 Muhammad Azrai1*
Muhammad Azrai1* Muhammad Aqil2
Muhammad Aqil2 Roy Efendi2Bunyamin Zainuddin2Elkawakib Syam’un1Muhammad Fuad Anshori1Muhammad Riadi1Slamet Bambang Priyanto2Nining Nurini Andayani2Muhammad Yasin2Reynaldi Laurenze1Amiruddin Syam3
Roy Efendi2Bunyamin Zainuddin2Elkawakib Syam’un1Muhammad Fuad Anshori1Muhammad Riadi1Slamet Bambang Priyanto2Nining Nurini Andayani2Muhammad Yasin2Reynaldi Laurenze1Amiruddin Syam3- 1Department of Agronomy, Faculty of Agriculture, Hasanuddin University, Makassar, Indonesia
- 2Research Center for Food Crops, National Research and Innovation Agency (BRIN), Bogor, Indonesia
- 3Research Center for Macroeconomics and Finance, National Research and Innovation Agency (BRIN), Jakarta, Indonesia
Despite substantial increases in planting density to enhance maize grain yield, productivity at the plant level has remained stagnant. Although leaf-related traits have been extensively studied in commercial hybrids, they remain underexplored in tropical maize breeding programs. This study aimed to: (1) investigate genotype-specific yield responses under standard, intermediate, and intensive planting densities; (2) evaluate correlations among key leaf physiological traits across density changes; and (3) identify resilient hybrids for high-density cultivation through a multi-trait genotype-ideotype distance index (MGIDI) assessment. The research was conducted at the Bajeng Experimental Station in Indonesia using a split-plot randomized complete block design with three replications. The trial assigned 11 maize genotypes and two upright-leaf commercial hybrids to the main plots, with subplots testing three densities: standard (±71,000 plants ha−1), intermediate (±81,000 plants ha−1), and intensive (±95,000 plants ha−1). The results indicated that genotype (G) and population density (D) significantly influenced yield, while the G × D interaction had no significant effect (p = 0.2981). Intermediate density achieved the highest average yield (12.85 t ha−1), surpassing both standard (11.54 t ha−1) and intensive (11.79 t ha−1) planting densities. The MGIDI model identified hybrids G4, G2, and G5 as broadly adaptable across densities, demonstrating stability in intermediate and intensive population densities. The MGIDI framework is recommended for integration into genotype–environment interaction analyses, enabling targeted identification of stress-resilient cultivars by balancing trait trade-offs.
1 Introduction
Maize (Zea mays L.) is a cornerstone of global food security, serving as a vital source of nutrition and industrial raw materials. With escalating demand, innovative agronomic practices are critical to enhancing productivity, particularly in tropical regions where environmental constraints and suboptimal agricultural practices hinder yields. Among these strategies, high-density planting systems, where plants are spaced closely to maximize land use, have emerged as a promising solution. While this approach intensifies competition for resources, which drives vertical growth and sturdier stalks to optimize light access, it also underscores the necessity of breeding maize varieties resilient to density-induced stress.
A pivotal challenge in high population density lies in optimizing photosynthesis. Enhancing productivity hinges on selecting stress-tolerant genotypes, refining planting densities, and adopting precision water and nutrient management. Canopy architecture plays a central role in regulating the interception of solar radiation. Studies suggest that a vertically oriented leaf arrangement improves photosynthetic efficiency under high-light conditions by minimizing mutual shading and maximizing light capture (Greveniotis et al., 2019). Such structural adaptations enable plants to allocate resources more effectively, directly influencing biomass accumulation and yield. However, the relationship between plant density and yield is non-linear and heavily mediated by environmental factors. For instance, elevated planting densities can deplete soil moisture, leading to variable yield outcomes depending on seasonal rainfall patterns and water availability.
During the productive growth phase, photosynthesis serves as the biochemical linchpin, generating carbohydrates essential for plant survival and ecosystem functioning. It acts as the primary pathway for carbon fixation, producing non-structural carbohydrates (NSCs) like soluble sugars and starch. These NSCs are critical for balancing carbon input and usage, particularly during stress, helping plants manage energy demands and maintain physiological stability (Sun et al., 2021). NSCs also play a key role in post-stress recovery. Following drought, for instance, rewatering allows plants to rapidly replenish their NSC reserves, facilitating growth restoration and reactivation of photosynthetic processes (Gori et al., 2023). Photosynthetic efficiency during the generative phase is a complex interplay of various photosynthetic pigments (Simkin et al., 2022), stomatal conductance, and the balance between carbon fixation and photorespiration (Bloom and Lancaster, 2018). Optimizing these factors can significantly enhance photosynthetic performance and plant productivity under varying environmental conditions.
As competition for light intensifies under high population density, it drives morphological adaptations such as stem elongation and leaf reorientation. However, excessive crowding can trigger shade avoidance responses, morphological and physiological adjustments initiated in response to vegetative shading, often triggered by changes in the red to far-red light ratio. While these responses may promote vertical growth to outcompete neighbours in natural ecosystems, they can negatively impact yield in high-density agricultural settings. In maize, shade avoidance responses lead to a reduced root-to-shoot ratio, resulting in diminished root biomass and excessive shoot elongation, which may compromise anchorage and resource uptake (Page et al., 2011; Afifi and Swanton, 2011). Stem elongation is often exaggerated, increasing susceptibility to lodging and reducing plant stability (Shi et al., 2024; Yu et al., 2018). Additionally, these responses tend to produce narrower leaf angles which, although improving light penetration through the canopy, can reduce overall light interception efficiency and yield.
Maize ideotype cultivars with upright upper and middle leaves improve light distribution within dense canopies, supporting prolonged photosynthesis, delayed senescence, greater dry matter accumulation, and higher grain yields compared to flat-leaf types. Under high population density planting, compact ideotype cultivars tend to develop more upright middle and upper leaves, enhancing light penetration through the canopy. This trait helps sustain green leaf area, slows leaf senescence, boosts dry matter accumulation during grain filling, and contributes to superior yield performance compared to flatter-leafed varieties. Moreover, the number of kernels per plant may also decline as shade avoidance redirects biomass away from reproductive development toward vegetative structures, leading to fewer resources available for ear and grain formation (Zheng et al., 2024; Chibane et al., 2021).
Nitrogen and water are among the other critical factors influencing maize productivity, particularly under high planting densities where competition intensifies stress on both resources. Limited nitrogen availability can lead to early canopy senescence, reduced dry matter accumulation, and ultimately lower yields (Tilman et al., 2011). Simultaneously, water scarcity further constrains growth, as maize yield responses to density are highly dependent on water availability throughout the season. Excessive nitrogen input, while intended to counter nutrient limitations, may worsen shading and reduce light-use efficiency, besides causing environmental concerns (Ju et al., 2009). Therefore, integrated strategies such as fertigation, which combines split nitrogen applications with precise water delivery, offer a promising solution. This approach enhances nitrogen use efficiency, supports balanced dry matter production and allocation, and helps sustain high yields even under high population planting and water-limited conditions (Guo et al., 2021).
The selection of maize genotypes suited for high-density planting has evolved from traditional single-trait methods to more integrative multi-trait approaches, enabling breeders to better capture the complex interactions among yield components and stress-adaptive traits. Recent advancements in phenotyping tools, particularly digital imaging meters, have significantly enhanced the ability to assess physiological traits in maize. These tools leverage various imaging technologies to provide high-throughput, non-destructive, and precise measurements of plant traits, including prediction and classification tasks (Coswosk et al., 2024; Zainuddin and Aqil, 2021; Aqil et al., 2025). Additionally, the integration of modern analytical techniques like the Multi-trait Genotype-Ideotype Distance Index (MGIDI) has enhanced the precision and speed of genotype selection by processing large datasets and identifying ideotype-driven candidates with superior performance (Yue et al., 2022; Azrai et al., 2023). These tools not only accelerate breeding pipelines but also support the development of climate-resilient, resource-efficient maize hybrids tailored to the demands of intensive cropping systems.
The study aimed to (1) investigate genotype-specific yield responses under standard, intermediate, and intensive planting densities; (2) evaluate correlations among key leaf physiological traits across density levels; and (3) identify resilient hybrids for high-density cultivation through MGIDI assessment.
2 Materials and methods
2.1 Site description and experimental design
The experiment was conducted during the 2021 growing season at two adjacent sites within the Bajeng Experimental Station (5.3167° S, 119.7667° E), Gowa Regency, Indonesia, a region characterized by humid tropical conditions with an annual rainfall of 2,500 mm and temperatures ranging between 24 and 32 °C. The Bajeng soil sample has a loam texture (40% sand, 50% silt, 10% clay) with slightly acidic pH (5.62 H₂O), low organic carbon (0.98%), and nitrogen (0.14%), resulting in a C/N ratio of 7. Though organic matter is limited, nutrient levels are high, with Olsen/Bray P₂O₅ at 114 ppm and K₂O at 146 ppm, indicating good phosphorus and potassium availability. Proper pH and nitrogen management can further enhance productivity. A split-plot randomized complete block design with three replications was implemented to evaluate genotype by density interactions.
The main plot treatments involved three maize planting densities: standard/SD (± 71,000 plant populations ha−1), intermediate/ID (± 81,000 plant populations ha−1), and intensive/XD (± 95,000 plant populations ha−1). Subplots consisted of 13 hybrid maize genotypes, including 11 advanced experimental hybrids (coded G01–G11) selected through strict criteria prioritizing upright leaf architecture (≤30° leaf angle) and compact canopy structure. Two commercial check hybrids were incorporated: Bisi-18 (G12), an upright-leaved cultivar (leaf angle <25°) and Nasa-29 (G13), a conventional hybrid with a drooping leaf phenotype (leaf angle >45°), serving as contrasting canopy models. Parental lineage and breeding pedigrees of experimental genotypes are detailed in Table 1. Each genotype × density combination was replicated three times, resulting in nine experimental plots per genotype (3 densities × 3 replicates) and a total of 117 plots (13 genotypes × 3 densities × 3 replicates).
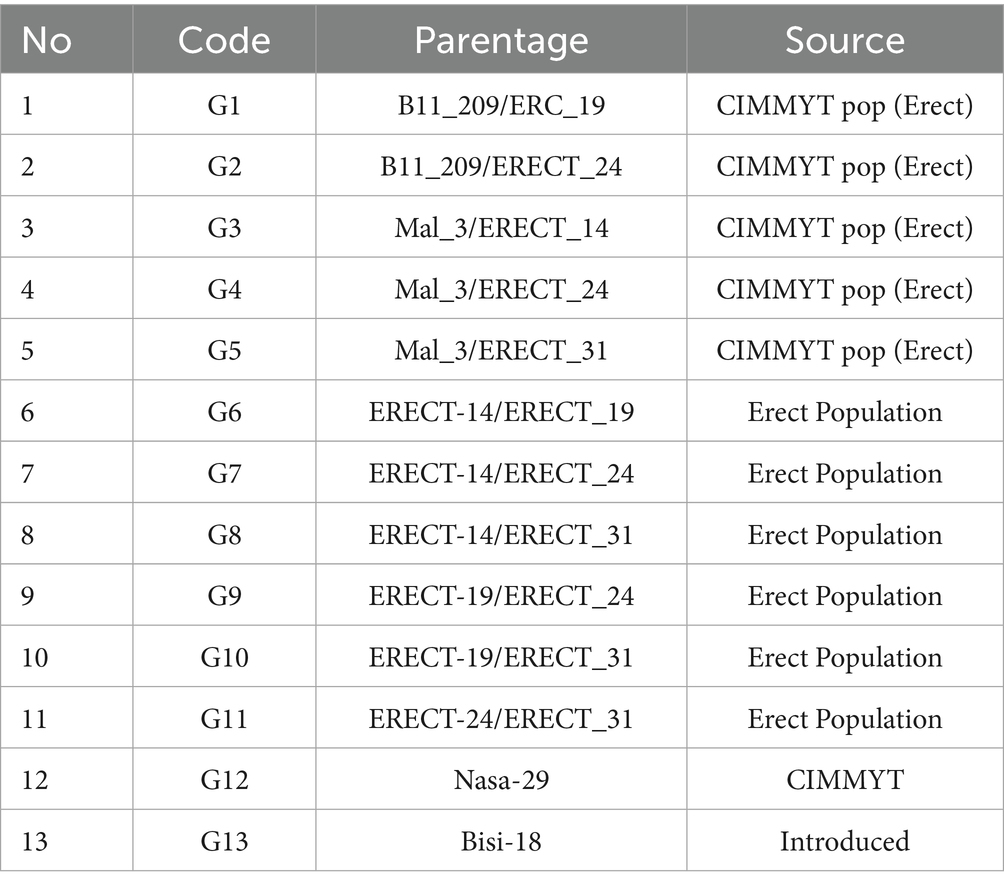
Table 1. Parentage and origin information of genotypes used in the experiment.
Crops were managed under optimal agronomic conditions, including irrigation scheduled to maintain soil moisture. A basal dose of 350 kg ha−1 of compound fertilizer (15:15:15) was incorporated before planting, followed by split applications of nitrogen fertilizer (46% N), applied at a rate of 200 kg ha−1 at 10 and 30 days after planting.
2.2 Phenotypic and yield trait quantification
Canopy architecture and physiological traits were assessed using standardized protocols. Leaf angle (LA) was measured at the midrib of the ear leaf using a digital inclinometer (Suunto PM-5/360PC), with leaf orientation (LO) classified as upright (<30°), semi-erect (30–45°), or drooping (>45°). The formula used for calculating LA non-destructively using the Radford coefficient method:
Where LA represents leaf area in square centimeters (cm2), K is a constant value (0.75), L denotes leaf length measured in centimeters (cm), and W stands for maximum leaf width also measured in centimeters (cm). Measurements began at the 7th leaf and extended upward to the plant apex. Another leaf area indices including ear leaf area (ELA), upper leaf area (ULA), and total leaf area per plant (TLDT). To determine how much ULA is needed to produce one gram of grain per plant, the total ULA per plant (LAG.) was divided by the grain yield (in grams) per plant.
2.3 Yield component analysis
At harvest, ears were collected from a 5 m central row segment per plot. Ear length (EL) and diameter (ED) were measured using digital callipers. KPP refers to the number of kernels produced per plant, while GPP denotes the total grain weight, measured in grams, produced by each plant. These parameters are essential indicators of individual plant productivity and are commonly used in evaluating yield components in maize. The kernel number per plant (KPP) was counted manually from three representative ears. Shelling percentage (SP) was calculated as grain weight cob weight × 100 cob weight. 1,000-kernel weight (1000KW) was adjusted to 15% moisture using a digital grain moisture tester (Grainer DMC-700, ±0.05% resolution).
The selection of optimal genotypes was conducted using the multi trait Genotype-Ideotype Distance Index (MGIDI) to integrate multiple trait data (Olivoto and Nardino, 2021). For standard and intermediate planting densities, grain yield was double weighted, and other traits were weighted single. For intensive planting density, grain yield and leaf orientation were weighed double, and the others single. Traits were rescaled using a transformation equation, where new values were defined based on trait-specific gains: for positive gains, traits were scaled between 0 and 100, while negative gains inverted this range. Factor Analysis (FA) reduced data dimensionality, employing the model:
Where F (genotype scores), Z (rescaled data), A (loadings), and R (correlation matrix). An ideotype matrix was established, and MGIDI was computed as the Euclidean distance between genotype factor scores and the ideotype:
with lower values indicating proximity to the ideotype. Selection employed a 20% intensity threshold.
2.4 Statistical analysis
Data were analysed using a hierarchical split-plot ANOVA model:
where Yijk = observed value, μ = grand mean, τi = planting density effect, βj = genotype effect, and ϵijk = residual error. Post-hoc comparisons employed Fisher’s LSD test (α = 0.05) to identify significant differences among treatments. Relationships between traits were assessed using Spearman’s rank correlation to account for non-parametric data distributions. Analyses were conducted in R v4.2.1 (lme4, agricolae packages). MGIDI implementation was performed via the R package metan.1
3 Results
3.1 Combined analysis of variance
A combined analysis of variance was thoroughly performed across three distinct population densities to assess a range of agronomic traits, yield components, and overall yield performance. Variance analysis of agronomic and yield component interactions is presented in Table 2. The results revealed that the density treatment significantly affected all observed traits, except kernel number per plant, 1,000 kernel weight and shelling percentage. This indicates that while plant density generally exerts a considerable impact on various aspects of plant growth and development, the kernel attributed traits remain relatively stable under such conditions. On the other hand, the genotype effect was significant for all observed traits except for grain yield per plant. This suggests that the hybrids evaluated in this study exhibit a high degree of phenotypic diversity in terms of canopy area, leaf size, leaf angle, yield components, and maize yield.
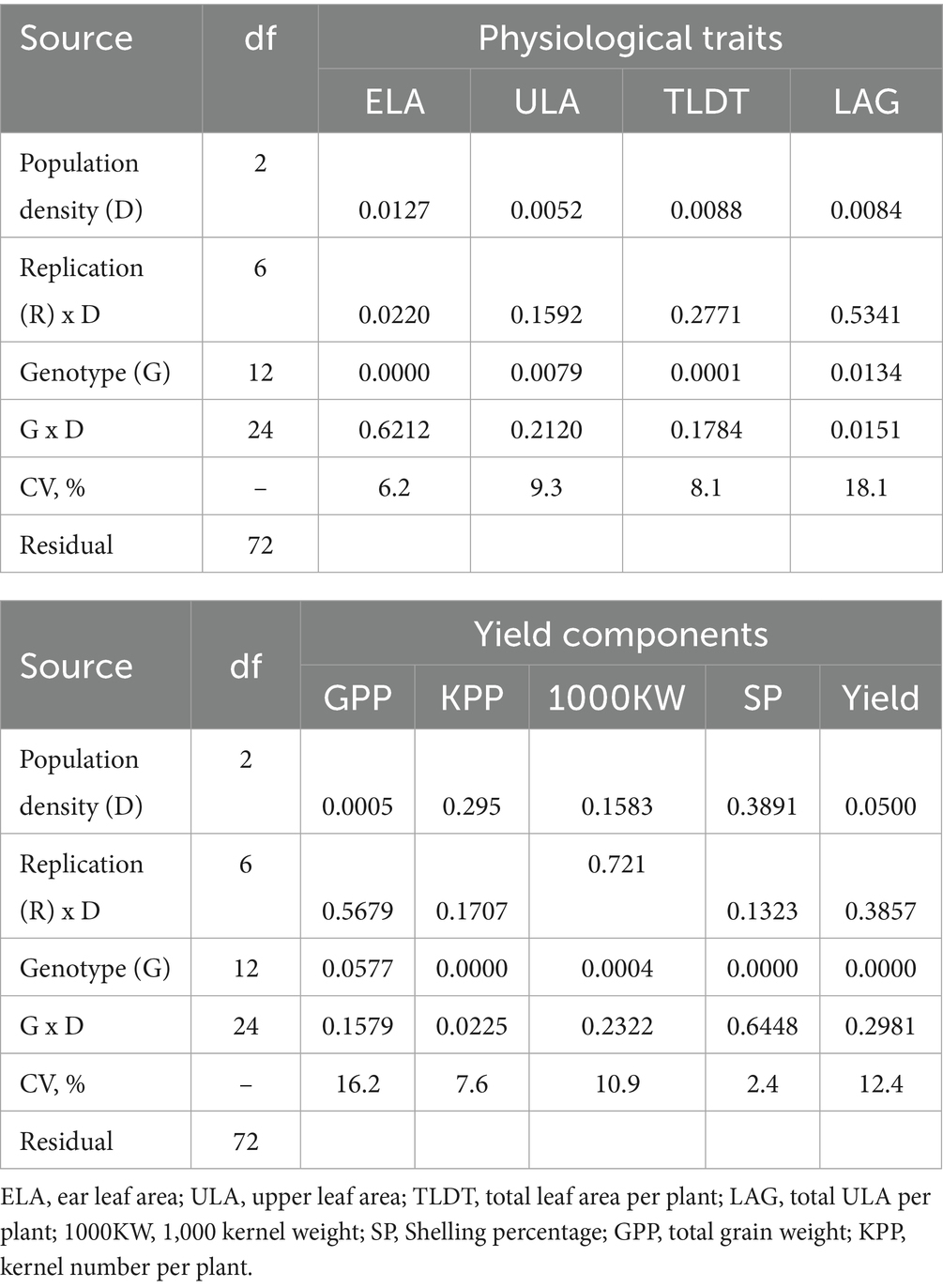
Table 2. Variance analysis of physiological and yield component interactions.
Analysis of variance showed that population density and genotype each exert strong main effects on both canopy architecture and yield components. Increasing planting density significantly affects all four physiological traits ELA, ULA TLDT and LAG. In addition, genotypes had highly significant effects on these traits (all p < 0.05). However, the density × genotype interaction was significant only for total upper leaf area per plant (LAG; p = 0.0151). In terms of yield components, increasing density markedly boosted total grain weight per plant (GPP; p = 0.0005) and grain yield that was on the threshold of significance (p = 0.0500). Density did not significantly affect kernel number per plant (KPP; p = 0.295), thousand-kernel weight (1000KW; p = 0.1583), and shelling percentage (SP; p = 0.3891). Genotype shows highly significant effects on KPP, 1000KW, SP, and grain yield (all p < 0.001). However, the density × genotype interaction was significant only for kernel number per plant (KPP; p = 0.0225). The significant genotype × density interactions for the physiological trait LAG and the yield component KPP demonstrate that different hybrids control their upper canopy leaf area and kernel set in distinct ways under crowding stress.
3.2 Mean performance against different population density
Analysis of mean performance indicated that grain yield varied significantly with increasing population density. The evaluation of maize hybrids across three population densities: 71,000 plants ha−1 (SD), 81,000 plants ha−1 (ID), and 95,000 plants ha−1 (XD) revealed significant differences in agronomic traits, which are critical for maximizing yield (Figure 1). Specifically, the ID population at 81,000 plants ha−1 outperformed both the SD and XD populations, producing 11% more yield. Statistical analysis showed that the ID population achieved an average yield of 12.85 t ha−1, which was significantly higher than the yields of the SD (11.54 t ha−1) and XD (11.79 t ha−1) populations. These results indicated that an intermediate plant population density is optimal for maximizing maize yield.
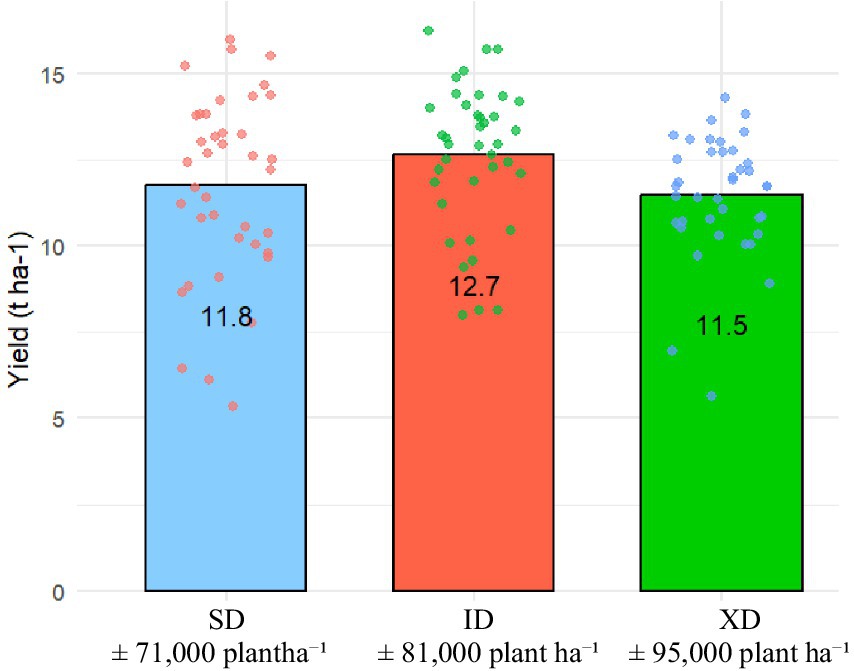
Figure 1. Boxplot of grain yield under SD, ID and XD.
Under ideal growing conditions, maize population density strongly influences nearly every growth parameter and drives competition among plants. Grain yield per plant decreases with reduced light and other environmental resources. Analyzing the responses of individual genotypes to different densities revealed distinct variations in grain yield as the population density increased from 71,000 to 81,000 plants ha−1 (Table 3). Ten hybrids, including G1/B11_209 × ERECT_19, G2/B11_209 × ERECT_24, G3/Mal_3 × ERECT_14, G4/Mal_3 × ERECT_24, G5/Mal_3 × ERECT_31, G6/ERECT_14 × ERECT_19, G7/ERECT_14 × ERECT_24, G8/ERECT_14 × ERECT_31, G9/ERECT_19 × ERECT_24, and G13/Bisi-18, showed a positive response with increased yield. In contrast, three hybrids G10/ERECT_19 × ERECT_31, G11/ERECT_24 × ERECT_31, and G12/Nasa-29, exhibited a negative response with decreased yield. At 95,000 plant ha−1, only three hybrids (B11_209 × ERECT_19, B11_209 × ERECT_24, and Mal_3 × ERECT_14) showed a positive response with higher yields. Ten hybrids, including Mal_3 × ERECT_24, Mal_3 × ERECT_31, ERECT_14 × ERECT_19, ERECT_14 × ERECT_24, ERECT_14 × ERECT_31, ERECT_19 × ERECT_24, ERECT_19 × ERECT_31, ERECT_24 × ERECT_1, Nasa-29, and Bisi-18, exhibited decreased yields. Under intensive population density, certain hybrids demonstrate inadequate adaptation to higher population densities, potentially leading to reduced yields or other agronomic challenges.
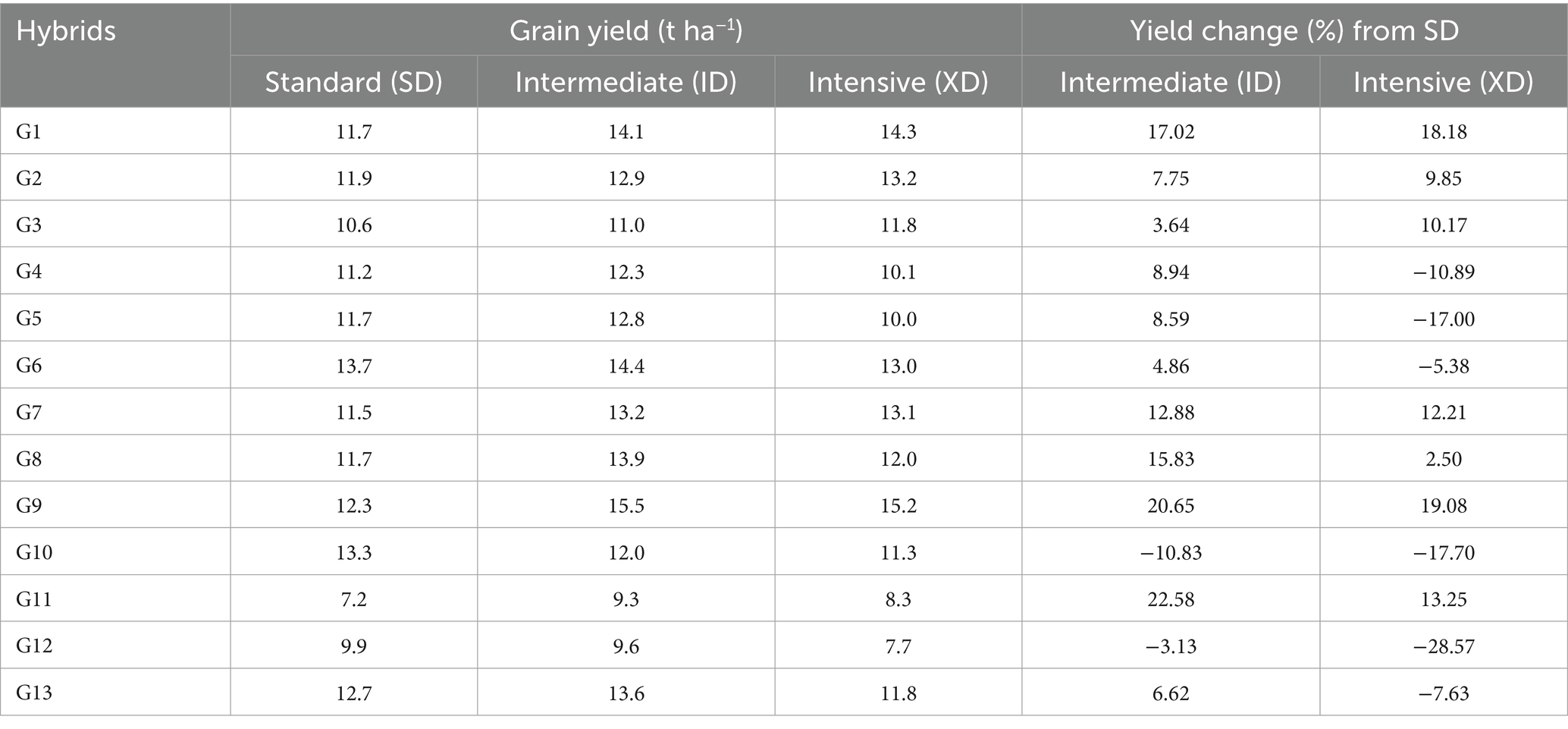
Table 3. Grain yield and percentage of yield change (%) under varying planting densities.
3.3 Leaf and ear characteristics
The analysis of the 13 maize hybrids at three different population densities reveals significant trends in leaf area and yield (Table 4). At the standard, the average yield was 11.49 t ha−1, with an average total leaf area of 6048.94 cm2 and upper leaf area of 2893.06 cm2, respectively. Among these hybrids, G6 stood out with the highest yield of 13.70 t ha−1, despite having moderate upper leaf area (2584.5 cm2) and total leaf area (5619.41 cm2). Conversely, G9 exhibited the highest total (6312.11 cm2) and upper leaf area (3121.23 cm2), yet its yield was not the highest (12.29 t ha−1). In addition to agronomic traits, yield components such as kernel number per plant, 1,000-kernel weight, and shelling percentage significantly affect the grain yield of tested hybrid candidates with varying magnitudes. At the normal density, hybrids like G6, G10, and G9 exhibit varying combinations of these traits. G6, with a kernel number per plant of 427.36 and a high 1,000-kernel weight of 300.98 g, achieves the highest yield, indicating the balance between kernel number and weight is crucial. G10, despite having the highest kernel number per plant (498.59), yields slightly less due to a lower 1,000-kernel weight (259.33 g). Conversely, G9, which has the highest 1,000-kernel weight recorded at 311.08 g, demonstrates a moderate yield performance.
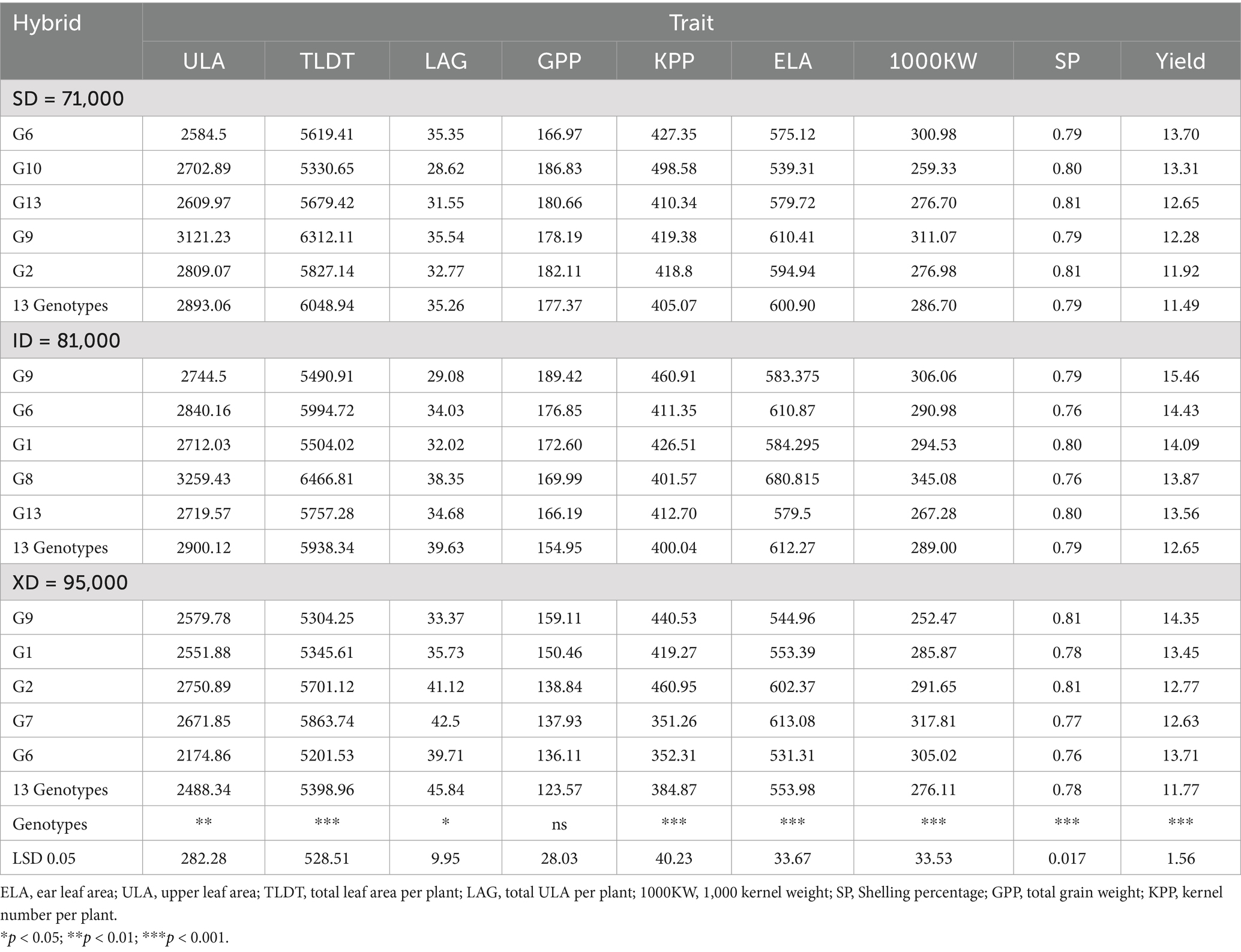
Table 4. Comparison of the top five grain yields with the average for 13 hybrids grown at normal, medium, and high plant densities.
Under intermediate population density, the 13 maize genotypes averaged a total leaf area of 5,938 cm2 and an upper leaf area of 2,900 cm2, producing a mean grain yield of 12.65 t ha−1. Genotype G9 achieved the highest yield (15.46 t ha−1) with moderate leaf areas (upper: 2744.5 cm2; total: 5490.9 cm2). By contrast, G8 exhibited the largest leaf areas (upper: 3259.4 cm2; total: 6466.8 cm2) but yielded 13.87 t ha−1. Genotype G6, which had a total leaf area above the mean (5994.7 cm2), delivered 14.43 t ha−1. Furthermore, as for yield components, G9’s performance was driven by its high kernel number per plant (460.9) and robust 1,000-kernel weight (306.1 g). G6 maintained a strong 1,000-kernel weight (291.0 g) with a slightly lower kernel count (411.4). Although G1 and G8 had fewer kernels per plant both achieved competitive yields through favorable 1,000-kernel weights and efficient shelling percentages (80.7 and 76.3%).
At the intensive population density, the average yield was 11.77 t ha−1, with an average total leaf area of 5398.96 cm2 and upper leaf area of 2488.34 cm2. G9 again showed robust performance with the highest yield of 15.15 t ha−1, despite having moderate leaf areas (upper: 2579.78 cm2, total: 5304.25 cm2). G2 and G7, with relatively higher total leaf areas (5701.12 cm2 and 5863.74 cm2, respectively), yielded lower (13.22 and 13.14 t ha−1, respectively). As for yield components, the stress from high population density is evident, with a general decrease in kernel number per plant and 1,000-kernel weight across hybrids. However, G9 continues to perform well, with a kernel number per plant of 440.53 and a 1,000-kernel weight of 252.47 g, supported by a high shelling percentage (81%). This hybrid’s consistent performance across densities underlines the importance of a stable kernel number and shelling efficiency in mitigating the stress of high density. G2, with the highest kernel number per plant (460.95) but a moderate 1,000-kernel weight (291.66 g), and G1, with a balanced 1,000-kernel weight (285.88 g) and shelling percentage (78.33%).
3.4 Trait–yield correlations across planting densities
Correlation analysis of morpho-physiological traits of maize under three population densities is shown in Figure 2. Under normal planting density, yield gains center on maximizing photosynthetic capacity (via ELA and ULA) and kernel weight (1000KW), reflecting sufficient resource availability for both canopy expansion and grain filling. The persistence of LAG as a negative predictor reinforces its utility as a universal efficiency metric, even in low-stress environments. Breeding efforts should emphasize leaf-area expansion and kernel plumpness, as competition-driven trade-offs (e.g., kernel number vs. size) are less pronounced. However, the moderate KPP–yield correlation suggests that kernel number remains a complementary target, albeit secondary to kernel weight.
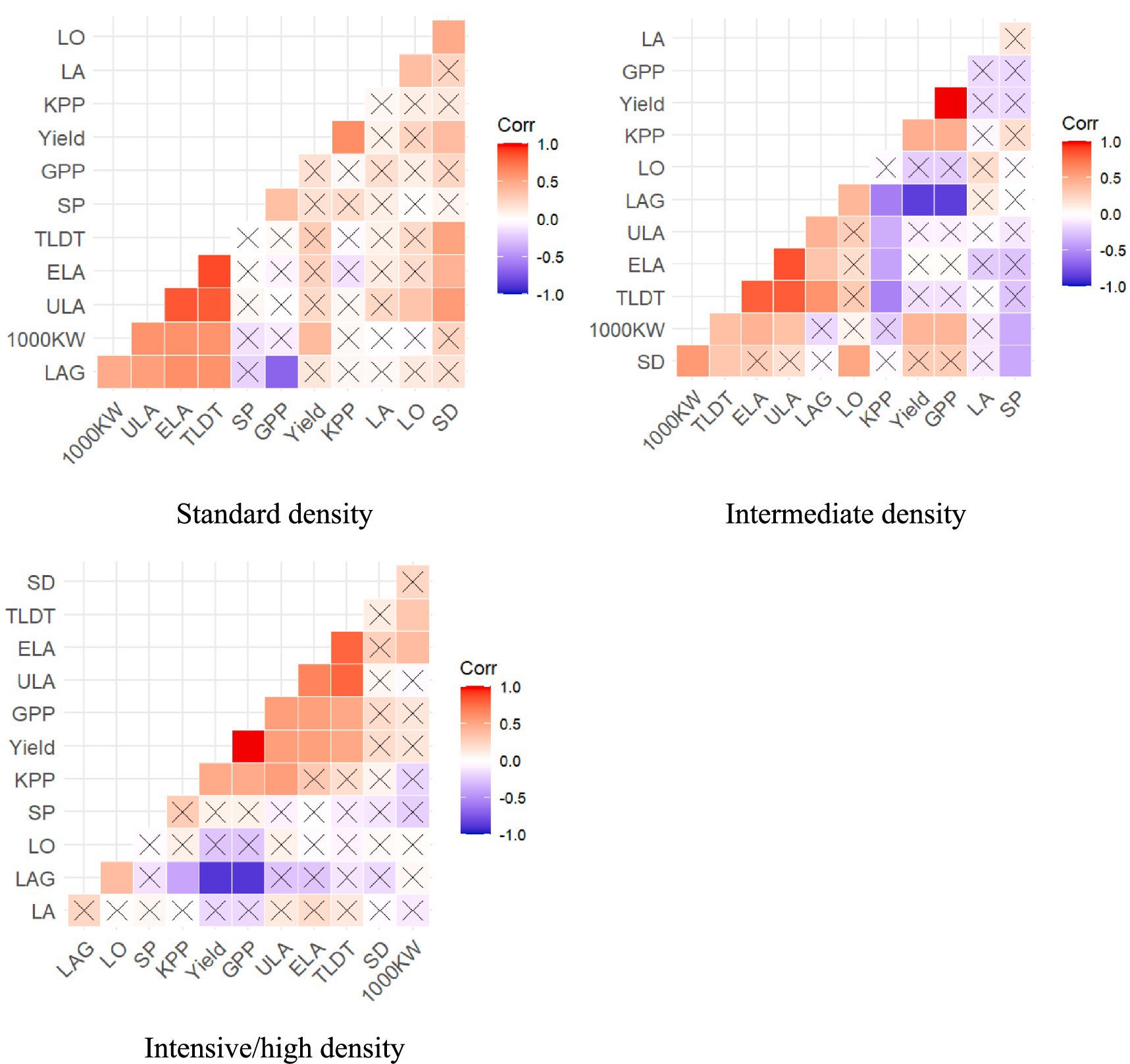
Figure 2. Correlation analysis of morphophysiological traits of maize under three population densities. LAG, total ULA per plant; LO, leaf orientation; SD, stem diameter; 1000KW, 1000 kernel weight; SP, Shelling percentage; GPP, total grain weight; KPP, kernel number per plant; ELA, ear leaf area; ULA, upper leaf area; TLDT, total leaf area per plant.
The dominance of KPP over GPP in yield determination under high density aligns with resource limitation dynamics, which drive plants to prioritize kernel survival over grain initiation under stress. The strong negative correlation of LAG with yield emphasizes that efficient leaf-area allocation, not merely maximizing foliage is critical in dense stands. This supports the conclusion that high-LAG genotypes waste resources on non-productive biomass under competition. While leaf-area traits (ELA, ULA) and 1000KW contribute positively, their intermediate effects suggest breeders should prioritize reducing LAG and enhancing kernel set efficiency to mitigate crowding penalties. The negative SP and LO associations further underscore the need for compact canopies and optimized assimilate partitioning. These results support density-specific trait prioritization, where high-density breeding should emphasize reducing LAG and increasing KPP to mitigate resource limitations, while normal-density programs should focus on enhancing leaf-area potential and kernel weight.
3.5 Principal component analysis across population densities
Principal Component Analysis (PCA) of the standard density trial identified four components (eigenvalues >1) explaining 85.0% of total variance (PC1: 41.5%, PC2: 18.1%, PC3: 14.6%, PC4: 10.8%), with a mean communality of 0.85 indicating robust trait covariance capture (Figure 3). Factor 1 (41.5%) represented photosynthetic capacity and kernel-filling potential, marked by strong negative loadings for total leaf area (TLDT = −0.96), ear leaf area (ELA = −0.96), and 1,000-kernel weight (1000KW = −0.81). Factor 2 (18.1%) captured yield output through negative associations with grain yield (−0.92) and kernels per plant (KPP = −0.87). Architectural traits dominated Factor 3 (14.6%), with leaf orientation (LO = −0.89) and angle (LA = −0.72) as key drivers, while Factor 4 (10.8%) emphasized shelling percentage (SP = 0.95). These axes provide a framework for the MGIDI index, enabling selection of genotypes balancing canopy size, yield potential, and harvest efficiency under non-crowded conditions.
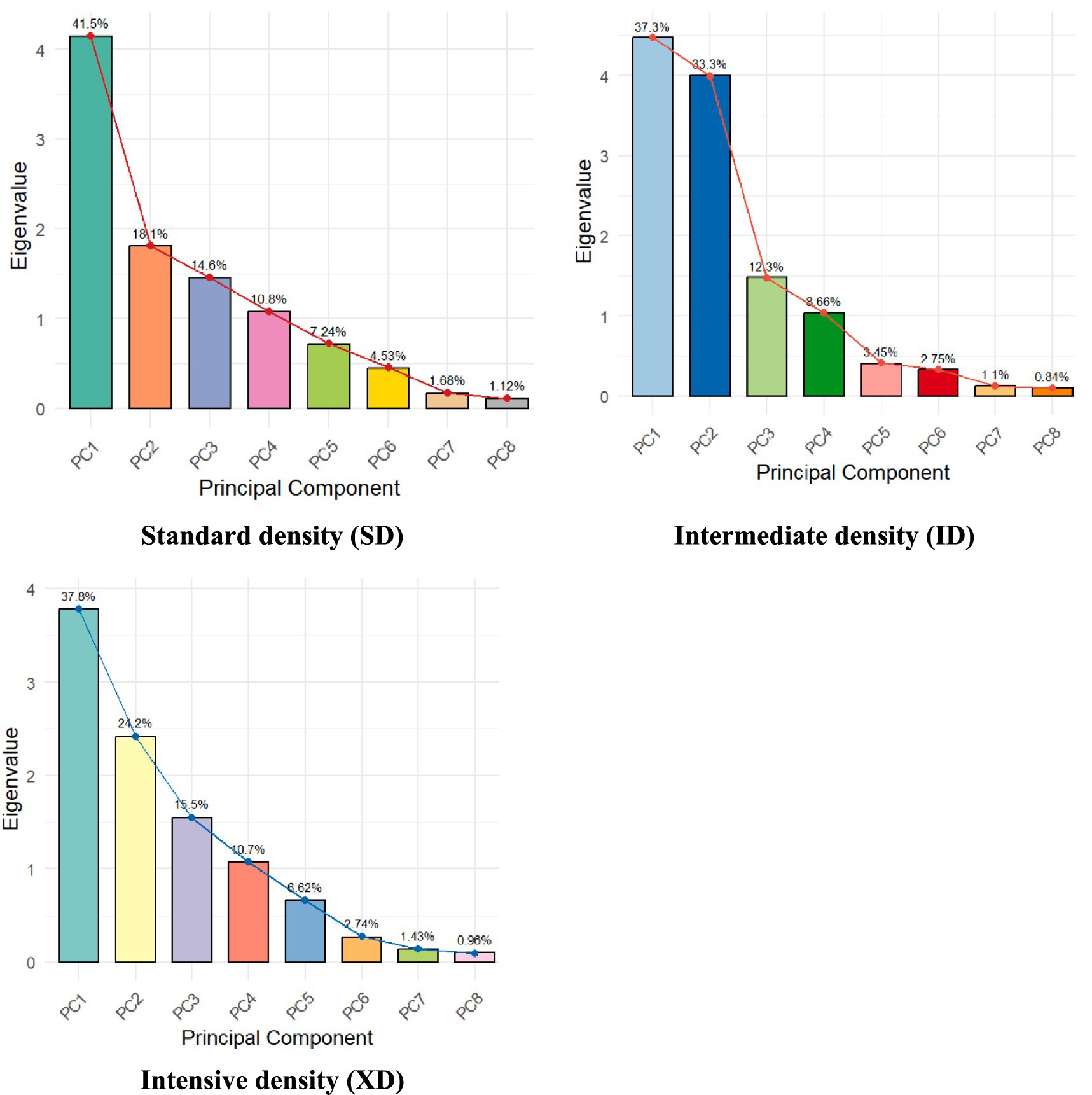
Figure 3. Eigenvalue and explained variance of morphophysiological traits of maize under three population densities.
Under moderate density, PCA revealed four components accounting for 91.6% of variance (PC1: 37.3%, PC2: 33.3%, PC3: 12.3%, PC4: 8.7%), with a mean communality of 0.92 confirming comprehensive trait representation. Factor 1 (37.3%) integrated photosynthetic capacity via positive loadings for ear leaf area (ELA = 0.97) and upper leaf area (ULA = 0.95). Factor 2 (33.3%) contrasted yield (−0.90) and kernel number (KPP = −0.83), reflecting resource allocation trade-offs. Architectural traits (LO = −0.90, LA = −0.83) defined Factor 3 (12.3%), while Factor 4 (8.7%) linked shelling percentage (SP = 0.90) with density adaptation. This supports MGIDI-driven selection for genotypes optimizing photosynthesis, yield, and canopy architecture under intermediate crowding.
At high density, PCA extracted four components explaining 88.2% of variance (PC1: 37.8%, PC2: 24.2%, PC3: 15.5%, PC4: 10.7%), with communality (0.882) affirming robust dimensionality reduction. Factor 1 (37.8%) highlighted yield efficiency through negative loadings for leaf-area-per-gram (LAG = −0.99) and yield (−0.94). Factor 2 (24.2%) combined photosynthetic capacity (TLDT = 0.94, ELA = 0.92) with kernel weight (1000KW = 0.79), while Factor 3 (15.5%) emphasized density tolerance (SD = 0.86) and shelling efficiency (SP = 0.71). Factor 4 (10.7%) revealed an architectural trade-off (LO = 0.77 vs. KPP = −0.80).
The PCA-derived MGIDI framework prioritized biologically critical trait synergies across densities. At normal density, selection emphasizes photosynthetic capacity and kernel plumpness, reflecting resource abundance. Under moderate density, architectural optimization and yield-component trade-offs gain prominence, while high-density conditions demand efficiency traits (e.g., low LAG) and stress resilience. The orthogonal separation of canopy architecture from yield components in all analyses underlines the need for balanced breeding strategies. By retaining >85% variance in four components, MGIDI ensures selection accounts for both major drivers and nuanced trade-offs, enabling development of density-specific ideotypes tailored to light capture, resource allocation, and stress adaptation.
3.5.1 Evaluation of factors and genetic gains
A Varimax-rotated factor analysis of ten agronomic traits under standard planting density (67,000 plants ha−1) consistently extracted four orthogonal factors (communalities 0.54–0.96, mean 0.85) that together explain the bulk of observed variation (Table 5). Under these conditions, Factor 1 contrasted maize stem strength, indicated by a strong positive loading on stem diameter, with leaf and grain size traits (ear leaf area, total leaf area, upper leaf area, and 1,000-kernel weight) all exhibiting negative loadings. Factor 2, the grain production, defined by substantial negative loadings on grain yield and kernels per plant, isolating reproductive output from structural reserves. Factor 3 represented canopy architecture, characterized by negative associations with leaf orientation and leaf area metrics, reflecting variation in leaf angle and spread. Factor 4 was dominated by shelling percentage, underlining harvest efficiency.
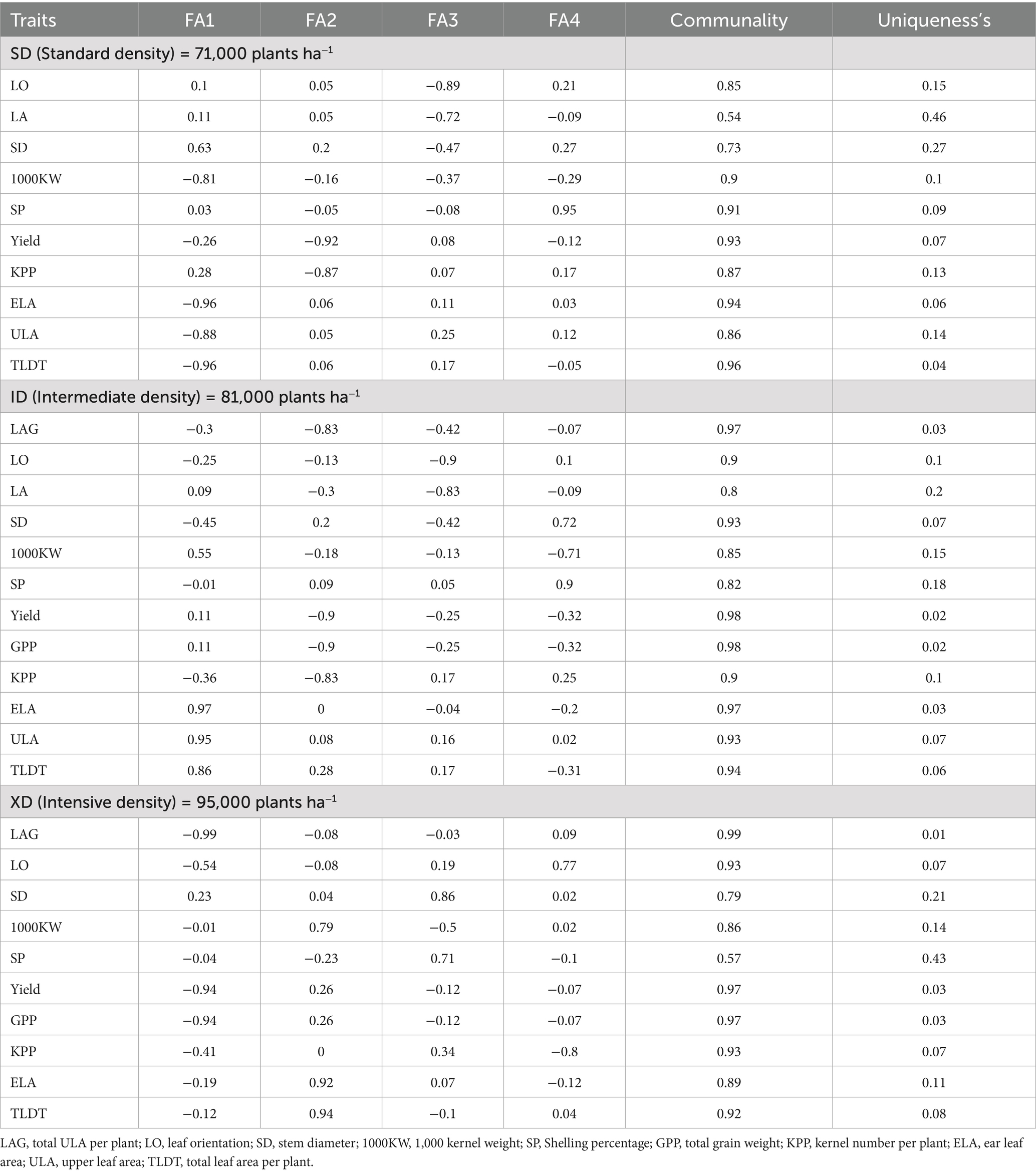
Table 5. Factor loadings and shared variances from the analysis of factor.
At an intermediate density of 81, 000 plants ha−1, the communalities range 0.80 to 0.98, even if with shifted trait loadings. Factor 1 evolved into a leaf area dimension, with very high positive loadings on ear leaf area, upper leaf area, and total leaf area, plus a moderate association with 1,000-kernel weight. This emphasizes canopy size under moderate competition. Factor 2 became a productive capacity, marked by strong negative loadings on upper leaf area, grain yield, total grain weight, and kernels per plant, thus separating genotypes that allocate more to grain set than to foliage. Factor 3 again captured canopy geometry, blending negative leaf orientation and leaf area loadings with moderate positive contributions from stem diameter and upper leaf area. Factor 4 combined shelling percentage and stem diameter versus kernel weight.
Under high density conditions (95,000 plants ha−1), communalities remained high (0.57–0.99), and four factors persisted with further realignments. Factor 1 turned into a source sink gradient, indicating large negative loadings on total upper leaf area and yield traits with moderate negative leaf orientation, thus reflecting the balance between photosynthetic source and grain-filling. Factor 2 highlighted leaf and kernel size, while Factor 3 joined stem diameter and shelling percentage into a structural crop harvest efficiency. Factor 4 contrasted positive leaf orientation against kernels per plant, again reflecting canopy geometry versus sink capacity.
Across the three planting densities, MGIDI-based selection consistently achieved meaningful genetic gains alongside high heritability’s, although both metrics shifted with plant population (Table 6). At the standard density, SG% ranged from modest reductions in canopy-related traits to nearly 6% improvements in yield components, with h2 values between 0.42 and 0.92. At 81,000 plants ha−1, gains narrowed due to source–sink limits, though h2 stayed high (0.21–0.93), supporting reliable SG% inheritance. At 95,000 plants ha−1, reduced canopy size and lower h2 (~0.08) tempered gains and limited selection under crowding.
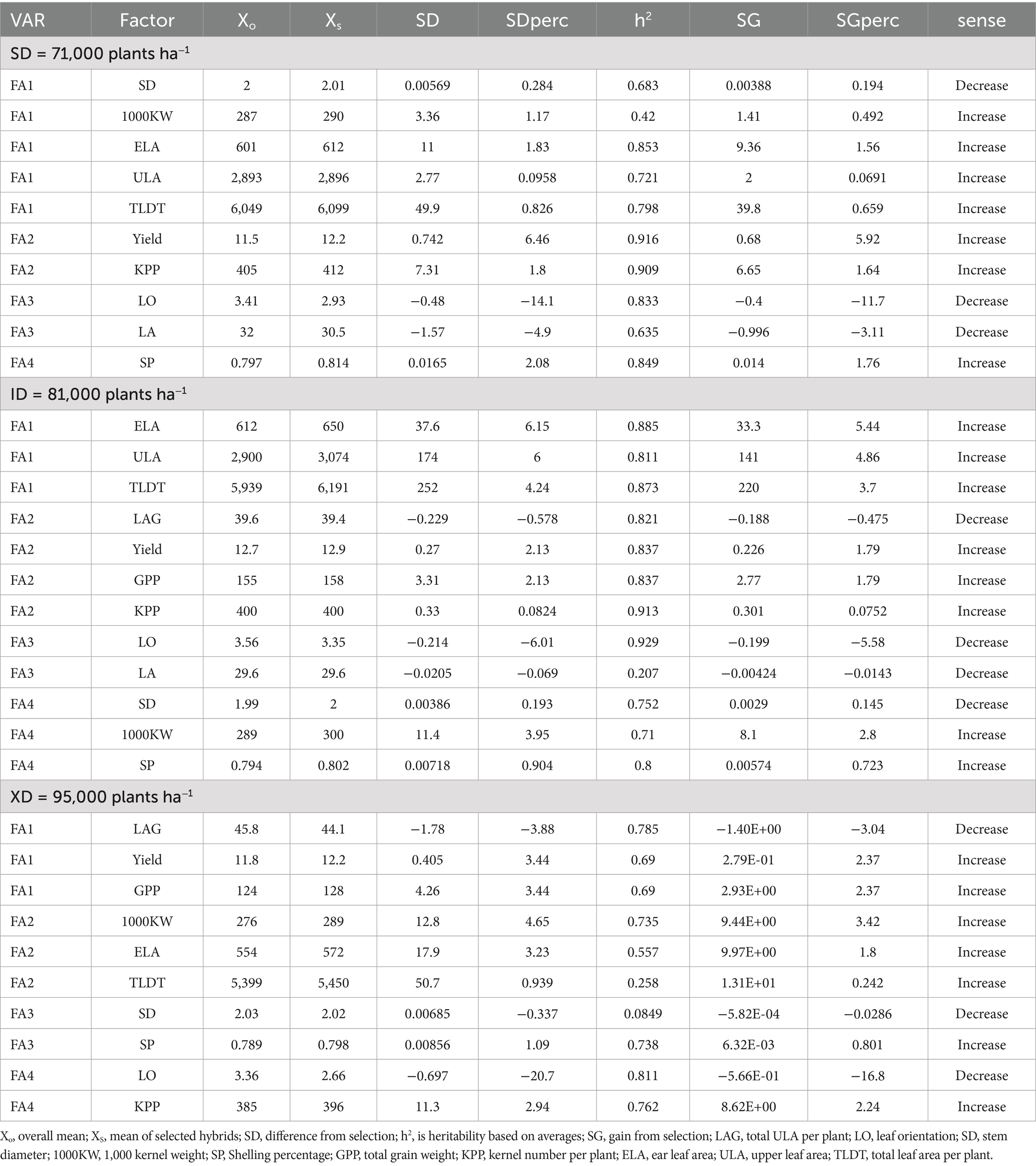
Table 6. Estimated genetic gains for the selected traits based on the MGIDI index.
These trends highlight the dual importance of SG and h2 in density-specific breeding strategies. High h2 for most traits at lower and moderate densities suggests strong potential for cumulative genetic improvement, particularly for leaf-area expansion and yield components. Conversely, the decline in h2 for structural traits at high density warns of diminishing returns when selecting for canopy robustness under competition. By combining SG and h2, the MGIDI index allows breeders to adjust selection by planting density. It emphasizes sink traits where heritability remains high under crowding, or source traits when canopy gene transmission is more reliable, optimizing genetic gains across environments.
Figure 4 shows a visual comparison of genotype rankings, derived from MGIDI index values, across three population density regimes: Distinct genotypes performance under each density level underscores their adaptability to specific environmental pressures, providing actionable insights for breeding and cultivation strategies. At standard density (SD), genotypes G4, G13, and G6 emerged as optimal candidates, combining robust adaptability with advantageous trait profiles under moderate competition. Notably, G4 reappeared as a top performer under intermediate density (ID), alongside G5 and G2, signaling its consistent stability across varying pressures. This recurrence highlights G4’s broad adaptability, suggesting its suitability for environments with fluctuating planting intensities. Under intensive density (XD), where resource competition peaks, genotypes G7, G2, and G5 demonstrated better resilience. The repeated success of G2 and G5 across both ID and XD conditions further underscores their genetic robustness, positioning them as prime candidates for high-stress agricultural systems. The MGIDI index effectively integrates multi-trait performance, streamlining genotype selection by balancing strengths and weaknesses (Olivoto and Nardino, 2021). While this approach efficiently identifies promising genotypes (e.g., G4, G2, G5), environmental factors may constrain the full expression of genetic potential, necessitating field validation to confirm resilience.
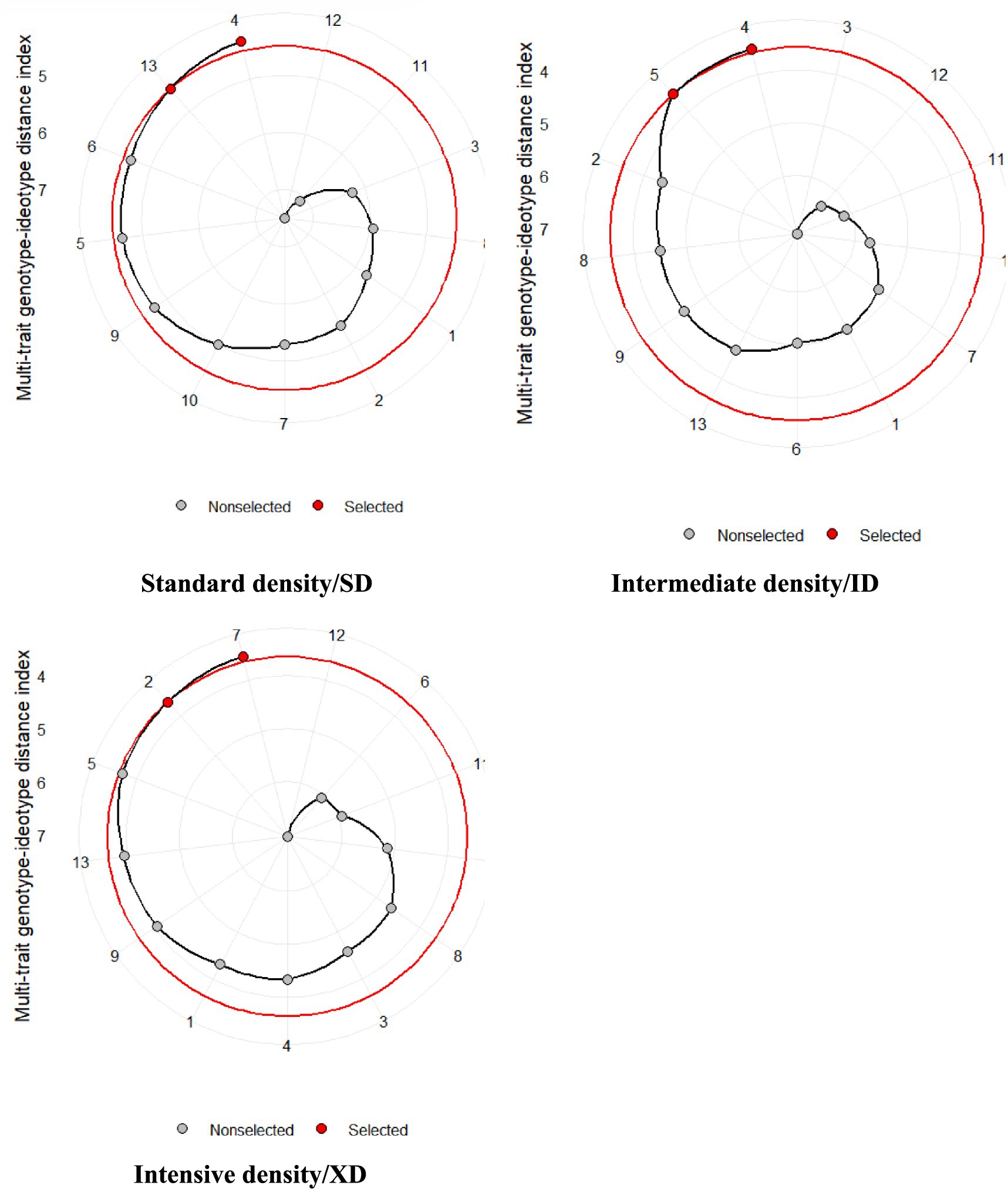
Figure 4. Selected hybrid based on MGIDI index under three population densities.
3.6 Evaluation of the strengths and weaknesses of hybrid
The strengths and weaknesses analysis of selected genotypes under standard population density (Figure 5A) revealed that hybrids G4 and G6 exhibited strong performance in traits related to leaf area growth, grain yield, grain weight per plant, and 1,000-kernel weight. Additionally, hybrids G5, G13, and G4 showed notable strengths in traits related to leaf orientation and kernel number per plant. Furthermore, hybrids G4, G6, and G13 demonstrated proficiency in traits associated with stem diameter and shelling percentage. At intermediate population density (Figure 5B), hybrid G4 continued to dominate in traits related to leaf area growth, grain yield, grain weight per plant, and 1,000-kernel weight, while G4, G5, G8, and G12 exhibited strong associations with stem diameter and shelling percentage. Meanwhile, hybrids G2 and G5 stood out for their strengths in leaf orientation and kernel number per plant. Under intensive density conditions (Figure 5C), hybrids G4 and G5 maintained their superiority in leaf area growth, grain yield, grain weight per plant, and 1,000-kernel weight, while G8 emerged as the leading performer in traits related to stem diameter and shelling percentage. Notably, hybrids G4, G5, and G13 once again demonstrated their adaptability by showing strong associations with leaf orientation and kernel number per plant.
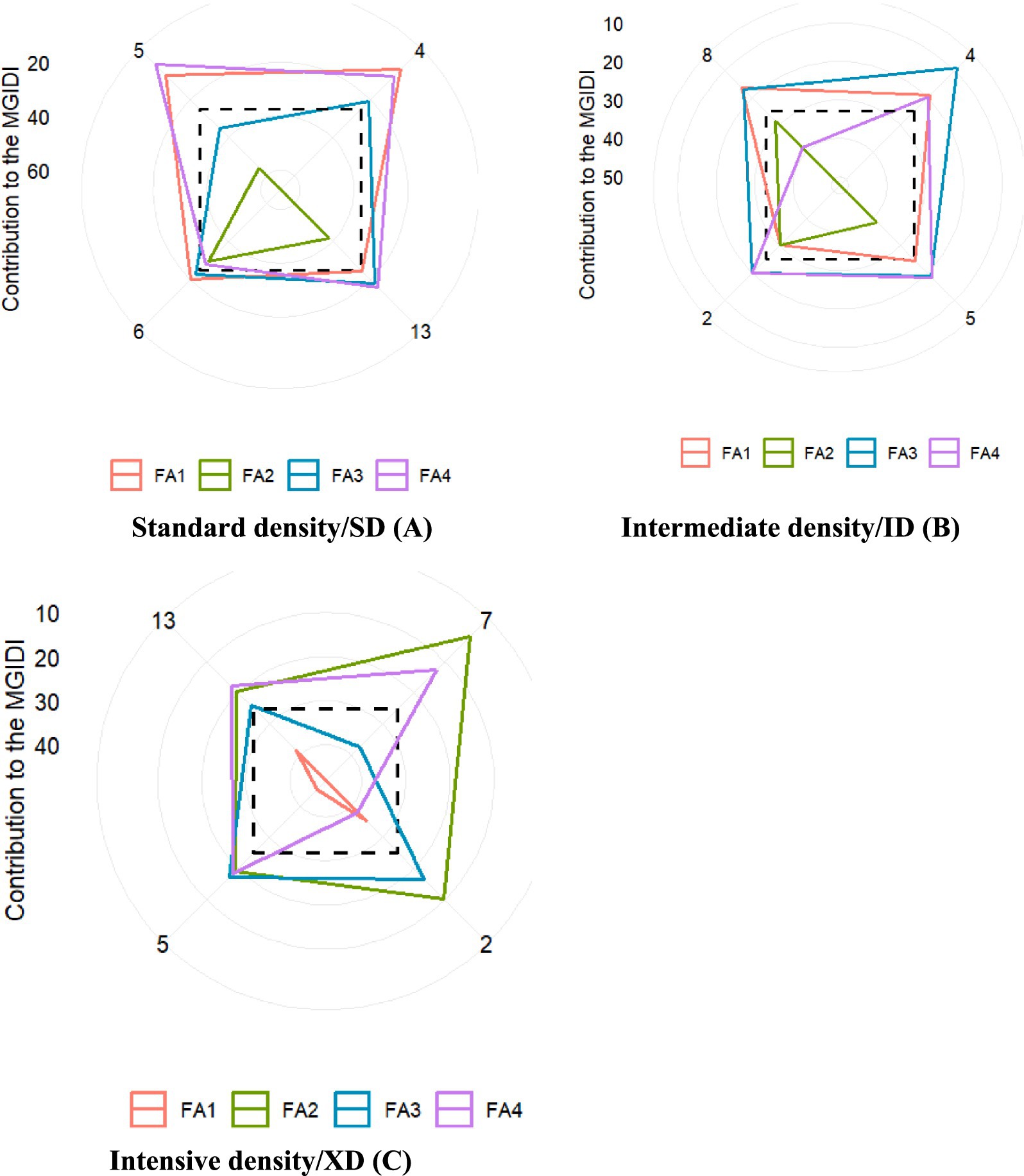
Figure 5. Correlation analysis of morphophysiological traits of maize under three population densities. FA1, ear leaf area /ELA, upper leaf area/ULA, total leaf area per plant /TLDT; FA2: total ULA per plant /LAG, Yield, total grain weight/GPP, kernel number per plant /KPP; FA3: leaf orientation, /LO, leaf area/LA; FA4: stem diameter/SD, 1000 kernel weight/1000KW, Shelling percentage/SP.
4 Discussion
4.1 Genotypic responses to increased planting density
Increasing maize productivity through enhanced planting density remains a critical strategy for optimizing yield per hectare. However, identifying the optimal plant density must consider genetic characteristics of the hybrid and its interaction with environmental factors. Our findings indicated that genotype effects outweighed planting density and genotype × density interaction effects. The variance attributed to hybrid genotype was 1.71-fold greater than that of planting density and 9.41-fold greater than the genotype × density interaction. This emphasizes the importance of hybrid selection directed to specific planting densities, rather than solely adjusting density. Prior research supports an optimal maize population density in the range of 83,000–97,000 plants ha−1 for maximum yield under favorable conditions (Liu et al., 2023). Our results align closely with the intermediate density (81,000 plants ha−1) yielding highest grain yield (12.65 t ha−1). Moderate increases in population density have been reported to significantly enhance grain yield compared to traditional densities. In our study, the intermediate density outperformed both the standard (71,000 ha−1) and the intensive high density (95,000 ha−1) in grain yield. This suggests that increasing density elevates inter-plant competition for light, water, and nutrients, so beyond an optimal threshold the gains in yield per area are negated by stress on individual plants. Over planting densities can detrimentally affect maize plants such as stalk strength and internode diameter that may decrease under crowding, leading to lodging and lower yield (Yang et al., 2022).
4.2 Biological basis of trait interactions
Plant morphology, particularly vertical (erect) leaf orientation allows deeper light penetration into the canopy, ensuring that lower leaves receive more sunlight than they would under a horizontal-leaf hybrid (Kiniry et al., 2004). Vertical leaf angles improve canopy light penetration and photosynthesis, but angles narrower than 10° can reduce light interception by individual leaves, so a moderate upright posture is ideal (Sandhu and Dhillon, 2021). In our experiment at 71,000 plants ha−1, hybrids with larger leaf area sometimes underperformed due to self-shading and higher respiratory costs. Instead, genotypes like G6 could balancing kernel number and kernel weight, thus achieved higher yields. Under intermediate density (81,000 ha−1), hybrids that sustained both kernel set, and grain fill (G6, G4) proved most resilient to crowding stress.
The uniform light distribution increases overall photosynthetic capacity of the stand. However, an optimal architecture must be maintained extremely narrow leaf angles could reduce a leaf’s individual light interception at certain sun angles, indicating diminishing returns if leaves are too erect. A moderate upright posture is therefore ideal to minimize mutual shading without significantly limiting the light capture of each leaf. Prior studies have reported that while a small leaf angle is desirable under high-density stress, extremely small leaf angles (<10°) are not beneficial for productivity, likely due to insufficient light interception by overly vertical leaves (Sandhu and Dhillon, 2021). Our results indicated that G9 hybrid had larger total leaf area yet did not consistently produce the highest yields. This indicates that leaf area alone does not guarantee productivity, and excess leaf area can lead to self-shading and increased respiratory load (Liu et al., 2023). Hybrids G6, which achieved a favorable balance between kernel number (sink size) and kernel weight (sink filling), out-yielded others even if their leaf area was not maximal. Thus, the plants that translated assimilates into grain most effectively were top yielders. As planting density increased to an intermediate level (81,000 ha−1), hybrids capable of maintaining high kernel number without sacrificing kernel weight performed best. High density typically imposes stress that can reduce kernel set or grain filling, but G6 and G4 genotypes showed resilience by sustaining kernel counts and adequate grain weight under crowding. At the highest density. The average yield declined slightly, reflecting the intensified competition and possible stress.
Upper canopy leaves, particularly those at or just above the ear, contribute most to grain yield by capturing light and directly fueling ear development. In contrast, overly long lower leaves receive little light in dense stands, diverting nutrients and screening sunlight from upper leaves. Therefore, an ideal ideotype features large, active upper leaves and reduced lower foliage, with narrow upper angles and slightly broader lower angles to maximize light use and minimize self-shading. Leaf stay-green (the ability to retain chlorophyll during grain filling) extends photosynthesis under dense planting, boosting kernel weight by sustaining assimilate supply. Cooler, greener leaves under heat stress improve radiation use efficiency, as seen in legumes with higher chlorophyll density. Modern breeding has leveraged post-silking photosynthetic duration to enhance yield, so combining stay-green with efficient canopy architecture and robust kernel set yields the greatest gains under high-density stress. Stay-green leaf prolongs chlorophyll retention in dense planting, sustaining photosynthesis and increasing kernel weight (Zaidi et al., 2003). Moreover, the prolonged retention of chlorophyll in elite maize hybrids supports their enhanced photosynthetic efficiency (Zainuddin and Aqil, 2021).
4.3 Genotype × environment interactions and trait heritability
It is essential that G × E interactions broadly can have a profound impact on trait expression and yield stability. The relatively small and non-significant genotype × density interactions in our analysis suggest that hybrids responded almost equally to changes in planting density. In maize, G × E interactions can contribute as much to phenotypic variation as the genotypic effects themselves for key agronomic traits (Hudson et al., 2022). A hybrid that is superior in one environment might lose its advantage in a different environment if its performance is not stable. By testing genotype performance in diverse environmental conditions, breeders can identify cultivars that are not only density-tolerant but also broadly adaptable, exhibiting yield stability despite environmental fluctuations. In our findings, G4, G2, and G5 as top performers across the tested planting densities. Multi-environment trials will help confirm if these hybrids maintain their yield advantage under different climate scenarios. If a hybrid shows low G × E interaction, it would be ideal for large scale recommendation. On the other hand, significant G × E effects might indicate a hybrid is specifically adapted.
An important aspect connected to G × E is trait heritability under varying conditions. The efficiency of selecting and improving a trait depends on how much of its variation is genetically controlled (heritable) versus how much it is due to environment. Our research indicated that many traits including leaf angles, plant height, kernel number, etc. showed clear genetic differences among hybrids. Leaf angle is known to be a highly heritable trait, indicating that it is predominantly governed by genetic factors and can be reliably passed to the next generation. Such high heritability is encouraging for breeders because it means selection for upright leaves would be effective and not easily undermined by environmental variation. Similarly, kernel weight and plant height often exhibit moderate to high heritability in well-managed (Magar et al., 2021), suggesting additive gene action plays a major role in these traits. For instance, heritability of 0.99 for thousand-kernel weight and 0.93 for grain yield under controlled conditions, indicating that substantial genetic gains are possible when environmental noise is minimized. High heritability coupled with high genetic advance is usually indicative of traits governed by additive genes (Alam et al., 2022).
Grain yield is a complex trait integrating many physiological processes, and it tends to have lower stability across environments than a morphological trait like leaf angle. When grown across diverse environments, genotypic variance for yield can be matched or even overshadowed by G × E variance. This means additional approach is needed including genomic selection or multi-environment phenotypic selection to capture stable genetic gains in yield. Traits that maintain high heritability under stress would be prime candidates for selection in breeding programs targeting climate resilience.
Integrating multiple traits for optimal genotype selection, the MGIDI index effectively identified superior hybrids across planting densities. Hybrids G4, G2, and G5 consistently ranked highest; G4 demonstrated broad adaptability at standard and intermediate densities, while G2 and G5 excelled under intensive conditions. Although the MGIDI approach shows great promise, additional field validation across diverse environments is essential to fully confirm genotype stability and performance. Consequently, breeders should prioritize maize hybrids with erect leaf posture, reduced leaf angles, and optimized leaf area for densely planted tropical cropping systems (Sandhu and Dhillon, 2021). Identified key traits include sustained active leaf area duration, essential for maintaining productivity throughout grain filling stages, regardless of environmental variability (Zaidi et al., 2003). Such traits enhance canopy efficiency, increase photosynthetic rates, and contribute significantly to maize yield under dense populations (Szulc et al., 2021). Indeed, strategic selection of leaf ideotypes not only optimizes light interception but also maintains chlorophyll content and manages leaf thermal dynamics, further boosting maize productivity in dense tropical planting systems (Lin et al., 2012).
Despite the deep physiological and yield-component analysis, our study also possesses several limitations, including evaluations at a single site during one season that restrict generalization across different soils, climates, and management practices. Uniform field conditions may have gentle genotype-specific adaptive responses, potentially overestimating the apparent stability of certain hybrids. In addition, the absence of molecular or genomic validation means that observed trait differences cannot be directly linked to underlying genetic factors, reducing the potential for marker-assisted breeding applications. Our findings also did not account for temporal variability such as shifts in performance under early versus late season stress events, which potentially alter genotype rankings. Future work should therefore incorporate multi-location and multi-season trials to capture a broader spectrum of environmental variability, integrate high throughput genotyping to map traits to specific loci, and explore advanced selection frameworks to accommodate nonlinear relationships among traits.
5 Conclusion
Our findings indicate a dynamic relationship between planting density and grain yield in hybrid maize, emphasizing that the optimal density has increased even though individual plant productivity remains unchanged. The trials indicated that both population density and genotype significantly affected yield, whereas the G × D interaction was not significant. Furthermore, intermediate density produced the highest average yield (12.85 t ha−1), outperforming standard (11.54 t ha−1) and intensive (11.79 t ha−1) planting densities. By applying the MGIDI framework, this study identified genotypes G4, G2, and G5 as resilient performers across varying density conditions, with intermediate density offering the highest yield advantage. These findings underline the value of incorporating multi-trait selection tools like MGIDI into breeding pipelines to enhance the development of density tolerant, high yielding tropical maize cultivars. Future breeding efforts should prioritize trait integration and genotype stability across planting intensities.
Data availability statement
The raw datasets supporting this article are available from the corresponding author upon reasonable request (YXpyYWlAYWdyaS51bmhhcy5hYy5pZA==).
Author contributions
MAz: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – review & editing. MAq: Formal analysis, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing. RE: Conceptualization, Formal analysis, Methodology, Software, Validation, Writing – original draft. BZ: Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. ES: Conceptualization, Supervision, Validation, Writing – review & editing. MAn: Formal analysis, software, Validation, Writing – review & editing. MR: Formal analysis, Supervision, Conceptualization, Writing – review & editing. SP: Conceptualization, Data curation, Formal analysis, Software, Writing – original draft. NA: Data curation, Formal analysis, Methodology, Software, Writing – original draft. MY: Conceptualization, Visualization, Supervision, Writing – review & editing. RL: Formal analysis, software, Validation, Writing – review & editing. AS: Conceptualization, Visualization, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by Hasanuddin University Indonesia.
Acknowledgments
We gratefully acknowledge the partial funding support provided by Hasanuddin University and National Research and Innovation Agency (BRIN), Indonesia, which contributed to the completion of this work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
Afifi, M., and Swanton, C. (2011). Maize seed and stem roots differ in response to neighbouring weeds. Weed Res. 51, 442–450. doi: 10.1111/j.1365-3180.2011.00865.x
Alam, M. A., Rahman, M., Ahmed, S., Jahan, N., Khan, M. A., Islam, M. R., et al. (2022). Genetic variation and genotype by environment interaction for agronomic traits in maize (Zea mays L.) hybrids. Plants 11:1522. doi: 10.3390/plants11111522
Aqil, M., Azrai, M., Efendi, R., Andayani, N. N., Zainuddin, B., Damayanti, A. I., et al. (2025). Deep learning-based stacking ensembles for tropical sorghum classification. J. Agric. Food Res. 21:101931. doi: 10.1016/j.jafr.2025.101931
Azrai, M., Aqil, M., Efendi, R., Andayani, N. N., Makkulawu, A. T., Iriany, R. N., et al. (2023). A comparative study on single and multiple trait selections of equatorial grown maize hybrids. Front. Sustain. Food Syst. 7:1185102. doi: 10.3389/fsufs.2023.1185102
Bloom, A. J., and Lancaster, K. M. (2018). Manganese binding to rubisco could drive a photorespiratory pathway that increases the energy efficiency of photosynthesis. Nat Plants 4, 414–422. doi: 10.1038/s41477-018-0191-0
Chibane, N., Caicedo, M., Martinez, S., Marcet, P., Revilla, P., and Ordás, B. (2021). Relationship between delayed leaf senescence (stay-green) and agronomic and physiological characters in maize (Zea mays L.). Agronomy 11:276. doi: 10.3390/agronomy11020276
Coswosk, G. G., Gonçalves, V. M. L., de Lima, V. J., de Souza, G. A. R., Teixeira do Amaral Junior, A., Pereira, M. G., et al. (2024). Utilizing visible band vegetation indices from unmanned aerial vehicle images for maize phenotyping. Remote Sens. 16:3015. doi: 10.3390/rs16163015
Gori, A., Moura, B. B., Sillo, F., Alderotti, F., Pasquini, D., Balestrini, R., et al. (2023). Unveiling resilience mechanisms of Quercus ilex seedlings to severe water stress: changes in non-structural carbohydrates, xylem hydraulic functionality and wood anatomy. Sci. Total Environ. 878:163124. doi: 10.1016/j.scitotenv.2023.163124
Greveniotis, V., Zotis, S., Sioki, E., and Ipsilandis, C. (2019). Field population density effects on field yield and morphological characteristics of maize. Agriculture 9, 1–11. doi: 10.3390/agriculture9070160
Guo, Q., Huang, G., Guo, Y., Zhang, M., Zhou, Y., and Duan, L. (2021). Optimizing irrigation and planting density of spring maize under mulch drip irrigation system in the arid region of Northwest China. Field Crop Res. 266:108141. doi: 10.1016/j.fcr.2021.108141
Hudson, A. I., Odell, S. G., Dubreuil, P., Tixier, M. H., Praud, S., Runcie, D. E., et al. (2022). Analysis of genotype-by-environment interactions in a maize mapping population. G3 12:jkac 013. doi: 10.1093/g3journal/jkac013
Ju, X. T., Xing, G. X., Chen, X. P., Zhang, S. L., Zhang, L. J., Liu, X. J., et al. (2009). Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 106, 3041–3046. doi: 10.1073/pnas.0813417106
Kiniry, J. R., Bean, B., Xie, Y., and Chen, P. Y. (2004). Maize yield potential: critical processes and simulation modeling in a high-yielding environment. Agric. Syst. 82, 45–56. doi: 10.1016/j.agsy.2003.11.006
Lin, Y. S., Medlyn, B. E., and Dewar, D. (2012). Temperature responses of leaf net photosynthesis: the role of component processes. Tree Physiol. 32, 219–231. doi: 10.1093/treephys/tpr141
Liu, Z., Wei, S., Li, M., Zhang, Q., Zong, R., and Li, Q. (2023). Response of water radiation utilization of summer maize to planting density and genotypes in the North China plain. Agronomy 13:68. doi: 10.3390/agronomy13010068
Magar, B. T., Acharya, S., Gyawali, B., Timilsena, K., Upadhayaya, J., and Shrestha, J. (2021). Genetic variability and trait association in maize (Zea mays L.) varieties for growth and yield traits. Heliyon 7:e07939. doi: 10.1016/j.heliyon.2021.e07939
Olivoto, T., and Nardino, M. (2021). MGIDI: toward an effective multivariate selection in biological experiments. Bioinformatics 37, 1383–1389. doi: 10.1093/bioinformatics/btaa981
Page, E. R., Liu, W., Cerrudo, D., Lee, E. A., and Swanton, C. J. (2011). Shade avoidance influences stress tolerance in maize. Weed Sci. 59, 326–334. doi: 10.1614/WS-D-10-00159.1
Sandhu, S., and Dhillon, B. S. (2021). Breeding plant type for adaptation to high plant density in tropical maize—a step towards productivity enhancement. Plant Breed. 140, 509–518. doi: 10.1111/pbr.12949
Shi, Q., Xia, Y., Wang, Q., Lv, K., Yang, H., Cui, L., et al. (2024). Phytochrome B interacts with LIGULELESS1 to control plant architecture and density tolerance in maize. Mol. Plant 17, 1255–1271. doi: 10.1016/j.molp.2024.06.014
Simkin, A. J., Kapoor, L., Doss, C. G. P., Hofmann, T. A., Lawson, T., and Ramamoorthy, S. (2022). The role of photosynthesis-related pigments in light harvesting, photoprotection and enhancement of photosynthetic yield in planta. Photosynth. Res. 152, 23–42. doi: 10.1007/s11120-021-00892-6
Sun, X., He, M., Zhou, B., Li, J., and Chen, N. (2021). Non-structural carbohydrates and C: N: P stoichiometry of roots, stems, and leaves of Zygophyllum xanthoxylon in response to xeric condition. Arid Land Geography 44, 240–249. doi: 10.12118/j.issn.1000-6060.2021.01.25
Szulc, P., Bocianowski, J., Nowosad, K., Zielewicz, W., and Kozak, J. K. (2021). SPAD leaf greenness index: green mass yield indicator of maize (Zea mays L.), genetic and agriculture practice relationship. Plants 10:830. doi: 10.3390/plants10050830
Tilman, D., Balzer, C., Hill, J., and Befort, B. L. (2011). Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 108, 20260–20264. doi: 10.1073/pnas.1116437108
Yang, J., Geng, W., Zhang, J., Ren, B., and Wang, L. (2022). Responses of the lodging resistance of summer maize with different gene types to plant density. Agronomy 12:10. doi: 10.3390/agronomy12010010
Yu, H. Q., Sun, F. A., Lu, F. Z., Feng, W. Q., Zhang, Y. Y., Liu, B. L., et al. (2018) Positive regulation of phytochrome a on shade avoidance in maize Pak. J. Bot. 50 1433–1440. Available online at: https://www.scopus.com/inward/record.uri?eid=2-s2.0-85046137665
Yue, H., Wei, J., Xie, J., Chen, S., Peng, H., Cao, H., et al. (2022). A study on genotype-by-environment interaction analysis for agronomic traits of maize genotypes across Huang-Huai-Hai region in China. Phyton (Buenos Aires) 91, 57–81. doi: 10.32604/phyton.2022.017308
Zaidi, P. H., Srinivasan, G., and Sanchez, C. (2003). Morpho-physiological traits associated with variable field performance of different maize germplasm across multiple environments. Maydica, 48, 207–220. Available online at: https://www.scopus.com/inward/record.uri?eid=2-s2.0-0347659442
Zainuddin, B., and Aqil, M. (2021). Analysis of the relationship between leaf color spectrum and soil plant analysis development. IOP Conf. Ser. Earth Environ. Sci. 911:012045. doi: 10.1088/1755-1315/911/1/012045
Keywords: population density, hybrid, yield, MGIDI, maize
Citation: Azrai M, Aqil M, Efendi R, Zainuddin B, Syam'un E, Anshori MF, Riadi M, Priyanto SB, Andayani NN, Yasin M, Laurenze R and Syam A (2025) Canopy architectural and physiological traits for optimizing maize yield under high planting density. Front. Sustain. Food Syst. 9:1469305. doi: 10.3389/fsufs.2025.1469305
Edited by:
Mohamed Ait-El-Mokhtar, University of Hassan II Casablanca, MoroccoReviewed by:
Eastonce Tendayi Gwata, University of Venda, South AfricaR. Yashaswini, University of Agricultural Sciences Raichur, India
Copyright © 2025 Azrai, Aqil, Efendi, Zainuddin, Syam’un, Anshori, Riadi, Priyanto, Andayani, Yasin, Laurenze and Syam. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Muhammad Azrai, YXpyYWlAYWdyaS51bmhhcy5hYy5pZA==