 Jinjia Fan1,2†
Jinjia Fan1,2† Feng Gao
Feng Gao Xiaobo Du
Xiaobo Du- 1Departmant of Oncology, National Health Commission Key Laboratory of Nuclear Technology Medical Transformation (Mianyang Central Hospital), Mianyang Central Hospital, School of Medicine, University of Electronic Science and Technology, Mianyang, China
- 2Department of Oncology, Affiliated Hospital of North Sichuan Medical College, Nan Chong, China
Radiation enteritis (Re) is one of the most common complications of radiation therapy for abdominal tumors. The efficacy of cancer treatment by radiation is often limited by the side effects of Re. Re can be acute or chronic. Treatment of acute Re is essentially symptomatic. However, chronic Re usually requires surgical procedures. The underlying mechanisms of Re are complex and have not yet been elucidated. The purpose of this review is to provide an overview of the pathogenesis of Re. We reviewed the role of intestinal epithelial cells, intestinal stem cells (ISCs), vascular endothelial cells (ECs), intestinal microflora, and other mediators of Re, noting that a better understanding of the pathogenesis of Re may lead to better treatment modalities.
Introduction
The intestine is particularly sensitive to ionizing radiation (IR).Vomiting, diarrhea, abdominal pain, bleeding, obstruction, perforation, and nutrient absorption disorder are common radiation toxicities in the intestine, which may lead to the decline of the patients’ quality of life and even death (1). Several therapy-related factors such as single-fraction dose, total dose, irradiated volume of the intestine, and the use of concurrent chemotherapy or biotherapy will influence the incidence and severity of intestinal radiation toxicity (2, 3). Moreover, patient-related factors like previous abdominal surgery, inflammatory bowel disease, diabetes, and vascular disorders may also affect the occurrence of Re (3). To solve the complications caused by radiotherapy, many strategies have been developed to relieve symptoms, including limiting intestinal irradiation dose and using a lower fractionated dose. However, these compromises may reduce the anti-tumor effect (3, 4). An in-depth study of the mechanism of Re is very important for finding new and effective strategies to prevent and treat Re. This review mainly summarized the current research mechanism and intervention measures related to intestinal injury caused by radiation therapy.
Structure and function of the intestinal barrier
The function of the intestinal barrier is for the absorption of nutrients through the mucosa and the prevention of injury from other toxic substances (5). The mucous layer is the first physical line of defense of the intestinal barrier, preventing bacteria and viruses from directly contacting epithelial cells (6). The main component of the mucous layer is highly glycosylated mucus, forming a gelatinous sieve structure outside the intestinal epithelium. Mucin 2 (MUC2) secreted by goblet cells is the most abundant mucine in the mucous layer, and the expression of MUC2 is closely related to the occurrence of enteritis, but the specific mechanism is still unclear (7, 8). The tight junctions (TJs) between the intestinal epithelial cells (IECs) constitute the second barrier of the intestinal barrier (9, 10), which are the determinant of the intestinal barrier function (5). The TJs’ structure is composed of transmembrane proteins such as claudin, occludin, tricellulin, and junction adhesion molecules (11, 12). TJs connect IECs to form a continuous polarized single-layer structure, separating the lumen from the lamina propria (13, 14). The lamina propria forms the last layer of the intestinal barrier, which is composed of immune cells, endothelial cells (ECs), myofibroblasts, matrix components, etc. In addition, the intestinal microbiota is also involved in the formation of the intestinal barrier (15). It should be noted that the intestinal barrier is not a static structure; it is always in dynamic equilibrium (Figure 1).
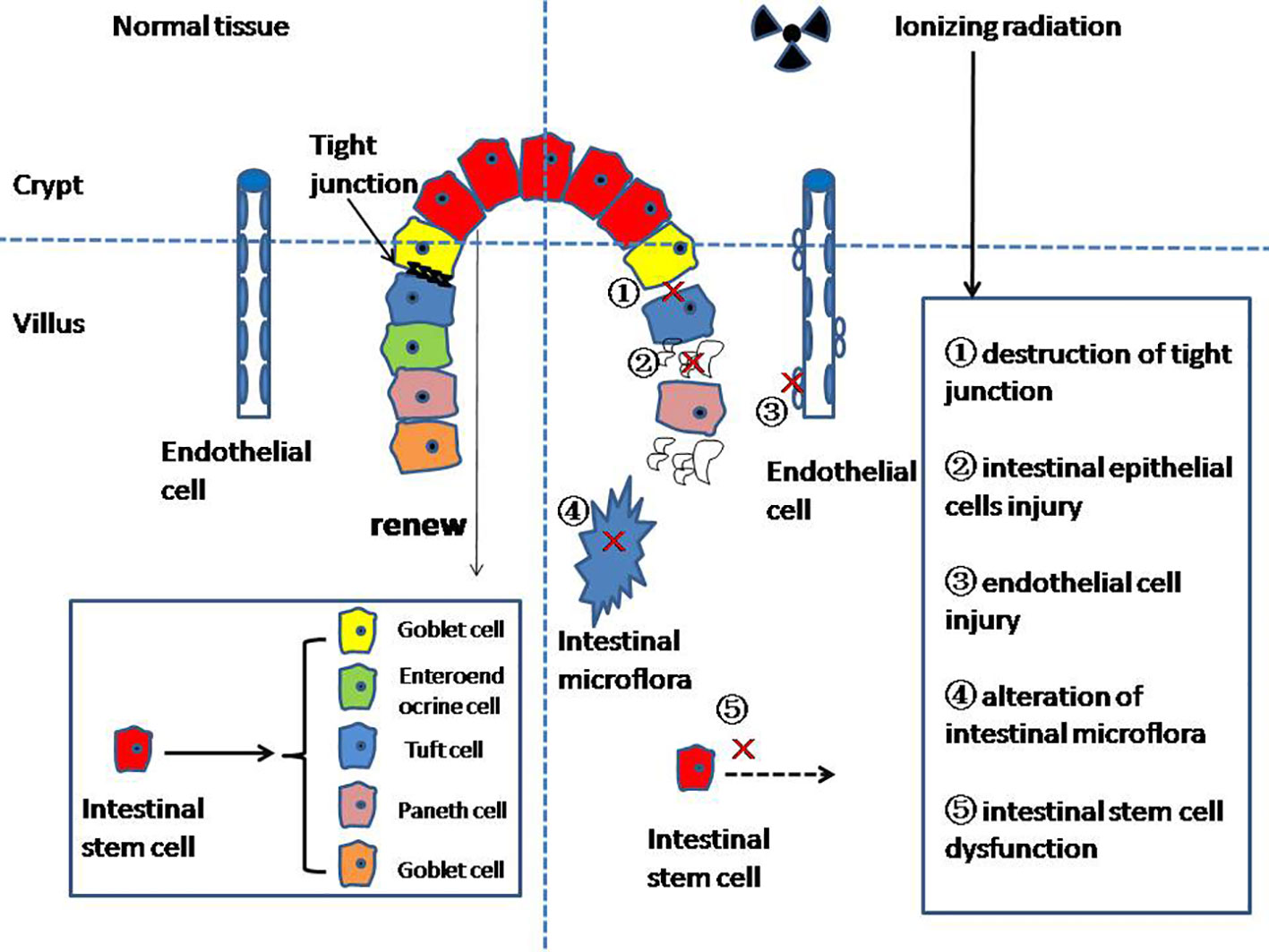
Figure 1 The mechanism of radiation enteritis.
At present, studies on the mechanism of Re focus on the following aspects: the destruction of the intestinal epithelium, intestinal stem cell (ISC) injury, intestinal microvascular changes, and intestinal microflora disruption, among others.
Intestinal epithelium injury
Radiotherapy can lead to an increase in intestinal epithelium barrier and permeability, which is closely related to the destruction of the TJs’ structure by IR. Morini et al. found that the expression of occludin, claudin, ZO-1, and ZO-2 was related to Re. They believed that when Re occurred, ZO-1, claudin-1, and occludin in TJs would recombine or break, resulting in the destruction of the intestinal barrier (16).
The adherent junctions (AJs) located below TJs are multiprotein complexes that indirectly regulate the TJs’ maturation and integrity. Gupta et al. (17) found that the AJs’ structure between IECs of mice disintegrated, expanded, and ruptured after exposure to IR. The amino acid-based oral rehydration solution (AA-ORS), including threonine, valine, serine, tyrosine, and aspartic acid, reduced dilation within AJs and reversed radiation-induced functional and structural disruption of the intestinal barrier (17).
Autophagy plays an important role in maintaining intestinal barrier homeostasis and regulates the apoptosis and necrosis of IECs (18). It has been proven that intracellular mitochondria produce a large number of intracellular reactive oxygen species (ROS), and when autophagy is insufficient, ROS levels will increase (19). ROS can cause structural damage and dysfunction in DNA. Ionizing radiation, on the other hand, inhibits autophagy, leading to damage to the intestinal barrier (20).
Resveratrol is a polyphenol synthesized in grape leaves and grape skins (21). Qin et al. (22) reported that the apoptosis level of IECs in the resveratrol-pretreated group was significantly lower than that in the irradiation group alone 24 h after exposure to ionizing radiation. Resveratrol can promote autophagy by activating the SIRT1 pathway to protect IECs and prevent the occurrence of Re (22).
Intestinal stem cell injury
IECs can renew themselves rapidly every 4–5 days (23). The ability to renew itself in the long-term can only be maintained by intestinal stem cells (ISCs) in the crypt (24). ISCs mainly form two differentiated epithelial lineages: (1) the enterocyte lineage, and (2) the secretory lineage. The enterocyte lineage is mainly responsible for absorbing nutrients. The secretory lineage consists of Paneth cells, which regulate the maintenance and differentiation of Lgr5+ ISCs; the mucus-secreting goblet cells; enteroendocrine cell (EEC); and tuft cells (Figure 1).
The main cause of Re is ISC death, which leads to the loss of key cells and destruction of crypt structures (25, 26).
Thiazolidine hydrochloride (TCZC01) is a novel compound synthesized by Zingerone. This paper concludes that pretreatment with TZC01 can significantly improve intestinal crypt apoptosis, increase the number of Lgr5+ ISCs, and even reduce intestinal cell apoptosis, thus protecting the intestinal barrier from IR damage. However, the mechanism of TZC01 is not explained in this paper (27).
The ISCs’ niche is not a constant, but a complex and dynamic environment. The stem cell zone is surrounded by enteric neurons, endothelial cells (ECs), smooth muscle cells (SMCs), intraepithelial lymphocytes, macrophages, and fibroblasts/myofibroblasts together with the extracellular matrix (ECM). Wnt, Notch, bone morphogenetic protein (BMP), and Hedgehog are the major signaling pathways involved in the maintenance of the ISCs’ niche (28).
Wnt/β-catenin pathway
The Wnt/β-catenin pathway plays an important regulatory role in intestinal tissue homeostasis (29). The stability of the environment within the intestinal tissue is coordinated and controlled by the self-renewal, regeneration, and reprogramming of stem cells (30). The typical Wnt signaling pathway is mediated by β-catenin, and β-catenin increases rapidly when Wnt ligands bind. After β-catenin enters the nucleus, it acts as a transcription co-activator of transcription factor 4 (TCF4), leading to the transcription and expression of a series of genes (31). A previous study showed that the Wnt/β-catenin signaling pathway played an important role in the self-renewal and proliferation of ISCs after radiation injury (32). However, we still do not know how the Wnt signal is transmitted to the target cell as the intercellular signal.
As reported by Li et al., epicatechin can salvage the ISCs’ activity and activate the Wnt/β-catenin pathway to induce crypt regeneration (33).
Podophyllotoxin combined with rutin (G-003M) is considered to reduce intestinal damage caused by radiation, and the main mechanism may enhance β-catenin nuclear translocation-promoted Lgr5 (+) ISC renewal through the Wnt/β-catenin signaling pathway (34).
Pretreatment with heat-killed Salmonella typhimurium (HKST) upregulated the nuclear localization of ß-catenin through the Wnt/b-catenin pathway. Moreover, pretreatment with HKST greatly increased the value of intestinal cells, significantly improved the structure and function of crypts, and reduced intestinal damage caused by ionizing radiation to prevent radiation enteritis (35).
Notch pathway
The Notch pathway is one of the key signaling pathways that maintain the balance of intestinal epithelial cell proliferation and differentiation. The Notch signaling pathway relies on cell-to-cell signaling, in which a cell provides a Notch ligand to adjacent cells expressing the Notch receptor (36). However, it usually results in the opposite fate of neighboring cells (lateral inhibition) (37). When the Notch receptor and ligand binding are activated, the Notch intracellular domain (NCID) is hydrolyzed and released into the nucleus and changes the gene expression in coordination with transcription factors, especially recombination signal binding protein J (RBP-J) (19).
Ghrelin is a hormone mainly produced by gastrointestinal endocrine cells (38). Recently, Kwak et al. (39) found that ghrelin could retain the proliferative function of IECs after irradiation by activating the Notch pathway in vitro. Further in vivo experiments confirmed that ghrelin could alleviate acute intestinal injury caused by radiation. The authors speculated that ghrelin is a potential strategy for the treatment of Re by activating the Notch pathway to retain the proliferative ability of IECs and repair intestinal barrier injury.
Park et al. (25) found that Valproic acid (VPA) was an effective intestinal protective agent. The results showed that IR reduced the activity of intestinal organoid by 70%, while pretreatment with VPA only reduced the activity of intestinal organoid by 30%. Further studies demonstrated that VPA significantly upregulated NOTCH1 mRNA level, activated the Notch pathway to reduce IR damage to LGR5+ cells, and improved crypt regeneration.
Bone morphogenetic protein signaling pathway
The BMP is the most important part of the transforming growth factor β (TGFβ) superfamily (40). BMP binds to the complex on the cell membrane (composed of serine/threonine kinase) and activates the intracellular heteromeric Smad complex, thereby regulating gene expression (41–43). On the other hand, the BMP signaling pathway has been shown to negatively regulate the self-renewal of Lgr5+ ISCs by inhibiting Wnt signaling (42, 44, 45). The BMP signaling pathway is negatively regulated by Gremlin, which is secreted by trophoblast cells (46, 47).
Martín-Alonso et al. (48)found that radiation-induced intestinal epithelial injury could not be repaired without the membrane-bound matrix metalloproteinase-17 (MMP17) expressed by smooth muscle cells. MMP17, an antagonist of the BMP signaling pathway, promotes the proliferation of ISCs.
Hedgehog signaling pathway
The Hedgehog (Hh) signaling pathway is a paracrine in the strict sense. In humans, three Hh ligands are expressed: Sonic Hedgehog (Shh), Indian Hedgehog (Ihh), and Desert Hedgehog (Dhh). Ihh is one of the important ligands of the Hedgehog pathway and maintains the stability of the ISCs’ niche (49). Ihh is secreted by IECs and acts on mesenchymal cells. Mesenchymal cells produce signaling factors that negatively regulate the proliferation of ISCs. Moreover, the BMP pathway plays a synergistic role in this process (50).
Intestinal vascular endothelial cell injury
Endothelial cell damage mediated by IR is also a pathophysiology of Re. Radiation intestinal epithelial injury is the main cause of acute radiation enteropathy, while chronic radiation enteropathy is caused by vascular endothelial injury. Vascular endothelial cells are sensitive to IR (51). Irradiation of the vascular system can rupture blood vessels and induce a pro-inflammatory response (52). After endothelial injury, subendothelial extracellular matrix (ECM) components are exposed to platelets, which initiate the hemostatic mechanism by forming the thrombus (53). Due to the excessive secretion of the von Willebrand factor (vWF) by damaged endothelial cells, the coagulation cascade is over-activated, resulting in vascular occlusion (54). This can then lead to hyperemia or hemorrhage at the site of injury due to increased vascular permeability. This is why the irradiated gut has a poor blood supply (55). The progression of Re can be improved by reducing IR damage to the vascular endothelium (56).
Shao et al. (57) reported that ferulic acid (FA) can reduce the oxidative damage of radiation to endothelial cells. The thrombomodulin (THBD) pathway may be an important mechanism of FA against radiation injury. Endothelial acid sphingomyelinase can catalyze ceramide production, which leads to endothelial cell apoptosis. Rotolo et al. (58) reported that IR can lead to the activation of the endothelial acid sphingomyelinase, thus initiating cell apoptosis. The 2A2 is an anti-ceramide IgM, which can prevent endothelial cell apoptosis in the lamina propria of the small intestine. In addition, the 2A2 can promote the recovery of ISCs.
Endothelial thrombomodulin (TM) is a multi-domain transmembrane receptor protein with anti-inflammatory, cytoprotective, antifibrinolytic, antioxidant, and anticoagulant functions (59). The high expression of TGF-β means endothelial damage, permeability destruction, and endothelial dysfunction (60, 61). TM suppresses the TGF-β signaling pathway by inhibiting extracellular signal-regulated kinase (ERK) activation (62). Pathak et al. (63) proved that TM treatment significantly ameliorated Re.
Yan et al. (64) reported that ionizing radiation can lead to reduced intestinal blood supply, resulting in intestinal ischemia and induced Re. BH4 can improve intestinal blood perfusion, which is mainly achieved through the Gh1/BH4/eNOS pathway. After intestinal exposure to ionizing radiation, guanosine triphosphate (GTP) cyclic hydrolase 1 (Gch1) will decrease, and BH4 is regulated by Gch1 and will decrease with the decrease of Gch1. Endothelial nitric oxide synthase (eNOS) must be completely saturated with BH4 to synthesize nitric oxide (NO). NO can relax vascular smooth muscle and maintain blood perfusion. Exogenous BH4 supplementation significantly improved the function of intestinal endothelial cells and intestinal blood perfusion, and alleviated pathological injury, thus preventing radiation enteritis. They further studied that ligustilide (LIG) can also prevent Re through the Gch1/BH4/eNOS pathway (65). The mechanism of action of LIG is to ameliorate the decrease of Gch1 protein level, thereby increasing BH4 and NO content. Compared with the control group, the LIG group significantly increased the length of intestinal villi. In addition, pretreatment with LIG improved weight loss and diarrhea caused by radiation. These results can reflect that LIG pretreatment has a positive effect on the prevention of Re.
Intestinal microflora
The human intestinal microflora contains 1014 species of resident microorganisms that live in the human intestinal tract with bacteria, viruses, fungi, and protozoa (66). Some studies have shown that intestinal microflora helps break down other indigestible polysaccharides in our diet, regulates the storage of calories extracted from our diet in fat cells, metabolizes foreign substances including carcinogens, regulates intestinal epithelial cell turnover, and educates the immune system how to respond to external stimuli (67). Metabolites produced by microbial communities play an important role in maintaining homeostasis and internal environment stability (68). The destruction of intestinal microflora is closely related to Re (69).
Touchefeu et al. believed that there are significant changes in intestinal microflora in patients receiving radiotherapy, with the most common being a decrease in the Clostridium cluster XIVa, Bifidobacterium, Faecalibacterium prausnitzii, and an increase in Enterobacteriaceae. These modifications may lead to mucositis, bacteremia, and diarrhea (70). Johnson et al. (71) believed that pseudo-intestinal obstruction and bacterial overgrowth may occur after abdominal radiotherapy. Impaired motor function is one of the causes of gastrointestinal colonization of Gram-negative bacilli. The results of Crawford and Gordon’ showed that compared with ordinary mice, germ-free mice received a lethal dose of total body irradiation, the survival rate was significantly higher, and the survival time was significantly longer (67).
Urolithin A (UroA) is a metabolite of intestinal microflora. Zhang et al. (72) found that UroA at 2 mg/kg significantly improved the survival and regeneration of intestinal structure and intestinal epithelium in rats exposed to ionizing radiation. In addition, UroA can regulate the structure of intestinal microbiome. IR can further increase the abundance of Escherichia shigella, Proteobacteria, Alphaproteobacteria, and Erysipelotrichaceae. The proliferation of these microflora leads to the destruction of the structure and function of the intestinal barrier and promotes intestinal inflammation, but UroA can reverse this result and prevent the occurrence of Re.
Conclusion
Cancer therapy continues to improve, but radiation therapy remains an important part of cancer treatment, and Re is an inevitable side effect of radiotherapy. Interventions for Re often determine the efficacy of radiation therapy in patients. The occurrence of Re is usually not determined by unilateral factors, and the complex interaction between intestinal epithelium, ISCs, capillary endothelium, and luminal bacteria is considered to be the basis of Re pathogenesis. Resveratrol, TZC01, HKST, etc. could reduce Re in animal experiments, which needs to be confirmed by clinical trials.
Author contributions
JF and BL drafted the manuscript, and MF, TN, FG and BT participated in the data review and collection for the study. XD conceived the study and participated in its design and coordination. All authors read and approved the final manuscript.
Funding
This work was financially supported by the NHC Key Laboratory of Nuclear Technology Medical Transformation (Mianyang Central Hospital, grant no. 2022HYX008) and Natural Science Foundation of Sichuan Province (grant no. 2022NSFSC0849).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Bhutta BS, Fatima R, Aziz M. Radiation enteritis. In: StatPearls. Treasure Island (FL: StatPearls Publishing LLC (2022).
2. Kountouras J, Zavos C. Recent advances in the management of radiation colitis. World J Gastroenterol (2008) 14(48):7289–301. doi: 10.3748/wjg.14.7289
3. Hauer-Jensen M, Denham JW, Andreyev HJ. Radiation enteropathy–pathogenesis, treatment and prevention. Nat Rev Gastroenterol Hepatol (2014) 11(8):470–9. doi: 10.1038/nrgastro.2014.46
4. Li Y, Yan H, Zhang Y, Li Q, Yu L, Li Q, et al. Alterations of the gut microbiome composition and lipid metabolic profile in radiation enteritis. Front Cell Infect Microbiol (2020) 10:541178. doi: 10.3389/fcimb.2020.541178
5. Keita AV, Söderholm JD. The intestinal barrier and its regulation by neuroimmune factors. Neurogastroenterol Motil (2010) 22(7):718–33. doi: 10.1111/j.1365-2982.2010.01498.x
6. Turner JR. Intestinal mucosal barrier function in health and disease. Nat Rev Immunol (2009) 9(11):799–809. doi: 10.1038/nri2653
7. Pelaseyed T, Bergström JH, Gustafsson JK, Ermund A, Birchenough GM, Schütte A, et al. The mucus and mucins of the goblet cells and enterocytes provide the first defense line of the gastrointestinal tract and interact with the immune system. Immunol Rev (2014) 260(1):8–20. doi: 10.1111/imr.12182
8. Van der Sluis M, De Koning BA, De Bruijn AC, Velcich A, Meijerink JP, Van Goudoever JB, et al. Muc2-deficient mice spontaneously develop colitis, indicating that MUC2 is critical for colonic protection. Gastroenterology (2006) 131(1):117–29. doi: 10.1053/j.gastro.2006.04.020
9. Wang K, Ding Y, Xu C, Hao M, Li H, Ding L. Cldn-7 deficiency promotes experimental colitis and associated carcinogenesis by regulating intestinal epithelial integrity. Oncoimmunology (2021) 10(1):1923910. doi: 10.1080/2162402X.2021.1923910
10. Qu W, Zhang L, Ao J. Radiotherapy induces intestinal barrier dysfunction by inhibiting autophagy. ACS Omega (2020) 5(22):12955–63. doi: 10.1021/acsomega.0c00706
11. Suzuki T. Regulation of the intestinal barrier by nutrients: The role of tight junctions. Anim Sci J (2020) 91(1):e13357. doi: 10.1111/asj.13357
12. Du H, Yang X, Fan J, Du X. Claudin 6: Therapeutic prospects for tumours, and mechanisms of expression and regulation (Review). Mol Med Rep (2021) 24(3). doi: 10.3892/mmr.2021.12316
13. Vancamelbeke M, Vermeire S. The intestinal barrier: a fundamental role in health and disease. Expert Rev Gastroenterol Hepatol (2017) 11(9):821–34. doi: 10.1080/17474124.2017.1343143
14. Hou Q, Huang J, Ayansola H, Masatoshi H, Zhang B. Intestinal stem cells and immune cell relationships: Potential therapeutic targets for inflammatory bowel diseases. Front Immunol (2020) 11:623691. doi: 10.3389/fimmu.2020.623691
15. Troy EB, Kasper DL. Beneficial effects of bacteroides fragilis polysaccharides on the immune system. Front Biosci (Landmark Ed) (2010) 15(1):25–34. doi: 10.2741/3603
16. Morini J, Babini G, Barbieri S, Baiocco G, Ottolenghi A. The interplay between radioresistant caco-2 cells and the immune system increases epithelial layer permeability and alters signaling protein spectrum. Front Immunol (2017) 8:223. doi: 10.3389/fimmu.2017.00223
17. Gupta R, Yin L, Grosche A, Lin S, Xu X, Guo J, et al. An amino acid-based oral rehydration solution regulates radiation-induced intestinal barrier disruption in mice. J Nutr (2020) 150(5):1100–8. doi: 10.1093/jn/nxaa025
18. Foerster EG, Mukherjee T, Cabral-Fernandes L, Rocha JDB, Girardin SE, Philpott DJ. How autophagy controls the intestinal epithelial barrier. Autophagy (2022) 18(1):86–103. doi: 10.1080/15548627.2021.1909406
19. Asano J, Sato T, Ichinose S, Kajita M, Onai N, Shimizu S, et al. Intrinsic autophagy is required for the maintenance of intestinal stem cells and for irradiation-induced intestinal regeneration. Cell Rep (2017) 20(5):1050–60. doi: 10.1016/j.celrep.2017.07.019
20. Datta K, Suman S, Fornace AJ Jr. Radiation persistently promoted oxidative stress, activated mTOR via PI3K/Akt, and downregulated autophagy pathway in mouse intestine. Int J Biochem Cell Biol (2014) 57:167–76. doi: 10.1016/j.biocel.2014.10.022
21. Galiniak S, Aebisher D, Bartusik-Aebisher D. Health benefits of resveratrol administration. Acta Biochim Pol (2019) 66(1):13–21. doi: 10.18388/abp.2018_2749
22. Qin H, Zhang H, Zhang X, Zhang S, Zhu S, Wang H. Resveratrol protects intestinal epithelial cells against radiation-induced damage by promoting autophagy and inhibiting apoptosis through SIRT1 activation. J Radiat Res (2021) 62(4):574–81. doi: 10.1093/jrr/rrab035
23. Rees WD, Sly LM, Steiner TS. How do immune and mesenchymal cells influence the intestinal epithelial cell compartment in inflammatory bowel disease? let’s crosstalk about it! J Leukoc Biol (2020) 108(1):309–21. doi: 10.1002/JLB.3MIR0120-567R
24. Beumer J, Clevers H. Regulation and plasticity of intestinal stem cells during homeostasis and regeneration. Development (2016) 143(20):3639–49. doi: 10.1242/dev.133132
25. Park M, Kwon J, Youk H, Shin US, Han YH, Kim Y. Valproic acid protects intestinal organoids against radiation via NOTCH signaling. Cell Biol Int (2021) 45(7):1523–32. doi: 10.1002/cbin.11591
26. Saha S, Aranda E, Hayakawa Y, Bhanja P, Atay S, Brodin NP, et al. Macrophage-derived extracellular vesicle-packaged WNTs rescue intestinal stem cells and enhance survival after radiation injury. Nat Commun (2016) 7:13096. doi: 10.1038/ncomms13096
27. Wu J, Duan Y, Cui J, Dong Y, Li H, Wang M, et al. Protective effects of zingerone derivate on ionizing radiation-induced intestinal injury. J Radiat Res (2019) 60(6):740–6. doi: 10.1093/jrr/rrz065
28. Moussa L, Usunier B, Demarquay C, Benderitter M, Tamarat R, Sémont A, et al. Bowel radiation injury: Complexity of the pathophysiology and promises of cell and tissue engineering. Cell Transplant (2016) 25(10):1723–46. doi: 10.3727/096368916X691664
29. Nusse R, Clevers H. Wnt/β-catenin signaling, disease, and emerging therapeutic modalities. Cell (2017) 169(6):985–99. doi: 10.1016/j.cell.2017.05.016
30. Huang P, Yan R, Zhang X, Wang L, Ke X, Qu Y. Activating wnt/β-catenin signaling pathway for disease therapy: Challenges and opportunities. Pharmacol Ther (2019) 196:79–90. doi: 10.1016/j.pharmthera.2018.11.008
31. Young MA, Daly CS, Taylor E, James R, Clarke AR, Reed KR. Subtle deregulation of the wnt-signaling pathway through loss of Apc2 reduces the fitness of intestinal stem cells. Stem Cells (2018) 36(1):114–22. doi: 10.1002/stem.2712
32. Bhanja P, Norris A, Gupta-Saraf P, Hoover A, Saha S. BCN057 induces intestinal stem cell repair and mitigates radiation-induced intestinal injury. Stem Cell Res Ther (2018) 9(1):26. doi: 10.1186/s13287-017-0763-3
33. Li Y, Ma S, Zhang Y, Yao M, Zhu X, Guan F. (-)-Epicatechin mitigates radiation-induced intestinal injury and promotes intestinal regeneration via suppressing oxidative stress. Free Radic Res (2019) 53(8):851–64. doi: 10.1080/10715762.2019.1635692
34. Kalita B, Ranjan R, Gupta ML. Combination treatment of podophyllotoxin and rutin promotes mouse Lgr5(+ ve) intestinal stem cells survival against lethal radiation injury through wnt signaling. Apoptosis (2019) 24(3-4):326–40. doi: 10.1007/s10495-019-01519-x
35. Chen Y, Cao K, Liu H, Liu T, Liu L, Qin H, et al. Heat killed salmonella typhimurium protects intestine against radiation injury through wnt signaling pathway. J Oncol (2021) 2021:5550956. doi: 10.1155/2021/5550956
36. Hori K, Sen A, Artavanis-Tsakonas S. Notch signaling at a glance. J Cell Sci (2013) 126(Pt 10):2135–40. doi: 10.1242/jcs.127308
37. Sancho R, Cremona CA, Behrens A. Stem cell and progenitor fate in the mammalian intestine: Notch and lateral inhibition in homeostasis and disease. EMBO Rep (2015) 16(5):571–81. doi: 10.15252/embr.201540188
38. Lv Y, Liang T, Wang G, Li Z. Ghrelin, a gastrointestinal hormone, regulates energy balance and lipid metabolism. Biosci Rep (2018) 38(5). doi: 10.1042/BSR20181061
39. Kwak SY, Shim S, Park S, Kim H, Lee SJ, Kim MJ. Ghrelin reverts intestinal stem cell loss associated with radiation-induced enteropathy by activating notch signaling. Phytomedicine (2021) 81:153424. doi: 10.1016/j.phymed.2020.153424
40. Auclair BA, Benoit YD, Rivard N, Mishina Y, Perreault N. Bone morphogenetic protein signaling is essential for terminal differentiation of the intestinal secretory cell lineage. Gastroenterology (2007) 133(3):887–96. doi: 10.1053/j.gastro.2007.06.066
41. Hardwick JC, Van Den Brink GR, Bleuming SA, Ballester I, Van Den Brande JM, Keller JJ, et al. Bone morphogenetic protein 2 is expressed by, and acts upon, mature epithelial cells in the colon. Gastroenterology (2004) 126(1):111–21. doi: 10.1053/j.gastro.2003.10.067
42. Wang S, Chen YG. BMP signaling in homeostasis, transformation and inflammatory response of intestinal epithelium. Sci China Life Sci (2018) 61(7):800–7. doi: 10.1007/s11427-018-9310-7
43. Feng XH, Derynck R. Specificity and versatility in tgf-beta signaling through smads. Annu Rev Cell Dev Biol (2005) 21:659–93. doi: 10.1146/annurev.cellbio.21.022404.142018
44. He XC, Zhang J, Tong WG, Tawfik O, Ross J, Scoville DH, et al. BMP signaling inhibits intestinal stem cell self-renewal through suppression of wnt-beta-catenin signaling. Nat Genet (2004) 36(10):1117–21. doi: 10.1038/ng1430
45. Tian Y, Ma X, Lv C, Sheng X, Li X, Zhao R, et al. Stress responsive miR-31 is a major modulator of mouse intestinal stem cells during regeneration and tumorigenesis. Elife (2017) 6. doi: 10.7554/eLife.29538
46. Biswas S, Davis H, Irshad S, Sandberg T, Worthley D, Leedham S. Microenvironmental control of stem cell fate in intestinal homeostasis and disease. J Pathol (2015) 237(2):135–45. doi: 10.1002/path.4563
47. McCarthy N, Manieri E, Storm EE, Saadatpour A, Luoma AM, Kapoor VN, et al. Distinct mesenchymal cell populations generate the essential intestinal BMP signaling gradient. Cell Stem Cell (2020) 26(3):391–402.e5. doi: 10.1016/j.stem.2020.01.008
48. Martín-Alonso M, Iqbal S, Vornewald PM, Lindholm HT, Damen MJ, Martínez F, et al. Smooth muscle-specific MMP17 (MT4-MMP) regulates the intestinal stem cell niche and regeneration after damage. Nat Commun (2021) 12(1):6741. doi: 10.1038/s41467-021-26904-6
49. Walton KD, Gumucio DL. Hedgehog signaling in intestinal development and homeostasis. Annu Rev Physiol (2021) 83:359–80. doi: 10.1146/annurev-physiol-031620-094324
50. Büller NV, Rosekrans SL, Westerlund J, van den Brink GR. Hedgehog signaling and maintenance of homeostasis in the intestinal epithelium. Physiol (Bethesda) (2012) 27(3):148–55. doi: 10.1152/physiol.00003.2012
51. Stewart FA, Akleyev AV, Hauer-Jensen M, Hendry JH, Kleiman NJ, Macvittie TJ, et al. ICRP publication 118: ICRP statement on tissue reactions and early and late effects of radiation in normal tissues and organs–threshold doses for tissue reactions in a radiation protection context. Ann ICRP (2012) 41(1-2):1–322. doi: 10.1016/j.icrp.2012.02.001
52. Ebrahimian T, Le Gallic C, Stefani J, Dublineau I, Yentrapalli R, Harms-Ringdahl M, et al. Chronic gamma-irradiation induces a dose-Rate-Dependent pro-inflammatory response and associated loss of function in human umbilical vein endothelial cells. Radiat Res (2015) 183(4):447–54. doi: 10.1667/RR13732.1
53. Wynn TA. Integrating mechanisms of pulmonary fibrosis. J Exp Med (2011) 208(7):1339–50. doi: 10.1084/jem.20110551
54. Giblin JP, Hewlett LJ, Hannah MJ. Basal secretion of von willebrand factor from human endothelial cells. Blood (2008) 112(4):957–64. doi: 10.1182/blood-2007-12-130740
55. Chang PY, Qu YQ, Wang J, Dong LH. The potential of mesenchymal stem cells in the management of radiation enteropathy. Cell Death Dis (2015) 6(8):e1840. doi: 10.1038/cddis.2015.189
56. Wang Y, Boerma M, Zhou D. Ionizing radiation-induced endothelial cell senescence and cardiovascular diseases. Radiat Res (2016) 186(2):153–61. doi: 10.1667/RR14445.1
57. Shao S, Gao Y, Liu J, Tian M, Gou Q, Su X, et al. Ferulic acid mitigates radiation injury in human umbilical vein endothelial cells In vitro via the thrombomodulin pathway. Radiat Res (2018) 190(3):298–308. doi: 10.1667/RR14696.1
58. Rotolo J, Stancevic B, Zhang J, Hua G, Fuller J, Yin X, et al. Anti-ceramide antibody prevents the radiation gastrointestinal syndrome in mice. J Clin Invest (2012) 122(5):1786–90. doi: 10.1172/JCI59920
59. Ito T, Maruyama I. Thrombomodulin: protectorate God of the vasculature in thrombosis and inflammation. J Thromb Haemost (2011) 9(Suppl)1:168–73. doi: 10.1111/j.1538-7836.2011.04319.x
60. Kruse JJ, Floot BG, te Poele JA, Russell NS, Stewart FA. Radiation-induced activation of TGF-beta signaling pathways in relation to vascular damage in mouse kidneys. Radiat Res (2009) 171(2):188–97. doi: 10.1667/RR1526.1
61. Feng W, Ying WZ, Aaron KJ, Sanders PW. Transforming growth factor-β mediates endothelial dysfunction in rats during high salt intake. Am J Physiol Renal Physiol (2015) 309(12):F1018–25. doi: 10.1152/ajprenal.00328.2015
62. Shang L, Jia SS, Jiang HM, Wang H, Xu WH, Lv CJ. Simvastatin downregulates expression of TGF-βRII and inhibits proliferation of A549 cells via ERK. Tumour Biol (2015) 36(6):4819–24. doi: 10.1007/s13277-015-3134-7
63. Pathak R, Wang J, Garg S, Aykin-Burns N, Petersen KU, Hauer-Jensen M. Recombinant thrombomodulin (Solulin) ameliorates early intestinal radiation toxicity in a preclinical rat model. Radiat Res (2016) 186(2):112–20. doi: 10.1667/RR14408.1
64. Yan T, Zhang T, Mu W, Qi Y, Guo S, Hu N, et al. Ionizing radiation induces BH(4) deficiency by downregulating GTP-cyclohydrolase 1, a novel target for preventing and treating radiation enteritis. Biochem Pharmacol (2020) 180:114102. doi: 10.1016/j.bcp.2020.114102
65. Yan T, Guo S, Zhang T, Zhang Z, Liu A, Zhang S, et al. Ligustilide prevents radiation enteritis by targeting Gch1/BH(4)/eNOS to improve intestinal ischemia. Front Pharmacol (2021) 12:629125. doi: 10.3389/fphar.2021.629125
66. Singh RK, Chang HW, Yan D, Lee KM, Ucmak D, Wong K, et al. Influence of diet on the gut microbiome and implications for human health. J Transl Med (2017) 15(1):73. doi: 10.1186/s12967-017-1175-y
67. Crawford PA, Gordon JI. Microbial regulation of intestinal radiosensitivity. Proc Natl Acad Sci U.S.A. (2005) 102(37):13254–9. doi: 10.1073/pnas.0504830102
68. Li Y, Zhang Y, Wei K, He J, Ding N, Hua J, et al. Review: Effect of gut microbiota and its metabolite SCFAs on radiation-induced intestinal injury. Front Cell Infect Microbiol (2021) 11:577236. doi: 10.3389/fcimb.2021.577236
69. Visich KL, Yeo TP. The prophylactic use of probiotics in the prevention of radiation therapy-induced diarrhea. Clin J Oncol Nurs (2010) 14(4):467–73. doi: 10.1188/10.CJON.467-473
70. Touchefeu Y, Montassier E, Nieman K, Gastinne T, Potel G, Bruley des Varannes S, et al. Systematic review: the role of the gut microbiota in chemotherapy- or radiation-induced gastrointestinal mucositis - current evidence and potential clinical applications. Aliment Pharmacol Ther (2014) 40(5):409–21. doi: 10.1111/apt.12878
71. Johnson LB, Riaz AA, Adawi D, Wittgren L, Bäck S, Thornberg C, et al. Radiation enteropathy and leucocyte-endothelial cell reactions in a refined small bowel model. BMC Surg (2004) 4:10. doi: 10.1186/1471-2482-4-10
Keywords: Radiation enteritis, mechanism, intestinal epithelial cells, intestinal stem cells, vascular endothelial cell, intestinal microflora
Citation: Fan J, Lin B, Fan M, Niu T, Gao F, Tan B and Du X (2022) Research progress on the mechanism of radiation enteritis. Front. Oncol. 12:888962. doi: 10.3389/fonc.2022.888962
Received: 03 March 2022; Accepted: 10 August 2022;
Published: 05 September 2022.
Edited by:
Shuyu Zhang, Sichuan University, ChinaReviewed by:
Guy Robert Orangio, Louisiana State University, United StatesArnaud Alves, Centre Hospitalier Universitaire de Caen, France
Copyright © 2022 Fan, Lin, Fan, Niu, Gao, Tan and Du. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaobo Du, ZHV4aWFvYm8yMDA1QDEyNi5jb20=
†These authors have contributed equally to this work