 Pablo Cárdenas-Camacho1
Pablo Cárdenas-Camacho1 Catalina Fernández-Díaz
Catalina Fernández-Díaz- 1IFAPA Centro El Toruño, Andalusia Research and Training Institute for Fisheries and Agriculture, Cádiz, Spain
- 2Associate Research Unit “Blue Growth” CSIC-IFAPA, Cádiz, Spain
Mysids have a great potential as the new high nutritional, sustainable and functional natural resources aquaculture require globally. Despite being an excellent source of proteins and essential fatty acids, their potential in aquaculture has barely been evaluated. We have characterized the nutritional value of Mesopodopsis slabberi in the Guadalquivir estuary, where constitutes a key prey for fish juveniles. Seasonal and ontogenic biochemical composition variation was studied. M. slabberi shows high protein per dry mass (60%) with a slight a significant (p < 0.05) decline in summer. Glutamic +glutamine, lysine and aspartic +asparagine were the dominant aminoacids, with relatively high levels of leucine, alanine, glycine, methionine, phenylalanine and tryptophane, all of great relevance for aquaculture feeds. Total lipids content is high in M. slabberi, representing PUFAs 44.15 ± 1.12% of the total fatty acids. The highest concentrations of EPA and DHA fatty acids were found in spring. Most important the ratio DHA/EPA was high and similar in all season with an average value of 0.83 ± 0.03. DHA/AA and EPA/AA ratio were 4.92 ± 0.38 and 6.10 ± 0.48, respectively. Highest n3/n6 ratio (p < 0.05) were detected in spring 7.83 ± 0.77. M. slabberi show an adequate antioxidant balance with TAC/LP ratio greater than 90 in spring and summer reaching the highest levels (132) in autumn. In Guadalquivir estuary, M. slabberi presents high zinc and selenium levels. Our results demonstrate how M. slabberi has high nutritional profile and potential to be used as a valuable functional ingredient in aquaculture, both inert and as a living resource. More research in relation to culture technology and functional assess is necessary to open new ways to reach aquaculture long-term sustainability.
1 Introduction
Aquaculture is one of the fastest-growing food-producing industries in the world and it is actually demanding alternative high-quality feed ingredients to support its sustainable growth (Eroldoğan et al., 2023). Blue Foods (fish and other aquatic foods) have become a strong perspective on finding new critical nutritional resources and ecologically sustainable to a growing world population (Hoegh-Guldberg et al., 2019; Willett et al., 2019; Ziegler et al., 2019) according to Sustainable Development Goals (SDG) set by FAO (2020). Aquaculture challenge is to replace fishmeal and fish oil (FAO, 2020) in aquafeed industry, providing diets that incorporate ingredients or functional additives to improve the health status of aquatic organisms (Copeman et al., 2009) and product quality. To achieve this goal is necessary to find and produce new nutritionally functional, cheaper and efficient live foods sources of marine protein n-3 LC-PUFA, among others- more sustainable and successfully than those currently used (Hoegh-Guldberg et al., 2019; Phillips et al., 2019).
A recent review about aquafeed industry current status (Eroldoğan et al., 2023) highlights the importance of providing new functional feed alternatives by two main lines of innovation: (1) addressing the potential of circular aquaculture bio-economy by the re-use of biological resources (both directly by valorization of waste and indirectly by integrated multitrophic aquaculture IMTA) and (2) innovation on the potential use of marine organisms-from microorganisms to macroalgae and macroinvertebrates in novel aquafeeds as alternative nutrient sources and more importantly as promoters of growth and health.
An example of circular aquaculture with macroinvertebrates is the utilization of shrimp fisheries remains to add as ingredient of alternative feeds for other shrimp or fish aquaculture, showing in general substantial amounts of key nutrients and high nutrient bioavailability and digestibility, good growth performance, adequate amino acid profiles and good levels of macro- and micro minerals to meet dietary requirements (Fox et al., 1994; Yang et al., 2009; Terrazas-Fierro et al., 2010; Salas-Leiton et al., 2020; Fricke et al., 2022).
The present works is focused on the second innovation line exposed and on invertebrates. Despite the fact that marine invertebrates have long been used as live feed or additives, only for a reduced group culturing protocols, nutritional valorization and suitability have been evaluated. As natural live-preys, brachiopod Artemia sp. and rotifers have been long used for the culture of fish larvae worldwide (Hernandez-Alarcon, 2016). Unfortunately, they do not always satisfy the nutritional requirements of cultured fish species during their different stages of development, due to the unfavorable biochemical composition of Artemia, requiring their enrichment before using them as first feeds (Conceição et al., 2010; Planas et al., 2020; Eroldoğan et al., 2023) and making necessary to explore new more nutritional alternatives to overcome the lack of adequate functional foods and contribute to the diversification of the species cultivated in aquaculture (Southgate, 2019).
In the search of better alternatives at lower trophic levels invertebrates, the potential use of other crustaceans like copepods and amphipods has been explored, proposing them as high-quality alternative feed live resources due to their nutritional value and high content of PUFAs, proteins and essential amino acids (Næss et al., 1995; Baeza-Rojano et al., 2010; Jiménez-Prada et al., 2018; Alberts-Hubatsch et al., 2019). Both offer good DHA/EPA ratios, and DHA + EPA/total fatty acids ratios, closer to those needed by fish than those of Artemia and rotifers, which make them more easily digestible for fish larvae and with a higher nutritional and functional value for fish (Eroldoğan et al., 2023) and have been successfully tested as live feed for seahorses (Planas et al., 2020; Vargas-Abúndez et al., 2021) and cephalopods (Baeza-Rojano et al., 2010). DHA and EPA fatty acids are main structural components of cellular membranes and essentials for marine fish larvae development (Glencross, 2009).
Though mysid shrimps -abundant in nature and an essential natural prey for many fish and invertebrate species, playing a key role in estuaries and coastal ecosystems food webs.
Azeiteiro et al. (2001), Vilas et al. (2008), and Biju et al. (2009) represent an important source of high-quality nutrients (Guevara et al., 2005), they have not been used widespread or considered as alternative functional feed. Mysids have not even been mentioned in the vast recent review of Eroldoğan et al. (2023) on invertebrates as alternative aqua feeds.
Andam et al. (2024) have recently demonstrated that even 100% fishmeal substitution by mysid meal (Mesopodopsis sp.) is possible in the Pacific white shrimp (Penaeus vannamei) postlarvae diet, finding an optimum 65.50% of mysid meal replacement to attain maximum growth. Other studies found mysid shrimps are naturally rich in lipids, including EPA and DHA (Dalsgaard et al., 2003; Fink, 2013; Griffin et al., 2020), similar to those found in Antarctic krill (Liu et al., 2018). Despite its potential, mysid culture may not turn out to be optimized (Guevara et al., 2005; Eusebio et al., 2010; Herrera et al., 2011). Up to date, most attempts to culture mysids shrimps have been performed under laboratory conditions with the purpose to produce model organisms for toxicological studies (Sardo et al., 2005; Verlecar et al., 2012; Lentes, 2018; Min et al., 2018). Overall, the optimal mysid culture conditions are still not defined and further research is needed to optimize the culturing techniques, taking into consideration the available and uploaded knowledge on mysid ecology, biology, feeding patterns and reproduction. A better understanding of mysid contribution as value-added products and their ability to improve animal performance, nutrient availability, palatability, and feed digestibility is needed to successfully integrate them as vital resources in aquaculture feeds.
Improving the health status of animals in culture environments becomes one of the main premises to take into account when looking for and incorporating new potentially functional foods, either as raw material or as live feeds. The evaluation and optimization of the antioxidant status of mysid shrimps as a potential functional food would allow a better understanding of mysid contribution as value-added products and their ability to improve animal performance.
The study of metal contents of M. slabberi is a valuable field of research, which is indispensable for both safety assessment as well as knowledge of their nutritional value. The capacity of mysid to accumulate metals depends on a variety of factors such as location, temperature, salinity, light, pH, nitrogen availability, season and metabolic processes.
Mysid Mesopodopsis slabberi is a common and abundant component of estuary and coastal ecosystems from North Atlantic to Mediterranean Sea (Biju et al., 2009) where plays a key ecological role, especially for nursery function and also as a main prey for nekton fish and shrimp species, making a substantial contribution to the stock of numerous organisms (Azeiteiro et al., 2001; Azeiteiro et al., 2003). These species present an opportunistic and omnivore feeding behavior, that feed on phytoplankton, algae, detritus, and organic matter from water and sediments which in turn form food for animals at higher trophic levels (Mauchline, 1980). Thus, mysids may contribute to the transfer of trace metals to higher trophic levels due to their significant capacity to bioaccumulate metals by direct absorption from water and assimilation through food substances (Biju and George, 2021).
In the Guadalquivir River estuary (Southern Spain), an important nursery area for many Gulf of Cádiz marine ecosystem species, M. slabberi contribute the most to total plankton and nekton biomass, but no information is available on its biochemical composition and antioxidant status. M. slabberi also couples benthic with pelagic realm and channels energy from low to high trophic levels (Vilas et al., 2008). Because it is exposed to continuous physicochemical changes and anthropogenic pressures in the estuary, mysids develop a series of adaptive strategies against the effects of stress on their biological status (Smith et al., 2003) which causes the oxidation of macromolecules and deregulating homeostatic processes (Gutteridge and Halliwell, 2018).
As a key trophic organism and source of basal nutrients in fish and shrimp diet, M. slabberi could offer a considerable contribution of elemental and bioactive compounds to diet. The present study characterizes M. slabberi biochemical composition present in the Guadalquivir Estuary and evaluates the influence of seasonality on nutritional profile and antioxidant status in order to know their potential to be used as aquafeed.
2 Materials and methods
2.1 Description of sampling procedures
Samples were collected seasonally from spring 2019 to fall 2020 at the Guadalquivir River from a wharf at Trebujena (Cádiz, Spain) (36°89′95”N-6°23′11”W), located in the mesohaline region of the Guadalquivir River estuary. Water physical–chemical parameters and measurements of weight and length of adult and juvenile individuals were registered (Table 1). In the present work there is no record of winter data because the population of M. slabberi falls exponentially during the colder months and it is not possible to obtain enough biomass during sampling. M. slabberi specimens were collected using plankton net of 40 cm mouth and 150 cm length with a mesh size of 500 μm. Collected organisms were transported in 25 L containers fulfilled of filtered in situ estuarine water with an oxygen pumping system. At the lab mysids were separated from other organisms by trawling in an aquarium a 1 mm piece of plastic net slowly -to minimize possible damage during handling- and introduced into 15 L aquariums with seawater filtered (at same salinity than collected) until unwanted material which passed beyond mesh size were removed from the bottom of aquarium to prevent contamination of samples. Individuals were separated into each ontogenic stages Mauchline (1980) and stored at −80°C until further biochemical analysis.
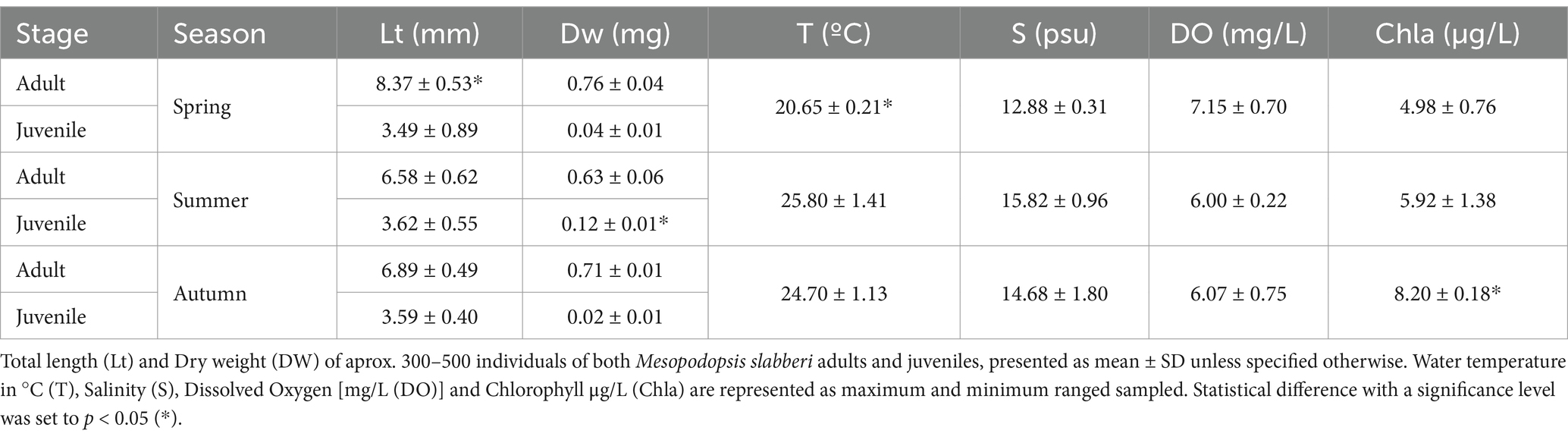
Table 1. Physical–chemical water properties at sampling site Trebujena dock at the Guadalquivir estuary through study period spring 2019 to autumn 2020.
2.2 Biochemical analyses
2.2.1 Total protein and amino acid composition
Total protein analyses were determined using the Lowry method (Lowry et al., 1951) increasing the concentration of NaOH (1.5 M) and heating at 100°C for 60 min. Aminoacids were extracted by an acidic and basic hydrolysis to release individual amino acids from peptides and proteins. The corresponding derivatives N-ethoxycarbonylethyl esters has been determined by GC–MS through 2 acid hydrolysis, using 6 M HCl in an atmosphere of N2 for 70 min at 150 ° C, and on the other, using 4.2 M NaOH in an atmosphere with N2 for 24 h at 110 ° C. A 1 μL aliquot of the derived extracts was injected in Splitless injection port of a gas chromatograph coupled to a triple mass spectrometer, Shimadzu GC–MS (GCMS-TQ8030), equipped with an Agilent HP-5MS fused silica capillary column (30 m x 0.25 mm i.d., 0.25 μm film 150 thickness), with a temperature ramp from 40°C for 5 min to 270/152°C for 20 min by increasing the temperature at 5°C min−1. Identification of derivative amino acids were achieved comparing the gas 156 chromatograph retention times and mass spectra with those of the pure standard compounds. All mass spectra were also compared with the data system library (NIST 11).
2.2.2 Carbohydrates
To estimate the total carbohydrate content, we used the methodology proposed by DuBois et al. (1956) adding 100 microliters of homogenised samples (5 mg/mL) were included in 900 microliters, 25 microliters of phenol (81%) and 2.5 mL of sulphuric acid (95–98%), shaken with vortex and maintained in darkness for 30 min, after this time, absorbance was measured at 485 nm following Kochert’s techniques (Kochert, 1978).
2.2.3 Ash content
The ash content was gravimetrically determined after burning up at 550 ° C for 4 h in a muffle furnace.
2.2.4 Total lipids and fatty acid content
Total lipids (TL) extraction was performed following the methodology proposed by Cañavate et al. (2019) using chloroform:methanol (2:1 v/v) containing 0.01% of butylated hydroxytoluene (BHT) as antioxidant. The organic solvent was evaporated under a stream of nitrogen and the lipid content was determined gravimetrically. TL extracts were subjected to acid catalyzed transmethylation for 16 h at 50°C, using 1 mL of toluene and 2 mL of 1% sulphuric acid (v/v) in methanol (Christie, 2003). The resultant FA methyl esters (FAME) and dimethylacetals (DMA) formed from ether lipids were purified by thin layer chromatography (TLC) and visualized by spraying with 1% (w/v) iodine in CHCl3 (Christie, 2003).
Fatty acid methyl esters (FAME) were separated in n-hexane and quantified by using a Shimadzu GC 2010-Plus gas chromatograph equipped with a flame ionization detector (280°C) and a fused silica capillary column SUPRAWAX-280 (15 m × 0.1 mm I.D.)Hydrogen was used as carrier gas and the oven initial temperature was 100°C for 0.5 min., followed by an increase at a rate of 20°C min − 1 to a final temperature of 250°C for 8 min. Individual FAME and DMA were identified by reference to authentic standards and to well-characterized fish oil. Fatty acid results were expressed both as μg fatty acids mg−1 dry weight and % of total fatty acids (Jiménez-Prada et al., 2018; Cañavate et al., 2019).
2.3 Determination of antioxidant status of M. slabberi
The total antioxidant capacity (TAC) of M. slabberi, expressed as Equivalent trolox (mM), was evaluated using the AIOR assay method as previously reported (Ching et al., 2006) and as described (Jiménez-Fernández et al., 2015). Lipid peroxidation (LP) expressed as malondialdehyde (MDA), was measured as described in Canavate et al. (2007) and Jiménez-Fernández et al. (2015). Ratio TAC/LP was estimated using TAC (μgTrolox Equivalent mgDW−1)/ LP (μgMDA mgDW−1).
2.4 Determination of macro and microminerals
Samples were weighed on a precision balance and oven dried at 60°C for 24 h. Metal analysis was carried out by the CSIC–ICMAN laboratory (Puerto Real, Cadiz) by analytical platforms based on elemental (inductively coupled plasma mass spectrometry, ICP-MS) and molecular (quadrupole time-of-flight mass spectrometry, QTOF-MS) mass spectrometry coupled to different chromatographic devices. This instrumental arrangement is based on a two-dimensional chromatographic device with ICP-MS detection to trace metal-tagged biomolecules. Generally, in the first-dimension metal-biomolecules are separated by the size using size-exclusion chromatography (SEC-ICP-MS) followed by the purification of the selected fraction by anion- or cation-exchange chromatography with ICP-MS detection (AEC/ CEC-ICP-MS). A total of eighteen metals (macroelements, essential and trace metals as well as heavy metals) were analyzed.
2.5 Statistical procedures
Before statistical analysis, all data were checked for normality using a Kolmogorov–Smirnov test and homogeneity of variance using Bartlett’s test and, when necessary, they were log-transformed before further statistical analysis. The results given in percentage values were arcsine transformed. Biochemical data are presented as mean ± SD. Fatty acids results obtained as a percentage of fatty acids were transformed using the internal standard 23:0 as a reference and as a function of the dry weight of the sample regarding to the total heavy lipids. Significant differences in function of development stage and season were analysed by a two-way ANOVA followed by Tukey’s post hoc analysis for pairwise comparisons when applicable using IBM SSPS 25 Software. Finally, we examined Pearson correlations between Total Antioxidant Capacity and Lipid Peroxidation among seasons and Pearson correlation between physical–chemical parameters and mineral composition. The significance level was set to p < 0.05.
3 Results
3.1 Water physical–chemical characteristics and measure sizes
During the study period temperature followed typical temperate seasonal pattern showing highest temperatures during summer and autumn (24–26°C), while lowest temperature was found during spring (20–24°C) (p < 0.05). Salinity remained relatively stable in this mesohaline estuary water mass throughout the entire study period (12.6–17.6 on average), without significant seasonal variations (p-value<0.05). Dissolved oxygen (DO) remained relatively stable throughout the study period (5.5–7.6 mg/L average; p > 0.05) (Table 1).
The average dry weight of the adult M. slabberi did not vary significantly throughout the study period (p < 0.05), remaining between 0.6 to 0.7 mg, while in the case of juvenile’s significant variations were observed, being the juvenile individuals captured in summer the heavier (0.12 mg). On the other hand, the average length of adult individuals was greater during spring (8.37 ± 0.53 mm), while juveniles maintained a similar length during the study period (3.5 mm) on average.
3.2 Biochemical composition
M. slabberi is characterized by high levels of total proteins (56–61%) and total lipids (13–16%) and lower levels of carbohydrates (1.7–2.6%) (Table 2). As no significant differences were detected between the elemental composition or the antioxidant status between adults and juveniles within each season of the year sampled, the data are shown as a whole for the entire study period. Significant differences were observed among seasons (p < 0.05), being summer the season with lowest significant total proteins values (56.01 ± 2.35%) (p < 0.05). Significant differences were found for carbohydrates, total lipids and ashes during autumn, with a decrease in carbohydrate and lipids content and an increase in ash values (p > 0.05). Aminoacids composition was only analysed for spring and summer are shown in Table 3. On average, aminoacids did not show significant differences among seasons, highlighting Lysine (12.40 ± 0.00%), Leucine (8.20 to 9.20%) and Methionine (6.0 to 7.6%) were the most abundant essential amino acids (EAA). Non-essential amino acids Glutamic Acid (GA) (18.0% on average) and Aspartic Acid (SPA) (10.5 to 11.4%) were found in the highest proportion.
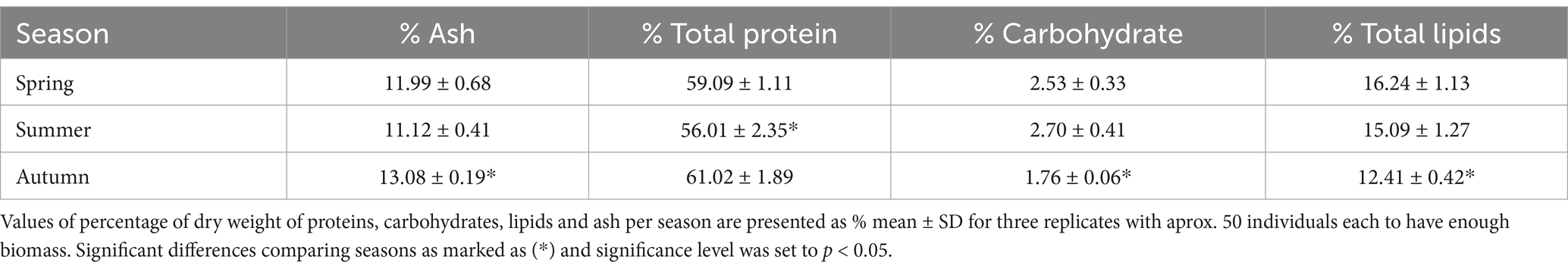
Table 2. Averaged elemental composition of Mesopodopsis slabberi for the study period 2019 to 2020 by season.
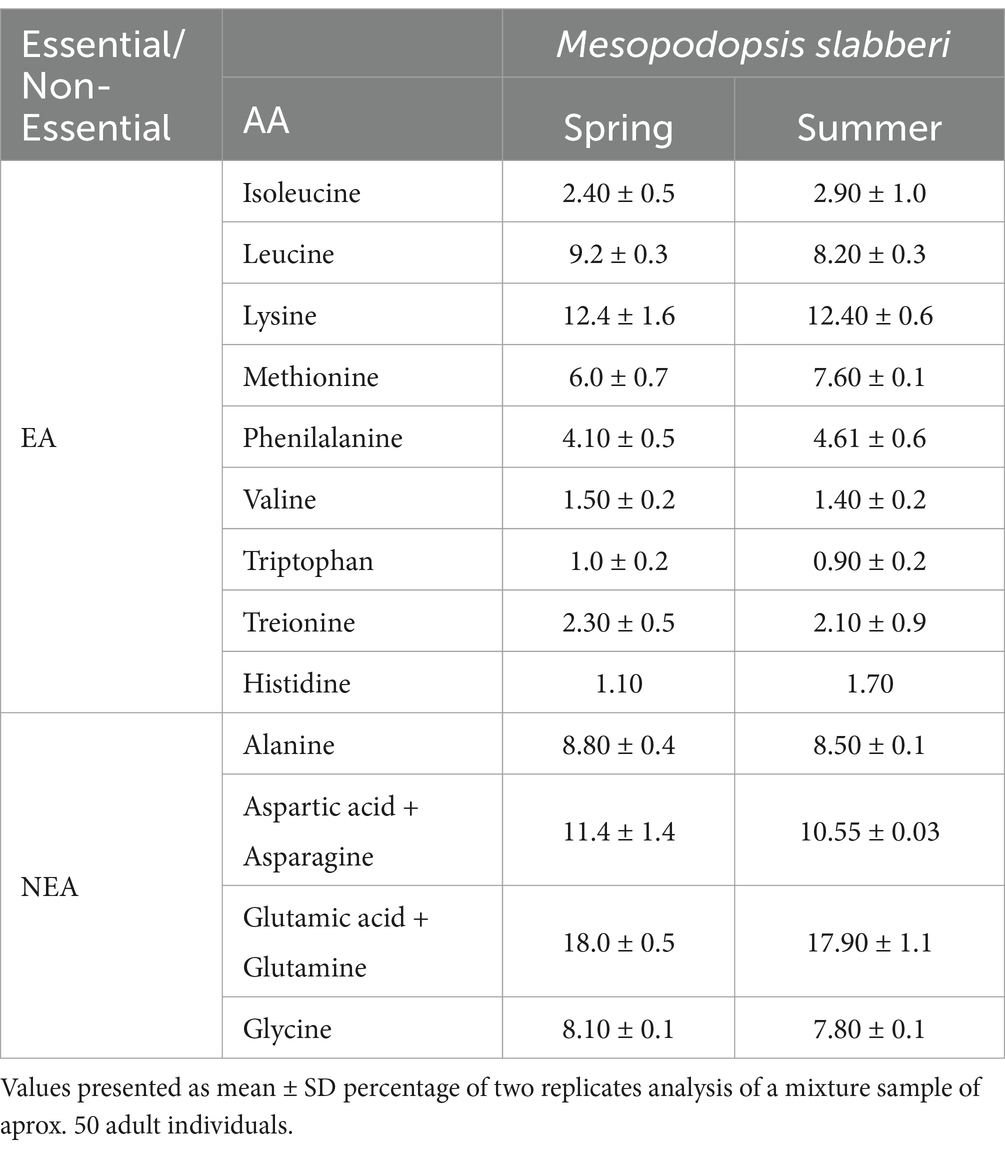
Table 3. Essential (EA) and non-essential (NEA) amino acids expressed as concentration (%) of identified amino acids of the Mesopodopsis slabberi.
Fatty acid profile of Mesopodopsis slabberi by seasons is shown in Table 4. Polyunsaturated fatty acids (PUFA) and saturated fatty acids (SFA) were the most abundant in proportion. Among monounsaturated fatty acids (MUFA), found in a lesser extent than saturated and polyunsaturated fatty acids, 18:1n-9, 18:1n-7 and 16:1n-7 were present at highest concentration, being 18:1n-9 and 18:1n-7 significantly higher during autumn (p < 0.05). It is worth mentioning how ubiquitous saturated 16:0 fatty acid was the most abundant, followed by polyunsaturated 20:5n-3 EPA and 22:6n-3 DHA. The content of EPA observed ranges from average 7.62 to 8.91 μg FA mg DW−1, the content of DHA were detected in a range of 5.61 to 7.10 μg FA mgDW−1, while arachidonic acid AA showed values between 0.79 to 1.81 μg FA mgDW−1. PERMANOVA analysis showed seasonal differences in essential PUFA fatty acid composition (p < 0.05), finding that autumn season present the lowest PUFA total fatty acid composition (40.60 ± 1.01%), in comparison to spring (47.2 ± 2.72%) and summer (44.80 ± 2.14%), 20:4n-6 (F = 3.450, p = 0.048), 20:5n-3 (F = 9.971, p = 0.001) and 22:6n-3 (F = 6.379, p = 0.006).
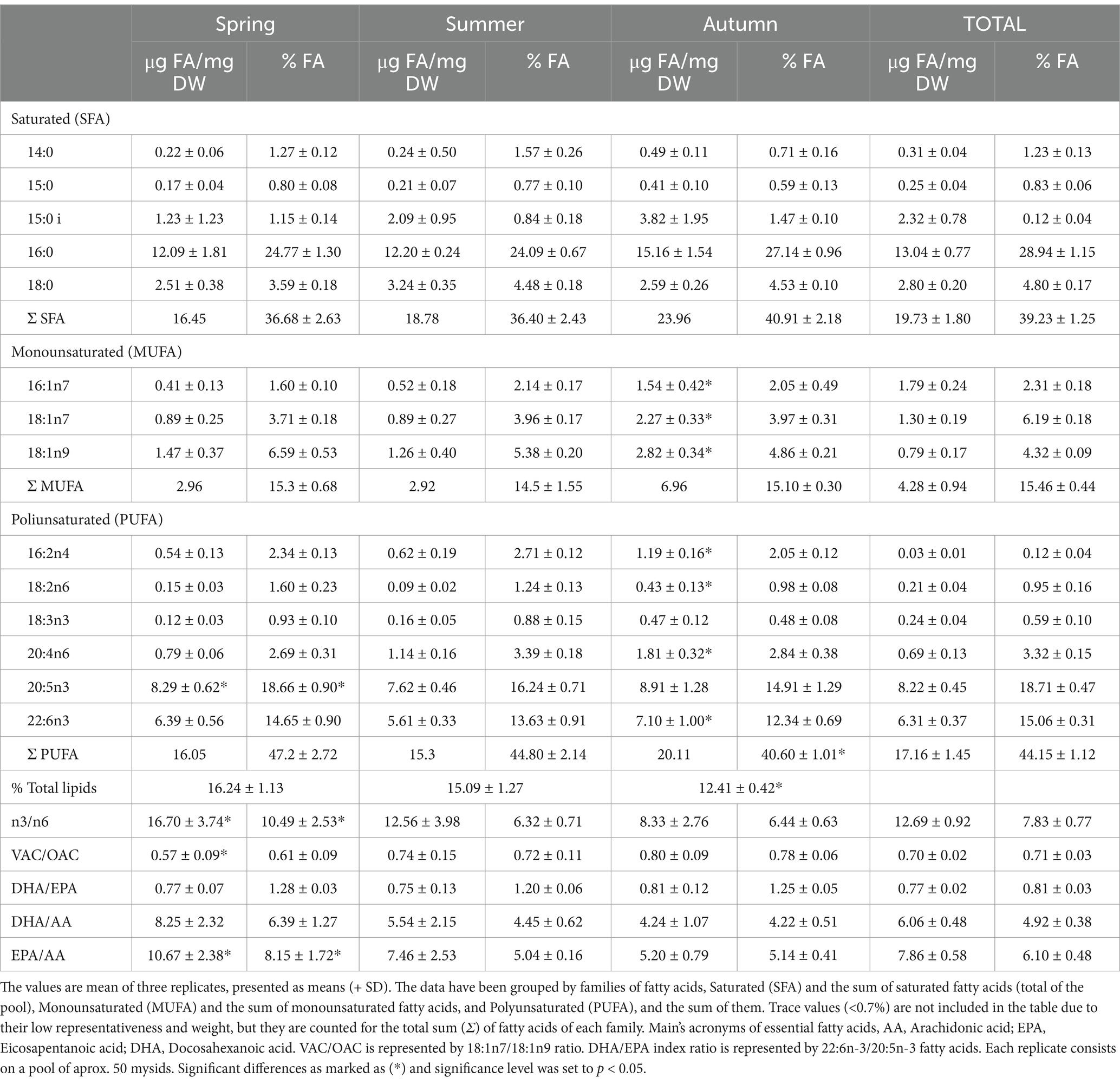
Table 4. Mesopodopsis slabberi fatty acids composition (μg/mg of DW and % of Total FA) averaged per seasons for the study period spring 2019 to autumn 2020 at the Guadalquivir estuary.
PUFA n3/n6 ratio represents the relationship between the sum of the n-3 and n-6 PUFA, which presented significative highest ratios observed during the spring (16.70 ± 3.74), and we were able to observe that there was a decrease in the ratios throughout the seasons, finding the lowest n3/n6 ratio values during the autumn (8.33 ± 2.76) (Table 4). The relationship given by the DHA/EPA ratio oscillates between 0.75 and 0.81 without observe differences among seasons (p > 0.05). Fatty acid VAC/OAC ratio proposed by Magnone et al. (2015), represented by 18:1n7/18:1n9, was found ranged from 0.6 to 0.9, finding lowest VAC/OAC ratios during spring period. The relationship given by the DHA/EPA ratio did not show a seasonal variability during study period (0.75 to 0.81). DHA/AA did not present a seasonal differentiation in their ratios observed, while, Fatty acid EPA/AA ratio marks significative higher ratio during spring (10.67 ± 2.38), than summer and autumn (p < 0.05).
Concentration of macroelements, essential and trace metal and heavy metal found in M. slabberi tissues are showed in Table 5. Similar values of different metal analyzed were detected in M. slabberi during the sampling period without statistical difference due to season (p > 0.05). Zn was found at highest concentration, ranging widely from 84.52 to 89.79 ug/g, without observing a seasonal variability during study period (p > 0.05). The lowest iron values found in M. slabberi were observed in summer (43.36 ± 4.81 ug/g) and highest in spring (52.92 ± 27.33 ug/g) and autumn (70.45 ± 26.05 ug/g), although no significant differences were detected due to seasonality for this metal. M. slabberi contains high values of selenium (4–6 ug/g), observing a seasonal variability with lower levels during spring.
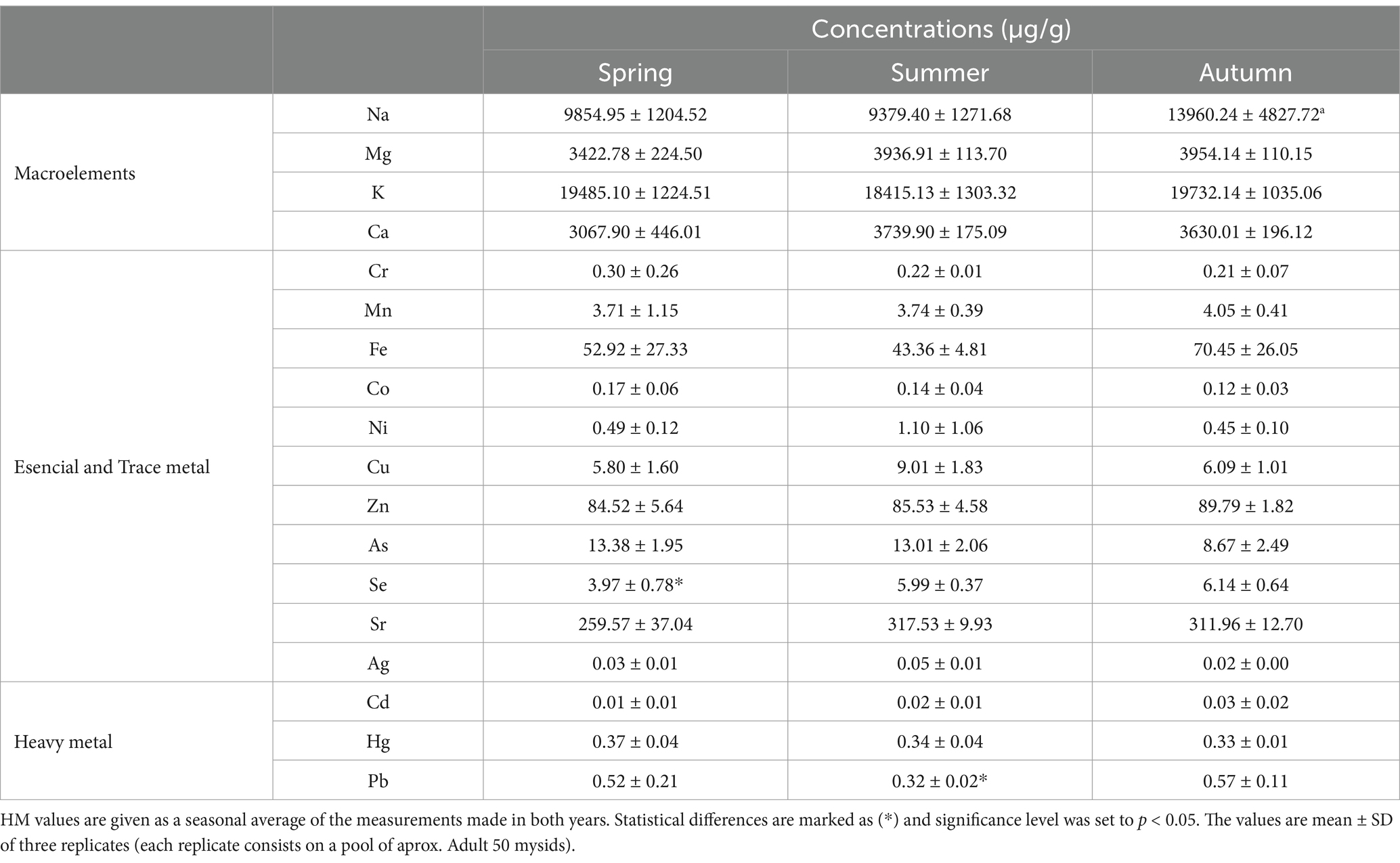
Table 5. Principal macronutrient, trace metal and heavy metal (μg/g) concentrations in Mesopodopsis slabberi tissue analyzed from each season, spring–summer-autumn of the 2 years study period 2019–20 in the Guadalquivir River estuary.
3.3 Antioxidant status
Antioxidant status of M. slabbery was enhanced as determined by TAC and MDA content (Table 6). A significant drop in MDA levels was detected in autumn (3.23 ± 0.33 MDA mgDW−1), coinciding with a lower percentage of total lipids (13.29 ± 0.52%) and an increase in the TAC values (69.99 ± 4.58 μgTrolox Equivalent mgDW−1), respect to spring and summer seasons. Positive antioxidant balance (TAC/LP ratio) was found to M. slabberi in all season sampled increased from spring to autumn (Table 6).
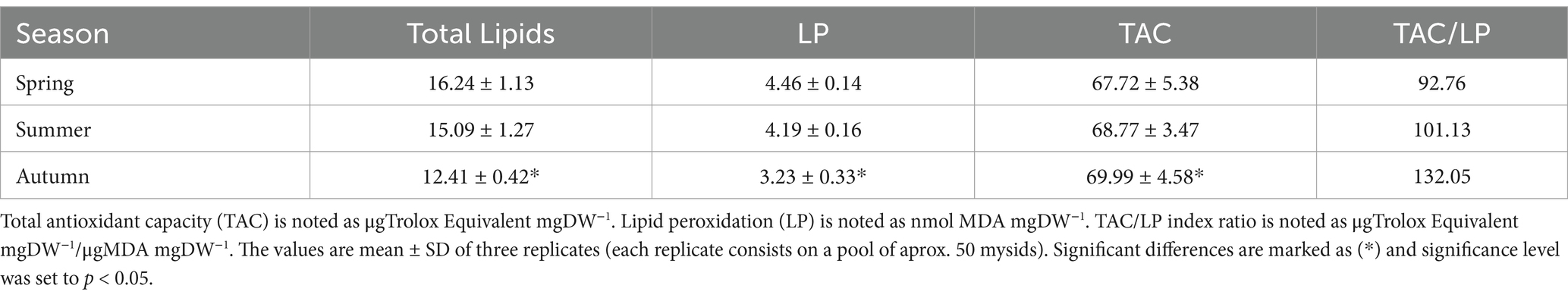
Table 6. Total antioxidant capacity and lipid peroxidation analysis of Mesopodopsis slabberi per season.
4 Discussion
Aquaculture actually requires new nutritional natural resources, demanding alternative high-quality feed ingredients but sustainable over time, where Blue Foods have become a new strong perspective on finding new critical nutritional resources and ecologically sustainable to a growing world population.
Despite being an excellent source of proteins and essential fatty acids, Mysids shrimps, which are abundant and a key prey of fish and shrimp species in many marines, coastal and freshwater ecosystems, becoming an excellent source of proteins and essential fatty acids, where their nutritional value as a potential new resource in aquaculture has barely been evaluated.
Mysid Mesopodopsis slabberi which is abundant and essential natural prey for many fish and invertebrate species in Guadalquivir river estuary, plays a key ecological role, especially for nursery function and also as a main prey for nekton fish and shrimp species, making a substantial contribution to the stock of numerous organisms with commercial interest (Mauchline, 1980; Azeiteiro et al., 2001; Jumars, 2007; Vilas et al., 2008), and contribute to transfer trace metals to higher trophic levels (Biju and George, 2021). In this way, knowing its nutritional status not only provides information of interest to deepen the study of its role in the trophic web of the estuary, but also advances in culture technology, knowing the environmental conditions in which it has been sampled allows us to infer parameters of the medium and to be able to carry out its controlled cultivation to produce biomass in the laboratory.
A better understanding of mysid contribution as value-added products and their ability to improve animal performance, nutrient availability, palatability, and feed digestibility is needed to successfully integrate them as vital resources in aquaculture feeds. The nutritional profile of mysid Mesopodopsis slabberi has been studied to determine their potential to be used as prey sustainable alternative for aquaculture. We results support this argue and are agree with previous reports to other mysid species (Guevara et al., 2005; Eusebio et al., 2010), evaluated as food for fish.
Protein values were similar to other mysid species described in the bibliography (Azeiteiro et al., 2003; Guevara et al., 2005; Eusebio et al., 2010). Present results suggest that main energy storage is not in the form of carbohydrate chains, but rather mainly as proteins or, most likely, as lipids, which were similar to other mysid species and amphipods as shown in Table 7.
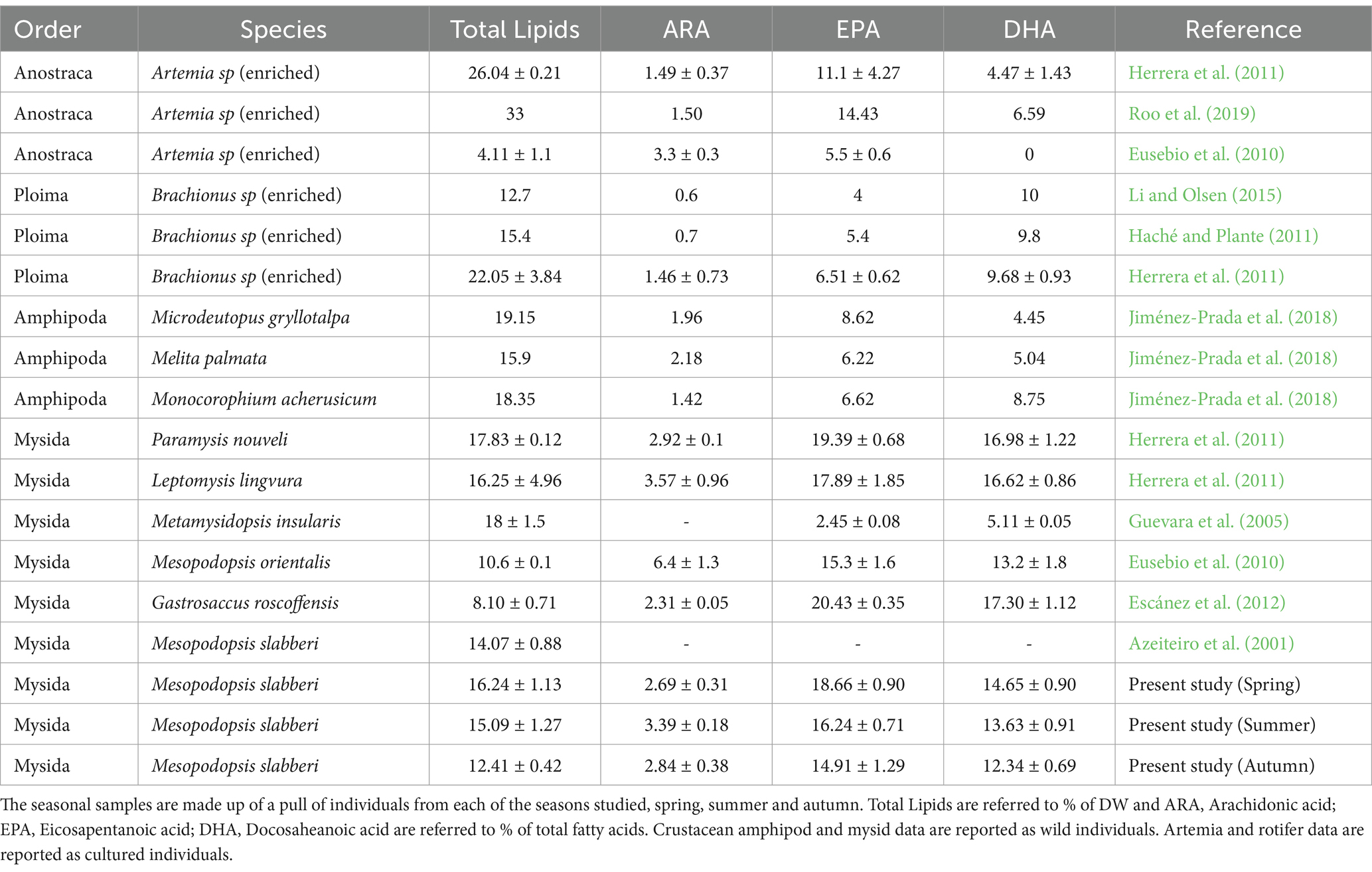
Table 7. Comparison table of seasonal fatty acids and total lipids of this study Mesopodopsis slabberi with others live preys used as alternative feed for aquaculture in literature.
In aquaculture, the content of aminoacids is considered an important nutritional value of the diet. Alanine and Aspartic Acid are among the major glucogenic precursors and important energy substrates for fish, and also can stimulate feeding response (Li et al., 2009), being interesting as functional ingredients for fish feeding in aquaculture, provided that the ideal dietary nutrient requirements of essential amino acids (EAAs) are supplied for the target species (Fricke et al., 2022). The composition of natural aminoacids of zooplankton depends on environmental conditions. Biotic and abiotic factors have the greatest influence on the composition of free AAs, not on the AA bound in proteins. Each element in the AA composition, along with biogenic elements (C, N, P), sterols, and essential polyunsaturated fatty acids, can play a key role in the physiology of plankton organisms and represent an irreplaceable or deficient food element for animals that occupy the highest levels of the aquatic ecosystem food chain (Kolmakova and Kolmakov, 2019).
Prevalent essential amino acids in M. slabberi total protein composition were Lysine, and Leucine, while least abundant were Isoleucine, Treionine, Valine, Histidine and Tryptophan. By the other hand, the highest non-essential AA content found in M. slabberi was detected for Aspartic acid + aspargine acid and Glutamic acid + glutamine, which, on the one hand, aspartic acid is an amino acid that has the ability to increase the absorption, circulation and use of minerals such as calcium, magnesium, zinc and potassium, while glutamic acid acts as a stimulant of the immune system reducing infections. The high level both of essential non-essential amino acids, especially glutamic acid, alanine, and glycine, give M. slabberi the ability to be a potential product to stimulate feed intake, in comparison to fishmeal diets low formulated in protein and amino acids most commonly used (FAO, 2020).
Considering fatty acids as trophic biomarker we can elucidate main components of M. slabberi diet demonstrating it is an omnivorous primary consumer with a great potential as a nutritive and sustainable natural resource for aquaculture which relays on basal trophic levels, detritus, bacterial and diatoms components (Guo et al., 2020; Gardade et al., 2021).
Regarding to lipids, ranging 13–16% of total lipids content respect to dry weight (DW), these values are similar to those reported by Azeiteiro et al. (2001) for M. slabberi, and relatively higher than the lipid concentrations for M. orientalis (10.6%) reported by Eusebio et al. (2010), in comparison to the lipid concentration of Artemia (4%) reported in the same (Table 7). Further, studies reported for Herrera et al. (2011) and Roo et al. (2009) for enriched Artemia salina, presented a total lipid content ranging from 26 to 33%, significantly higher taking account the natural proportion of lipids from various species of mysid shrimp; while using M. slabberi, an enrichment would not be necessary thanks to its lipid composition. The incorporation of these lipids in a natural way would contribute to a better assimilation of the same and would facilitate the work within an aquaculture facility. Analysis of the fatty acid profile reveals a predominance spectrum of polyunsaturated and saturated over monounsaturated fatty acids. Together, saturated and polyunsaturated fatty acids account for about 80% of the total fatty acid content. By the one hand, Saturated fatty acids (16:0; 18:0) and MUFA 18:1n7 values found for M. slabberi are similar to gammarids SFA values collected by Jiménez-Prada et al. (2018), with similar trophic role and founded in marsh ponds. Index 18:1n7/18:1n9 trophic ratio reported values close to unity, which would be suggesting that M. slabberi is an omnivorous organism not strictly herbivorous, and which may seasonally feed on more carnivorous sources (Magnone et al., 2015).
On the other hand, the highest proportion of M. slabberi fatty acids were found in the form of PUFA’s, around 50% of the total fatty acids, which were highly superior to the amphipod PUFAs (38.3%), reported by Baeza-Rojano et al. (2010) as a suitable natural aquafeed. The high proportion of PUFAs found in M. slabberi remained a similar percentage basis consistent with the results for mysids P. nouveli and L. lingvura described by Herrera et al. (2011), further, being significantly higher to PUFA’s percentage basis of M. insularis (30.41%), reported by Guevara et al. (2005), where the highest proportion of fatty acids was in the form of n-3 fatty acids (27.49%), compared to PUFA n-3 composition acids of M. slabberi (36–40%), being typically markers of a high consumption of microalgae, either to a greater extent different species of the genus diatoms (Σ C16; 20:5n3), or dinoflagellates (Σ C18; 22:6n3).
The high content of polyunsaturated fatty acids, especially the essential fatty acids EPA (6–10 μg FA mg DW−1) and DHA (5–8.5 μg FA mg DW−1) in M. slabberi, were significantly higher than those observed in comparison to other live preys compiled in Table 7, and presented a significant positive effect on animal growth for mysids (Viherluoto et al., 2000). It would be interesting to know if mysid shrimps could perform a “trophic upgrading” in the dietary availability of PUFA for higher trophic levels through the accumulation and/or de novo synthesis of essential fatty acids, EPA, DHA and AA, obtained from the consumption of phytoplankton and macroalgae (Hiltunen et al., 2017; Pairohakul et al., 2021), as have been described in gammarids and amphipods (Jiménez-Prada et al., 2018) and in polychaetes (Pairohakul et al., 2021). In this context, the use of mysids capable of trophically improving dietary fatty acids could have positive effects on the aquaculture fish healthcare (Eroldoğan et al., 2023), such as improving functions in the immune system, improving growth, as well as the maintenance of membrane fluidity in ectothermic species (Fink, 2013).
As demonstrated in this study, M. slabberi is an organism rich in PUFAs and other essential nutrients and is therefore a new and potentially high-quality resource for use in aquafeeds, potentially consuming it to produce dietary quality considerably higher in terms of essential PUFAs for consumers compared to (lower quality) live foods that have typically been used for decades, and allows to improve the assessment of M. slabberi as a functional product and high nutritional value as aquafeed.
The study of metal contents of M. slabberi is a valuable field of research, which is indispensable for both safety assessment as well as knowledge of their nutritional value. The capacity of mysid to accumulate metals depends on a variety of factors such as location, temperature, salinity, light, pH, nitrogen availability, season and metabolic processes. Trace elements such as zinc, iron, copper and selenium are required as coenzymes for metalloenzymes and are vital for maintenance of cellular functions in the immune system of higher vertebrates. In fish little is known about the effects of trace elements on immune function. Some studies dealing with the inclusion of different forms of trace elements such as Zn, Mn and Se in plant protein-rich diets suggest that dietary minerals supplemented in the organic form could be reasonably considered more effective than the inorganic and encapsulated forms of supply (Domínguez et al., 2017).
M. slabberi presented a high proportion of macronutrient (K > Na > Mg > Ca, over 3,000 μg/g) in their tissues regardless of the season of the year or the stage of development. Among the main trace elements of interest in aquaculture founded in M. slabberi, zinc was notable for appearing in relatively high concentrations. Zinc is the second most abundant trace element, after Fe, essential to all cells in most living organisms (Vallee and Falchuk, 1993; Dos Santos et al., 2019). Zn acts as a cofactor for many enzymes that aid in metabolism, as well as being involved in several metabolic pathways, such as nucleic acids and protein synthesis (Chatterjee et al., 2019). Like other trace elements, the main routes of zinc uptake from water are through the gills and gastrointestinal tract; however, the major route of Zn absorption is the gastrointestinal tract both in FW and SW (Spry et al., 1988; Dos Santos et al., 2019). A meta-analysis of published information on Zn requirements of several fish species showed estimates ranging from 15–115 mg kg−1 (Dos Santos et al., 2019).
Food is regarded as the main source of Fe for metabolic purposes (Bury et al., 2003). Iron in feeds is found in two forms, heme iron and non-heme iron, being heme iron found as the major sources of animal origin feed ingredients. Iron plays too an important role as fatty acid desaturases (Wood et al., 2012), being found in high proportions in M. slabberi regardless of the season, finding oscillations in the fish requirements according to species between 30 and 160 mg kg−1, consistent with the data obtained in the present work, being within the limiting values established by Dos Santos et al. (2019). Therefore, iron could influence the degree of FA saturation in marine plankton (Nechev et al., 2006), presumably metabolized more rapidly than sterols, which have a greater regulatory and structural function, being susceptible to Fe nutrition for the production and metabolism of fatty acids.
On the other hand, it is widely accepted that Cu is an essential trace element required for cellular functioning of all living organisms. Fish absorb Cu via the gills and digestive tract; however, the diet is considered a major source of Cu for growth, development and essential physiological functions (Kamunde et al., 2002; Bury et al., 2003). Cu, which plays a relevant role in the formation of respiratory pigments, was found in an average proportion in M. slabberi to cover the nutritional requirements of Cu of the fish (3–10 mg kg−1).
Selenium is present at high concentration in M. slabberi. Fish absorb limited amounts of Se from the environment via the gills and skin under certain conditions; however, the gastrointestinal tract is the primary site for Se absorption (Janz et al., 2010; Janz, 2012). Se plays an important role in the early life stages of fish, during their growth and allow them to generate a certain antioxidant capacity (Younis et al., 2015). Considering that levels of Se required for fish varies between 0.1 to 1.8 mg kg−1 (Dos Santos et al., 2019), the incorporation of a certain biomass of M. slabberi in aquaculture feed will mean a relevant Se source. It is necessary to highlight the increase in selenium levels detected in M. slabberi during the summer and autumn period. The increase detected in the water temperature in the Guadalquivir estuary may have influenced a greater bioavailability of this metal. The higher selenium values observed in M. slabberi can modulate the cellular oxidative balance as this metal has recognized antioxidant functions. M. slabberi must develop an optimal and seasonal antioxidant state, thus the determination of the origin of antioxidant enzyme drives the need to study in greater depth the oxidative stress state of estuarine organisms, which develop adaptive mechanisms to regulate the oxidative stress generated (Smith et al., 2003; Smith, 2015; Karthikeyan et al., 2019).
Mysid shrimps present the notorious ability to accumulate and transfer nutrients and essential elements to higher trophic levels; Biju and George (2021) also highlighted that mysids Mesopodopsis orientalis and Rhoplaophtalmus indicus tend to accumulate high levels of trace metals rarely above natural levels due to the absorption of metals from environmental compartments (water, suspended particulate matter, sediments, phytoplankton and benthic organisms) that are chronically exposed with industrial effluents (George et al., 2016). The seasonal concentrations of trace metals detected in M. slabberi in the present study are probably related to the seasonal uptake of metals in mysids due to variable competition for food during the seasons (Biju and George, 2021), and the high influx of metals as result of anthropic activities located on the banks of the Guadalquivir River, and the consequent increase in the bioavailability of metals for mysids, leading to a state of oxidative stress.
Oxygen is an essential component for almost all life; however, it is frequently converted to toxic reactive forms called reactive oxygen species (ROS). To overcome cell damage induced by oxidative stress, aquatic organisms have evolved antioxidant defense systems through evolutionary processes that employ non-enzymatic and antioxidants enzymes to remove excess ROS (Zhou et al., 2008; Haque et al., 2018).
Oxidative damage was evaluated by the formation of malondialdehyde (MDA) as an indication of lipid peroxidation (LP). Contrary to found by others authors like Bury et al. (2003) and Haque et al. (2018) in others mysid species, no significant variations in MDA values between stages of development were detected in M. slabberi. Differences in lipid peroxidation associated with the seasons of the year were founded in M. slabberi. These differences may be marked by environmental variables (temperature, salinity, food…) that may affect physiology response and generate an increase in ROS. Enhanced LP could be due to auto-oxidation of the PUFA as a consequence of increased oxygen presence due to higher metabolism (Jiménez-Fernández et al., 2018). The higher LP level detected in spring season is likely to be due to the prominent presence of PUFA rather than to be inefficiency of the oxidative stress defenses.
TAC measurement has become a useful tool to acquire a picture of the imbalance between free radical levels and antioxidant stores (Bartosz, 2003). TAC was used as a proxy to identify the antioxidant status of M. slabberi and our results revealing that this specie showed high TAC levels. A TAC/LP balance is a useful biomarker to determine the physiological state of the individual and also provides useful information about a potential food source and its contribution of antioxidant compounds (Jiménez-Fernández et al., 2018). The antioxidant activity evaluated in mysid demonstrated a specific compensatory mechanism to neutralize peroxides. When TAC value was plotted against LP a positive balance of TAC was detected. This fact evidences an adequate antioxidant status to mysid in all season sampled. Our data reveal that by showing the M. slabberi a positive balance of antioxidants, this makes them a suitable product for use in aquaculture feed.
The results of the present study allow us to obtain a good approach of the biochemical composition of M. slabberi and highlight the important concentration of basal elements and functional components, essential for an aquaculture feed, this species offer. The improvement on knowledge about the M. slabberi nutritional capacity will allow us to design an adequate culture system, adapted to the ideal conditions for its maintenance and development, in perspective of the potential functional capacity of M. slabberi as live food in aquaculture. These results demonstrate M. slabberi has considerable potential to be used as a functional ingredient in aquaculture. It is necessary to investigate its contribution as immune-nutrient and advance on suitable culturing protocols and technology to achieve a constant supply.
5 Conclusion
The current study concludes that mysid M. slabberi has a high percentage of proteins, is relatively high on lipid content, presents a relatively enriching proportion of essential amino acids and an adequate nutritional value fatty acids composition (high PUFAs) for applications in aquaculture, relatively rich in essential fatty acids EPA and DHA. M. slabberi also shows high proportion of macronutrient and main trace elements of interest in aquaculture, highlighting the importance elements such as selenium which are of great importance during the early stages of fish development, emphasizing the importance of these organically supplied trace elements is much more appropriate than those currently introduced as inorganic supplements. Based on its antioxidant capacity M. slabberi can be considered a powerful functional food. On summary, M. slabberi high nutritional value makes it a high low-trophic value natural resource with great potential as inert or live prey for fish and crustaceans post-larvae under culture conditions.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
PC-C: Data curation, Formal analysis, Visualization, Writing – original draft, Writing – review & editing. CV: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing. CF-D: Conceptualization, Data curation, Investigation, Methodology, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work has been supported by the INIA project RTA2014-00023-CO2–01 co-financed 80% with FEDER Operational Program 2014–2020 funds and 75% co-financed by the European Maritime and Fisheries Fund 2014–2020 and by projects GUADALQUIVIR-LTER-PP.FEM.PPA201700.5, GUADACONECT-PR.FEM. PPA201900.005 and PR.FEMPA.DIP2023B.006-GUADALQUIVIR_LTEES, 75% co-funded by the European Maritime and Fisheries Fund (EMFF). Pablo Cárdenas-Camacho has been the beneficiary of a pre-doctoral grant-contract belonging to a Training Program for research staff managed by the Andalusian Agricultural and Fisheries Research Institute (IFAPA) and financed by the National Institute of Food Research and Technology (INIA), Ministry of Economy and Competitiveness, and IFAPA (Junta de Andalucía).
Acknowledgments
Special thanks to the researchers Inmaculada Giralde from the UHU and Julian Blasco from the ICMAN-CSIC for the analysis of amino acids and metals, which would not have been possible without their collaboration.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alberts-Hubatsch, H., Slater, M. J., and Beermann, J. (2019). Effect of diet on growth, survival and fatty acid profile of marine amphipods: implications for utilisation as a feed ingredient for sustainable aquaculture. Aquac. Environ. Interact. 11, 481–491. doi: 10.3354/aei00329
Andam, M. B., Traifalgar, R. F. M., Laureta, L. V. Jr., Nuñal, S. N., and Nillos, M. G. G. (2024). Mysid meal as a dietary replacement for fishmeal in the diets of Pacific white shrimp Penaeus vannamei (Boone, 1931) postlarvae. Front. Sustain. Food Syst. 8:1322314. doi: 10.3389/fsufs.2024.1322314
Azeiteiro, U., Fonseca, J., and Marques, J. (2001). Biometry, estimates of production and seasonal variation in the biochemical composition of Mesopodopsis slabberi (Van Beneden, 1861)(Crustacea: Mysidacea). Bol. Inst. Esp. Oceanogr. 17, 15–26.
Azeiteiro, U. M., Fonseca, J. C., Pastorinho, R., Morgado, F., and Marques, J. C. (2003). Patterns of variation in the biochemical composition of Mesopodopsis slabberi (Van Beneden, 1861) (Crustacea: Mysidacea). Bol. Inst. Esp. Oceanogr. 19, 433–442.
Baeza-Rojano, E., García, S., Garrido, D., Guerra-García, J. M., and Domingues, P. (2010). Use of amphipods as alternative prey to culture cuttlefish (Sepia officinalis) hatchlings. Aquaculture 300, 243–246. doi: 10.1016/j.aquaculture.2009.12.029
Bartosz, G. (2003). Total antioxidant capacity. Adv. Clin. Chem. 37, 219–292. doi: 10.1016/S0065-2423(03)37010-6
Biju, A., and George, R. (2021). Trace metal dynamics in mysids from the Cochin estuary. Environ. Forensic 22, 56–62. doi: 10.1080/15275922.2020.1836077
Biju, A., Gireesh, R., Jayalakshmi, K., Haridevi, C., and Panampunnayil, S. (2009). Seasonal abundance, ecology, reproductive biology, and biochemical composition of Mesopodopsis orientalis WM tattersall (Mysida) from a tropical estuary (Cochin backwater) in India. Crustaceana, 82, 981–996.
Bury, N. R., Walker, P. A., and Glover, C. N. (2003). Nutritive metal uptake in teleost fish. J. Exp. Biol. 206, 11–23. doi: 10.1242/jeb.00068
Canavate, J. P., Prieto, A., Zerolo, R., Sole, M., Sarasquete, C., and Fernandez-Diaz, C. (2007). Effects of light intensity and addition of carotene rich Dunaliella salina live cells on growth and antioxidant activity of Solea senegalensis Kaup (1858) larval and metamorphic stages. J. Fish Biol. 71, 781–794. doi: 10.1111/j.1095-8649.2007.01544.x
Cañavate, J.-P., Van Bergeijk, S., Giráldez, I., González-Ortegón, E., and Vílas, C. (2019). Fatty acids to quantify phytoplankton functional groups and their spatiotemporal dynamics in a highly turbid estuary. Estuar. Coasts 42, 1971–1990. doi: 10.1007/s12237-019-00629-8
Chatterjee, A., Bhattacharya, R., and Saha, N. C. (2019). Zinc oxide (ZnO) induced toxicity and behavioural changes to oligochaete worm Tubifex tubifex (Muller). Int. J. Sci. Res. Biol. Sci. 6, 35–42.
Ching, S. Y., Hall, J., Croft, K., Beilby, J., Rossi, E., and Ghisalberti, E. (2006). Antioxidant inhibition of oxygen radicals for measurement of total antioxidant capacity in biological samples. Anal. Biochem. 353, 257–265. doi: 10.1016/j.ab.2006.02.027
Christie, W. W. (2003). Lipid analysis: Isolation, separation, identification and structural analysis of lipids. Bridgewater: The Oily Press.
Conceição, L. E., Yúfera, M., Makridis, P., Morais, S., and Dinis, M. T. (2010). Live feeds for early stages of fish rearing. Aquac. Res. 41, 613–640. doi: 10.1111/j.1365-2109.2009.02242.x
Copeman, L., Parrish, C. C., Gregory, R., Jamieson, R., Wells, J., and Whiticar, M. (2009). Fatty acid biomarkers in Coldwater eelgrass meadows: elevated terrestrial input to the food web of age-0 Atlantic cod Gadus morhua. Mar. Ecol. Prog. Ser. 386, 237–251. doi: 10.3354/meps08063
Dalsgaard, J., St John, M., Kattner, G., Müller-Navarra, D., and Hagen, W. (2003). Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 46, 225–340. doi: 10.1016/S0065-2881(03)46005-7
Domínguez, D., Rimoldi, S., Robaina, L. E., Torrecillas, S., Terova, G., Zamorano, M. J., et al. (2017). Inorganic, organic, and encapsulated minerals in vegetable meal based diets for Sparus aurata (Linnaeus, 1758). PeerJ 5:e3710. doi: 10.7717/peerj.3710
Dos Santos, C. C. M., Da Costa, J. F. M., Dos Santos, C. R. M., and Amado, L. L. (2019). Influence of seasonality on the natural modulation of oxidative stress biomarkers in mangrove crab Ucides cordatus (Brachyura, Ucididae). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 227, 146–153. doi: 10.1016/j.cbpa.2018.10.001
Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., and Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350–356.
Eroldoğan, O. T., Glencross, B., Novoveska, L., Gaudêncio, S. P., Rinkevich, B., Varese, G. C., et al. (2023). From the sea to aquafeed: a perspective overview. Rev. Aquac. 15, 1028–1057. doi: 10.1111/raq.12740
Escánez, A., Riera, R., Márquez, L., Skalli, A., Felipe, B. C., García-Herrero, I., et al. (2012). A general survey of the feasibility of culturing the mysid Gastrosaccus roscoffensis (Peracarida, Mysida): Growth, survival, predatory skills, and lipid composition. Ciencias marinas 38, 475–490.
Eusebio, P., Coloso, R., and Gapasin, R. (2010). Nutritional evaluation of mysids Mesopodopsis orientalis (Crustacea: Mysida) as live food for grouper Epinephelus fuscoguttatus larvae. Aquaculture 306, 289–294. doi: 10.1016/j.aquaculture.2010.05.004
Fink, P. (2013). Invasion of quality: high amounts of essential fatty acids in the invasive Ponto-Caspian mysid Limnomysis benedeni. J. Plankton Res. 35, 907–913. doi: 10.1093/plankt/fbt029
Fox, C. J., Blow, P., Brown, J. H., and Watson, I. (1994). The effect of various processing methods on the physical and biochemical-properties of shrimp head meals and their utilization by juvenile Penaeus-Monodon fab. Aquaculture 122, 209–226. doi: 10.1016/0044-8486(94)90511-8
Fricke, E., Koch, M., Dietz, H., Slater, M. J., and Saborowski, R. (2022). Brown shrimp (Crangon crangon) processing remains as ingredient for Litopenaeus vannamei feeds: biochemical characterisation and digestibility. Aquac. Rep. 25:101225. doi: 10.1016/j.aqrep.2022.101225
Gardade, L., Khandeparker, L., Desai, D. V., Atchuthan, P., and Anil, A. C. (2021). Fatty acids as indicators of sediment organic matter dynamics in a monsoon-influenced tropical estuary. Ecol. Indic. 130:108014. doi: 10.1016/j.ecolind.2021.108014
George, R., Martin, G., Nair, S., Thomas, S. P., and Jacob, S. (2016). Geochemical assessment of trace metal pollution in sediments of the Cochin backwaters. Environ. Forensic 17, 156–171. doi: 10.1080/15275922.2016.1163623
Glencross, B. D. (2009). Exploring the nutritional demand for essential fatty acids by aquaculture species. Rev. Aquac. 1, 71–124. doi: 10.1111/j.1753-5131.2009.01006.x
Griffin, J. E., O’malley, B. P., and Stockwell, J. D. (2020). The freshwater mysid Mysis diluviana (Audzijonyte & Väinölä, 2005)(Mysida: Mysidae) consumes detritus in the presence of Daphnia (Cladocera: Daphniidae). J. Crustac. Biol. 40, 520–525. doi: 10.1093/jcbiol/ruaa053
Guevara, M., Lodeiros, C., Donato, M., Graciani, C., and Freites, L. (2005). Nutritional quality of Metamysidopsis insularis Brattegard (Crustacea: Mysidacea). Aquac. Nutr. 11, 315–319. doi: 10.1111/j.1365-2095.2005.00361.x
Guo, F., Lee, S. Y., Kainz, M. J., and Brett, M. T. (2020). Fatty acids as dietary biomarkers in mangrove ecosystems: current status and future perspective. Sci. Total Environ. 739:139907. doi: 10.1016/j.scitotenv.2020.139907
Gutteridge, J. M., and Halliwell, B. (2018). Mini-review: oxidative stress, redox stress or redox success? Biochem. Biophys. Res. Commun. 502, 183–186. doi: 10.1016/j.bbrc.2018.05.045
Haché, R., and Plante, S. (2011). The relationship between enrichment, fatty acid profiles and bacterial load in cultured rotifers (Brachionus plicatilis L-strain) and Artemia (Artemia salina strain Franciscana). Aquaculture 311, 201–208.
Haque, M. N., Lee, D.-H., Kim, B.-M., Nam, S.-E., and Rhee, J.-S. (2018). Dose-and age-specific antioxidant responses of the mysid crustacean Neomysis awatschensis to metal exposure. Aquat. Toxicol. 201, 21–30. doi: 10.1016/j.aquatox.2018.05.023
Hernandez-Alarcon, I. (2016). Efecto de la dieta en la composición de ácidos grasos del alimento vivo utilizado en la crianza larvaria de peces marinos. La Paz, BCS, México: Maestría en Ciencias, Instituto Politecnico Nacional, Centro Interdisciplinario de Ciencias Marinas.
Herrera, A., Packard, T., Santana, A., and Gómez, M. (2011). Effect of starvation and feeding on respiratory metabolism in Leptomysis lingvura (GO Sars, 1866). J. Exp. Mar. Biol. Ecol. 409, 154–159. doi: 10.1016/j.jembe.2011.08.016
Hiltunen, M., Honkanen, M., Taipale, S., Strandberg, U., and Kankaala, P. (2017). Trophic upgrading via the microbial food web may link terrestrial dissolved organic matter to Daphnia. J. Plankton Res. 39, 861–869. doi: 10.1093/plankt/fbx050
Hoegh-Guldberg, O., Jacob, D., Taylor, M., Guillén Bolaños, T., Bindi, M., Brown, S., et al. (2019). The human imperative of stabilizing global climate change at 1.5 C. Science 365:eaaw6974. doi: 10.1126/science.aaw6974
Janz, D. (2012). “Selenium” in Fish physiology, in homeostasis and toxicology of essential metals. eds. C. M. Wood, A. P. Farrell, and C. J. Brauner (Cambridge, MA, USA: Elsevier/Academic Press), 329–374.
Janz, D. M., Deforest, D. K., Brooks, M. L., Chapman, P. M., Gilron, G., Hoff, D., et al. (2010). “Selenium toxicity to aquatic organisms” in Ecological assessment of selenium in the aquatic environment, vol. 1. eds. P. M. Chapman, W. J. Adams, M. L. Brooks, C. G. Delos, S. N. Luoma, and W. A. Maher, et al. (Boca Raton, FL, USA: CRC Press), 141–231.
Jiménez-Fernández, E., Ponce, M., Rodríguez-Rúa, A., Manchado, M., and Fernández-Díaz, C. (2018). Assessing the role of vitamin C and iron in early larvae stages of Solea senegalensis fed enriched Artemia. Aquaculture 488, 145–154. doi: 10.1016/j.aquaculture.2018.01.021
Jiménez-Fernández, E., Ponce, M., Rodriguez-Rúa, A., Zuasti, E., Manchado, M., and Fernández-Díaz, C. (2015). Effect of dietary vitamin C level during early larval stages in Senegalese sole (Solea senegalensis). Aquaculture 443, 65–76. doi: 10.1016/j.aquaculture.2015.03.013
Jiménez-Prada, P., Hachero-Cruzado, I., Giráldez, I., Fernández-Diaz, C., Vilas, C., Cañavate, J. P., et al. (2018). Crustacean amphipods from marsh ponds: a nutritious feed resource with potential for application in integrated multi-trophic aquaculture. PeerJ 6:e4194. doi: 10.7717/peerj.4194
Jumars, P. A. (2007). “Habitat coupling by mid-latitude, subtidal, marine mysids: import-subsidised omnivores” in Oceanography and marine biology: an annual review. eds. R. Gibson, R. Atkinson, and J. Gordon, (Boca Raton, FL, USA: CRC Press), 9–138.
Kamunde, C., Grosell, M., Higgs, D., and Wood, C. M. (2002). Copper metabolism in actively growing rainbow trout (Oncorhynchus mykiss): interactions between dietary and waterborne copper uptake. J. Exp. Biol. 205, 279–290. doi: 10.1242/jeb.205.2.279
Karthikeyan, P., Marigoudar, S. R., Nagarjuna, A., and Sharma, K. V. (2019). Toxicity assessment of cobalt and selenium on marine diatoms and copepods. Environ. Chem. Ecotoxicol. 1, 36–42. doi: 10.1016/j.enceco.2019.06.001
Kochert, G. (1978). "Carbohydrate determination by the phenol sulfuric acid method". in Handbook of Phycological Methods-Physiological and Biochemical Methods, eds. J. A. Hellebust and J. S. Craigie. (New York, USA: Cambridge University Press), 95–97.
Kolmakova, A. A., and Kolmakov, V. I. (2019). Amino acid composition of green microalgae and diatoms, Cyanobacteria, and zooplankton (review). Inland Water Biol. 12, 452–461. doi: 10.1134/S1995082919040060
Lentes, G. (2018). Aquaculture production of the zooplankton species Americamysis bahia: Comparing on-site production at the Waikīkī aquarium to importation : Maste of Science, University of Hawaii.
Li, P., Mai, K., Trushenski, J., and Wu, G. (2009). New developments in fish amino acid nutrition: towards functional and environmentally oriented aquafeeds. Amino Acids 37, 43–53. doi: 10.1007/s00726-008-0171-1
Li, K., and Olsen, Y. (2015). Effect of enrichment time and dietary DHA and non-highly unsaturated fatty acid composition on the efficiency of DHA enrichment in phospholipid of rotifer (Brachionus Cayman). Aquaculture 446, 310–317.
Liu, Y., Cong, P., Li, B., Song, Y., Liu, Y., Xu, J., et al. (2018). Effect of thermal processing towards lipid oxidation and non-enzymatic browning reactions of Antarctic krill (Euphausia superba) meal. J. Sci. Food Agric. 98, 5257–5268. doi: 10.1002/jsfa.9064
Lowry, O. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J. (1951). Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193, 265–275. doi: 10.1016/S0021-9258(19)52451-6
Magnone, L., Bessonart, M., Gadea, J., and Salhi, M. (2015). Trophic relationships in an estuarine environment: a quantitative fatty acid analysis signature approach. Estuar. Coast. Shelf Sci. 166, 24–33. doi: 10.1016/j.ecss.2014.12.033
Min, B. H., Ravikumar, Y., Lee, D. H., Choi, K. S., Kim, B. M., and Rhee, J. S. (2018). Age-dependent antioxidant responses to the bioconcentration of microcystin-LR in the mysid crustacean, Neomysis awatschensis. Environ. Pollut. 232, 284–292. doi: 10.1016/j.envpol.2017.09.050
Næss, T., Germain-Henry, M., and Naas, K. E. (1995). First feeding of Atlantic halibut (Hippoglossus hippoglossus) using different combinations of Artemia and wild zooplankton. Aquaculture 130, 235–250. doi: 10.1016/0044-8486(94)00323-G
Nechev, J., Stefanov, K., and Popov, S. (2006). Effect of cobalt ions on lipid and sterol metabolism in the marine invertebrates Mytilus galloprovincialis and Actinia equina. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 144, 112–118. doi: 10.1016/j.cbpa.2006.02.022
Pairohakul, S., Olive, P. J. W., Bentley, M. G., and Caldwell, G. S. (2021). Trophic upgrading of long-chain polyunsaturated fatty acids by polychaetes: a stable isotope approach using Alitta virens. Mar. Biol. 168, 1–13. doi: 10.1007/s00227-021-03874-3
Phillips, H. R. P., Guerra, C. A., Bartz, M. L. C., Briones, M. J. I., Brown, G., Crowther, T. W., et al. (2019). Global distribution of earthworm diversity. Science 366, 480–485. doi: 10.1126/science.aax4851
Planas, M., Olivotto, I., González, M. J., Laurà, R., and Zarantoniello, M. (2020). A multidisciplinary experimental study on the effects of breeders diet on newborn seahorses (Hippocampus guttulatus). Front. Mar. Sci. 7:638. doi: 10.3389/fmars.2020.00638
Roo, J., Hernández-Cruz, C. M., Mesa-Rodriguez, A., Fernández-Palacios, H., and Izquierdo, M. (2019). Effect of increasing n-3 HUFA content in enriched Artemia on growth, survival and skeleton anomalies occurrence of greater amberjack Seriola dumerili larvae. Aquaculture 500, 651–659.
Roo, F. J., Hernández-Cruz, C. M., Socorro, J. A., Fernández-Palacios, H., Montero, D., and Izquierdo, M. S. (2009). Effect of DHA content in rotifers on the occurrence of skeletal deformities in red porgy Pagrus pagrus (Linnaeus, 1758). Aquaculture 287, 84–93. doi: 10.1016/j.aquaculture.2008.10.010
Salas-Leiton, E., Hachero-Cruzado, I., Asensio, E., Vilas, C., Zerolo, R., and Cañavate, J. P. (2020). Valorisation and enhanced sustainability of Senegalese sole (Solea senegalensis) aquaculture by dietary use of ditch shrimp (Palaemonetes varians) meal. Aquaculture 522:735104. doi: 10.1016/j.aquaculture.2020.735104
Sardo, A. M., Morgado, F., and Soares, A. M. (2005). Mesopodopsis slabberi (Crustacea: Mysidacea): can it be used in toxicity tests? Ecotoxicol. Environ. Saf. 60, 81–86. doi: 10.1016/j.ecoenv.2003.12.017
Smith, S. D. (2015). Tracing organic matter pathways in marine food webs using fatty acids and compound specific stable isotope analysis 1–102 : Master of Science in Marine Science, The University of Texas at Austin.
Smith, S. V., Swaney, D. P., Talaue-Mcmanus, L., Bartley, J. D., Sandhei, P. T., Mclaughlin, C. J., et al. (2003). Humans, hydrology, and the distribution of inorganic nutrient loading to the ocean. Bioscience 53, 235–245. doi: 10.1641/0006-3568(2003)053[0235:HHATDO]2.0.CO;2
Southgate, P. (2019). “Hatchery and larval foods” in Aquaculture: farming aquatic animals and plants. eds. J. Lucas, P. Southgate, and C. Tucker. 3rd ed (New Jersey, USA: Wiley-Blackwell), 183–201.
Spry, D. J., Hodson, P. V., and Wood, C. M. (1988). Relative contributions of dietary and waterborne zinc in the rainbow trout, Salmo gairdneri. Can. J. Fish. Aquat. Sci. 45, 32–41. doi: 10.1139/f88-005
Terrazas-Fierro, M., Civera-Cerecedo, R., Ibarra-Martínez, L., Goytortúa-Bores, E., Herrera-Andrade, M., and Reyes-Becerra, A. (2010). Apparent digestibility of dry matter, protein, and essential amino acid in marine feedstuffs for juvenile whiteleg shrimp Litopenaeus vannamei. Aquaculture 308, 166–173. doi: 10.1016/j.aquaculture.2010.08.021
Vallee, B. L., and Falchuk, K. H. (1993). The biochemical basis of zinc physiology. Physiol. Rev. 73, 79–118. doi: 10.1152/physrev.1993.73.1.79
Vargas-Abúndez, J. A., López-Vázquez, H. I., Mascaró, M., Martínez-Moreno, G. L., and Simões, N. (2021). Marine amphipods as a new live prey for ornamental aquaculture: exploring the potential of Parhyale hawaiensis and Elasmopus pectenicrus. PeerJ 9:e10840. doi: 10.7717/peerj.10840
Verlecar, X. N., Das, P. B., Jena, K., Maharana, D., and Desai, S. R. (2012). Antioxidant responses in Mesopodopsis zeylanica at varying salinity to detect mercury influence in culture ponds. Turk. J. Biol. 36, 711–718. doi: 10.3906/biy-1203-63
Viherluoto, M., Kuosa, H., Flinkman, J., and Viitasalo, M. (2000). Food utilisation of pelagic mysids, Mysis mixta and M. relicta, during their growing season in the northern Baltic Sea. Mar. Biol. 136, 553–559. doi: 10.1007/s002270050715
Vilas, C., Drake, P., and Fockedey, N. (2008). Feeding preferences of estuarine mysids Neomysis integer and Rhopalophthalmus tartessicus in a temperate estuary (Guadalquivir estuary, SW Spain). Estuar. Coast. Shelf Sci. 77, 345–356. doi: 10.1016/j.ecss.2007.09.025
Willett, W., Rockström, J., Loken, B., Springmann, M., Lang, T., Vermeulen, S., et al. (2019). Food in the Anthropocene: the EAT-lancet commission on healthy diets from sustainable food systems. Lancet 393, 447–492. doi: 10.1016/S0140-6736(18)31788-4
Wood, C. M., Farrell, A. P., and Brauner, C. J. (2012). Homeostasis and toxicology of essential metals. Fish Physiology, London: Academic press.
Yang, Q., Zhou, X., Zhou, Q., Tan, B., Chi, S., and Dong, X. (2009). Apparent digestibility of selected feed ingredients for white shrimp Litopenaeus vannamei, Boone. Aquac. Res. 41, 78–86. doi: 10.1111/j.1365-2109.2009.02307.x
Younis, A. M., Amin, H. F., Alkaladi, A., and Mosleh, Y. Y. I. (2015). Bioaccumulation of heavy metals in fish, squids and crustaceans from the Red Sea, Jeddah coast, Saudi Arabia. Open J. Mar. Sci. 5, 369–378. doi: 10.4236/ojms.2015.54030
Zhou, Q., Zhang, J., Fu, J., Shi, J., and Jiang, G. (2008). Biomonitoring: an appealing tool for assessment of metal pollution in the aquatic ecosystem. Anal. Chim. Acta 606, 135–150. doi: 10.1016/j.aca.2007.11.018
Keywords: biochemical composition, mysid, Mesopodopsis slabberi , fatty acids, antioxidant status, aquaculture
Citation: Cárdenas-Camacho P, Vilas C and Fernández-Díaz C (2025) Biochemical characterization and antioxidant status of mysid Mesopodopsis slabberi: potential use as natural resource to aquaculture. Front. Sustain. Food Syst. 9:1568203. doi: 10.3389/fsufs.2025.1568203
Edited by:
Roberto Anedda, Porto Conte Ricerche, ItalyReviewed by:
Tonmoy Ghosh, Durban University of Technology, South AfricaAnselmo Miranda-Baeza, State University of Sonora, Mexico
Copyright © 2025 Cárdenas-Camacho, Vilas and Fernández-Díaz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cesar Vilas, Y2VzYXIudmlsYXNAanVudGFkZWFuZGFsdWNpYS5lcw==