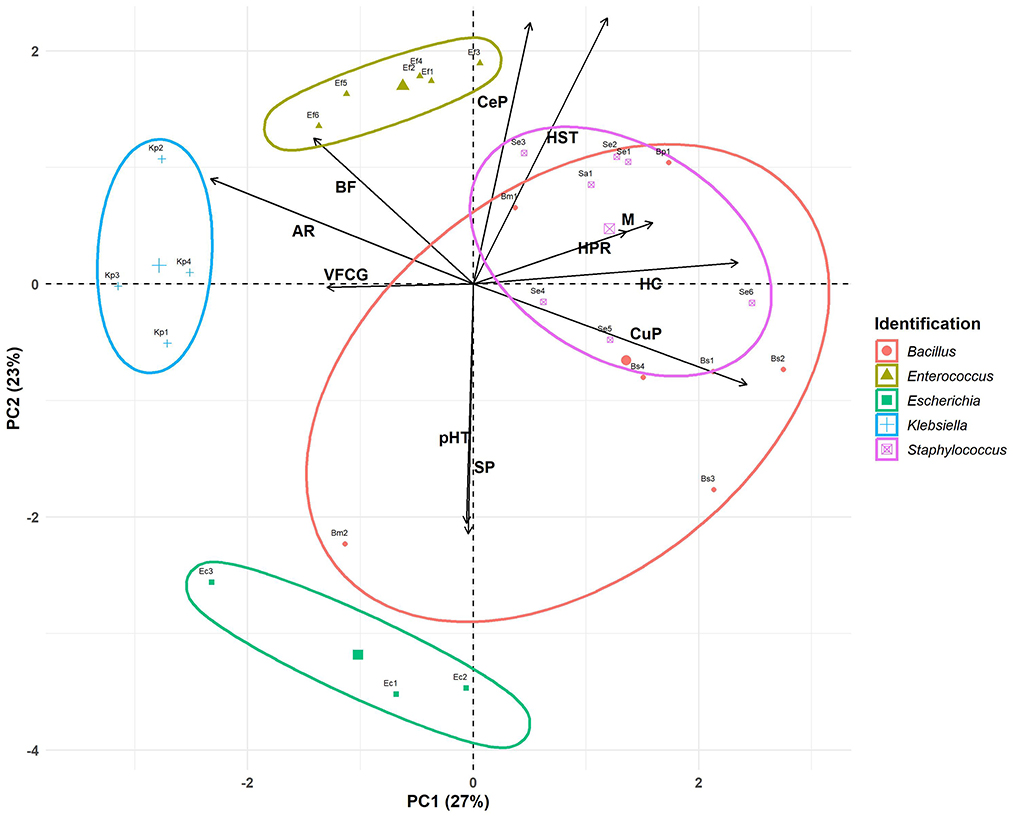
Figure 2. Principal components analysis. Antimicrobial resistance (AR), biofilm formation (BF), hemolytic capacity (HC), motility (M), siderophore production (SP), virulence factor coding genes (VFCG), curli production (CuP), cellulose production (CeP), H2O2 resistance (HPR), pH 2.5 resistance (pHR), and human serum resistance (HSR).
Based on PCA loadings, PC1 primarily distinguished bacteria by traits such as antimicrobial resistance (−0.45), biofilm formation (−0.27), and virulence factor coding genes (−0.25) on the left side vs. curli production (0.47), hemolytic capacity (0.46), motility (0.31), and H2O2 resistance (0.26) on the right side. PC2 differentiates bacteria based on human serum resistance (0.48), cellulose production (0.47), and biofilm formation (0.27; upper quadrant) vs. siderophore production (−0.45) and pH 2.5 resistance (−0.43; lower quadrant). It is important to highlight that biofilm formation contributes equally to both components of the PCA. Both components may reflect a balance between pathogenic potential and the host's defenses.
Each bacterial genus was enclosed within shaded ellipses, representing the clustering tendency for each bacterial group. The within-cluster sum of squares (WSS) for each group was calculated to reflect the internal variability of the clusters formed by the algorithm.
PC1 segregated the bacteria along the x-axis, positioning the clusters of Klebsiella (WSS: 1.53), Enterococcus (WSS: 1.56), and Escherichia (WSS: 3.12) to the left. These clusters were associated with antimicrobial resistance and the presence of virulence factor-coding genes. The distinction among these genera was marked by PC2, which separated the bacteria along the y-axis. Enterococcus and Klebsiella were located at the top of the graph, linked to biofilm formation and cellulose production. In contrast, Escherichia was positioned at the bottom of the graph associated with siderophore production and resistance to pH 2.5. Our results confirmed that Klebsiella and Enterococcus isolates, with robust and strong biofilm-forming abilities, respectively, were resistant to multiple antimicrobials, suggesting a direct relationship between antimicrobial resistance and biofilm-formation capacity (see Tables 1, 3). Although the UP isolates were derived from patients who had not received antibiotic therapy in the 10 days preceding the device removal procedure, it is essential to consider the potential impact of previous extended antimicrobial treatments, which may have contributed to the observed resistance. Antibiotic therapy is crucial for the treatment of UTI, but the rising prevalence of MDR UP in recent years represented a significant challenge to their effective management (Peng et al., 2018). On the other hand, the combination of increased motility with a high prevalence of virulence factors observed in Enterococcus strains suggested that these isolates may be particularly difficult to treat, becoming a significant risk in clinical settings.
Staphylococcus (WSS: 2.84) was clustered in the right center of the graph. According to PC1, this genus is associated with hemolytic capacity, motility, and curli production. Regarding PC2, this cluster was associated with cellulose production, resistance to human serum, and biofilm formation.
The most heterogeneous group was Bacillus (WSS: 3.99), with its members distributed across three quadrants of the biplot. Using PC1 as a reference, all members were associated with curli production (except Bm2). Regarding PC2, all members clustered based on cellulose production (except Bm2) and human serum resistance (except Bm2 and Bs3). B. subtilis was linked in the bottom right, associated with siderophore production and pH 2.5 resistance (PC2). Additionally, they were associated with peroxide resistance, motility, and total hemolytic capacity. Due to the variability of the Bacillus cluster, a key characteristic that cannot be fully appreciated is the strong biofilm formation, as all bacteria of this genus exhibit this trait. This ability could also allow it to persist in medical or industrial environments, though it is rarely associated with human infections. The high curli production and strong biofilm formation observed in both Staphylococcus and Bacillus suggest a robust capacity for adhesion and colonization on both abiotic and biotic surfaces. These microbial attributes may have implications for their virulence, potentially increasing the risk of systemic infections.
Bacillus spp. modulate uropathogen behavior in polymicrobial interactions
Polymicrobial interactions can modify the pathogenic potential of one organism over others. Therefore, microbes co-existing in complex communities must employ diverse mechanisms, such as cross-feeding, cooperation, competition, and immune modulation, to not only shape the composition of the bacterial community but also influence interactions with the host, affecting the progression from colonization to infection (Gaston et al., 2021). Although an exhaustive characterization of each isolate grown as a pure culture was conducted, it is essential to study their behavior in polymicrobial systems. As mentioned before, in five of the eight catheters, more than one bacterial species was isolated (Table 1). Notably, Bacillus spp. was present in four catheters, allowing us to study its role in interactions with microorganisms typically regarded as pathogens. Therefore, we investigate the interaction between four co-isolated pairs obtained simultaneously from the same catheter: Ef5/Bp1, Kp2/Bm2, Se3/Bs1, and Se4/Bs2 (see Table 1). First, we performed interaction assays in a solid medium, with deferred and simultaneous inoculation of the isolates. As shown in Figures 3A, B, Bp1 inhibits the growth of Ef5 when inoculated simultaneously and when inoculated first in deferred assays. However, when Ef5 was first inoculated, a change in the colony complexity was observed in the interaction zone, denoted by the loss of the characteristic wrinkling of the Bp1 colony. Regarding Kp2/Bm2, Kp2 induced slight transparency in the Bm2 colony within the interaction zone, which can be interpreted as an inhibition of Bm2 during the simultaneous growth of both bacteria. This phenotype was exacerbated when Kp2 growth first occurred in the deferred assay (Figure 3A).
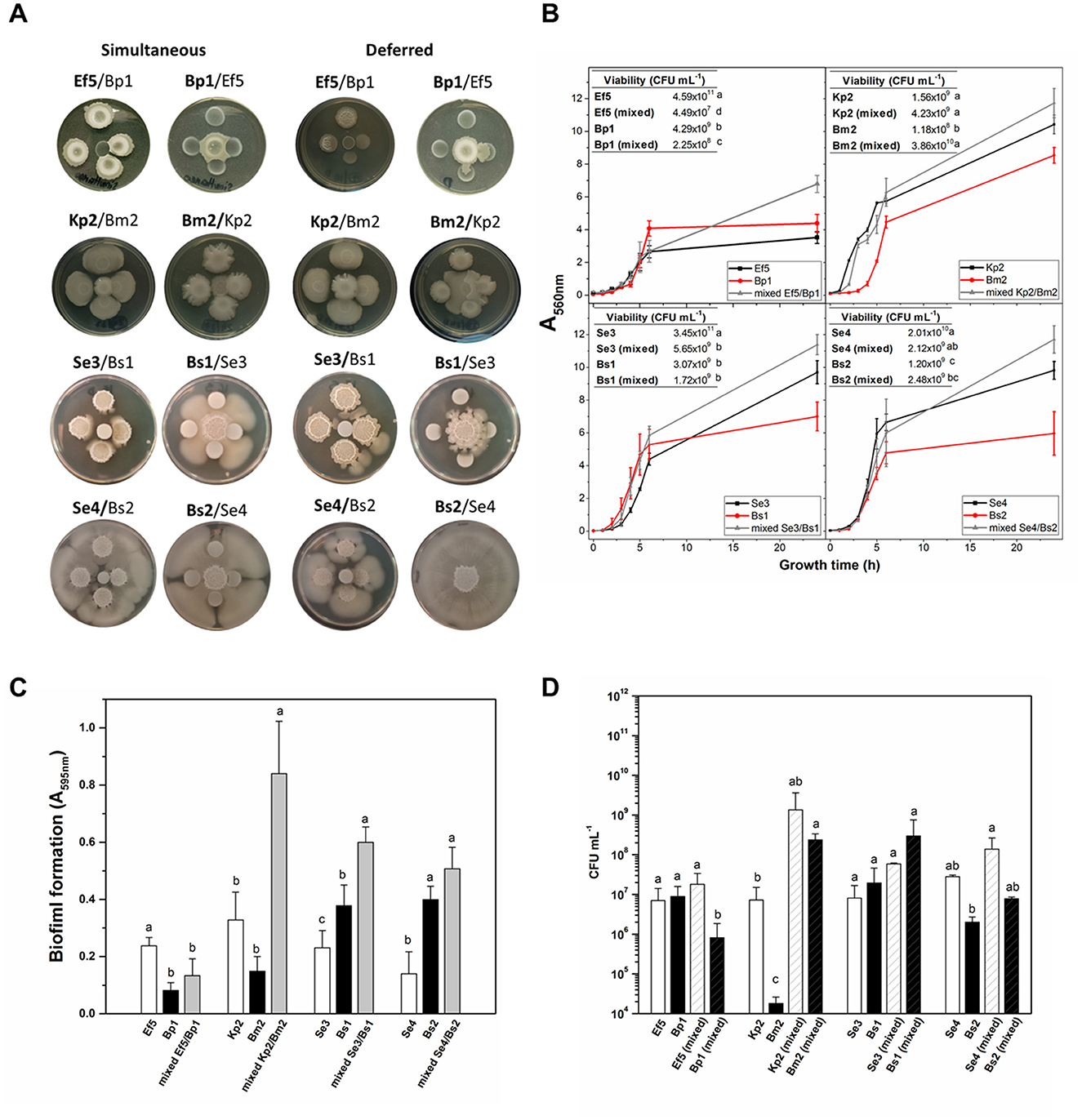
Figure 3. Interaction of co-isolated pairs. (A) In the colony interaction assay, aliquots of the co-isolated pair were inoculated simultaneously or 24 h post-growth at 30°C (deferred) on BHI-agar plates. Bold letters in the name pair indicate the central bacterium. (B) Growth curves (A560nm) of axenic or mixed (Ef5/Bp1, Kp2/Bm2, Se3/Bs1, and Se4/Bs2) were conducted in BHI with agitation for 24 h. Inset: Isolate viability in axenic or mixed conditions. (C) Biofilm formation (A595nm) and (D) cell viability (CFU mL−1) were determined for the four co-isolated pairs after 24 h of growth. Data represent the mean ± SD of at least four independent experiments. For each co-pair interaction assay, different letters indicate significant differences, as determined by one-way ANOVA performed with the Fisher test, with a p-value of 0.05.
When S. epidermidis/B. subtilis interactions were analyzed, the results showed that, in both cases, S. epidermidis enhanced the motility of B. subtilis when the bacteria grew simultaneously. Notably, this effect was more robust in the Se4/Bs2 pair. When B. subtilis strains Bs1 and Bs2 were grown 24 h prior to their counterparts, they significantly inhibited the growth of Se3 and Se4, respectively (Figure 3A). The observed behaviors between these bacteria were not surprising since it was recently described that B. subtilis actively responds to the presence of S. epidermidis in its proximity by two strategies: antimicrobial production and the development of a subpopulation with a migratory response (Hernandez-Valdes et al., 2020). However, in this study, these behaviors depend on the moment in which the interaction occurs. Control of single colonies' growth at 24 and 48 h is shown in Supplementary Figure S5. Interspecies interactions vary significantly depending on whether the bacterial strains grow individually or share the same growth location. In deferred assays (e.g., Ef5, Bp1, and Bs1), the potential production of metabolites, including antimicrobial compounds or other agents that alter the local environment, may lead to the observed inhibition of the counterpart upon its arrival. However, this potential metabolite production may be modified in the presence of the other strain when both grow simultaneously, resulting in a different type of interaction. This observation raises the possibility that colonization order influences physiological adaptations and virulence expression in each species, potentially affecting the progression and severity of infection in the host.
Since bacteria in clinical settings are generally found in mixed cultures or communities that share a common environment, we decided to perform co-culture assays in agitated and static liquid media. Growth curves (A560nm) and viable cell counts (CFU mL−1) for both mixed and monocultures are shown in Figure 3B. In general, mixed cultures in all pairs, except for the Kp2/Bm2, displayed higher A560nm values than monocultures after 24 h (p < 0.05). However, different results were observed when viable cell quantification was assessed. In mixed culture, Ef5 and Bp1 exhibited a lower viability compared to their respective monocultures (see inset tables). In the Se3/Bs1 and Se4/Bs2 pairs, the viable cells of S. epidermidis strains in mixed culture decreased by at least one order of magnitude, suggesting that B. subtilis may exert an inhibitory effect on the growth of S. epidermidis. This inhibition may be attributed to the production of secondary metabolites by B. subtilis, such as surfactins or antimicrobial peptides, which have been previously reported as antimicrobial substances (Gonzalez et al., 2011; Hernandez-Valdes et al., 2020). Conversely, in the Kp2/Bm2 pair, Kp2 viability remained unaffected when grown in mixed culture after 24 h, while Bm2 CFU mL−1 increased in mixed cultures.
Considering that co-isolated pairs were attached to the catheters, the biofilm formation of the four co-isolated pairs was evaluated on polystyrene plates after 24 h in mixed and axenic cultures (Figure 3C). The results showed that biofilm formation was enhanced in three of the four co-isolate pairs (Kp2/Bm2, Se3/Bs1, and Se4/Bs2) compared to single-species biofilms (Figure 3C). To assess the contribution of each isolate to biofilm formation, bacterial viability was determined in the biofilms (Figure 3D). As expected, in the co-cultures, where increased biofilm formation was observed, the number of viable cells was higher than in the monocultures. Different studies demonstrated that B. subtilis often facilitates biofilm development by producing extracellular polymeric substances (EPS), which provide structural support and protection to cohabitating species within the biofilm matrix (Vlamakis et al., 2013). Regarding the Ef5/Bp1 pair, Ef5 viability remained similar in mixed and single-species biofilms, while the number of viable Bp1 cells was reduced in the mixed culture compared to the monoculture (Figure 3D). This may be due to the production of antimicrobial peptides since Enterococcus spp. is one of the most frequent producers of bacteriocins (Almeida-Santos et al., 2021). Our results showed that interactions between the different co-isolated pairs reflected a combination of competitive, inhibitory, and cooperative behaviors in biofilm formation, consistent with the description of microbial cohabitation in this lifestyle (Luo et al., 2021).
The role of Bacillus spp. in these dynamics, particularly its ability to enhance biofilm formation and inhibit certain pathogens, highlights the complexity of polymicrobial interactions in complex environments, such as double-J catheters. The order of colonization may not only determine which microorganism dominates the niche but also have a direct impact on the patient's clinical outcome. If a virulent bacterium colonizes first, it may alter the environment, either promoting or limiting subsequent colonization by other species, thereby influencing the overall virulence of the infection. Conversely, simultaneous colonization could lead to complex interspecies interactions with potentially unpredictable consequences for infection progression and treatment efficacy. These findings underscore the need to investigate the role of polymicrobial colonization in clinical infections and its impact on host response and the efficacy of antimicrobial treatments.
Conclusion
The characterization of phenotypic and genotypic traits of UP isolates obtained from double-J catheters enhances the understanding of the potential virulence of these bacteria in clinical settings. The observed virulence factors and interactions of Bacillus spp. suggest that this genus may serve as an opportunistic pathogen, highlighting its potential clinical relevance in co-infections of the urinary tract. Overall, this study provides a foundation for advancing knowledge of bacterial pathogenesis associated with indwelling devices, providing crucial insights for the effective clinical management and treatment of multi-bacterial urinary infections.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for this work because according to the CIOMS – International Ethical Guidelines for Health-related Research Involving Humans, Biological material refers to materials derived from the human body, such as organs, tissues, blood, cells, DNA, and bodily fluids. Discarded medical devices or materials not containing such identifiable biological components may be exempt from ethics review. Based on this, the double J catheters used in this study do not fall under the category of biological material requiring ethical approval. The samples used in this study were acquired from a by-product of routine care. Written informed consent to participate in this study was not required from the participants or the participants' legal guardians/next of kin in accordance with the national legislation and the institutional requirements
Author contributions
JF: Validation, Methodology, Supervision, Conceptualization, Writing – original draft, Visualization, Writing – review & editing, Investigation. EC: Visualization, Formal analysis, Writing – original draft, Methodology, Data curation, Investigation. JTA-E: Methodology, Investigation, Writing – original draft. JA: Validation, Data curation, Software, Writing – original draft, Visualization. ACV: Writing – original draft, Methodology. EMH: Writing – original draft, Formal analysis, Visualization, Methodology. LS: Formal analysis, Writing – original draft, Visualization, Methodology. VR: Writing – review & editing, Investigation, Project administration, Visualization, Resources, Funding acquisition, Validation. JMV: Conceptualization, Funding acquisition, Writing – review & editing, Validation, Methodology, Writing – original draft, Investigation, Formal analysis. MG-P: Writing – review & editing, Supervision, Writing – original draft, Funding acquisition, Software, Resources, Investigation, Project administration, Formal analysis, Conceptualization, Validation, Visualization, Methodology, Data curation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by Argentinean grants of the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) (PIP 3262-2021, 2021–2023) and the Agencia Nacional de Promoción Científica y Técnica (ANPCyT) (PICT, PICT 2021-GRF-TI-00306, 2022–2023; PICT-2021-CAT-I-00159, 2022–2023; and PICT 02977, 2020–2021).
Acknowledgments
We gratefully acknowledge Dr. Martin Lescano for providing the catheters.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Correction note
A correction has been made to this article. Details can be found at: 10.3389/fmicb.2025.1674834.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1624743/full#supplementary-material
References
Adamski, P., Byczkowska-Rostkowska, Z., Gajewska, J., Zakrzewski, A. J., and Klebukowska, L. (2023). Prevalence and antibiotic resistance of Bacillus sp. isolated from raw milk. Microorganisms 11:1065. doi: 10.3390/microorganisms11041065
Agace, W. W., Patarroyo, M., Svensson, M., Carlemalm, E., and Svanborg, C. (1995). Escherichia coli induces transuroepithelial neutrophil migration by an intercellular adhesion molecule-1-dependent mechanism. Infect. Immun. 63, 4054–4062. doi: 10.1128/iai.63.10.4054-4062.1995
Almeida-Santos, A. C., Novais, C., Peixe, L., and Freitas, A. R. (2021). Enterococcus spp. as a producer and target of bacteriocins: a double-edged sword in the antimicrobial resistance crisis context. Antibiotics 10:1215. doi: 10.3390/antibiotics10101215
Al-Qahtani, M., Safan, A., Jassim, G., and Abadla, S. (2019). Efficacy of anti-microbial catheters in preventing catheter associated urinary tract infections in hospitalized patients: a review on recent updates. J. Infect. Public Health 12, 760–766. doi: 10.1016/j.jiph.2019.09.009
Arciola, C. R., Campoccia, D., Gamberini, S., Cervellati, M., Donati, E., and Montanaro, L. (2002). Detection of slime production by means of an optimised Congo red agar plate test based on a colourimetric scale in Staphylococcus epidermidis clinical isolates genotyped for ica locus. Biomaterials 23, 4233–4239. doi: 10.1016/S0142-9612(02)00171-0
Arias, C. A., and Murray, B. E. (2012). The rise of the Enterococcus: beyond vancomycin resistance. Nat. Rev. Microbiol. 10, 266–278. doi: 10.1038/nrmicro2761
Azevedo, A. S., Almeida, C., Melo, L. F., and Azevedo, N. F. (2017). Impact of polymicrobial biofilms in catheter-associated urinary tract infections. Crit. Rev. Microbiol. 43, 423–439. doi: 10.1080/1040841X.2016.1240656
Badhan, R., Singh, D. V., Badhan, L. R., and Kaur, A. (2016). Evaluation of bacteriological profile and antibiotic sensitivity patterns in children with urinary tract infection: a prospective study from a tertiary care center. Indian J. Urol. 32, 50–56. doi: 10.4103/0970-1591.173118
Bianco, A., Capozzi, L., Monno, M. R., Del Sambro, L., Manzulli, V., Pesole, G., et al. (2020). Characterization of Bacillus cereus group isolates from human bacteremia by whole-genome sequencing. Front. Microbiol. 11:599524. doi: 10.3389/fmicb.2020.599524
Blanco, J., Blanco, M., Blanco, J. E., Mora, A., Alonso, M. P., González, E. A., et al. (2002). “Enterobacterias: características generales. Género Escherichia,” in Manual de Microbiología Veterinaria, eds. S. Vadillo, S. Píriz, and E. Mateos (España: McGraw-Hill Interamericana España), 301–325.
Bohning, J., Ghrayeb, M., Pedebos, C., Abbas, D. K., Khalid, S., Chai, L., et al. (2022). Donor-strand exchange drives assembly of the TasA scaffold in Bacillus subtilis biofilms. Nat. Commun. 13:7082. doi: 10.1038/s41467-022-34700-z
Bokranz, W., Wang, X., Tschape, H., and Romling, U. (2005). Expression of cellulose and curli fimbriae by Escherichia coli isolated from the gastrointestinal tract. J. Med. Microbiol. 54(Pt 12), 1171–1182. doi: 10.1099/jmm.0.46064-0
Bossa, L., Kline, K., McDougald, D., Lee, B. B., and Rice, S. A. (2017). Urinary catheter-associated microbiota change in accordance with treatment and infection status. PLoS ONE 12:e0177633. doi: 10.1371/journal.pone.0177633
Branda, S. S., Chu, F., Kearns, D. B., Losick, R., and Kolter, R. (2006). A major protein component of the Bacillus subtilis biofilm matrix. Mol. Microbiol. 59, 1229–1238. doi: 10.1111/j.1365-2958.2005.05020.x
Busscher, H. J., van de Belt-Gritter, B., Dijkstra, R. J., Norde, W., and van der Mei, H. C. (2008). Streptococcus mutans and Streptococcus intermedius adhesion to fibronectin films are oppositely influenced by ionic strength. Langmuir 24, 10968–10973. doi: 10.1021/la8016968
Buxton, R. (2005). Blood agar plates and hemolysis protocols. Am. Soc. Microbiol. 1–9. Available online at: https://asm.org/protocols/blood-agar-plates-and-hemolysis-protocols
Cavalcanti, F. C. N., Rodrigues, J. F., Cabral, A. B., Azarias, L. C. B., Morais Junior, M. A., Castro, C., et al. (2019). Relationships between phagocytosis, mucoid phenotype, and genetic characteristics of Klebsiella pneumoniae clinical isolates. Rev. Soc. Bras. Med. Trop. 52:e20190089B. doi: 10.1590/0037-8682-0089-2019
Cepas, V., Lopez, Y., Munoz, E., Rolo, D., Ardanuy, C., Marti, S., et al. (2019). Relationship between biofilm formation and antimicrobial resistance in gram-negative bacteria. Microb. Drug Resist. 25, 72–79. doi: 10.1089/mdr.2018.0027
Chapman, M. R., Robinson, L. S., Pinkner, J. S., Roth, R., Heuser, J., Hammar, M., et al. (2002). Role of Escherichia coli curli operons in directing amyloid fiber formation. Science 295, 851–855. doi: 10.1126/science.1067484
Chatterjee, S., Maiti, P., Dey, R., Kundu, A., and Dey, R. (2014). Biofilms on indwelling urologic devices: microbes and antimicrobial management prospect. Ann. Med. Health Sci. Res. 4, 100–104. doi: 10.4103/2141-9248.126612
Chen, Y. H., Ko, W. C., and Hsueh, P. R. (2013). Emerging resistance problems and future perspectives in pharmacotherapy for complicated urinary tract infections. Expert Opin. Pharmacother. 14, 587–596. doi: 10.1517/14656566.2013.778827
Chua, R. Y. R., Lim, K., Leong, S. S. J., Tambyah, P. A., and Ho, B. (2017). An in-vitro urinary catheterization model that approximates clinical conditions for evaluation of innovations to prevent catheter-associated urinary tract infections. J. Hosp. Infect. 97, 66–73. doi: 10.1016/j.jhin.2017.05.006
CLSI (2020). Performance Standards for Antimicrobial Susceptibiity Testing, 30th Edn. CLSIsupplement M100. Wayne, PA: CLSI.
Cotter, P. D., and Hill, C. (2003). Surviving the acid test: responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 67, 429–453. doi: 10.1128/MMBR.67.3.429-453.2003
Croxall, G., Weston, V., Joseph, S., Manning, G., Cheetham, P., and McNally, A. (2011). Increased human pathogenic potential of Escherichia coli from polymicrobial urinary tract infections in comparison to isolates from monomicrobial culture samples. J. Med. Microbiol. 60(Pt 1), 102–109. doi: 10.1099/jmm.0.020602-0
Cui, H., Zhang, C., Li, C., and Lin, L. (2020). Inhibition of Escherichia coli O157:H7 biofilm on vegetable surface by solid liposomes of clove oil. LWT 117:108656. doi: 10.1016/j.lwt.2019.108656
Da Re, S., and Ghigo, J. M. (2006). A CsgD-independent pathway for cellulose production and biofilm formation in Escherichia coli. J. Bacteriol. 188, 3073–3087. doi: 10.1128/JB.188.8.3073-3087.2006
Dalhoff, A. (2012). Resistance surveillance studies: a multifaceted problem–the fluoroquinolone example. Infection 40, 239–262. doi: 10.1007/s15010-012-0257-2
Danese, P. N., Pratt, L. A., Dove, S. L., and Kolter, R. (2000). The outer membrane protein, antigen 43, mediates cell-to-cell interactions within Escherichia coli biofilms. Mol. Microbiol. 37, 424–432. doi: 10.1046/j.1365-2958.2000.02008.x
Del Pozo, J. L. (2018). Biofilm-related disease. Expert Rev. Anti Infect. Ther. 16, 51–65. doi: 10.1080/14787210.2018.1417036
Delcaru, C., Alexandru, I., Podgoreanu, P., Grosu, M., Stavropoulos, E., Chifiriuc, M. C., et al. (2016). Microbial biofilms in urinary tract infections and prostatitis: etiology, pathogenicity, and combating strategies. Pathogens 5:65. doi: 10.3390/pathogens5040065
Drobniewski, F. A. (1993). Bacillus cereus and related species. Clin. Microbiol. Rev. 6, 324–338. doi: 10.1128/CMR.6.4.324
Dunne, W. M. Jr. (2002). Bacterial adhesion: seen any good biofilms lately? Clin. Microbiol. Rev. 15, 155–166. doi: 10.1128/CMR.15.2.155-166.2002
Eberly, A. R., Beebout, C. J., Carmen Tong, C. M., Van Horn, G. T., Green, H. D., Fitzgerald, M. J., et al. (2020). Defining a molecular signature for uropathogenic versus urocolonizing Escherichia coli: the status of the field and new clinical opportunities. J. Mol. Biol. 432, 786–804. doi: 10.1016/j.jmb.2019.11.008
Echverría-Zarate, J., Sarmiento, A. E, and Osores-Plenge, F. (2006). Urinary tract infection and antibiotic treatment. Acta Méd. Peru. 23, 26–31.
El Abed, S., Ibnsouda, S. K., Latrache, H., Meftah, H., Tahri, N. J., and Hamadi, F. (2012). Environmental Scanning Electron Microscopy characterization of the adhesion of conidia from Penicillium expansum to cedar wood substrata at different pH values. World J. Microbiol. Biotechnol. 28, 1707–1713. doi: 10.1007/s11274-011-0980-3
Fayanju, A. M., Akinyele, B. J., Oladejo, B. O., and Osunla, A. C. (2024). Genetic composition, virulence factors, and antimicrobial resistance profiles of Bacillus cereus and Bacillus subtilis isolates from food vendors in Ondo State, Nigeria: implications for food safety. Bull. Natl. Res. Centre 48:90. doi: 10.1186/s42269-024-01245-8
Fekete, T. (2015). “Bacillus species and related genera other than Bacillus anthracis,” in Mandell, Douglas, and Bennett's Principles and Practice of Infectious Diseases, 8th Edn., eds. J. E. Bennett, R. Dolin, and M. J. Blaser (Philadelphia, PA: W. B. Saunders), 2410–2414.e2411. doi: 10.1016/B978-1-4557-4801-3.00210-1
Flemming, H. C., and Wingender, J. (2010). The biofilm matrix. Nat. Rev. Microbiol. 8, 623–633. doi: 10.1038/nrmicro2415
Flores-Mireles, A., Hreha, T. N., and Hunstad, D. A. (2019). Pathophysiology, treatment, and prevention of catheter-associated urinary tract infection. Top. Spinal Cord Inj. Rehabil. 25, 228–240. doi: 10.1310/sci2503-228
Flores-Mireles, A. L., Walker, J. N., Caparon, M., and Hultgren, S. J. (2015). Urinary tract infections: epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 13, 269–284. doi: 10.1038/nrmicro3432
Foster, J. W. (2004). Escherichia coli acid resistance: tales of an amateur acidophile. Nat. Rev. Microbiol. 2, 898–907. doi: 10.1038/nrmicro1021
Garau, J. (2008). Other antimicrobials of interest in the era of extended-spectrum beta-lactamases: fosfomycin, nitrofurantoin and tigecycline. Clin. Microbiol. Infect. 14(Suppl. 1), 198–202. doi: 10.1111/j.1469-0691.2007.01852.x
Garcia, E. C., Brumbaugh, A. R., and Mobley, H. L. (2011). Redundancy and specificity of Escherichia coli iron acquisition systems during urinary tract infection. Infect. Immun. 79, 1225–1235. doi: 10.1128/IAI.01222-10
Gaston, J. R., Andersen, M. J., Johnson, A. O., Bair, K. L., Sullivan, C. M., Guterman, L. B., et al. (2020). Enterococcus faecalis polymicrobial interactions facilitate biofilm formation, antibiotic recalcitrance, and persistent colonization of the catheterized urinary tract. Pathogens 9:835. doi: 10.3390/pathogens9100835
Gaston, J. R., Johnson, A. O., Bair, K. L., White, A. N., and Armbruster, C. E. (2021). Polymicrobial interactions in the urinary tract: is the enemy of my enemy my friend? Infect. Immun. 89, 10–1128. doi: 10.1128/IAI.00652-20
Gauhar, V., Pirola, G. M., Scarcella, S., De Angelis, M. V., Giulioni, C., Rubilotta, E., et al. (2022). Nephrostomy tube versus double J ureteral stent in patients with malignant ureteric obstruction. A systematic review and meta-analysis of comparative studies. Int. Braz. J. Urol. 48, 903–914. doi: 10.1590/S1677-5538.IBJU.2022.0225
Gerhardt, P., Murray, R. G. E., Murray, R. G. E., Krieg, N. R., and Wood, W. A. (1994). Methods for General and Molecular Bacteriology. Washington, DC: American Society for Microbiology. doi: 10.1002/food.19960400226
Gonzalez, D. J., Haste, N. M., Hollands, A., Fleming, T. C., Hamby, M., Pogliano, K., et al. (2011). Microbial competition between Bacillus subtilis and Staphylococcus aureus monitored by imaging mass spectrometry. Microbiology 157(Pt 9), 2485–2492. doi: 10.1099/mic.0.048736-0
Gould C. V. Umscheid C. A. Agarwal R. K. Kuntz G. Pegues D. A. Healthcare Infection Control Practices Advisory C. (2010). Guideline for prevention of catheter-associated urinary tract infections 2009. Infect. Control Hosp. Epidemiol. 31, 319–326. doi: 10.1086/651091
Grillo-Puertas, M., Martinez-Zamora, M. G., Rintoul, M. R., Soto, S. M., and Rapisarda, V. A. (2015). Environmental phosphate differentially affects virulence phenotypes of uropathogenic Escherichia coli isolates causative of prostatitis. Virulence 6, 608–617. doi: 10.1080/21505594.2015.1059561
Guyer, D. M., Radulovic, S., Jones, F. E., and Mobley, H. L. (2002). Sat, the secreted autotransporter toxin of uropathogenic Escherichia coli, is a vacuolating cytotoxin for bladder and kidney epithelial cells. Infect. Immun. 70, 4539–4546. doi: 10.1128/IAI.70.8.4539-4546.2002
Hancock, L. E., and Gilmore, M. S. (2006). “Pathogenicity of Enterococci,” in Gram-Positive Pathogens, eds. V. A. Fischetti, R. P. Novick, J. J. Ferretti, D. A. Portnoy and J. I. Rood (Washington, DC: ASM Press), 299–311. doi: 10.1128/9781555816513.ch25
Hancock, L. E., and Perego, M. (2004). The Enterococcus faecalis fsr two-component system controls biofilm development through production of gelatinase. J. Bacteriol. 186, 5629–5639. doi: 10.1128/JB.186.17.5629-5639.2004
Hansen, L. T., and Vogel, B. F. (2011). Desiccation of adhering and biofilm Listeria monocytogenes on stainless steel: survival and transfer to salmon products. Int. J. Food Microbiol. 146, 88–93. doi: 10.1016/j.ijfoodmicro.2011.01.032
Henrichsen, J. (1997). “Streptococcus milleri” strains exhibit not gliding motility but sliding. Int. J. Syst. Bacteriol. 47:604. doi: 10.1099/00207713-47-2-604
Hernandez-Valdes, J. A., Zhou, L., de Vries, M. P., and Kuipers, O. P. (2020). Impact of spatial proximity on territoriality among human skin bacteria. NPJ Biofilms Microbiomes 6:30. doi: 10.1038/s41522-020-00140-0
Hood, M. I., and Skaar, E. P. (2012). Nutritional immunity: transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 10, 525–537. doi: 10.1038/nrmicro2836
Hooton, T. M. (2012). Clinical practice. Uncomplicated urinary tract infection. N. Engl. J. Med. 366, 1028–1037. doi: 10.1056/NEJMcp1104429
Houry, A., Briandet, R., Aymerich, S., and Gohar, M. (2010). Involvement of motility and flagella in Bacillus cereus biofilm formation. Microbiology 156(Pt 4), 1009–1018. doi: 10.1099/mic.0.034827-0
Imlay, J. A. (2013). The molecular mechanisms and physiological consequences of oxidative stress: lessons from a model bacterium. Nat. Rev. Microbiol. 11, 443–454. doi: 10.1038/nrmicro3032
Jacobsen, S. M., Stickler, D. J., Mobley, H. L., and Shirtliff, M. E. (2008). Complicated catheter-associated urinary tract infections due to Escherichia coli and Proteus mirabilis. Clin. Microbiol. Rev. 21, 26–59. doi: 10.1128/CMR.00019-07
Jessberger, N., Krey, V. M., Rademacher, C., Bohm, M. E., Mohr, A. K., Ehling-Schulz, M., et al. (2015). From genome to toxicity: a combinatory approach highlights the complexity of enterotoxin production in Bacillus cereus. Front. Microbiol. 6:560. doi: 10.3389/fmicb.2015.00560
Johnson, J. R. (2002). “Evolution of pathogenic Escherichia coli,” in Escherichia coli: Virulence Mechanisms of a Versatile Pathogen, eds. C. M. Thorpe, J. M. Ritchie, and K. Acheson (Amsterdam: Academic Press), 55–77. doi: 10.1016/B978-012220751-8/50003-3
Johnson, J. R., Scheutz, F., Ulleryd, P., Kuskowski, M. A., O'Bryan, T. T., and Sandberg, T. (2005). Phylogenetic and pathotypic comparison of concurrent urine and rectal Escherichia coli isolates from men with febrile urinary tract infection. J. Clin. Microbiol. 43, 3895–3900. doi: 10.1128/JCM.43.8.3895-3900.2005
Josenhans, C., and Suerbaum, S. (2002). The role of motility as a virulence factor in bacteria. Int. J. Med. Microbiol. 291, 605–614. doi: 10.1078/1438-4221-00173
Juarez, G. E., and Galvan, E. M. (2018). Role of nutrient limitation in the competition between uropathogenic strains of Klebsiella pneumoniae and Escherichia coli in mixed biofilms. Biofouling 34, 287–298. doi: 10.1080/08927014.2018.1434876
Kaiser, T. D., Pereira, E. M., Dos Santos, K. R., Maciel, E. L., Schuenck, R. P., and Nunes, A. P. (2013). Modification of the Congo red agar method to detect biofilm production by Staphylococcus epidermidis. Diagn. Microbiol. Infect. Dis. 75, 235–239. doi: 10.1016/j.diagmicrobio.2012.11.014
Kaito, C., and Sekimizu, K. (2007). Colony spreading in Staphylococcus aureus. J. Bacteriol. 189, 2553–2557. doi: 10.1128/JB.01635-06
Kalsi, J., Arya, M., Wilson, P., and Mundy, A. (2003). Hospital-acquired urinary tract infection. Int. J. Clin. Pract. 57, 388–391. doi: 10.1111/j.1742-1241.2003.tb10513.x
Kanamaru, S., Kurazono, H., Ishitoya, S., Terai, A., Habuchi, T., Nakano, M., et al. (2003). Distribution and genetic association of putative uropathogenic virulence factors iroN, iha, kpsMT, ompT and usp in Escherichia coli isolated from urinary tract infections in Japan. J. Urol. 170(6 Pt 1), 2490–2493. doi: 10.1097/01.ju.0000094185.48467.dc
Kandi, V., Palange, P., Vaish, R., Bhatti, A. B., Kale, V., Kandi, M. R., et al. (2016). Emerging bacterial infection: identification and clinical significance of Kocuria species. Cureus 8:e731. doi: 10.7759/cureus.731
Kart, D., Kustimur, A. S., Sagiroglu, M., and Kalkanci, A. (2017). Evaluation of antimicrobial durability and anti-biofilm effects in urinary catheters against Enterococcus faecalis clinical isolates and reference strains. Balkan Med. J. 34, 546–552. doi: 10.4274/balkanmedj.2016.1853
Kavanaugh, J. S., and Horswill, A. R. (2016). Impact of environmental cues on staphylococcal quorum sensing and biofilm development. J. Biol. Chem. 291, 12556–12564. doi: 10.1074/jbc.R116.722710
Kazi, M. M., Harshe, A., Sale, H., Mane, D., Yande, M., and Chabukswar, S. (2015). Catheter associated urinary tract infections (CAUTI) and antibiotic sensitivity pattern from confirmed cases of CAUTI in a tertiary care hospital: a prospective study. Clin. Microbiol. 4:2. doi: 10.4172/2327-5073.1000193
Kearns, D. B., and Losick, R. (2003). Swarming motility in undomesticated Bacillus subtilis. Mol. Microbiol. 49, 581–590. doi: 10.1046/j.1365-2958.2003.03584.x
Kehinde, E. O., Rotimi, V. O., Al-Hunayan, A., Abdul-Halim, H., Boland, F., and Al-Awadi, K. A. (2004). Bacteriology of urinary tract infection associated with indwelling J ureteral stents. J. Endourol. 18, 891–896. doi: 10.1089/end.2004.18.891
Klein, R. D., and Hultgren, S. J. (2020). Urinary tract infections: microbial pathogenesis, host-pathogen interactions and new treatment strategies. Nat. Rev. Microbiol. 18, 211–226. doi: 10.1038/s41579-020-0324-0
Kline, K. A., and Lewis, A. L. (2016). Gram-positive uropathogens, polymicrobial urinary tract infection, and the emerging microbiota of the urinary tract. Microbiol. Spectr. 4, 1–31. doi: 10.1128/microbiolspec.UTI-0012-2012
Klis, R., Korczak-Kozakiewicz, E., Denys, A., Sosnowski, M., and Rozanski, W. (2009). Relationship between urinary tract infection and self-retaining double-J catheter colonization. J. Endourol. 23, 1015–1019. doi: 10.1089/end.2008.0518
Klis, R., Szymkowiak, S., Madej, A., Blewniewski, M., Krzeslak, A., Forma, E., et al. (2014). Rate of positive urine culture and double-J catheters colonization on the basis of microorganism DNA analysis. Cent. Euro. J. Urol. 67, 81–85.
Kong, C., Neoh, H. M., and Nathan, S. (2016). Targeting Staphylococcus aureus toxins: a potential form of anti-virulence therapy. Toxins 8:72. doi: 10.3390/toxins8030072
Lazar, V., and Chifiriuc, M. C. (2010). Medical significance and new therapeutical strategies for biofilm associated infections. Roum. Arch. Microbiol. Immunol. 69, 125–138.
Learman, B. S., Brauer, A. L., Eaton, K. A., and Armbruster, C. E. (2019). A rare opportunist, Morganella morganii, decreases severity of polymicrobial catheter-associated urinary tract infection. Infect. Immun. 88, 1–17. doi: 10.1128/IAI.00691-19
Lenchenko, E., Blumenkrants, D., Sachivkina, N., Shadrova, N., and Ibragimova, A. (2020). Morphological and adhesive properties of Klebsiella pneumoniae biofilms. Vet. World 13, 197–200. doi: 10.14202/vetworld.2020.197-200
Li, Y., Jiang, L., Luo, S., Hu, D., Zhao, X., Zhao, G., et al. (2023). Analysis of characteristics, pathogens and drug resistance of urinary tract infection associated with long-term indwelling double-J stent. Infect. Drug Resist. 16, 2089–2096. doi: 10.2147/IDR.S392857
Lin, C., Liang, A., Liang, S., Wang, X., Meng, L., and Chen, M. (2022). Laparoscopic surgery combined with the double-J tube implantation for the rare cystic-solid schwannoma of seminal vesicle: a case report and literature review. Medicine 101:e29352. doi: 10.1097/MD.0000000000029352
Logan, L. K., and Weinstein, R. A. (2017). The epidemiology of carbapenem-resistant enterobacteriaceae: the impact and evolution of a global menace. J. Infect. Dis. 215, S28–S36. doi: 10.1093/infdis/jiw282
Luo, A., Wang, F., Sun, D., Liu, X., and Xin, B. (2021). Formation, development, and cross-species interactions in biofilms. Front. Microbiol. 12:757327. doi: 10.3389/fmicb.2021.757327
Mandakhalikar, K. D., Rahmat, J. N., Chiong, E., Neoh, K. G., Shen, L., and Tambyah, P. A. (2018). Extraction and quantification of biofilm bacteria: method optimized for urinary catheters. Sci. Rep. 8:8069. doi: 10.1038/s41598-018-26342-3
Martin-Rodriguez, A. J., Villion, K., Yilmaz-Turan, S., Vilaplana, F., Sjoling, A., and Romling, U. (2021). Regulation of colony morphology and biofilm formation in Shewanella algae. Microb. Biotechnol. 14, 1183–1200. doi: 10.1111/1751-7915.13788
Matz, C., Webb, J. S., Schupp, P. J., Phang, S. Y., Penesyan, A., Egan, S., et al. (2008). Marine biofilm bacteria evade eukaryotic predation by targeted chemical defense. PLoS ONE 3:e2744. doi: 10.1371/journal.pone.0002744
Mazzariol, A., Bazaj, A., and Cornaglia, G. (2017). Multi-drug-resistant Gram-negative bacteria causing urinary tract infections: a review. J. Chemother. 29, 2–9. doi: 10.1080/1120009X.2017.1380395
Monds, R. D., and O'Toole, G. A. (2009). The developmental model of microbial biofilms: ten years of a paradigm up for review. Trends Microbiol. 17, 73–87. doi: 10.1016/j.tim.2008.11.001
Moraveji, Z., Tabatabaei, M., Shirzad Aski, H., and Khoshbakht, R. (2014). Characterization of hemolysins of Staphylococcus strains isolated from human and bovine, southern Iran. Iran. J. Vet. Res. 15, 326–330.
Mulcahy, H., Charron-Mazenod, L., and Lewenza, S. (2008). Extracellular DNA chelates cations and induces antibiotic resistance in Pseudomonas aeruginosa biofilms. PLoS Pathog. 4:e1000213. doi: 10.1371/journal.ppat.1000213
Muscholl-Silberhorn, A., Rozdzinski, E., and Wirth, R. (2000). Aggregation substance of Enterococcus faecalis: a multifunctional adhesin. Adv. Exp. Med. Biol. 485, 75–83. doi: 10.1007/0-306-46840-9_10
Nallapareddy, S. R., Wenxiang, H., Weinstock, G. M., and Murray, B. E. (2005). Molecular characterization of a widespread, pathogenic, and antibiotic resistance-receptive Enterococcus faecalis lineage and dissemination of its putative pathogenicity island. J. Bacteriol. 187, 5709–5718. doi: 10.1128/JB.187.16.5709-5718.2005
Nicolle, L. E. (2000). Urinary tract infection in long-term-care facility residents. Clin. Infect. Dis. 31, 757–761. doi: 10.1086/313996
Nicolle, L. E. (2012). Urinary catheter-associated infections. Infect. Dis. Clin. North Am. 26, 13–27. doi: 10.1016/j.idc.2011.09.009
Novick, R. P., and Geisinger, E. (2008). Quorum sensing in staphylococci. Annu. Rev. Genet. 42, 541–564. doi: 10.1146/annurev.genet.42.110807.091640
Oliveira, W. F., Silva, P. M. S., Silva, R. C. S., Silva, G. M. M., Machado, G., Coelho, L., et al. (2018). Staphylococcus aureus and Staphylococcus epidermidis infections on implants. J. Hosp. Infect. 98, 111–117. doi: 10.1016/j.jhin.2017.11.008
Ostensvik, O., From, C., Heidenreich, B., O'Sullivan, K., and Granum, P. E. (2004). Cytotoxic Bacillus spp. belonging to the B. cereus and B. subtilis groups in Norwegian surface waters. J. Appl. Microbiol. 96, 987–993. doi: 10.1111/j.1365-2672.2004.02226.x
O'Toole, G. A., and Kolter, R. (1998). Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: a genetic analysis. Mol. Microbiol. 28, 449–461. doi: 10.1046/j.1365-2958.1998.00797.x
Otto, M. (2009). Staphylococcus epidermidis–the 'accidental' pathogen. Nat. Rev. Microbiol. 7, 555–567. doi: 10.1038/nrmicro2182
Ozturk, H. (2015). Tubeless versus standard PCNL in geriatric population. Actas Urol. Esp. 39, 494–501. doi: 10.1016/j.acuro.2015.02.010
Paczosa, M. K., and Mecsas, J. (2016). Klebsiella pneumoniae: going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 80, 629–661. doi: 10.1128/MMBR.00078-15
Paterson, D. L. (2006). Resistance in gram-negative bacteria: Enterobacteriaceae. Am. J. Infect. Control 34(5 Suppl. 1), S20–28; discussion: S64–73. doi: 10.1016/j.ajic.2006.05.238
Pendleton, J. N., Gorman, S. P., and Gilmore, B. F. (2013). Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti Infect. Ther. 11, 297–308. doi: 10.1586/eri.13.12
Peng, D., Li, X., Liu, P., Luo, M., Chen, S., Su, K., et al. (2018). Epidemiology of pathogens and antimicrobial resistanceof catheter-associated urinary tract infections in intensivecare units: a systematic review and meta-analysis. Am. J. Infect. Control 46, e81–e90. doi: 10.1016/j.ajic.2018.07.012
Pérez, E., Uyan, B., Dzubay, D. P., and Fenton, S. H. (2017). Catheter-associated urinary tract infections: challenges and opportunities for the application of systems engineering. Health Syst. 6, 68–76. doi: 10.1057/s41306-016-0017-0
Perilla, M. J., Ajello, G., Bopp, C., Elliott, J., Facklam, R., Knapp, J. S., et al. (2009). Manual de laboratorio para la identificación y prueba de susceptibilidad a los antimicrobianos de patógenos bacterianos de importancia para la salud pública en el mundo en desarrollo. Medicina and Laboratorio. Switzerland: World Health Organization.
Pigrau, C. (2013). Nocosomial urinary tract infections. Enferm. Infecc. Microbiol. Clin. 31, 614–624. doi: 10.1016/j.eimc.2012.11.015
Pollitt, E. J. G., and Diggle, S. P. (2017). Defining motility in the Staphylococci. Cell. Mol. Life Sci. 74, 2943–2958. doi: 10.1007/s00018-017-2507-z
Pratt, L. A., and Kolter, R. (1998). Genetic analysis of Escherichia coli biofilm formation: roles of flagella, motility, chemotaxis and type I pili. Mol. Microbiol. 30, 285–293. doi: 10.1046/j.1365-2958.1998.01061.x
Quan, K., Hou, J., Zhang, Z., Ren, Y., Peterson, B. W., Flemming, H. C., et al. (2022). Water in bacterial biofilms: pores and channels, storage and transport functions. Crit. Rev. Microbiol. 48, 283–302. doi: 10.1080/1040841X.2021.1962802
Ramarao, N., and Sanchis, V. (2013). The pore-forming haemolysins of Bacillus cereus: a review. Toxins 5, 1119–1139. doi: 10.3390/toxins5061119
Ramos, Y., and Morales, D. K. (2019). Exopolysaccharide-mediated surface penetration as new virulence trait in Enterococcus faecalis. Commun. Integr. Biol. 12, 144–147. doi: 10.1080/19420889.2019.1657373
Ranganathan, M., Akbar, M., Ilham, M. A., Chavez, R., Kumar, N., and Asderakis, A. (2009). Infective complications associated with ureteral stents in renal transplant recipients. Transplant. Proc. 41, 162–164. doi: 10.1016/j.transproceed.2008.10.022
Rendon, M. A., Saldana, Z., Erdem, A. L., Monteiro-Neto, V., Vazquez, A., Kaper, J. B., et al. (2007). Commensal and pathogenic Escherichia coli use a common pilus adherence factor for epithelial cell colonization. Proc. Natl. Acad. Sci. U.S.A. 104, 10637–10642. doi: 10.1073/pnas.0704104104
Rivera, A. M., and Boucher, H. W. (2011). Current concepts in antimicrobial therapy against select gram-positive organisms: methicillin-resistant Staphylococcus aureus, penicillin-resistant pneumococci, and vancomycin-resistant enterococci. Mayo Clin. Proc. 86, 1230–1243. doi: 10.4065/mcp.2011.0514
Rodrigues, D. F., and Elimelech, M. (2009). Role of type 1 fimbriae and mannose in the development of Escherichia coli K12 biofilm: from initial cell adhesion to biofilm formation. Biofouling 25, 401–411. doi: 10.1080/08927010902833443
Romero, D., Aguilar, C., Losick, R., and Kolter, R. (2010). Amyloid fibers provide structural integrity to Bacillus subtilis biofilms. Proc. Natl. Acad. Sci. U.S.A. 107, 2230–2234. doi: 10.1073/pnas.0910560107
Ronald, A. (2002). The etiology of urinary tract infection: traditional and emerging pathogens. Am. J. Med. 113(Suppl. 1A) 14S−19S. doi: 10.1016/S0002-9343(02)01055-0
Saint, S., and Chenoweth, C. E. (2003). Biofilms and catheter-associated urinary tract infections. Infect. Dis. Clin. North Am. 17, 411–432. doi: 10.1016/S0891-5520(03)00011-4
Salm, J., Salm, F., Arendarski, P., and Kramer, T. S. (2023). High frequency of Enterococcus faecalis detected in urinary tract infections in male outpatients - a retrospective, multicenter analysis, Germany 2015 to 2020. BMC Infect. Dis. 23:812. doi: 10.1186/s12879-023-08824-6
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual, 2nd Edn. Long Island, NY: Cold Spring Harbor Laboratory Press.
Senesi, S., Salvetti, S., Celandroni, F., and Ghelardi, E. (2010). Features of Bacillus cereus swarm cells. Res. Microbiol. 161, 743–749. doi: 10.1016/j.resmic.2010.10.007
Sharma, S., Bhatnagar, R., and Gaur, D. (2020). Complement evasion strategies of human pathogenic bacteria. Indian J. Microbiol. 60, 283–296. doi: 10.1007/s12088-020-00872-9
Shea, A. E., Frick-Cheng, A. E., Smith, S. N., and Mobley, H. L. T. (2022). Phenotypic assessment of clinical Escherichia coli isolates as an indicator for uropathogenic potential. mSystems 7:e0082722. doi: 10.1128/msystems.00827-22
Siegel J. D. Rhinehart E. Jackson M. Chiarello L. the Healthcare Infection Control Practices Advisory Committee. (2006). Management of Multidrug-Resistant Organisms in Healthcare Settings, 2006. Available online at: https://www.cdc.gov/infection-control/hcp/mdro-management
Sievert, D. M., Ricks, P., Edwards, J. R., Schneider, A., Patel, J., Srinivasan, A., et al. (2013). Antimicrobial-resistant pathogens associated with healthcare-associated infections: summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009-2010. Infect. Control Hosp. Epidemiol. 34, 1–14. doi: 10.1086/668770
Singh, K. V., Nallapareddy, S. R., Sillanpaa, J., and Murray, B. E. (2010). Importance of the collagen adhesin ace in pathogenesis and protection against Enterococcus faecalis experimental endocarditis. PLoS Pathog. 6:e1000716. doi: 10.1371/annotation/1ccae8f8-d274-4ff8-a295-815037ce9cc6
Socohou, A., Sina, H., Degbey, C., Nanoukon, C., Chabi-Sika, K., Ahouandjinou, H., et al. (2020). Antibiotics resistance and biofilm formation capacity of Staphylococcus spp. strains isolated from surfaces and medicotechnical materials. Int. J. Microbiol. 2020:6512106. doi: 10.1155/2020/6512106
Soto, S. M., Smithson, A., Martinez, J. A., Horcajada, J. P., Mensa, J., and Vila, J. (2007). Biofilm formation in uropathogenic Escherichia coli strains: relationship with prostatitis, urovirulence factors and antimicrobial resistance. J Urol 177, 365–368. doi: 10.1016/j.juro.2006.08.081
Stenfors Arnesen, L. P., Fagerlund, A., and Granum, P. E. (2008). From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 32, 579–606. doi: 10.1111/j.1574-6976.2008.00112.x
Stepanovic, S., Vukovic, D., Hola, V., Di Bonaventura, G., Djukic, S., Cirkovic, I., et al. (2007). Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 115, 891–899. doi: 10.1111/j.1600-0463.2007.apm_630.x
Stepien-Pysniak, D., Hauschild, T., Kosikowska, U., Dec, M., and Urban-Chmiel, R. (2019). Biofilm formation capacity and presence of virulence factors among commensal Enterococcus spp. from wild birds. Sci. Rep. 9:11204. doi: 10.1038/s41598-019-47602-w
Subashchandrabose, S., and Mobley, H. L. (2015). Back to the metal age: battle for metals at the host-pathogen interface during urinary tract infection. Metallomics 7, 935–942. doi: 10.1039/C4MT00329B
Sydow, K., Eger, E., Schwabe, M., Heiden, S. E., Bohnert, J. A., Franzenburg, S., et al. (2022). Geno- and phenotypic characteristics of a Klebsiella pneumoniae ST20 isolate with unusual colony morphology. Microorganisms 10:2063. doi: 10.3390/microorganisms10102063
Torres-Rodríguez, M. T., Pinheiro Queiroz Viana, A., Gleire Rodrigues De Meneses, F., Silva do Fernandes Vieira, R. H., and Viana De Sousa, O. (2020). Factores de virulencia presentes en estirpes bacterianas del género Enterococcus aisladas en el litoral de Fortaleza (Ceará, Brasil). Higiene y Sanidad Ambiental. 20, 1915–1922.
Totsika, M., Moriel, D. G., Idris, A., Rogers, B. A., Wurpel, D. J., Phan, M. D., et al. (2012). Uropathogenic Escherichia coli mediated urinary tract infection. Curr. Drug Targets 13, 1386–1399. doi: 10.2174/138945012803530206
Turnbull, P. C. B. (1996). “Bacillus,” in Medical Microbiology, ed. S. Baron (Galveston, TX: University of Texas Medical Branch at Galveston).
Ubeda, C., Taur, Y., Jenq, R. R., Equinda, M. J., Son, T., Samstein, M., et al. (2010). Vancomycin-resistant Enterococcus domination of intestinal microbiota is enabled by antibiotic treatment in mice and precedes bloodstream invasion in humans. J. Clin. Invest. 120, 4332–4341. doi: 10.1172/JCI43918
Ulett, G. C., Webb, R. I., and Schembri, M. A. (2006). Antigen-43-mediated autoaggregation impairs motility in Escherichia coli. Microbiology 152(Pt 7), 2101–2110. doi: 10.1099/mic.0.28607-0
Uruen, C., Chopo-Escuin, G., Tommassen, J., Mainar-Jaime, R. C., and Arenas, J. (2020). Biofilms as promoters of bacterial antibiotic resistance and tolerance. Antibiotics 10:3. doi: 10.3390/antibiotics10010003
van der Heijden, K. M., van der Heijden, I. M., Galvao, F. H., Lopes, C. G., Costa, S. F., Abdala, E., et al. (2014). Intestinal translocation of clinical isolates of vancomycin-resistant Enterococcus faecalis and ESBL-producing Escherichia coli in a rat model of bacterial colonization and liver ischemia/reperfusion injury. PLoS ONE 9:e108453. doi: 10.1371/journal.pone.0108453
Vernet, V., Philippon, A., Madoulet, C., Vistelle, R., Jaussaud, R., and Chippaux, C. (1995). Virulence factors (aerobactin and mucoid phenotype) in Klebsiella pneumoniae and Escherichia coli blood culture isolates. FEMS Microbiol. Lett. 130, 51–57. doi: 10.1111/j.1574-6968.1995.tb07697.x
Vlamakis, H., Chai, Y., Beauregard, P., Losick, R., and Kolter, R. (2013). Sticking together: building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 11, 157–168. doi: 10.1038/nrmicro2960
Vuotto, C., Longo, F., Pascolini, C., Donelli, G., Balice, M. P., Libori, M. F., et al. (2017). Biofilm formation and antibiotic resistance in Klebsiella pneumoniae urinary strains. J. Appl. Microbiol. 123, 1003–1018. doi: 10.1111/jam.13533
Walker, M. J., Hollands, A., Sanderson-Smith, M. L., Cole, J. N., Kirk, J. K., Henningham, A., et al. (2007). DNase Sda1 provides selection pressure for a switch to invasive group A streptococcal infection. Nat. Med. 13, 981–985. doi: 10.1038/nm1612
Wang, J., Cao, Y., Zhang, L., Liu, G., and Li, C. (2021). Pathogen distribution and risk factors for urinary tract infection in infants and young children with retained double-J catheters. J. Int. Med. Res. 49:03000605211012379. doi: 10.1177/03000605211012379
Whelan, S., Lucey, B., and Finn, K. (2023). Uropathogenic Escherichia coli (UPEC)-associated urinary tract infections: the molecular basis for challenges to effective treatment. Microorganisms 11:2169. doi: 10.3390/microorganisms11092169
White, A. P., Gibson, D. L., Kim, W., Kay, W. W., and Surette, M. G. (2006). Thin aggregative fimbriae and cellulose enhance long-term survival and persistence of Salmonella. J. Bacteriol. 188, 3219–3227. doi: 10.1128/JB.188.9.3219-3227.2006
Wiles, T. J., Kulesus, R. R., and Mulvey, M. A. (2008). Origins and virulence mechanisms of uropathogenic Escherichia coli. Exp. Mol. Pathol. 85, 11–19. doi: 10.1016/j.yexmp.2008.03.007
Wu, H., Moser, C., Wang, H. Z., Hoiby, N., and Song, Z. J. (2015). Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 7, 1–7. doi: 10.1038/ijos.2014.65
Wyres, K. L., Nguyen, T. N. T., Lam, M. M. C., Judd, L. M., van Vinh Chau, N., Dance, D. A. B., et al. (2020). Genomic surveillance for hypervirulence and multi-drug resistance in invasive Klebsiella pneumoniae from South and Southeast Asia. Genome Med. 12:11. doi: 10.1186/s13073-019-0706-y
Yue, W. W., Grizot, S., and Buchanan, S. K. (2003). Structural evidence for iron-free citrate and ferric citrate binding to the TonB-dependent outer membrane transporter FecA. J. Mol. Biol. 332, 353–368. doi: 10.1016/S0022-2836(03)00855-6
Zboromyrska, Y., de Cueto López, M., Alonso-Tarrés, C., and Sánchez-Hellín, V. (2019). “Diagnóstico microbiológico de las infecciones del tracto urinario”, in Procedimientos en Microbiología Clínica, eds. E. Cercenado Mansilla and R. Cantón Moreno: Sociedad Española de Enfermedades Infecciosas y Microbiología Clínica), 1–78.
Zou, Z., Potter, R. F., McCoy, W. H., Wildenthal, J. A., Katumba, G. L., Mucha, P. J., et al. (2023). E. coli catheter-associated urinary tract infections are associated with distinctive virulence and biofilm gene determinants. JCI Insight 8:1461. doi: 10.1172/jci.insight.161461
Keywords: uropathogens, catheters, biofilm, virulence, bacterial interaction, antibiotic resistance
Citation: Farizano JV, Castagnaro E, Arroyo-Egea JT, Aparicio JD, Vallejos AC, Hebert EM, Saavedra L, Rapisarda VA, Villegas JM and Grillo-Puertas M (2025) Virulence traits and bacterial interactions within the complex microbial population in urinary double-J catheters. Front. Microbiol. 16:1624743. doi: 10.3389/fmicb.2025.1624743
Received: 07 May 2025; Accepted: 11 June 2025;
Published: 10 July 2025; Corrected: 13 August 2025.
Edited by:
Zhangnv Yang, Zhejiang Center for Disease Control and Prevention (Zhejiang CDC), ChinaReviewed by:
Parisa Noorian, University of Technology Sydney, AustraliaNayab Batool, University of Agriculture, Faisalabad, Pakistan
Benjamin Hunt, University at Buffalo, United States
Copyright © 2025 Farizano, Castagnaro, Arroyo-Egea, Aparicio, Vallejos, Hebert, Saavedra, Rapisarda, Villegas and Grillo-Puertas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariana Grillo-Puertas, bWFyaWFuYS5ncmlsbG9AZmJxZi51bnQuZWR1LmFy