 Aadil A. Rahman1*†Keren Brooks2†Shawn P. Conley3†John M. Gaska3†Trent Irby4†Carrie Knott5Chad Lee6
Aadil A. Rahman1*†Keren Brooks2†Shawn P. Conley3†John M. Gaska3†Trent Irby4†Carrie Knott5Chad Lee6 Laura E. Lindsey7†Spyros Mourtzinis3†
Laura E. Lindsey7†Spyros Mourtzinis3† Seth Naeve8
Seth Naeve8 W. Jeremy Ross9†
W. Jeremy Ross9† Maninder P. Singh10†Rachel A. Vann11
Maninder P. Singh10†Rachel A. Vann11 Emma G. Matcham1,7*†
Emma G. Matcham1,7*†- 1Department of Agronomy, University of Florida, Gainesville, FL, United States
- 2Department of Applied Agriculture and Food Studies, Morningside University, Sioux City, IA, United States
- 3Department of Plant and Agroecosystem Sciences, University of Wisconsin, Madison, WI, United States
- 4Department of Plant and Soil Sciences, Mississippi State University, Starkville, MS, United States
- 5Plant and Soil Sciences, University of Kentucky, Princeton, KY, United States
- 6Plant and Soil Sciences, University of Kentucky, Lexington, KY, United States
- 7Department of Horticulture and Crop Science, The Ohio State University, Columbus, OH, United States
- 8Department of Agronomy and Plant Genetics, University of Minnesota, St. Paul, MN, United States
- 9Department of Crop, Soil, and Environmental Sciences, University of Arkansas System Division of Agriculture, Little Rock, AR, United States
- 10Department of Plant, Soil and Microbial Sciences, Michigan State University, East Lansing, MI, United States
- 11Department of Crop and Soil Sciences, North Carolina State University, Raleigh, NC, United States
U.S. soybean (Glycine max [L.] Merr) is an important source of nutrition worldwide, with a 2022 export value of $34.3 billion. Nitrogen (N) and sulfur (S) fertilizer inputs can influence nutritional quality of seed due to their roles in amino acid biosynthesis. However, the role of N and S fertilizers needs to be further evaluated due to high nutrient demands from increased crop yields. This study examines the impact of N and S fertilizer applications on soybean composition in diverse environments across the U.S. Two S sources, ammonium sulfate (AMS) and gypsum (CaSO4), were tested at three rates (11, 22, and 33 kg S/ha). Three additional treatments included urea applied at nitrogen (N) rates of 10, 20, and 29 kg N/ha, equivalent to the N rates supplied by AMS, to assess the impact of N in AMS. All fertilizers were manually broadcast at planting, and a control treatment with no fertilizer was included. Data were collected from 40 sites in 9 states over 2019 and 2020. The effect of the 10 fertilizer treatments on soybean seed protein, oil, and sulfur-containing amino acid content was assessed. ANOVAs were performed with linear fixed effects models separately for each year of the study (2019 and 2020). Results showed that fertilizer rates had a significant effect on protein, oil, cysteine and methionine content which varied by growing environment. Our findings suggest that S and N fertilization, particularly with high rates of AMS or medium rates of gypsum, can significantly enhance cysteine and methionine content in soybeans across various environments, improving the nutritional value for human and animal consumption.
Introduction
Soybeans (Glycine max [L.] Merr) are a critical component of the U.S. agricultural sector. Since 2002, soybean yields have increased by a total 30% from 2,585 kg ha−1 to 3,331 kg ha−1 in 2022 (Vaiknoras and Hubbs, 2023). During this period, soybean production area fluctuated between 30.8 million and 35.4 million hectares [USDA National Agricultural Statistics Service (USDA-NASS), 2021]. As soybean yields increase, the crop's demand for sulfur (S) on a per-hectare basis also increases; as a result, crop S demand now exceeds annual S atmospheric deposition rates (Hinckley and Driscoll, 2022; Bender et al., 2015), suggesting that use of S fertilizer may be necessary to maintain high yield. With the need to sustain high yields, maintaining not only grain quantity but also quality, particularly through adequate nitrogen (N) and sulfur (S) availability, has become important for determining seed composition.
The effect of N application on plant proteins is characterized by an increased concentration in the content of various amino acids, particularly glutamine, asparagine, and serine (Stitt and Krapp, 2002). This increase in amino acid content plays a crucial role in enhancing the synthesis of grain proteins, which is vital for the nutritional quality and overall protein yield of the crop (Wan et al., 2023). Notably, protein quality is contingent upon sufficient S availability. Since S in plants exhibits lower mobility compared to N, its efficiency in transitioning from vegetative to reproductive tissues is reduced (Borja Reis et al., 2021). Sulfate-based fertilizers are necessary for maintaining an adequate supply of cysteine within the grain, a precursor of sulfur-containing organic compounds that are important for protein synthesis (Bouranis and Chorianopoulou, 2023).
Previous studies on N and S fertilization in the United States have demonstrated inconsistent impacts of N and S management on yield and soybean quality (Brooks et al., 2023b; Chiluwal et al., 2021; Wesley et al., 2013; Wood et al., 1993; Brown et al., 1981). However, many of these studies are geographically limited or lack data on SAA content. Additionally, suboptimal levels of these nutrients may impact soybean seed composition due to their importance in protein formation. This presents a challenge to ensure that soybean crops receive adequate S and N to meet their growing demands and maintain seed composition, while also managing environmental and economic factors in a sustainable manner. Therefore, our study aims to assess the impact of S and N applications on soybean quality, including protein, oil, and SAAs, across a wide range of environments in the United States.
Materials and methods
Field methods
The experiment was conducted across 40 sites in 9 states during the 2019 and 2020 growing seasons (Figure 1). A randomized complete block design (RCBD) with four to six replications per site was employed to account for field heterogeneity. Blocking factors were determined on a site-specific basis to reduce experimental error. The study evaluated ten treatments (Table 1), including two S sources—AMS and gypsum—applied at three different S rates (11, 22, and 33 kg S/ha), as well as three urea treatments with N rates that match the N applied by the AMS treatments (10, 20, and 29 kg N/ha). A zero-fertilizer control was also included. Fertilizer treatments were uniformly broadcast by hand at planting and not incorporated into the soil. The fertilizer application rates were determined using typical crop removal S rates based on average U.S. soybean yield levels (16–19 kg S/ha), while also testing rates above and below this range. Weed control, pest management, water management, and other plot maintenance practices were kept consistent across all plots within each site to ensure that observed treatment effects could not be confounded by differences in plot management. These background management methods align with local best management recommendations for each site, and more information can be found in Brooks et al. (2023b).
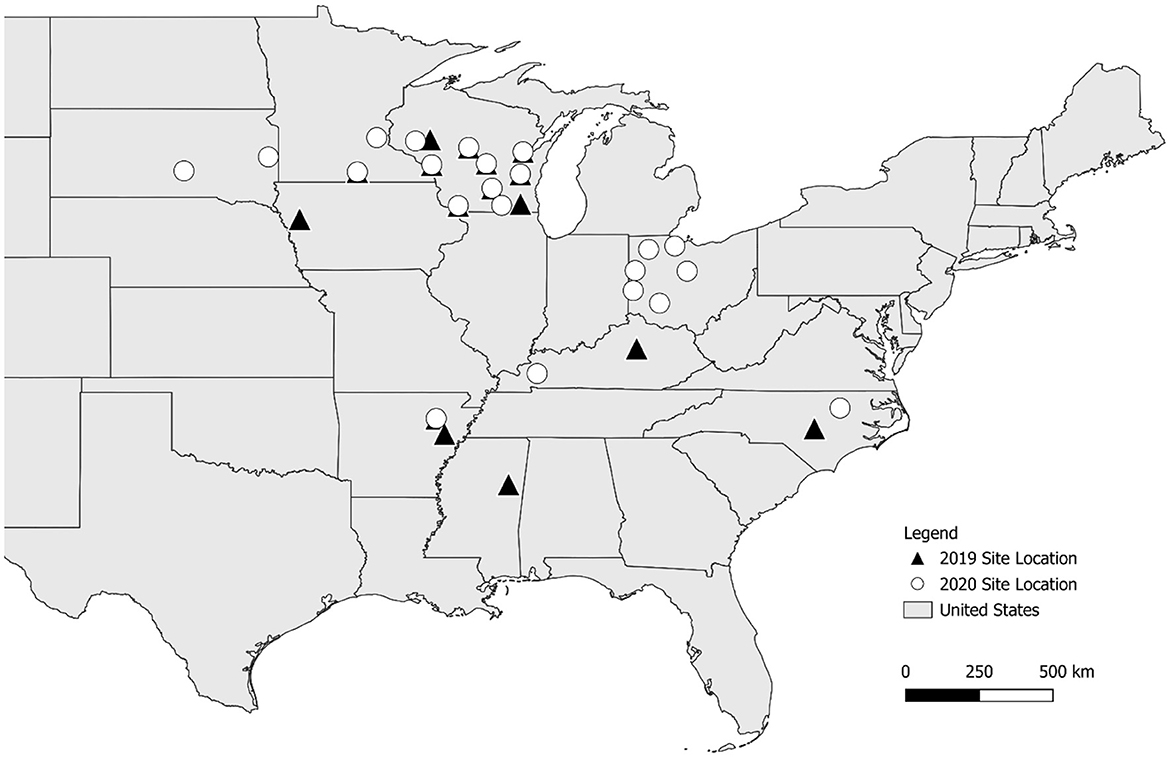
Figure 1. Trial sites in 2019 (black triangles) and 2020 (white circles).
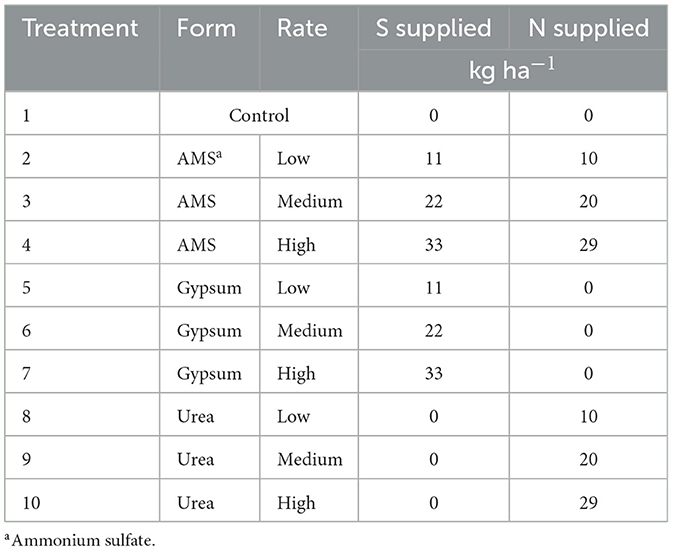
Table 1. List of treatments and nutrient rates that each treatment supplied.
Plots were harvested for grain yield with a plot combine at each location, and 150–200 g of seed were collected from each plot for grain analysis. Samples were stored in individually sealed bags and stored at room temperature. Further information about soybean yield response to S and N additions can be found in Brooks et al. (2023b). This publication includes data from 40 of the 52 sites in the Brooks et al. (2023b) paper that were able to collect grain samples during harvest.
Laboratory methods
The grain samples were dried and sent to the University of Minnesota Soybean Quality Lab, where near-infrared spectroscopy (NIRS) (PerkinElmer DA7250®) was used to analyze seed constituents. Calibration equations were developed in collaboration between the University of Minnesota and PerkinElmer using an annual validation procedure where a subset of samples are scanned in triplicate and cross-checked with wet chemistry analyses of protein, oil, moisture, and amino acids conducted at Eurofins Nutrition Analysis Center and the University of Missouri Agricultural Experiment Station Chemical Laboratories. At the end of each soybean growing season, matched NIR spectra and wet chemistry data were used by PerkinElmer to update calibration profiles, which were implemented in the following season's testing. Concentrations of 18 amino acids were measured, including methionine, cysteine, lysine, threonine, tryptophan, isoleucine, leucine, histidine, phenylalanine, valine, alanine, arginine, aspartic acid (representing aspartate and asparagine), glutamic acid (representing glutamine and glutamate), glycine, proline, serine, and tyrosine (Pfarr et al., 2018). Oil content and total protein content were also estimated by NIRS.
Seed constituents were expressed at 130 g kg−1 seed moisture content in all cases. Relative cysteine and relative methionine were calculated by dividing the methionine and cysteine content by total protein for each grain sample. While cysteine and methionine are often represented together as a sum (M+C) due to their sparing effect in the diet, we opted to analyze them separately to better understand their individual contributions to the overall amino acid profile and protein quality. This approach allows for a more detailed assessment of each amino acid's response to fertilization, providing insights that could be masked if they were grouped together.
Statistical analysis
Analysis was conducted in R 4.3.1 (R Core Team, 2023). For all response variables (protein, oil, relative cysteine content, and relative methionine content), ANOVAs were performed with linear fixed effects models separately for each year of the study (2019 and 2020) in order to improve model fit as compared to running models across both years of the study. The large size of the dataset (859 observation in 2019 and 1,023 observations in 2020) provides a good estimate of the variability in soybean grain qualities in our study area. Observations where the response variable fell outside of three standard deviations from the mean value were considered outliers, likely due to sampling error, and removed from the dataset. The total proportion of outliers removed was <1% in 2019 and <2% in 2020.
In all ANVOA models, treatment, site, and their interaction, as well as replication (nested within site), were considered fixed effects. Replication was included as a fixed effect; given the repeated use of these sites across different trials, we aimed to determine whether certain sites exhibited larger replication effects than others for use when planning future projects among our team. This analysis choice also allowed us to better determine whether blocks were arranged in a way that reduced or increased error overall in the trial, since blocking parallel to field variation patterns can sometimes reduce trial power. Kenward-Roger degrees of freedom adjustment was used for all ANOVAs. Additionally, the normality of residuals in each model was assessed using histograms and QQ-plots. Post hoc tests were performed for pairwise comparisons between different levels of factors using the package agricolae, and Bonferroni correction was applied to models (de Mendiburu, 2023).
Results
The analysis of variance (ANOVA) results for protein, oil, cysteine and methionine content indicated that there were significant differences across sites in both 2019 and 2020 (Table 2). Oil content did not vary among treatments in either year; however, there was limited evidence of a treatment × site interaction impacting oil content in 2019 (Table 2). The interaction between treatment and site impacted protein content in both years. Treatment impacted relative cysteine content in both years, and the interaction between treatment and site was significant in 2019 but not in 2020 (Table 2). For relative methionine content, differences were found among treatments in both years. The interaction between treatment and site was not significant in 2020 nor in 2019 (Table 2). Replication (nested within site) predicted differences in oil and protein levels in both years, indicating that plots were arranged at sites in a way that reduced overall trial error (Table 2). Additional post-hoc test results are available in Supplementary Table 1 (2019 ANOVAs) and Supplementary Table 2 (2020 ANOVAs).
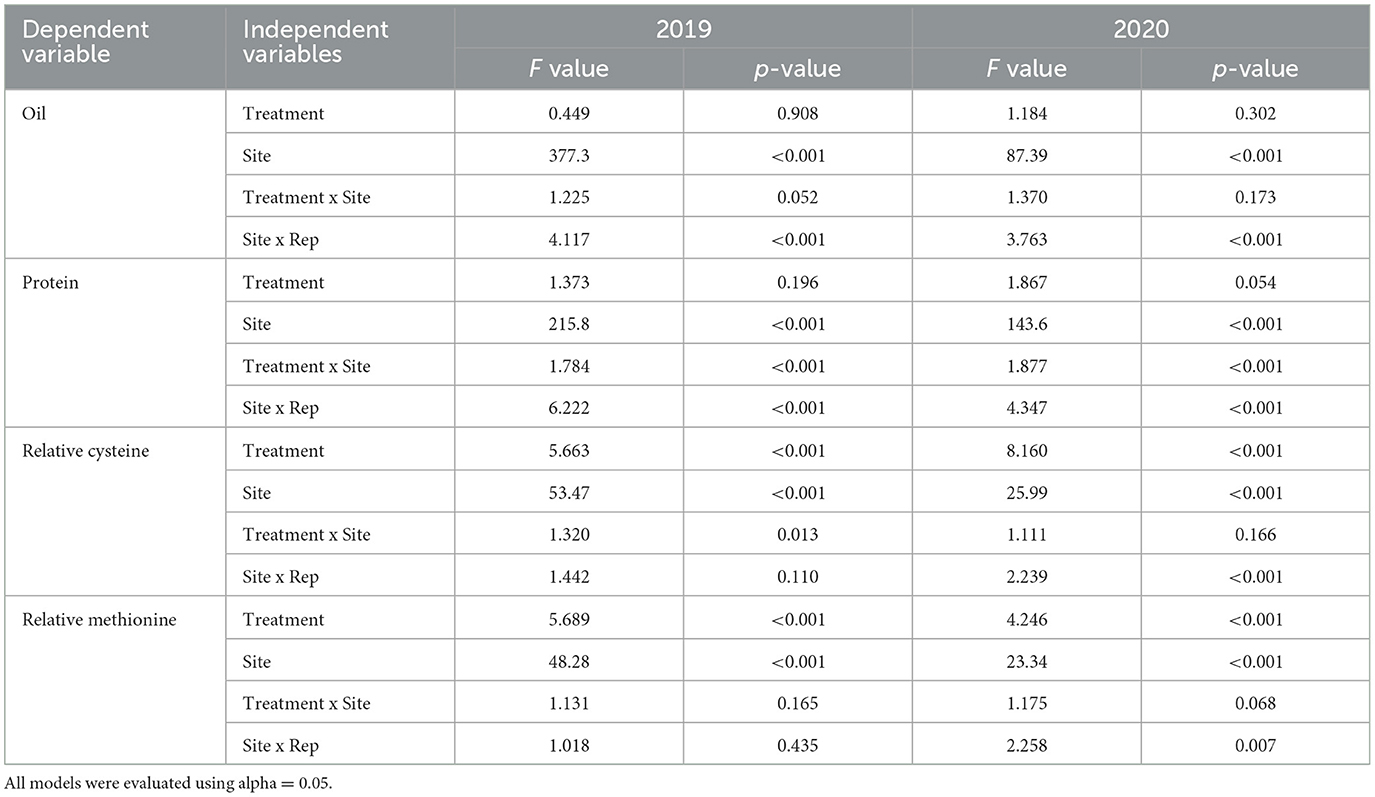
Table 2. ANOVA results for fixed-effect models where oil, protein, relative methionine content, and relative cystine content were considered dependent variables, and independent variables were treatment, site, the treatment by site interaction, and replication nested within site.
In 2019, relative methionine content was higher in soybean treated with a high rate of AMS or medium rate of gypsum as compared to the control (Figure 2a). Relative cysteine content was higher than the control in plots treated with the high rate of AMS and low Urea (Figure 2b).
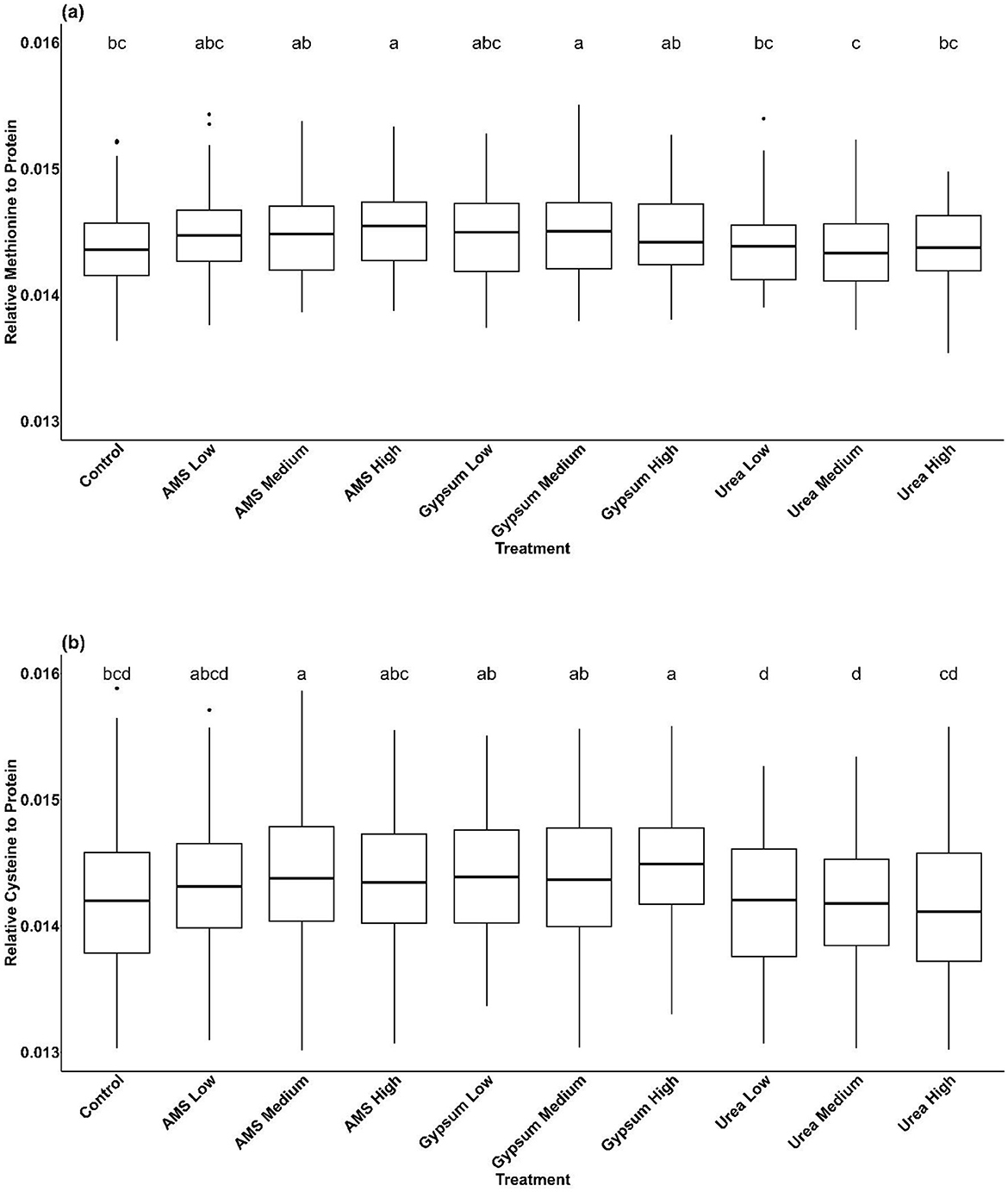
Figure 2. Boxplots for response variables with significant treatment effects (p < 0.05) identified by the overall ANOVA, post-hoc letters indicate significant differences according to Bonferroni's test. (a) is for the response variable relative methionine content in 2019, and (b) summarizes relative cysteine content in 2020. Treatments are identified on the x-axis, with the abbreviation AMS used for ammonium sulfate. Boxes delimit first and third quartiles. Solid lines inside the box indicate median. Upper and lower whiskers represent maximum and minimum values. Outliers within a given treatment are denoted with closed circles above or below the whisker lines.
In 2019 at Minnesota Lake location, plots treated with high rates of AMS had higher proportions of cysteine than plots treated with medium rates of urea (Supplementary Table 1). Similarly, for the same site and year, protein level in the urea high treatment was different from protein level in the AMS medium, but both treatments were not different than the control. In 2020 at Hancock, Wisconsin, low rates of gypsum had higher protein than medium AMS.
Discussion
The availability and assimilation of S into soybean plants significantly influences the expression of SAA genes across vegetative and reproductive organs, thereby impacting the quality of seed storage proteins (Borja Reis et al., 2021). Maintaining optimal S levels throughout plant development is critical for enhancing seed nutritional value. However, S fertilization does not consistently influence soybean production across different growing environments, particularly yields (Brooks et al., 2023b). This could be due to a variety of factors, including environmental differences such as soil type or varying amounts of atmospheric deposition of S between fields (Brooks et al., 2023b) or management factors such as utilizing P or K sources that include high concentrations of sulfur (Camberato et al., 2023).
In our study, the increases in protein content related to fertilizer application were minimal, with 38 out of 40 sites showing no changes in protein content associated with N or S fertilizers application. This aligns with other studies on N application in the U.S., including Wood et al. (1993) who concluded that N fertilization is ineffective for altering protein and oil content of soybean grown in Alabama. Similarly, Wesley et al. (2013) found no significant effect of N rates on soybean seed protein across eight sites in Kansas, and Brooks et al. (2023a) found no significant impact of late-season N application on protein content in immature soybeans (edamame) in the mid-Atlantic region. Our results showed that in Minnesota (2019), the most responsive treatment (medium AMS) resulted in a 5.3% higher protein content compared to the lowest treatment (high urea). In Wisconsin (2020), the highest treatment (high urea) exhibited a 4.3% increase in protein content compared to the lowest treatment (low urea). Given that other rates of AMS at the Minnesota site and urea at the Wisconsin site did not affect seed composition, these findings indicate that the effects of S and/or N fertilization on protein content are variable, especially in environments where nutrient application rates are a small proportion of overall quantities of available nutrients even when applications exceed crop removal rates of N and S.
Similar to N, S application did not significantly affect protein and oil content across treatments and locations. Variations in protein content for specific treatments at certain sites and years were not consistent or substantial enough to indicate a significant overall impact. This differs from both a meta-analysis across eight states, which reported a 0.3% increase in seed protein concentration with S application at planting (Borja Reis et al., 2021), and studies outside the US where S fertilization has increased soybean protein and oil content (Islam et al., 2010). Site-specific variability in soil sulfur levels and organic matter may have influenced responses in our study, as yield and protein benefits from sulfur fertilization have been shown to occur primarily in environments with intermediate soil S and SOM levels (Fleuridor et al., 2023; Borja Reis et al., 2021). Our results showed no significant effects of S application on soybean oil concentration, whereas research in Stoneville, Mississippi, found that S and S combined with N fertilizers increased soybean seed oil content, but despite these findings, our study did not detect similar effects, likely due to differences in environmental conditions, particularly water availability, which was a key factor influencing seed composition in their study (Bellaloui et al., 2011). Site-specific factors are critical to interpreting the impact of fertilizer on soybean quality, as local climate data can accurately predict variations in regional soybean seed quality (Naeve and Huerd, 2008).
Previous research has shown that incidental application of S contributes to the total sulfur available in the system, ensuring that sulfur availability is sufficient throughout the crop's development (Camberato et al., 2023). Our results align with research suggesting that when sulfur availability is not severely limiting and remobilization from vegetative tissues supplies much of the seed's sulfur demand during reproduction, sulfur applications may have minimal impact on overall seed protein and oil concentrations but can selectively enhance sulfur-containing amino acids such as methionine and cysteine (Naeve and Shibles, 2005; Sexton et al., 2008).
Across all sites, high AMS rates in 2019 raised relative methionine by 0.87%, and high gypsum rates in 2020 raised relative cysteine by 1.84%. These findings align with a multilevel meta-analysis conducted across eight states which demonstrated an ~1% increase in SAA concentration regardless of fertilization timing (Borja Reis et al., 2021). Similarly, a comprehensive study across the main soybean-producing regions of the U.S., including Kansas, Minnesota, Arkansas, Illinois, Iowa, South Dakota, and Indiana, confirmed that SAA content was enhanced after S application, irrespective of fertilizer application timing (Moro Rosso et al., 2020). Furthermore, research in Maryland and Pennsylvania soybean fields showed that S application increased the proportion of methionine and cysteine in both extracted protein and seeds (Rushovich and Weil, 2021). These consistent findings across differing regions suggest a response of soybean SAA content to S fertilization. Across our study, as well as in previous research, incorporating S at rates higher than soil available S levels can increase in SAA content in many environments. The decline in atmospheric sulfur deposition across the Upper Midwest, combined with spatial variability in soil sulfur pools and mineralization capacity, creates potential for localized deficiencies even when regional averages appear sufficient; future fertilization strategies should therefore account for the non-uniformity of sulfur inputs and crop responses, as regional trends may obscure meaningful site-level variation (Hinckley and Driscoll, 2022).
Sulfur-containing amino acids (SAAs), specifically cysteine and methionine, significantly impact protein quality and are essential for both human and animal nutrition. Soybean meal, a common animal feed ingredient, falls short of the recommended 3.5 g of SAAs per 100 g of protein, and SAA deficiencies in animal diets can impair health and development (Krishnan and Jez, 2018). With the growing demand for meat substitutes, soybeans have also gained prominence as a key protein source in human diets (Smetana et al., 2023; Zhang et al., 2021). Although soybean seed composition is typically around 40% protein, sulfur-containing amino acids make up <1.5% of that total, falling below recommended dietary levels (Assefa et al., 2018). In our study, sulfur fertilization increased the relative concentration of methionine by 0.87% in 2019 (with a high AMS rate) and cysteine by 1.84% in 2020 (with a high gypsum rate). Given the low baseline of <1.5% SAAs, these increases represent a substantial proportional improvement in nutritional quality.
While increasing SAA concentrations in soybeans through fertilizer application may be beneficial to consumers, their increased concentration in soybean due to fertilizer application may not justify the additional costs to farmers, especially considering the efficiency and affordability of direct amino acid supplementation in both livestock and human nutrition (Neubauer and Landecker, 2021). Additionally, the relationship between yield and S fertilization is variable and influenced by factors such as soil type, organic matter content, and geographical region (Borja Reis et al., 2021; Brooks et al., 2023b). This cost-benefit consideration has been explored in other agronomic crops, including wheat. Previous research on nitrogen fertilization in wheat shows that while N significantly enhances crude protein content and amino acid composition, farmers must carefully weigh these benefits against the added fertilizer costs (Gunthardt and McGinnis, 1957; Brooks et al., 2023b).
A previous study in 1995 found that a 1% increase in methionine adds 0.77 cents per kilogram to soybean value (McVey et al., 1995), highlighting its significance for organic livestock production due to feed restrictions (USDA National List of Allowed Prohibited Substances, 2024). When adjusted for inflation using the consumer price index (CPI), this equates to 1.32 cents per kilogram in the year 2020 (U.S. Bureau of Labor Statistics, 2024). Additionally, based on price estimates in the study area from 2019 to 2020 for fertilizer without application costs, the cost of high rates of AMS and gypsum would be ~53.82 and $39.85 per hectare, respectively. The profitability impact of these treatments would vary based on annual and geographic variation in fertilizer costs.
The deficiency of SAAs in soybean meal and many common feeds necessitates the use of synthetic supplements to support optimal growth therefore managing soybean for increased SAA content may be more economically viable for organic livestock feed production. Our study found high AMS rates increased methionine by 0.87%, while in 2020, high gypsum rates increased cysteine by 1.84%. Cysteine can partially replace methionine in livestock feed and reduce the need for methionine when adequately supplied (Shoveller et al., 2003; Di Buono et al., 2001). Based on these average increased SAA levels, the 0.87% increase in methionine alone would add ~1.15 cents per kilogram (0.87 * 1.32 cents), translating to a considerable added value per hectare when scaled up to typical yields. The organic livestock market (meat, dairy, eggs, poultry, fish) remains a critical segment of the organic sector, with dairy and eggs alone accounting for 8.4% of total organic retail sales (U.S. Department of Agriculture, 2025). Given that organic livestock feed cannot contain lab-produced SAAs, exploring the use of fertilizers, especially organic-approved sulfur sources like gypsum, presents a viable option to increase SAAs for these markets.
Currently, most soybean farmers are compensated based on yield rather than composition, though some markets are starting to shift toward rewarding the quality and nutritional profile of the beans. For example, high oleic soybeans, with higher levels of monounsaturated fats and lower saturated fat levels compared to traditional varieties, are specifically cultivated to meet the demands of certain markets (Nicholson et al., 2024). This study suggests there may be economic and practical advantages in transitioning to a composition-based approach in soybean management, at least in niche markets. By focusing on sulfur amino acids (SAA), essential for livestock feed, this research highlights ways to enhance the nutritional value of soybeans. For instance, specific fertilizer treatments like ammonium sulfate (AMS) and gypsum led to increases in methionine content by 0.87% and cysteine by 1.84%, thereby improving soybean's nutritional profile. Since cysteine can partially replace methionine in feed and reduce the need for methionine supplementation when sufficiently supplied (Shoveller et al., 2003; Di Buono et al., 2001), the increased SAA levels are valuable. Additionally, financially rewarding farmers can help mitigate soybean production challenges related to seed quality by providing economic motivation (Borja Reis et al., 2022).
Conclusions
Our findings indicate that while S and N applications can consistently increase SAA content of soybean grown in diverse environments, the impact of fertilizer on overall protein and oil content is variable. While increasing SAAs can add value to soybean crops, the economic viability of these fertilization practices must be considered. The additional costs associated with high rates of S and N fertilizers may be justified in specific contexts, such as organic livestock feed markets where use of synthetic amino acids are restricted.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
AR: Data curation, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. KB: Data curation, Writing – original draft, Writing – review & editing. SC: Supervision, Writing – original draft, Writing – review & editing. JG: Data curation, Supervision, Writing – original draft, Writing – review & editing. TI: Writing – original draft, Writing – review & editing. CK: Data curation, Conceptualization, Investigation, Writing – review & editing. CL: Data curation, Conceptualization, Investigation, Writing – review & editing. LL: Writing – original draft, Writing – review & editing. SM: Methodology, Writing – original draft, Writing – review & editing. SN: Writing – original draft, Writing – review & editing. WR: Writing – original draft, Writing – review & editing. MS: Writing – original draft, Writing – review & editing. RV: Writing – original draft, Writing – review & editing. EM: Data curation, Investigation, Methodology, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors would like to thank the Qualified State Soybean Boards who funded data collection in some states.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2025.1572255/full#supplementary-material
References
Assefa, Y., Bajjalieh, N., Archontoulis, S., Casteel, S., Davidson, D., Kovács, P., et al. (2018). Spatial characterization of soybean yield and quality (amino acids, oil, and protein) for United States. Sci. Rep. 8:14653. doi: 10.1038/s41598-018-32895-0
Bellaloui, N., Ebelhar, M. W., Gillen, A. M., Fisher, D. K., Abbas, H. K., Mengistu, A., et al. (2011). Soybean seed, protein, oil, and fatty acids are altered by s, and, s, + n, fertilizers under irrigated or non-irrigated environments, Agric. Sci. 2, 465–476. doi: 10.4236/as.2011.24060
Bender, R. R., Haegele, J. W., and Below, F. E. (2015). Nutrient uptake, partitioning, and remobilization in modern soybean varieties. Agron. J. 107, 563–573. doi: 10.2134/agronj14.0435
Borja Reis, A. F., Rosso, L., Davidson, D., Kovács, P., Purcell, L. C., Below, F. E., et al. (2022). Soybean management for seed composition: the perspective of U.S. farmers. Agron. J. 114, 2608–2617. doi: 10.1002/agj2.21082
Borja Reis, A. F., Rosso, L. H. M., Davidson, D., Kovács, P., Purcell, L. C., Below, F. E., et al. (2021). Sulfur fertilization in soybean: a meta-analysis on yield and seed composition. Eur. J. Agron. 127:126285. doi: 10.1016/j.eja.2021.126285
Bouranis, D. L., and Chorianopoulou, S. N. (2023). Foliar application of sulfur-containing compounds—pros and cons. Plants 12:3794. doi: 10.3390/plants12223794
Brooks, K., Mourtzinis, S., Conley, S. P., Reiter, M. S., Gaska, J., Holshouser, D. L., et al. (2023b). Soybean yield response to sulfur and nitrogen additions across diverse U.S. environments. Agron. J. 115, 370–383. doi: 10.1002/agj2.21216
Brooks, K., Reiter, M., Zhang, B., and Mott, J. (2023a). Edamame yield and quality response to nitrogen and sulfur fertilizers. Agronomy 13:1865. doi: 10.3390/agronomy13071865
Brown, J. R., Thom, W. O., and Wall, L. L. Sr. (1981). Effects of sulfur application on yield and composition of soybeans and soil sulfur. Commun. Soil Sci. Plant Anal. 12, 247–261. doi: 10.1080/00103628109367146
Camberato, J. J., Li, P., and Nielsen, R. L. (2023). Potentially significant amounts of sulfate-S found in phosphorus fertilizers. Crop Forage Turfgrass Manag. 9:e20248. doi: 10.1002/cft2.20248
Chiluwal, A., Haramoto, E. R., Hildebrand, D., Naeve, S., Poffenbarger, H., Purcell, L. C., et al. (2021). Late-season nitrogen applications increase soybean yield and seed protein concentration. Front. Plant Sci. 12:715940. doi: 10.3389/fpls.2021.715940
de Mendiburu, F. (2023). Agricolae: Statistical Procedures for Agricultural Research (R package version 1.3–7). Available online at: https://CRAN.R-project.org/package=agricolae (accessed August 2023).
Di Buono, M., Wykes, L. J., Ball, R. O., and Pencharz, P. B. (2001). Dietary cysteine reduces the methionine requirement in men. Am. J. Clin. Nutr. 74, 761–766. doi: 10.1093/ajcn/74.6.761
Fleuridor, L., Fulford, A., Lindsey, L. E., Lentz, E., Watters, H., Dorrance, A., et al. (2023). Ohio grain crop response to sulfur fertilization. Agron. J. 115, 1282–1295. doi: 10.1002/agj2.21328
Gunthardt, H., and McGinnis, J. (1957). Effect of nitrogen fertilization on amino acids in whole wheat. J. Nutr. 61, 167–176. doi: 10.1093/jn/61.2.167
Hinckley, E. -L. S., and Driscoll, C. T. (2022). Sulfur fertiliser use in the Midwestern US increases as atmospheric sulfur deposition declines with improved air quality. Commun. Earth Environ. 3:1. doi: 10.1038/s43247-022-00662-9
Islam, M., Farhad, S., Islam, M., Hoque, S., and Bhuiyan, M. (2010). Role of potassium and sulphur on the growth, yield and oil content of soybean (Glycine max L.). 3. doi: 10.15373/2249555X/JUNE2013/162
Krishnan, H. B., and Jez, J. M. (2018). Review: the promise and limits for enhancing sulfur-containing amino acid content of soybean seed. Plant Sci. 272, 14–21. doi: 10.1016/j.plantsci.2018.03.030
McVey, M. J., Pautsch, G. R., and Baumel, C. P. (1995). Estimated domestic producer and end user benefits from genetically modifying U.S. Soybeans. J. Prod. Agric. 8, 209–216. doi: 10.2134/jpa1995.0209
Moro Rosso, L., Carciochi, W., Naeve, S., Kovács, P., Casteel, S., and Ciampitti, I. (2020). Nitrogen and sulfur fertilization in soybean: impact on seed yield and quality. Kansas Agric. Exp. Station Res. Rep. 6. doi: 10.4148/2378-5977.7934
Naeve, S. L., and Huerd, S. C. (2008). Year, region, and temperature effects on the quality of Minnesota's soybean crop. Agron. J. 100, 690–695. doi: 10.2134/agronj2007.0204
Naeve, S. L., and Shibles, R. M. (2005). Distribution and mobilization of sulfur during soybean reproduction. Crop Sci. 45, 2261–2269. doi: 10.2135/cropsci2005.0155
Neubauer, C., and Landecker, H. (2021). A planetary health perspective on synthetic methionine. Lancet Planetary Health 5, e560–e569. doi: 10.1016/S2542-5196(21)00138-8
Nicholson, C. F., Stephenson, M. W., Armentano, L., and Harvatine, K. (2024). Economic analysis of high-oleic soybeans in dairy rations. J Dairy Sci. 107, 3642–3650. doi: 10.3168/jds.2023-23738
Pfarr, M. D., Kazula, M. J., Miller-Garvin, J. E., and Naeve, S. L. (2018). Amino acid balance is affected by protein concentration in soybean. Crop Sci. 58, 1521–1530. doi: 10.2135/cropsci,2017.11.0703
R Core Team (2023). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online at: https://www.R-project.org/ (accessed August 2023).
Rushovich, D., and Weil, R. (2021). Sulfur fertility management to enhance methionine and cysteine in soybeans. J. Sci. Food Agric. 101, 6595–6601. doi: 10.1002/jsfa.11307
Sexton, P. J., Paek, N. C., Naeve, S. L., and Shibles, R. M. (2008). Sulfur metabolism and protein quality of soybean. J. Plant Nutr. 5, 285–308. doi: 10.1300/J144v05n01_12
Shoveller, A. K., Brunton, J. A., House, J. D., Pencharz, P. B., and Ball, R. O. (2003). Dietary cysteine reduces the methionine requirement by an equal proportion in both parenterally and enterally fed piglets. J. Nutr. 133, 4215–4224. doi: 10.1093/jn/133.12.4215
Smetana, S., Ristic, D., Pleissner, D., Tuomisto, H. L., Parniakov, O., and Heinz, V. (2023). Meat substitutes: resource demands and environmental footprints. Resources, conservation, and recycling 190:106831. doi: 10.1016/j.resconrec.2022.106831
Stitt, M., and Krapp, A. (2002). The interaction between elevated carbon dioxide and nitrogen nutrition: the physiological and molecular background. Plant Cell Environ. 22, 583–621. doi: 10.1046/j.1365-3040.1999.00386.x
U.S. Bureau of Labor Statistics (2024). CPI inflation calculator. Available online at: https://data.bls.gov/cgi-bin/cpicalc.pl
U.S. Department of Agriculture (2025). Organic Situation Report, 2025 Edition (Report No. EIB-281). Economic Research Service. Available online at: https://www.ers.usda.gov/ (accessed April 17, 2025).
USDA National Agricultural Statistics Service (USDA-NASS). (2021). Prices recieved: Soybean prices received by month. USDA-NASS. Available online at: https://www.nass.usda.gov/Charts_and_Maps/Agricultural_Prices/pricesb.php (accessed August 2023).
USDA National List of Allowed and Prohibited Substances (2024). Available: https://www.ams.usda.gov/rules-regulations/organic/national-list (accessed July 6, 2024).
Vaiknoras, K., and Hubbs, T. (2023). Characteristics and trends of U.S. soybean production practices, costs, and returns since 2002 (Report No. ERR-316). U.S. Department of Agriculture, Economic Research Service. doi: 10.32747/2023.8023698.ers
Wan, C., Gao, L., Wang, J., Lei, X., Tao, J., Feng, B., et al. (2023). Effects of nitrogen fertilizer on protein synthesis, accumulation, and physicochemical properties in common buckwheat. Crop J. 11, 941–950. doi: 10.1016/j.cj.2023.01.002
Wesley, T. L., Lamond, R. E., Martin, V. L., and Duncan, S. R. (2013). Effects of late-season nitrogen fertilizer on irrigated soybean yield and composition. J. Prod. Agric. 11, 331–336. doi: 10.2134/jpa1998.0331
Wood, C. W., Torbert, H. A., and Weaver, D. (1993). Nitrogen Fertilizer Effects on Soybean Growth, Yield and Seed Composition. J. Prod. Agric. 3, 354–360. doi: 10.2134/jpa1993.0354
Keywords: soybean, sulfur, nitrogen, sulfur containing amino acids, protein, fertilizer
Citation: Rahman AA, Brooks K, Conley SP, Gaska JM, Irby T, Knott C, Lee C, Lindsey LE, Mourtzinis S, Naeve S, Ross WJ, Singh MP, Vann RA and Matcham EG (2025) Impact of sulfur and nitrogen fertilization on seed composition of soybean. Front. Sustain. Food Syst. 9:1572255. doi: 10.3389/fsufs.2025.1572255
Received: 06 February 2025; Accepted: 21 April 2025;
Published: 26 May 2025.
Edited by:
Pushp Sheel Shukla, Sea6 Energy Private Limited, IndiaReviewed by:
Shuchi Singh, University of Illinois at Urbana-Champaign, United StatesPurabi Banerjee, Bidhan Chandra Krishi Viswavidyalaya, India
Copyright © 2025 Rahman, Brooks, Conley, Gaska, Irby, Knott, Lee, Lindsey, Mourtzinis, Naeve, Ross, Singh, Vann and Matcham. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aadil A. Rahman, YWFkaWwucmFobWFuQHVmbC5lZHU=; Emma G. Matcham, bWF0Y2hhbS4zQG9zdS5lZHU=
†ORCID: Aadil A. Rahman orcid.org/0009-0002-8423-2935
Keren Brooks orcid.org/0000-0003-1745-7077
Shawn P. Conley orcid.org/0000-0002-8413-1088
John M. Gaska orcid.org/0000-0002-4272-6649
Trent Irby orcid.org/0000-0002-8195-2462
Laura E. Lindsey orcid.org/0000-0001-7026-0949
Spyros Mourtzinis orcid.org/0000-0002-7302-5482
W. Jeremy Ross orcid.org/0000-0001-8319-8515
Maninder P. Singh orcid.org/0000-0001-6166-2452
Emma G. Matcham orcid.org/0000-0002-9896-2253