 Jiaqiang Wang
Jiaqiang Wang Chien-shan Cheng2,3†
Chien-shan Cheng2,3†- 1Department of Anesthesiology, The Obstetrics and Gynecology Hospital of Fudan University, Shanghai, China
- 2Department of Integrative Oncology, Fudan University Shanghai Cancer Center, Shanghai, China
- 3Department of Oncology, Shanghai Medical College, Fudan University, Shanghai, China
- 4Department of Anesthesiology, Shanghai First Maternity and Infant Hospital, Tongji University School of Medicine, Shanghai, China
Volatile anesthetics are widely used inhalation anesthetics in clinical anesthesia. In recent years, the regulation of anti-cancer relevant signaling of volatile anesthetics has drawn the attention of investigators. However, their underlying mechanism remains unclear. This review summarizes the research progress on the regulation of anti-cancer relevant signaling of volatile anesthetics, including sevoflurane, desflurane, xenon, isoflurane, and halothane in vitro, in vivo, and clinical studies. The present review article aims to provide a general overview of regulation of anti-cancer relevant signaling and explore potential underlying molecular mechanisms of volatile anesthetics. It may promote promising insights of guiding clinical anesthesia procedure and instructing enhance recovery after surgery (ERAS) with latent benefits.
Introduction
Cancer describes diseases characterized by uncontrolled cell division and tissue invasion. Cancer hallmarks include maintaining proliferation signals, evading cell death, resisting treatment, enabling invasion, inducing angiogenesis, and activating metastasis (1). Cancer treatment strategies include traditional methods, such as surgery, chemo- and radio-therapy; and newer methods such as a ligand or receptor-based target therapy, stem cell therapy, and various forms of novel drug delivery systems (2). Until now, cancer is still an insurmountable problem worldwide, leading to high morbidity and mortality (3).
Volatile anesthetics, including sevoflurane, desflurane, xenon, isoflurane, halothane and others, are used for inhalational anesthesia in clinical practice. Volatile anesthetics target specific central nervous system receptors to perform anesthetic functions, such as the neuronal GABAA receptor, NMDA receptor and glutamate receptor subtypes. Volatile anesthetics can also affect cells by changing transcriptional elements, thereby changing specific characteristics of cell function. Previous studies have shown that volatile anesthetics have organ protection effects (4, 5). Recently, researchers have focused on the regulation of anti-cancer relevant signaling of volatile anesthetics on different kinds of cancer in vitro, in vivo, and clinical studies, but the specific mechanism remains unclear.
Everything has two sides. Some studies reported that sevoflurane (6–12), isoflurane (7, 10, 11) and halothane (13–15) may play tumor-promoting effects. We compared the articles and found that the controversy may come from different cancer types, cell lines, incubation concentrations and other conditions. More research on differences should be studied to discover potential conditions for volatile anesthetics and suitable cancer types. Analysis articles aimed at comparing opposite findings are also welcome. More in vivo and clinical studies should be conducted to further determine the regulation of anti-cancer relevant signaling of volatile anesthetics to guide clinical anesthesia procedures.
This review summarized the regulation of anti-cancer relevant signaling, including anti-proliferation, anti-migration and invasion, anti-metastasis, apoptosis-inducing effects, and the underlying mechanisms of volatile anesthetics. It may be instructive for future clinical inhalation anesthesia and beneficial for ERAS.
Regulation of Anti-Cancer Relevant Signaling
Sevoflurane
Sevoflurane (C4H3F7O, Table 1) is one of the most commonly used volatile anesthetics. It is a colorless and sweet-smelling inhalation anesthetic used to induce and maintain general anesthesia. For induction and maintenance of general anesthesia, sevoflurane concentration ranges from 0.5%–5% and less than 4%, respectively. In the electrophysiological study of neurons and recombinant receptors, sevoflurane is a positive allosteric modulator of the GABAA receptors (16–18). However, it can also act as a NMDA receptor antagonist (19), enhancing glycine receptor electro-currents (20) and inhibiting nAChR (21) and 5-HT3 receptor currents (22, 23). Sevoflurane is particularly non-irritating to the respiratory tract, so it is particularly suitable for asthma patients’ anesthesia.
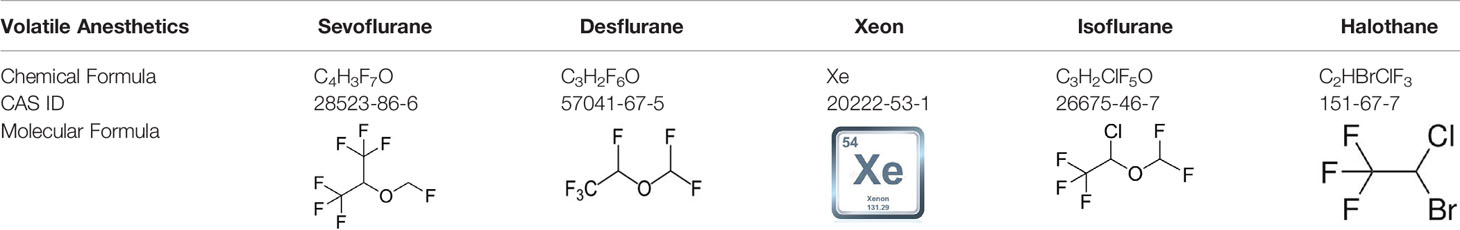
Table 1 The volatile anesthetics.
Sevoflurane and miRNAs
miRNAs are single-stranded, highly conserved small non-coding RNAs. More and more studies have shown that miRNAs affect cancer proliferation, metastasis, and invasion. In addition, miRNA expression can also determine the pathogenesis, diagnosis, and diseases prognosis of cancer (24). Recently, miRNAs are proposed to function both as oncogenes and tumor suppressors by regulating various target gene expressions (25, 26). Oncogenes are genes that may cause cancer and are often mutated and expressed at high cancer cell levels. On the other hand, tumor suppressor genes, or anti-oncogenes, are genes that protect cells from malignant transformation (27). Recent studies have found that Sevoflurane can regulate miRNA expression.
miR-203 has been implicated to play an essential role cancer proliferation regulation and are of potential diagnostic value. It is reported that miR-203 can act both as an oncogene and tumor suppressor gene in the development of different types of cancers (28–30). Fan and colleagues treated colorectal cancer cells (CRC, cell lines: SW620 and HCT116) with 1%, 2%, and 4% sevoflurane for 6 h to investigate the regulation of anti-cancer signaling of sevoflurane in CRC cell lines. The study demonstrated a proliferation suppression effect of sevoflurane, along with its migration and invasion inhibitory effects, by regulating the ERK/MMP-9 pathway through miR-203/Robo1 (31). In another study, MDA-MB-231 and MCF-7 breast cancer cells were exposed to 2% sevoflurane for 6 h. Results demonstrated that clinical concentration of sevoflurane could significantly suppress the proliferation of breast cancer cells via up-regulation of miR-203 (32).
miRNA-637 is as a tumor suppressor effect and plays crucial role in carcinogenesis and cancer progression (33–35). Emerging evidence suggested that miRNA-637 regulates the migration and invasion of glioma cells (36). In glioma in vitro models, U251 cells were treated with sevoflurane (1.7%, 3.4%, 5.1%) for 6 h. Yi and colleagues reported that sevoflurane inhibited glioma migration and invasion by up-regulating miRNA-637 and suppression of downstream Akt1 expression and activity (37).
In lung cancer, the regulation of anti-cancer signaling of sevoflurane by regulating miRNA has also been investigated (38). A study aimed at elucidating sevoflurane’s effect on the miRNA in lung cancer cells showed that A549 cells pretreated with 3% sevoflurane for 0.5 h caused an increase in apoptosis, thereby significantly reduced the risk of cancer cell metastasis and improving patients’ postoperative survival rate. Sevoflurane pretreatment up-regulated tumor suppressor miRNA-21, miRNA-221 and down-regulated oncogenic miRNA-34a in A549 cancer cells (39).
miR-124 is widely expressed in the nervous system (40). Rho-associated coiled-coil–containing protein kinase (ROCK1) plays essential roles in regulating tumorigenesis, cell apoptosis, invasion and migration (41). As reported by Cao et al., 4.1% sevoflurane pretreatment for 4 h inhibits glioma proliferation, invasion, and metastasis in U251and U87 cells through enhancing miR-124-3p level, thereby suppressed tumor malignancy-related ROCK1 signaling pathway (42).
Sevoflurane and MMPs
Matrix metalloproteinases (MMPs) are proteolytic enzymes that contribute to the degradation of extracellular matrix and basement membrane and are associated with cancer cell invasion. Among them, MMP-2 and MMP-9 are remarkably up-regulated in malignant tumors and contribute to cancer invasion (43).
Compared with normal brain tissue, MMP-2 is highly expressed in gliomas. MMP-2 has shown multiple effects in tumor progression, and promoted glioma malignancies (44, 45). Research by Hurmath et al. suggested that 2.5% sevoflurane incubation for 1.5 h suppressed the migration capability of U87MG glioma cells by down-regulation MMP-2 activity (46).
Degrading extracellular matrix is considered first step of tumor cell progression. Prior to tumor invasion into blood vessels or lymph nodes, tumor cells degrade the extracellular matrix, such as MMP-9 (47). Research evidence showed that in an in vitro reperfusion injury model, preconditioning with 2.2% sevoflurane for 45 min reduced MMP-9 release from human neutrophils by interfering with its downstream CXCR2 and its upstream PKC. By down-regulating MMP-9, sevoflurane suppressed MC-38 colon cancer cells migration (48).
Sevoflurane also demonstrated growth and invasion inhibitory effects in lung adenocarcinoma A549 cell line (49). The mechanism of its growth inhibition may be related to the synergistic down-regulation of X-linked inhibitor of apoptosis (XIAP) and survivin. Furthermore, its the synergistic effect of invasion inhibition may be related to the down-regulation of MMP-2 and MMP-9.
Sevoflurane and Cell Cycle
Abnormal cell proliferation is most related to the influence of cell cycle regulation (50). 2.5% Sevoflurane treatment for 4 h significantly inhibited A549 cells’ proliferation, invasion and induce cellular apoptosis and arrest the cell cycle at the G2/M phase (51). Furthermore, 2% sevoflurane preconditioning for 6 h possessed anti-proliferative and pro-apoptotic effects, possibly related to the down-regulation of XIAP and survivin expression and caspase-3 activation. Cell cycle arrest in the G2/M phase is associated with the down-regulation of cyclin A, cyclin B1, and cdc2 kinase expression. Sevoflurane can significantly inhibit breast cancer cells’ proliferation by blocking the cell cycle in the G1 phase (32).
Sevoflurane and Cell Apoptosis
In colonic cancer, 3% sevoflurane incubation for 1 or 2 h induced late apoptosis in Caco-2 cells in vitro (52). The study reported that sevoflurane intervention increases CYP2E1, caspase-3, and p53 expression. Furthermore, sevoflurane also facilitates an early increase of de novo ceramide synthesis. These results suggested that sevoflurane acts on both signaling pathways and metabolic pathways in vitro.
In neck squamous cell carcinoma (HNSCC) cancer, 2% and 4% sevoflurane pretreatment for 2, 4, 6, and 8 h inhibited proliferation, invasion, migration, and induced cellular apoptosis of FaDu and CAL-27 cell lines (53). The anti-proliferation effect of sevoflurane was associated with the downregulation p-Akt expression, and the cell apoptosis effect was associated HIF-1α activation, which regulated the Fas/FasL signaling pathway.
Sevoflurane and HIF-1α
The HIF is a family of transcription factors that involved in crucial aspects of cancer biology, such as cell proliferation, angiogenesis, metabolomic adaptation, and metastasis (54). Sevoflurane preconditioning (1.5%, 2.5%, or 3.5% sevoflurane incubation of A549 cells for 4 h) can inhibit the proliferation and invasion of lung cancer A549 cells induced by hypoxia, which may be related to the down-regulation of HIF-1α and its downstream genes XIAP, survivin, fascin, and HPA (55). Under hypoxia conditions, HIF-1α activation is dependent on the activation of the p38 MAPK signaling pathway. Also, the study proved that sevoflurane partially reversed the hypoxia induced p38 MAPK activity.
Activation of HIF-1α by sevoflurane regulates the Fas/FasL signaling pathway to exert cell apoptosis as demonstrated above (43).
Sevoflurane and VEGF
Hypoxia regulates transcriptional factor HIF-1, which regulates hypoxia-inducible angiogenic factor VEGF (56). VEGF is an important survival factor for endothelial and tumor cells. In tongue squamous cell carcinoma cell (TSCC), SCC-4 cells incubated with 4.1% sevoflurane for 24 and 72 h was shown attenuated VEGF level via increasing the DNA methylation of the VEGF promoter region in vitro (37).
Sevoflurane and Wnt/β-Catenin Signaling
Sevoflurane was found to significantly inhibited the growth of a panel of chronic myeloid leukemia (CML) cell lines (KCL22, K562, KU812, LAMA84 and KBM-7) (57). It also inhibited proliferation, differentiation and self-renewal capacities of CML CD34 cells. Mechanistically, it is purposed that 2%, 4%, or 8% sevoflurane preconditioning for 24 h dose-dependently decreases β-catenin and c-Myc expressions and activities in K562 and CML CD34 cells. The findings also reveal the Wnt/β-catenin pathway may be important targets of volatile anesthetics in cancer treatment.
Sevoflurane and Platelets Activation
It has been demonstrated that activated platelets contribute to tumor cells’ metastatic ability and protect circulating tumor cells from immune cells (58, 59). Furthermore, surgery stress potentiates platelets activation. Thus, a promising therapeutic strategy of preventing platelets-induced metastasis during cancer surgery procedure is much needed. Previous study suggested that sevoflurane attenuates platelet activation in lung cancer patients by reducing GPIIb/IIIa, CD62P, and PAR levels and these effects are further validated in vitro. It is indicated that sevoflurane at 1 MAC reduces platelets-induced invasive potential of lung cancer A549 cells through decreasing platelets activity (60).
Desflurane
Desflurane (C3H2F6O, Table 1) is widely used for anesthesia maintenance in contemporary clinical work. Characterized by low blood solubility, it functions as the fastest in acting and revival of volatile anesthetics. Desflurane is prohibited for anesthesia induction in infants and young children due to its potential of causing adverse reactions.
Desflurane and MMPs
In an in vitro reperfusion injury model, MC-38 colon cancer cells were incubated with 6% of desflurane for 45 min. It was demonstrated that desflurane could reduce the deliverance of MMP-9 by intervening downstream of the CXCR2 pathway. By down-regulating MMP-9, desflurane reduced the degradation of matrigel and the migration of colorectal cancer cells (48).
Desflurane and DFS
One study conducted a historical cohort study in which all patients received the primary cytoreductive surgery for stage III epithelial ovarian cancer, and the evaluation factor was disease-free survival (DFS). Studies have found that, compared with other volatile anesthetics, desflurane decreased the relative risk of cancer recurrences and is associated with improved DFS after surgery (61).
Desflurane and the Immune System
According to a randomized trial, during the perioperative period, desflurane anesthesia for breast cancer surgery can induce an adequate immune response in terms of maintaining the ratio of CD4(+)/CD8(+) T cells (62). Regarding leukocytes and NK cells, desflurane anesthesia’s adverse immune response is less than that of propofol.
Xenon
Xenon (Xe, Table 1), is the most stable gas of noble gas, which targets the glycine binding site of the NMDA receptor and the KATP channel. Xenon gas can dissolve in the fat of cells, causing cell anesthesia and swelling, thereby temporarily stopping the function of nerve endings. Owing to that xenon does not increase the sensitivity of myocardium to catecholamines-induced arrhythmia and it inhibits myocardial contraction but with minimal inhibitory effect on cardiovascular function, xenon is suitable for and widely used in cardiovascular surgery.
Xenon and RANTES
Regulated on activation, normal T cell expressed and secreted (RANTES), also known as CCL5 and functioning on receptor CCR5, is a cytokine that continues to increase in breast cancer subtypes (63) and is associated with promoting breast cancer metastasis and progression (64, 65). Ash et al. investigated the effect of xenon on migration and oncogene expression in human breast adenocarcinoma cells (66). It was demonstrated that 70% xenon incubation for 20 min inhibited the migration of estrogen receptor-positive (MCF-7) and negative (MDA-MB-231) breast cancer cells and reduce the pro-angiogenic factor’s release.
Xenon and PMCA
It is reported that xenon, at partial pressures ranging from 0.5 to 1.5 atm (equivalent to 0.5 to 1.6 MAC) for 30 min, can inhibit the pumping of plasma membrane Ca2+-ATPase (PMCA) in synaptic plasma membrane vesicles in rat C6 glioma cells. This mechanism may inhibit the physiological functions of cancer cells (67).
Owning to the inertness of xenon, it can only be extracted and liquefied, but cannot be synthesized artificially. Therefore, it is costly to utilize xenon. However, with the development of novel manufactured techniques, xenon is gradually adopted in various countries. Although it has not been used clinically, it is suggested that xenon may regulate anti-cancer relevant signaling and is worthy of further exploration.
Isoflurane
Isoflurane (C3H2ClF5O, Table 1) is colorless and of pungent odor. It is used for anesthesia maintenance and has the properties of reducing pain sensitivity and relaxing muscles. Isoflurane may bind to GABA and Glycine receptors, but has different effects. However, the clinical application of isoflurane is gradually replaced by sevoflurane and desflurane due to its potential complication of inducing epileptiform EEG.
Isoflurane and Glutamate
Glutamate is the primary excitatory neurotransmitter and acts as an effective neurotoxin when overexcited. Therefore, the extracellular glutamate concentration must be kept low to carry out neurotransmission and prevent damage effectively. Isoflurane incubation of C6 glioma cells can increase the expression and activity of type 3 excitatory amino acid transporter (EAAT3) through a pathway that depends on PKC and PI3K, thereby exhibiting higher glutamine in a time- and concentration-dependent manner (68). In addition, it has been reported that potential treatments targeting glutamine metabolism can be used to treat many types of cancer (69).
Isoflurane and COX-2
Cyclooxygenase (COX), also known as prostaglandin H2 (PGH2) synthase, is an essential enzyme for converting arachidonic acid to PGH2. Studies have shown that inhibition of COX is related to tumor behavior. A research report pointed out that 1.4% isoflurane treatment for 0.5 h significantly reduced the enhancement of COX-2 and the release of PGE2 of human laryngeal papilloma cells. By inhibiting the activation of p38 MAPK, isoflurane inhibited cell proliferation and apoptosis evasion (70).
Isoflurane and Cell Apoptosis
An in vitro and in vivo study reported that isoflurane incubation (2 mg/ml, 48 h) could not only inhibited liver cancer growth, but also decreased cell viability in liver cancer patient. The specific mechanism involves upregulating the expression levels of proapoptotic genes (caspase-3 and caspase-8) and downregulating anti-apoptotic (Bcl-2 and Bax) mRNA expression. Furthermore, isoflurane treatment inhibited migration and invasion of hepatic carcinoma cells. The molecular mechanisms underlying the tumor aggressiveness suppressive role of isoflurane involved regulation of NF-κB activity, and the PI3K/AKT signaling pathway (71).
Halothane
Halothane (C2HBrClF3, Table 1) is liquid anesthetic with colorless, clear, volatile and scented properties. It is unstable in nature and can be slowly decomposed by light and heat. Similar to other volatile anesthetics, halothane performs its anesthetic function by activating GABA-A, glycine, and NMDA receptors (72–74).
Halothane and Energy Metabolism
In the presence of halothane, glucose uptake and lactate output increase and oxygen consumption is inhibited, 0.9% halothane incubation showed 50% inhibition in the heteroploid strain and 0.35% halothane in the mouse sarcoma I strain. Also, population growth and high-energy phosphate production are diminished. A variety of biochemical mechanisms implicating the mitochondrial mechanisms may be involved (75).
Halothane and DNA or RNA Synthesis
Jackson et al. reported that halothane treatment varied in 0.1%–5.0% for 24 h was found to inhibit cell multiplication and cell growth in rat hepatoma cells, with 2.5% halothane pretreatment for 6 h being the most significant (76). Another study suggested that 0.1%–5.0% halothane preconditioning for 2 h inhibited the incorporation of extracellular thymidine into DNA, thus inhibiting DNA synthesis on hepatoma HTC cells (77).
Studies on cytotoxicity and anti-proliferative effects indicate that the anti-tumor ability of inhaled anesthetics may be halothane> sevoflurane> isoflurane. In human colon cancer (Caco-2), laryngeal cancer (HEp-2), and poorly differentiated cells from lymph node metastasis of colon carcinoma (SW-620), 1.5% halothane preconditioning for 2, 4, and 6 h showed significantly growth inhibitory effect. Among the cell lines studied, halothane significantly reduced the DNA and RNA synthesis in Caco-2 and Hep-2 cells. Furthermore, decrease in DNA, RNA and protein synthesis were observed in Caco-2 and Hep-2 cells. In SW620 cells, protein synthesis were decreased. A DNA fragmentation was observed in MIA PaCa-2 and Caco-2 cells (78).
Halothane and Morphology
In a study, cultured neuroblastoma cells were incubated of halothane (100 or 1,000 ppm) for 4, 12, or 24 h in vitro. Exposure to halothane resulted in significant changes in the actin distribution pattern of neuroblastoma cells, and the cells exhibited characteristic morphological changes (79).
A clinical study showed that the type of anesthesia influenced the end results of therapy of cancer patients, the survival rates of patients receiving halothane anesthesia were much higher than ether-anaesthetized ones. The mechanism may involve influences of anesthetics on the pituitary-adrenal cortex system and carcinoma development and the role of immunity in tumor cell implantation and growth of metastases (80).
It is reported that 1.5 and 2.21 mM halothane incubation induced genotoxic and cytotoxic effects in lung cancer A549 cells in vitro. Consequences of the regulation of anti-cancer relevant effects involved reducing cell viability, inhibiting mitotic activity, and destroying the nucleus and nucleolus structure (81).
Investigations of volatile anesthetics on cellular morphological differentiation unveiled that 0.3%–2.1% halothane preconditioning up to 72 h inhibited neuroblastoma cells (clone NB2a) differentiation, the inhibition of neurite extension dose dependently and virtually abolished microspike formation even at the lowest concentration incubated. The mechanism was demonstrated that halothane inhibited neurite extension and abolished microspike formation (82).
Halothane and ICAM-1
Intercellular adhesion molecule 1 (ICAM-1) is a cell surface glycoprotein commonly expressed on endothelial cells and immune cells. Recent studied have reported that ICAM-1 is expressed in several tumors, and high expression has been positively correlated with metastatic potential. In human melanoma SK-MEL-37 cells, 4% halothane incubation for 3, 6, 12, or 24 h was demonstrated to perform lower ICAM-1 expression. Thus it was concluded that halothane possessed tumor metastasis inhibiting property by down-regulating ICAM-1 expression in vitro (83).
Halothane and PMCA
The plasma membrane Ca2+-ATPase (PMCA), is a ubiquitously expressed Ca2+ pump that releases Ca2+ from the calcium reservoir to the cytoplasm, regulating physiological functions including cell movement, growth and differentiation. Halothane, at concentration ranging from 0.5% to 1.75% (0.5 to 1.6 MAC), significantly inhibited plasma membrane vesicles Ca2+ uptake dose-dependently in rat C6 glioma cells, B104 neuroblastoma cells and PC12 pheochromocytoma cells (67).
Discussion
Recent studies have shown that volatile anesthetics regulate of anti-cancer relevant signaling in human cancers. Specifically, exposure to volatile anesthetics can change the biological response of cancer cells or regulate the gene expression of cancer cells, thereby exerting apoptosis induction, anti-invasion, anti-migration and other anti-cancer properties. There are many studies on the regulate of anti-cancer relevant signaling of sevoflurane, but more research on desflurane, which is also commonly used in clinical practice, is needed. As for the promising new type of inhaled anesthesia xenon, due to the difficulty of production and high price, it has not yet been widely used in clinical practice. The prospect of scientific research is worth exploring. There lacks of research articles concerning the regulation of anti-cancer relevant signaling of enflurane, methoxyhalothane, and ether. These documents are no longer in clinical use, so we did not discuss them in this review.
Although volatile anesthetics are not traditionally regarded as anti-cancer drugs, more and more research have focused on the potential anti-cancer properties. Volatile anesthetics mainly act on NMDA and GABA receptors. Although it is still unclear why anesthetics can regulate of anti-cancer relevant signaling, it has been reported that activated receptors can exert regulate anti-cancer-related signaling in cancer cells (84–87). Therefore, studying the regulation of anti-cancer relevant signaling of volatile anesthetics and their related receptors is a new and enlightening insight with important significance, and therefore may make outstanding contributions to cancer biology. Considering different types of cancer have different sensitivity to volatile anesthetics, this current review may guide the choice of volatile anesthetics to best improve the clinical prognosis of cancer patients and improve their postoperative recovery (ERAS).
The shortcomings of contemporary researches relatively lack of animal studies, clinical trials, genomics analysis and big data analysis. Volatile anesthetics exert anesthetic functions via passing through the respiratory tracts and blood-brain barrier and then acting on the receptors. Do volatile anesthetics demonstrate exceptional sensitivity of anti-cancer relevant signaling in lung cancer and brain tumor? More specific and compelling trials are needed, especially those related to sevoflurane and desflurane which is clinically widely used, to clarify the relationship between anesthetics and tumor prognosis, and to provide more precise guidance for anesthesia management.
Conclusion
From the above research and investigation, it can be concluded that volatile anesthetics could regulate anti-cancer relevant signaling (Table 2). The underlying mechanism involves miRNA, transcription factors, apoptotic pathway, MMP, etc. Figure 1. Although the current research may shortcomings, more in depth studies, especially clinical research, is warranted to clarify the regulation of anti-cancer relevant signaling of volatile anesthetics.
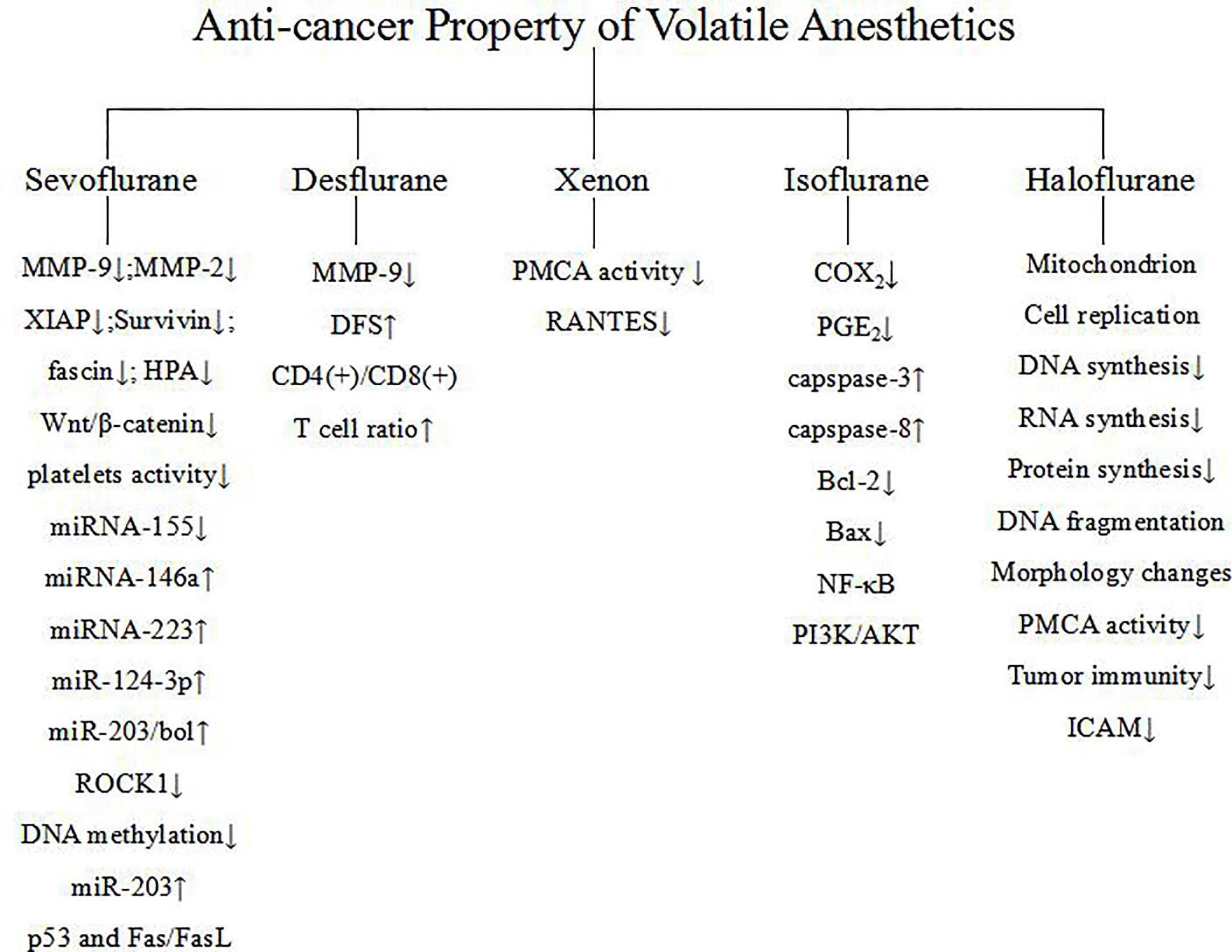
Figure 1 Mechanisms of regulation of anti-cancer relevant signaling of volatile anesthetics.
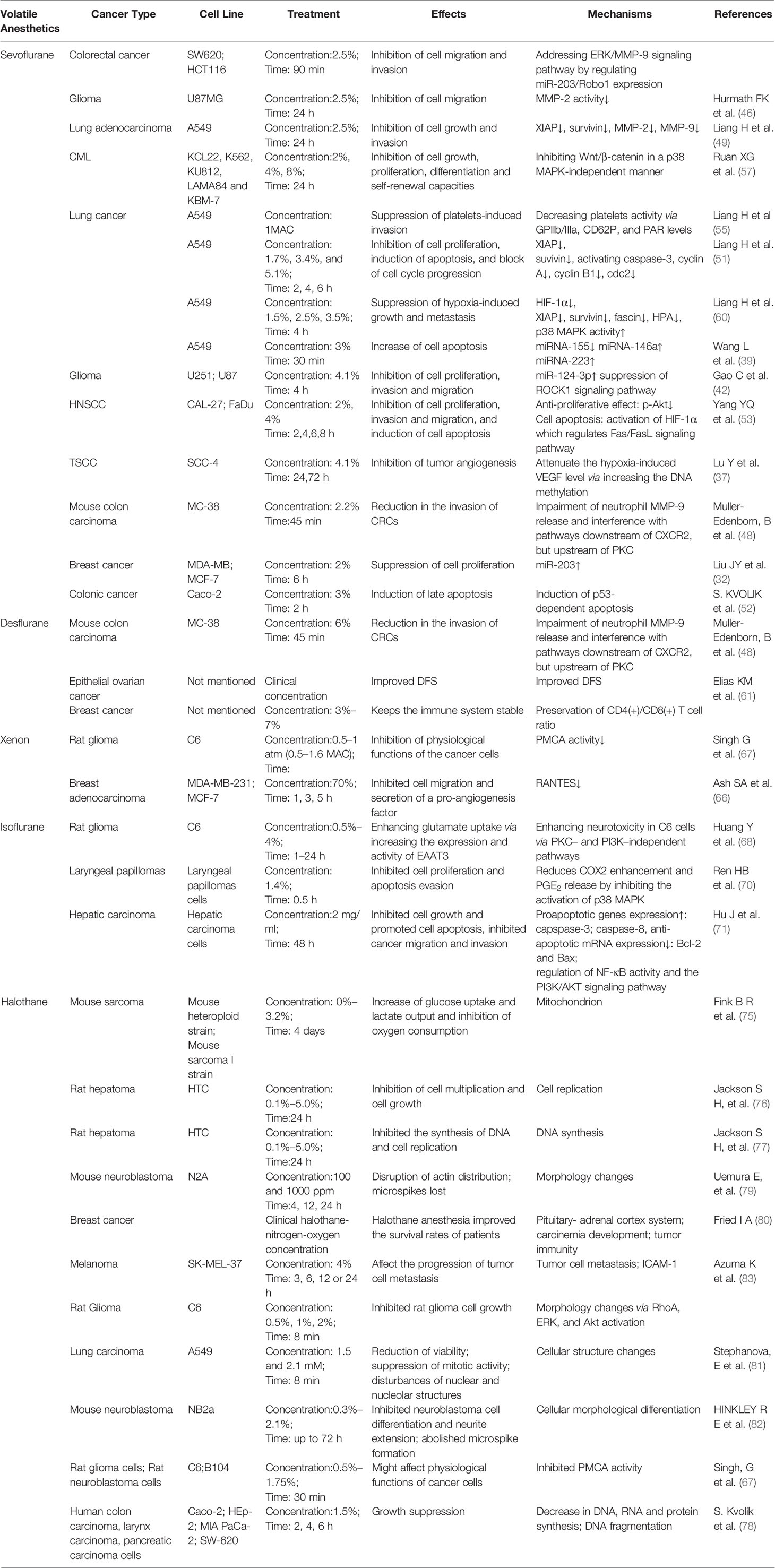
Table 2 Regulation of anti-cancer relevant signaling of volatile anesthetics.
Author Contributions
JW was in charge of the writing. C-sC was responsible for the pictures and editing. YL compiled the table and inserted the references. SS reconstructed and redesigned the work. SH made final agreement and approval of the work to be published. All authors contributed to the article and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell (2011) 144(5):646–74. doi: 10.1016/j.cell.2011.02.013
2. Mitra AK, Agrahari V, Mandal A, Cholkar K, Natarajan C, Shah S, et al. Novel delivery approaches for cancer therapeutics. J Control Release (2015) 219:248–68. doi: 10.1016/j.jconrel.2015.09.067
3. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin (2019) 69(1):7–34. doi: 10.3322/caac.21551
4. Wu L, Zhao H, Wang T, Pac-Soo C, Ma D. Cellular signaling pathways and molecular mechanisms involving inhalational anesthetics-induced organoprotection. J Anesth (2014) 28(5):740–58. doi: 10.1007/s00540-014-1805-y
5. Luo Y, Ma D, Ieong E, Sanders RD, Yu B, Hossain M, et al. Xenon and sevoflurane protect against brain injury in a neonatal asphyxia model. Anesthesiology (2008) 109(5):782–9. doi: 10.1097/ALN.0b013e3181895f88
6. Zhu M, Li M, Zhou Y, Dangelmajer S, Kahlert UD, Xie R, et al. Isoflurane enhances the malignant potential of glioblastoma stem cells by promoting their viability, mobility in vitro and migratory capacity in vivo. Brit J Anaesth (2016) 116(6):870–7. doi: 10.1093/bja/aew124
7. Zhang W, Shao X. Isoflurane Promotes Non-Small Cell Lung Cancer Malignancy by Activating the Akt-Mammalian Target of Rapamycin (mTOR) Signaling Pathway. Med Sci Monit (2016) 22:4644–50. doi: 10.12659/MSM.898434
8. Luo X, Zhao H, Hennah L, Ning J, Liu J, Tu H, et al. Impact of isoflurane on malignant capability of ovarian cancer in vitro. Brit J Anaesth (2015) 114(5):831–9. doi: 10.1093/bja/aeu408
9. Huang H, Benzonana LL, Zhao H, Watts HR, Perry NJS, Bevan C, et al. Prostate cancer cell malignancy via modulation of HIF-1α pathway with isoflurane and propofol alone and in combination. Br J Cancer (2014) 111(7):1338–49. doi: 10.1038/bjc.2014.426
10. Benzonana LL, Perry NJ, Watts HR, Yang B, Perry IA, Coombes C, et al. Isoflurane, a commonly used volatile anesthetic, enhances renal cancer growth and malignant potential via the hypoxia-inducible factor cellular signaling pathway in vitro. Anesthesiology (2013) 119(3):593–605. doi: 10.1097/ALN.0b013e31829e47fd
11. Kawaraguchi Y, Horikawa YT, Murphy AN, Murray F, Miyanohara A, Ali SS, et al. Volatile Anesthetics Protect Cancer Cells against Tumor Necrosis Factor-related Apoptosis-inducing Ligand-induced Apoptosis via Caveolins. Anesthesiology (2011) 115(3):499–508. doi: 10.1097/ALN.0b013e3182276d42
12. Jun R, Gui-he Z, Xing-xing S, Hui Z, Li-xian X. Isoflurane enhances malignancy of head and neck squamous cell carcinoma cell lines: A preliminary study in vitro. Oral Oncol (2011) 47(5):329–33. doi: 10.1016/j.oraloncology.2011.03.002
13. Moudgil GC, Singal DP. Halothane and isoflurane enhance melanoma tumour metastasis in mice. Can J Anaesth (1997) 44(1):90–4. doi: 10.1007/BF03014331
14. Melamed R, Bar-Yosef S, Shakhar G, Shakhar K, Ben-Eliyahu S. Suppression of Natural Killer Cell Activity and Promotion of Tumor Metastasis by Ketamine, Thiopental, and Halothane, but Not by Propofol: Mediating Mechanisms and Prophylactic Measures. Anesth Analg (2003) 97(5):1331–9. doi: 10.1213/01.ANE.0000082995.44040.07
15. Shapiro J, Jersky J, Katzav S, Feldman M, Segal S. Anesthetic Drugs Accelerate the Progression of Postoperative Metastases of Mouse Tumors. Survey Anesthesiol (1982) 26(4):231. doi: 10.1097/00132586-198226040-00041
16. Jenkins A, Franks NP, Lieb WR. Effects of temperature and volatile anesthetics on GABA(A) receptors. Anesthesiology (1999) 90(2):484–91. doi: 10.1097/00000542-199902000-00024
17. Wu J, Harata N, Akaike N. Potentiation by sevoflurane of the gamma-aminobutyric acid-induced chloride current in acutely dissociated CA1 pyramidal neurones from rat hippocampus. Br J Pharmacol (1996) 119(5):1013–21. doi: 10.1111/j.1476-5381.1996.tb15772.x
18. Krasowski MD, Harrison NL. The actions of ether, alcohol and alkane general anaesthetics on GABAA and glycine receptors and the effects of TM2 and TM3 mutations. Br J Pharmacol (2000) 129(4):731–43. doi: 10.1038/sj.bjp.0703087
19. Brosnan RJ, Thiesen R. Increased NMDA receptor inhibition at an increased Sevoflurane MAC. BMC Anesthesiol (2012) 12:9. doi: 10.1186/1471-2253-12-9
20. Schüttler J, Schwilden S. Modern Anesthetics. Springer-Verlag Berlin Heidelberg, Springer Science & Business Media (2008). 32 p.
21. Van Dort CJ. Regulation of Arousal by Adenosine A(1) and A(2A) Receptors in the Prefrontal Cortex of C57BL/6J Mouse. ProQuest (2008), 120.
22. Hang LH, Shao DH, Wang H, Yang JP. Involvement of 5-hydroxytryptamine type 3 receptors in sevoflurane-induced hypnotic and analgesic effects in mice. Pharmacol Rep (2010) 62(4):621–6. doi: 10.1016/s1734-1140(10)70319-4
23. Suzuki T, Koyama H, Sugimoto M, Uchida I, Mashimo T. The diverse actions of volatile and gaseous anesthetics on human-cloned 5-hydroxytryptamine3 receptors expressed in Xenopus oocytes. Anesthesiology (2002) 96(3):699–704. doi: 10.1097/00000542-200203000-00028
24. Hill M, Tran N. MicroRNAs Regulating MicroRNAs in Cancer. Trends Cancer (2018) 4(7):465–8. doi: 10.1016/j.trecan.2018.05.002
25. Zhang B, Pan X, Cobb GP, Anderson TA. microRNAs as oncogenes and tumor suppressors. Dev Biol (2007) 302(1):1–12. doi: 10.1016/j.ydbio.2006.08.028
26. Chen CZ. MicroRNAs as oncogenes and tumor suppressors. N Engl J Med (2005) 353(17):1768–71. doi: 10.1056/NEJMp058190
27. Wang J, Cheng CS, Lu Y, Ding X, Zhu M, Miao C, et al. Novel Findings of Anti-cancer Property of Propofol. Anticancer Agents Med Chem (2018) 18(2):156–65. doi: 10.2174/1871520617666170912120327
28. Gao G, Ge R, Li Y, Liu S. Luteolin exhibits anti-breast cancer property through up-regulating miR-203. Artif Cells Nanomed Biotechnol (2019) 47(1):3265–71. doi: 10.1080/21691401.2019.1646749
29. Zhang LS, Ma HG, Sun FH, Zhao WC, Li G. MiR-203 inhibits the malignant behavior of prostate cancer cells by targeting RGS17. Eur Rev Med Pharmacol Sci (2019) 23(13):5667–74. doi: 10.26355/eurrev_201907_18303
30. Chi Y, Jin Q, Liu X, Xu L, He X, Shen Y, et al. miR-203 inhibits cell proliferation, invasion, and migration of non-small-cell lung cancer by downregulating RGS17. Cancer Sci (2017) 108(12):2366–72. doi: 10.1111/cas.13401
31. Fan L, Wu Y, Wang J, He J, Han X. Sevoflurane inhibits the migration and invasion of colorectal cancer cells through regulating ERK/MMP-9 pathway by up-regulating miR-203. Eur J Pharmacol (2019) 850:43–52. doi: 10.1016/j.ejphar.2019.01.025
32. Liu J, Yang L, Guo X, Jin G, Wang Q, Lv D, et al. Sevoflurane suppresses proliferation by upregulating microRNA-203 in breast cancer cells. Mol Med Rep (2018) 18(1):455–60. doi: 10.3892/mmr.2018.8949
33. Leivonen SK, Sahlberg KK, Makela R, Due EU, Kallioniemi O, Borresen-Dale AL, et al. High-throughput screens identify microRNAs essential for HER2 positive breast cancer cell growth. Mol Oncol (2014) 8(1):93–104. doi: 10.1016/j.molonc.2013.10.001
34. Stokowy T, Wojtas B, Fujarewicz K, Jarzab B, Eszlinger M, Paschke R. miRNAs with the potential to distinguish follicular thyroid carcinomas from benign follicular thyroid tumors: results of a meta-analysis. Horm Metab Res (2014) 46(3):171–80. doi: 10.1055/s-0033-1363264
35. Zhang JF, He ML, Fu WM, Wang H, Chen LZ, Zhu X, et al. Primate-specific microRNA-637 inhibits tumorigenesis in hepatocellular carcinoma by disrupting signal transducer and activator of transcription 3 signaling. Hepatology (2011) 54(6):2137–48. doi: 10.1002/hep.24595
36. Que T, Song Y, Liu Z, Zheng S, Long H, Li Z, et al. Decreased miRNA-637 is an unfavorable prognosis marker and promotes glioma cell growth, migration and invasion via direct targeting Akt1. Oncogene (2015) 34(38):4952–63. doi: 10.1038/onc.2014.419
37. Lu Y, Wang J, Yan J, Yang Y, Sun Y, Huang Y, et al. Sevoflurane attenuate hypoxia-induced VEGF level in tongue squamous cell carcinoma cell by upregulating the DNA methylation states of the promoter region. BioMed Pharmacoth (2015) 71:139–45. doi: 10.1016/j.biopha.2015.02.032
38. Chen X, Hu Z, Wang W, Ba Y, Ma L, Zhang C, et al. Identification of ten serum microRNAs from a genome-wide serum microRNA expression profile as novel noninvasive biomarkers for nonsmall cell lung cancer diagnosis. Int J Cancer (2012) 130(7):1620–8. doi: 10.1002/ijc.26177
39. Wang L, Wang T, Gu J, Su H. Volatile anesthetic sevoflurane suppresses lung cancer cells and miRNA interference in lung cancer cells. Oncotarg Ther (2018) 11:5689–93. doi: 10.2147/OTT.S171672
40. An F, Gong G, Wang Y, Bian M, Yu L, Wei C. MiR-124 acts as a target for Alzheimer’s disease by regulating BACE1. Oncotarget (2017) 8(69):114065–71. doi: 10.18632/oncotarget.23119
41. Liu X, Kang J, Sun S, Luo Y, Ji X, Zeng X, et al. iASPP, a microRNA124 target, is aberrantly expressed in astrocytoma and regulates malignant glioma cell migration and viability. Mol Med Rep (2018) 17(1):1970–8. doi: 10.3892/mmr.2017.8097
42. Gao C, Shen J, Meng Z, He X. Sevoflurane Inhibits Glioma Cells Proliferation and Metastasis through miRNA-124-3p/ROCK1 Axis. Pathol Oncol Res (2019) 262:2. doi: 10.1007/s12253-019-00597-1
43. Kessenbrock K, Plaks V, Werb Z. Matrix metalloproteinases: regulators of the tumor microenvironment. Cell (2010) 141(1):52–67. doi: 10.1016/j.cell.2010.03.015
44. Choe G, Park JK, Jouben-Steele L, Kremen TJ, Liau LM, Vinters HV, et al. Active matrix metalloproteinase 9 expression is associated with primary glioblastoma subtype. Clin Cancer Res (2002) 8(9):2894–901. doi: 10.1159/000063870
45. Forsyth PA, Wong H, Laing TD, Rewcastle NB, Morris DG, Muzik H, et al. Gelatinase-A (MMP-2), gelatinase-B (MMP-9) and membrane type matrix metalloproteinase-1 (MT1-MMP) are involved in different aspects of the pathophysiology of malignant gliomas. Br J Cancer (1999) 79(11-12):1828–35. doi: 10.1038/sj.bjc.6690291
46. Hurmath FK, Mittal M, Ramaswamy P, Umamaheswara Rao GS, Dalavaikodihalli Nanjaiah N. Sevoflurane and thiopental preconditioning attenuates the migration and activity of MMP-2 in U87MG glioma cells. Neurochem Int (2016) 94:32–8. doi: 10.1016/j.neuint.2016.02.003
47. Nicoud IB, Jones CM, Pierce JM, Earl TM, Matrisian LM, Chari RS, et al. Warm hepatic ischemia-reperfusion promotes growth of colorectal carcinoma micrometastases in mouse liver via matrix metalloproteinase-9 induction. Cancer Res (2007) 67(6):2720–8. doi: 10.1158/0008-5472.CAN-06-3923
48. Muller-Edenborn B, Roth-Z’Graggen B, Bartnicka K, Borgeat A, Hoos A, Borsig L, et al. Volatile anesthetics reduce invasion of colorectal cancer cells through down-regulation of matrix metalloproteinase-9. Anesthesiology (2012) 117(2):293–301. doi: 10.1097/ALN.0b013e3182605df1
49. Liang H, Wang HB, Liu HZ, Wen XJ, Zhou QL, Yang CX. The effects of combined treatment with sevoflurane and cisplatin on growth and invasion of human adenocarcinoma cell line A549. BioMed Pharmacoth (2013) 67(6):503–9. doi: 10.1016/j.biopha.2013.03.005
50. Gerard C, Goldbeter A. The balance between cell cycle arrest and cell proliferation: control by the extracellular matrix and by contact inhibition. Interface Focus (2014) 4(3):20130075. doi: 10.1098/rsfs.2013.0075
51. Liang H, Gu MN, Yang CX, Wang HB, Wen XJ, Zhou QL. Sevoflurane inhibits proliferation, induces apoptosis, and blocks cell cycle progression of lung carcinoma cells. Asian Pac J Cancer Prev (2011) 12(12):3415–20. doi: 10.1159/000328275
52. Kvolik S, Dobrosevic B, Marczi S, Prlic L, Glavas-Obrovac L. Different apoptosis ratios and gene expressions in two human cell lines after sevoflurane anaesthesia. Acta Anaesth Scand (2009) 53(9):1192–9. doi: 10.1111/j.1399-6576.2009.02036.x
53. Yang Y, Hu R, Yan J, Chen Z, Lu Y, Jiang J, et al. Sevoflurane inhibits the malignant potential of head and neck squamous cell carcinoma via activating the hypoxiainducible factor-1alpha signaling pathway in vitro. Int J Mol Med (2018) 41(2):995–1002. doi: 10.3892/ijmm.2017.3306
54. Semenza GL. Targeting HIF-1 for cancer therapy. Nat Rev Cancer (2003) 3(10):721–32. doi: 10.1038/nrc1187
55. Liang H, Yang CX, Zhang B, Wang HB, Liu HZ, Lai XH, et al. Sevoflurane suppresses hypoxia-induced growth and metastasis of lung cancer cells via inhibiting hypoxia-inducible factor-1α. J Anesth (2015) 29(6):821–30. doi: 10.1007/s00540-015-2035-7
56. Guillemin K, Krasnow MA. The hypoxic response: huffing and HIFing. Cell (1997) 89(1):9–12. doi: 10.1016/S0092-8674(00)80176-2
57. Ruan X, Jiang W, Cheng P, Huang L, Li X, He Y, et al. Volatile anesthetics sevoflurane targets leukemia stem/progenitor cells via Wnt/β-catenin inhibition. BioMed Pharmacoth (2018) 107:1294–301. doi: 10.1016/j.biopha.2018.08.063
58. Miyashita T, Tajima H, Makino I, Nakagawara H, Kitagawa H, Fushida S, et al. Metastasis-promoting role of extravasated platelet activation in tumor. J Surg Res (2015) 193(1):289–94. doi: 10.1016/j.jss.2014.07.037
59. Goubran HA, Stakiw J, Radosevic M, Burnouf T. Platelet-cancer interactions. Semin Thromb Hemost (2014) 40(3):296–305. doi: 10.1055/s-0034-1370767
60. Liang H, Yang CX, Zhang B, Zhao ZL, Zhong JY, Wen XJ. Sevoflurane attenuates platelets activation of patients undergoing lung cancer surgery and suppresses platelets-induced invasion of lung cancer cells. J Clin Anesth (2016) 35:304–12. doi: 10.1016/j.jclinane.2016.08.008
61. Elias KM, Kang S, Liu X, Horowitz NS, Berkowitz RS, Frendl G. Anesthetic Selection and Disease-Free Survival Following Optimal Primary Cytoreductive Surgery for Stage III Epithelial Ovarian Cancer. Ann Surg Oncol (2015) 22(4):1341–8. doi: 10.1245/s10434-014-4112-9
62. Woo JH, Baik HJ, Kim CH, Chung RK, Kim DY, Lee GY, et al. Effect of Propofol and Desflurane on Immune Cell Populations in Breast Cancer Patients: A Randomized Trial. J Korean Med Sci (2015) 30(10):1503. doi: 10.3346/jkms.2015.30.10.1503
63. Gonzalez RM, Daly DS, Tan R, Marks JR, Zangar RC. Plasma biomarker profiles differ depending on breast cancer subtype but RANTES is consistently increased. Cancer Epidemiol Biomarkers Prev (2011) 20(7):1543–51. doi: 10.1158/1055-9965.EPI-10-1248
64. Lv D, Zhang Y, Kim HJ, Zhang L, Ma X. CCL5 as a potential immunotherapeutic target in triple-negative breast cancer. Cell Mol Immunol (2013) 10(4):303–10. doi: 10.1038/cmi.2012.69
65. Zhang Y, Lv D, Kim HJ, Kurt RA, Bu W, Li Y, et al. A novel role of hematopoietic CCL5 in promoting triple-negative mammary tumor progression by regulating generation of myeloid-derived suppressor cells. Cell Res (2013) 23(3):394–408. doi: 10.1038/cr.2012.178
66. Ash SA, Valchev GI, Looney M, Ni MA, Crowley PD, Gallagher HC, et al. Xenon decreases cell migration and secretion of a pro-angiogenesis factor in breast adenocarcinoma cells: comparison with sevoflurane. Br J Anaesth (2014) 113 Suppl 1:i14–21. doi: 10.1093/bja/aeu191
67. Singh G, Janicki PK, Horn JL, Janson VE, Franks JJ. Inhibition of plasma membrane Ca(2+)-ATPase pump activity in cultured C6 glioma cells by halothane and xenon. Life Sci (1995) 56(10):L219–24. doi: 10.1016/0024-3205(95)00011-T
68. Huang Y, Zuo Z. Isoflurane enhances the expression and activity of glutamate transporter type 3 in C6 glioma cells. Anesthesiology (2003) 99(6):1346–53. doi: 10.1097/00000542-200312000-00016
69. Choi YK, Park KG. Targeting Glutamine Metabolism for Cancer Treatment. Biomol Ther (Seoul) (2018) 26(1):19–28. doi: 10.4062/biomolther.2017.178
70. Ren H, Shi X, Li Y. Reduction of p38 mitogen-activated protein kinase and cyclooxygenase-2 signaling by isoflurane inhibits proliferation and apoptosis evasion in human papillomavirus-infected laryngeal papillomas. Exp Ther Med (2016) 12(5):3425–32. doi: 10.3892/etm.2016.3776
71. Hu J, Hu J, Jiao H, Li Q. Anesthetic effects of isoflurane and the molecular mechanism underlying isoflurane−inhibited aggressiveness of hepatic carcinoma. Mol Med Rep (2018) 18(1):184–92. doi: 10.3892/mmr.2018.8945
72. Olney JW, Farber NB, Wozniak DF, Jevtovic-Todorovic V, Ikonomidou C. Environmental agents that have the potential to trigger massive apoptotic neurodegeneration in the developing brain. Environ Health Perspect (2000) 108 Suppl 3:383–8. doi: 10.1289/ehp.00108s3383
73. Komatsu H, Nogaya J, Ogli K. Volatile anaesthetics as central nervous system excitants. Ann Acad Med Singapore (1994) 23(6 Suppl):130–8.
74. Brunner EA, Cheng SC, Berman ML. Effects of anesthesia on intermediary metabolism. Annu Rev Med (1975) 26:391–401. doi: 10.1146/annurev.me.26.020175.002135
75. Fink BR, Kenny GE. Metabolic effects of volatile anesthetics in cell culture. Anesthesiology (1968) 29(3):505–16. doi: 10.1097/00000542-196805000-00024
76. Jackson SH. The metabolic effects of halothane on mammalian hepatoma cells in vitro. I. Inhibition of cell replication. Anesthesiology (1972) 37(5):489–92. doi: 10.1097/00000542-197211000-00005
77. Jackson SH. The metabolic effects of halothane on mammalian hepatoma cells in vitro. II. Inhibition of DNA synthesis. Anesthesiology (1973) 39(4):405–9. doi: 10.1097/00000542-197310000-00013
78. Kvolik S, Glavas-Obrovac L, Bares V, Karner I. Effects of inhalation anesthetics halothane, sevoflurane, and isoflurane on human cell lines. Life Sci (2005) 77(19):2369–83. doi: 10.1016/j.lfs.2004.12.052
79. Uemura E, Levin ED. The effect of halothane on cultured fibroblasts and neuroblastoma cells. Neurosci Lett (1992) 145(1):33–6. doi: 10.1016/0304-3940(92)90196-E
80. Fried IA. The influence of the anaesthetic on survival rates of breast cancer patients after surgery. Int J Cancer (1977) 20(2):213–8. doi: 10.1002/ijc.2910200208
81. Stephanova E, Topouzova-Hristova T, Hazarosova R, Moskova V. Halothane-induced alterations in cellular structure and proliferation of A549 cells. Tissue Cell (2008) 40(6):397–404. doi: 10.1016/j.tice.2008.04.001
82. Hinkley RE, Telser AG. The effects of halothane on cultured mouse neuroblastoma cells. I. Inhibition of morphological differentiation. J Cell Biol (1974) 63(2 Pt 1):531–40. doi: 10.1083/jcb.63.2.531
83. Azuma K, Mike N, Fujiwara Y, Shimada Y, Watanabe T. Effect of halothane on intercellular adhesion molecule-1 (ICAM-1) in melanoma cells. J Anesth (1993) 7(4):442–6. doi: 10.1007/s0054030070442
84. Taylor RA, Watt MJ. Unsuspected Protumorigenic Signaling Role for the Oncometabolite GABA in Advanced Prostate Cancer. Cancer Res (2019) 79(18):4580–1. doi: 10.1158/0008-5472.CAN-19-2182
85. Chen X, Wu Q, You L, Chen S, Zhu M, Miao C. Propofol attenuates pancreatic cancer malignant potential via inhibition of NMDA receptor. Eur J Pharmacol (2017) 795:150–9. doi: 10.1016/j.ejphar.2016.12.017
86. Chen X WQSP. Propofol Disrupts Aerobic Glycolysis in Colorectal Cancer Cells via Inactivation of the NMDAR-CAMKII-ERK Pathway. Cell Physiol Biochem (2018) 462(2). doi: 10.1159/000488617
Keywords: volatile anesthetics, inhalation anesthesia, anti-cancer, volatile anesthesia, mechanism
Citation: Wang J, Cheng C-s, Lu Y, Sun S and Huang S (2021) Volatile Anesthetics Regulate Anti-Cancer Relevant Signaling. Front. Oncol. 11:610514. doi: 10.3389/fonc.2021.610514
Received: 26 September 2020; Accepted: 22 January 2021;
Published: 26 February 2021.
Edited by:
Jiang-Jiang Qin, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Cheng Zhou, Sichuan University, ChinaQiyang Shou, Zhejiang Chinese Medical University, China
Copyright © 2021 Wang, Cheng, Lu, Sun and Huang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shen Sun, c3Vuc2hlbjE5ODBAMTI2LmNvbQ==; Shaoqiang Huang, dGltcm9iYmluc0AxNjMuY29t
†These authors have contributed equally to this work