 Xianquan Zhan
Xianquan Zhan Jiajia Li2,4
Jiajia Li2,4- 1Shandong Key Laboratory of Radiation Oncology, Cancer Hospital of Shandong First Medical University, Jinan, China
- 2Science and Technology Innovation Center, Shandong First Medical University, Jinan, China
- 3Department of Oncology, Shandong Provincial Hospital Affiliated to Shandong First Medical University, Jinan, China
- 4Department of Thoracic Surgery, Xiangya Hospital, Central South University, Changsha, China
Oxidative stress and oxidative damage are the common pathophysiological characteristics in pituitary adenomas (PAs), which have been confirmed with many omics studies in PA tissues and cell/animal experimental studies. Nuclear factor erythroid 2 p45-related factor 2 (Nrf2), the core of oxidative stress response, is an oxidative stress sensor. Nrf2 is synthesized and regulated by multiple factors, including Keap1, ERK1/2, ERK5, JNK1/2, p38 MAPK, PKC, PI3K/AKT, and ER stress, in the cytoplasm. Under the oxidative stress status, Nrf2 quickly translocates from cytoplasm into the nucleus and binds to antioxidant response element /electrophile responsive element to initiate the expressions of antioxidant genes, phases I and II metabolizing enzymes, phase III detoxifying genes, chaperone/stress response genes, and ubiquitination/proteasomal degradation proteins. Many Nrf2 or Keap1 inhibitors have been reported as potential anticancer agents for different cancers. However, Nrf2 inhibitors have not been studied as potential anticancer agents for PAs. We recommend the emphasis on in-depth studies of Nrf2 signaling and potential therapeutic agents targeting Nrf2 signaling pathways as new therapeutic strategies for PAs. Also, the use of Nrf2 inhibitors targeting Nrf2 signaling in combination with ERK inhibitors plus p38 activators or JNK activators targeting MAPK signaling pathways, or drugs targeting mitochondrial dysfunction pathway might produce better anti-tumor effects on PAs. This perspective article reviews the advances in oxidative stress and Nrf2-mediated oxidative stress response signaling pathways in pituitary tumorigenesis, and the potential of targeting Nrf2 signaling pathways as a new therapeutic strategy for PAs.
Introduction
Pituitary adenoma (PA) is a common intracranial neoplasm that occurs in the central regulatory organ pituitary gland in the hypothalamic-pituitary-target organ axis system, which seriously affects human endocrine system and health. PAs account for 10–25% of all intracranial tumors, and are classified into benign (∼65%), invasive (∼35%), and malignant (carcinoma; only 0.1–0.2%) PAs according to the malignancy level (Stalla et al., 2019). PAs are divided into macroadenomas (≥10 mm) and microadenomas (<10 mm) according to tumor size (Lopes, 2017). They are also divided into clinically functional and nonfunctional PAs (FPAs and NFPAs) according to the level of hormone secretion (Zhan et al., 2016). FPAs are hormone-secreting PAs, which result in hyperpituitarism, including acromegaly derived from growth hormone (GH)-secreting PAs, hyperprolactinemia derived from prolactin (PRL)-secreting PAs, and Cushing’s syndrome derived from adrenocorticotropin (ACTH)-secreting PAs. NFPAs are non-hormone-secreting PAs (Qian et al., 2018). The main clinical symptoms of PAs include inappropriate hormone secretion syndrome, and compression of the neighboring tissues and structures such as headache, visual field defect, and increased intracranial pressure (Reimondo et al., 2019). PA is a multi-factor, multi-process, and multi-consequence complex disease, which is involved in a series of molecular alterations at the levels of genome, transcriptome, proteome, peptidome, metabolome, and radiome; and these molecules mutually associate and function in a molecular network system (Zhan and Desiderio, 2010b; Hu et al., 2013; Grech et al., 2015; Cheng and Zhan, 2017; Lu and Zhan, 2018). Thus, one must shift the research and practice strategy from a single-factor model to a multi-parameter systematic model for predictive, preventive, and personalized medicine in PAs (Hu et al., 2013; Grech et al., 2015; Cheng and Zhan, 2017). Multiomics is an effective approach to realize this multi-parameter systematic strategy model shift, which can establish signaling pathway systems for in-depth understanding of molecular mechanisms of PAs, identify molecular network-based biomarkers for prediction, diagnosis, and prognostic assessment of PAs, and discover signaling pathway network-based therapeutic targets for effective treatment of PAs (Grech et al., 2015; Cheng and Zhan, 2017; Lu and Zhan, 2018).
A series of omics analyses have been performed in PAs to reach our long-term goals that clarify molecular mechanisms and discover effective biomarkers and therapeutic targets for PAs (Zhan and Desiderio, 2010a; Long et al., 2019; Cheng et al., 2019; Wang Y. et al., 2019), including NFPA quantitative transcriptomics (differentially expressed genes, DEGs) (Moreno et al., 2005; Cheng et al., 2019), NFPA quantitative proteomics (differentially expressed proteins, DEPs) (Moreno et al., 2005), NFPA proteomic mapping (Zhan and Desiderio, 2003; Wang X. et al., 2015; Cheng et al., 2019), NFPA nitroproteomics (Zhan and Desiderio, 2006), invasive NFPA quantitative transcriptomics (Galland et al., 2010; Zhou et al., 2011; Wang Y. et al., 2019), invasive NFPA quantitative proteomics (Zhan et al., 2014b), control pituitary proteomic mapping (Beranova-Giorgianni et al., 2002; Giorgianni et al., 2003; Zhao et al., 2005), pituitary control nitroproteomics (Zhan and Desiderio, 2004; Zhan and Desiderio, 2007), control pituitary phosphoproteomics (Giorgianni et al., 2004; Beranova-Giorgianni et al., 2006), PRL-secreting adenoma proteomics and transcriptomics (Evans et al., 2008), and ACTH-secreting adenoma proteomics and metabolomics (Feng et al., 2018). Integrative analysis of these omics data has revealed some important signaling pathway network alterations in PA pathogenesis, including mitochondrial dysfunction, oxidative stress, cell cycle dysregulation, and mitogen-activated protein kinase (MAPK) signaling pathway alteration (Zhan and Desiderio, 2010a; Long et al., 2019). Mitochondrial dysfunction pathway network and mitochondrial dynamics (Li and Zhan, 2019), and MAPK signaling pathway-based drug therapeutic targets (Lu et al., 2019) have been discussed in detailed in PAs. It is well-known that mitochondria are the energy factories of the body, and mitochondrial metabolism is the source of reactive oxygen species (ROS). The imbalance between free radicals reactive oxygen/nitrogen species (ROS/RNS) and antioxidant system leads to oxidative stress, which plays an important role in diseases. Many studies focus on oxidative stress system as therapeutic strategy; for example, benfotiamine is an efficient antioxidant, which could prevent oxidative stress in the anterior tibialis muscle and heart of mice (Gonçalves et al., 2019). Another research shows that pancreatic oxidative damage in the diabetic state is caused by ROS, and scavenging the various ROS generated in the disease is one of effective ways to treat this disease (Afolabi et al., 2018). Studies have clearly demonstrated that mitochondrial dysfunction and oxidative stress pathway changes operate in PAs (Zhan and Desiderio, 2010a), and nuclear factor erythroid 2 p45-related factor 2 (Nrf2)-mediated oxidative stress response significantly impacts the pathogenesis of PAs and modulates the energy metabolism reprogramming for PAs (Sabatino et al., 2018). It is well-known that PAs can lead to abnormal hormone secretion, which might affect oxidative stress and Nrf2 signaling in PAs; for example, human growth hormone (hGH) can attenuate inflammation and oxidative stress attained by Cisplatin probably through inhibition of Nrf2/heme oxygenase 1 (HO-1) pathway (Mahran, 2020). More studies show that Nrf2 signaling and oxidative stress can be regulated by cortisol (Wu et al., 2019), thyroid hormone (Mishra et al., 2019), follicle-stimulating hormone (FSH) (Li et al., 2020), luteinizing hormone (LH) (Li et al., 2020), GH (Mahran, 2020), ACTH (Benlloch et al., 2016), and PRL (Ebokaiwe et al., 2020). These findings clearly demonstrate the importance of oxidative stress in PAs. This present review article will focus on oxidative stress response signing pathway network in PA pathogenesis.
Redox Homeostasis and Nrf2 as the Heart of Oxidative Stress Response
Oxidative stress is derived from the imbalance between the upload of free radicals ROS/RNS from in vivo and in vitro environmental approaches and the ability of endogenous antioxidants to detoxify these ROS/RNS (Prasad et al., 2016; Klaunig, 2018; Sajadimajd and Khazaei, 2018). It results in the injuries of multiple biomacromolecules such as DNAs, RNAs, proteins, and membrane lipids to significantly associate with a wide spectrum of diseases including cancers. Many studies demonstrate that the increased ROS/RNS productions promote carcinogenesis development (Kudryavtseva et al., 2016; Kruk and Aboul-Enein, 2017), and oxidative stress-medicated chronic inflammation is the risk factor of tumorigenesis (Reuter et al., 2010; Qian et al., 2019). The oxidative phosphorylation system in mitochondrial respiratory chain is the central machine that generates ROS products such as superoxide radical (O2.-). One study shows that ROS levels and signs of oxidative damage are significantly increased in PAs (Sabatino et al., 2018). One of the most important RNS, nitric oxide (NO), is generated by inducible nitric synthase (iNOS) in many pathogenesis conditions, which can rapidly react with superoxide radical (O2.-) to generate more toxic peroxynitrite anion (ONOO-) and highly reactive hydroxyl radical (OH.) to attack DNAs, RNAs, proteins, and membrane lipids. iNOS has been extensively found in rat and human pituitaries (Ceccatelli et al., 1993; Lloyd et al., 1995; Ueta et al., 1998; Kruse et al., 2002; Pawlikowshi et al., 2003) and has the elevated activities in PAs compared to those in controls (Vankelecom et al., 1997; Kruse et al., 2002). Another study shows that NO functions in the hypothalamic-pituitary-adrenocortical axis (Riedel, 2002) by promoting the release of follicle-stimulating hormone-releasing hormone (FSHRH) and luteinizing hormone-releasing hormone (LHRH) from hypothalamus (McCann et al., 2001; Pinilla et al., 2001; McCann et al., 2003), and regulating secretion of PRL (Duvilanski et al., 1995) and GH in pituitaries and PAs (Cuttica et al., 1997; Pinilla et al., 1999; Bocca et al., 2000). Peroxynitrite anion (ONOO−) is a key factor in vivo that causes protein tyrosine nitration and alters protein functions. Nine nitrotyrosine-containing proteins have been identified in NFPA tissues, and tyrosine nitration occurs in important structural and functional domains to change protein functions (Zhan and Desiderio, 2006).
With the generation of ROS/RNS, the in vivo antioxidant detoxification system is correspondingly initiated to adapt against the increased ROS/RNS (Valko et al., 2006; Obrador et al., 2019). The endogenous antioxidant detoxification system is a very complex system, including i) enzymatic antioxidants such as superoxide dismutases (CuZnSOD and MnSOD), glutathione peroxidase, and catalase; ii) non-enzymatic antioxidants such as vitamin E, vitamin C, carotenoid, flavonoid, selenium, thiol antioxidant (thioredoxin, lipoic acid, and glutathione), and others; and iii) multiple regulatory factors [Nrf2, NF-kB (nuclear factor kB), and AP-1 (activator protein-1), etc.] that interact with antioxidants (Valko et al., 2006; Obrador et al., 2019). CuZnSOD exists in most parts of cells, while MnSOD is only found in mitochondrial matrix; and both of them are able to effectively scavenge O2.- and generate H2O2 (Li et al., 1995; Melov et al., 2001; Elchuri et al., 2005). H2O2 can be scavenged by GPX's (glutathione peroxidases) and peroxiredoxins (thioredox-independent peroxidases) (Chu et al., 2004; Kang et al., 2005). Studies have found that the levels of CuZnMOD and MnSOD are significantly lower in PAs compared to those of controls (Kurisaka et al., 2004; Yang et al., 2012; Ilhan et al., 2018). The abnormal activities of these antioxidant enzymes and non-enzymatic antioxidants are directly associated with carcinogenesis (Neumann et al., 2003; Chu et al., 2004; Harris et al., 2015). The transcription factor Nrf2 is pivotal to the antioxidant response, which is a sensor of oxidative stress in redox homeostasis, and is mainly located in the cytoplasm under basal conditions (Li and Kong, 2009; Furfaro et al., 2016a). When the upload of free radicals ROS/RNS is increased to cause oxidative stress, Nrf2 quickly translocates from cytoplasm into the nucleus to initiate the antioxidant response, protecting against oxidative/nitrative damages (Dhakshinamoorthy and Porter, 2004; Osburn et al., 2006; Mann et al., 2007; Pi et al., 2008). The Nrf2 signaling regulatory system contains at least four components, including Nrf2, Kelch-like ECH-associated protein 1 (Keap1), small musculoaponeurotic fibrosarcoma (Maf), and antioxidant response element (ARE) or electrophile responsive element (EpRE), which in combination are necessary for the antioxidant response (Kwak and Kensler, 2010; Furfaro et al., 2016; de la Vega et al., 2018). Nrf2 signaling pathways regulate multiple biological processes, including i) the expressions of antioxidant genes, ii) ubiquitin-proteasome system, iii) molecular chaperone/stress-response system, and iv) anti-inflammatory response (Kwak and Kensler, 2010; Furfaro et al., 2016). The accumulated evidence clearly demonstrates that Nrf2 signaling pathways are involved in 12 hallmarks of cancer, including sustained proliferative signaling, insensitivity to antigrowth signals, resistance to apoptosis, limitless replicative potential, sustained angiogenesis, tissue invasion and metastasis, metabolic reprogramming, avoiding immune destruction, tumor-promoting inflammation, genome instability, altered redox homeostasis, and proteotoxic stress (de la Vega et al., 2018). Thereby, any decreased capability of the antioxidant protective system in the redox homeostasis might cause more susceptibility to carcinogen toxicity, tumor inflammatory response, oxidative stress, and carcinogenesis (Yates and Kensler, 2007).
Multiuomics Reveals Oxidative Stress-Related Pathway Alterations in PAs
Our multiomics studies in PAs (Zhan and Desiderio, 2010a; Long et al., 2019) clearly demonstrate oxidative stress-related pathway changes in PAs. For example, i) Nrf2-mediated oxidative stress response pathway is significantly changed in NFPAs with evidence of upregulation of key molecules [upregulated DEPs: GST (glutathione S-transferase) or GSTM2 (glutathione S-transferase mu 2), and ERP29 (endoplasmic reticulum protein 29], and downregulation of key molecules [downregulated DEPs: HSP22 (heat shock protein 22), HSP27, and HSP90 or GRP94 (94 kD glucose-regulated protein)] in this pathway. ii) Mitochondrial dysfunction pathway is significantly changed in NFPAs with evidence of upregulation of key molecules [upregulated DEPs: NDUFS8 (NADH ubiquinone oxidoreductase core subunit S8), COX6B (cytochrome c oxidase subunit 6B), CAT (catalase), β-secret2, and ATP5B (ATP synthase, H+ transporting mitochondrial F1 complex, beta subunit)], and downregulation of key molecules [downregulated DEPs: GPX4 (glutathione peroxidase 4), and ATP5A1] in this pathway. Mitochondrial dysfunction can increase ROS production in cancer cells to mediate tumor-related signaling pathways and activate pro-oncogenic signaling (Li and Zhan, 2019). iii) Oxidative phosphorylation pathway is significantly changed in NFPAs with evidence of upregulation of key molecules (upregulated DEPs: NDUFS8, COX6B, and ATP5B) in this pathway. Mitochondrial oxidative phosphorylation system contains mitochondrial complexes I, II, III, IV, and V, which are the major sites that produce endogenous ROS such as OH. and O−2; among these, complexes I, II, and III play a crucial role in the generation of mitochondrial ROS, because the electrons tend to be leaky at complexes I and III, which results in an incomplete reduction of oxygen and thus generates a free radical such as superoxide radical (Li and Zhan, 2019). iv) Glutathione redox reaction I pathway is significantly changed in NFPAs with evidence of downregulation of key molecule (downregulated DEP: GPX4) in this pathway. GPX’s (glutathione peroxidases) are important components in the antioxidant defense system: the downregulation of GPX’s can decrease the capability of the antioxidant defense system. v) The superoxide radical degradation pathway is significantly changed in NFPAs with evidence of upregulation of key molecule (upregulated DEP: CAT) in this pathway. vi) Aryl hydrocarbon receptor signaling is significantly changed in NFPAs with evidence of upregulation of key molecules [upregulated DEP: GST; upregulated DEGs: HSPCA (heat shock protein 90 alpha family class A member 1), HSPCB (heat shock protein 90 alpha family class B member 1), ESR1 (estrogen receptor 1), and Bax (BCL2 associated X, apoptosis regulator)], and downregulation of key molecules [downregulated DEPs: HSP27, HSP90 or GRP94, and TGM2 (transglutaminase 2); downregulated DEG: ESR2 (estrogen receptor 2)] in this pathway. vii) Glucocorticoid receptor signaling is significantly changed in NFPAs with evidence of upregulation of key molecule [upregulated DEG: PI3K (phosphatidylinositol 3 kinase)], and downregulation of key molecules [downregulated DEGs: HSP70, c-Fos, CCL2 (C-C motif chemokine ligand 2), BCL2, PRL, and POMC (proopiomelanocortin)] in this pathway. viii) Corticotropin-releasing hormone signaling is significantly changed in NFPAs with evidence of upregulation of key molecules [upregulated DEGs: CALM (calmodulin), and IP3R)], and downregulation of key molecules [downregulated DEGs: ACTH, Nur77 (NR4A1 nuclear receptor subfamily 4 group A member 1), and c-FOS)] in this pathway. ix) Melatonin signaling is significantly changed in NFPAs with evidence of nitration of key molecule (PKA) in this pathway. x) Methylglyoxal degradation III pathway is significantly changed in NFPAs with evidence of upregulation of key molecules [upregulated DEPs: aldose reductase or AKR1B1(aldo-keto reductase family 1 member B)] in this pathway. xi) AMPK signaling is significantly changed in NFPAs with evidence of upregulation of key molecules [upregulated DEGs: PP2C (putative protein phosphatase), and PFK (phosphofructokinase)], and downregulation of key molecules [downregulated DEGs: PI3K, PKA, and PDK1 (pyruvate dehydrogenase kinase 1)] in this pathway. Thereby, these signaling pathway changes clearly demonstrate that the disturbance in redox homeostasis, the imbalance between generation and detoxification of free radicals ROS/RNS, results in oxidative stress and damage in human PAs. Recently, these findings are also confirmed with experiments in cell models and animal models, which demonstrate that increased mitochondrial fusion results in bigger mitochondria, increased ROS levels, and oxidative damage in PAs, and that Nrf2 signaling pathway is activated in PAs as an antioxidant response (Sabatino et al., 2018). Thus, it suggests that Nrf2 is the master regulator of the cellular antioxidant response (de la Vega et al., 2018).
Nrf2-Mediated Oxidative Stress Response Signaling Pathways in PAs
Nrf2 signaling pathway in response to oxidative stress is shown (Figure 1). Multiple in vivo and in vitro environmental factors, including inflammatory cytokines, prostaglandins, growth factors, low-density lipoproteins, bacterial and viral infection, heavy metals, ultraviolet (UV) radiation, ionizing radiation, drugs, xenobiotics, antioxidants, oxidants, and chemopreventive agents, cause the increased upload of free radicals ROS/RNS and electrophiles to result in oxidative stress (Hetland et al., 2020; Mehnati et al., 2020). The increased ROS or electrophiles will activate the Nrf2/Keap1 complex in the cytoplasm through ERK1/2, ERK5, JNK1/2, p38 MAPK, PKC, and PI3K-AKT signaling pathways, and these signaling pathways will communicate with each other (Roy Chowdhury et al., 2014; Tian et al., 2014; Wang K.-C. et al., 2019). The activated Nrf2 is phosphorylated and separated from Keap1 (Hambright et al., 2015; Sánchez-Martín et al., 2020). The separated and phosphorylated Nrf2 quickly translocates into the nucleus to interact with ARE or EpRE, which will initiate at least five types of gene expressions to exert the corresponding biological functions (Furfaro et al., 2016; Sánchez-Martín et al., 2020): i) reduction of the oxidative damage via antioxidant proteins such as NRF2, small MAF, ATF4, SQSTM1, HO-1, PRDX1, FTL, FTH1, CAT, GPX’s, SOD, TXN, GSR, and TRXR1 (Sun et al., 2019; Saad El-Din et al., 2020; Yu et al., 2020); ii) detoxification and metabolism of xenobiotics to regulate cell survival, or production of reactive metabolites to promote tumorigenesis via phase I and II metabolizing enzymes such as CYP1A/2A/3A/4A/2C, FMO, GST, NQD, UGT, AFAR, EPHX1, GCLC, GCLN, CBR4, AKR, and AOX4 (Zhao et al., 2015; Huang et al., 2018); iii) transportation of xenobiotics and metabolites via phase III detoxifying proteins such as SR-B1 and MRP1 (Sivils et al., 2013; Lubelska et al., 2016); iv) repairment and removal of the damaged proteins via chaperone and stress response proteins such as HSP22/40/90, STIP1, PTPLAD1, HERPUD1, CCT7, CLPP, FKBP5, PPIB, and ERP29 (Niture and Jaiswal, 2010; Sahin et al., 2012); and v) repairment and removal of the damaged proteins via ubiquitination and proteasomal degradation proteins such as PSM, UB2R1, VCP, USP14, UBB, and HIP2 (Liu et al., 2019; Song et al., 2019). This clearly demonstrates that while the Nrf2-mediated oxidative stress response signaling pathways are regulated by multiple factors, Nrf2 is the essential component. In the cytoplasm, Keap1, the main regulator of Nrf2, is a substrate adaptor protein for the Cul3-Keap1-E3 ligase complex that ubiquinates Nrf2, marking it for proteasomal degradation in the cytoplasm under basal conditions (Baird and Yamamoto, 2020; Dayalan Naidu and Dinkova-Kostova, 2020). To reduce its inhibitory effects on Nrf2, Keap1 can be ubiquitinated for degradation, leading to an increase in Nrf2 phosphorylation (activation) (Villeneuve et al., 2010). The phosphorylated Nrf2 can then interact with actin to form an Nrf2/actin complex that then translocates into the nucleus. After Nrf2 translocates into the nucleus, there are additional regulatory systems in place that include multiple factors such as ATF4, JUN, ERK1/2-CBP/P300, small MAF, BACH1, c-FOS, FRA1, and c-MAF, to influence the binding of Nrf2 and ARE/EpRE. The detailed regulatory mechanism system of Nrf2 has been extensively reviewed (Kwak and Kensler, 2010; Hybertson et al., 2011; Furfaro et al., 2016a; Lu et al., 2016; Menegon et al., 2016; Taguchi and Yamamoto, 2017; Bellezza et al., 2018; Chen and Maltagliati, 2018; de la Vega et al., 2018; Ryoo and Kwak, 2018; Sajadimajd and Khazaei, 2018; Cloer et al., 2019; Cuadrado et al., 2019; Qin et al., 2019) response signaling pathways have also been studied in pituitaries and PAs. One study shows that Nrf2, phosphorylated Nrf2 (p-Nrf2) protein, and mRNA expressions are increased in PAs, and the Nrf2 downstream effector HO-1 is also increased in PAs (Sabatino et al., 2018). This clearly demonstrates the activation of the Nrf2 signaling pathway, likely causing the extensive surviving capability of pituitary tumor cells. The Nrf2/PTEN-induced putative kinase protein 1 (PINK1)/Parkin pathway and mitophagy are activated in T-2 toxin-induced toxicities in rat pituitary GH3 cells (Deyu et al., 2018). Antioxidants N-acetylcysteine (NAC) and vitamin E can decrease the expressions of Nrf2 and HO-1 in rat pituitaries (Prevatto et al., 2017). Genetically induced Nrf2 overexpression in melanoma cells promotes tumor growth and increases antioxidant defense in malignant cells, which can be inhibited by anticancer agent pterostilbene (Pter, a natural dimethoxylated analog of resveratrol) through the downregulation of pituitary production of ACTH, plasma corticosterone, and the glucocorticoid receptor- and Nrf2-dependent antioxidant defense systems in growing melanomas (Benlloch et al., 2016). Irradiation can result in oxidative damage in C57/BL6 mice via activation of Nrf2 and HO-1 expressions, which can be blocked by antioxidant agent pituitary adenylate cyclase-activating polypeptide 38 (PACAP38) through inhibiting Nrf2 expression (Li et al., 2019). Chronic restraint stress (CRS) and acute restraint stress (ARS) can upregulate the mRNA expressions of oxidative stress molecules (gp91phox, iNOS, and Nrf2) and inflammation-related molecules (IL-1β, IL-6, TNFα, and TLR4) in the mouse hypothalamus, which can be alleviated by Iptakalim (Ipt), an ATP-sensitive potassium (K-ATP) channel opener (Zhao et al., 2017). The loss-of-function mutations of the aryl hydrocarbon receptor-interacting protein gene (AIP) are well-recognized in PAs (Hernández-Ramírez et al., 2018). The aryl hydrocarbon receptor signaling is also revealed by multiomics as an oxidative stress-related signaling pathway in PAs (Long et al., 2019). Further studies show that AIP interacts with antioxidants, chaperone and stress response-related proteins, and cytoskeletal proteins, including HSPA5, HSPA9, HSP90AA1, HSP90AB1, HSPA8, SOD1, TUBB, TUBB2A, and NME1; AIP variants show the impaired interaction of AIP with HSPA8, HSP90AB1, NME1, SOD1, TUBB, and TUBB2A; AIP-mutated PAs show the reduced expression of TUBB2A (Cuadrado et al., 2019). The levels of MnSOD and total antioxidant capability (TAC) are significantly decreased in GH-secreting PAs (Ilhan et al., 2018). The frequencies of micronuclei (MN), nucleoplasmic bridges, nuclear buds, apoptotic and necrotic cells, and plasma 8-hydroxy-2′-deoxyguanosine (8-OHdG) levels in peripheral blood lymphocytes are significantly increased in PRL-secreting PAs, which indicates the increased oxidative damage in PRL-secreting PAs (Bitgen et al., 2016). Oxidative stress and mitochondrial dysfunction have been revealed by multiple proteomics and nitroproteomics studies in human PAs (Zhan and Desiderio, 2010a; Zhan et al., 2013; Zhan et al., 2014a; Zhan et al., 2014b; Wang X. et al., 2015; Long et al., 2019). Also, tumor inflammation is an important pathophysiological characteristic in human PAs, which is always tightly associated with oxidative stress and chronic inflammation. The relationship among age-related disease, chronic inflammation, and oxidative stress has also been discussed (Pizza et al., 2011; Liguori et al., 2018; Qian et al., 2019). Oxidative stress is also involved in the processes of anti-proliferative effect and cell death induced by dopamine in the pituitary tumor cells via dopamine D2 receptors through p38 MAPK, and ERK pathways (An et al., 2003). Therefore, oxidative stress and antioxidative stress response extensively exist in PA pathogenesis. Nrf2, as the core of oxidative stress response, could be the novel target used to develop effective therapeutic agents for human PAs (Kwak and Kensler, 2010; Furfaro et al., 2016; de la Vega et al., 2018).
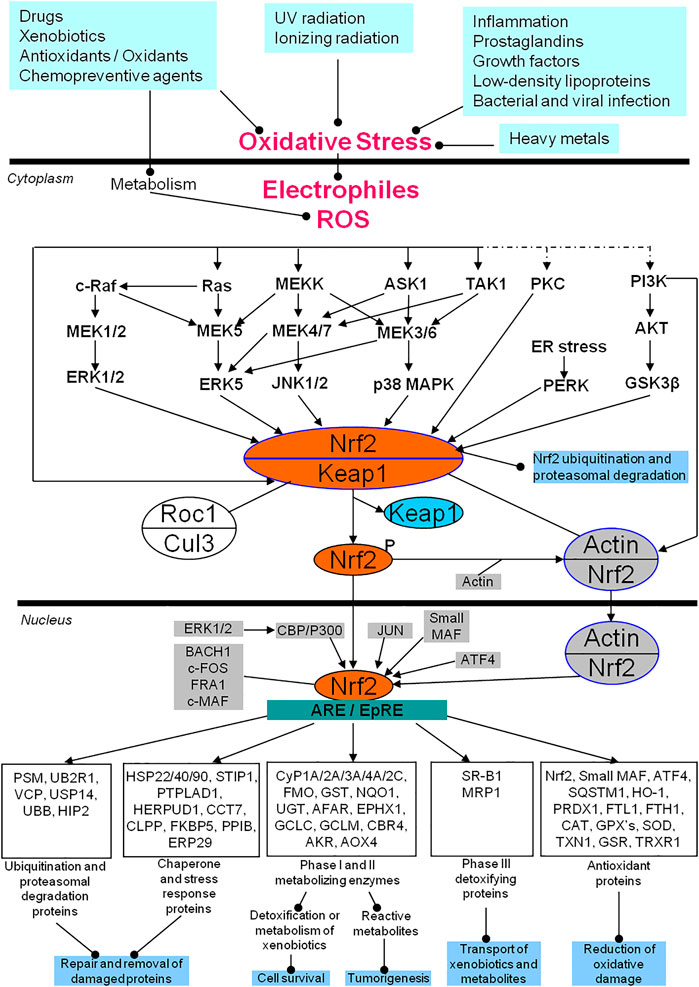
FIGURE 1. Nrf2-mediated oxidative stress response signaling pathways in human pituitary adenomas. AKR, Palmitoyltransferase; AKT, Protein kinase B; AOX4, Aldehyde oxidase 4; ARE, Antioxidant response element; ASK1, Apoptosis signal-regulating kinase 1; ATF4, Activating transcription factor 4; BACH1, Transcription regulator protein BACH1; CAT, catalase; CBP, CREB-binding protein; CBR4, carbonyl reductase 4; CCT7, T-complex protein 1 subunit eta; c-FOS, Proto-oncogene protein c-FOS; CLPP, Caseinolytic protease; Cul3, Cullin 3-based ubiquitin E3 ligase complex; Cyp, cytochrome P; EPHX1, Epoxide hydrolase 1; EpRE, Electrophile responsive element; ER, endoplasmic reticulum; ERK: Extracellular signal-related kinase; ERP29: endoplasmic reticulum protein 29; FKBP5, FK506-binding protein 5; FMO, Dimethylaniline monooxygenase [N-oxide-forming]; FRA1, Fos-related antigen 1; FTH1, Ferritin heavy polypeptide 1; FTL1, ferritin light polypeptide; GCLC, glutamate-cysteine ligase catalytic subunit; GCLM, glutamate-cysteine ligase modifier subunit; GPX's, Glutathione peroxidases; GSK3β, glycogen synthase kinase 3β; GSR, glutathione reductase; GST, glutathione S-transferase; HERPUD1, Homocysteine-responsive endoplasmic reticulum-resident ubiquitin-like domain member 1 protein; HIP2, Ubiquitin-conjugating enzyme E2 K; HO-1, heme oxygenase 1; HSP22/40/90, heat shock proteins 22, 40 and 90; JNK, Jun N-terminal kinase; Keap1, Kelch-like ECH-associated protein 1; Maf, Musculoaponeurotic fibrosarcoma; MAPK, Mitogen-activated protein kinase; MEK, Mitogen-activated protein kinase kinase (MAPKK); MEKK, Mitogen-activated protein kinase kinase kinase (MAPKKK); MRP1, multidrug-resistant protein-1; NQO1, NAD(P)H:quinine oxidoreductase 1; Nrf2, Nuclear factor erythroid 2 p45-related factor 2; PERK: the double-stranded RNA (PKR)-activated protein kinase-like eukaryotic initiation factor 2 kinase; PI3K, Phosphatidylinositol 3 kinase; PKC, protein kinase C; PPIB, Peptidyl-prolyl cis-trans isomerase B; PRDX1, peroxiredoxin 1; PSM: multiple subunits of the 20S proteasome; PTPLAD1, 3-hydroxyacyl-CoA dehydratase 3; c-Raf, RAF proto-oncogene serine/threonine-protein kinase; Ras, GTPase Ras; ROS, reactive oxygen species; SOD, Superoxide dismutase; SQSTM1, sequestosome-1 protein; SR-B1, Scavenger receptor class B member 1; STIP1, stress induced phosphoprotein 1; TAK1, TGF beta-Activated Kinase 1; TXN1: thioredoxin; TRXR1, thioredoxin reductase 1; UBB, Polyubiquitin-B; UB2R1, Ubiquitin-conjugating enzyme E2 R1; UGT, UDP glucuronosyl transferase; USP14, ubiquitin-specific peptidase 14; and VCP, valosin-containing protein. Modified from Zhan X et al. (2010) (Zhan and Desiderio, 2010a), copyright permission from BioMed Central publisher open-access article, copyright 2010; and modified from Long Y et al. (2019) (Long et al., 2019), copyright permission from Frontiersin publisher open-access article, copyright 2019.
Therapeutic Status Targeting Nrf2 Signaling Pathways in Cancers
Nrf2 signaling, as the heart of oxidative stress response, is extensively related to cancer pathogenesis, which has attracted tremendous attention as possible anticancer therapeutic target. Nrf2 signaling-based anticancer therapeutic studies have been extensively carried out in multiple cancers, including acute myeloid leukemia, gallbladder cancer, renal carcinoma, pancreatic cancer, melanoma, hepatocellular carcinoma, lung cancer, colon cancer, ovarian cancer, breast cancer, esophageal cancer, and glioblastoma (Table 1). i) In acute myeloid leukemia, studies found that Nrf2 activators [dimethyl fumarate (DMF), tert-butylhydroquinone, or carnosic acid] and vitamin D derivatives can cooperatively induce acute myeloid leukemia cell differentiation to inhibit leukemia progression in a xenograft mouse model via activating the Nrf2/ARE signaling pathway (Nachliely et al., 2019). Novel pyrazolyl hydroxamic acid derivative (4f) can inhibit Nrf2 activity to induce apoptosis of human acute myeloid leukemia cells (Zhang et al., 2017). ii) In gallbladder cancer, one study found that atypical protein kinase Cι (aPKCι) can promote gallbladder tumorigenesis and chemoresistance of anticancer agent gemcitabine by competing with Nrf2 for binding to Keap1, implying that inhibiting the aPKC1-Keap1-Nrf2 axis might overcome drug resistance for the treatment of gallbladder cancer (Tian et al., 2019). iii) In renal carcinoma, one study found that the natural product chitosan oligosaccharide (COS) can inhibit human renal carcinoma cell proliferation in vitro and in vivo by promoting the expressions of Nrf2 and Nrf2 target genes such as HO-1, the modifier subunit of glutamate cysteine ligase, solute carrier family 7 member 11, glucose-regulated protein 78, protein RNA-like endoplasmic reticulum kinase, and cytochrome C. (Zhai et al., 2019). iv) In pancreatic cancer, one study found that anticancer agent resveratrol enhances the sensitivity of pancreatic cancer cells to gemcitabine via suppressing NAF-1 (nutrient-deprivation autophagy factor-1) expression, inducing ROS accumulation, and activating Nrf2 signaling pathways (Cheng et al., 2018). v) In melanoma, the co-treatment of Nrf2 inhibitor (brusatol, BR) and UVA irradiation can effectively inhibit melanoma growth by regulating AKT-Nrf2 pathway (Wang et al., 2018). vi) In hepatocellular carcinoma, one study found that potential Nrf2 inhibitors can sensitize chemotherapy drugs in hepatocellular carcinoma (Tian et al., 2018). Cordycepin (CA) can activate the Nrf2/HO-1/NF-κB pathway for its anti-hepatocarcinoma effect in N-nitrosodiethylamine (NDEA)-induced mouse hepatocellular carcinomas (Zeng et al., 2017). The novel indazolo[3,2-b]quinazolinone (IQ) derivatives, IQ-7 and IQ-12, can induce apoptosis of human hepatoma cells Hep3B and inhibit the Nrf2/ARE signaling pathway in Hep3B cells, and IQ-7 is suggested as a degree of specificity against cancer cells (Zhang et al., 2016). Also, dibenzoylmethane (DBM) can protect against carbon tetrachloride (CCl4)-induced liver injury by activating Nrf2 signaling via JNK, AMPK, and calcium signaling (Cao et al., 2017). vii) In lung cancer, one study found that the potent anticancer agent isodeoxyelephantopin can induce protective autophagy in lung cancer cells via the Nrf2-p62-keap1 pathway (Wang et al., 2017). The Nrf2 activators, DMF and the synthetic oleanane triterpenoids, activate the Nrf2 pathway as well as regulate different subsets of Nrf2 target genes and Nrf2-independent genes in lung cancer (Chian et al., 2014; To et al., 2015). viii) In colon cancer, one study found that anticancer agent sulforaphane (SFN) can activate Nrf2 signaling to suppress human colon cancer (Johnson et al., 2017). Also, taxifolin (TAX) can induce antioxidant response pathway and enhance level of Nrf2 protein, and act as effective chemopreventive agent capable of modulating inflammation in colon cancer (Manigandan et al., 2015). ix) In ovarian cancer, one study found that Nrf2 can mediate the response of cancer cells to the anti-HER2 drugs, trastuzumab and pertuzumab, in ovarian cancer cells (Khalil et al., 2016). Also, activation of Nrf2 pathway in ovarian cancer seems to be related to Keap1 mutations within highly conserved domains of Keap1 gene and that Nrf2 may serve as an important therapeutic target for novel drugs capable of preventing or reversing resistance to chemotherapy in ovarian cancer (Konstantinopoulos et al., 2011). x) In breast cancer, Nrf2 serves as a key regulator in chemotherapeutic resistance under hypoxia through ROS-Nrf2-GCLC-GSH pathway and can be a potential treatment for hypoxia induced drug resistance in breast cancer cells (Song et al., 2011; Syu et al., 2016). xi) In esophageal cancer, C-28 methyl ester of 2-cyano-3,12-dioxoolean-1,9-dien-28-oic acid (CDDO-Me) can protect the cells against oxidative stress via inhibition of ROS generation, while CDDO-Me at low micromolar concentrations induces apoptosis by increasing ROS and decreasing intracellular glutathione levels in esophageal squamous cancer cells (Wang Y. Y. et al., 2015). xii) In glioblastoma, there are many potent anti-cancer agents targeting Nrf2 signaling for chemotherapy and chemoresistance in glioblastoma (Zhu et al., 2014). xiii) In osteosarcoma, the bioengineered Nrf2-siRNA can effectively interfere with the Nrf2 signaling pathway to improve chemosensitivity of human cancer cells (Li et al., 2018). Moreover, the PIM (proviral integration site for moloney murine leukemia virus) kinase inhibitors can reduce Nrf2 signaling and increase ROS to kill hypoxic tumor cells such as prostate cancer cells (PC4-LN4), colon cancer cells (HCT-116), and breast cancer cells (MB-MDA-231 and MB-MDA-231-ARE-Luc) (Warfel et al., 2016). One study shows that proteasome biogenesis is dependent on the Nrf2 transcriptional factor, thus proteasome inhibitors have been actively developed as potential anticancer drugs (Albornoz et al., 2019). Gallic acid (GA), Z-ligustilide (LIG), and senkyunolide A (SA) can individually or cooperatively target Nrf2/ARE pathway to prevent cancer (Liu et al., 2018). Therefore, it can be said that Keap1-Nrf2 signaling pathways have different roles at different stages of cancer (Leinonen et al., 2014; Furfaro et al., 2016; de la Vega et al., 2018). Multiple Nrf2 or Keap1 inhibitors have been reported; and some of them are in the stages of pre- and clinical trial towards the Nrf2 signaling for cancers. For example, sulforaphane can target Nrf2 and the Nrf2 target genes NQO1 and GCLC to prevent oral cancer, and a preclinical trail has been performed to study its chemopreventive activity for oral cancer (Bauman et al., 2016). A single centre, single arm prospective phase II clinical trial has been performed for phytosome complex of curcumin targeting Nrf2 signaling as a the complementary therapy of gemcitabine on pancreatic cancer (Pastorelli et al., 2018). However, none of these Nrf2 or Keap1 inhibitors have currently entered into real clinical applications, which suggests that the sole inhibition of Nrf2 might not be sufficient for anticancer. A rational combination of Nrf2 inhibitors with other chemical agents would be a better strategy to treat cancers (Zhang et al., 2019).
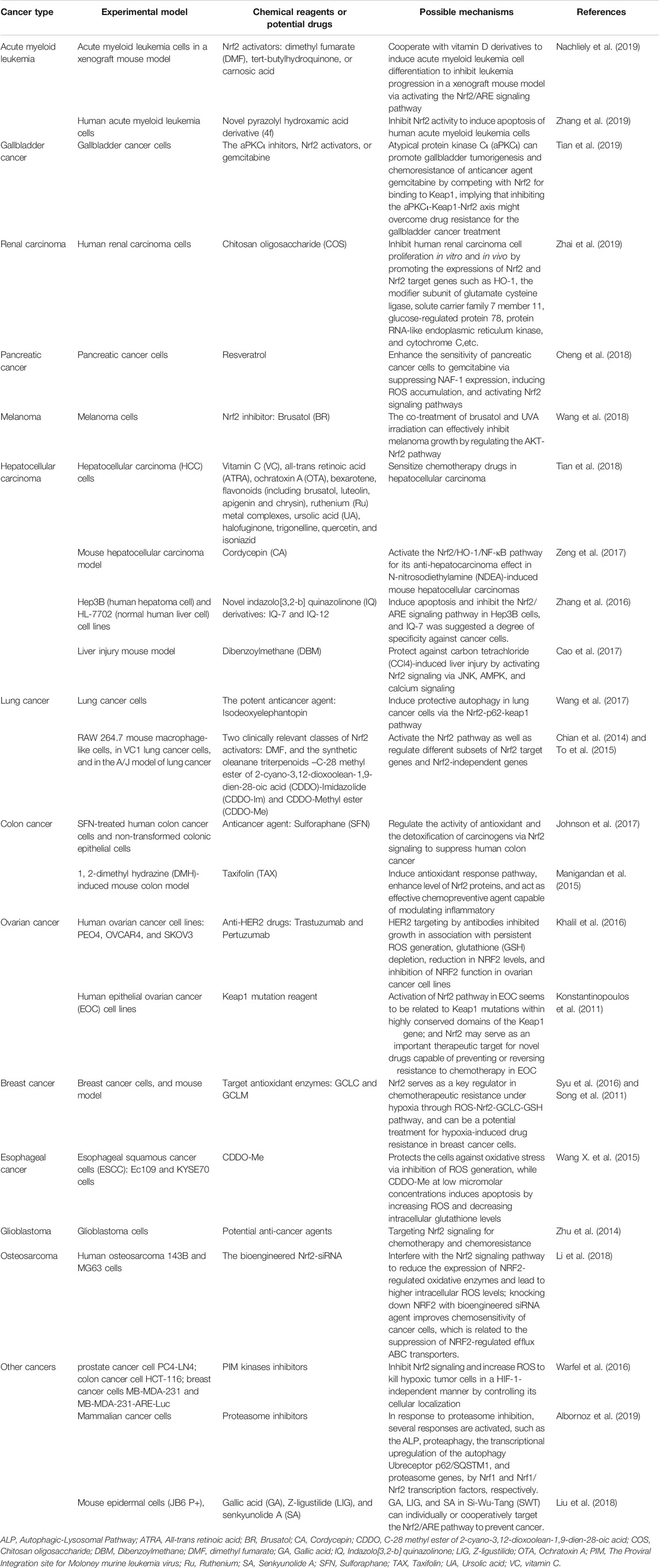
TABLE 1. Current research status of therapeutic potentials targeting Nrf2-mediated oxidative stress response signaling pathways in different cancers.
Potential of Targeting Nrf2 Signaling as New Therapeutic Strategy for PAs
As described above, many omics studies in human PA tissues and experimental studies in PA cells and animal models demonstrate that oxidative stress and oxidative damage is the important hallmark of PA pathogenesis. Nrf2-mediated oxidative stress response signaling pathways are at the heart of oxidative stress response, and many chemical agents targeting Nrf2 signaling pathways have been developed and tested as potential anticancer drugs for different cancers. This clearly demonstrates the potential of targeting Nrf2 signaling pathways as new therapeutic strategies for PAs. However, the use of Nrf2 signaling as a therapeutic target for PAs has not been studied. We strongly believe that the Nrf2-mediated oxidative stress response signaling pathways are the promising targets for novel therapeutic strategies for PAs. Furthermore, MAPK signaling pathways including ERK, JNK, and p38 MAPK clearly regulate Nrf2 signaling (Figure 1). Moreover, MAPK signaling pathways have been recognized as potential therapeutic targets for PAs (Lu et al., 2019). The combined use of Nrf2 inhibitors targeting Nrf2 signaling and ERK inhibitors [e.g., somatostatin analogs pasireotide (SOM230) and octreotide (OCT), or dopamine] plus p38 activators (e.g., cabergoline, bromocriptine, and fulvestrant) or JNK activators (e.g., ursolic acid, UA) targeting MAPK signaling pathways (Lu et al., 2019) might produce better anti-tumor effects on PAs. In addition, oxidative stress is tightly associated with mitochondrial dysfunctions, both operate in PAs (Zhan and Desiderio, 2010a; Li and Zhan, 2019; Long et al., 2019). Some drugs targeting mitochondria are also recognized as a therapeutic strategy for PAs, including pyrimethamine, temozolomide, melatonin, melatonin inhibitors, gossypol acetate, 18 beta-glycyrrhetinic acid, T-2 toxin, Yougui pill, cyclosporine A, grifolic acid, paeoniflorin, and dopamine agonists (Li and Zhan, 2019). Therefore, the combined use of Nrf2 inhibitors targeting Nrf2 signaling and drugs targeting metochondria could be another way to generate better anti-tumor effects on PAs.
Conclusion
Pituitary adenoma (PA) is a common and important disease that occurs in the hypothalamic-pituitary-target organ axis system and seriously affects human endocrine system and health. The imbalance between oxidative stress and the antioxidant defense system is an important pathophysiological characteristic in PAs, which has been evidenced by many omics analysis in PA tissues and experimental studies in PA cells and animal models. Nrf2 signaling is at the heart of oxidative stress response signaling pathways. Multiple anticancer agents targeting Nrf2-mediated oxidative stress response pathways have been developed and tested as potential therapeutic drugs for different cancers. However, Nrf2 signaling and targeting Nrf2 signaling as a therapeutic strategy has not yet been extensively studied in PAs. We strongly recommend the emphasis on in-depth studies of Nrf2 signaling and potential therapeutic agents targeting Nrf2 signaling pathways in PAs. Furthermore, the combined use of Nrf2 inhibitors targeting Nrf2 signaling and ERK inhibitors plus p38 activators or JNK activators targeting MAPK signaling pathways, or drugs targeting mitochondria dysfunction pathway might produce better anti-tumor effects on PAs.
Author Contributions
XZ conceived the concept, collected and analyzed literature, designed, coordinated, wrote and revised manuscript, and was responsible for its financial supports and the corresponding works. JL and TZ participated in literature collection and analysis, and prepared figures. All authors approved the final manuscript.
Funding
The financial supports from the Shandong First Medical University Talent Introduction Funds (to XZ), and the Hunan Provincial Hundred Talent Plan (to XZ).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest
Acknowledgments
The authors acknowledge the financial supports from the Shandong First Medical University Talent Introduction Funds (to XZ), and the Hunan Provincial Hundred Talent Plan (to XZ). We acknowledge that Xiaohan Zhan from Wesleyan University in United States critically revised the manuscript.
Glossary
ACTH Adrenocorticotropin
AFAR aldo-keto reductase family 7 member A2
AIP Aryl hydrocarbon receptor interacting protein gene
AKR Palmitoyltransferase AKT
AKR1B1 aldo-keto reductase family 1 member B
AKT Protein kinase B
AOX4 Aldehyde oxidase 4
AP-1 Activator protein-1
aPKCι Atypical protein kinase Cι
ARE Antioxidant response element
ARS Acute restraint stress
ASK1 Apoptosis signal-regulating kinase 1
ATF4 Activating transcription factor 4
ATP5A1 ATP synthase subunit alpha, mitochondrial
ATP5B ATP synthase, H+ transporting mitochondrial F1 complex, beta subunit
BACH1 Transcription regulator protein BACH1
Bax BCL2 associated X, apoptosis regulator
BCL2 BCL2 apoptosis regulator
BR Brusatol
CALM calmodulin
CAT catalase
CBP CREB-binding protein
CBR4 carbonyl reductase 4
CCL2 C-C motif chemokine ligand 2
CCT7 T-complex protein 1 subunit eta
c-FOS Proto-oncogene protein c-FOS
CLPP Caseinolytic protease
COS Chitosan oligosaccharide
COX6B cytochrome c oxidase subunit 6B
c-MAF (MAF) MAF bZIP transcription factor
c-Raf RAF proto-oncogene serine/threonine-protein kinase
CRS Chronic restraint stress
Cul3 Cullin 3-based ubiquitin E3 ligase complex
Cyp cytochrome P
CYP1A cytochrome P450 family 1 subfamily A
CYP2A cytochrome P450 family 2 subfamily A
CYP2C cytochrome P450 family 2 subfamily C
CYP3A cytochrome P450 family 3 subfamily A
CYP4A cytochrome P450 family 4 subfamily A
DBM Dibenzoylmethane
DEG Differentially expressed gene
DEP Differentially expressed protein
DMF Dimethyl fumarate
EPHX1 Epoxide hydrolase 1
EpRE Electrophile responsive element
ER endoplasmic reticulum
ERK Extracellular signal-related kinase
ERK1/2 mitogen-activated protein kinase
ERK5 mitogen-activated protein kinase
ERP29 endoplasmic reticulum protein 29
ESR1 estrogen receptor 1
ESR2 estrogen receptor 2
FKBP5 FK506-binding protein 5
FMO Dimethylaniline monooxygenase [N-oxide-forming]
FPA Functional pituitary adenoma
FRA1 Fos-related antigen 1
FSH Follicle-stimulating hormone
FSHRH Follicle-stimulating hormone-releasing hormone
FTH1 Ferritin heavy polypeptide 1
FTL1 ferritin light polypeptide
GA Gallic acid
GCLC glutamate-cysteine ligase catalytic subunit
GCLM glutamate-cysteine ligase modifier subunit
GH Growth hormone
GPX4 glutathione peroxidase 4
GPX's Glutathione peroxidases
GRP94 94 kD glucose-regulated protein
GSK3β glycogen synthase kinase 3β
GSR glutathione reductase
GST glutathione S-transferase
GSTM2 glutathione S-transferase mu 2
HERPUD1 Homocysteine-responsive endoplasmic reticulum-resident ubiquitin-like domain member 1 protein
HIP2 Ubiquitin-conjugating enzyme E2 K
HO-1 heme oxygenase 1
HSP22/40/90 heat shock proteins 22, 40 and 90
HSP27 heat shock protein 27
HSP70 (HSPA4) heat shock protein family A member 4
HSP90 heat shock protein 90
HSP90AA1 heat shock protein 90 alpha family class A member 1
HSP90AB1 heat shock protein 90 alpha family class B member 1
HSP94 heat shock protein 94
HSPA5 heat shock protein family A (Hsp70) member 5
HSPA8 heat shock protein family A (Hsp70) member 8
HSPA9 heat shock protein family A (Hsp70) member 9
HSPCA (HSP90AA1) heat shock protein 90 alpha family class A member 1
HSPCB (HSP90AB1) heat shock protein 90 alpha family class B member 1
IL-1β interleukin 1 beta
IL-6 interleukin 6
iNOS Inducible nitric synthase
IP3R (ITPR1) inositol 1,4,5-trisphosphate receptor type 1
Ipt Iptakalim
JNK Jun N-terminal kinase
JNK1 (MAPK8) mitogen-activated protein kinase 8
JNK 2 (MAPK9) mitogen-activated protein kinase 9
JUN Jun proto-oncogene, AP-1 transcription factor subunit
K-ATP ATP-sensitive potassium
Keap1 Kelch-like ECH-associated protein 1
LH Luteinizing hormone
LHRH Luteinizing hormone-releasing hormone
LIG Z-ligustilide
Maf Musculoaponeurotic fibrosarcoma
MAPKs Mitogen-activated protein kinases
MEK Mitogen-activated protein kinase kinase (MAPKK)
MEKK Mitogen-activated protein kinase kinase kinase (MAPKKK)
MN Micronucleus
MRP1 multidrug-resistant protein-1
NAC N-acetylcysteine
NAF-1 Nutrient-deprivation autophagy factor-1
NDEA N-nitrosodiethylamine
NDUFS8 (NADH) ubiquinone oxidoreductase core subunit S8
NF-kB Nuclear factor kB
NFPA Nonfunctional pituitary adenoma
NME1 NME/NM23 nucleoside diphosphate kinase 1
NO Nitric oxide
NQO1 NAD(P)H:quinine oxidoreductase 1
Nrf2 Nuclear factor erythroid 2 p45-related factor 2
Nur77 (NR4A1) nuclear receptor subfamily 4 group A member 1
O2.- Superoxide radical
8-OHdG 8-hydroxy-2’-deoxyguanosine
OCT octreotide
OH hydroxyl radical
ONOO- Peroxynitrite anion
PA Pituitary adenoma
PACAP38 Pituitary adenylate cyclase-activating polypeptide 38
PDK1 pyruvate dehydrogenase kinase 1
PERK the double-stranded RNA (PKR)-activated protein kinase-like eukaryotic initiation factor 2 kinase
PFK phosphofructokinase
PI3K Phosphatidylinositol 3 kinase
PINK1 PTEN-induced putative kinase protein 1
PKA cAMP dependent protein kinase
PKC protein kinase C
POMC proopiomelanocortin
PPIB Peptidyl-prolyl cis-trans isomerase B
PP2C putative protein phosphatase
PRDX1 peroxiredoxin 1
PRL Prolactin
PSM multiple subunits of the 20S proteasome
Pter Pterostilbene
PTPLAD1 3-hydroxyacyl-CoA dehydratase 3
Ras GTPase Ras
RNS Reactive nitrogen species
ROS Reactive oxygen species
SA Senkyunolide A
SFN Sulforaphane
SOD Superoxide dismutase
SOD1 superoxide dismutase 1
SOM230 somatostatin analogs pasireotide
SQSTM1 sequestosome-1 protein
SR-B1 Scavenger receptor class B member 1
STIP1 stress induced phosphoprotein 1
TAC Total antioxidant capability
TAK1 TGF beta-Activated Kinase 1
TGM2 transglutaminase 2
TLR4 toll like receptor 4
TNFα tumor necrosis factor alpha
TRXR1 thioredoxin reductase 1
TUBB tubulin beta class I
TUBB2A tubulin beta 2A class IIa
TXN1 thioredoxin
UBB Polyubiquitin-B
UB2R1 Ubiquitin-conjugating enzyme E2 R1
UGT UDP glucuronosyl transferase
USP14 ubiquitin-specific peptidase 14
UV ultraviolet
VCP valosin-containing protein
References
Afolabi, O. B., Oloyede, O. I., and Agunbiade, S. O. (2018). Inhibitory potentials of phenolic-rich extracts from Bridelia ferruginea on two key carbohydrate-metabolizing enzymes and Fe2+-induced pancreatic oxidative stress. J. Integrat Med. 16, 192–198. doi:10.1016/j.joim.2018.04.006
Albornoz, N., Bustamante, H., Soza, A., and Burgos, P. (2019). Cellular responses to proteasome inhibition: molecular mechanisms and beyond. Int. J. Mol. Sci. 20 (14), E3379. doi:10.3390/ijms20143379
An, J. J., Cho, S. R., Jeong, D. W., Park, K. W., Ahn, Y. S., and Baik, J. H. (2003). Anti-proliferative effects and cell death mediated by two isoforms of dopamine D2 receptors in pituitary tumor cells. Mol. Cell Endocrinol. 206 (1-2), 49–62. doi:10.1016/s0303-7207(03)00236-3
Baird, L., and Yamamoto, M. (2020). The molecular mechanisms regulating the KEAP1-NRF2 pathway. Mol. Cell Biol. 40 (13), e00099. doi:10.1128/MCB.00099-20
Bauman, J. E., Zang, Y., Sen, M., Li, C., Wang, L., Egner, P. A., et al. (2016). Prevention of carcinogen-induced oral cancer by sulforaphane. Cancer Prev. Res. 9 (7), 547–557. doi:10.1158/1940-6207.CAPR-15-0290
Bellezza, I., Giambanco, I., Minelli, A., Donato, R., Bellezza, I., et al. (2018). Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta Mol. Cel. Res. 1865 (5), 721–733. doi:10.1016/j.bbamcr.2018.02.010
Benlloch, M., Obrador, E., Valles, S. L., Rodriguez, M. L., Sirerol, J. A., Alcácer, J., et al. (2016). Pterostilbene decreases the antioxidant defenses of aggressive cancer cells in vivo: a physiological glucocorticoids- and Nrf2-dependent mechanism. Antioxid. Redox Signal 24 (17), 974–990. doi:10.1089/ars.2015.6437
Beranova-Giorgianni, S., Giorgianni, F., and Desiderio, D. M. (2002). Analysis of the proteome in the human pituitary. Proteomics 2, 534–542. doi:10.1002/1615-9861(200205)2:5<534::AID-PROT534>3.0.CO;2-K
Beranova-Giorgianni, S., Zhao, Y., Desiderio, D. M., and Giorgianni, F. (2006). Phosphoproteomic analysis of the human pituitary. Pituitary 9, 109–120. doi:10.1007/s11102-006-8916-x
Bitgen, N., Donmez-Altuntas, H., Bayram, F., Cakir, I., Hamurcu, Z., Diri, H., et al. (2016). Increased micronucleus, nucleoplasmic bridge, nuclear bud frequency and oxidative DNA damage associated with prolactin levels and pituitary adenoma diameters in patients with prolactinoma. Biotech. Histochem. 91 (2), 128–136. doi:10.3109/10520295.2015.1101163
Bocca, L., Valenti, S., Cuttica, C. M., Spaziante, R., Giordano, G., and Giusti, M. (2000). Nitric oxide biphasically modulates GH secretion in cultured cells of GH-secreting human pituitary adenomas. Minerva. Endocrinol. 25 (3-4), 55–59.
Cao, M., Wang, H., Guo, L., Yang, S., Liu, C., Khor, T. O., et al. (2017). Dibenzoylmethane protects against CCl4-induced acute liver injury by activating Nrf2 via JNK, AMPK, and calcium signaling. AAPS J. 19 (6), 1703–1714. doi:10.1208/s12248-017-0133-1
Ceccatelli, S., Hulting, A. L., Zhang, X., Gustafsson, L., Villar, M., and Hökfelt, T. (1993). Nitric oxide synthase in the rat anterior pituitary gland and the role of nitric oxide in regulation of LH secretion. Proc. Natl. Acad. Sci. U.S.A. 90 (23), 11292–11296. doi:10.1073/pnas.90.23.11292
Chen, Q. M., and Maltagliati, A. J. (2018). Nrf2 at the heart of oxidative stress and cardiac protection. Physiol. Genomics 50 (2), 77–97. doi:10.1152/physiolgenomics.00041.2017
Cheng, L., Yan, B., Chen, K., Jiang, Z., Zhou, C., Cao, J., et al. (2018). Resveratrol-induced downregulation of NAF-1 enhances the sensitivity of pancreatic cancer cells to gemcitabine via the ROS/Nrf2 signaling pathways. Oxid Med. Cell Longev. 2018, 9482018. doi:10.1155/2018/9482018
Cheng, T., Wang, Y., Lu, M., Zhan, X., Zhou, T., Li, B., et al. (2019). Quantitative analysis of proteome in non-functional pituitary adenomas: clinical relevance and potential benefits for the patients. Front. Endocrinol. 10, 854. doi:10.3389/fendo.2019.00854
Cheng, T., and Zhan, X. (2017). Pattern recognition for predictive, preventive, and personalized medicine in cancer. EPMA J. 8, 51–60. doi:10.1007/s13167-017-0083-9
Chian, S., Thapa, R., Chi, Z., Wang, X. J., and Tang, X. (2014). Luteolin inhibits the Nrf2 signaling pathway and tumor growth in vivo. Biochem. Biophys. Res. Commun. 447 (4), 602–608. doi:10.1016/j.bbrc.2014.04.039
Chu, F. F., Esworthy, R. S., Chu, P. G., Longmate, J. A., Huycke, M. M., Wilczynski, S., et al. (2004). Bacteria-induced intestinal cancer in mice with disrupted GPx1 and Gpx2 genes. Cancer Res. 64 (3), 962–968. doi:10.1158/0008-5472.can-03-2272
Cloer, E. W., Goldfarb, D., Schrank, T. P., Weissman, B. E., and Major, M. B. (2019). NRF2 activation in cancer: from DNA to protein. Cancer Res. 79 (5), 889–898. doi:10.1158/0008-5472.CAN-18-2723
Cuadrado, A., Rojo, A. I., Wells, G., Hayes, J. D., Cousin, S. P., Rumsey, W. L., et al. (2019). Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nat. Rev. Drug Discov. 18 (4), 295–317. doi:10.1038/s41573-018-0008-x
Cuttica, C. M., Giusti, M., Bocca, L., Sessarego, P., De Martini, D., Valenti, S., et al. (1997). Nitric oxide modulates in vivo and in vitro growth hormone release in acromegaly. Neuroendocrinology 66 (6), 426–431. doi:10.1159/000127268
Dayalan Naidu, S., and Dinkova-Kostova, A. T. (2020). KEAP1, a cysteine-based sensor and a drug target for the prevention and treatment of chronic disease. Open Biol. 10 (6), 200105. doi:10.1098/rsob.200105.106
de la Vega, M. R., Chapman, E., and Zhang, D. D. (2018). NRF2 and the hallmarks of cancer. Cancer Cell 34 (1), 21–43. doi:10.1016/j.ccell.2018.03.022
Deyu, H., Luqing, C., Xianglian, L., Pu, G., Qirong, L., Xu, W., et al. (2018). Protective mechanisms involving enhanced mitochondrial functions and mitophagy against T-2 toxin-induced toxicities in GH3 cells. Toxicol. Lett. 295, 41–53. doi:10.1016/j.toxlet.2018.05.041
Dhakshinamoorthy, S., and Porter, A. G. (2004). Nitric oxide-induced transcriptional up-regulation of protective genes by Nrf2 via the antioxidant response element counteracts apoptosis of neuroblastoma cells. J. Biol. Chem. 279 (19), 20096–20107. doi:10.1074/jbc.M312492200
Duvilanski, B. H., Zambruno, C., Seilicovich, A., Pisera, D., Lasaga, M., Diaz, M. C., et al. (1995). Role of nitric oxide in control of prolactin release by the adenohypophysis. Proc. Natl. Acad. Sci. U.S.A. 92 (1), 170–174. doi:10.1073/pnas.92.1.170
Ebokaiwe, A. P., Obeten, K. E., Okori, S. O., David, E. E., Olusanya, O., Chukwu, C. J., et al. (2020). Co-administration of selenium nanoparticles and metformin abrogate testicular oxidative injury by suppressing redox imbalance, augmenting sperm quality and Nrf2 protein expression in streptozotocin-induced diabetic rats. Biol. Trace Elem. Res. 198, 544. doi:10.1007/s12011-020-02082-2
Elchuri, S., Oberley, T. D., Qi, W., Eisenstein, R. S., Jackson Roberts, L., et al. (2005). CuZnSOD deficiency leads to persistent and widespread oxidative damage and hepatocarcinogenesis later in life. Oncogene 24 (3), 367–380. doi:10.1038/sj.onc.1208207
Evans, C. O., Moreno, C. S., Zhan, X., Mccabe, M. T., Vertino, P. M., Desiderio, D. M., et al. (2008). Molecular pathogenesios f human prolactinomasi dentified by genee xpressionp rofiling, RT-qPCR, and proteomic analyses. Pituitlry 11, 231–245. doi:10.1007/s11102-007-0082-2
Feng, J., Zhang, Q., Zhou, Y., Yu, S., Hong, L., Zhao, S., et al. (2018). Integration of proteomics and metabolomics revealed metabolite-protein networks in ACTH-secreting pituitary adenoma. Front. Endocrinol. 9, 678. doi:10.3389/fendo.2018.00678
Furfaro, A. L., Traverso, N., Domenicotti, C., Piras, S., Moretta, L., Marinari, U. M., et al. (2016). The Nrf2/HO-1 axis in cancer cell growth and chemoresistance. Oxid Med. Cell 2016, 1958174. doi:10.1155/2016/1958174
Galland, F., Lacroix, L., Saulnier, P., Dessen, P., Meduri, G., Bernier, M., et al. (2010). Differential gene expression profiles of invasive and non-invasive non-functioning pituitary adenomas based on microarray analysis. Endocr. Relat. Cancer 17, 361–371. doi:10.1677/ERC-10-0018
Giorgianni, F., Beranova-Giorgianni, S., and Desiderio, D. M. (2004). Identification and characterization of phosphorylated proteins in the human pituitary. Proteomics 4, 587–598. doi:10.1002/pmic.200300584
Giorgianni, F., Desiderio, D. M., and Beranova-Giorgianni, S. (2003). Proteome analysis using isoelectric focusing in immobilized pH gradient gels followed by mass spectrometry. Electrophoresis 24, 253–259. doi:10.1002/elps.200390021
Gonçalves, Á. d. C., Moreira, E. J. S., and Portari, G. V. (2019). Benfotiamine supplementation prevents oxidative stress in anterior tibialis muscle and heart. J. Integr. Med. 17, 423–429. doi:10.1016/j.joim.2019.07.001
Grech, G., Zhan, X., Yoo, B. C., Bubnov, R., Hagan, S., Danesi, R., et al. (2015). EPMA position paper in cancer: current overview and future perspectives. EPMA J. 6, 9. doi:10.1186/s13167-015-0030-6
Hambright, H. G., Meng, P., Kumar, A. P., and Ghosh, R. (2015). Inhibition of PI3K/AKT/mTOR axis disrupts oxidative stress-mediated survival of melanoma cells. Oncotarget 6 (9), 7195–7208. doi:10.18632/oncotarget.3131
Harris, I. S., Treloar, A. E., Inoue, S., Sasaki, M., Gorrini, C., Lee, K. C., et al. (2015). Glutathione and thioredoxin antioxidant pathways synergize to drive cancerinitiation and progression. Cancer Cell 27 (2), 211–222. doi:10.1016/j.ccell.2014.11.019
Hernández-Ramírez, L. C., Morgan, R. M. L., Barry, S., D'Acquisto, F., Prodromou, C., and Korbonits, M. (2018). Multi-chaperone function modulation and association with cytoskeletal proteins are key features of the function of AIP in the pituitary gland. Oncotarget 9 (10), 9177–9198. doi:10.18632/oncotarget.24183
Hetland, G., Tangen, J.-M., Mahmood, F., Mirlashari, M. R., Nissen-Meyer, L. S. H., Nentwich, I., et al. (2020). Antitumor, anti-Inflammatory and antiallergic effects of Agaricus blazei Mushroom extract and the related medicinal basidiomycetes mushrooms, hericium erinaceus and Grifola frondosa: a Review of Preclinical and Clinical Studies. Nutrients 12 (5), 1339. doi:10.3390/nu12051339
Hu, R., Wang, X., and Zhan, X. (2013). Multi-parameter systematic strategies for predictive, preventive and personalised medicine in cancer. EPMA J. 4, 2. doi:10.1186/1878-5085-4-2
Huang, C. S., Chen, H. W., Lin, T. Y., Lin, A. H., and Lii, C. K. (2018). Shikonin upregulates the expression of drug-metabolizing enzymes and drug transporters in primary rat hepatocytes. J. Ethnopharmacol. 216, 18–25. doi:10.1016/j.jep.2018.01.026
Hybertson, B. M., Gao, B., Bose, S. K., and McCord, J. M. (2011). Oxidative stress in health and disease: the therapeutic potential of Nrf2 activation. Mol. Aspects Med. 32 (4-6), 234–246. doi:10.1016/j.mam.2011.10.006
Ilhan, M., Turgut, S., Turan, S., Demirci Cekic, S., Ergen, H. A., Korkmaz Dursun, G., et al. (2018). The assessment of total antioxidant capacity and superoxide dismutase levels, and the possible role of manganese superoxide dismutase polymorphism in acromegaly. Endocr. J. 65 (1), 91–99. doi:10.1507/endocrj.EJ17-0300
Johnson, G. S., Li, J., Beaver, L. M., Dashwood, W. M., Sun, D., Rajendran, P., et al. (2017). A functional pseudogene, NMRAL2P, is regulated by Nrf2 and serves as a coactivator of NQO1 in sulforaphane-treated colon cancer cells. Mol. Nutr. Food Res. 61 (4). doi:10.1002/mnfr.201600769
Kang, S. W., Rhee, S. G., Chang, T. S., Jeong, W., and Choi, M. H. (2005). 2-Cys peroxiredoxin function in intracellular signal transduction: therapeutic implications. Trends Mol. Med. 11 (12), 571–578. doi:10.1016/j.molmed.2005.10.006
Khalil, H. S., Langdon, S. P., Goltsov, A., Soininen, T., Harrison, D. J., Bown, J., et al. (2016). A novel mechanism of action of HER2 targeted immunotherapy is explained by inhibition of NRF2 function in ovarian cancer cells. Oncotarget 7 (46), 75874–75901. doi:10.18632/oncotarget.12425
Klaunig, J. E. (2018). Oxidative stress and cancer. Curr. Pharm. Des. 24 (40), 4771–4778. doi:10.2174/1381612825666190215121712
Konstantinopoulos, P. A., Spentzos, D., Fountzilas, E., Francoeur, N., Sanisetty, S., Grammatikos, A. P., et al. (2011). Keap1 mutations and Nrf2 pathway activation in epithelial ovarian cancer. Cancer Res. 71 (15), 5081–5089. doi:10.1158/0008-5472.CAN-10-4668
Kruk, J., and Aboul-Enein, H. Y. (2017). Reactive oxygen and nitrogen species in carcinogenesis: implications of oxidative stress on the progression and development of several cancer types. Mini Rev. Med. Chem. 17 (11), 904–919. doi:10.2174/1389557517666170228115324
Kruse, A., Broholm, H., Rubin, I., Schmidt, K., and Lauritzen, M. (2002). Nitric oxide synthase activity in human pituitary adenomas. Acta Neurol. Scand. 106 (6), 361–366. doi:10.1034/j.1600-0404.2002.01138.x
Kudryavtseva, A. V., Krasnov, G. S., Dmitriev, A. A., Alekseev, B. Y., Kardymon, O. L., Sadritdinova, A. F., et al. (2016). Mitochondrial dysfunction and oxidative stress in aging and cancer. Oncotarget 7 (29), 44879–44905. doi:10.18632/oncotarget.9821
Kurisaka, M., Nakajo, T., and Mori, K. (2004). Immunohistochemical study on the expression of copper and zinc-superoxide dismutase (Cu, Zn-SOD) in human adenohypophysis and pituitary adenomas. No To Shinkei 46 (10), 948–954.
Kwak, M. K., and Kensler, T. W. (2010). Targeting NRF2 signaling for cancer chemoprevention. Toxicol. Appl. Pharmacol. 244 (1), 66–76. doi:10.1016/j.taap.2009.08.028
Leinonen, H. M., Kansanen, E., Pölönen, P., Heinäniemi, M., and Levonen, A. L. (2014). Role of the Keap1-Nrf2 pathway in cancer. Adv. Cancer Res. 122, 281–320. doi:10.1016/B978-0-12-420117-0.00008-6
Li, H., Cao, L., Yi, P. Q., Xu, C., Su, J., Chen, P. Z., et al. (2019). Pituitary adenylate cyclase-activating polypeptide ameliorates radiation-induced cardiac injury. Am. J. Transl. Res. 11 (10), 6585–6599. www.ajtr.org/ISSN:1943-8141/AJTR0102651.
Li, N., and Zhan, X. (2019). Mitochondrial dysfunction pathway networks and mitochondrial dynamics in the pathogenesis of pituitary adenomas. Front. Endocrinol. 10, 690. doi:10.3389/fendo.2019.00690
Li, P. C., Tu, M. J., Ho, P. Y., Jilek, J. L., Duan, Z., et al. (2018). Bioengineered NRF2-siRNA is effective to interfere with NRF2 pathways and improve chemosensitivity of human cancer cells. Drug Metab. Dispos. 46 (1), 2–10. doi:10.1124/dmd.117.078741
Li, W., Dong, S., Chen, Q., Chen, C., and Dong, Z. (2020). Selenium may suppress peripheral blood mononuclear cell apoptosis by modulating HSP70 and regulate levels of SIRT1 through reproductive hormone secretion and oxidant stress in women suffering fluorosis. Eur. J. Pharmacol. 878, 173098. doi:10.1016/j.ejphar.2020.173098
Li, W., and Kong, A. N. (2009). Molecular mechanisms of Nrf2-mediated antioxidant response. Mol. Carcinog. 48 (2), 91–104. doi:10.1002/mc.20465
Li, Y., Huang, T. T., Carlson, E. J., Melov, S., Ursell, P. C., Olson, J. L., et al. (1995). Dilated cardiomyopathy and neonatal lethality in mutant mice lacking mangese superoxide dismutase. Nat. Genet. 11 (4), 376–381. doi:10.1038/ng1295-376
Liguori, I., Russo, G., Curcio, F., Bulli, G., Aran, L., Della-Morte, D., et al. (2018). Oxidative stress, aging, and diseases. Clin. Interv. Aging 13, 757–772. doi:10.2147/CIA.S158513
Liu, M. M., Huang, K. M., Qian, L., Chatterjee, P., Zhang, S., Li, R., et al. (2018). Effects of bioactive constituents in the Traditional Chinese Medicinal formula Si-Wu-Tang on Nrf2 signaling and neoplastic cellular transformation. Phytomedicine 40, 1–9. doi:10.1016/j.phymed.2017.12.031
Liu, P., Tian, W., Tao, S., Tillotson, J., Wijeratne, E. M. K., Gunatilaka, A. A. L., et al. (2019). Non-covalent NRF2 activation confers greater cellular protection than covalent activation. Cell Chem Biol. 26 (10), 1427–1435. doi:10.1016/j.chembiol.2019.07.011
Lloyd, R. V., Jin, L., Qian, X., Zhang, S., and Scheithauer, B. W. (1995). Nitric oxide synthase in the human pituitary gland. Am. J. Pathol. 146 (1), 86–94.
Long, Y., Lu, M., Cheng, T., Zhan, X., and Zhan, X. (2019). Multiomics-based signaling pathway network alterations in human non-functional pituitary adenomas. Front. Endocrinol. 10, 835. doi:10.3389/fendo.2019.00835
Lopes, M. B. S. (2017). The 2017 World Health Organization classification of tumors of the pituitary gland: a summary. Acta Neuropathol. 134, 521–535. doi:10.1007/s00401-017-1769-8
Lu, M., Wang, Y., and Zhan, X. (2019). The MAPK pathway-based drug therapeutic targets in pituitary adenomas. Front. Endocrinol. 10, 330. doi:10.3389/fendo.2019.00330
Lu, M., and Zhan, X. (2018). The crucial role of multiomic approach in cancer research and clinically relevant outcomes. EPMA J. 9, 77–102. doi:10.1007/s13167-018-0128-8
Lu, M. C., Ji, J. A., Jiang, Z. Y., and You, Q. D. (2016). The Keap1-Nrf2-ARE pathway as a potential preventive and therapeutic target: an update. Med. Res. Rev. 36 (5), 924–963. doi:10.1002/med.21396
Lubelska, K., Wiktorska, K., Mielczarek, L., Milczarek, M., Zbroińska-Bregisz, I., and Chilmonczyk, Z. (2016). Sulforaphane regulates NFE2L2/Nrf2-Dependent xenobiotic metabolism phase II and phase III enzymes differently in human colorectal cancer and untransformed epithelial colon cells. Nutr. Cancer 68 (8), 1338–1348. doi:10.1080/01635581.2016.1224369
Mahran, Y. F. (2020). New insights into the protection of growth hormone in cisplatin-induced nephrotoxicity: the impact of IGF-1 on the keap1-nrf2/HO-1 signaling. Life Sci. 253, 117581. doi:10.1016/j.lfs.2020.117581
Manigandan, K., Manimaran, D., Jayaraj, R. L., Elangovan, N., Dhivya, V., and Kaphle, A. (2015). Taxifolin curbs NF-κB-mediated Wnt/β-catenin signaling via up-regulating Nrf2 pathway in experimental colon carcinogenesis. Biochimie 119, 103–112. doi:10.1016/j.biochi.2015.10.014
Mann, G. E., Rowlands, D. J., Li, F. Y., de Winter, P., and Siow, R. C. (2007). Activation of endothelial nitric oxide synthase by dietary isoflavones: role of NO in Nrf2-mediated antioxidant gene expression. Cardiovasc. Res. 75 (2), 261–274. doi:10.1016/j.cardiores.2007.04.004
McCann, S. M., Haens, G., Mastronardi, C., Walczewska, A., Karanth, S., Rettori, V., et al. (2003). The role of nitric oxide (NO) in control of LHRH release that mediates gonadotropin release and sexual behavior. Curr. Pharm. Des. 9 (5), 381–390. doi:10.2174/1381612033391766
McCann, S. M., Karanth, S., Mastronardi, C. A., Dees, W. L., Childs, G., Miller, B., et al. (2001). Control of gonadotropin secretion by follicle-stimulating hormone-releasing factor, luteinizing hormone-releasing hormone, and leptin. Arch. Med. Res. 32 (6), 476–485. doi:10.1016/s0188-4409(01)00343-5
Mehnati, P., Baradaran, B., Vahidian, F., and Nadiriazam, S. (2020). Functional response difference between diabetic/normal cancerous patients to inflammatory cytokines and oxidative stresses after radiotherapy. Rep. Pract. Oncol. Radiother. 25 (5), 730–737. doi:10.1016/j.rpor.2020.06.008
Melov, S., Doctrow, S. R., Schneider, J. A., Haberson, J., Patel, M., Coskun, P. E., et al. (2001). Lifepan extension and rescue of spongiform encephalopathy in superoxide dismutase 2 nullizygous mice treated with superoxide dismutasecatalase mimetics. J. Neurosci. 21 (21), 8348–8353. doi:10.1523/jneurosci.21-21-08348.2001
Menegon, S., Columbano, A., and Giordano, S. (2016). The dual roles of NRF2 in cancer. Trends Mol. Med. 22 (7), 578–593. doi:10.1016/j.molmed.2016.05.002
Mishra, P., Paital, B., Jena, S., Swain, S. S., Kumar, S., Yadav, M. K., et al. (2019). Possible activation of NRF2 by Vitamin E/Curcumin against altered thyroid hormone induced oxidative stress via NFĸB/AKT/mTOR/KEAP1 signalling in rat heart. Sci. Rep. 9, 7408. doi:10.1038/s41598-019-43320-5
Moreno, C. S., Evans, C. O., Zhan, X., Okor, M., Desiderio, D. M., and Oyesiku, N. M. (2005). Novel molecular signaling and classification of human clinically nonfunctional pituitary adenomas identified by gene expression profiling and proteomic analyses. Cancer Res. 65, 10214–10222. doi:10.1158/0008-5472.CAN-05-0884
Nachliely, M., Trachtenberg, A., Khalfin, B., Nalbandyan, K., Cohen-Lahav, M., Yasuda, K., et al. (2019). Dimethyl fumarate and vitamin D derivatives cooperatively enhance VDR and Nrf2 signaling in differentiating AML cells in vitro and inhibit leukemia progression in a xenograft mouse model. J. Steroid Biochem. Mol. Biol. 188, 8–16. doi:10.1016/j.jsbmb.2018.11.017
Neumann, C. A., Krause, D. S., Carman, C. V., Das, S., Dubey, D. P., Abraham, J. L., et al. (2003). Essential role for the peroxiredoxin Prdx1 in erythrocyte antioxidant defence and tumour suppression. Nature 424 (6948), 561–565. doi:10.1038/nature01819
Niture, S. K., and Jaiswal, A. K. (2010). Hsp90 interaction with INrf2(Keap1) mediates stress-induced Nrf2 activation. J. Biol. Chem. 285 (47), 36865–36875. doi:10.1074/jbc.M110.175802
Obrador, E., Liu-Smith, F., Dellinger, R. W., Salvador, R., Meyskens, F. L., and Estrela, J. M. (2019). Oxidative stress and antioxidants in the pathophysiology of malignant melanoma. Biol. Chem. 400 (5), 589–612. doi:10.1515/hsz-2018-0327
Osburn, W. O., Wakabayashi, N., Misra, V., Nilles, T., Biswal, S., Trush, M. A., et al. (2006). Nrf2 regulates an adaptive response protecting against oxidative damage following diquat-mediated formation of superoxide anion. Arch. Biochem. Biophys. 454 (1), 7–15. doi:10.1016/j.abb.2006.08.005
Pastorelli, D., Fabricio, A. S. C., Giovanis, P., D'Ippolito, S., Fiduccia, P., Soldà, C., et al. (2018). Phytosome complex of curcumin as complementary therapy of advanced pancreatic cancer improves safety and efficacy of gemcitabine: results of a prospective phase II trial. Pharmacol. Res. 132, 72–79. doi:10.1016/j.phrs.2018.03.013
Pawlikowshi, M., Winczyk, K., and Jaranowska, M. (2003). Immunohistochemical demonstration of nitric oxide synthase (NOS) in the normal rat pituitary gland, estrogen-induced rat pituitary tumor and human pituitary adenomas. Folia Histochem. Cytobiol 41 (2), 87–90.
Pi, J., Zhang, Q., Woods, C. G., Wong, V., Collins, S., and Andersen, M. E. (2008). Activation of Nrf2-mediated oxidative stress response in macrophages by hypochlorous acid. Toxicol. Appl. Pharmacol. 226 (3), 236–243. doi:10.1016/j.taap.2007.09.016
Pinilla, L., González, L. C., Tena-Sempere, M., Bellido, C., and Aguilar, E. (2001). Effects of systemic blockade of nitric oxide synthases on pulsatile LH, prolactin, and GH secretion in adult male rats. Horm. Res. 55 (5), 229–235. doi:10.1159/000050001
Pinilla, L., Tena-Sempere, M., and Aguilar, E. (1999). Nitric oxide stimulates growth hormone secretion in vitro through a calcium- and cyclic guanosine monophosphate-independent mechanism. Horm. Res. 51 (5), 242–247. doi:10.1159/000023378
Pizza, V., Agresta, A., D’Acunto, C. W., Festa, M., and Capasso, A. (2011). Neuroinflammation and ageing: current theories and an overview of the data. Rev. Recent Clin. Trials 6 (3), 189–203. doi:10.2174/157488711796575577
Prasad, S., Gupta, S. C., Pandey, M. K., Tyagi, A. K., and Deb, L. (2016). Oxidative stress and cancer: advances and challenges. Oxid Med. Cell Longev. 2016, 5010423. doi:10.1155/2016/5010423
Prevatto, J. P., Torres, R. C., Diaz, B. L., Silva, P. M. R. E., Martins, M. A., and Carvalho, V. F. (2017). Antioxidant treatment induces hyperactivation of the HPA Axis by upregulating ACTH receptor in the adrenal and downregulating glucocorticoid receptors in the pituitary. Oxid Med. Cell Longev. 2017, 4156361. doi:10.1155/2017/4156361
Qian, S., Golubnitschaja, O., and Zhan, X. (2019). Chronic inflammation: key player and biomarker-set to predict and prevent cancer development and progression based on individualized patient profiles. EPMA J. 10, 365–381. doi:10.1007/s13167-019-00194-x
Qian, S., Yang, Y., Li, N., Cheng, T., Wang, X., Liu, J., et al. (2018). Prolactin variants in human pituitaries and pituitary adenomas identified with two-dimensional gel electrophoresis and mass spectrometry. Front. Endocrinol. 9, 468. doi:10.3389/fendo.2018.00468
Qin, J. J., Cheng, X. D., Zhang, J., and Zhang, W. D. (2019). Dual roles and therapeutic potential of Keap1-Nrf2 pathway in pancreatic cancer: a systematic review. Cell Commun. Signal 17 (1), 121. doi:10.1186/s12964-019-0435-2
Reimondo, G., Puglisi, S., Pia, A., and Terzolo, M. (2019). Autonomous hypercortisolism: definition and clinical implications. Minerva. Endocrinol. 44, 33–42. doi:10.23736/s0391-1977.18.02884-5
Reuter, S., Gupta, S. C., Chaturvedi, M. M., and Aggarwal, B. B. (2010). Oxidative stress, inflammation, and cancer: how are they linked? Free Radic. Biol. Med. 49 (11), 1603–1616. doi:10.1016/j.freeradbiomed.2010.09.006
Riedel, W. (2002). Role of nitric oxide in the control of the hypothalamic-pituitary-adrenocortical axis. Z. Rheumatol. 59 (Suppl. 2), 42. doi:10.1007/s003930070016
Roy Chowdhury, S., Sengupta, S., Biswas, S., Sinha, T. K., Sen, R., Basak, R. K., et al. (2014). Bacterial fucose-rich polysaccharide stabilizes MAPK-mediated Nrf2/Keap1 signaling by directly scavenging reactive oxygen species during hydrogen peroxide-induced apoptosis of human lung fibroblast cells. PLoS One 9 (11), e113663. doi:10.1371/journal.pone.0113663
Ryoo, I. G., and Kwak, M. K. (2018). Regulatory crosstalk between the oxidative stress-related transcription factor Nfe2l2/Nrf2 and mitochondria. Toxicol. Appl. Pharmacol. 359, 24–33. doi:10.1016/j.taap.2018.09.014
Saad El-Din, S., Rashed, L., Medhat, E., Emad Aboulhoda, B., Desoky Badawy, A., Mohammed ShamsEldeen, A., et al. (2020). Active form of vitamin D analogue mitigates neurodegenerative changes in Alzheimer's disease in rats by targeting Keap1/Nrf2 and MAPK-38p/ERK signaling pathways. Steroids 156, 108586. doi:10.1016/j.steroids.2020.108586
Sabatino, M. E., Grondona, E., Sosa, L. D. V., Mongi Bragato, B., Carreño, L., Juarez, V., et al. (2018). Oxidative stress and mitochondrial adaptive shift during pituitary tumoral growth. Free Radic. Biol. Med. 120, 41–55. doi:10.1016/j.freeradbiomed.2018.03.019
Sahin, K., Orhan, C., Akdemir, F., Tuzcu, M., Iben, C., and Sahin, N. (2012). Resveratrol protects quail hepatocytes against heat stress: modulation of the Nrf2 transcription factor and heat shock proteins. J. Anim. Physiol. Anim. Nutr. 96 (1), 66–74. doi:10.1111/j.1439-0396.2010.01123.x
Sajadimajd, S., and Khazaei, M. (2018). Oxidative stress and cancer: the role of Nrf2. Curr. Cancer Drug Targets 18 (6), 538–557. doi:10.2174/1568009617666171002144228
Sánchez-Martín, P., Sou, Y. S., Kageyama, S., Koike, M., Waguri, S., and Komatsu, M. (2020). NBR1-mediated p62-liquid droplets enhance the Keap1-Nrf2 system. EMBO Rep. 21 (3), e48902. doi:10.15252/embr.201948902
Sivils, J. C., Ancrum, T. M., and Bain, L. J. (2013). LOSS of Mrp1 alters detoxification enzyme expression in a tissue- and hormonal-status-specific manner. J. Appl. Toxicol. 33 (8), 766–773. doi:10.1002/jat.2727
Song, M. K., Lee, J. H., Ryoo, I. G., Lee, S. H., Ku, S. K., and Kwak, M. K. (2019). Bardoxolone ameliorates TGF-β1-associated renal fibrosis through Nrf2/Smad7 elevation. Free Radic. Biol. Med. 138, 33–42. doi:10.1016/j.freeradbiomed.2019.04.033
Song, N. Y., Kim, D. H., Kim, E. H., Na, H. K., Kim, N. J., Suh, Y. G., et al. (2011). Multidrug resistance-associated protein 1 mediates 15-deoxy-Δ(12,14)-prostaglandin J2-induced expression of glutamate cysteine ligase expression via Nrf2 signaling in human breast cancer cells. Chem. Res. Toxicol. 24 (8), 1231–1241. doi:10.1021/tx200090n
Stalla, G. K., Ciato, D., and Dimopoulou, C. (2019). “The adrenal gland: central relay in health and disease - current challenges and perspectives 2018” - Cushing’s disease. Exp. Clin. Endocrinol. Diabetes 127, 147–155. doi:10.1055/a-0664-7632
Sun, T., Gao, J., Han, D., Shi, H., and Liu, X. (2019). Fabrication and characterization of solid lipid nano-formulation of astraxanthin against DMBA-induced breast cancer via Nrf-2-Keap1 and NF-kB and mTOR/Maf-1/PTEN pathway. Drug Deliv. 26 (1), 975–988. doi:10.1080/10717544.2019.1667454
Syu, J. P., Chi, J. T., and Kung, H. N. (2016). Nrf2 is the key to chemotherapy resistance in MCF7 breast cancer cells under hypoxia. Oncotarget 7 (12), 14659–14672. doi:10.18632/oncotarget.7406
Taguchi, K., and Yamamoto, M. (2017). The KEAP1-NRF2 system in Cancer. Front. Oncol. 7, 85. doi:10.3389/fonc.2017.00085
Tian, B., Lu, Z. N., and Guo, X. L. (2018). Regulation and role of nuclear factor-E2-related factor 2 (Nrf2) in multidrug resistance of hepatocellular carcinoma. Chem. Biol. Interact 280, 70–76. doi:10.1016/j.cbi.2017.12.014
Tian, H., Zhang, D., Gao, Z., Li, H., Zhang, B., Zhang, Q., et al. (2014). MDA-7/IL-24 inhibits Nrf2-mediated antioxidant response through activation of p38 pathway and inhibition of ERK pathway involved in cancer cell apoptosis. Cancer Gene Ther. 21 (10), 416–426. doi:10.1038/cgt.2014.45
Tian, L., Lu, Y., Yang, T., Deng, Z., Xu, L., Yao, W., et al. (2019). aPKCι promotes gallbladder cancer tumorigenesis and gemcitabine resistance by competing with Nrf2 for binding to Keap1. Redox Biol. 22, 101149. doi:10.1016/j.redox.2019.101149
To, C., Ringelberg, C. S., Royce, D. B., Williams, C. R., Risingsong, R., Sporn, M. B., et al. (2015). Dimethyl fumarate and the oleanane triterpenoids, CDDO-imidazolide and CDDO-methyl ester, both activate the Nrf2 pathway but have opposite effects in the A/J model of lung carcinogenesis. Carcinogenesis 36 (7), 769–781. doi:10.1093/carcin/bgv061
Ueta, Y., Levy, A., Powell, M. P., Lightman, S. L., Kinoshita, Y., Yokota, A., et al. (1998). Neuronal nitric oxide synthase gene expression in human pituitary tumours: a possible association with somatotroph adenomas and growth hormone-releasing hormone gene expression. Clin. Endocrinol. 49 (1), 29–38. doi:10.1046/j.1365-2265.1998.00399.x
Valko, M., Rhodes, C. J., Moncol, J., Izakovic, M., and Mazur, M. (2006). Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact 160 (1), 1–40. doi:10.1016/j.cbi.2005.12.009
Vankelecom, H., Matthys, P., and Denef, C. (1997). Inducible nitric oxide synthase in the anterior pituitary gland: induction by interferon-r in a subpopulation of folliculostellate cells and in an unidentifiable population of nonhormone-secreting cells. J. Histochem. Cytochem. 45 (6), 847–857. doi:10.1177/002215549704500609
Villeneuve, N. F., Lau, A., and Zhang, D. D. (2010). Regulation of the Nrf2–Keap1 antioxidant response by the ubiquitin proteasome system: an insight into cullin-ring ubiquitin ligases. Antioxid. Redox Signal. 13 (11), 1699–1712. doi:10.1089/ars.2010.3211
Wang, K.-C., Liu, Y.-C., El-Shazly, M., Shih, S.-P., Du, Y.-C., Hsu, Y.-M., et al. (2019). The antioxidant from ethanolic extract of Rosa cymosa fruits activates phosphatase and tensin homolog in vitro and in vivo: a new insight on its antileukemic effect. Int. J. Mol. Sci. 20 (8), 1935. doi:10.3390/ijms20081935
Wang, M., Shi, G., Bian, C., Nisar, M. F., Guo, Y., Wu, Y., et al. (2018). UVA irradiation enhances brusatol-mediated inhibition of melanoma growth by downregulation of the Nrf2-mediated antioxidant response. Oxid Med. Cell Longev. 2018, 9742154. doi:10.1155/2018/9742154
Wang, X., Guo, T., Peng, F., Long, Y., Mu, Y., Yang, H., et al. (2015). Proteomic and functional profiles of a follicle-stimulating hormone positive human nonfunctional pituitary adenoma. Electrophoresis 36, 1289–1304. doi:10.1002/elps.201500006
Wang, Y., Cheng, T., Lu, M., Mu, Y., Li, B., Li, X., et al. (2019). TMT-based quantitative proteomics revealed follicle-stimulating hormone (FSH)-related molecular characterizations for potentially prognostic assessment and personalized treatment of FSH-positive non-functional pituitary adenomas. EPMA J. 10, 395–414. doi:10.1007/s13167-019-00187-w
Wang, Y., Zhang, J., Huang, Z. H., Huang, X. H., Zheng, W. B., Yin, X. F., et al. (2017). Isodeoxyelephantopin induces protective autophagy in lung cancer cells via Nrf2-p62-keap1 feedback loop. Cell Death Dis. 8 (6), e2876. doi:10.1038/cddis.2017.265
Wang, Y. Y., Yang, Y. X., Zhao, R., Pan, S. T., Zhe, H., He, Z. X., et al. (2015). Bardoxolone methyl induces apoptosis and autophagy and inhibits epithelial-to-mesenchymal transition and stemness in esophageal squamous cancer cells. Drug Des. Devel Ther. 9, 993–1026. doi:10.2147/DDDT.S73493
Warfel, N. A., Sainz, A. G., Song, J. H., and Kraft, A. S. (2016). PIM kinase inhibitors kill hypoxic tumor cells by reducing Nrf2 signaling and increasing reactive oxygen species. Mol. Cancer Ther. 15 (7), 1637–1647. doi:10.1158/1535-7163.MCT-15-1018
Wu, C., Zhang, H., Zhang, J., Zhang, H., Zeng, Y., Fang, S., et al. (2019). Increased oxidative stress, inflammation and fibrosis in perirenal adipose tissue of patients with cortisol-producing adenoma. Adipocyte 8 (1), 347–356. doi:10.1080/21623945.2019.1690834
Yang, J., Zhang, C., Zhang, W., Shi, R., and Zhang, Z. (2012). Extracellular superoxide dismutase, a potential extracellular biomarker candidate for prolactinoma. West Indian Med. J. 61 (7), 665–669.
Yates, M. S., and Kensler, T. W. (2007). Chemopreventive promise of targeting the Nrf2 pathway. Drug News Perspect. 20 (2), 109–117. doi:10.1358/dnp.2007.20.2.108343
Yu, Z., Dai, Z. Y., Qin, G. X., Li, M. Y., and Wu, L. F. (2020). Alleviative effects of dietary microbial floc on copper-induced inflammation, oxidative stress, intestinal apoptosis and barrier dysfunction in Rhynchocypris lagowski Dybowski. Fish Shellfish Immunol. 106, 120–132. doi:10.1016/j.fsi.2020.07.070
Zeng, Y., Lian, S., Li, D., Lin, X., Chen, B., Wei, H., et al. (2017). Anti-hepatocarcinoma effect of cordycepin against NDEA-induced hepatocellular carcinomas via the PI3K/Akt/mTOR and Nrf2/HO-1/NF-κB pathway in mice. Biomed. Pharmacother. 95, 1868–1875. doi:10.1016/j.biopha.2017.09.069
Zhai, X., Yuan, S., Yang, X., Zou, P., Li, L., Li, G., et al. (2019). Chitosan oligosaccharides induce apoptosis in human renal carcinoma via reactive-oxygen-species-dependent endoplasmic reticulum stress. J. Agric. Food Chem. 67 (6), 1691–1701. doi:10.1021/acs.jafc.8b06941
Zhan, X., and Desiderio, D. M. (2003). A reference map of a human pituitary adenoma proteome. Proteomics 3, 699–713. doi:10.1002/pmic.200300408
Zhan, X., and Desiderio, D. M. (2006). Nitroproteins from a human pituitary adenoma tissue discovered with a nitrotyrosine affinity column and tandem mass spectrometry. Anal. Biochem. 354, 279–289. doi:10.1016/j.ab.2006.05.024
Zhan, X., and Desiderio, D. M. (2010a). Signaling pathway networks mined from human pituitary adenoma proteomics data. BMC Med. Genomics 3, 13. doi:10.1186/1755-8794-3-13
Zhan, X., and Desiderio, D. M. (2010b). The use of variations in proteomes to predict, prevent, and personalize treatment for clinically nonfunctional pituitary adenomas. EPMA J. 1, 439–459. doi:10.1007/s13167-010-0028-z
Zhan, X., and Desiderio, D. M. (2004). The human pituitary nitroproteome: detection of nitrotyrosyl-proteins with two-dimensional Western blotting, and amino acid sequence determination with mass spectrometry. Biochem. Biophys. Res. Commun. 325, 1180–1186. doi:10.1016/j.bbrc.2004.10.169
Zhan, X., Wang, X., and Cheng, T. (2016). Human pituitary adenoma proteomics: new progresses and perspectives. Front. Endocrinol. 7, 54. doi:10.3389/fendo.2016.00054
Zhan, X., Wang, X., and Desiderio, D. M. (2013). Pituitary adenoma nitroproteomics: current status and perspectives. Oxid Med. Cel Longev. 2013, 580710. doi:10.1155/2013/580710
Zhan, X., Wang, X., Long, Y., and Desiderio, D. M. (2014a). Heterogeneity analysis of the proteomes in clinically nonfunctional pituitary adenomas. BMC Med. Genomics 7, 69. doi:10.1186/s12920-014-0069-6
Zhan, X., Desiderio, D. M., Wang, X., Zhan, X., Guo, T., and Li, M. (2014b). Identification of the proteomic variations of invasive relative to non-invasive non-functional pituitary adenomas. Electrophoresis 35, 2184–2194. doi:10.1002/elps.201300590
Zhan, X., and Desiderio, D. M. (2007). Linear ion-trap mass spectrometric characterization of human pituitary nitrotyrosine-containing proteins. Int. J. Mass. Spectrom. 259, 96–104. doi:10.1016/j.ijms.2006.06.009
Zhang, B., Ma, Z., Tan, B., and Lin, N. (2019). Targeting the cell signaling pathway Keap1-Nrf2 as a therapeutic strategy for adenocarcinomas of the lung. Expert Opin. Ther. Targets 23 (3), 241–250. doi:10.1080/14728222.2019.1559824
Zhang, J., Su, L., Ye, Q., Zhang, S., Kung, H., Jiang, F., et al. (2017). Discovery of a novel Nrf2 inhibitor that induces apoptosis of human acute myeloid leukemia cells. Oncotarget 8 (5), 7625–7636. doi:10.18632/oncotarget.13825
Zhang, Y., Qiao, R., He, D., Zhao, Z., Yang, S., Zou, H., et al. (2016). Indazolo[3,2-b]quinazolinones attack hepatocellular carcinoma Hep3B cells by inducing mitochondrial-dependent apoptosis and inhibition of Nrf2/ARE signaling pathway. Curr. Mol. Med. 16 (9), 820–828. doi:10.2174/1566524016666161128114444
Zhao, L., Liu, Z., Jia, H., Feng, Z., Liu, J., and Li, X. (2015). Lipoamide acts as an indirect antioxidant by simultaneously stimulating mitochondrial biogenesis and phase II antioxidant enzyme systems in ARPE-19 cells. PLoS One 10 (6), e0128502. doi:10.1371/journal.pone.0128502
Zhao, X. J., Zhao, Z., Yang, D. D., Cao, L. L., Zhang, L., Ji, J., et al. (2017). Activation of ATP-sensitive potassium channel by iptakalim normalizes stress-induced HPA axis disorder and depressive behaviour by alleviating inflammation and oxidative stress in mouse hypothalamus. Brain Res. Bull. 130, 146–155. doi:10.1016/j.brainresbull.2017.01.026
Zhao, Y., Giorgianni, F., Desiderio, D. M., Fang, B., and Beranova-Giorgianni, S. (2005). Toward a global analysis of the human pituitary proteome by multiple gel-based technology. Anal. Chem. 77, 5324–5331. doi:10.1021/ac050354e
Zhou, W., Song, Y., Xu, H., Zhou, K., Zhang, W., Chen, J., et al. (2011). Nonfunctional pituitary adenomas, estrogen receptors and slug contribute to development of invasiveness. J. Clin. Endocrinol. Metab. 96, E1237–E1245. doi:10.1210/jc.2010-3040
Keywords: pituitary adenoma, oxidative stress, Nrf2, signaling pathway, biomarker, therapeutic target and drug
Citation: Zhan X, Li J and Zhou T (2021) Targeting Nrf2-Mediated Oxidative Stress Response Signaling Pathways as New Therapeutic Strategy for Pituitary Adenomas. Front. Pharmacol. 12:565748. doi: 10.3389/fphar.2021.565748
Received: 26 May 2020; Accepted: 12 February 2021;
Published: 24 March 2021.
Edited by:
Feng Zhu, Zhejiang University, ChinaReviewed by:
Chen Ling, Fudan University, ChinaQingxia Yang, Nanjing University of Posts and Telecommunications, China
Copyright © 2021 Zhan, Li and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xianquan Zhan, eWp6aGFuMjAxMUBnbWFpbC5jb20=