 Katherine E. Larrimore
Katherine E. Larrimore Yevgeniy V. Serebrenik
Yevgeniy V. Serebrenik- Department of Biochemistry and Molecular Biology, Robert Wood Johnson Medical School, Rutgers University, Piscataway, NJ, United States
Multifunctional ligand-binding domains have revolutionized cell biology by enabling the precise visualization and manipulation of fused proteins of interest through small molecule probes. Employing these domains is labor-intensive and low-throughput, limiting studies to only small subsets of proteins at a time. However, advancements in high-throughput technologies like pooled protein tagging have enabled the tagging of proteins across the entire proteome with ligand-binding domains. This allows for the generation of complex cell libraries where each cell expresses a different protein fused to a generic, ligandable handle. These libraries unlock opportunities to explore the proteome with a versatile toolbox of small molecule probes designed to interact with the fused tags, allowing researchers to visualize protein localization, induce protein misfolding, manipulate protein-protein interactions, modulate protein stability, and more in a scalable, systematic manner. This review explores recent studies employing multifunctional domains at proteome scale, delving into how the associated chemical probes have been employed to enable insights into endogenous protein function, cellular processes, and disease mechanisms. Additional multifunctional ligand-binding domains are discussed that can be used in pooled protein tagging, as well as their potential strengths and weaknesses. We also discuss potential applications in drug discovery such as high-throughput screening for therapeutic targets with insights for bifunctional ligand optimization. By integrating pooled protein tagging with ligand-binding domains and chemical probes, we highlight how the fusion of chemical biology and functional genomics is paving the way for innovative research avenues and transformative advances in cell biology and pharmacology.
1 Introduction
Genetic tagging of cellular proteins has transformed molecular and cellular biology research by enabling precise control over protein behavior, allowing researchers to study protein localization, function, interactions, and local environment. Ligand-binding domains are a family of protein tags that bind small molecule ligands with a broad range of applications, enabling precise dissection of protein function in living cells based on the chosen ligand. Despite the versatility of ligand-binding domains, their large-scale application has been hindered by the challenges associated with single-gene tagging approaches. Traditional methods for endogenous tagging, such as homologous recombination or CRISPR-mediated knock-in, are labor-intensive and inherently low-throughput, requiring individual genetic modifications for each target protein followed by screening for properly-modified clones. As a result, studies employing multifunctional ligand-binding domains have typically been restricted to focused experiments studying individual proteins. To address some of these limitations, arrayed tagging of endogenous genes by homology directed repair (HDR) with a small protein tag has provided an effective solution for systematic protein labeling (Cho et al., 2022). However, even with these advancements, the need for gene-specific donor designs has remained a bottleneck. Overexpression-based protein fusions that use ectopically expressed proteins fused to tags reveal insight into protein function at scale (Yen et al., 2008; Poirson et al., 2024) although time-intensive molecular cloning and differences in expression levels relative to endogenous contexts should be taken into consideration.
Recent advancements in genomic engineering have enabled endogenous pooled protein tagging approaches, facilitating the systematic study of protein function at scale. Such strategies, when combined with multifunctional ligand-binding domains, allow for proteome-wide interrogation of endogenous protein dynamics using a common set of chemical probes. This review explores key studies that have leveraged protein tagging to enable high-throughput imaging, protein destabilization, induced proximity, and targeted protein degradation. Additionally, we examine the latest technological developments in genome-scale tagging and their implications for studying protein function in complex cellular systems. Finally, we consider future directions, including promising ligandable domains for use in pooled systems, as well as the application of these tools for drug discovery. By integrating chemical biology with pooled screening methodologies, this emerging field is poised to provide transformative insights into proteome organization, protein function, and therapeutic target discovery.
2 Proteome-scale pooled tagging approaches
CRISPR-based endogenous gene tagging methods and ORFeome libraries have made it possible to move beyond single-protein experiments and systematically study thousands of proteins in a pool. These large-scale approaches are vital for systematic functional studies, especially when combined with chemical probes to interrogate protein behavior, localization, and interactions on a proteome-wide level. Tagging endogenous genes allows for the opportunity to investigate proteins under their native regulatory control, allowing capture of physiological expression patterns, stoichiometries, and post-transcriptional regulation. While the aforementioned properties are often lost in overexpression systems, better control over the expression and composition of the expressed genes provides alternative benefits. In this section, we review the state-of-the-art approaches for generating pooled tag libraries.
2.1 Retroviral-based tagging systems
Early generic tagging strategies used retroviral vectors to insert synthetic exons carrying a fluorescent protein flanked by splice acceptor and donor sites to a random location in the genome (Jarvik et al., 1996, Jarvik et al., 2002; Sigal et al., 2006a; Sigal et al., 2006b; Cohen et al., 2008). Although cell lines generated in these initial approaches were not studied in a pooled fashion, they laid groundwork for methods such as ORFtag (Nemčko et al., 2024), which uses a splice donor in a retroviral cassette to achieve internal fusions under an exogenous promoter. These systems can be pooled, and antibiotic markers can be included in the cassette to enable straightforward selection of cells that have integrated the tag (Figure 1A). Identification of tagged sites is performed by inverse PCR-related methods (Nemčko et al., 2024).
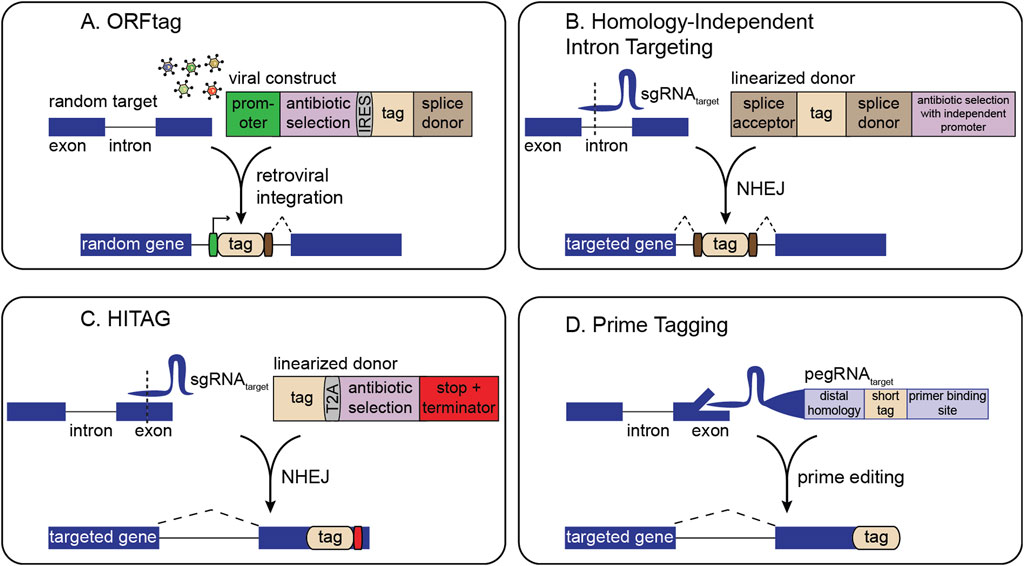
Figure 1. Overview of strategies for pooled gene tagging. (A) ORFtag utilizes viral integration of a synthetic exon with a splice donor to drive the expression of protein fragments. (B) Homology-independent intron targeting utilizes site-specific integration of a synthetic exon for internal protein tagging. (C) HITAG utilizes site-specific integration of a tag targeting exonic regions near stop codons. (D) Prime tagging uses prime editing to precisely insert short tags directly at protein termini (C-terminal tagging depicted here).
2.2 CRISPR-based pooled tagging
Several methodologies harness CRISPR-induced double-strand breaks (DSBs) combined with non-homologous end-joining (NHEJ) for tag insertion. Here, the only gene-specific reagent required is a single-guide RNA (sgRNA) to direct Cas9 to the locus of interest. A second sgRNA can linearize a generic donor plasmid, facilitating homology-independent tag integration (Figures 1B,C). This design allows the generation of complex cell libraries in which each cell carries a distinct, endogenously tagged protein. For instance, homology-independent intron targeting from Shalem and colleagues integrates synthetic exons within introns, leveraging the fact that introns often harbor many viable CRISPR target sites (Figure 1B) (Serebrenik et al., 2019; Serebrenik et al., 2024; Reicher et al., 2020; Reicher et al., 2024; Sansbury et al., 2025). The resulting fusions are scarless, as even if indels occur, they are outside the splice sites and thus restricted to the intron rather than the coding sequence. Selection markers can also be placed outside the splice sites for easy enrichment of genomic integration events without disrupting the tagged protein. This approach predominantly generates internal gene fusions, thus enabling screening of many fusion varieties for each gene, increasing the likelihood of finding non-disruptive insertion points.
Other NHEJ-driven methods favor inserting tags within exons at or near protein termini (Lackner et al., 2015; Schmid-Burgk et al., 2016; Suzuki et al., 2016). Although NHEJ-based tag integration tends to be high fidelity, exon tagging may result in frameshifts at either end of the tag. Thus, current pooled NHEJ-based exon tagging approaches, such as the ‘high-throughput insertion of tags across the genome’ (HITAG) system, rely on C-terminal tagging where in-frame fusion at the 5′ end is ensured by downstream selection markers, and an exogenous stop codon and termination sequence is used at the 3′ end (Figure 1C) (Kim et al., 2024). NHEJ-based tagging libraries can be analyzed either by inverse PCR or by sgRNA sequencing, making them easily compatible with methods such as single cell RNA sequencing or in situ sequencing (Feldman et al., 2019; Sansbury et al., 2025).
Precise, indel-free N- or C-terminal tagging of endogenous genes can be achieved by prime editing-based pooled tagging, which avoids NHEJ and provides flexibility in choosing the integration site relative to the target sequence (Figure 1D) (Sanchez et al., 2024). Although powerful, pooled prime tagging is currently limited to tags that can be encoded within a pegRNA. Future pooled prime editing approaches can incorporate recombinase landing pads to integrate larger tags (Anzalone et al., 2022; Yarnall et al., 2023; Pandey et al., 2025).
2.3 ORF libraries and overexpression approaches
For studying proteins that are not natively expressed in a given cell line, or where robust overexpression is desired, researchers can turn to exogenous expression libraries (Yang et al., 2011; ORFeome Collaboration, 2016; Tycko et al., 2020; Tycko et al., 2024; Alerasool et al., 2022; DelRosso et al., 2023; Lacoste et al., 2024; Moon et al., 2024; Poirson et al., 2024; Wilson et al., 2024). These libraries, often driven by strong promoters, can also be tagged with fluorophores or multifunctional domains. However, overexpression can introduce non-physiological artifacts, underscoring their complementary use with endogenous tagging approaches. Although exogenous expression can lead to supraphysiological protein levels, expression can sometimes approximate endogenous levels depending on the protein and system used. Therefore, investigators should empirically assess expression in their experimental context. Beyond standard biochemical and high-throughput methods, cytometry-based and live-cell pulse-chase strategies have been utilized for quantifying ligand-binding domain fusion protein abundance and turnover (Cattoglio et al., 2019; Layer et al., 2020).
Regardless of the tagging strategy, the capacity to produce large libraries of tagged proteins opens the door to systematically deploying small molecule probes against the proteome. By coupling these pooled tag systems with chemical modulators or fluorescent ligands, investigators can map subcellular localization changes, affect protein stability, or induce non-native protein-protein interactions in a high-throughput manner.
3 HaloTag-based pooled tagging enables insights into proteome dynamics
HaloTag, derived from a bacterial haloalkane dehalogenase, is one of the most versatile multifunctional ligand-binding domains (Los et al., 2008; Encell et al., 2012; England et al., 2015). Its covalent reaction with chloroalkane ligands proceeds efficiently in mammalian cells, comparable to binding rates exhibited by biotin-streptavidin, providing fast bio-orthogonal labeling that resists hydrolysis and washout (Encell et al., 2012). Multiple HaloTag variants exist, including the stable HaloTag7 and metastable HaloTag2 (Encell et al., 2012; Tae et al., 2012; Huppertz et al., 2024). This flexibility has led to a broad palette of HaloTag ligands for cellular imaging, protein purification, targeted protein modulation, and more. When combined with pooled gene tagging, HaloTag can be fused to many proteins in parallel, creating libraries of endogenously tagged proteins as demonstrated by Scalable POoled Targeting with a LIgandable Tag at Endogenous Sites (SPOTLITES) (Serebrenik et al., 2024; Sansbury et al., 2025). These cell libraries can then be treated with a toolbox of cell-permeable HaloTag ligands to enable visualization and perturbation of the proteome in a variety of ways (Figure 2) (Hoelzel and Zhang, 2020).
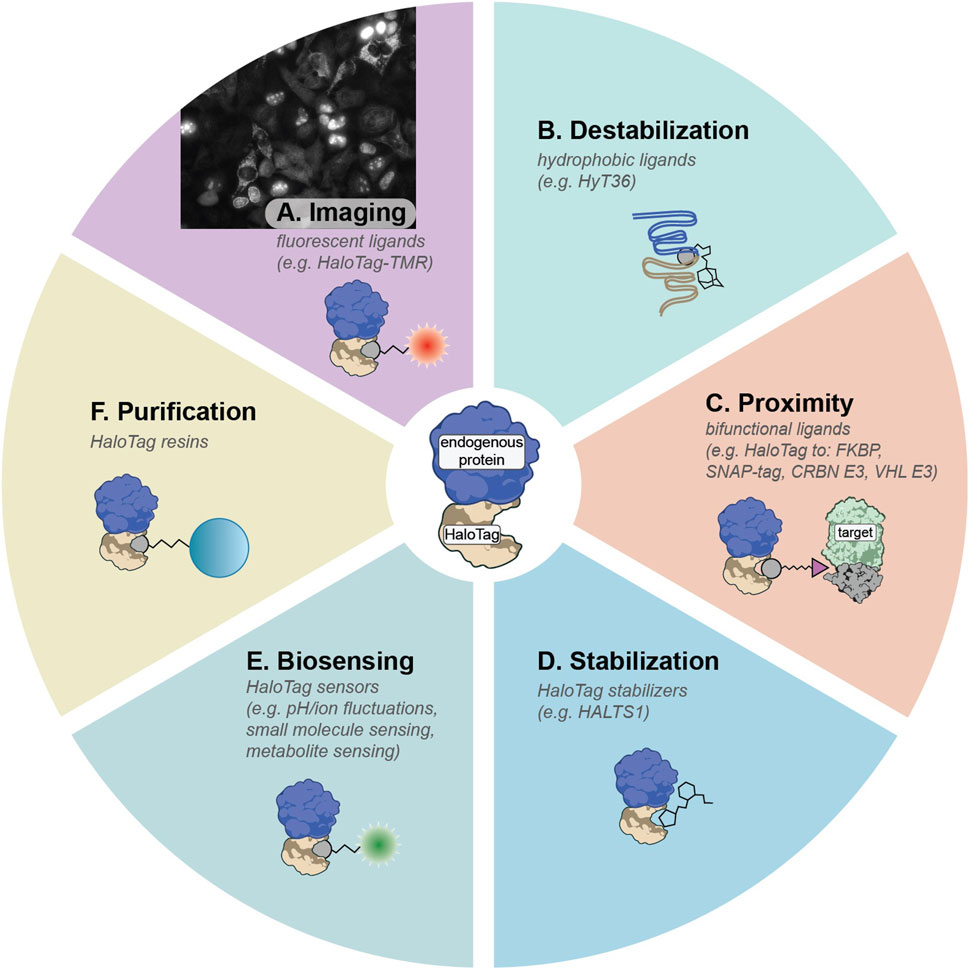
Figure 2. Summary of HaloTag ligands that can be deployed on SPOTLITES libraries. (A) Fluorescent HaloTag ligands like HaloTag-TMR can be used for visualization of protein levels and localization patterns. (B) Hydrophobic tags can induce destabilization of HaloTag2 which can lead to proteotoxic stress responses. (C) Proximity between HaloTag and other ligand-binding domains or other proteins such as E3 ligases can be induced with a suite of bifunctional ligands. (D) The metastable HaloTag2 can be stabilized by treatment with non-covalent binder HALTS1. (E) A large toolbox of biosensors has been developed for HaloTag. (F) Tagged proteins can be easily purified with chloroalkane resins.
3.1 Pooled labeling with fluorescent HaloTag ligands to visualize proteome dynamics
By treating SPOTLITES-derived cell libraries with fluorescent HaloTag ligands, researchers can quickly assess multitudes of protein localization patterns across various cell compartments at single-cell resolution. A range of fluorescent HaloTag ligands has been developed, offering diverse spectral properties and compatibility with different biochemical environments (Figure 2A) (Los et al., 2008; Grimm et al., 2015). Recent work has demonstrated robust visualization of proteins in most organelles and subcellular structures in HAP1, HEK 293, and HeLa cells, and has enabled insight into the endogenous localization patterns of poorly annotated proteins (Sansbury et al., 2025). Beyond proteome imaging at baseline conditions, pooled imaging is well suited for exploring proteome remodeling at the level of expression and localization induced by various stressors or disease models.
3.2 Pooled protein destabilization provides insights into compartment-specific proteostasis mechanisms
Another distinctive advantage of HaloTag arises from its capacity to respond to hydrophobic tagging. When metastable HaloTag2 binds small molecules appended with hydrophobic moieties like adamantane, it mimics partially misfolded proteins, inducing proteotoxic stress and becoming engaged by proximal protein quality control factors (Figure 2B) (Neklesa et al., 2011; Neklesa et al., 2013; Tae et al., 2012). Targeting HaloTag2 to various compartments followed by induced misfolding has illuminated compartment-specific stress responses in the cytosol (Neklesa et al., 2013), ER (Raina et al., 2014), and the Golgi apparatus (Serebrenik et al., 2018; Hellerschmied et al., 2019). This approach has been recently scaled to a pooled format with SPOTLITES, enabling systematic insight into compartment- and even subcompartment-specific proteotoxic stress responses (Sansbury et al., 2025). Treatment of the cell pool with a hydrophobic HaloTag ligand followed by single-cell transcriptomics to identify both the tagged protein and the distinct transcriptional response in each cell provided a map of spatially-restricted proteotoxic stress responses, and revealed proteostasis-related interactions between organelles (Sansbury et al., 2025). By integrating SPOTLITES with genetic models of diseases like proteinopathies, pooled hydrophobic tagging will allow for detailed dissection of proteostasis states during disease progression and identify new opportunities for therapeutic intervention.
3.3 Pooled protein recruitment to profile molecular functions and develop next-generation induced proximity therapies
Pooled HaloTag fusion libraries treated with generic bifunctional ligands can be leveraged to screen induced protein-protein interactions at scale, enabling unique functional genomics studies linking function to proximity (Figure 2C). These screens additionally hold translational significance for chemically induced proximity (CIP) therapies, which harness a small molecule to bridge a “target” protein to an “effector” protein, as exemplified by targeted protein degradation (TPD) where E3 ligases like VHL or CRBN are recruited to degrade neo-substrates (Bondeson et al., 2015; Winter et al., 2015; Burslem and Crews, 2020). Existing CIP technologies rely on just a few known effectors, utilized primarily for TPD; however, recent proteome-scale proximity screens demonstrate that many other effectors can degrade, stabilize, or otherwise modify target proteins, vastly broadening the CIP landscape (Poirson et al., 2024; Serebrenik et al., 2024).
Pooled recruitment screens based on the SPOTLITES platform used HaloTag-fused proteins recruited to a fluorescent target or an essential target to identify previously unrecognized effectors by fluorescence- or cell growth-based enrichment, respectively (Serebrenik et al., 2024). This work converged on known TPD effectors as well as members of the C-terminal-to-LisH (CTLH) complex and other components of the proteostasis machinery as potent degraders. SPOTLITES-based recruitment offers multiple advantages: (1) endogenous effector tagging preserves physiological expression and stoichiometry, (2) internal tagging allows screening of multiple insertion sites that may favor specific orientations between target and effector, and (3) modular ligands facilitate testing different linker designs and chemical warheads for optimal activity. These pooled approaches accommodate diverse readouts such as fluorescence, viability, or complex phenotypic assays (Kahnwald et al., 2024), and importantly, can be applied not only to TPD but also to protein stabilization, disaggregation, and other post-translational modifications.
By systematically interrogating novel effector–target pairs, recruitment screens uncover both protein function and highlight the wealth of potent modulators in the proteome with unique properties such as cell type-specific expression (Jevtić et al., 2021) or high essentiality in disease contexts (Ottis et al., 2019; Zhang et al., 2019; Kurimchak et al., 2022; Hanzl et al., 2023). These datasets will thus provide a starting point for uncovering new biological pathways and targeting a variety of disease mechanisms with selective and resistance-tolerant CIP therapies.
3.4 Other HaloTag ligands and their applications for pooled labeling
An expanding suite of functional HaloTag ligands stands poised for diverse high-throughput applications (Hoelzel and Zhang, 2020). Bifunctional molecules that recruit HaloTag to orthogonal ligand-binding domains such as FKBP, DHFR, or SNAP-tag (see Section 4 for discussion of other ligand-binding domains), can enable multiplexed control of protein-protein interactions, some of which can be triggered or reversed by light-activated chemical groups (Ballister et al., 2014; Zimmermann et al., 2014; Parvez et al., 2016; Zhang et al., 2017). Moreover, targeted protein depletion can be achieved using HaloPROTACs recruiting VHL or CRBN E3 ligases, offering an acute protein depletion alternative to genomic- or transcript-based knockdown methods like CRISPR or RNAi and reducing the confounding effects of genetic adaptation (Buckley et al., 2015; Tovell et al., 2019; Ody et al., 2023). Conversely, ligands such as HALTS1 can stabilize HaloTag2-fused proteins, allowing bidirectional pharmacological control of protein levels (Figure 2D) (Neklesa et al., 2013). Additionally, environment-sensitive fluorophores permit real-time readouts of metabolites, ion concentrations, and protein folding states in living cells (Figure 2E) (Liu et al., 2018; Wang et al., 2020). Ligands fused to resins can be used to capture HaloTag-fused proteins of interest (Figure 2F). By combining these ligands with pooled HaloTag libraries and sequencing-based readouts, researchers gain the ability to probe myriad aspects of protein function and regulation systematically, from rapid degradation studies to detailed investigations of local chemical microenvironments.
3.5 Limitations for pooled ligand-binding assays
Despite the flexibility offered by HaloTag and other ligand-binding domains, several practical and conceptual limitations must be considered. First, ligands may have inconsistent labeling efficiencies when faced with targets expressed at varying levels and cellular environments. Furthermore, downstream effects of labeling can vary: in hydrophobic tagging applications, metastable fusion proteins may completely misfold, whereas more stable fusions may be less acutely affected. Bifunctional ligands designed for induced proximity can also miss certain protein-protein interactions if linker length and flexibility are not properly tuned. Additional concerns, like off-target binding and other ligand-specific considerations such as membrane permeability must be accounted for with different tag systems (see Section 4). Second, the gene tagging process itself can disrupt protein stability or function, particularly with larger tags, though this can be alleviated by testing multiple insertion sites per protein as part of the pooled tagging approach. In diploid cells, heterozygous integration is likely to occur depending on the tagging method used, which may complicate methods such as pooled degradation screens that would rely on perturbation of the entirety of a given protein. Finally, no single tagging platform suits all genes: intron-based methods fail when genes lack introns, while certain proteins cannot tolerate terminal tagging. While these issues do not negate the advantages of pooled ligand-binding assays, they highlight the importance of careful experimental design and follow-up validation.
4 Alternative domains and probes
A widely used strategy for visualizing and targeting specific proteins of interest (POIs) involves fusions with fluorescent or split fluorescent proteins (Feng et al., 2017; Romei and Boxer, 2019), affinity tags (Kimple et al., 2013), nanobodies (de Beer and Giepmans, 2020; Stevens et al., 2024), and degron tags (Natsume and Kanemaki, 2017). Recently, the use of ligand-binding domains has emerged as a “swiss army knife” approach for manipulating POIs (Table 1). By switching among different probes, one can readily repurpose a single domain for numerous applications such as fluorescent labeling, protein-protein interaction studies, stabilization or destabilization, targeted degradation, quantitative proteomics, purification, drug screening, biosensing, and scaffolding. Several alternatives orthogonal to the widely-used HaloTag have been developed over the years, each offering distinct advantages and drawbacks depending on the experimental context.
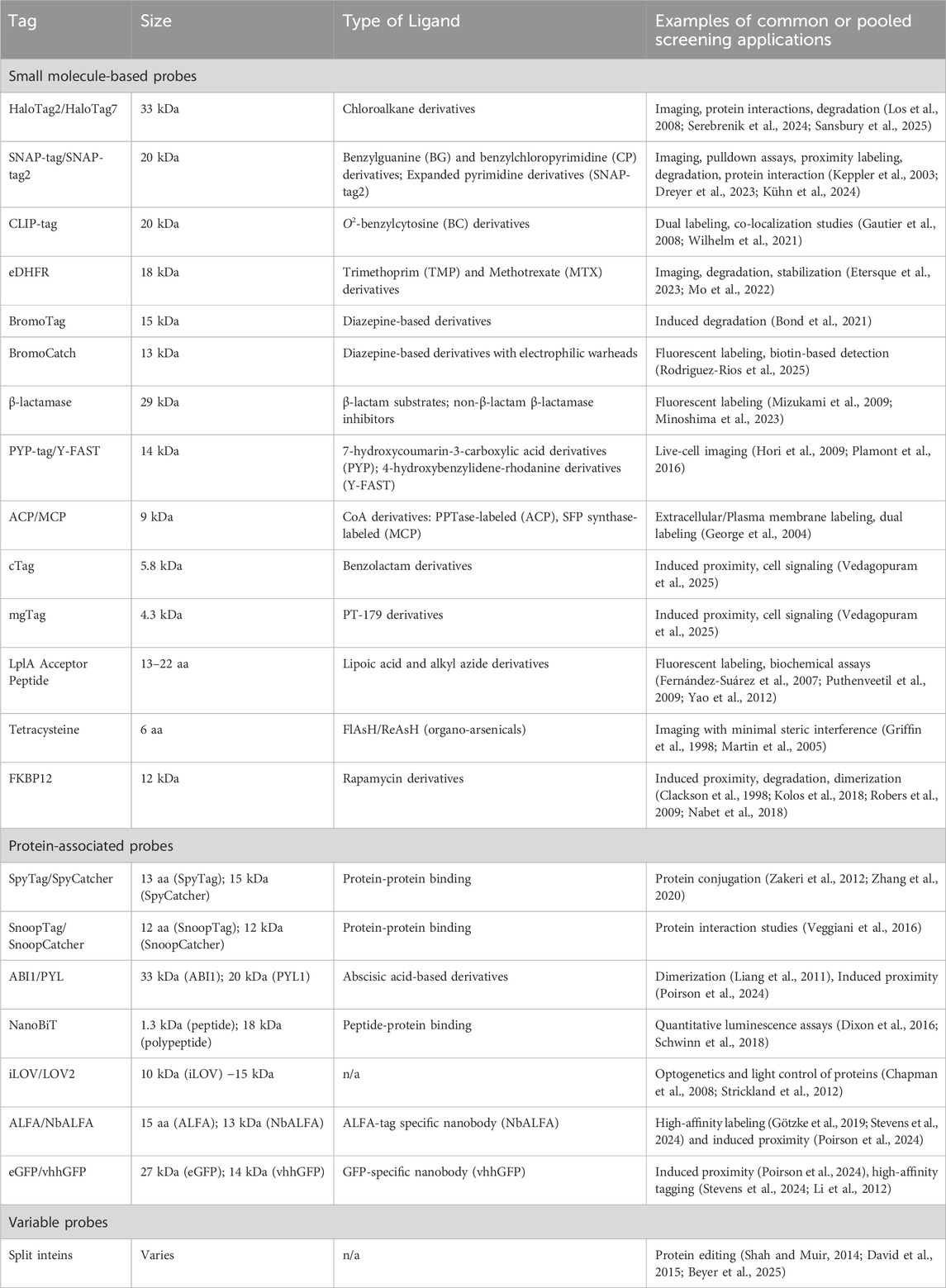
Table 1. Summary of common protein tags amenable to pooled knock-in.
4.1 Small molecule–based probes
Domains for binding probes can be broadly categorized into self-labeling domains that directly form covalent bonds with their ligands, enzyme-mediated systems in which exogenous enzymes catalyze the bond formation, and non-covalent domains that rely on high-affinity binding rather than covalent linkage. Self-labeling protein (SLP) tags provide simple schemes that require only the tag and its cognate ligand. SNAP-tag is an SLP derived from human O6-alkylguanine-DNA alkyltransferase (AGT) and binds covalently to O6-benzyl -guanine and -chloropyrimidine derivatives (Keppler et al., 2003; Corrêa et al., 2013). SNAP-tag is smaller (20 kDa) than HaloTag (33 kDa), making it less likely to interfere with the tagged protein. However, the SNAP-tag has slower labeling kinetics and some SNAP-tag ligands suffer from limited cell permeability, restricting its use in live-cell imaging (Dreyer et al., 2023). The engineered SNAP-tag2 improves labeling speed and fluorescence brightness and can be used in conjugation with pyrimidine-based substrates optimized for improved kinetics and cell permeability (Kühn et al., 2024). SNAP-tag has been used in many applications from targeted degradation to induction of protein-protein interactions (Kindermann et al., 2003; Bottanelli et al., 2016; Yu et al., 2019; McCutcheon et al., 2020; Dotson and Ngo, 2022; Pol et al., 2024; Seabrook et al., 2024). The CLIP-tag, also derived from AGT (Gautier et al., 2008) binds O2-benzylcytosine derivatives, enabling orthogonal labeling alongside SNAP-tag for co-localization studies (Wilhelm et al., 2021). Comparable in size to SNAP-tag, the 18 kDa Escherichia coli dihydrofolate reductase (eDHFR) binds trimethoprim (TMP)- and methotrexate-based ligands to allow bioorthogonal labeling in mammalian cells for applications from fluorescent imaging to degradation (Baker et al., 1981; Etersque et al., 2023; Rana et al., 2024). Further improvements to eDHFR TMP now allow for fast, covalent labeling (Chen et al., 2012; Mo et al., 2022). The BromoTag system is a 15 kDa inducible degron platform that employs a BRD4 bromodomain L387A variant tag to enable rapid target degradation via heterobifunctional bumped PROTACs (Bond et al., 2021). Complementing this, the 13 kDa SLP-based BromoCatch system utilizes a bromodomain-derived tag compatible with functionalized diazepine probes with an electrophilic group being developed for a wide range of applications, including protein-protein interaction assays, purification, targeted degradation, and live-cell fluorescence imaging (Rodriguez-Rios et al., 2025).
Additional self-labeling tags include the 29 kDa TEM-1 β-lactamase inhibitor-based tags (Mizukami et al., 2009; Minoshima et al., 2023), as well as the 14 kDa photoactive yellow protein PYP-tag (Hori et al., 2009) and its reversible derivative Y-FAST (Plamont et al., 2016). Among the smaller tags that covalently bind small molecules, ACP (Acyl Carrier Protein) and MCP (mutated ACP D36T/D39G), 9 and 8 kDa respectively, allow selective dual labeling on the cell surface using CoA derivatives with respective phosphopantetheinyl transferases (George et al., 2004). Two compact covalent tags, cTag (5.8 kDa) and mgTag (4.3 kDa), mediate group-transfer reactions with their respective ligands (Vedagopuram et al., 2025). cTag employs an engineered C1 domain containing a reactive cysteine residue, while mgTag features an engineered zinc-finger domain with a cysteine for reactivity. cTag specifically binds ligands bearing cysteine-reactive benzolactam scaffolds, whereas mgTag engages immunomodulatory drug (IMiD) derivatives, such as the molecular glue PT-179 (Mercer et al., 2024) modified with a group-transfer linker, in a cereblon-dependent manner.
A variety of short peptide tags exist that rely on exogenous delivery of an enzyme to catalyze covalent bond formation between the tag and its ligand; many of these tags are suitable only for cell surface labeling and have been reviewed elsewhere (Wolf et al., 2021). The Escherichia coli Lipoic Acid Ligase (LplA) Acceptor Peptides, as small as 13 amino acids, are notable for enabling specific intracellular protein tagging with a variety of probes (Fernández-Suárez et al., 2007; Puthenveetil et al., 2009; Yao et al., 2012); although the requirement of LplA to physically interact with its substrates may limit the compartments amenable for pooled protein tagging. In addition, a minimal six-amino acid tetracysteine motif binds FlAsH/ReAsH dyes for fluorescence imaging (Griffin et al., 1998) but its specificity is limited (Stroffekova et al., 2001). Optimized tetracysteine sequences with improved ligand affinity and fluorescence have since been developed (Martin et al., 2005).
Non-covalent small molecule interactions are exemplified by mutant forms of human FKBP12 (e.g., FKBPF36V, 12 kDa), which bind rapamycin-derived probes (Choi et al., 1996; Clackson et al., 1998). This system is routinely used for chemically induced proximity, targeted protein degradation, intracellular imaging, and more (Kohler and Bertozzi, 2003; Inoue et al., 2005; Banaszynski et al., 2006a; Banaszynski et al., 2006b; Robers et al., 2009; Kolos et al., 2018; Nabet et al., 2018; Du et al., 2023).
4.2 Protein-associated probes
Many fusion tags utilize peptide-based ligands or specifically bind other proteins which can be genetically encoded yet lack the chemical flexibility and minimal size of small molecule ligands. Nevertheless, these systems can still be functionalized with a variety of probes and delivered extracellularly. SpyTag/SpyCatcher, derived from the Streptococcus pyogenes CnaB2 domain of a fibronectin-binding protein, FbaB (Zakeri et al., 2012; Zhang et al., 2020) and SnoopTag/SnoopCatcher, derived from a Streptococcus pneumoniae adhesin, RrgA (Veggiani et al., 2016) form spontaneous isopeptide bonds, creating robust covalent linkages used for protein conjugation with options for chemical diversification. Likewise, ABA-based PYR/PYL domains (Liang et al., 2011) enable ligand-dependent dimerization for induced proximity or degradation (Poirson et al., 2024; Han et al., 2025). NanoBiT (Dixon et al., 2016) and its variants (Schwinn et al., 2018) reconstitutes a split-luciferase signal, yielding sensitive, quantitative assays of protein interactions. For optogenetics, small flavin-binding domains iLOV/LOV2 (Chapman et al., 2008; Strickland et al., 2012) regulate protein function via light but demand specialized setups.
Nanobody-binding domains represent another large category of tags, with prominent examples including ALFA/NbALFA (Götzke et al., 2019) and eGFP/vhhGFP (Li et al., 2012) which exploit short peptide epitopes recognized by high-affinity nanobodies. These systems have been used for many applications from imaging to inducing protein-protein interactions, even in pooled systems (de Beer and Giepmans, 2020; Poirson et al., 2024; Stevens et al., 2024). Finally, split intein-based tags allow for specific and flexible protein editing through a covalent trans splicing reaction (Shah and Muir, 2014). Proteins can be tagged with split inteins terminally (David et al., 2015) or internally by using two orthogonal split intein pairs (Beyer et al., 2025). After splicing, inteins leave behind a minimal footprint (≤10 amino acids) defined by the intein-specific “extein” sequence along with a peptide tag of choice that can include non-canonical amino acids for compatibility with a wide variety of probes (Yi et al., 2024; Beyer et al., 2025).
4.3 Ideal next-generation domains and probes
Although existing ligand-binding domains offer an impressive array of functionalities, no single system perfectly satisfies all research needs. An ideal “next-generation” fusion domain for large-scale tagging approaches would have the following attributes: 1) orthogonal to endogenous cellular machinery, preventing unintended labeling, interactions, or interference; 2) monomeric with a minimal size and footprint to help preserve the proper fold, function, and binding interfaces of the fused protein; 3) high tag stability would further minimize perturbation of the structure of the protein it is fused to; 4) the tag would have a high-quality monoclonal antibody for orthogonal detection and biochemical isolation. On the ligand side, 5) a chemically versatile scaffold with rapid, high-affinity, and specific binding to ensure quantitative labeling; 6) such ligands should be easily customized for imaging, degradation, recruitment, or other modifications, and 7) would not require additional enzymes or components for binding. Such a tag system would be highly effective for proteome-scale pooled tag libraries.
5 Conclusions and perspectives
Pooled protein tagging with multifunctional ligand-binding domains has enabled new approaches for cell biology that combine chemical biology and functional genomics for high-throughput protein interrogation at both endogenous and exogenous expression levels. By coupling pooled tag libraries with specialized ligands ranging from fluorescent probes to bifunctional molecules, researchers can systematically investigate protein localization, stability, interactions, and more in diverse cell models. This convergence of technologies offers unique insights into proteostasis and disease mechanisms, laying the groundwork for next-generation therapeutic strategies, including those utilizing induced proximity.
Looking ahead, there remains ample opportunity to expand existing pooled tagging approaches into new disease contexts, such as cancer and neurodegeneration, to gain unique insights on the relevant protein dynamics, interactions, or responses to protein misfolding. Furthermore, exploring so-far untested domains and probes in a proteome-wide context will enable additional systematic insights into cellular environments. Finally, continued large-scale tagging efforts will iteratively pave the way for comprehensive proteome-wide libraries of high-fidelity fusions which can be utilized in subsequent tagging experiments. Incorporating multiple tag fusion variations for each protein will be especially valuable for protein–protein interaction studies, where proper alignment can make the difference between a fleeting contact and a functionally relevant complex. In tandem with the ongoing development of new domains and ligands, these integrative strategies promise to push the frontier of systematic studies of the proteome, bridging functional genomics and chemical biology and opening up exciting avenues for both fundamental discovery and translational research.
Author contributions
KL: Writing – review and editing, Writing – original draft. YS: Writing – review and editing, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by R00AG075256 from NIH/NIA, awarded to Y.V.S.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alerasool, N., Leng, H., Lin, Z.-Y., Gingras, A.-C., and Taipale, M. (2022). Identification and functional characterization of transcriptional activators in human cells. Mol. Cell 82, 677–695.e7. doi:10.1016/j.molcel.2021.12.008
Anzalone, A. V., Gao, X. D., Podracky, C. J., Nelson, A. T., Koblan, L. W., Raguram, A., et al. (2022). Programmable deletion, replacement, integration and inversion of large DNA sequences with twin prime editing. Nat. Biotechnol. 40, 731–740. doi:10.1038/s41587-021-01133-w
Baker, D. J., Beddell, C. R., Champness, J. N., Goodford, P. J., Norrington, F. E., Smith, D. R., et al. (1981). The binding of trimethoprim to bacterial dihydrofolate reductase. FEBS Lett. 126, 49–52. doi:10.1016/0014-5793(81)81030-7
Ballister, E. R., Aonbangkhen, C., Mayo, A. M., Lampson, M. A., and Chenoweth, D. M. (2014). Localized light-induced protein dimerization in living cells using a photocaged dimerizer. Nat. Commun. 5, 5475. doi:10.1038/ncomms6475
Banaszynski, L. A., Chen, L.-C., Maynard-Smith, L. A., Ooi, A. G. L., and Wandless, T. J. (2006a). A rapid, reversible, and tunable method to regulate protein function in living cells using synthetic small molecules. Cell 126, 995–1004. doi:10.1016/j.cell.2006.07.025
Banaszynski, L. A., Liu, C. W., and Wandless, T. J. (2006b). Characterization of the FKBP·rapamycin·FRB ternary complex [J. am. Chem. Soc. 2005, 127, 4715−4721]. J. Am. Chem. Soc. 128, 15928. doi:10.1021/ja0699788
Beyer, J. N., Serebrenik, Y. V., Toy, K., Najar, M. A., Feierman, E., Raniszewski, N. R., et al. (2025). Intracellular protein editing enables incorporation of noncanonical residues in endogenous proteins. Science 388, eadr5499. doi:10.1126/science.adr5499
Bond, A. G., Craigon, C., Chan, K.-H., Testa, A., Karapetsas, A., Fasimoye, R., et al. (2021). Development of BromoTag: A “bump-and-hole”-PROTAC system to induce potent, rapid, and selective degradation of tagged target proteins. J. Med. Chem. 64, 15477–15502.
Bondeson, D. P., Mares, A., Smith, I. E. D., Ko, E., Campos, S., Miah, A. H., et al. (2015). Catalytic in vivo protein knockdown by small-molecule PROTACs. Nat. Chem. Biol. 11, 611–617. doi:10.1038/nchembio.1858
Bottanelli, F., Kromann, E. B., Allgeyer, E. S., Erdmann, R. S., Wood Baguley, S., Sirinakis, G., et al. (2016). Two-colour live-cell nanoscale imaging of intracellular targets. Nat. Commun. 7, 10778. doi:10.1038/ncomms10778
Buckley, D. L., Raina, K., Darricarrere, N., Hines, J., Gustafson, J. L., Smith, I. E., et al. (2015). HaloPROTACS: use of small molecule PROTACs to induce degradation of HaloTag fusion proteins. ACS Chem. Biol. 10, 1831–1837. doi:10.1021/acschembio.5b00442
Burslem, G. M., and Crews, C. M. (2020). Proteolysis-targeting chimeras as therapeutics and tools for biological discovery. Cell 181, 102–114. doi:10.1016/j.cell.2019.11.031
Cattoglio, C., Pustova, I., Walther, N., Ho, J. J., Hantsche-Grininger, M., Inouye, C. J., et al. (2019). Determining cellular CTCF and cohesin abundances to constrain 3D genome models. Elife 8, e40164.
Chapman, S., Faulkner, C., Kaiserli, E., Garcia-Mata, C., Savenkov, E. I., Roberts, A. G., et al. (2008). The photoreversible fluorescent protein iLOV outperforms GFP as a reporter of plant virus infection. Proc. Natl. Acad. Sci. U. S. A. 105, 20038–20043. doi:10.1073/pnas.0807551105
Chen, Z., Jing, C., Gallagher, S. S., Sheetz, M. P., and Cornish, V. W. (2012). Second-generation covalent TMP-tag for live cell imaging. J. Am. Chem. Soc. 134, 13692–13699. doi:10.1021/ja303374p
Cho, N. H., Cheveralls, K. C., Brunner, A.-D., Kim, K., Michaelis, A. C., Raghavan, P., et al. (2022). OpenCell: endogenous tagging for the cartography of human cellular organization. Science 375, eabi6983. doi:10.1126/science.abi6983
Choi, J., Chen, J., Schreiber, S. L., and Clardy, J. (1996). Structure of the FKBP12-rapamycin complex interacting with the binding domain of human FRAP. Science 273, 239–242. doi:10.1126/science.273.5272.239
Clackson, T., Yang, W., Rozamus, L. W., Hatada, M., Amara, J. F., Rollins, C. T., et al. (1998). Redesigning an FKBP-ligand interface to generate chemical dimerizers with novel specificity. Proc. Natl. Acad. Sci. U. S. A. 95, 10437–10442. doi:10.1073/pnas.95.18.10437
Cohen, A. A., Geva-Zatorsky, N., Eden, E., Frenkel-Morgenstern, M., Issaeva, I., Sigal, A., et al. (2008). Dynamic proteomics of individual cancer cells in response to a drug. Science 322, 1511–1516. doi:10.1126/science.1160165
Corrêa, I. R., Baker, B., Zhang, A., Sun, L., Provost, C. R., Lukinavičius, G., et al. (2013). Substrates for improved live-cell fluorescence labeling of SNAP-tag. Curr. Pharm. Des. 19, 5414–5420. doi:10.2174/1381612811319300011
David, Y., Vila-Perelló, M., Verma, S., and Muir, T. W. (2015). Chemical tagging and customizing of cellular chromatin states using ultrafast trans-splicing inteins. Nat. Chem. 7, 394–402. doi:10.1038/nchem.2224
de Beer, M. A., and Giepmans, B. N. G. (2020). Nanobody-based probes for subcellular protein identification and visualization. Front. Cell. Neurosci. 14, 573278. doi:10.3389/fncel.2020.573278
DelRosso, N., Tycko, J., Suzuki, P., Andrews, C., Mukund, A., Liongson, I., et al. (2023). Large-scale mapping and mutagenesis of human transcriptional effector domains. Nature 616, 365–372. doi:10.1038/s41586-023-05906-y
Dixon, A. S., Schwinn, M. K., Hall, M. P., Zimmerman, K., Otto, P., Lubben, T. H., et al. (2016). NanoLuc complementation reporter optimized for accurate measurement of protein interactions in cells. ACS Chem. Biol. 11, 400–408. doi:10.1021/acschembio.5b00753
Dotson, H. L., and Ngo, J. T. (2022). SNAP-tag and HaloTag fused proteins for HaSX8-inducible control over synthetic biological functions in engineered mammalian cells. bioRxiv. doi:10.1101/2022.08.12.503781
Dreyer, R., Pfukwa, R., Barth, S., Hunter, R., and Klumperman, B. (2023). The evolution of SNAP-tag labels. Biomacromolecules 24, 517–530. doi:10.1021/acs.biomac.2c01238
Du, Z., Wang, W., Luo, S., Zhang, L., Yuan, S., Hei, Y., et al. (2023). Self-renewable tag for photostable fluorescence imaging of proteins. J. Am. Chem. Soc. 145, 18968–18976. doi:10.1021/jacs.3c06102
Encell, L. P., Friedman Ohana, R., Zimmerman, K., Otto, P., Vidugiris, G., Wood, M. G., et al. (2012). Development of a dehalogenase-based protein fusion tag capable of rapid, selective and covalent attachment to customizable ligands. Curr. Chem. Genomics 6, 55–71. doi:10.2174/1875397301206010055
England, C. G., Luo, H., and Cai, W. (2015). HaloTag technology: a versatile platform for biomedical applications. Bioconjug. Chem. 26, 975–986. doi:10.1021/acs.bioconjchem.5b00191
Etersque, J. M., Lee, I. K., Sharma, N., Xu, K., Ruff, A., Northrup, J. D., et al. (2023). Regulation of eDHFR-tagged proteins with trimethoprim PROTACs. Nat. Commun. 14, 7071. doi:10.1038/s41467-023-42820-3
Feldman, D., Singh, A., Schmid-Burgk, J. L., Carlson, R. J., Mezger, A., Garrity, A. J., et al. (2019). Optical pooled screens in human cells. Cell 179, 787–799.e17. doi:10.1016/j.cell.2019.09.016
Feng, S., Sekine, S., Pessino, V., Li, H., Leonetti, M. D., and Huang, B. (2017). Improved split fluorescent proteins for endogenous protein labeling. Nat. Commun. 8, 370. doi:10.1038/s41467-017-00494-8
Fernández-Suárez, M., Baruah, H., Martínez-Hernández, L., Xie, K. T., Baskin, J. M., Bertozzi, C. R., et al. (2007). Redirecting lipoic acid ligase for cell surface protein labeling with small-molecule probes. Nat. Biotechnol. 25, 1483–1487. doi:10.1038/nbt1355
Gautier, A., Juillerat, A., Heinis, C., Corrêa, I. R., Kindermann, M., Beaufils, F., et al. (2008). An engineered protein tag for multiprotein labeling in living cells. Chem. Biol. 15, 128–136. doi:10.1016/j.chembiol.2008.01.007
George, N., Pick, H., Vogel, H., Johnsson, N., and Johnsson, K. (2004). Specific labeling of cell surface proteins with chemically diverse compounds. J. Am. Chem. Soc. 126, 8896–8897. doi:10.1021/ja048396s
Götzke, H., Kilisch, M., Martínez-Carranza, M., Sograte-Idrissi, S., Rajavel, A., Schlichthaerle, T., et al. (2019). The ALFA-tag is a highly versatile tool for nanobody-based bioscience applications. Nat. Commun. 10, 4403. doi:10.1038/s41467-019-12301-7
Griffin, B. A., Adams, S. R., and Tsien, R. Y. (1998). Specific covalent labeling of recombinant protein molecules inside live cells. Science 281, 269–272. doi:10.1126/science.281.5374.269
Grimm, J. B., English, B. P., Chen, J., Slaughter, J. P., Zhang, Z., Revyakin, A., et al. (2015). A general method to improve fluorophores for live-cell and single-molecule microscopy. Nat. Methods 12 (244–50), 244–250. doi:10.1038/nmeth.3256
Han, M., Fu, M. L., Zhu, Y., Choi, A. A., Li, E., Bezney, J., et al. (2025). Programmable control of spatial transcriptome in live cells and neurons. Nature, 1–11. doi:10.1038/s41586-025-09020-z
Hanzl, A., Casement, R., Imrichova, H., Hughes, S. J., Barone, E., Testa, A., et al. (2023). Functional E3 ligase hotspots and resistance mechanisms to small-molecule degraders. Nat. Chem. Biol. 19, 323–333. doi:10.1038/s41589-022-01177-2
Hellerschmied, D., Serebrenik, Y. V., Shao, L., Burslem, G. M., and Crews, C. M. (2019). Protein folding state-dependent sorting at the Golgi apparatus. Mol. Biol. Cell 30, 2296–2308. doi:10.1091/mbc.E19-01-0069
Hoelzel, C. A., and Zhang, X. (2020). Visualizing and manipulating biological processes by using HaloTag and SNAP-tag technologies. Chembiochem 21, 1935–1946. doi:10.1002/cbic.202000037
Hori, Y., Ueno, H., Mizukami, S., and Kikuchi, K. (2009). Photoactive yellow protein-based protein labeling system with turn-on fluorescence intensity. J. Am. Chem. Soc. 131, 16610–16611. doi:10.1021/ja904800k
Huppertz, M. C., Wilhelm, J., Grenier, V., Schneider, M. W., Falt, T., Porzberg, N., et al. (2024). Recording physiological history of cells with chemical labeling. Science 383 (6685), 890–897. doi:10.1126/science.adg0812
Inoue, T., Heo, W. D., Grimley, J. S., Wandless, T. J., and Meyer, T. (2005). An inducible translocation strategy to rapidly activate and inhibit small GTPase signaling pathways. Nat. Methods 2, 415–418. doi:10.1038/nmeth763
Jarvik, J. W., Adler, S. A., Telmer, C. A., Subramaniam, V., and Lopez, A. J. (1996). CD-tagging: a new approach to gene and protein discovery and analysis. Biotechniques 20, 896–904. doi:10.2144/96205rr03
Jarvik, J. W., Fisher, G. W., Shi, C., Hennen, L., Hauser, C., Adler, S., et al. (2002). In vivo functional proteomics: mammalian genome annotation using CD-tagging. Biotechniques 33 (852–4), 852–856. doi:10.2144/02334rr02
Jevtić, P., Haakonsen, D. L., and Rapé, M. (2021). An E3 ligase guide to the galaxy of small-molecule-induced protein degradation. Cell Chem. Biol. 28, 1000–1013. doi:10.1016/j.chembiol.2021.04.002
Kahnwald, M., Mählen, M., Oost, K. C., and Liberali, P. (2024). Advances in optical pooled screening to map spatial complexity. Nat. Biotechnol., 1–3. doi:10.1038/s41587-024-02434-6
Keppler, A., Gendreizig, S., Gronemeyer, T., Pick, H., Vogel, H., and Johnsson, K. (2003). A general method for the covalent labeling of fusion proteins with small molecules in vivo. Nat. Biotechnol. 21, 86–89. doi:10.1038/nbt765
Kim, J., Kratz, A. F., Chen, S., Sheng, J., Kim, H. K., Zhang, L., et al. (2024). High-throughput tagging of endogenous loci for rapid characterization of protein function. Sci. Adv. 10, eadg8771. doi:10.1126/sciadv.adg8771
Kimple, M. E., Brill, A. L., and Pasker, R. L. (2013). Overview of affinity tags for protein purification. Curr. Protoc. Protein Sci. 73, 9.9.1–9.9.23. doi:10.1002/0471140864.ps0909s73
Kindermann, M., George, N., Johnsson, N., and Johnsson, K. (2003). Covalent and selective immobilization of fusion proteins. J. Am. Chem. Soc. 125, 7810–7811. doi:10.1021/ja034145s
Kohler, J. J., and Bertozzi, C. R. (2003). Regulating cell surface glycosylation by small molecule control of enzyme localization. Chem. Biol. 10, 1303–1311. doi:10.1016/j.chembiol.2003.11.018
Kolos, J. M., Voll, A. M., Bauder, M., and Hausch, F. (2018). FKBP ligands-where we are and where to go? Front. Pharmacol. 9, 1425. doi:10.3389/fphar.2018.01425
Kühn, S., Nasufovic, V., Wilhelm, J., Kompa, J., de Lange, E. M. F., Lin, Y.-H., et al. (2024). SNAP-tag2: faster and brighter protein labeling. bioRxiv. doi:10.1101/2024.08.28.610127
Kurimchak, A. M., Herrera-Montávez, C., Montserrat-Sangrà, S., Araiza-Olivera, D., Hu, J., Neumann-Domer, R., et al. (2022). The drug efflux pump MDR1 promotes intrinsic and acquired resistance to PROTACs in cancer cells. Sci. Signal. 15, eabn2707. doi:10.1126/scisignal.abn2707
Lackner, D. H., Carré, A., Guzzardo, P. M., Banning, C., Mangena, R., Henley, T., et al. (2015). A generic strategy for CRISPR-Cas9-mediated gene tagging. Nat. Commun. 6, 10237. doi:10.1038/ncomms10237
Lacoste, J., Haghighi, M., Haider, S., Reno, C., Lin, Z.-Y., Segal, D., et al. (2024). Pervasive mislocalization of pathogenic coding variants underlying human disorders. Cell 187, 6725–6741.e13. doi:10.1016/j.cell.2024.09.003
Layer, J. H., Christy, M., Placek, L., Unutmaz, D., Guo, Y., Davé, U. P., et al. (2020). LDB1 enforces stability on direct and indirect oncoprotein partners in leukemia. Mol. Cell. Biol. 40, 1–28.
Li, T., Bourgeois, J.-P., Celli, S., Glacial, F., Le Sourd, A.-M., Mecheri, S., et al. (2012). Cell-penetrating anti-GFAP VHH and corresponding fluorescent fusion protein VHH-GFP spontaneously cross the blood-brain barrier and specifically recognize astrocytes: application to brain imaging. FASEB J. 26, 3969–3979. doi:10.1096/fj.11-201384
Liang, F.-S., Ho, W. Q., and Crabtree, G. R. (2011). Engineering the ABA plant stress pathway for regulation of induced proximity. Sci. Signal. 4, rs2. doi:10.1126/scisignal.2001449
Liu, Y., Miao, K., Li, Y., Fares, M., Chen, S., and Zhang, X. (2018). A HaloTag-based multicolor fluorogenic sensor visualizes and quantifies proteome stress in live cells using solvatochromic and molecular rotor-based fluorophores. Biochemistry 57, 4663–4674. doi:10.1021/acs.biochem.8b00135
Los, G. V., Encell, L. P., McDougall, M. G., Hartzell, D. D., Karassina, N., Zimprich, C., et al. (2008). HaloTag: a novel protein labeling technology for cell imaging and protein analysis. ACS Chem. Biol. 3, 373–382. doi:10.1021/cb800025k
Martin, B. R., Giepmans, B. N. G., Adams, S. R., and Tsien, R. Y. (2005). Mammalian cell-based optimization of the biarsenical-binding tetracysteine motif for improved fluorescence and affinity. Nat. Biotechnol. 23, 1308–1314. doi:10.1038/nbt1136
McCutcheon, D. C., Lee, G., Carlos, A., Montgomery, J. E., and Moellering, R. E. (2020). Photoproximity profiling of protein-protein interactions in cells. J. Am. Chem. Soc. 142, 146–153. doi:10.1021/jacs.9b06528
Mercer, J. A. M., DeCarlo, S. J., Roy Burman, S. S., Sreekanth, V., Nelson, A. T., Hunkeler, M., et al. (2024). Continuous evolution of compact protein degradation tags regulated by selective molecular glues. Science 383, eadk4422. doi:10.1126/science.adk4422
Minoshima, M., Umeno, T., Kadooka, K., Roux, M., Yamada, N., and Kikuchi, K. (2023). Development of a versatile protein labeling tool for live-cell imaging using fluorescent ß-lactamase inhibitors. Angew. Chem. Int. Ed. Engl. 62, e202301704. doi:10.1002/anie.202301704
Mizukami, S., Watanabe, S., Hori, Y., and Kikuchi, K. (2009). Covalent protein labeling based on noncatalytic beta-lactamase and a designed FRET substrate. J. Am. Chem. Soc. 131, 5016–5017. doi:10.1021/ja8082285
Mo, J., Chen, J., Shi, Y., Sun, J., Wu, Y., Liu, T., et al. (2022). Third-generation covalent TMP-tag for fast labeling and multiplexed imaging of cellular proteins. Angew. Chem. Int. Ed. Engl. 61, e202207905. doi:10.1002/anie.202207905
Moon, H. C., Herschl, M. H., Pawluk, A., Konermann, S., and Hsu, P. D. (2024). A combinatorial domain screening platform reveals epigenetic effector interactions for transcriptional perturbation. bioRxiv. doi:10.1101/2024.10.28.620683
Nabet, B., Roberts, J. M., Buckley, D. L., Paulk, J., Dastjerdi, S., Yang, A., et al. (2018). The dTAG system for immediate and target-specific protein degradation. Nat. Chem. Biol. 14, 431–441. doi:10.1038/s41589-018-0021-8
Natsume, T., and Kanemaki, M. T. (2017). Conditional degrons for controlling protein expression at the protein level. Annu. Rev. Genet. 51, 83–102.
Neklesa, T. K., Noblin, D. J., Kuzin, A. P., Lew, S., Seetharaman, J., Acton, T. B., et al. (2013). A bidirectional system for the dynamic small molecule control of intracellular fusion proteins. ACS Chem. Biol. 8, 2293–2300. doi:10.1021/cb400569k
Neklesa, T. K., Tae, H. S., Schneekloth, A. R., Stulberg, M. J., Corson, T. W., Sundberg, T. B., et al. (2011). Small-molecule hydrophobic tagging-induced degradation of HaloTag fusion proteins. Nat. Chem. Biol. 7, 538–543. doi:10.1038/nchembio.597
Nemčko, F., Himmelsbach, M., Loubiere, V., Yelagandula, R., Pagani, M., Fasching, N., et al. (2024). Proteome-scale tagging and functional screening in mammalian cells by ORFtag. Nat. Methods 21, 1668–1673. doi:10.1038/s41592-024-02339-x
Ody, B. K., Zhang, J., Nelson, S. E., Xie, Y., Liu, R., Dodd, C. J., et al. (2023). Synthesis and evaluation of cereblon-recruiting HaloPROTACs. Chembiochem 24, e202300498. doi:10.1002/cbic.202300498
ORFeome Collaboration (2016). The ORFeome Collaboration: a genome-scale human ORF-clone resource. Nat. Methods 13, 191–192. doi:10.1038/nmeth.3776
Ottis, P., Palladino, C., Thienger, P., Britschgi, A., Heichinger, C., Berrera, M., et al. (2019). Cellular resistance mechanisms to targeted protein degradation converge toward impairment of the engaged ubiquitin transfer pathway. ACS Chem. Biol. 14, 2215–2223. doi:10.1021/acschembio.9b00525
Pandey, S., Gao, X. D., Krasnow, N. A., McElroy, A., Tao, Y. A., Duby, J. E., et al. (2025). Efficient site-specific integration of large genes in mammalian cells via continuously evolved recombinases and prime editing. Nat. Biomed. Eng. 9, 22–39. doi:10.1038/s41551-024-01227-1
Parvez, S., Long, M. J. C., Lin, H.-Y., Zhao, Y., Haegele, J. A., Pham, V. N., et al. (2016). T-REX on-demand redox targeting in live cells. Nat. Protoc. 11, 2328–2356. doi:10.1038/nprot.2016.114
Plamont, M.-A., Billon-Denis, E., Maurin, S., Gauron, C., Pimenta, F. M., Specht, C. G., et al. (2016). Small fluorescence-activating and absorption-shifting tag for tunable protein imaging in vivo. Proc. Natl. Acad. Sci. U. S. A. 113, 497–502. doi:10.1073/pnas.1513094113
Poirson, J., Cho, H., Dhillon, A., Haider, S., Imrit, A. Z., Lam, M. H. Y., et al. (2024). Proteome-scale discovery of protein degradation and stabilization effectors. Nature 628, 878–886. doi:10.1038/s41586-024-07224-3
Pol, S. A., Liljenberg, S., Barr, J., Simon, G., Wong-Dilworth, L., Paterson, D. L., et al. (2024). Induced degradation of SNAP-fusion proteins. RSC Chem. Biol. 5, 1232–1247. doi:10.1039/d4cb00184b
Puthenveetil, S., Liu, D. S., White, K. A., Thompson, S., and Ting, A. Y. (2009). Yeast display evolution of a kinetically efficient 13-amino acid substrate for lipoic acid ligase. J. Am. Chem. Soc. 131, 16430–16438. doi:10.1021/ja904596f
Raina, K., Noblin, D. J., Serebrenik, Y. V., Adams, A., Zhao, C., and Crews, C. M. (2014). Targeted protein destabilization reveals an estrogen-mediated ER stress response. Nat. Chem. Biol. 10, 957–962. doi:10.1038/nchembio.1638
Rana, S., Dranchak, P., Dahlin, J. L., Lamy, L., Li, W., Oliphant, E., et al. (2024). Methotrexate-based PROTACs as DHFR-specific chemical probes. Cell Chem. Biol. 31, 221–233.e14. doi:10.1016/j.chembiol.2023.09.014
Reicher, A., Koren, A., and Kubicek, S. (2020). Pooled protein tagging, cellular imaging, and in situ sequencing for monitoring drug action in real time. Genome Res. 30, 1846–1855. doi:10.1101/gr.261503.120
Reicher, A., Reiniš, J., Ciobanu, M., Růžička, P., Malik, M., Siklos, M., et al. (2024). Pooled multicolour tagging for visualizing subcellular protein dynamics. Nat. Cell Biol. 26, 745–756. doi:10.1038/s41556-024-01407-w
Robers, M., Pinson, P., Leong, L., Batchelor, R. H., Gee, K. R., and Machleidt, T. (2009). Fluorescent labeling of proteins in living cells using the FKBP12 (F36V) tag. Cytom. A 75, 207–224. doi:10.1002/cyto.a.20649
Rodriguez-Rios, M., Craigon, C. G., Nakasone, M. A., Bond, A. G., Dorward, M., Edmonds, A. K., et al. (2025). BromoCatch: a self-labelling tag platform for protein analysis and live cell imaging. bioRxiv, 647551. doi:10.1101/2025.04.07.647551
Romei, M. G., and Boxer, S. G. (2019). Split green fluorescent proteins: scope, limitations, and outlook. Annu. Rev. Biophys. 48, 19–44. doi:10.1146/annurev-biophys-051013-022846
Sanchez, H. M., Lapidot, T., and Shalem, O. (2024). High-throughput optimized prime editing mediated endogenous protein tagging for pooled imaging of protein localization. bioRxiv, 2024.09.16.613361. doi:10.1101/2024.09.16.613361
Sansbury, S. E., Serebrenik, Y. V., Lapidot, T., Smith, D. G., Burslem, G. M., and Shalem, O. (2025). Pooled tagging and hydrophobic targeting of endogenous proteins for unbiased mapping of unfolded protein responses. Mol. Cell 85, 1868–1886.e12. doi:10.1016/j.molcel.2025.04.002
Schmid-Burgk, J. L., Höning, K., Ebert, T. S., and Hornung, V. (2016). CRISPaint allows modular base-specific gene tagging using a ligase-4-dependent mechanism. Nat. Commun. 7, 12338. doi:10.1038/ncomms12338
Schwinn, M. K., Machleidt, T., Zimmerman, K., Eggers, C. T., Dixon, A. S., Hurst, R., et al. (2018). CRISPR-mediated tagging of endogenous proteins with a luminescent peptide. ACS Chem. Biol. 13, 467–474. doi:10.1021/acschembio.7b00549
Seabrook, L. J., Franco, C. N., Loy, C. A., Osman, J., Fredlender, C., Zimak, J., et al. (2024). Methylarginine targeting chimeras for lysosomal degradation of intracellular proteins. Nat. Chem. Biol. 20, 1566–1576. doi:10.1038/s41589-024-01741-y
Serebrenik, Y. V., Hellerschmied, D., Toure, M., López-Giráldez, F., Brookner, D., and Crews, C. M. (2018). Targeted protein unfolding uncovers a Golgi-specific transcriptional stress response. Mol. Biol. Cell 29, 1284–1298. doi:10.1091/mbc.E17-11-0693
Serebrenik, Y. V., Mani, D., Maujean, T., Burslem, G. M., and Shalem, O. (2024). Pooled endogenous protein tagging and recruitment for systematic profiling of protein function. Cell Genom 4, 100651. doi:10.1016/j.xgen.2024.100651
Serebrenik, Y. V., Sansbury, S. E., Kumar, S. S., Henao-Mejia, J., and Shalem, O. (2019). Efficient and flexible tagging of endogenous genes by homology-independent intron targeting. Genome Res. 29, 1322–1328. doi:10.1101/gr.246413.118
Shah, N. H., and Muir, T. W. (2014). Inteins: nature’s gift to protein chemists. Chem. Sci. 5, 446–461. doi:10.1039/C3SC52951G
Sigal, A., Milo, R., Cohen, A., Geva-Zatorsky, N., Klein, Y., Alaluf, I., et al. (2006a). Dynamic proteomics in individual human cells uncovers widespread cell-cycle dependence of nuclear proteins. Nat. Methods 3, 525–531. doi:10.1038/nmeth892
Sigal, A., Milo, R., Cohen, A., Geva-Zatorsky, N., Klein, Y., Liron, Y., et al. (2006b). Variability and memory of protein levels in human cells. Nature 444, 643–646. doi:10.1038/nature05316
Stevens, T. A., Tomaleri, G. P., Hazu, M., Wei, S., Nguyen, V. N., DeKalb, C., et al. (2024). A nanobody-based strategy for rapid and scalable purification of human protein complexes. Nat. Protoc. 19, 127–158. doi:10.1038/s41596-023-00904-w
Strickland, D., Lin, Y., Wagner, E., Hope, C. M., Zayner, J., Antoniou, C., et al. (2012). TULIPs: tunable, light-controlled interacting protein tags for cell biology. Nat. Methods 9, 379–384. doi:10.1038/nmeth.1904
Stroffekova, K., Proenza, C., and Beam, K. G. (2001). The protein-labeling reagent FLASH-EDT2 binds not only to CCXXCC motifs but also non-specifically to endogenous cysteine-rich proteins. Pflugers Arch. 442, 859–866. doi:10.1007/s004240100619
Suzuki, K., Tsunekawa, Y., Hernandez-Benitez, R., Wu, J., Zhu, J., Kim, E. J., et al. (2016). In vivo genome editing via CRISPR/Cas9 mediated homology-independent targeted integration. Nature 540, 144–149. doi:10.1038/nature20565
Tae, H. S., Sundberg, T. B., Neklesa, T. K., Noblin, D. J., Gustafson, J. L., Roth, A. G., et al. (2012). Identification of hydrophobic tags for the degradation of stabilized proteins. Chembiochem 13, 538–541. doi:10.1002/cbic.201100793
Tovell, H., Testa, A., Maniaci, C., Zhou, H., Prescott, A. R., Macartney, T., et al. (2019). Rapid and reversible knockdown of endogenously tagged endosomal proteins via an optimized HaloPROTAC degrader. ACS Chem. Biol. 14, 882–892. doi:10.1021/acschembio.8b01016
Tycko, J., DelRosso, N., Hess, G. T., Banerjee, A., Mukund, A., Van, M. V., et al. (2020). High-throughput discovery and characterization of human transcriptional effectors. Cell 183, 2020–2035. doi:10.1016/j.cell.2020.11.024
Tycko, J., Van, M. V., DelRosso, N., Ye, H., Yao, D., Valbuena, R., et al. (2024). Development of compact transcriptional effectors using high-throughput measurements in diverse contexts. Nat. Biotechnol., 1–14. doi:10.1038/s41587-024-02442-6
Veggiani, G., Nakamura, T., Brenner, M. D., Gayet, R. V., Yan, J., Robinson, C. V., et al. (2016). Programmable polyproteams built using twin peptide superglues. Proc. Natl. Acad. Sci. U. S. A. 113, 1202–1207. doi:10.1073/pnas.1519214113
Vedagopuram, S., Sindi, S. H., Chaudhary, S. K., Pergu, R., Singh, P., Karaj, E., et al. (2025). Ultrasmall chemogenetic tags with group-transfer ligands. bioRxiv, 653252. doi:10.1101/2025.05.10.653252
Wang, L., Hiblot, J., Popp, C., Xue, L., and Johnsson, K. (2020). Environmentally sensitive color-shifting fluorophores for bioimaging. Angew. Chem. Int. Ed. Engl. 59, 21880–21884. doi:10.1002/anie.202008357
Wilhelm, J., Kühn, S., Tarnawski, M., Gotthard, G., Tünnermann, J., Tänzer, T., et al. (2021). Kinetic and structural characterization of the self-labeling protein tags HaloTag7, SNAP-tag, and CLIP-tag. Biochemistry 60, 2560–2575. doi:10.1021/acs.biochem.1c00258
Wilson, C. M., Pommier, G. C., Richman, D. D., Sambold, N., Hussmann, J. A., Weissman, J. S., et al. (2024). Combinatorial effector targeting (COMET) for transcriptional modulation and locus-specific biochemistry. bioRxiv, 2024.10.28.620517. doi:10.1101/2024.10.28.620517
Winter, G. E., Buckley, D. L., Paulk, J., Roberts, J. M., Souza, A., Dhe-Paganon, S., et al. (2015). DRUG DEVELOPMENT. Phthalimide conjugation as a strategy for in vivo target protein degradation. Science 348, 1376–1381. doi:10.1126/science.aab1433
Wolf, P., Gavins, G., Beck-Sickinger, A. G., and Seitz, O. (2021). Strategies for site-specific labeling of receptor proteins on the surfaces of living cells by using genetically encoded peptide tags. Chembiochem 22, 1717–1732. doi:10.1002/cbic.202000797
Yang, X., Boehm, J. S., Yang, X., Salehi-Ashtiani, K., Hao, T., Shen, Y., et al. (2011). A public genome-scale lentiviral expression library of human ORFs. Nat. Methods 8, 659–661. doi:10.1038/nmeth.1638
Yao, J. Z., Uttamapinant, C., Poloukhtine, A., Baskin, J. M., Codelli, J. A., Sletten, E. M., et al. (2012). Fluorophore targeting to cellular proteins via enzyme-mediated azide ligation and strain-promoted cycloaddition. J. Am. Chem. Soc. 134, 3720–3728. doi:10.1021/ja208090p
Yarnall, M. T. N., Ioannidi, E. I., Schmitt-Ulms, C., Krajeski, R. N., Lim, J., Villiger, L., et al. (2023). Drag-and-drop genome insertion of large sequences without double-strand DNA cleavage using CRISPR-directed integrases. Nat. Biotechnol. 41, 500–512. doi:10.1038/s41587-022-01527-4
Yen, H.-C. S., Xu, Q., Chou, D. M., Zhao, Z., and Elledge, S. J. (2008). Global protein stability profiling in mammalian cells. Science 322, 918–923. doi:10.1126/science.1160489
Yi, H. B., Lee, S., Seo, K., Kim, H., Kim, M., and Lee, H. S. (2024). Cellular and biophysical applications of genetic code expansion. Chem. Rev. 124, 7465–7530. doi:10.1021/acs.chemrev.4c00112
Yu, Q., Pourmandi, N., Xue, L., Gondrand, C., Fabritz, S., Bardy, D., et al. (2019). A biosensor for measuring NAD+ levels at the point of care. Nat. Metab. 1, 1219–1225. doi:10.1038/s42255-019-0151-7
Zakeri, B., Fierer, J. O., Celik, E., Chittock, E. C., Schwarz-Linek, U., Moy, V. T., et al. (2012). Peptide tag forming a rapid covalent bond to a protein, through engineering a bacterial adhesin. Proc. Natl. Acad. Sci. U. S. A. 109, E690–E697. doi:10.1073/pnas.1115485109
Zhang, H., Aonbangkhen, C., Tarasovetc, E. V., Ballister, E. R., Chenoweth, D. M., and Lampson, M. A. (2017). Optogenetic control of kinetochore function. Nat. Chem. Biol. 13, 1096–1101. doi:10.1038/nchembio.2456
Zhang, L., Riley-Gillis, B., Vijay, P., and Shen, Y. (2019). Acquired resistance to BET-PROTACs (proteolysis-targeting chimeras) caused by genomic alterations in core components of E3 ligase complexes. Mol. Cancer Ther. 18, 1302–1311. doi:10.1158/1535-7163.MCT-18-1129
Zhang, Y., Sohn, C., Lee, S., Ahn, H., Seo, J., Cao, J., et al. (2020). Detecting protein and post-translational modifications in single cells with iDentification and qUantification sEparaTion (DUET). Commun. Biol. 3, 420. doi:10.1038/s42003-020-01132-8
Keywords: pooled protein tagging, SPOTLITES, HaloTag, induced proximity, hydrophobic tagging, protein degradation, CRISPR knock-in
Citation: Larrimore KE and Serebrenik YV (2025) Utilizing small molecules to probe and harness the proteome by pooled protein tagging with ligandable domains. Front. Pharmacol. 16:1593844. doi: 10.3389/fphar.2025.1593844
Received: 14 March 2025; Accepted: 03 June 2025;
Published: 23 June 2025.
Edited by:
Madhu Sudhan Ravindran, Bristol Myers Squibb, United StatesReviewed by:
Bingbing Li, Oregon Health and Science University, United StatesUtpal P. Davé, Indiana University Bloomington, United States
Copyright © 2025 Larrimore and Serebrenik. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yevgeniy V. Serebrenik, eWV2Z2VuaXkuc2VyZWJyZW5pa0BydXRnZXJzLmVkdQ==