 Figen Taşcı Durgut
Figen Taşcı Durgut Funda Eryılmaz Açıkgöz
Funda Eryılmaz Açıkgöz Fatma Funda Özdüven2
Fatma Funda Özdüven2 Seydi Yıkmış
Seydi Yıkmış- 1Department of Machinery and Metal Technology, Vocational College of Technical Sciences, University of Tekirdag Namik Kemal, Tekirdag, Türkiye
- 2Department of Plant and Animal Production, Vocational College of Technical Sciences, University of Tekirdag Namik Kemal, Tekirdag, Türkiye
- 3Department of Food Technology, Tekirdag Namık Kemal University, Tekirdag, Türkiye
This study aimed to evaluate the performance of a fully automated microsprout production cabinet system developed with a sustainable food processing perspective and to compare the nutritional and functional properties of microsprouts produced in different growing environments. Leek, amaranth, and purslane microsprouts were grown in fully automated (confetti cabinet) and standard laboratory environments. Microsprout samples were analyzed for dry matter, protein, vitamin C, macro- and microelement contents, and total antioxidant capacity. According to the results, leek samples grown in the cabinet showed significantly higher vitamin C content than those grown in the laboratory environment (74.27 ppm vs. 62.68 ppm, p < 0.05). The highest antioxidant capacity was detected in purslane samples, with an EC50 value of 174.64 μg/mL. Furthermore, the nitrogen content was measured at high levels in all species, ranging from 6.14 to 7.62%. PCA and correlation analyses revealed significant differences in the distribution of certain micronutrients (e.g., Na, Mg, B) across different production environments. These findings demonstrate that fully automated production cabinets provide effective, sustainable, and traceable systems for producing micro sprouts with high nutritional quality, and have significant potential for future applications in functional food production.
1 Introduction
The growing global demand for food, coupled with the challenges of climate change, depletion of soil and water resources, and the need to adapt agricultural practices to a changing climate, has highlighted the importance of environmental protection and biodiversity conservation. Meeting the increasing food demands with limited resources requires a skilled workforce and advanced technology. Additional technology must be developed to meet this requirement, and information transfer in agriculture must be facilitated. Additionally, systems that promote the efficient utilization of resources should be created.
Understanding climate change projections that affect ecosystems at the local level forms the basis for developing nature conservation strategies resilient to climatic changes (Pearce-Higgins et al., 2022). The vital importance of agricultural production in providing sufficient food for the growing world population is increasing, but threats of attack are not significantly affecting this support (Fasciolo et al., 2024). The growing global demand for food, coupled with the challenges of climate change, depletion of soil and water resources, and the need to adapt agricultural practices to a changing climate, has highlighted the importance of environmental protection and biodiversity conservation. Meeting the increasing food demands with limited resources requires a skilled workforce and advanced technology. Additional technology must be developed to meet this requirement, and information transfer in agriculture must be facilitated. Additionally, systems that promote the efficient utilization of resources should be created (Liu et al., 2021; Yap and Al-Mutairi, 2024).
Furthermore, the “Internet of Things (IoT)” has been integrated into the system by installing sensors in agricultural machinery. IoT increases productivity while minimizing loss and damage by integrating wired sensor networks, artificial intelligence, and cloud computing (Choudhary et al., 2025). This digitalization allows for the analysis of data collected through intelligent tools. The integration of smart technologies into agricultural systems has gained increasing attention in response to climate change, resource limitations, and the demand for sustainable food production. Among these technologies, the Internet of Things (IoT) has emerged as a transformative tool across all stages of agricultural production. During the development of agricultural technologies, the adoption of IoT enables precision planting in open fields, controlled environment facilities, and greenhouses; ensures the supervision of product quality and safety; and facilitates real-time monitoring of environmental parameters such as soil temperature and moisture, nitrogen and oxygen levels, ambient temperature and humidity, light intensity, and CO2 concentration. These data streams provide a scientific basis for informed crop management decisions, optimize growing conditions, and enhance both productivity and sustainability. As a result, IoT technologies significantly promote the automation and intelligence level in agriculture, contributing to the advancement of controlled environment agriculture and digital food systems (Ojo and Zahid, 2022). These technologies effectively address the requirements of global agricultural systems by overseeing agrarian production. They promote environmentally sustainable practices, facilitate production planning, enable traceability through data analysis at different stages of the production process, ensure sustainability, promote digitalization, encourage innovative agricultural practices, optimize the use of limited resources, and facilitate quality management (Elijah et al., 2018). Furthermore, the rapid living conditions prevalent in our era, together with the rise in education and income levels, have resulted in the emergence of discerning customers and a surge in the need for dependable food. Given this consciousness, people prioritize agricultural products that can be consumed in meals without being processed or preserved. Ensuring productivity, sustainability, and food safety in a farming system relies heavily on providing optimal conditions for plant growth. By maintaining these conditions, the nutritional value of the plants is preserved, and harvesting operations can be carried out effectively. Implementing regulated environmental conditions in plant production enhances yield, nutritional quality, disease and insect resistance, and guarantees the safety of food consumption (McClements et al., 2021).
Microgreens have gained popularity in recent years because they can increase the aesthetic appeal of professional kitchens and are nutritious greens used in diet programs. Because of their abundant bioactive ingredients, microgreens are now commonly used as a garnish for food. Furthermore, there is an increasing interest in commercial production. Microgreens are collected once the cotyledon leaves have fully matured, typically 7–14 days following germination, even though genuine leaves have not yet appeared. True leaves exhibit an extended growth period of 20 days or more and typically necessitate the application of fertilizers and agrochemicals. They are harvested once they have fully matured. Microgreens, on the other hand, are cultivated in a controlled environment using either soil or other growing materials. They rely on a combination of natural and/or artificial light sources for their growth (Kyriacou et al., 2017; Lenzi et al., 2019; Pinto et al., 2015; Xiao et al., 2019).
Microgreens possess many health-enhancing substances, such as antioxidants, phenols, vitamins, and minerals, which classify them as functional foods or superfoods. Micro-greens are known to have a greater abundance of nutrients compared to vegetables that are harvested at the usual growing stage (Düzgüneş et al., 1987; Ebert et al., 2014; Pinto et al., 2015).
In this investigation, we developed and implemented a novel, fully automated microgreen growing cabinet—the confetti cabinet—designed to establish an optimized and precisely regulated environment for microgreen cultivation. Unlike traditional or semi-automated systems, this prototype integrates real-time sensor-supported monitoring and PLC-based automation to control critical parameters such as humidity, light spectrum, CO2 concentration, temperature, irrigation, and ventilation. What sets this system apart is its capacity to enable autonomous environmental regulation, continuous data acquisition, and traceable production conditions. This approach ensures consistency and high nutritional value in microgreen production and represents a significant advancement in the digitalization and sustainability of controlled-environment agriculture systems.
2 Materials and methods
2.1 Materials used in growing microgreens
The microgreen plants selected for the research project were leek from the Alliaceae family, amaranth from the Amaranthaceae family, and purslane from the Portulacaceae family. The seeds utilized in the study were conventional seeds.
The study utilized peat (Klasmann-Deilmann, Potground H, Germany) as the growing medium, which possessed the following chemical properties: nitrogen (N) content ranging from 160 to 260 mg/L, phosphorus pentoxide (P2O5) content ranging from 180 to 280 mg/L, potassium oxide (K2O5) content ranging from 200 to 150 mg/L, magnesium (Mg) content ranging from 80 to 150 mg/L, pH level of 6, nitrogen (N) content of 0.8%, organic matter content of 70%, and carbon (C) content of 35%.
2.2 Fully automated micro foliage plant growing cabinet
A prototype microgreen plant growing cabinet, capable of full automation, was designed and produced specifically for conducting trials of microgreen plant growing (confetti cabinet). An LED lighting system (5050 RGB LED Strip, 60 LED/m Shenzhen Ledmy Co., Ltd., Shenzhen, China) along with temperature, humidity, CO2 sensors (MH-Z19C, EMS Control Electronics and Machinery Inc., Izmir, Türkiye) a camera (Shenzhen Jınshuoshi Industrial Co., LTD), a 7″ touch monitor, and an IoT system, was installed in the confetti cabinet. This setup allowed for automation management and ensured that the microgreen species grown in the fully automated microgreen growing cabinet could develop optimally. The microgreen plant growth cabinet incorporates production trays that house the growing environment. The production trays are constructed from stainless steel with 18/10 composition and are specifically engineered with a 15° angle to facilitate the drainage of any surplus water in the plant root zone within the plant growth environment (Figure 1).

Figure 1. Microgreens growing cabinet (confetti cabinet) and production tray layout.
2.3 Microgreen growing cabinet measurement and control equipment
The process conditions for the microgreen growing cabinet system were regulated using measurement, control, and automation devices and equipment controlled by a Programmable Logic Controller (PLC). The data acquired from the system was displayed and recorded using the same equipment. The system was equipped with analog and digital input/output channels, which can be expanded if needed. The switch regulated the initial power supply to the system.
The system utilized an SV2 Series DVP28SV11T2 type PLC (Delta Electronics Inc., Taipei, Taiwan). This module is equipped with 16 digital inputs and 12 transistor outputs. The Real Time Clock (RTC) capability allows the real-world clock to continue operating even after prolonged power interruptions to the PLC. The CPU is equipped with integrated RS-232 and RS-485 connections, allowing it to establish communication with all devices using the MODBUS protocol.
The Delta DVP-04AD functions as an analog signal reading card. Four analog input ports are available, each capable of reading ±10 V DC or ±20 mA analog signals. The Ladder programming language was employed with WPLSoft to program the PLC in the system. The touch screen was programmed using DOPSoft software.
This system monitors and regulates the microgreen growth compartment’s temperature, humidity, and carbon dioxide levels. It also controls the wavelength of LED illumination, ventilation, and irrigation scheduling. The equipment’s controls and settings are operated using a touchscreen interface with an HDMI monitor. The HDMI 7″ monitor features a TFT screen that displays 65.536 colors. It has 1 COM port, one internal COM port, USB host, and USB client capabilities.
The ESHT-102-W50 is a wall-mounted temperature and humidity sensor (Sisel Engineering Electronics Industry and Trade Inc., Istanbul, Türkiye) that weighs 125 g. It has a temperature range of −40°C to +125°C and a humidity range of 0 to 100% RH. The humidity accuracy is within ±4% RH for the entire 0 to 100% RH range, while the temperature accuracy is within ±1°C for the 0–70°C range. It can be powered by a 15–35 V DC or 10–25 V AC supply. The sensor can select the output as 0–20 mA DC or 0–10 V DC.
The carbon dioxide sensor accurately quantifies the carbon dioxide concentration in the surrounding environment and turns it into output signals according to specified standards (such as 0–10 V, 0–3 V, 4–20 mA). The sensor is powered by a direct current supply of 12–24 V. It has an output range of 0–10 V or 4–20 mA. The sensor’s measurement range is 0–5,000 parts per million, with a sensitivity of ± 10 and an accuracy of ± 25 parts per million.
RGB LEDs were utilized for the illumination system. The LED system is highly luminous and capable of emitting light on many wavelengths. It consists of 60 LEDs per meter.
The microgreens growing cabinet camera system was employed to monitor and observe the growth cycle of the plant species cultivated in the cabinet, starting from the seed planting stage. The HD 1080P wireless indoor WIFI IP camera allows for remote connection, provides night vision capabilities, and allows live viewing.
The irrigation system for the micro greenery growing cabinet utilized a medium-pressure diaphragm water pump (850-885-2526, 3.6LT/MIN Suction Discharge Pump Max. 15 Bar, Sismist Air Conditioning Industry Trade Co. Ltd., Bursa, Türkiye). Additionally, two high-pressure mist spray brass nozzle heads were employed to ensure uniform irrigation in each tray. The optimal operating pressure for the water pump is 9–10 bar, the sound level is 45 decibels, the working voltage is 220 V or 24 V direct current, the working current is 5 A, and the maximum working humidity is 95%.
The system utilized two AYNEFY 220 V 100 W ceramic air heaters to heat the cabinet. It also contains four superior-quality FZY12038HA2BL-type fans. Two of these fans heat, while the remaining two cool.
2.4 Operator panel screen design
The touch screen allows all settings and changes implemented in the created micro green cabinet (Power Automation Ltd. Co., Tekirdağ, Türkiye). The main page provides access to the following pages: automatic control, manual control, alarm page, system settings, and contact pages.
The automatic control tab is accessed to initiate or terminate the system’s automated functioning. From the system settings, the cabinet’s current temperature, humidity, and CO2 levels can be monitored, as well as the input predefined values. The desired CO2 level is input on this webpage. Initiating the system by pressing the Start button triggers operation based on the input values on the System Settings page. The devices inside the system can be manually controlled by accessing the manual control tab.
A system settings page allows users to configure the cabinet’s operating hours and the environmental conditions during those hours. This includes settings for the cabinet interior, lighting color, cabinet temperature setpoint, and plant watering time.
2.5 Determination of the performance of the microgreen plant growing cabinet
The research employed an experimental design to compare micro-greens production in a growing cabinet with production in an acclimatization laboratory, where production can be conducted under controlled conditions. The microgreen growth cabinet was kept under controlled conditions with a constant temperature of 22°C, constant humidity of 51%, and a maximum CO2 level of 400 ppm. The acclimation laboratory maintained a production environment with a temperature of 23°C during the day and 16°C at night, a humidity level of 60%, a photoperiodic pattern of 14 h of light and 10 h of darkness, and a light intensity of 400 μmol m−2 s−1.
The research was conducted in a microgreen cabinet and acclimatization laboratory, following a coincidence plot experimental design with three replications (Table 1). In the microgreens growing cabinet experiment, three plant species (P: purslane; A: amaranth; L: leek) were employed in each replication, as described by Düzgüneş et al. (1987).
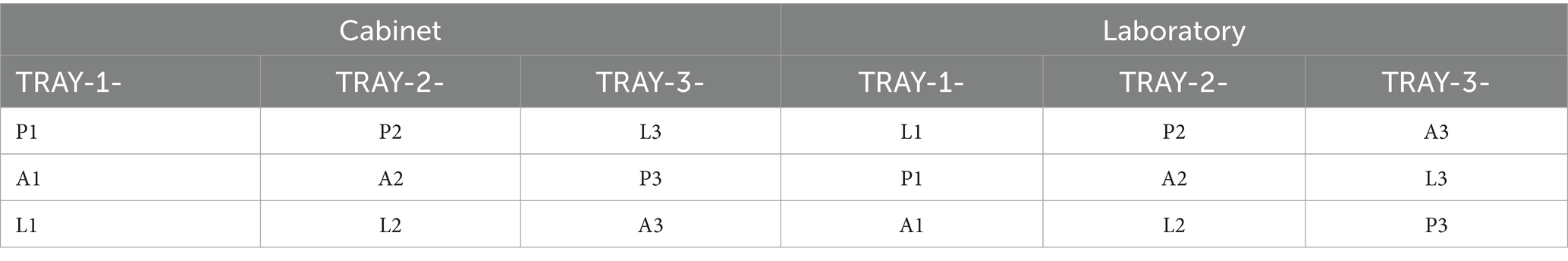
Table 1. Microgreen cabinet and laboratory experimental design.
During the research, the development stages of microgreens were examined daily, and data were recorded. The microgreens cultivated in the microgreen cabinet and acclimatization laboratory were typically harvested 7–10 days after planting the seeds. The harvesting process involved using a sharp and sterile knife to cut the microgreens from the plant’s root collar, as shown in Figures 2, 3.
![Panels showing sprouts of [a], [b], and [c] cultivated in trays within a growth cabinet under controlled conditions (top) and the corresponding harvested sprouts from each tray, packaged in plastic bags(bottom).](https://www.frontiersin.org/files/Articles/1620622/fsufs-09-1620622-HTML/image_m/fsufs-09-1620622-g002.jpg)
Figure 2. Microgreen growing cabinet conditions and harvested microgreens (a) Purslane-(Portulaca oleracea L.), (b) Amaranth-(Amaranthus hybridus L.), (c) Leek-(Allium ampeloprasum var. porrum L.).
![Panels showing sprouts of [a], [b], and [c] cultivated in trays under climate-controlled laboratory conditions (top) and the corresponding harvested sprouts from each tray, packaged in plastic bags (bottom).](https://www.frontiersin.org/files/Articles/1620622/fsufs-09-1620622-HTML/image_m/fsufs-09-1620622-g003.jpg)
Figure 3. Microgreen harvesting in the acclimatization laboratory (a) Purslane-(Portulaca oleracea L.), (b) Amaranth-(Amaranthus hybridus L.), (c) Leek-(Allium ampeloprasum var. porrum L.).
Following the harvest, plants were washed with purified water. The weight of the plants (in grams) was measured accurately and without delay using a precise balance. Subsequently, the plants were promptly stored in a deep freezer at a temperature of −20 ± 1°C and left under these conditions until further examination.
As illustrated in Figure 2, the harvested microgreens grown under fully automated cabinet conditions exhibited species-specific morphological characteristics. Purslane (Portulaca oleracea L.) (a) demonstrated compact and uniform canopy development, indicative of synchronized growth. Amaranth (Amaranthus hybridus L.) (b) was characterized by slender, reddish-hued hypocotyls and delicate leaf structures, while leek (Allium ampeloprasum var. porrum L.) (c) developed erect, elongated shoots with a notable consistency in stem height and pigmentation. These phenotypic attributes visually reinforce the quantitative biochemical and functional outcomes reported in this study, highlighting the efficacy of the automated system in ensuring controlled and reproducible growth dynamics across species. In particular, the morphological uniformity observed in purslane and leek samples suggests that the cabinet environment provided optimal microclimatic stability, contributing to reduced phenotypic variation. Such structural uniformity, beyond their visual appeal, plays a critical role in consumer acceptability and marketability, often exhibiting a positive correlation with biochemical quality parameters. The elevated vitamin C concentrations and antioxidant capacities measured in cabinet-grown samples further substantiate the nutritional advantages of precision-controlled cultivation (Di Gioia et al., 2023). Collectively, these morphological and compositional observations affirm the potential of the developed automated system as a robust platform for the sustainable production of high-quality microgreens in controlled-environment agriculture (Gupta et al., 2023).
2.6 Analytical methods for evaluating microgreen composition
The plant samples acquired at the end of the research were examined for vitamin C, protein content, various macro and micronutrients (nitrogen, phosphorus, potassium, calcium, magnesium, copper, manganese, iron, and zinc), and total antioxidant levels. Techniques employed in the corresponding analyses are listed below. Based on the study conducted by Benton et al. (1991), a modified micro Kjeldahl method was used to determine the crude protein content, while a titrimetric approach was employed to measure the vitamin C content. The Kjeldahl method was used to determine the total nitrogen content, as described by Kaçar and İnal (2010). The Ca and Mg contents were determined using the EDTA titration method, whereas the P and K contents were measured using an ICP-OES instrument (Sağlam, 2008). Lindsay and Norvell (1978) studied the elements Cu, Zn, Mn, and Fe. For the analysis, the plant samples underwent extraction and wet combustion, and the elements were then measured using an ICP-OES instrument. Total antioxidant capacity was calculated using the DPPH (1,1-Diphenyl-2-picrylhydrazyl radical; C18H12N5O6) radical scavenging activity, which uses a spectrophotometric approach.
As depicted in Figure 3, the microgreens cultivated in the laboratory displayed noticeable interspecies variation in their morphological development. Purslane (Portulaca oleracea L.) (a) appeared less compact compared to cabinet-grown samples, with less uniform canopy structure. Amaranth (Amaranthus hybridus L.) (b) showed increased etiolation and heterogeneous stem development, while leek (Allium ampeloprasum var. porrum L.) (c) exhibited inconsistent shoot height and curling in the apical regions, suggesting environmental stress. These phenotypic differences suggest that variations in temperature, humidity, and photo period in the laboratory environment may have led to suboptimal developmental regulation. Recent studies emphasize that morphological disparities under fluctuating environmental conditions can significantly impact phytochemical accumulation and physiological integrity of microgreens (Di Gioia et al., 2023). In particular, studies have demonstrated that relative instability in ambient growth conditions leads to increased morphological heterogeneity and reduced nutrient retention (Flores et al., 2024). In this context, Figure 3 not only illustrates the visual outcomes of environmental variability but also highlights the necessity of precision agriculture tools such as automated cabinets to ensure consistency in both structural and biochemical parameters of microgreen crops.
2.7 Statistical analysis
Statistical analyses were conducted to evaluate the nutritional and functional differences among microgreens grown under various conditions. One-way analysis of variance (ANOVA) was used to determine statistically significant differences between groups, followed by Tukey’s post-hoc test (p < 0.05). These tests were conducted using SPSS 20.0 software (SPSS Inc., Chicago, IL, USA). Additionally, multivariate statistical techniques, including Principal Component Analysis (PCA), Principal Coordinate Analysis (PCoA), Hierarchical Cluster Analysis (HCA), and Pearson Correlation Analysis, were employed to evaluate the patterns, similarities, and interrelationships among variables. All multivariate analyses were conducted using JMP Pro 17 software (SAS Institute Inc., Cary, NC, USA).
3 Results and discussion
3.1 Comparative analysis of nutritional and functional components in microgreens
Microgreens, a hypothetical term for the emerging food product developed from various commercial food crops such as vegetables, grains, and herbs, consist of developed cotyledons and partially expanded true leaves (Bhaswant et al., 2023). Microgreens are superfoods that can be produced in urban and peri-urban environments. They have a short growth cycle and minimal external nutrients (Wojdyło et al., 2020). They are grown in different light and growth media, resulting in fully developed cotyledons with one or two true leaves (Di Bella et al., 2020). Microgreens are cultivated in small-scale settings and alongside large-scale commercial production of vegetables and edible flowers. With their rapidly increasing popularity and consumer demand, various commonly grown vegetable species have been utilized for microgreen cultivation. These species belong to various botanical families, reflecting a rich biodiversity. For instance, members of the Amaranthaceae family (amaranth, chard, quinoa, spinach, buckwheat, beet), Amaryllidaceae (garlic, onion, leek), Apiaceae (parsley, carrot, fennel, celery, dill, chervil, coriander), Asteraceae (lettuce, radicchio, chicory, endive, tarragon, common dandelion), Boraginaceae (phacelia), Brassicaceae (radish, watercress, arugula, broccoli, cauliflower, cabbage, wild rocket), Convolvulaceae (water spinach), Cucurbitaceae (melon, cucumber, squash), Malvaceae (jute mallow), Poaceae (corn, lemongrass), Lamiaceae (chia), Leguminosae (chickpea, alfalfa, bean, green bean, fenugreek, fava bean, lentil, pea, clover), Onagraceae (evening primrose), and Portulacaceae (common purslane, moss-rose purslane) have all been employed in microgreen production. The diversity of these species contributes to the nutritional value of microgreens and enhances their functional properties, positioning them as valuable components within the category of functional foods (Di Bella et al., 2020; Ebert, 2022; Marchioni et al., 2021; Tan et al., 2020).
Environmental factors that affect plant growth are light (photoperiod, light intensity, and spectrum mix), relative humidity, temperature, electrical conductivity (EC), pH, nutrients, and moisture (Rusu et al., 2021). Of these factors, light is an important regulator of microgreen growth and quality (Frutos-Totosa et al., 2023). Microgreens are packed with nutrients. They have lots of essential elements such as iron (Fe), zinc (Zn), potassium (K), calcium (Ca), nitrogen (N), phosphorus (P), sulfur (S), manganese (Mn), selenium (Se), and molybdenum (Mo). They also have bioactive phytochemicals, which are good for health and could have therapeutic effects. They contain more bioactive compounds than mature plants. These include things like ascorbic acid, phylloquinone, and α-tocopherol. Their unique phytochemical profile makes them valuable as a food (Choe et al., 2018). The evaluation involved analyzing microgreens’ macro and micronutrient content grown in a laboratory setting and using a fully automated microgreen plant growing cabinet. The results are presented in Table 2. The data presented in Table 2 were statistically analyzed using a one-way analysis of variance (ANOVA) test at a 95% confidence interval. No statistically significant disparity was observed in the macro and micronutrient composition of microgreens cultivated in cabinet and laboratory settings. The macro and micronutrient contents of leek, amaranth, and purslane samples were grown in a fully automated micro-green production cabinet, and the laboratory was closed. Nitrogen (N) level was high in all species and ranged between 6.14 and 6.20% in leek, 6.86–6.92% in amaranth, and 7.50–7.62% in purslane. Phosphorus (P) and potassium (K) values were similarly highest in purslane; they were measured between 13.337 and 13,789 ppm and 28.233–27.340 ppm, respectively. Calcium (Ca) and magnesium (Mg) contents varied between 5.547 and 10,805 ppm and 2.791–9,418 ppm, respectively. In these parameters, the Mg level in leek differed significantly depending on the production environment (in the cabin: 2.791 ppm; in the laboratory: 4.978 ppm; p < 0.05), while no significant difference was found in other species. According to the correlation analysis presented in Figure 4, Mg is strongly positively correlated with P (r = 0.959), K (r = 0.867), and N (r = 0.966). This indicates that Mg transport, in particular, is coordinated with other macronutrients and that synergistic effects in the production environment may influence this relationship. The study results indicate that the macro and micronutrient levels of microgreens cultivated in cabinet and laboratory environments are within acceptable thresholds. The findings align with prior research conducted by Mir et al. (2017) and Xiao et al. (2015). Zhang et al. (2020) investigated the effects of LED light on phytochemical content in sprouts and microgreens. LED light enhanced the accumulation of various phytochemicals, including phenolic compounds, vitamins, glucosinolates, chlorophyll, and carotenoids. Antioxidant capacity was significantly increased under UV-B light. The accumulation of minerals such as Ca2+, Fe2+, and K+ also increased. LED lighting effectively produces sprouts and microgreens with higher nutritional value, depending on the plant species (Zhang et al., 2020). Similar natural interspecific variations were observed in microelements such as iron (Fe), copper (Cu), zinc (Zn), manganese (Mn), sodium (Na), and boron (B); however, environmental differences were limited. For example, Zn content in purslane was found to be quite high (217.53–236.84 ppm), whereas in amaranth this value remained at low levels, such as 38.85–40.17 ppm. Significant differences in some elements were observed in leek samples. While Na level was 1126.57 ppm in the laboratory environment, it was measured as 745.38 ppm in the cabin environment, and this difference was statistically significant (p < 0.05). Similarly, B content in leek was higher in the cabin environment (38.60 ppm) and lower in the laboratory (24.53 ppm) (p < 0.05). As shown in Figure 4, Cu was strongly negatively correlated with K (r = −0.978), Mg (r = −0.757), and N (r = −0.814). This suggests that Cu uptake may vary inversely with some macroelements and that ionic competition may play a role. Similarly, a very high positive correlation (r = 0.998) was found between Zn and Mn, suggesting that the uptake mechanisms of these two microelements are co-regulated. Vaštakaite and Viršile (2015), investigated whether different types of light lamps affect the nutritional quality of microgreens belonging to the Brassicaceae family. Higher anthocyanin and mineral element contents (Ca, K, Mg, Na, P, Fe, and Zn) were observed under 50 and 250 LED light conditions (Vaštakaite and Viršile, 2015). When the relationships between Dry Matter and macro–micro elements are analyzed, it is seen in Figure 4 that this parameter shows high negative correlation with N (r = −0.984), P (r = −0.973), K (r = −0.959), Mg (r = −0.933) and Fe (r = −0.637). This suggests that samples with higher mineral contents may also have higher water content, thus the percentage of dry matter may remain relatively lower. In this context, it is evaluated that humidity and temperature control in the production environment play a critical role in dry matter balance, and this balance is successfully achieved thanks to the automated cabin. At the same time, a positive correlation was observed between dry matter and protein (r = 0.882), and it is understood that protein concentration tends to increase as the dry matter ratio increases. This correlation confirms that the production cabinet provides a platform to support protein biosynthesis by controlling the physical environmental conditions. This system is an advantageous alternative for functional food production.
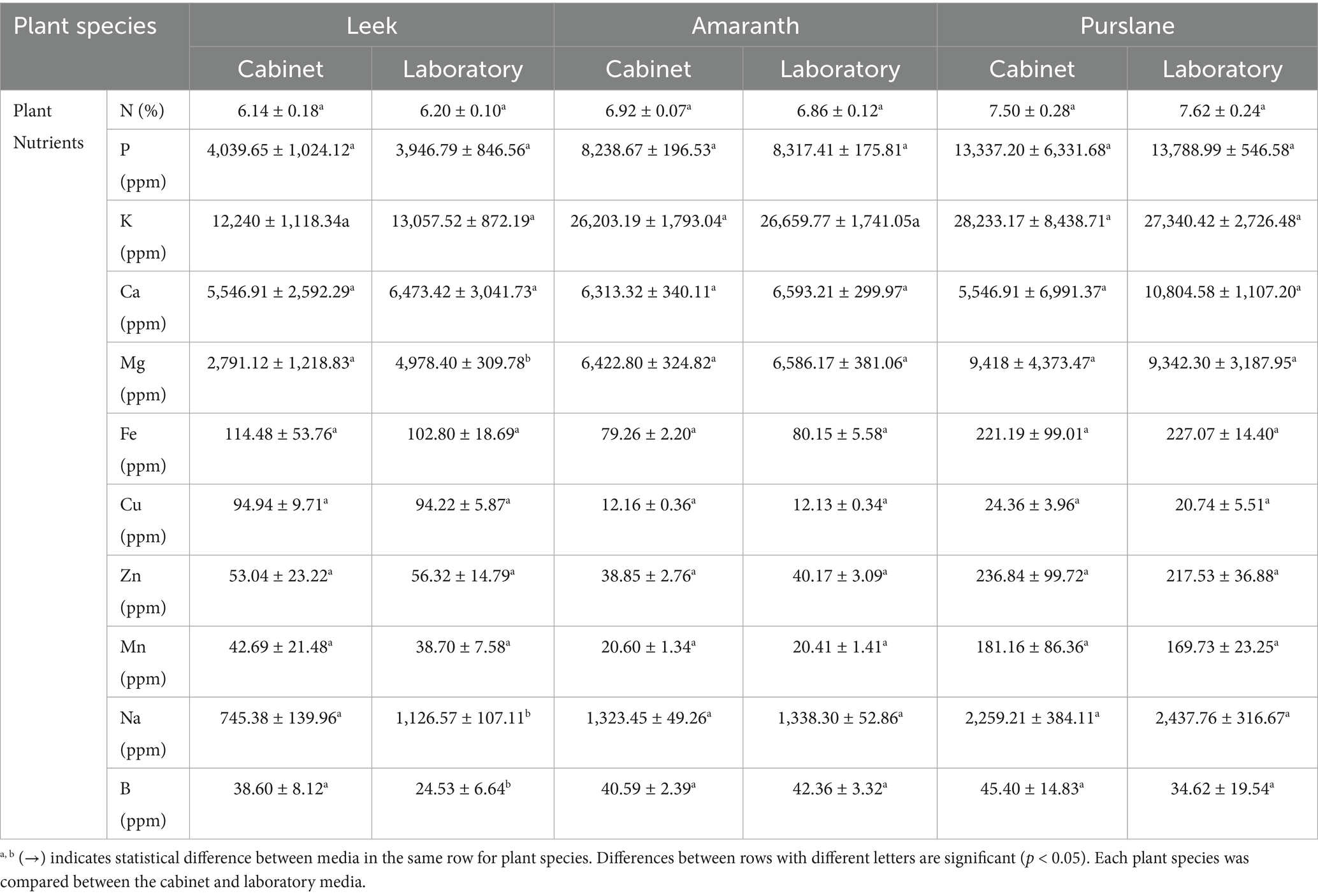
Table 2. Some macro-micronutrients (% and ppm) in microgreens grown under cabinet and laboratory conditions.
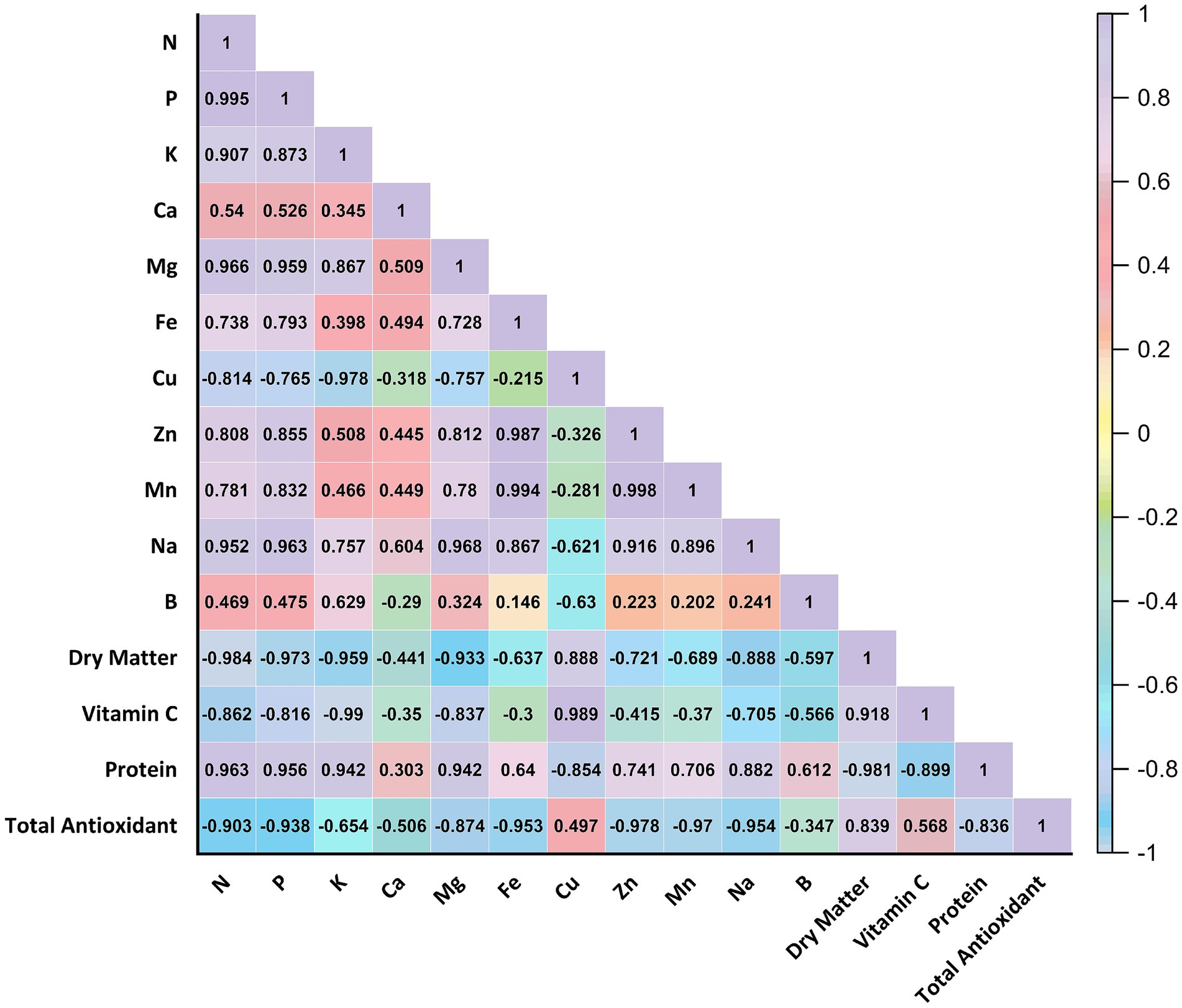
Figure 4. Pearson correlation matrix showing the relationships among macro- and micro-element content, dry matter, vitamin C, protein content, and total antioxidant capacity of microgreens. Strong positive and negative correlations are highlighted.
Recent advances in smart agriculture have introduced artificial intelligence and sensor-based systems into controlled-environment microgreen cultivation, which has increased substantially. These technologies enable the continuous monitoring and analysis of critical environmental parameters such as temperature, humidity, light spectrum, and CO2 concentration, thereby facilitating the optimization of growth efficiency and nutritional quality. These systems have been successfully employed to simulate plant environment interactions and to predict nutrient dynamics under varying light conditions. Furthermore, these digital infrastructures ensure consistency in the synthesis of functional compounds and allow for the implementation of adaptive management strategies through real-time feedback control mechanisms (Ojo and Zahid, 2022).
Due to their short growth cycles and high metabolic activity, microgreens are particularly sensitive to even minor fluctuations in environmental conditions, making them especially well-suited for data-driven precision cultivation systems. The integration of AI and automation improves productivity and quality control and contributes to the development of sustainable and traceable food systems. These advancements pave the way for more resilient agricultural practices that can respond swiftly to changing conditions. As a result, growers can optimize resource use, reduce waste, and ultimately enhance food security in a rapidly evolving climate landscape (Sharma and Shivandu, 2024). This holistic approach encourages the adoption of innovative technologies, empowering farmers to make informed decisions based on real-time data. By fostering collaboration between technology developers and agricultural experts, the industry can further enhance its capacity to meet global food demands while minimizing environmental impact (Kumari et al., 2025).
The investigation assessed the dry matter, vitamin C, protein, and total antioxidant values of microgreens cultivated in a laboratory using a fully automated microgreen plant growing cabinet. The results are presented in Table 3. The data were statistically analyzed using a one-way analysis of variance (ANOVA) test, with a 95% confidence interval. When the basic nutritional and functional components of microgreen species grown in different production environments (fully automated cabin and laboratory) were compared, no statistically significant differences were observed for most parameters due to production environment (p > 0.05). Dry matter contents of leek, amaranth, and purslane were 13.56–13.96%, 9.22–9.30%, and 6.74–6.83%, respectively. These values reflect the natural variations due to physiological differences among microgreen species and show that the production environment has no significant effect on this parameter. As shown in Figure 4, dry matter content is negatively correlated with many macroelements, especially nitrogen (r = −0.984) and phosphorus (r = −0.973). This strong inverse relationship suggests that the percentage of dry matter may decrease due to increased water content in tissues where minerals are dense; thus, high mineral intake may dilute tissue density. In contrast, the positive correlation between dry matter and protein (r = 0.882) suggests that drier tissues are also rich in protein, and this relationship is maintained regardless of the production environment.
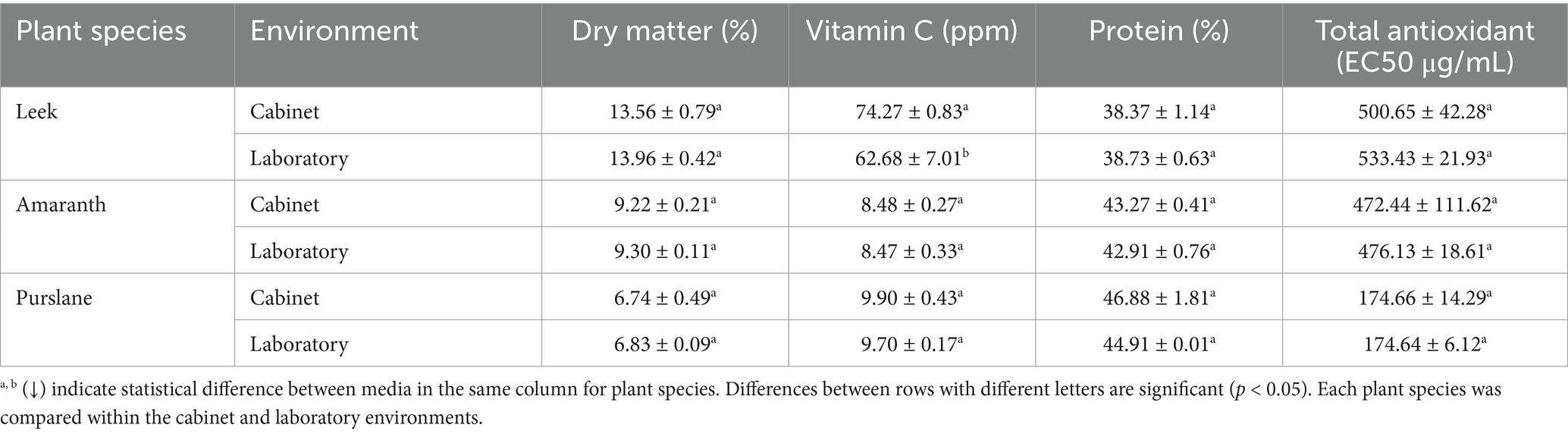
Table 3. Dry matter content (%), vitamin C content (ppm), protein content (%), and total antioxidant (EC50 μg/ml) values of microgreens grown under cabinet and laboratory conditions.
Regarding vitamin C content, only leek samples showed a significant difference (p < 0.05) depending on the production environment; the samples grown in the cabin environment had a higher level of 74.27 ppm, while the value in the laboratory environment remained at 62.68 ppm. This indicates that the controlled environmental conditions provided in the automated production cabinet increased vitamin C stability, since ascorbic acid is susceptible to external factors such as temperature, light, and oxygen. In one study, microgreens contained six times more vitamin C than their mature counterparts, while garnet amaranth had a significantly higher total ascorbic acid content than its mature stage (De la Fuente et al., 2019). Meas et al. (2020) observed that different light spectra changed the vitamin C content of amaranth microgreens (Meas et al., 2020). Figure 4 shows a robust positive correlation between vitamin C and Fe (r = 0.989). While this relationship can be explained by the role of ascorbic acid in enhancing iron absorption, it also indicates that these two compounds tend to accumulate together in plant tissue. The negative correlation between vitamin C and Zn (r = −0.415) and Mn (r = −0.705) indicates that high levels of these elements may have possible inhibitory effects on ascorbic acid metabolism. Therefore, it is necessary to optimize the production environment not only at the level of individual components but also in terms of their interactions with each other. According to a comprehensive review by Gupta et al. (2023), microgreens are emerging as “budding living functional foods”; their nutritional content can be significantly increased through genetic crossbreeding, biofortification and OMICS techniques (Gupta et al., 2023). The increase in vitamin C (or specific antioxidants) obtained in this study appears to be consistent with the phenomenon of “increased nutritional value in microgreen production systems” described by Gupta et al. In particular, as frequently emphasized in the literature regarding the effect of controlled environments (e.g., cabinet systems), our results also show an increase in bioavailability with a decrease in environmental variability.
Microgreens, which have grown in popularity in recent years, are rich in phytochemicals, including phenolic compounds that serve an antioxidant purpose (Gök et al., 2024). When total antioxidant capacity (EC50 μg/mL) was evaluated, this parameter measured at 174.64–174.66 levels in purslane samples showed high antioxidant activity with a lower EC50 value than other species. This value was found in leek between 500.65 and 533.43 μg/mL and in amaranth between 472.44 and 476.13 μg/mL. Although there was no significant difference depending on the production environment, the correlation analysis in Figure 4 sheds light on the relationship of this compound with other parameters. Total antioxidant capacity was moderately correlated with vitamin C (r = 0.568) and protein (r = −0.836), especially with Cu (r = 0.497). Interestingly, Zn (r = −0.978) was negatively correlated with Mn (r = −0.907) and Mg (r = −0.874). These findings suggest that high concentrations of some trace elements may be inversely related to free radical scavenging systems and reveal that antioxidant capacity is sensitive not only to phenolic compounds but also to the microelement composition of the medium. Similarly, Lee et al. (2025) reported that daily consumption of red beet and red cabbage microgreens is feasible and tolerable in healthy middle-aged individuals. In particular, red cabbage microgreen consumption improved symptoms associated with gastrointestinal inflammation (p = 0.047). This human intervention study demonstrates that microgreens are promising not only in terms of their content but also in terms of their functional effects, and supports the idea that these products can be provided with more consistent quality through controlled production systems (Lee et al., 2025). Overall, the fully automated production system can maintain microgreens’ nutritional quality and functional potential and optimize this potential in certain cases. Boonrat et al. (2025) successfully mapped the time-dependent changes in total phenolic and flavonoid content in the leaves and stems of sunflower microgreens using hyperspectral imaging and machine learning methods, showing that flavonoid accumulation peaked in the leaves on the 5th day, while the increase in the stems occurred later (Boonrat et al., 2025). These results, integrated with our digitally controlled cabinet system, provide a concrete foundation for determining the optimal harvest time. In addition, spectral signatures enable the separation of different tissue groups based on their nutritional components. This allows the system to be adjusted according to micro-nutrient profiles in precision production, thereby increasing the efficiency of functional food production. The analytical results indicate that the dry matter, vitamin C, protein, and total antioxidant values of microgreens cultivated in cabinet and laboratory conditions were within acceptable thresholds. The findings of this study are consistent with the results of prior research conducted by Kaçar and İnal (2010), Mir et al. (2017) and Xiao et al. (2015).
The PCA sample scatter plot in the left panel of Figure 5 shows how the microgreens are differentiated by different species (leek, amaranth, purslane) and production environments (cabin and laboratory). Principal component 1 (PC1) explains 74.4% of the variance, and Principal component 2 (PC2) explains 16.3%, totaling 90.7% of the variance represented by these two axes. It can be seen on the graph that purslane-cabin and purslane-laboratory samples are located quite far away from each other, indicating that the production environment has a significant effect on purslane samples. Similarly, leek-cabin and leek-laboratory samples are also separated along the axis, indicating that the environmental difference may also significantly affect leek. On the other hand, the amaranth-cabin and amaranth-lab samples are located very close to each other in the PCA plane, suggesting that amaranth is less affected by changes in the production environment and has a stable structure in terms of content.
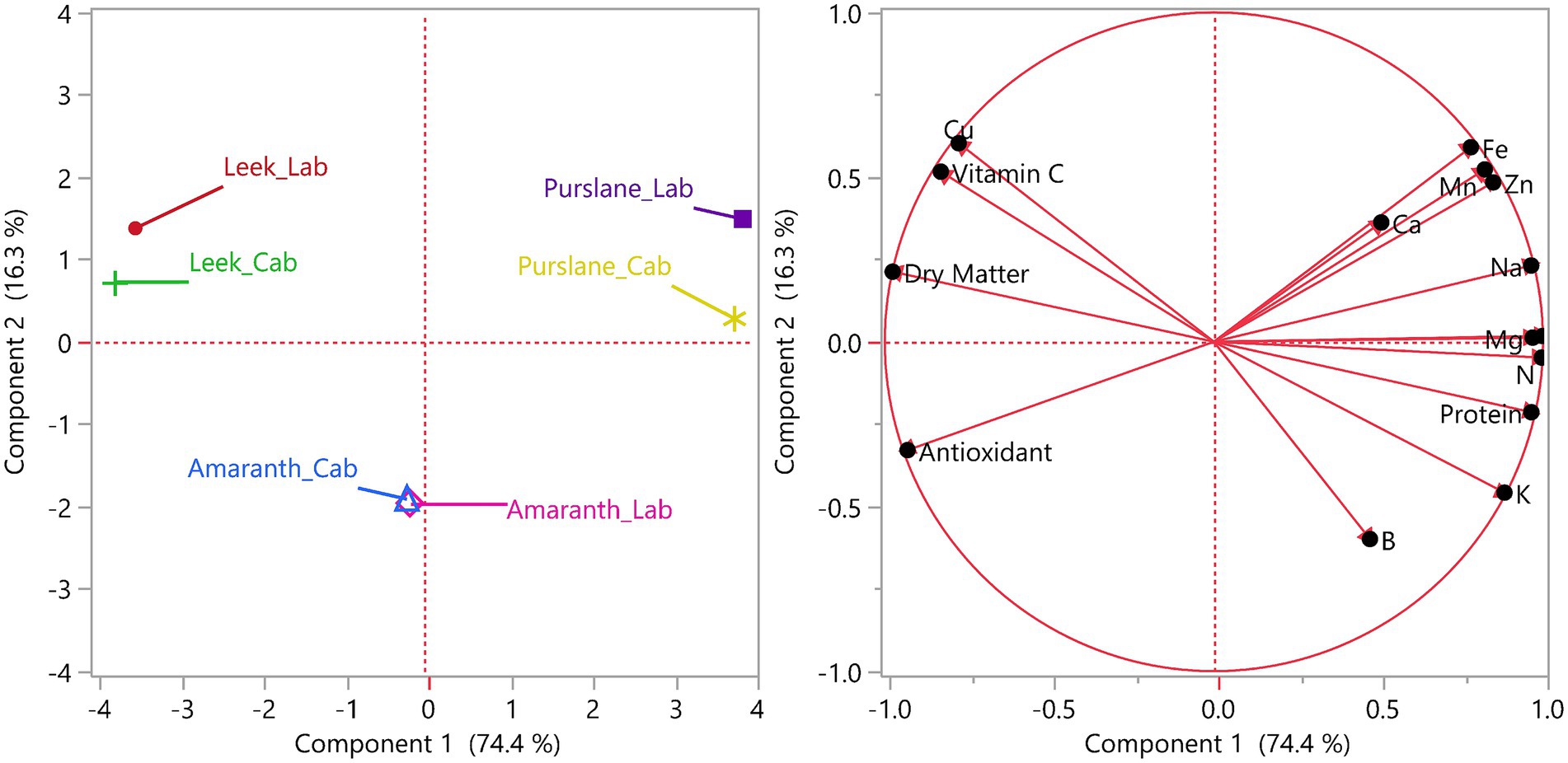
Figure 5. Results of principal component analysis (PCA) of microgreen samples by species (leek, amaranth, purslane) and production environments (cabin and laboratory).
The right panel of Figure 5 presents the PCA loadings plot, showing the contributions and orientations of the variables used in the analysis on PC1 and PC2. In the graph, nutrients such as Fe, Mn, Zn, Ca, Na, P, Mg, N, and Protein are grouped in the right half-plane, showing that they tend to change together and represent the positive direction of PC1. On the other hand, Cu, vitamin C, dry matter, and total antioxidant capacity were located in the left half plane and were associated with the negative direction of PC1. This distribution reveals that these parameters are related to the content profile arising from species differences and that these variables are decisive in the discrimination of samples. In particular, the fact that dry matter and antioxidant activity variables are located close to each other in the graph shows a positive relationship between these two parameters. At the same time, some elements such as boron (B) and potassium (K) exhibit more independent behavior by moving off-axis. Overall, Figure 5 shows that differences in content can be strongly differentiated based on both production environment and plant species, and that some essential compounds (e.g., vitamin C, Fe, protein) play a dominant role in this differentiation.
Figure 6 shows the hierarchical clustering analysis according to the similarities of macro and micronutrients and functional traits determined in microgreens. As a result of the study, parameters such as nitrogen (N), dry matter, protein, vitamin C, Cu, Fe, Zn, Mn, and total antioxidant capacity were found to be in the same central cluster with similar variation trends. The high positive correlations in Pearson correlation analysis in Figure 4 support this finding. In particular, the close positioning of Fe, Mn, and Zn with antioxidant capacity reflects the functions of these microelements in plant oxidative processes. In contrast, potassium (K) was separated from all other parameters, forming a singular cluster. This suggests that the variation of potassium is independent of the different components and may be more related to interspecific differences. In addition, P, Mg, and Ca were found in close subgroups, suggesting that these minerals are linked to standard biochemical and structural functions.
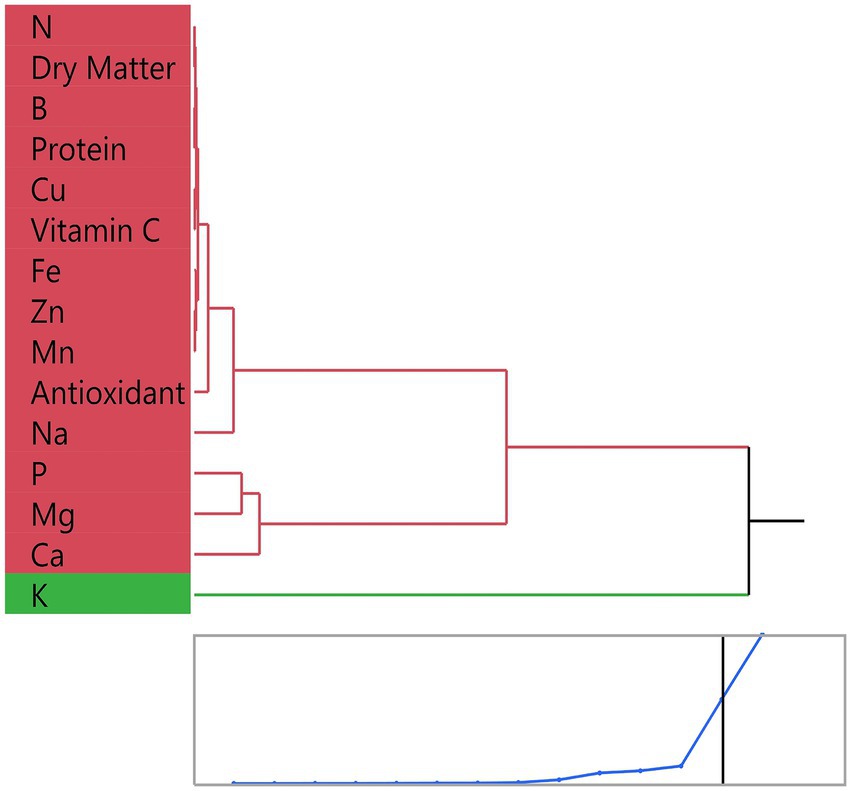
Figure 6. Multivariate analysis of the relationships among nutritional and functional components in microgreens. Hierarchical clustering dendrogram of measured parameters, showing grouping of traits based on similarity.
Through multivariate analyses, Figures 7, 8 visualize the relationships between measured parameters and the differences between sample groups. Figure 7, Principal Coordinate Analysis (PCoA) biplot, shows that parameters such as nitrogen, dry matter, protein, vitamin C, and total antioxidant capacity are located close in the analysis plane, indicating that these traits tend to vary together through common variance. On the other hand, potassium, calcium, and magnesium are located far away from the others, indicating that they have more independent sources of variation. Figure 8 presents the pairwise comparative distribution matrix of the samples (combinations of plant species and production media). This matrix shows that the purslane-cabin samples exhibit a different distribution from the other combinations, especially regarding calcium content. In contrast, leek-cabin and leek-laboratory samples clustered in a similar region, suggesting that the leek species gives stable results regarding nutritional and functional properties even if the production environment changes. This indicates that some species can maintain their intrinsic physiological balance more stably regardless of the production system. When Figures 6–8 are evaluated as a whole, they provide the opportunity to holistically analyze the biochemical relationships between parameters and the multidimensional structure of species-medium combinations.
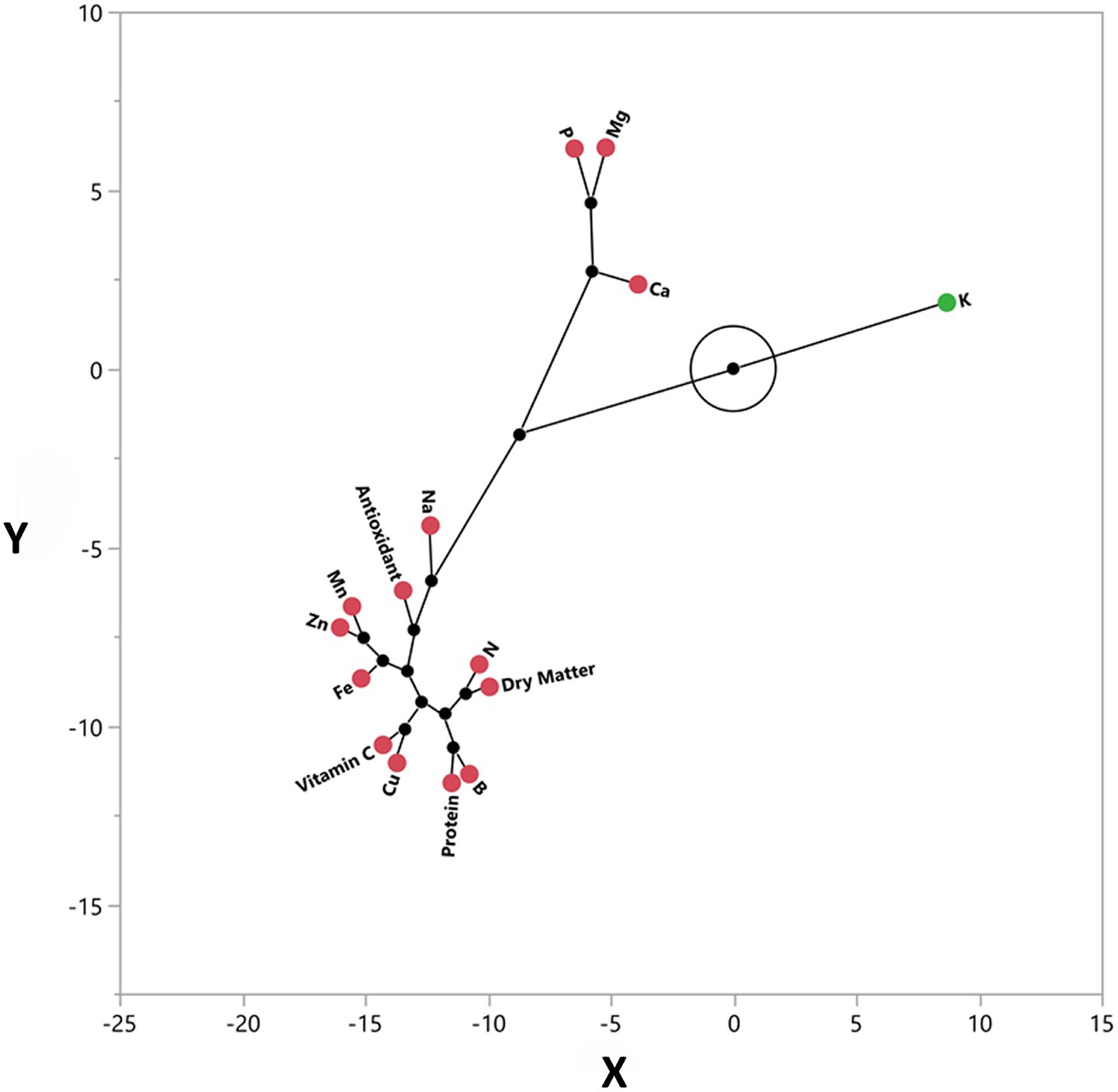
Figure 7. Multivariate analysis of the relationships among nutritional and functional components in microgreens. Principal Coordinate Analysis (PCoA) biplot displaying the distribution of key nutrients and bioactive components about each other; traits such as antioxidant activity, protein, and vitamin C were spatially close, indicating correlation.
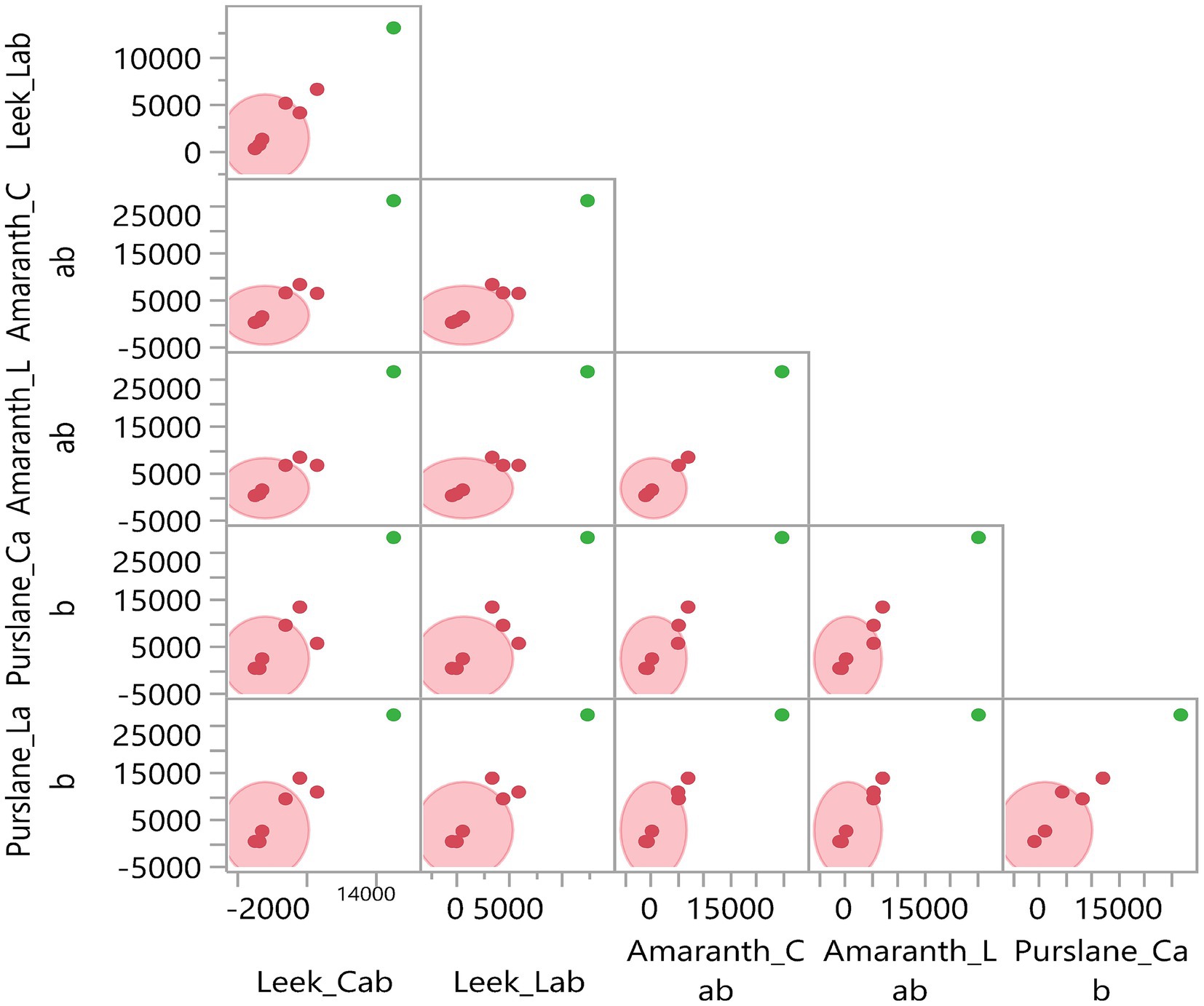
Figure 8. Multivariate analysis of the relationships among nutritional and functional components in microgreens. Pairwise comparison matrix among microgreen species and treatments, showing clustering patterns and variation among species (Leek, Amaranth, Purslane) and growing conditions (Cabinet and Laboratory).
4 Future work and limitations of study
Although this study aimed to demonstrate the potential of an automation-supported cabin system in microgreen production, it has some limitations. First, the study was conducted on only three different plant species (leek, amaranth and purslane), and the system’s performance could not be evaluated on other microgreen species. In addition, the trials were conducted only for a particular production period and in a single geographical location, and the system’s effectiveness was not tested under different climatic and environmental conditions. However, a detailed evaluation of the commercial viability of the system in terms of economic analysis and scalability was not made; data on long-term production cycles were not collected. Future studies can fill these gaps by examining the system’s performance under different climatic and environmental conditions and various plant species and detailing the economic dimensions.
5 Conclusion
The fully automated microgreen growing cabinet developed in this study successfully maintained controlled environmental conditions, enabling consistent microgreens production with high nutritional and functional quality. While most macro- and micro-nutrient values did not significantly differ between cabinet and laboratory conditions, cabinet-grown leek samples exhibited higher vitamin C content. Strong correlations between antioxidant capacity and specific microelements emphasized the role of mineral composition in functional quality. Multivariate analyses highlighted distinct content variations based on species and environment, with amaranth showing notable stability. These findings suggest that automation-supported cultivation systems hold promise for sustainable, traceable, and high-value microgreen production in controlled-environment agriculture.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors without undue reservation.
Author contributions
FT: Writing – review & editing, Conceptualization, Methodology, Investigation, Formal analysis, Writing – original draft, Funding acquisition. FE: Formal analysis, Writing – original draft, Investigation, Writing – review & editing. FÖ: Writing – review & editing, Investigation, Writing – original draft, Formal analysis. SY: Writing – review & editing, Writing – original draft, Software, Investigation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Research Fund of Tekirdag Namık Kemal University. Project Number: NKUBAP.42.GA.22.356.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Benton, J. Jr., Jones, B. W., and Mills, H. A. (1991). Plant analysis handbook: a practical sampling, preparation, analysis, and interpretation guide. Athens: Micro-Macro Publishing.
Bhaswant, M., Shanmugam, D. K., Miyazawa, T., Abe, C., and Miyazawa, T. (2023). Microgreens—a comprehensive review of bioactive molecules and health benefits. Molecules 28:867. doi: 10.3390/molecules28020867
Boonrat, P., Patel, M., Pengphorm, P., Detarun, P., and Daengngam, C. (2025). Hyperspectral imaging for the dynamic mapping of total phenolic and flavonoid contents in microgreens. AgriEngineering 7:107. doi: 10.3390/agriengineering7040107
Choe, U., Liangli Lucy, Y., and Wang, T. T. Y. (2018). The science behind microgreens as an exciting new food for the 21st century. J. Agric. Food Chem. 66, 11519–11530. doi: 10.1021/acs.jafc.8b03096
Choudhary, V., Guha, P., Pau, G., and Mishra, S. (2025). An overview of smart agriculture using internet of things (IoT) and web services. Environ. Sustain. Indicat. 26:100607. doi: 10.1016/j.indic.2025.100607
De la Fuente, B., López García, G., Máñez, V., Alegría, A., Barberá, R., and Cilla, A. (2019). Evaluation of the bioaccessibility of antioxidant bioactive compounds and minerals of four genotypes of Brassicaceae microgreens. Foods 8:250. doi: 10.3390/foods8070250
Di Bella, M. C., Niklas, A., Toscano, S., Picchi, V., Romano, D., Lo Scalzo, R., et al. (2020). Morphometric characteristics, polyphenols and ascorbic acid variation in Brassica Oleracea L. novel foods: sprouts, microgreens and baby leaves. Agronomy 10:782. doi: 10.3390/agronomy10060782
Di Gioia, F., Hong, J. C., Pisani, C., Petropoulos, S. A., Bai, J., and Rosskopf, E. N. (2023). Yield performance, mineral profile, and nitrate content in a selection of seventeen microgreen species. Front. Plant Sci. 14, 1–14. doi: 10.3389/fpls.2023.1220691
Düzgüneş, O., Kesici, T., Kavuncu, O., and Gürbüz, F. (1987). Araştırma ve Deneme Metodları. Ankara Üniversitesi, Ziraat Fakültesi Yayınları 60:1021.
Ebert, A. W. (2022). Sprouts and microgreens—novel food sources for healthy diets. Plants 11:571. doi: 10.3390/plants11040571
Ebert, A. W., Wu, T. H., and Yang, R. Y. (2014). Amaranth sprouts and microgreens–a homestead vegetable production option to enhance food and nutrition security in the rural-urban continuum. In Proceedings of the Regional Symposium on Sutaining Small Scale Vegetable Production and Marketing Systems for Food and Nutrition Security (SEAVEG 2014), Bangkok, Thailand, 233–244.
Elijah, O., Rahman, T. A., Orikumhi, I., Leow, C. Y., and Hindia, M. H. D. N. (2018). An overview of internet of things (IoT) and data analytics in agriculture: benefits and challenges. IEEE Internet Things J. 5, 3758–3773. doi: 10.1109/JIOT.2018.2844296
Fasciolo, B., Panza, L., and Lombardi, F. (2024). Exploring the integration of industry 4.0 technologies in agriculture: a comprehensive bibliometric review. Sustainability (Switzerland) 16:8948. doi: 10.3390/su16208948
Flores, M., Hernández-Adasme, C., Guevara, M. J., and Escalona, V. H. (2024). Effect of different light intensities on agronomic characteristics and antioxidant compounds of Brassicaceae microgreens in a vertical farm system. Front. Sustain. Food Syst. 8, 1–15. doi: 10.3389/fsufs.2024.1349423
Frutos-Totosa, A., Hernández Adasme, C., Martínez, V., Mestre, T., Díaz Mula, H. M., Botella, M. Á., et al. (2023). Light spectrum effects on rocket and lamb’s lettuce cultivated in a vertical indoor farming system. Sci. Hortic. 321:112221. Elsevier. doi: 10.1016/j.scienta.2023.112221
Gök, S. B., Özdüven, F., and Açıkgöz, F. E. (2024). The effect of different harvest times on phenolic content and antioxidant activity in some microgreens. Kahramanmaraş Sütçü İmam Üniversitesi Tarım ve Doğa Dergisi 27, 417–422. doi: 10.18016/ksutarimdoga.vi.1216114
Gupta, A., Sharma, T., Singh, S. P., Bhardwaj, A., Srivastava, D., and Kumar, R. (2023). Prospects of microgreens as budding living functional food: breeding and biofortification through OMICS and other approaches for nutritional security. Front. Genet. 14:1053810. doi: 10.3389/fgene.2023.1053810
Kumari, K., Nafchi, A. M., Mirzaee, S., and Abdalla, A. (2025). AI-driven future farming: achieving climate-smart and sustainable agriculture. AgriEngineering 7, 1–30. doi: 10.3390/agriengineering7030089
Kyriacou, M. C., De Pascale, S., Kyratzis, A., and Rouphael, Y. (2017). Microgreens as a component of space life support systems: a cornucopia of functional food. Front. Plant Sci. 8, 8–11. doi: 10.3389/fpls.2017.01587
Lee, S. Y., Michell, K. A., Butler, M. M., Smith, B. T., Woolf, E. K., Holmes, S. C., et al. (2025). Feasibility and tolerability of daily microgreen consumption in healthy middle-aged/older adults: a randomized, open-label, controlled crossover trial. Nutrients 17:467. doi: 10.3390/nu17030467
Lenzi, A., Orlandini, A., Bulgari, R., Ferrante, A., and Bruschi, P. (2019). Antioxidant and mineral composition of three wild leafy species: a comparison between microgreens and baby greens. Foods 8:487. doi: 10.3390/foods8100487
Lindsay, W. L., and Norvell, W. A. (1978). Development of a Dtpa Soil Test for Zinc, Iron, Manganese, and Copper. Soil Sci. Soc. Am. J. 42, 421–428. doi: 10.2136/sssaj1978.03615995004200030009x)
Liu, Y., Ma, X., Shu, L., Hancke, G. P., and Abu-Mahfouz, A. M. (2021). From industry 4.0 to agriculture 4.0: current status, enabling technologies, and research challenges. IEEE Trans. Industr. Inform. 17, 4322–4334. doi: 10.1109/TII.2020.3003910
Marchioni, I., Martinelli, M., Ascrizzi, R., Gabbrielli, C., Flamini, G., Pistelli, L., et al. (2021). Small functional foods: comparative phytochemical and nutritional analyses of five microgreens of the Brassicaceae family. Foods 10:427. doi: 10.3390/foods10020427
McClements, D. J., Barrangou, R., Hill, C., Kokini, J. L., Lila, M. A., Meyer, A. S., et al. (2021). Building a resilient, sustainable, and healthier food supply through innovation and technology. Annu. Rev. Food Sci. Technol. 12, 1–28. doi: 10.1146/annurev-food-092220-030824
Meas, S., Luengwilai, K., and Thongket, T. (2020). Enhancing growth and phytochemicals of two Amaranth microgreens by LEDs light irradiation. Sci. Hortic. 265:109204. doi: 10.1016/j.scienta.2020.109204
Mir, S. A., Shah, M. A., and Mir, M. M. (2017). Microgreens: production, shelf life, and bioactive components. Crit. Rev. Food Sci. Nutr. 57, 2730–2736. doi: 10.1080/10408398.2016.1144557
Ojo, M. O., and Zahid, A. (2022). Deep learning in controlled environment agriculture: a review of recent advancements, challenges and prospects. Sensors 22:7965. doi: 10.3390/s22207965
Pearce-Higgins, J. W., Antão, L. H., Bates, R. E., Bowgen, K. M., Bradshaw, C. D., Duffield, S. J., et al. (2022). A framework for climate change adaptation indicators for the natural environment. Ecol. Indic. 136:108690. doi: 10.1016/j.ecolind.2022.108690
Pinto, E., Almeida, A. A., Aguiar, A. A., and Ferreira, I. M. P. L. V. O. (2015). Comparison between the mineral profile and nitrate content of microgreens and mature lettuces. J. Food Compos. Anal. 37, 38–43. doi: 10.1016/j.jfca.2014.06.018
Rusu, T., Cowden, R. J., Moraru, P. I., Maxim, M. A., and Ghaley, B. B. (2021). Overview of multiple applications of basil species and cultivars and the effects of production environmental parameters on yields and secondary metabolites in hydroponic systems. Sustainability 13:11332. doi: 10.3390/su132011332
Sağlam, M. T. (2008). Toprak ve Suyun Kimyasal Analiz Yöntemleri. Tekirdağ: Namık Kemal Üniversitesi.
Sharma, K., and Shivandu, S. K. (2024). Integrating artificial intelligence and internet of things (IoT) for enhanced crop monitoring and management in precision agriculture. Sens. Int. 5:100292. doi: 10.1016/j.sintl.2024.100292
Tan, L., Nuffer, H., Feng, J., Kwan, S. H., Chen, H., Tong, X., et al. (2020). Antioxidant properties and sensory evaluation of microgreens from commercial and local farms. Food Sci. Human Wellness 9, 45–51. doi: 10.1016/j.fshw.2019.12.002
Vaštakaite, V., and Viršile, A. (2015). Light—emitting diodes (LEDs) for higher nutritional quality of Brassicaceae microgreens. Res. Rural Dev. 1, 111–117.
Wojdyło, A., Nowicka, P., Tkacz, K., and Turkiewicz, I. P. (2020). Sprouts vs. microgreens as novel functional foods: variation of nutritional and phytochemical profiles and their in vitro bioactive properties. Molecules 25:4648. doi: 10.3390/molecules25204648
Xiao, Z., Lester, G. E., Park, E., Saftner, R. A., Luo, Y., and Wang, Q. (2015). Evaluation and correlation of sensory attributes and chemical compositions of emerging fresh produce: microgreens. Postharvest Biol. Technol. 110, 140–148. doi: 10.1016/j.postharvbio.2015.07.021
Xiao, Z., Rausch, S. R., Luo, Y., Sun, J., Yu, L., Wang, Q., et al. (2019). Microgreens of Brassicaceae: genetic diversity of phytochemical concentrations and antioxidant capacity. LWT 101, 731–737. doi: 10.1016/j.lwt.2018.10.076
Yap, C. K., and Al-Mutairi, K. A. (2024). A conceptual model relationship between industry 4.0—food-agriculture nexus and agroecosystem: a literature review and knowledge gaps. Foods 13, 169–172. doi: 10.3390/foods13010150
Zhang, X., Bian, Z., Yuan, X., Chen, X., and Lu, C. (2020). A review on the effects of light-emitting diode (LED) light on the nutrients of sprouts and microgreens. Trends Food Sci. Technol. 99, 203–216. doi: 10.1016/j.tifs.2020.02.031
Nomenclature
ANOVA - Analysis of Variance
B - Boron (ppm)
Ca - Calcium (ppm)
CO2 - Carbon Dioxide
Cu - Copper (ppm)
DM - Dry Matter (%)
DPPH - 1,1-Diphenyl-2-picrylhydrazyl (used in antioxidant capacity assay)
EC50 - Effective concentration for 50% scavenging activity (μg/mL)
Fe - Iron (ppm)
HCA - Hierarchical Cluster Analysis
HDMI - High-Definition Multimedia Interface
ICP-OES - Inductively Coupled Plasma - Optical Emission Spectrometry
IoT - Internet of Things
K - Potassium (ppm)
Mg - Magnesium (ppm)
Mn - Manganese (ppm)
Mo - Molybdenum (ppm)
N - Nitrogen (%)
Na - Sodium (ppm)
P - Phosphorus (ppm)
PCA - Principal Component Analysis
PCoA - Principal Coordinate Analysis
PLC - Programmable Logic Controller
ppm - Parts per million
RGB LEDs - Red-Green-Blue Light-Emitting Diodes
RS-232, RS-485 - Serial Communication Standards
RTC - Real-Time Clock
Se - Selenium (ppm)
SPSS - Statistical Package for the Social Sciences (software)
TFT - Thin-Film Transistor (screen technology)
Zn - Zinc (ppm)
Keywords: sustainable food processing, controlled environment agriculture, fully automated growth cabinet, microgreen production, multivariate statistical analysis
Citation: Taşcı Durgut F, Eryılmaz Açıkgöz F, Özdüven FF and Yıkmış S (2025) Sustainable microgreen production through automated cultivation systems: nutritional and functional profiling via multivariate analysis. Front. Sustain. Food Syst. 9:1620622. doi: 10.3389/fsufs.2025.1620622
Edited by:
Mohammad U.H. Joardder, Rajshahi University of Engineering & Technology, BangladeshCopyright © 2025 Taşcı Durgut, Eryılmaz Açıkgöz, Özdüven and Yıkmış. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Seydi Yıkmış, c3lpa21pc0Bua3UuZWR1LnRy; Funda Eryılmaz Açıkgöz, ZmVyeWlsbWF6QG5rdS5lZHUudHI=