 Lauren R. Stubbs
Lauren R. Stubbs Surendra Singh
Surendra Singh Donald Wysocki
Donald Wysocki Garett C. Heineck
Garett C. Heineck Haly L. Neely
Haly L. Neely Shikha Singh
Shikha Singh- 1Lind Dryland Research Station, Washington State University, Lind, WA, United States
- 2Department of Crop and Soil Sciences, Washington State University, Pullman, WA, United States
- 3Oregon State University Extension, Pendleton, OR, United States
- 4Prosser Irrigated Agriculture Research and Extension Center, Prosser, WA, United States
Changing weather conditions are having negative impacts on dryland (rainfed) crop production systems such as those found in the inland Pacific Northwest (iPNW). This region is dominated by winter wheat (Triticum aestivum L.) production and also produces alternative crops such as canola (Brassica napus L.) and peas (Pisum sativum L.). Producers need crop rotations and agronomic management strategies that are equally productive and more sustainable than traditional winter wheat-fallow systems. New crop rotations should prioritize crop water use efficiency (WUE), protect soil health, and manage herbicide-resistant weeds. Intercropping is one practice that can promote agroecosystem biodiversity, soil health, drought resilience, and resource use efficiency (RUE), among other ecosystem services. Spring pea and spring canola intercropping, also referred to as “peaola,” is emerging as an alternative crop rotation and management strategy for the dryland iPNW mainly because it increases RUE and WUE, while reducing production inputs. However, little is known about producing peaola in the iPNW. Therefore, this review seeks to address a wide range of topics, including (but not limited to) ecological and agronomic aspects of intercropped systems, the impact of peaola production on soil health, the potential of peaola to reduce pest pressure and improve drought resilience, and examples of barriers that may prevent the adoption of peaola in the iPNW. Findings indicate that peaola can provide agroecosystem benefits such as improved water infiltration, soil organic matter turnover, nutrient cycling, and increased land use efficiency. However, complexity of management, a lack of region-specific research, and marketing constraints are legitimate challenges barring the immediate adoption of peaola. Nonetheless, peaola has the potential to improve cereal crop production and promote sustainability in dryland agricultural systems across the iPNW.
1 Introduction
1.1 Current challenges in inland Pacific Northwest dryland cropping systems
Recognized as the largest biome on Earth, drylands are responsible for producing nearly 60% of the world’s food (Prăvălie, 2016; Osborne et al., 2022; Yadav et al., 2024). In the instance of wheat, a commodity that is a staple in diets around the world, three-quarters of the global crop (approximately 620 million tons) is produced in dryland growing regions (Adil et al., 2024). Production challenges such as water scarcity and ecosystem vulnerability will only be amplified by climate change, and are projected to worsen in dryland production regions, including the inland Pacific Northwest (iPNW), which will lead producers to adopt cropping systems that prioritize high crop water use efficiency and drought resilience (Yu et al., 2024; Strauss et al., 2021). Despite historically reliable yields over the years, the cereal monoculture cropping system that dominates iPNW production is the subject of much criticism due to the lack of biodiversity and ecological instability of this system (Kirby et al., 2017), as well as its high demand for inputs, such as fertilizer nitrogen (N), and low resource use efficiency (Maaz et al., 2018).
Dryland (rainfed) agricultural production in the iPNW, a geographical region comprised of eastern Washington state and surrounding areas of Idaho and Oregon, occupies approximately 3.4 million ha of land and is dominated by wheat (Triticum aestivum L.) production systems (Maaz et al., 2018; Schillinger, 2020; Schillinger et al., 2006). From west to east, the iPNW is characterized by three annual precipitation zones, referred to as the low rainfall (<300 mm), intermediate rainfall (300–450 mm) and high rainfall (450–600 mm) zones (Schillinger, 2020). Crop rotations vary by precipitation zone and are often centered around winter cereal crops. In the low and intermediate rainfall zones, two-year winter wheat-fallow rotations (in which a wheat crop is established once every two years with approximately 14 months of fallow between each crop) are implemented to store precipitation over the fallow winter for the subsequent wheat crop (Karimi et al., 2017; Kaur et al., 2022; Schillinger et al., 2006). In the high rainfall zone, and certain locations of the intermediate rainfall zone, annual cropping is possible, and producers may include a spring or winter pulse or oilseed crop for diversification (Maaz et al., 2018). While pulse crops were once a popular rotation in the iPNW, modern government-subsidized commodity programs, in addition to changes in production technology, have long favored wheat as the area’s most widely produced crop (Maaz et al., 2018).
Relatively low crop residue returns and long fallow periods from traditional two-year wheat-fallow rotations (which often involve tillage), have put a strain on the iPNW’s resources for nearly 140 years, as tillage has left the region’s soils vulnerable to wind and water erosion for decades and led to a decrease in soil organic matter (SOM) content (Schillinger, 2020). While these rotations have promoted stable wheat crop yields, the wheat-fallow system has resulted in a loss of over 50% of inherent SOM from the top 60 cm of soil profiles (Ghimire et al., 2015; Awale et al., 2022). While the adoption of no-till farming in the iPNW has been increasing steadily and has helped to curb soil erosion and rebuild SOM, no-till practices still face many barriers to adoption, including the high cost of purchasing no-till drills and high-horsepower tractors (Gelardi et al., 2023), as well as the limited potential of chemical no-till fallow in the low rainfall zone and parts of the intermediate rainfall zone due to the increased risk of seed zone soil water loss during hot, dry summer months (Schillinger et al., 2006). Additionally, no-till practices rely solely on herbicides to manage weed populations (Young et al., 2006). Failure to diversify weed control beyond chemical means, due to the use of homogeneous crop rotations, or a lack of chemical stewardship, has contributed to the rise of herbicide-resistant weed species in conventional and no-till iPNW cropping systems (Renton and Flower, 2015).
The issues of soil erosion, declining SOM, herbicide resistance, and variability in precipitation create significant challenges for growers in the dryland iPNW. It is apparent that the diversification of cropping systems is needed in this region, but a new, multifarious cropping system of any merit cannot sacrifice short-term economic productivity for long-term sustainability. For new crop rotation methodologies to be successful in dryland systems, the rotations must be low-input, and promote qualities such as agroecosystem biodiversity, soil health, more effective weed control, drought resilience, improved water use efficiency (WUE), and enhanced profitability. However, growers in the iPNW are expressing interest in alternative cropping systems, as demonstrated by the responses collected in the Washington State Department of Agriculture (WSDA)’s soil management survey (Gelardi et al., 2023). The majority (72%) of producers surveyed reported the use of reduced- or no-till practices on their farms and 37% of producers reported the use of two or more conservation practices together (Gelardi et al., 2023).
1.2 Intercropping: an unconventional “fix” for the inland Pacific Northwest
One such practice not mentioned in the 2022 WSDA survey was intercropping, or the practice of growing two or more crop species at the same time in the same field. Globally, intercropping has been practiced for centuries across an array of agricultural systems that differ drastically in climate, crop types, and farm size (Tang et al., 2024; Stomph et al., 2020). Perhaps the most well-known system is the Three Sisters, an intercrop of maize (Zea mays L.), bean (Phaseolus vulgaris L.), and squash (Cucurbita pepo L.) grown by the Indigenous Peoples of the Americas (Ngapo et al., 2021). Today, these practices are often restricted in their modern-day applications to smallholder cropping systems in developing countries, as the mass-mechanization of agriculture and the development of economies of scale have forced a reduction in the complexity of cropping systems (Li et al., 2023; Bybee-Finley and Ryan, 2018; Khanal et al., 2021). Additionally, conflicts over best land use practices and high land prices, as noted by Khanal et al. (2021), are driving risk-averse agriculturists around the world toward monoculture production systems in an attempt to lower the per-unit costs of commodity production.
Nonetheless, intercropping and the concept of sustainable agriculture are growing in interest among agriculturists around the world, and current research efforts indicate that intercropped systems will continue to be employed globally to serve the analogous purposes of improving on-farm biodiversity, soil health, resource use efficiency, weed suppression, and climate change resilience (Table 1). In a meta-analysis of global intercropping systems, Gu et al. (2021) indicates there are three main categories of commonly utilized crop species-cereals (seven species); legumes (12 species); and “other” crops (seven species). However, in this review, a search of relevant literature revealed that the most prevalent species used in intercropped systems can be grouped into the following categories: cereals, fiber crops, forage grasses, legumes (including legumes for forage), oilseeds, and “other” species (Table 1). In Australia, Europe, and North America, intercropping systems with small-grain cereals, legumes, and small-grain oilseeds are typically grown (Khanal et al., 2021; Gu et al., 2021). Contrastingly, most intercropped systems that utilize cereal crops with C4 photosynthesis, such as maize (Zea mays L.) and sorghum (Sorghum bicolor L.) are grown predominantly in Asia (Gu et al., 2021; Khanal et al., 2021; Stomph et al., 2020). Intercrop species selection is contingent on many factors, including grower preference and desired outcome, production cost, and market viability. Therefore, the most promising intercrop combinations for the world’s drylands consist of crop species that are already being produced in those respective regions. For the iPNW specifically, examples of intercrops that could be produced (due to their preexisting prevalence as monocultures) include a winter cereal-winter legume combination, a winter oilseed-winter legume combination, or a spring oilseed-spring legume combination.
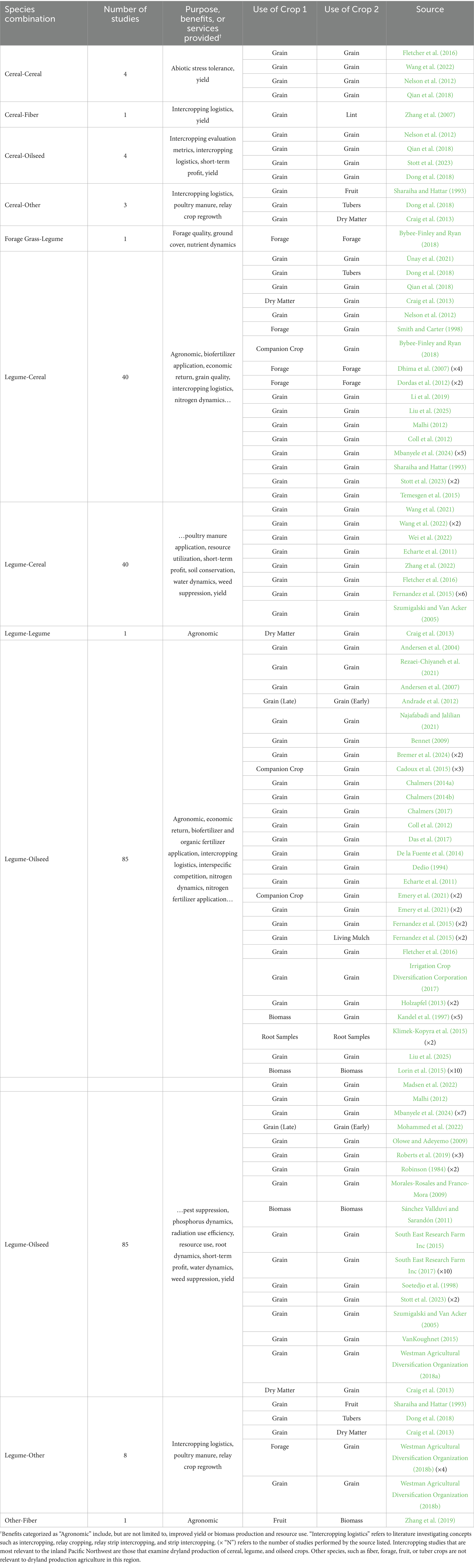
Table 1. A review of relevant intercropped species combinations (Modified from Dowling et al., 2021; Mirdoraghi et al., 2024).
In the western United States, and more specifically the iPNW, intercropping is not yet a common practice. While there have been several intercropping studies performed at local land-grant research institutions, the practice has been limited to plot-scale trials and intermittent, small-scale stands on producers’ farms (Oregon State University Columbia Basin Agricultural Research Center, 2020; Washington State University Wheat and Small Grains, 2024; Clark and Madsen, 2021; Madsen and Ford, 2021). That is not to say that intercropping in the iPNW is an improbable feat. In a region entrenched in cereal crop production, but also known for its oilseed and pulse crop production (Kaur et al., 2022), an intercropped rotation of peas (Pisum sativum L.) and canola (Brassica napus L.) (hereafter referred as “peaola”) would utilize two additional major crop functional groups (legumes and Brassica species), and complement cereal crop production systems. A spring peaola rotation would likely be the best fit for the majority of the iPNW’s dryland cropping systems (Karimi et al., 2017), as spring pea production already takes place on approximately 40,000 ha each year, and the majority of the 2019 canola crop in Washington state (29,000 ha) was spring-sown (Maaz et al., 2018; Schillinger, 2020).
Establishing peaola in lieu of monoculture spring pea or spring canola has the potential to increase land productivity and resource use efficiency (RUE), or the amount of biomass produced per unit of available resource (Hodapp et al., 2019). Peaola can also promote yield stability and WUE—the ratio of primary production to transpiration (Hodapp et al., 2019) and improve crop N use efficiency (NUE)—the ratio of biomass produced per unit of N consumed (Congreves et al., 2021). As an intercrop, peaola may serve as a cultural weed control method, reduce pest damage, and improve soil health, too (Duchene et al., 2017; Temesgen et al., 2015). However, intercropping is associated with a suite of logistical and agronomic challenges, and there are still numerous research questions that need to be answered regarding the implementation of peaola in the iPNW (Duchene et al., 2017; Madsen et al., 2022). Nonetheless, the agroecosystem services provided as a result of peaola intercropping not only have the potential to enhance cereal crop production in the iPNW, but also promote the sustainable intensification of production agriculture across the region.
The objective of this literature review is to synthesize current information about dryland pea-canola intercropped systems and determine if these systems are applicable to the iPNW. The review addresses challenges currently faced by producers in the iPNW, such as declining soil health and droughty conditions, and suggests that a peaola intercrop may be a more sustainable and viable option than current crop rotations. To support this optimistic approach, the review discusses both generalized and region-specific agronomic topics, including a brief overview on the ecological principles that drive successful intercropped systems, a comprehensive discussion of the feasibility of a pea-canola intercrop in the iPNW, the potential impacts of peaola on crop WUE and soil health, and the role of peaola in reducing pest pressure. More importantly, the review identifies several ways in which peaola can promote drought resilience and sustainability in the dryland iPNW. The penultimate section in this review discusses several barriers to peaola adoption as a crop rotation in the iPNW. To conclude, the review makes recommendations for specific research objectives and producer considerations.
2 Ecological principles of intercropping
The goals of all intercropped systems are as follows: to mitigate risk by growing two or more crop species, which increases the possibility of at least one species surviving to maturity (Madsen et al., 2022), to increase crop production by improving within-season production (Fletcher et al., 2016), and to improve land efficiency by enhancing crop RUE, which is quantified by a system’s land equivalency ratio (LER) value (Bybee-Finley and Ryan, 2018; Temesgen et al., 2015). The most common forms of intercropping used in production agriculture include relay-cropping, or planting a second crop into a prior established crop with the intent of harvesting at two different intervals, and interseeding, or planting one species directly into another, either at the same time or directly after the first species is established (Mohler and Stoner, 2009). In the iPNW, interseeding (intercropping) is likely to be the most feasible option for growers, due to the timing constraints in seeding and harvest operations in dryland production regions (Bybee-Finley and Ryan, 2018; Fletcher et al., 2016; Reddy et al., 2023).
While iPNW monoculture systems embody the principle of spatial diversification, or growing different crop species in separate fields, intercropping systems are an example of temporal diversification, which is the principle of growing two or more different crop species at the same time and in the same space (Bybee-Finley and Ryan, 2018; Fletcher et al., 2016). Temporal diversification, and by association, increased species richness and crop diversity (Maitra, 2020), leads to increased agroecosystem productivity, short-term increases in crop yield, and promotes long-term system stability (Reddy et al., 2023; Dowling et al., 2021). However, managing the many benign or detrimental interactions in crop stands with mixed species is challenging and requires the manipulation or avoidance of certain factors (Bybee-Finley and Ryan, 2018; Fletcher et al., 2016; Duchene et al., 2017), such as complementarity, competition, and facilitation between crop species (Naudin et al., 2010) (Figure 1).
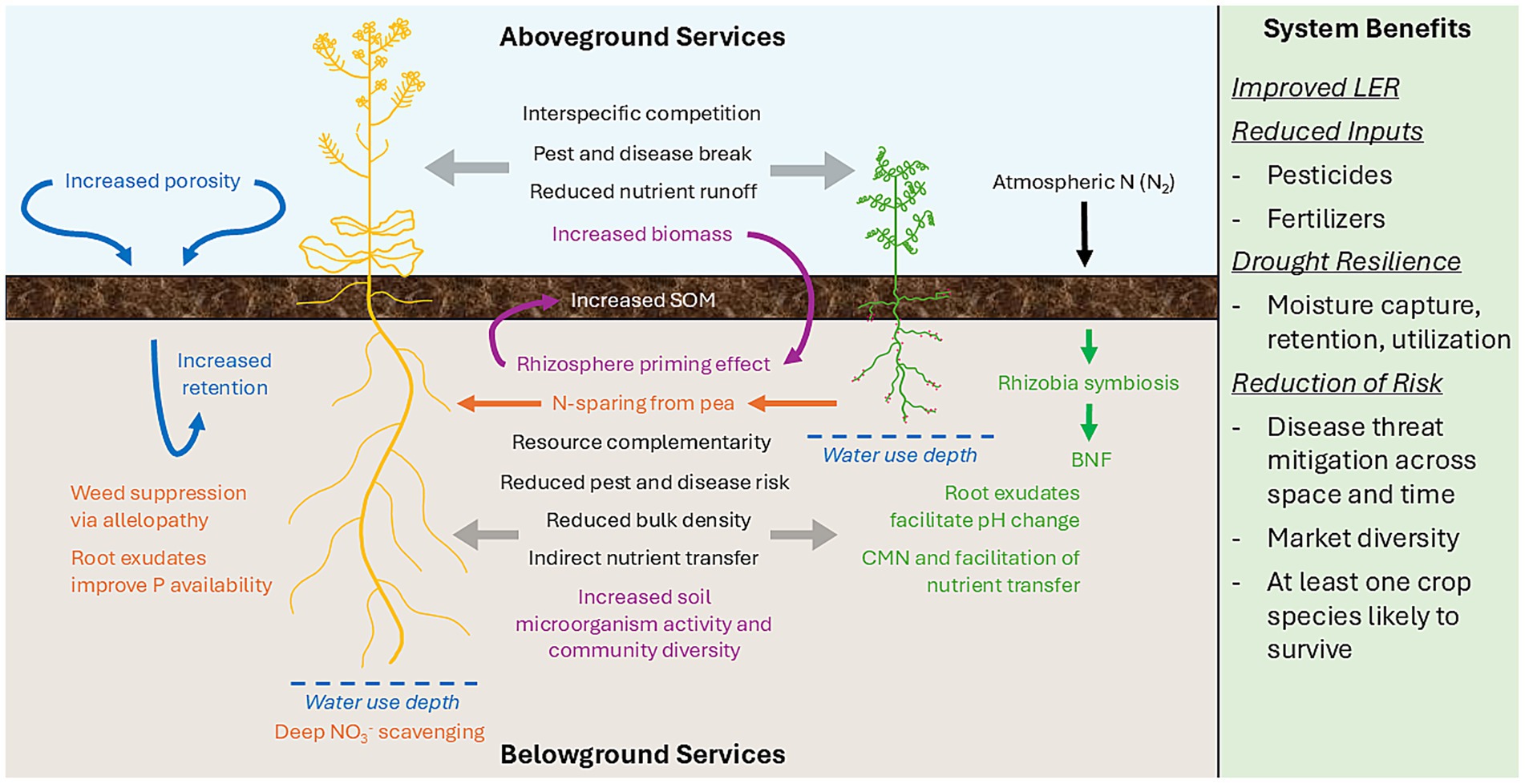
Figure 1. A diagram detailing the aboveground services of an intercropped peaola system in the upper blue tercile; the belowground services in the lower brown tercile; and the benefits to a cropping system in the green tercile on the right-hand side. SOM (soil organic matter); BNF (biological nitrogen fixation); CMN (common mycorrhizal network); LER (land equivalency ratio).
Interspecific complementarity, or niche partitioning, refers to the different partitioning and acquisition of resources based on a species’ unique needs (Duchene et al., 2017; Fridley, 2001). This concept outlines the importance of choosing the correct species for an intercropping rotation to reduce competition, especially if the producer plans to sow both species at the same time. In the iPNW, a pea-canola intercrop is likely to demonstrate some level of complementarity, as the rapid emergence rate of canola (Begna et al., 2021) allows for adequate establishment and resource acquisition before its pea counterpart. Ideally, at the time of pea emergence and development, the pea and canola would be accessing resources from differing niches in their environment (Figure 1). This is especially important for the iPNW, as growthier canola plants can serve as trellises for their pea counterparts—a fact that cannot be understated in this region, as ease of harvesting is crucial for growers that manage fields with steep, hilly terrain. Interspecific competition, while usually negatively associated with crop-weed interactions, can be manipulated in intercropping systems between complimentary crop species, especially if one is a legume, and the other species has a high N requirement (e.g., canola); competition for N between both crops can stimulate N fixation by the former, to the benefit of the latter (Maleziéux et al., 2009; Dowling et al., 2021). Facilitation in intercropping systems refers to the ability of one crop species to provide or make a limiting resource more accessible to its companion (Bybee-Finley and Ryan, 2018; Fletcher et al., 2016; Duchene et al., 2017; Andersen et al., 2004). Examples of interspecific facilitation that could occur in iPNW peaola systems include peas providing N to their canola counterparts (Bybee-Finley and Ryan, 2018; Louarn et al., 2020), hydraulic redistribution of soil moisture from a deep-rooted species (canola) to a shallow-rooted species (pea) (Caldwell and Richards, 1989; Burgess, 2011; Sekiya et al., 2011), and the acidification of the rhizosphere by one species’ root exudates (pea) to improve nutrient availability for its counterpart (canola) (Duchene et al., 2017; Hinsinger et al., 2003) (Figure 1).
3 The feasibility of peaola in the inland Pacific Northwest
In the iPNW, peas and canola are both grown as either fall- or spring-seeded crops in rotation with winter or spring cereals (Madsen et al., 2022; Maaz et al., 2018). Both pulse and oilseed crops are increasing in the number of acres grown per year (Kaur et al., 2022), and there are established markets, public and private breeding programs (Maaz et al., 2018; Schillinger, 2020), and processing facilities in the region for both crops (Maaz et al., 2018). Additionally, both pulse crops and oilseeds have existing state or regional commodity commissions that work to fund research, expand domestic and international markets, and engage in policy making on behalf of producers (Maaz et al., 2018).
Spring canola is one candidate for an intercropped rotation in the iPNW due to its favorable economic standing in the region as a biofuel feedstock (Schillinger, 2020), deep-rooting habits (Madsen et al., 2022), and ability to scavenge for nutrients such as N and P (Madsen et al., 2022; Andersen et al., 2004; Koenig et al., 2011) (Figure 1). Complementary to spring canola, spring peas are shallow-rooted plants and have moderate water requirements, making them ideal for the low, intermediate, and high rainfall zones (Schillinger, 2020; Kaur et al., 2022) (Figure 1). Spring canola cannot form symbiotic relationships with certain soil microbe functional groups, but spring peas can form symbiotic relationships with arbuscular mycorrhizal fungi (AMF) and rhizobia bacteria, which helps to facilitate system-wide nutrient transport, uptake, and biological nitrogen fixation (BNF) (Madsen et al., 2022; Hauggaard-Nielsen and Jensen, 2005) (Figure 1). The large-scale adoption of spring peaola is agronomically feasible for all three rainfall zones of the iPNW (Madsen et al., 2022), as Kaur et al. (2022) rated spring pea and spring canola as crops that are “intermediately stable” and “extremely stable” in their respective abilities to adapt to climate change. Additionally, there are various metrics associated with intercropped systems that can help producers and researchers quantify the agronomic and economic feasibility of peaola intercropping in the iPNW (Table 2).
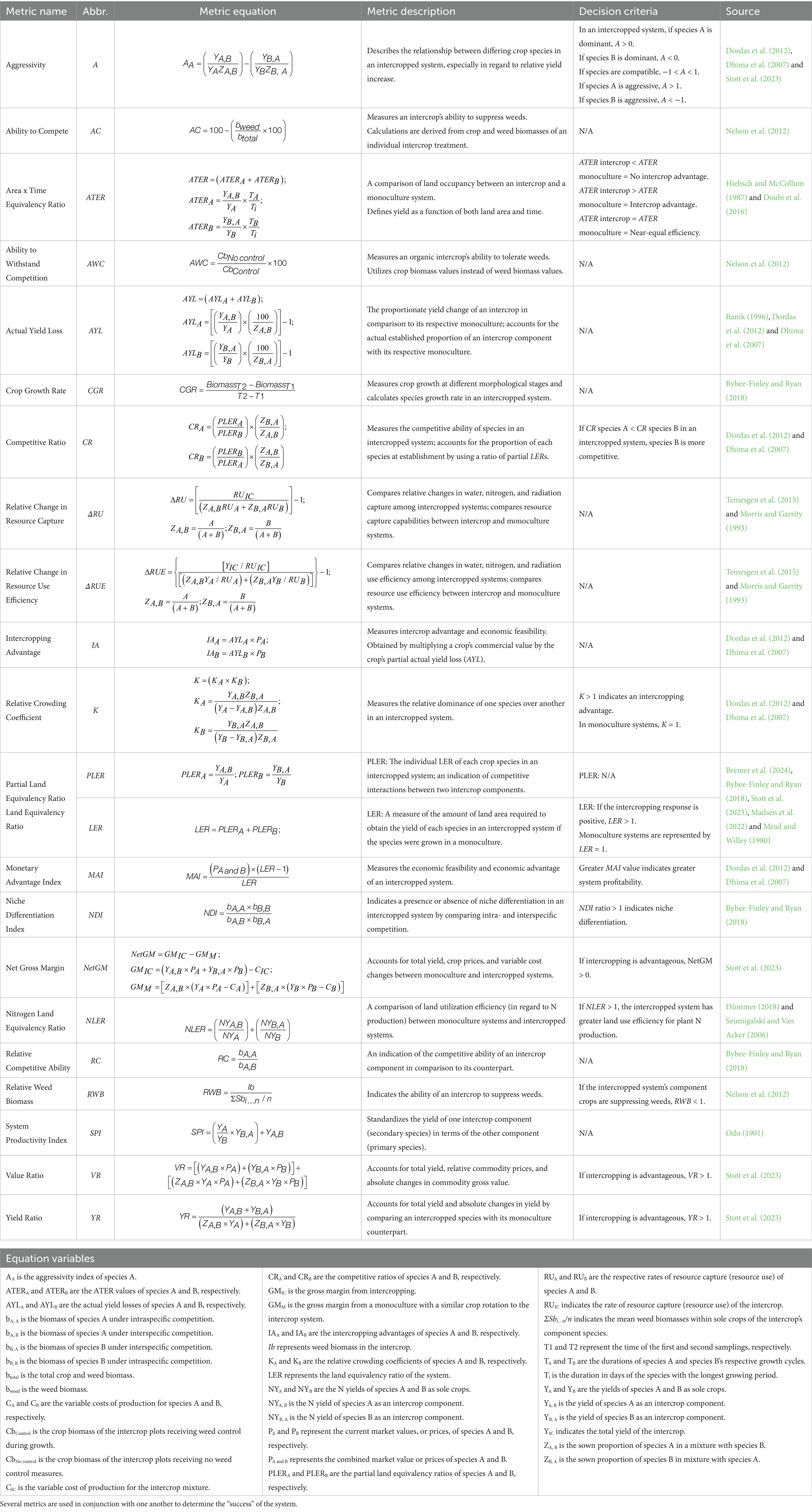
Table 2. Metrics used to evaluate the profitability, resource use efficiency, yield, and competitiveness of intercropped systems, and to assess a variety of agronomic, ecological, and economic factors
4 Peaola and crop water use efficiency
Intercropped systems are expected to succeed in challenging environments, such as those with limited water availability, poor soil fertility, or significant biotic stressors (Stomph et al., 2020). Soil water availability is the most limiting factor for crop growth and yield in the iPNW, and water deficits in dryland cropping systems negatively impact crop WUE (Kaur et al., 2022; Karimi et al., 2017). Therefore, risk-averse producers are more likely to include summer fallow periods and wait to plant a cool-season cereal instead of establishing a spring broadleaf crop, even though studies from the iPNW’s low-rainfall zone have determined the precipitation use efficiency of a winter wheat-summer fallow rotation averaged less than 30% (Kaur et al., 2022; Karimi et al., 2017; Williams et al., 2020a). Altered weather patterns also affect WUE and play a major role in cropping system decisions (Karimi et al., 2017). However, the WUE of peaola has the potential to be greater than the WUE of a pea, canola, or wheat monoculture in water-limited environments, as complementary root structure enables both species to exploit different volumes of soil (Duchene et al., 2017; Madsen et al., 2022). For example, Gan et al. (2009) reported that the roots of oilseed crops can reach depths of 80–100 cm, whereas that of pulse crops may only grow to 40–60 cm. Additionally, Cutforth et al. (2013) demonstrated that pea and canola crops withdraw their water requirements at different points in the growing season. Reportedly, oilseeds meet their water requirements earlier in the growing season, withdrawing only 42% of their moisture requirement post-anthesis at depths of 90 cm or greater, while pulses meet their moisture requirements later, withdrawing 52% of their moisture requirement post-anthesis from the upper 60 cm of the soil profile (Cutforth et al., 2013). Furthermore, the same study reported that wheat withdrew more water than either the oilseed or pulse crops, meeting nearly half its water requirement post-anthesis (Cutforth et al., 2013). However, the claim about intercropping enhancing WUE is based more in ecological theory and less in agronomic practice, as there are no preexisting iPNW-specific investigations that address this topic.
Due to its deep taproot and shallow lateral roots, spring canola may have the ability to redistribute moisture within the soil profile via hydraulic lift (Figure 1) (Neumann and Cardon, 2012; Koenig et al., 2011). Canola plants can have a hydraulic conductivity rate of approximately 0.11 m min−1 plant−1 at eight bars pressure, or more than double the hydraulic conductivity rate of pea plants (0.05 m min−1 plant−1at eight bars pressure) (Cutforth et al., 2013). Reportedly, the extraction of water from deep in the soil, and the subsequent release in upper profiles as a result of changes in matric pressure can redistribute small volumes (less than 1 mm) of water per day (Neumann and Cardon, 2012; Fletcher et al., 2016). While some studies indicate that this redistribution of water may be insignificant (e.g., Burgess, 2011), spring peas have a lower water requirement (approximately 350 mm) than spring canola (nearly 510 mm) (Stepanović et al., 2019; Bauder, n.d.; Schillinger, 2020); in instances of drought, where surface moisture reserves may be depleted, canola could potentially provide a small volume of water to its pea counterpart.
Differences in growth rate also help to stagger periods of high water demand throughout the growing season (Ehrmann and Ritz, 2014). Spring peas have aggressive periods of development early in the growing season, which can be beneficial for preliminary weed control via canopy closure, but may be detrimental to spring canola growth and development later in the season (Table 2) (Madsen et al., 2022). Concerns from producers regarding spring pea growth and interference with spring canola establishment are valid, and this issue can be alleviated by spring-sowing a winter pea cultivar, as demonstrated by Madsen et al. (2022).
It is thought that peaola may have the ability to improve the WUE of future crops by enhancing water infiltration and retention capabilities of the soil profile (Figure 1). The enhancement in soil water infiltration is due to the taproots of canola plants, which can grow to depths of nearly 1.2 m or greater (Williams et al., 2020b; Koenig et al., 2011), and create vertical channels for improved infiltration and percolation of precipitation (Madsen et al., 2022). Soil organic matter also plays a key role in soil water retention. In intercropped systems, SOM turnover rates are increased by the addition of labile organic matter from crop biomass and residues, as this initiates the rhizosphere priming effect (PE) among local microbial groups (Blagodatskaya and Kuzyakov, 2008; Fuhrmann and Zuberer, 2021) (Figure 1). Recent reports approximate this increased rate of SOM turnover resulting from PE to be nearly 37% on a generalized, global scale (Xu et al., 2024). Altogether, the presence of a living, diversified crop stand (and by extension, a diverse rhizosphere), promotes enhanced soil water retention and WUE through the maintenance of soil structure (Duchene et al., 2017).
5 Impacts of peaola on soil health
Soil health, or the capacity of a soil to function and sustain life, results from the confluence of physical, chemical, and biological factors (Pepper and Brooks, 2021). Increased biodiversity, including the addition of multiple plant species, is one way for agricultural producers to positively influence soil health and functioning. While managing interspecific competition between crop species can be challenging, there are many benefits to improving on-farm plant biodiversity (Maaz et al., 2018). For example, by growing both peas and canola (two crops from differing agronomic functional groups), producers can sustainably take advantage of a suite of agroecosystem services in a single growing season (Bybee-Finley and Ryan, 2018; Duchene et al., 2017; Madsen et al., 2022). To contrast, monoculture crop rotations require strategic considerations of year-to-year nutrient requirements and herbicide applications, in addition to predictive considerations of future weather, pest, and market conditions (Reddy et al., 2023; Maitra, 2020). Monoculture crop rotations are also limited to a narrow scope of agroecosystem services provided in a single growing season (Reddy et al., 2023; Maitra, 2020).
The prevention and reduction of soil erosion is another avenue for the maintenance of soil structure and health. An effectively established intercropped system promotes the growth of a dense plant cover, which prevents precipitation from striking bare soil (Reddy et al., 2023). By reducing the physical impact of rainfall on the soil surface, the occurrence of clogged surface pores and surface crusting is minimized, thereby promoting permeability and reducing surface runoff (Reddy et al., 2023; Lithourgidis et al., 2011). To add, peas have a high concentration of roots in the upper 30 cm of soil, which is beneficial for erosion control and enhancing soil aeration, while deeper-rooted canola serves to create vertically connected pore networks (Reddy et al., 2023; Lithourgidis et al., 2011). An increase in root activity at varying depths in the soil profile has also been demonstrated by Duchene et al. (2017) to improve soil aggregation and decrease bulk density, which can be soil health challenges in monoculture systems (Figure 1).
In instance of local soil fertility, studies conducted in the intermediate rainfall zone of the iPNW by Madsen et al. (2022) determined that, when compared to pea and canola monoculture yields, peaola outyielded both without the addition of fertilizer N. By eliminating the need for an N application, peaola can improve the long-term sustainability of production systems via the reduction of fertilizer (Stott et al., 2023; Chai et al., 2014), as well as decrease nonpoint source pollution from nitrate (NO3−) leaching (Whitmore and Schröder, 2007; Bybee-Finley and Ryan, 2018). In a balanced system, legumes fix sufficient quantities of plant-available N from atmospheric nitrogen (N2) to satisfy their own growth needs. If soil N is a limiting growth factor due to low availability or interspecific competition, the legumes will contribute excess mineral N to the system via the sparing effect (Chalk et al., 2014; Duchene et al., 2017; Ehrmann and Ritz, 2014) (Figure 1). As a highly competitive crop with an N demand comparable to wheat (Koenig et al., 2011), canola may initiate the sparing effect by decreasing the soil inorganic N concentration, thereby stimulating BNF in its pea counterpart (Schmidtke et al., 2004; Naudin et al., 2010). The sparing effect can improve agroecosystem NUE (Temesgen et al., 2015; Dowling et al., 2021), but competition between the canola and peas must reach a state of equilibrium in which BNF can be sustained without dominating the peas’ ability to symbiotically fix N (Dowling et al., 2021).
In addition to soil N enrichment, root exudates from legumes modify the chemical composition of the rhizosphere and facilitate the availability and mobilization of nutrients, including P, K, and Mg (Hauggaard-Nielsen et al., 2009; Duchene et al., 2017; Dowling et al., 2021) (Figure 1). In the instance of P, phosphatase enzymes and carboxylates exuded by legume roots improve P bioavailability and cycling throughout the soil profile via hydrolysis of organic and inorganic forms of P, rendering P more available for plant uptake under less-than ideal environmental conditions (Hinsinger et al., 2003; Dowling et al., 2021; Duchene et al., 2017). For example, after inducing P-deficient conditions on soybean-wheat intercrops and their respective monocultures, Bargaz et al. (2017) reported that soybean nodule acid phosphatase activity at the zero to 15 cm layer of soil was significantly stimulated under both the monoculture and intercropped system by 37 and 33%, respectively.
The ability of peas to improve P cycling is especially important in peaola systems, as canola does not benefit from P transport via the common mycorrhizal network (CMN). However, peas maintain the ability to form relationships with AMF and become symbionts in the CMN, shifting nutrients within the rhizosphere along a source-sink gradient from nutrient-rich to nutrient-poor plants (Duchene et al., 2017; Drijber and McPherson, 2021). Cropping systems with diverse plants result in AMF networks that are also diverse and abundant (Bybee-Finley and Ryan, 2018; Duchene et al., 2017); this not only benefits the plants, but also soil aggregate stability, as the binding nature of the CMN, paired with mycelial glomalin secretions, has been shown to improve soil stability (Duchene et al., 2017; Morton, 2021).
Peaola has also demonstrated the ability to alter the compositions of core microbiomes associated with pea and canola monocultures. According to Madsen et al. (2022), in dryland peaola intercropped systems, canola plants may associate with microorganisms that are not typically found in the core microbiomes of canola monoculture systems. Furthermore, Madsen et al. (2022) states that the microbiomes of peaola intercrops contain at least 10 bacterial core members that are not typically found in either canola or pea monoculture core microbiomes, suggesting that peaola has the ability to “recruit” microorganisms, and curate both diverse and niche microbiomes. Similarly, Bargaz et al. (2017) determined that members of bacterial families known to contribute to soil N cycling or serve as plant growth-promoting rhizobacteria, including Bradyrhizobiacaea and Rhodospirillaceae, were present in greater amounts under intercropped treatments than monoculture treatments, regardless of soil P availability.
6 Reducing pest pressure with peaola
Increasing plant biodiversity in a cropping system minimizes disease incidence, insect pressure, and weed pressure, which reduces input costs and boosts crop yield over the short- and long-term (Bybee-Finley and Ryan, 2018). In the iPNW, establishing peaola rotations would help break the insect, pest, and disease cycles by employing the dilution effect, which refers to reducing the number of susceptible host plants (Schillinger, 2020; Bybee-Finley and Ryan, 2018) (Figure 1). As a member of the Brassica family, canola produces glucosinolates (GSLs), which act as defense mechanisms against soil-borne pathogens when undergoing hydrolysis in the soil (Schillinger, 2020; Ehrmann and Ritz, 2014) (Figure 1). It has been determined that, on a per-plant basis, canola is a more effective emitter of allelochemicals—producing more GSLs when grown in tandem with a legume species than when grown in a monoculture (Couëdel et al., 2018). In their 2018 study, Couëdel et al. calculated GSLMIX:GSLSC, or the ratio of GSLs produced by a crucifer-legume mixture to the GSLs produced by a sole crop (μmol gDM−1). Reportedly, the canola-legume mixtures had a total root GSLMIX:GSLSC value of 1.22, and a total shoot GSLMIX:GSLSC value of 2.47, both of which were significantly greater than the normalized sole crop GSL value of 1.0 (Couëdel et al., 2018). Additionally, when compared to monoculture systems, peaola has been shown to contain greater populations of parasitoid insects (Clark and Madsen, 2021) and reduced populations of pest insects, such as pea aphid (Acyrthosiphon pisum) (Madsen et al., 2022; Dowling et al., 2021).
Peaola may also reduce weed pressure, as intercropping two plant species is a more effective way to ensure a crop stand competitively utilizes resources such as light, space, water, and nutrients (Duchene et al., 2017; Bybee-Finley and Ryan, 2018). By consuming these resources at an increased rate, which may stunt weed growth and vigor, peaola can serve as a form of cultural weed control. Subsequently, weed biomass, abundance, and reproductive vigor are also slowed by the lack of available resources, which, in the long-term, may help to deplete the soil weed seed bank (Poggio, 2005; Thorne et al., 2007). The variability of peaola growth and development ensures the crop receives adequate sunlight, while shading the soil surface and preventing weed germination (Reddy et al., 2023). Minimal or no applications of N, paired with the release of allelopathic compounds from the roots of canola plants, also help to suppress weed growth and biomass accumulation (Dowling et al., 2021; Maleziéux et al., 2009) (Figure 1). If needed, applications of herbicides selective to grass weed species can be used in peaola systems for an added control benefit (Madsen et al., 2022).
7 The ability of peaola to promote drought resilience and sustainability
Peaola rotations, and all intercropped rotations by extension, have the potential to boost productivity, promote on-farm drought resilience, and improve sustainability in the dryland areas of the iPNW by maximizing the use of resources on a per-acre basis (Reddy et al., 2023). In low productivity environments, such as those limited by soil moisture, LER values for intercropped rotations are higher than those of monoculture rotations due to the greater RUE of intercropped systems (Temesgen et al., 2015). In a multi-year winter and spring peaola study conducted at two locations in the iPNW’s intermediate rainfall zone, Madsen et al. (2022) calculated the overall LER of intercropped peaola to be 1.63, compared to the normalized 1.00 LER value for both pea and canola monocultures. In the same study, Madsen et al. (2022) also determined that LER values for peaola will remain relatively stable, even during the transition from a year that received average precipitation to a drought year. Peaola production is low-input and financially sustainable, as LER values have shown that peaola will outyield monoculture systems without the addition of synthetic N (Madsen et al., 2022). Little-to-no need for synthetic N applications, paired with a reduction or elimination in pesticide applications, reduces greenhouse gas emissions (Chai et al., 2014), promotes system-wide sustainability, and improves the system’s adaptive capacity to financial stressors (Dowling et al., 2021; Madsen et al., 2022) (Figure 1).
Ecological stability and drought resilience are achieved with peaola through the reduction of risk. Intercropped systems have been shown to improve crop yield stability and on-farm drought resilience over time and across locations, as demonstrated by the increase in peaola LER values from a normal year to a drought year (Bybee-Finley and Ryan, 2018; Madsen et al., 2022) (Figure 1). Peaola systems are considered less fragile than their monoculture counterparts under abiotic or biotic stress, as the ability of at least one species to survive harsh conditions and reach reproductive maturity is likely (Madsen et al., 2022; Whitmore and Schröder, 2007). In addition to improving sustainability and on-farm resilience, intercropped systems have the potential to improve cereal production in the iPNW, as wheat-following-canola or wheat-following-legume crops have been reported to exceed yields of wheat-following-wheat by 0.8 and 0.7 t ha−1, respectively (Angus et al., 2015). However, it is critical to note that (spring) wheat following (winter) canola has been investigated previously in the iPNW. Results from this multiyear study indicated a slight spring wheat yield hit, which was later linked to changes in microbial community composition and a decrease in microbial biomass under a winter canola crop (Schillinger and Paulitz, 2018; Hansen et al., 2019). The investigators concluded that the decrease in microbial biomass after winter canola was most likely due to the release of allelopathic compounds, such as GSLs and isothiocyanates (ITCs) from canola root residue (Hansen et al., 2019). Ultimately, the authors believe the yield-reduction scenario is not a long-term issue, especially if producers account for these findings when planning crop rotations by including a 13-month fallow period after a canola crop (Schillinger and Paulitz, 2018; Hansen et al., 2019).
8 Barriers to the adoption of peaola in the inland Pacific Northwest
There are several agronomic, economic, and logistical challenges associated with the adoption of intercropping in the iPNW. One reason as to why peaola and similar systems are yet to be attempted on a large scale in the region is due to the complexity of management required for successful intercropping (Stott et al., 2023; Fletcher et al., 2016; Madsen et al., 2022). Lack of available farmworkers (Fletcher et al., 2016), differences in nutrient requirements or applications between crop species (Dowling et al., 2021), and compatibility issues with chemical pest control are all considered barriers to adoption (Mamine and Farès, 2020; Madsen et al., 2022). Herbicide carryover is a concern in the iPNW, as soil-residual herbicides used to control weeds in cereal crop systems may persist in the environment for several years and negatively impact broadleaf crop production (Schillinger, 2020). Intercropped rotations also rely on interspecies competition to reduce weed pressure and promote crop yield. Successful manipulation of these innate plant characteristics requires understanding of plant–plant and plant–soil interactions (Bybee-Finley and Ryan, 2018), favorable climatic conditions (Duchene et al., 2017), and the careful selection of crop species and seeding rate (Duchene et al., 2017; Reddy et al., 2023).
While growing broadleaf crops in the iPNW has been historically viewed as a favorable disease mitigation practice, the establishment of a pea-canola intercrop may enable the development of pest and disease cycles that could affect subsequent wheat crops. Wheat grown in the iPNW is susceptible to root-lesion nematodes, with Pratylenchus neglectus and Pratylenchus thornei as the two most common species (Smiley et al., 2014). In a greenhouse assessment of 30 monocotyledonous and dicotyledonous plant species, Smiley et al. (2014) determined that canola was a “good” to “very good” host of P. neglectus, and that peas were generally “very good” hosts of P. thornei. Consequently, it can be inferred that a stand of peaola would be an effective host of both nematode species. To add, Paulitz (2006) states that diseases of wheat, such as Rhizoctonia bare patch and root rot, and Pythium root rot, have wide host ranges, and cannot be controlled with crop rotation alone.
Previously in the iPNW, economic and marketing constraints have limited the production of alternative crops, such as canola (Maaz et al., 2018). Although canola is considered a major crop in the iPNW now, any alternative crop that is to be produced on a commercial scale in this region must be agronomically feasible and economically competitive with wheat (Maaz et al., 2018). In the iPNW, producers may choose to grow a “less risky” crop to maintain relationships with absentee landlords and ensure access to federal financial assistance and crop insurance (Maaz et al., 2018). Additionally, growing a “risky” alternative crop is an issue regarding production, and presents the possibility of a significant financial burden for producers, as the equipment currently owned by a producer may not have the capability to plant, harvest, or process two crop species simultaneously (Stott et al., 2023), and there is currently no crop insurance option for intercrops. While recent iPNW peaola research has demonstrated that peaola can gross $143.53 ha−1, topping monoculture pea and canola crops at $137.58 ha−1 and $114.62 ha−1, respectively (Wysocki, 2021), these figures do not reflect grain sorting costs, which Stott et al. (2023) estimates to be at least $55 USD ($85 AUD) ton−1. According to Wysocki (2021) and Stott et al. (2023), a peaola crop yielding 1,477 kg ha−1 would be assessed with a total commodity separation fee of $88.62 ha−1, or 62% of the peaola crop’s gross revenue ha−1. Furthermore, iPNW warehouses or grain buyers may not have the facilities to dry, separate, and process two intermixed commodities (Mamine and Farès, 2020).
9 The future of peaola in the inland Pacific Northwest: conclusions and recommendations
The successful implementation of sustainable intercropped systems in the iPNW will require collaborative participation from researchers, producers, and government entities. Producers should consider their current management goals and determine which intercropping practices will best suit their farm (Bybee-Finley and Ryan, 2018). Additionally, producers who want to establish farm-scale experiments should consider taking advantage of either state- or federal-funded cost-share programs. Private and public research institutions will also play an important role in the future of intercropping. More region-specific research is needed to evaluate the resource-use dimensions of intercropping, in addition to short- and long-term benefits to the agroecosystem (Fletcher et al., 2016).
Research should also be conducted to identify genotypic adaptation within species for intercropping systems, similar to initiatives taken in previous decades to further the production of alternative crops, such as canola, in the iPNW (Brooker et al., 2015; Maaz et al., 2018). Currently, there is interest among producers and stakeholders in the iPNW to continue and ramp up peaola research efforts. For context, the investigation performed by Madsen et al. (2022) is considered to be a keystone dryland peaola study among the iPNW research community and has been the inspiration for novel research projects. To add, iPNW producers that want to learn more about intercropping have been engaging in facilitated meetings regarding the use of peaola in iPNW cropping systems. These meetings have discussed topics such as potential barriers to adoption, agronomic logistics, and marketing and processing peaola grain.
Altogether, the ability of producers to successfully integrate intercropped rotations on their farms, and the ability of researchers to conduct intercropping experiments will rely on the actions of policymakers. Historically, in the iPNW, the success of alternative crops has been dependent on factors outside the control of producers and researchers, such as favorable trade policies, access to markets, the establishment of grading standards, levy systems, and the expansion of handling and processing facilities (Maaz et al., 2018). Ultimately, policy support for sustainable agriculture practices will promote the adoption of intercropping in the iPNW and provide sufficient funding for research programs and producer resources (Bybee-Finley and Ryan, 2018).
The many challenges faced by the iPNW’s agricultural community will continue to negatively impact the region’s ability to produce high-yielding cereal crops. Peaola, with its potential to promote plant biodiversity, improve soil health, and increase land productivity, may be the solution. However, the implementation of this alternative crop in the iPNW may not look like other peaola systems around the world. Peaola utilization as either a winter or spring crop in the iPNW will likely vary due to the region’s three distinct rainfall zones and dynamic microclimates. Furthermore, peaola may not be an ideal grain crop for the region. If this alternative commodity proves to be too difficult to separate and market, peaola grazing or peaola silage production could become attractive, economically favorable contingencies for growers.
Author contributions
LS: Conceptualization, Data curation, Methodology, Writing – original draft. SuS: Conceptualization, Supervision, Writing – review & editing. DW: Writing – review & editing. GH: Writing – review & editing. HN: Writing – review & editing. ShS: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Washington Oilseed Cropping Systems funded the graduate student assistantship.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adil, M., Lv, F., Li, T., Chen, Y., Gul, I., Lu, H., et al. (2024). Long-term effects of management practices on soil water, yield and water use of dryland wheat: a global meta-analysis. Eur. J. Soil Sci. 75:e13541. doi: 10.1111/ejss.13541
Andersen, M. K., Hauggaard-Nielsen, H., Ambus, P., and Jensen, E. S. (2004). Biomass production, symbiotic nitrogen fixation and inorganic N use in dual and tri-component annual intercrops. Plant Soil 266, 273–287. doi: 10.1007/s11104-005-0997-1
Andersen, M. K., Hauggaard-Nielsen, H., Weiner, J., and Jensen, E. S. (2007). Competitive dynamics in two- and three-component intercrops. J. Appl. Ecol. 44, 545–551. doi: 10.1111/j.1365-2664.2007.01289.x
Andrade, J. F., Cerrudo, A., Rizzalli, R. H., and Monzon, J. P. (2012). Sunflower–soybean intercrop productivity under different water conditions and sowing managements. Agron. J. 104, 1049–1055. doi: 10.2134/agronj2012.0051
Angus, J. F., Kirkegaard, J. A., Hunt, J. R., Ryan, M. H., Ohlander, L., and Peoples, M. B. (2015). Break crops and rotations for wheat. Crop Pasture Sci. 66, 523–552. doi: 10.1071/CP14252
Awale, R., Singh, S., and Machado, S. (2022). Soil chemical properties and wheat yields under different tillage and nitrogen rates in eastern Oregon. Agrosyst. Geosci. Environ. 5:e20272. doi: 10.1002/agg2.20272
Banik, P. (1996). Evaluation of wheat (Triticum aestivum) and legume intercropping under 1:1 and 2:1 row-replacement series system. J. Agron. Crop Sci. 176, 289–294. doi: 10.1111/j.1439-037X.1996.tb00473.x
Bargaz, A., Noyce, G. L., Fulthorpe, R., Carlsson, G., Furze, J. R., Jensen, E. S., et al. (2017). Species interactions enhance root allocation, microbial diversity and P acquisition in intercropped wheat and soybean under P deficiency. Appl. Soil Ecol. 120, 179–188. doi: 10.1016/j.apsoil.2017.08.011
Bauder, J. W. (n.d.). The right strategy for irrigating your canola crop. Available online at: https://waterquality.montana.edu/farm-ranch/irrigation/other_crops/canola.html (Accessed May 12, 2025).
Begna, S., Angadi, S., Mesbah, A., Umesh, R. M., and Stamm, M. (2021). Forage yield and quality of winter canola–pea mixed cropping system. Sustainability. 13:2122. doi: 10.3390/su13042122
Bennet, M. (2009). Peola at Minnipa in 2009. Available online at: https://www.farmtrials.com.au/trial/20746 (Accessed February 20, 2025).
Blagodatskaya, Е., and Kuzyakov, Y. (2008). Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: critical review. Biol. Ferti. Soils 45, 115–131. doi: 10.1007/s00374-008-0334-y
Bremer, E., Ellert, B. H., Pauly, D., and Greer, K. J. (2024). Variation in over-yielding of pulse-oilseed intercrops. Field Crop Res. 305:109190. doi: 10.1016/j.fcr.2023.109190
Brooker, R. W., Bennett, A. E., Cong, W. F., Daniell, T. J., George, T. S., Hallett, P. D., et al. (2015). Improving intercropping: a synthesis of research in agronomy, plant physiology and ecology. New Phytol. 206, 107–117. doi: 10.1111/nph.13132
Burgess, S. S. O. (2011). Can hydraulic redistribution put bread on our table? Plant Soil 341, 25–29. doi: 10.1007/s11104-010-0638-1
Bybee-Finley, K. A., and Ryan, M. R. (2018). Advancing intercropping research and practices in industrialized agricultural landscapes. Agriculture 8:80. doi: 10.3390/agriculture8060080
Cadoux, S., Sauzet, G., Valantin-Morison, M., Pontet, C., Champolivier, L., Robert, C., et al. (2015). Intercropping frost-sensitive legume crops with winter oilseed rape reduces weed competition, insect damage, and improves nitrogen use efficiency. OCL 22:D302. doi: 10.1051/ocl/2015014
Caldwell, M. M., and Richards, J. H. (1989). Hydraulic lift: water efflux from upper roots improves effectiveness of water uptake by deep roots. Oecologia 79, 1–5. doi: 10.1007/BF00378231
Chai, Q., Qin, A., Gan, Y., and Yu, A. (2014). Higher yield and lower carbon emission by intercropping maize with rape, pea, and wheat in arid irrigation areas. Agron. Sustain. Dev. 34, 535–543. doi: 10.1007/s13593-013-0161-x
Chalk, P. M., Peoples, M. B., McNeill, A. M., Boddey, R. M., Unkovich, M. J., Gardener, M. J., et al. (2014). Methodologies for estimating nitrogen transfer between legumes and companion species in agro-ecosystems: a review of 15N-enriched techniques. Soil Biol. Biochem. 73, 10–21. doi: 10.1016/j.soilbio.2014.02.005
Chalmers, S. (2014a). Intercropping pea and canola based on row orientation and nitrogen rates final report 2011-2013. WADO 2014 Ann. Rep., 107–127.
Chalmers, S. (2017). Responses of pea and canola intercrops to nitrogen and phosphorus applications (year 2 of 2; final report). WADO 2017 Ann. Rep., 127–136.
Clark, R., and Madsen, I. (2021). “Peaola intercropping as a pest and beneficial insect management tactic” in 2021 dryland field day abstracts: highlights of research Progress (Pullman, WA, United States: Washington State University Department of Crop and Soil Sciences), 62–63.
Coll, L., Cerrudo, A., Rizzalli, R., Monzon, J. P., and Andrade, F. H. (2012). Capture and use of water and radiation in summer intercrops in the south-east pampas of Argentina. Field Crop Res. 134, 105–113. doi: 10.1016/j.fcr.2012.05.005
Congreves, K. A., Otchere, O., Ferland, D., Farzadfar, S., Williams, S., and Arcand, M. M. (2021). Nitrogen use efficiency definitions of today and tomorrow. Front. Plant Sci. 12:637108. doi: 10.3389/fpls.2021.637108
Couëdel, A., Alletto, L., Kirkegaard, J., and Justes, É. (2018). Crucifer glucosinolate production in legume-crucifer cover crop mixtures. Eur. J. Agron. 96, 22–33. doi: 10.1016/j.eja.2018.02.007
Craig, P. R., Coventry, D., and Edwards, J. H. (2013). Productivity advantage of crop–perennial pasture intercropping in southeastern Australia. Agron. J. 105, 1588–1596. doi: 10.2134/agronj2013.0196
Cutforth, H. W., Angadi, S. V., McConkey, B. G., Miller, P. R., Ulrich, D., Gulden, R., et al. (2013). Comparing rooting characteristics and soil water withdrawal patterns of wheat with alternative oilseed and pulse crops grown in the semiarid Canadian prairie. Can. J. Soil Sci. 93, 147–160. doi: 10.4141/cjss2012-081
Das, S., Nandini Devi, K., Singh Athokpam, H., Lhungdim, J., and Longjam, M. (2017). Chickpea (Cicer arietinum L.) based intercropping system with rapeseed (Brassica napus L.) on growth, yield and competition indices. Environ. Ecol. 35, 427–430. doi: 10.5555/20173083361
de la Fuente, E. B., Suárez, S. A., Lenardis, A. E., and Poggio, S. L. (2014). Intercropping sunflower and soybean in intensive farming systems: evaluating yield advantage and effect on weed and insect assemblages. NJAS Wageningen J. Life Sci. 70-71, 47–52. doi: 10.1016/j.njas.2014.05.002
Dhima, K. V., Lithourgidis, A. S., Vasilakoglou, I. B., and Dordas, C. A. (2007). Competition indices of common vetch and cereal intercrops in two seeding ratio. Field Crop Res. 100, 249–256. doi: 10.1016/j.fcr.2006.07.008
Dong, N., Tang, M. M., Zhang, W. P., Bao, X. G., Wang, Y., Christie, P., et al. (2018). Temporal differentiation of crop growth as one of the drivers of intercropping yield advantage. Sci. Rep. 8:3110. doi: 10.1038/s41598-018-21414-w
Dordas, C. A., Vlachostergios, D. N., and Lithourgidis, A. S. (2012). Growth dynamics and agronomic-economic benefits of pea–oat and pea–barley intercrops. Crop Pasture Sci. 63, 45–52. doi: 10.1071/CP11181
Doubi, B. T. S., Kouassi, K. I., Kouakou, K. L., Koffi, K. K., Baudoin, J. P., and Zoro, B. I. A. (2016). Existing competitive indices in the intercropping system of Manihot esculenta Crantz and Lagenaria siceraria (Molina) Standley. J. Plant Interact. 11, 178–185. doi: 10.1080/17429145.2016.1266042
Dowling, A., Sadras, V. O., Roberts, P., Doolette, A., Zhou, Y., and Denton, M. D. (2021). Legume-oilseed intercropping in mechanised broadacre agriculture – a review. Field Crop Res. 260:107980. doi: 10.1016/j.fcr.2020.107980
Drijber, R. A., and McPherson, M. R. (2021). “Mycorrhizal symbioses” in Applications of soil microbiology. eds. T. J. Gentry, J. J. Fuhrmann, and D. A. Zuberer (Cambridge, MA, United States: Elsevier Inc), 303–325.
Duchene, O., Vian, J. F., and Celette, F. (2017). Intercropping with legume for agroecological cropping systems: complementarity and facilitation processes and the importance of soil microorganisms. A review. Agric. Ecosyst. Environ. 240, 148–161. doi: 10.1016/j.agee.2017.02.019
Dümmer, R. (2018). Higher wheat yields with narrow strip pea-wheat intercropping [Master’s thesis]. Wageningen (NL): Wageningen University and Research.
Echarte, L., Maggiora, A. D., Cerrudo, D., Gonzalez, V. H., Abbate, P., Cerrudo, A., et al. (2011). Yield response to plant density of maize and sunflower intercropped with soybean. Field Crop Res. 121, 423–429. doi: 10.1016/j.fcr.2011.01.011
Ehrmann, J., and Ritz, K. (2014). Plant: soil interactions in temperate multi-cropping production systems. Plant Soil 376, 1–29. doi: 10.1007/s11104-013-1921-8
Emery, S. E., Anderson, P., Carlsson, G., Friberg, H., Larsson, M. C., Wallenhammar, A. C., et al. (2021). The potential of intercropping for multifunctional crop protection in oilseed rape (Brassica napus L.). Front. Agron. 3:782686. doi: 10.3389/fagro.2021.782686
Fernandez, A. L., Sheaffer, C. C., and Wyse, D. L. (2015). Productivity of field pea and lentil with cereal and brassica intercrops. Agron. J. 107, 249–256. doi: 10.2134/agronj14.0361
Fletcher, A. L., Kirkegaard, J. A., Peoples, M. B., Robertson, M. J., Whish, J., and Swan, A. D. (2016). Prospects to utilise intercrops and crop variety mixtures in mechanised, rain-fed, temperate cropping systems. Crop Pasture Sci. 67, 1252–1267. doi: 10.1071/CP16211
Fridley, J. D. (2001). The influence of species diversity on ecosystem productivity: how, where, and why? Oikos 93, 514–526. doi: 10.1034/j.1600-0706.2001.930318.x
Fuhrmann, J. J., and Zuberer, D. A. (2021). “Carbon transformations and soil organic matter formation” in Principles and applications of soil microbiology. eds. T. J. Gentry, J. J. Fuhrmann, and D. A. Zuberer (Cambridge, MA, United States: Elsevier Inc), 327–361.
Gan, Y. T., Campbell, C. A., Janzen, H. H., Lemke, R., Liu, L. P., Basnyat, P., et al. (2009). Root mass for oilseed and pulse crops: growth and distribution in the soil profile. Can. J. Plant Sci. 89, 883–893. doi: 10.4141/CJPS08154
Gelardi, D. L., Roy, M., and Hancock, J. (2023). Soil management in Washington’s dryland wheat: WSDA survey results. Olympia, WA, United States: WSDA Agriculture AGR Publication, 103–166.
Ghimire, R., Machado, S., and Rhinhart, K. (2015). Long-term crop residue and nitrogen management effects on soil profile carbon and nitrogen in wheat-fallow systems. Agron. J. 107, 2230–2240. doi: 10.2134/agronj14.0601
Gu, C., Bastiaans, L., Anten, N. P. R., Makowski, D., and van der Werf, W. (2021). Annual intercropping suppresses weeds: a meta-analysis. Agric. Ecosyst. Environ. 322:107658. doi: 10.1016/j.agee.2021.107658
Hansen, J. C., Schillinger, W. F., Sullivan, T. S., and Paulitz, T. C. (2019). Soil microbial biomass and fungi reduced with canola introduced into long-term monoculture wheat rotations. Front. Microbiol. 10:1488. doi: 10.3389/fmicb.2019.01488
Hauggaard-Nielsen, H., Gooding, M., Ambus, P., Corre-Hellou, G., Crozat, Y., Dahlmann, C., et al. (2009). Pea–barley intercropping for efficient symbiotic N2-fixation, soil N acquisition and use of other nutrients in European organic cropping systems. Field Crop Res. 113, 64–71. doi: 10.1016/j.fcr.2009.04.009
Hauggaard-Nielsen, H., and Jensen, E. S. (2005). Facilitative root interactions in intercrops. Plant Soil 274, 237–250. doi: 10.1007/s11104-004-1305-1
Hiebsch, C. K., and McCollum, R. E. (1987). Area-×−time equivalency ratio: a method for evaluating the productivity of intercrops. Agron. J. 79, 15–22. doi: 10.2134/agronj1987.00021962007900010004x
Hinsinger, P., Plassard, C., Tang, C., and Jaillard, B. (2003). Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: a review. Plant Soil 248, 43–59. doi: 10.1023/A:1022371130939
Hodapp, D., Hillebrand, H., and Striebel, M. (2019). “Unifying” the concept of resource use efficiency in ecology. Front. Ecol. Evol. 6:233. doi: 10.3389/fevo.2018.00233
Holzapfel, C. (2013). Exploring the merits of intercropping canola and field pea in south-east Saskatchewan and fine-tuning management practices for maximum yield and profit. Available online at: https://saskpulse.com/research/listing/exploring-the-merits-of-intercropping-canola-and-field-pea-in-south-east-saskatchewan-and-fine-tuning-management-practices-for-maximum-yield-and-profit/ (Accessed February 22, 2025).
Irrigation Crop Diversification Corporation (2017). Intercropping marrowfat pea and mustard. ICDC Research Demonstr. Rep. 2017, 154–155.
Kandel, H. J., Schneiter, A. A., and Johnson, B. L. (1997). Intercropping legumes into sunflower at different growth stages. Crop Sci. 37, 1532–1537. doi: 10.2135/cropsci1997.0011183X003700050020x
Karimi, T., Stöckle, C. O., Higgins, S. S., Nelson, R. L., and Huggins, D. (2017). Projected dryland cropping system shifts in the Pacific northwest in response to climate change. Front. Ecol. Evol. 5:20. doi: 10.3389/fevo.2017.00020
Kaur, H., Huggins, D. R., Carlson, B., Stockle, C., and Nelson, R. (2022). Dryland fallow vs flex-cropping decisions in inland Pacific Northwest of USA. Agric. Syst. 201:103432. doi: 10.1016/j.agsy.2022.103432
Khanal, U., Stott, K. J., Armstrong, R., Nuttall, J. G., Henry, F., Christy, B. P., et al. (2021). Intercropping—evaluating the advantages to broadacre systems. Agriculture 11:453. doi: 10.3390/agriculture11050453
Kirby, E., Pan, W., Huggins, D., Painter, K., and Bista, P. (2017). “Rotational diversification and intensification” in Advances in dryland farming in the inland Pacific Northwest. eds. G. Yorgey and C. Kruger (Pullman, WA, United States: Washington State University Extension), 163–236.
Klimek-Kopyra, A., Głąb, T., Zając, T., Stokłosa, A., and Kulig, B. (2015). Vertical distribution of the root system of linseed (Linum usitatissimum L.) and legumes in pure and mixed sowing. Acta Agrobot. 68, 43–52. doi: 10.5586/aa.2015.004
Koenig, R. T., Hammac, W. A., and Pan, W. L. (2011). Canola growth, development, and fertility. Pullman, WA, United States: WSU Extension FS045E.
Li, Y., Shi, D., Li, G., Zhao, B., Zhang, J., Liu, P., et al. (2019). Maize/peanut intercropping increases photosynthetic characteristics, 13C-photosynthate distribution, and grain yield of summer maize. J. Integr. Agric. 18, 2219–2229. doi: 10.1016/S2095-3119(19)62616-X
Li, C., Stomph, T. J., Makowski, D., Li, H., Zhang, C., Zhang, F., et al. (2023). The productive performance of intercropping. Proc. Natl. Acad. Sci. 120:e2201886120. doi: 10.1073/pnas.2201886120
Lithourgidis, A. S., Dordas, C. A., Damalas, C. A., and Vlachostergios, D. N. (2011). Annual intercrops: an alternative pathway for sustainable agriculture. Aust. J. Crop. Sci. 5, 396–410.
Liu, K., Choo-Foo, K., Wen, G., Schoenau, J., and Knight, J. D. (2025). Assessing crop productivity, grain quality, and soil labile carbon and nitrogen in pea-based intercrops under low nitrogen input. Can. J. Plant Sci. 105, 1–12. doi: 10.1139/cjps-2024-0136
Lorin, M., Jeuffroy, M. H., Butier, A., and Valantin-Morison, M. (2015). Undersowing winter oilseed rape with frost-sensitive legume living mulches to improve weed control. Eur. J. Agron. 71, 96–105. doi: 10.1016/j.eja.2015.09.001
Louarn, G., Barillot, R., Combes, D., and Escobar-Gutiérrez, A. (2020). Towards intercrop ideotypes: non-random trait assembly can promote overyielding and stability of species proportion in simulated legume-based mixtures. Ann. Bot. 126, 671–685. doi: 10.1093/aob/mcaa014
Maaz, T., Wulfhorst, J. D., McCracken, V., Kirkegaard, J., Huggins, D. R., Roth, I., et al. (2018). Economic, policy, and social trends and challenges of introducing oilseed and pulse crops into dryland wheat cropping systems. Agric. Ecosyst. Environ. 253, 177–194. doi: 10.1016/j.agee.2017.03.018
Madsen, I. J., and Ford, J. (2021). “Peaola yield and land equivalence ratio experiments” in 2021 Dryland field day abstracts: highlights of research progress (Pullman, WA, United States: Washington State University Department of Crop and Soil Sciences), 66–67.
Madsen, I. J., Parks, J. M., Friesen, M. L., and Clark, R. E. (2022). Increasing biodiversity and land-use efficiency through pea (Pisum aestivum)-canola (Brassica napus) intercropping (peaola). Front. Soil Sci. 2:818862. doi: 10.3389/fsoil.2022.818862
Maitra, S. (2020). Intercropping of small millets for agricultural sustainability in drylands: a review. CR 55, 162–171. doi: 10.31830/2454-1761.2020.025
Maleziéux, E., Crozat, Y., Dupraz, C., Laurans, M., Makowski, D., Ozier-Lafontaine, H., et al. (2009). Mixing plant species in cropping systems: concepts, tools and models. A review. Agron. Sustain. Dev. 29, 43–62. doi: 10.1051/agro:2007057
Malhi, S. S. (2012). Improving crop yield, N uptake and economic returns by intercropping barley or canola with pea. Agric. Sci. 3, 1023–1033. doi: 10.4236/as.2012.38124
Mamine, F., and Farès, M. (2020). Barriers and levers to developing wheat–pea intercropping in Europe: a review. Sustainability 12:6962. doi: 10.3390/su12176962
Mbanyele, V., Enesi, R. O., Shaw, L., and Gorim, L. Y. (2024). A review of intercropping systems in Western Canada. Agron. J. 116, 2089–2108. doi: 10.1002/agj2.21622
Mead, R., and Willey, R. W. (1980). The concept of a ‘land equivalent ratio’ and advantages in yields from intercropping. Exp. Agric. 16, 217–228. doi: 10.1017/S0014479700010978
Mirdoraghi, M., Maleki Farahani, S., and Rezazadeh, A. (2024). Oilseeds in intercropping systems: strategies to increase oil quality and fatty acid profile, a review. J. Agric. Food Res. 17:101229. doi: 10.1016/j.jafr.2024.101229
Mohammed, Y. A., Gesch, R. W., Matthees, H. L., and Wells, S. S. (2022). Maturity selection but not sowing date enhances soybean productivity and land use in a winter camelina–soybean relay system. Food Energy Secur. 11:e346. doi: 10.1002/fes3.346
Mohler, C. L., and Stoner, K. A. (2009). “Guidelines for intercropping” in Crop rotation on organic farms: a planning manual. eds. C. L. Mohler and S. E. Johnson (Brentwood, MD: NRAES Cooperative Extension), 95–100.
Morales-Rosales, E. J., and Franco-Mora, O. (2009). Biomass, yield, and land equivalent ratio of Helianthus annus L. in sole crop and intercropped with Phaseolus vulgaris L. in high valleys of Mexico. Trop. Subtrop. Agroecosystems 10, 431–439.
Morris, R. A., and Garrity, D. P. (1993). Resource capture and utilization in intercropping: water. Field Crop Res. 34, 303–317. doi: 10.1016/0378-4290(93)90119-8
Morton, J. B. (2021). “Fungi” in Principles and applications of soil microbiology. eds. T. J. Gentry, J. J. Fuhrmann, and D. A. Zuberer (Cambridge, MA, United States: Elsevier Inc), 149–170.
Najafabadi, A., and Jalilian, J. (2021). Impacts of high and low-input farming systems on the quality change of safflower oil while intercropped with bitter vetch. J. Agr. Sci. Tarim Bili. 28, 401–411. doi: 10.15832/ankutbd.857026
Naudin, C., Corre-Hellou, G., Pineau, S., Crozat, Y., and Jeuffroy, M. H. (2010). The effect of various dynamics of N availability on winter pea–wheat intercrops: crop growth, N partitioning and symbiotic N2 fixation. Field Crop Res. 119, 2–11. doi: 10.1016/j.fcr.2010.06.002
Nelson, A. G., Pswarayi, A., Quideau, S., Frick, B., and Spaner, D. (2012). Yield and weed suppression of crop mixtures in organic and conventional systems of the western Canadian prairie. Agron. J. 104, 756–762. doi: 10.2134/agronj2011.0374
Neumann, R. B., and Cardon, Z. G. (2012). The magnitude of hydraulic redistribution by plant roots: a review and synthesis of empirical and modeling studies. New Phytol. 194, 337–352. doi: 10.1111/j.1469-8137.2012.04088.x
Ngapo, T. M., Bilodeau, P., Arcand, Y., Charles, M. T., Diederichsen, A., Germain, I., et al. (2021). Historical indigenous food preparation using produce of the three sisters intercropping system. Foods 10:524. doi: 10.3390/foods10030524
Odo, P. E. (1991). Evaluation of short and tall sorghum varieties in mixtures with cowpea in the Sudan savanna of Nigeria: land equivalent ratio, grain yield and system productivity index. Exp. Agric. 27, 435–441. doi: 10.1017/S0014479700019426
Olowe, V. I. O., and Adeyemo, A. Y. (2009). Enhanced crop productivity and compatibility through intercropping of sesame and sunflower varieties. Ann. Appl. Biol. 155, 285–291. doi: 10.1111/j.1744-7348.2009.00340.x
Oregon State University Columbia Basin Agricultural Research Center (2020). 2020 field tour pea-Ola (pea/canola mixes) field tour. Available online at: https://agsci.oregonstate.edu/cbarc/outreach/2020-field-day (Accessed April 17, 2025).
Osborne, B. B., Bestelmeyer, B. T., Currier, C. M., Homyak, P. M., Throop, H. L., Young, K., et al. (2022). The consequences of climate change for dryland biogeochemistry. New Phytol. 236, 15–20. doi: 10.1111/nph.18312
Paulitz, T. C. (2006). Low input no-till cereal production in the Pacific northwest of the U.S.: the challenges of root diseases. Eur. J.Plant Pathol. 115, 271–281. doi: 10.1007/s10658-006-9023-6
Pepper, I. L., and Brooks, J. P. (2021). “Soil microbial influences on ‘one health’” in Principles and applications of soil microbiology. eds. T. J. Gentry, J. J. Fuhrmann, and D. A. Zuberer (Cambridge, MA, United States: Elsevier Inc), 681–700.
Poggio, S. L. (2005). Structure of weed communities occurring in monoculture and intercropping of field pea and barley. Agric. Ecosyst. Environ. 109, 48–58. doi: 10.1016/j.agee.2005.02.019
Prăvălie, R. (2016). Drylands extent and environmental issues. A global approach. Earth-Sci. Rev. 161, 259–278. doi: 10.1016/j.earscirev.2016.08.003
Qian, X., Zang, H., Xu, H., Hu, Y., Ren, C., Guo, L., et al. (2018). Relay strip intercropping of oat with maize, sunflower and mung bean in semi-arid regions of Northeast China: yield advantages and economic benefits. Field Crop Res. 223, 33–40. doi: 10.1016/j.fcr.2018.04.004
Reddy, M. G., Yernaidu, Y., Chittibomma, K., and Kumar, T. S. (2023). Intercropping – an approach towards sustainability in dry land agriculture. Int. J. Environ. Clim. Chang. 13, 182–190. doi: 10.9734/ijecc/2023/v13i92221
Renton, M., and Flower, K. C. (2015). Occasional mouldboard ploughing slows evolution of resistance and reduces long-term weed populations in no-till systems. Agric. Syst. 139, 66–75. doi: 10.1016/j.agsy.2015.06.005
Rezaei-Chiyaneh, E., Mahdavikia, H., Battaglia, M. L., Thomason, W. E., and Caruso, G. (2021). Intercropping and fertilizer type impact seed productivity and secondary metabolites of dragon’s head and fenugreek. Sci. Hortic. 287:110277. doi: 10.1016/j.scienta.2021.110277
Roberts, P., Moodie, M., and Wilhelm, N. (2019). Intercropping increases productivity in the south Australian Mallee. Proceedings of the 2019 Agronomy Australia Conference, Wagga Wagga, Australia, 1–4.
Robinson, R. G. (1984). Sunflower for strip, row, and relay intercropping. Agron. J. 76, 43–47. doi: 10.2134/agronj1984.00021962007600010012x
Sánchez Vallduví, G. E., and Sarandón, S. J. (2011). Effects of changes in flax (Linum usitatissimum L.) density and interseeding with red clover (Trifolium pratense L.) on the competitive ability of flax against brassica weeds. J. Sust. Agric. 35, 914–926. doi: 10.1080/10440046.2011.611745
Schillinger, W. F. (2020). New winter crops and rotations for the Pacific northwest low-precipitation drylands. Agron. J. 112, 3335–3349. doi: 10.1002/agj2.20354
Schillinger, W. F., Papendick, R. I., Guy, S. O., Rasmussen, P. E., and Van Kessel, C. (2006). “Dryland cropping in the western United States” in Agronomy monographs. eds. G. A. Peterson, P. W. Unger, and W. A. Payne (Madison, WI, United States: ASA, CSSA, SSSA), 365–393.
Schillinger, W. F., and Paulitz, T. C. (2018). Canola versus wheat rotation effects on subsequent wheat yield. Field Crop Res. 223, 26–32. doi: 10.1016/j.fcr.2018.04.002
Schmidtke, K., Neumann, A., Hof, C., and Rauber, R. (2004). Soil and atmospheric nitrogen uptake by lentil (Lens culinaris Medik.) and barley (Hordeum vulgare ssp. nudum L.) as monocrops and intercrops. Field Crop Res. 87, 245–256. doi: 10.1016/j.fcr.2003.11.006
Sekiya, N., Araki, H., and Yano, K. (2011). Applying hydraulic lift in an agroecosystem: forage plants with shoots removed supply water to neighboring vegetable crops. Plant Soil 341, 39–50. doi: 10.1007/s11104-010-0581-1
Sharaiha, R. K., and Hattar, B. (1993). Intercropping and poultry manure effects on yields of corn, watermelon and soybean grown in a calcareous soil in the Jordan Valley. J. Agron. Crop Sci. 171, 260–267. doi: 10.1111/j.1439-037X.1993.tb00138.x
Smiley, R. W., Yan, G. P., and Gourlie, J. A. (2014). Selected Pacific northwest crops as hosts of Pratylenchus neglectus and P. thornei. Plant Dis. 98, 1341–1348. doi: 10.1094/PDIS-12-13-1296-RE
Smith, M. A., and Carter, P. R. (1998). Strip intercropping corn and alfalfa. J. Prod. Agric. 11, 345–353. doi: 10.2134/jpa1998.0345
Soetedjo, P., Martin, L. D., and Tennant, D. (1998). Productivity and water use of intercrops of field pea and canola. Proceedings of the 1998 Agronomy Australia Conference.
South East Research Farm Inc (2017). Intercropping Brassica carinata with various pulse crops. Agricultural Demonstration of Practices and Technologies (ADOPT) Final Report 20150470.
Stepanović, S., Arsenijevic, N., and Ugljic, Z. (2019). Yield and water used of field peas and chickpeas under irrigation. Available online at: https://cropwatch.unl.edu/2019/yield-water-use-irrigated-field-peas-chickpeas/ (Accessed May 12, 2025).
Stomph, T., Dordas, C., Baranger, A., de Rijk, J., Dong, B., Evers, J., et al. (2020). “Designing intercrops for high yield, yield stability and efficient use of resources: are there principles?” in Advances in agronomy. ed. D. L. Sparks (Newark, DE, United States: Academic Press), 1–50.
Stott, K. J., Wallace, A. J., Khanal, U., Christy, B. P., Mitchell, M. L., Riffkin, P. A., et al. (2023). Intercropping—towards an understanding of the productivity and profitability of dryland crop mixtures in southern Australia. Agronomy 13:2510. doi: 10.3390/agronomy13102510
Strauss, J., Swanepoel, P., Laker, M., and Smith, H. (2021). Conservation agriculture in rainfed annual crop production in South Africa. S. Afr. J. Plant Soil 38, 217–230. doi: 10.1080/02571862.2021.1891472
Szumigalski, A., and Van Acker, R. (2005). Weed suppression and crop production in annual intercrops. Weed Sci. 53, 813–825. doi: 10.1614/WS-05-014R.1
Szumigalski, A. R., and Van Acker, R. C. (2006). Nitrogen yield and land use efficiency in annual sole crops and intercrops. Agron. J. 98, 1030–1040. doi: 10.2134/agronj2005.0277
Tang, Y., Qiu, Y., Li, Y., Xu, H., and Li, X. F. (2024). Research on intercropping from 1995 to 2021: a worldwide bibliographic review. Plant Soil 506, 7–25. doi: 10.1007/s11104-024-06542-9
Temesgen, A., Fukai, S., and Rodriguez, D. (2015). As the level of crop productivity increases: is there a role for intercropping in smallholder agriculture. Field Crop Res. 180, 155–166. doi: 10.1016/j.fcr.2015.06.003
Thorne, M. E., Young, F. L., and Yenish, J. P. (2007). Cropping systems alter weed seed banks in Pacific northwest semi-arid wheat region. Crop Prot. 26, 1121–1134. doi: 10.1016/j.cropro.2006.10.021
Ünay, A., Sabanci, İ., and Çinar, V. M. (2021). The effect of maize (Zea mays L.) / soybean (Glycine max (L.) Merr.) intercropping and biofertilizer (Azotobacter) on yield, leaf area index and land equivalent ratio. J. Agr. Sci. Tarim Bili. 27, 76–82. doi: 10.15832/ankutbd.572495
VanKoughnet, B. (2015). On-farm evaluation of peaola intercropping. Available online at: https://manitobapulse.ca/wp-content/uploads/2018/02/On-Farm-Evaluation-of-Peaola-Intercropping-2015.pdf (Accessed February 20, 2025).
Wang, Q., Bai, W., Sun, Z., Zhang, D., Zhang, Y., Wang, R., et al. (2021). Does reduced intraspecific competition of the dominant species in intercrops allow for a higher population density? Food Energy Secur. 10:e270. doi: 10.1002/fes3.270
Wang, W., Li, M. Y., Gong, D. S., Zhou, R., Khan, A., Zhu, Y., et al. (2022). Water use of intercropped species: maize-soybean, soybean-wheat and wheat-maize. Agric. Water Manag. 269:107690. doi: 10.1016/j.agwat.2022.107690
Washington State University Wheat and Small Grains (2024). Intercropping on the Palouse with Dr. Garett Heineck. Available online at: https://smallgrains.wsu.edu/wbp194/ (Accessed April 17, 2025).
Wei, W., Liu, T., Shen, L., Wang, X., Zhang, S., and Zhang, W. (2022). Effect of maize (Zea mays) and soybean (Glycine max) intercropping on yield and root development in Xinjiang, China. Agriculture 12:996. doi: 10.3390/agriculture12070996
Westman Agricultural Diversification Organization (2018a). Effect of applied urea and agrotain treated urea in soybean and flax intercrop. WADO 2018 Ann. Rep., 63–67.
Westman Agricultural Diversification Organization (2018b). Relay crop/intercrop legumes in hemp grain production. WADO 2018 Ann. Rep., 68–70.
Whitmore, A. P., and Schröder, J. J. (2007). Intercropping reduces nitrate leaching from under field crops without loss of yield: a modelling study. Eur. J. Agron. 27, 81–88. doi: 10.1016/j.eja.2007.02.004
Williams, J. D., Long, D. S., and Reardon, C. L. (2020a). Productivity and water use efficiency of intensified dryland cropping systems under low precipitation in Pacific northwest, USA. Field Crop Res. 254:107787. doi: 10.1016/j.fcr.2020.107787
Williams, J. D., Reardon, C., Wuest, S. B., and Long, D. S. (2020b). Soil water infiltration after oilseed crop introduction into a Pacific northwest winter wheat–fallow rotation. J. Soil Water Conserv. 75, 739–745. doi: 10.2489/jswc.2020.00165
Wysocki, D. (2021). Peaola sowing rates for intercropping of peas and spring canola. Washington Oilseeds Commission Research Report for 2020-2021, 1–5.
Xu, S., Delgado-Baquerizo, M., Kuzyakov, Y., Wu, Y., Liu, L., Yang, Y., et al. (2024). Positive soil priming effects are the rule at a global scale. Glob. Chang. Biol. 30:e17502. doi: 10.1111/gcb.17502
Yadav, M., Vashisht, B. B., Jalota, S. K., Jyolsna, T., Singh, S. P., Kumar, A., et al. (2024). Improving water efficiencies in rural agriculture for sustainability of water resources: a review. Water Resour. Manag. 38, 3505–3526. doi: 10.1007/s11269-024-03836-6
Young, F. L., Thorne, M. E., and Young, D. L. (2006). Nitrogen fertility and weed management critical for continuous no-till wheat in the Pacific northwest. Weed Technol. 20, 658–669. doi: 10.1614/WT-05-133R.1
Yu, J., Long, A., Lai, X., Elbeltagi, A., Deng, X., Gu, X., et al. (2024). Evaluating sustainable intensification levels of dryland agriculture: a focus on Xinjiang, China. Ecol. Indic. 158:111448. doi: 10.1016/j.ecolind.2023.111448
Zhang, Y., Sun, Z., Su, Z., Du, G., Bai, W., Wang, Q., et al. (2022). Root plasticity and interspecific complementarity improve yields and water use efficiency of maize/soybean intercropping in a water-limited condition. Field Crop Res. 282:108523. doi: 10.1016/j.fcr.2022.108523
Zhang, L., van der Werf, W., Zhang, S., Li, B., and Spiertz, J. H. J. (2007). Growth, yield and quality of wheat and cotton in relay strip intercropping systems. Field Crop Res. 103, 178–188. doi: 10.1016/j.fcr.2007.06.002
Keywords: intercropping, peaola, sustainability, pea, canola
Citation: Stubbs LR, Singh S, Wysocki D, Heineck GC, Neely HL and Singh S (2025) Improving sustainability of inland Pacific Northwest dryland agriculture systems with pea-canola intercropping: a review. Front. Sustain. Food Syst. 9:1639908. doi: 10.3389/fsufs.2025.1639908
Edited by:
Dinesh Jinger, Indian Institute of Soil and Water Conservation (ICAR), IndiaReviewed by:
Hero T. Gollany, United States Department of Agriculture, United StatesRam Swaroop Bana, Indian Agricultural Research Institute (ICAR), India
Venkatesh Paramesha, Central Coastal Agricultural Research Institute (ICAR), India
Copyright © 2025 Stubbs, Singh, Wysocki, Heineck, Neely and Singh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shikha Singh, c2hpa2hhLnNpbmdoQHdzdS5lZHU=