 Orsolya Papp1*†
Orsolya Papp1*† Tamás Kocsis2*†Dániel Ganszky1,3Éva Borbélyné Hunyadi1László Mezőfi1Balázs Labus1
Tamás Kocsis2*†Dániel Ganszky1,3Éva Borbélyné Hunyadi1László Mezőfi1Balázs Labus1 Nora Szigeti1Dóra Selmeczi1Ferenc Tóth1Geoffrey Darbon4,5
Nora Szigeti1Dóra Selmeczi1Ferenc Tóth1Geoffrey Darbon4,5 Dóra Drexler1
Dóra Drexler1- 1Hungarian Research Institute of Organic Agriculture (ÖMKi), Budapest, Hungary
- 2Department of Food Microbiology, Hygiene and Safety, Institute of Food Science and Technology, Hungarian University of Agriculture and Life Sciences, Budapest, Hungary
- 3Geonardo Environmental Technologies Ltd., Budapest, Hungary
- 4Plant Production Systems, Cultivation Techniques and Varieties in Arable Farming, Agroscope, Nyon, Switzerland
- 5Applied Microbiology, Mycology, Earth and Life Institute, Université Catholique de Louvain, Louvain-la-Neuve, Belgium
Potato holds significant importance in human nutrition. However, climatic stress factors such as prolonged drought have increasingly challenged potato production in recent years. Various research efforts are underway to mitigate the adverse effects of abiotic stress in potato agrotechnology, including the use of soil inoculation with beneficial microorganisms. Despite the promising results reported in the literature, there is still limited knowledge about the appropriate technology and microbial strains for field application, particularly in organic potato production. Therefore, a 2-year study was conducted to evaluate the effects of Pseudomonas brassicacearum 3Re2-7, Trichoderma asperelloides A, Rhizophagus irregularis MUCL41833, and Paraburkholderia phytofirmans PsJN strains under two irrigation strategies (irrigated and rainfed) in an organic potato field, using two varieties. Concerning the effects of microbial inoculations, significant differences were observed only in plant stomatal conductance among all measured parameters: inoculation with T. asperelloides significantly decreased stomatal conductance in potato plants. For the other studied parameters (yield, incidence of tuber pathogens, starch and phosphorus content of tubers, and SPAD values), the effects of variety, irrigation, and year were more significant than those of inoculation, regardless of whether strains were applied alone or in combination. The lack of significant inoculation effects may be attributed to open-field environmental conditions, limitations of soil nutrient status, application methodology, and resilience of the rich microbial communities associated with organic agriculture.
1 Introduction
Potato (Solanum tuberosum L.), a member of the Solanaceae family, is the fourth most important food crop (after rice, wheat, and maize) and the primary non-grain food crop in the world. It provides a crucial source of nourishment for more than one billion people worldwide (Sagili et al., 2022). Its cultivation is fertilizer-intensive and water-demanding due to its high nutrient demand and shallow root system (Peralta and Stockle, 2002).
Potato production is subject to various stressors, including heat, drought, salinity, and pest-related stress, which reduce crop performance (Zaki and Radwan, 2022). Given that the frequency and influence of these stressors continuously increase due to climate change, research into agrotechnical practices that enhance the stress tolerance of potatoes is becoming increasingly important (Dahal et al., 2019). Research has explored the impact of drought stress on potatoes and the role of soil microbial strains in enhancing drought tolerance. Studies have investigated various aspects, including the rhizosphere microbiome composition in different potato cultivars under dry conditions (Martins et al., 2023) and the impact of drought stress on soil microbial communities and their role in plant responses to stress conditions (Shaffique et al., 2022).
The use of plant growth-promoting rhizobacteria (PGPR) in sustainable agriculture is gaining widespread adoption worldwide (Santos et al., 2019). For example, one of the aims of the European Commission’s Green Deal is to reduce the use of synthetic fertilizers and support alternative solutions, including treatments with microorganisms that contribute to plant development, increase yields, and mitigate production risks. The use of PGPR as biofertilizers has been highlighted as a sustainable approach to crop production, fostering nutrient acquisition, disease control, and stress tolerance in plants (Hamid et al., 2021; Kumar and Verma, 2018; Lugtenberg et al., 1991), thereby ameliorating drought stress and improving overall agronomic sustainability (Raklami et al., 2019; Shaffique et al., 2022). Based on the major results of four meta-analysis studies (Herrmann et al., 2022; Li et al., 2022; Nkebiwe et al., 2024; Schütz et al., 2018), beneficial microbial inoculants can improve agricultural productivity; however, the overall assessment of their significant effects is made difficult by the selective publication of results (Herrmann et al., 2022; Nkebiwe et al., 2024). The primary mechanism by which microbial inoculants enhanced crop yield is by alleviating stress. The Pseudomonas genus was found to be the most effective in enhancing crop yield through stress alleviation, whereas the Enterobacter genus was the most effective in improving plant nutrient availability (Li et al., 2022). Considering both mechanisms together, the combined inoculant containing Pseudomonas, Enterobacter, and Bacillus was the most effective in enhancing crop yield.
The potato crop hosts numerous communities of native rhizobacteria, which can play a significant role in promoting its growth, as observed in other plants. More than 60 bacterial genera have been isolated from the potato rhizosphere (Diallo et al., 2011); however, understanding the potential and actual effects of beneficial microorganisms on potato production remains limited, especially under field conditions. Inoculation of Rhizophagus irregularis in two field trials could reduce the infection severity of Phytophthora infestans; however, no mycorrhizal effect was observed on tuber yield (Alaux et al., 2018). Larkin (2016) compared commercially available biocontrol formulations to chemical seed treatments in a one-time in-furrow field application. The biocontrol formulations (mainly Bacillus subtilis) reduced Rhizoctonia solani infection and influenced soil microbial community characteristics. The study by Hijri (2016), which analyzed 231 field trials where the same Arbuscular Mycorrhizal Fungi (AMF) inoculant (Rh. irregularis) was applied, reports a significant increase in marketable potato yield.
Organic cultivation of potato accounted for 2.46% area share in the European Union, covering 37.992 hectares in 2022 (FiBL Statistics, 2024). Organic agriculture has been recognized as a potential solution to mitigate the impacts of drought stress on crop production. For instance, soil structure-preserving techniques and reduced tillage can help reduce drought vulnerability and improve resilience to water stress (Knutson et al., 2011). However, research on organic potato production primarily focuses on disease and nutrient management, often in comparison with integrated and conventional systems. Only a few studies investigated the effect of microbial inoculation on organic potato cultivation. According to these studies, AMF inoculation improved the marketable yield and mineral content of potato plants (Lombardo et al., 2020, 2021). In-furrow application of Trichoderma harzianum could reduce the incidence of Rhizoctonia solani infection, but the total yield remained unaffected (Tsror et al., 2001). Papp et al. (2021a) observed that the effects of two combined PGPR consortia on the severity of Rhizoctonia and Streptomyces infections depended on the growing season and the infection pressure from the pathogens.
Our research was performed to assess the efficacy of microbial inoculants in organic potato farming. The tested microbial strains were selected in previous pot experiments: Mamun et al. (2024) investigated the drought protection effects of six fungal and bacterial inoculants and ten consortia on potato under pot and field conditions. Based on the results, Pseudomonas brassicacearum 3Re2-7, Trichoderma asperelloides A, Rhizophagus irregularis MUCL41833, and Paraburkholderia phytofirmans PsJN strains were selected for open-field testing on two sites under two irrigation strategies: one in Switzerland (under a conventional farming system) and the other in Hungary (under organic production), for 3 years. In the Swiss test, Rh. irregularis and P. brassicacearum showed the best pathogen control in combination. This treatment reduced the incidence of common scab and silver scurf-associated symptoms under high pathogen pressure. However, the inoculants did not affect yield (Darbon et al., 2024). In the Hungarian test, the same inoculation treatments were used. The results of the first year (2019) showed no major effects of the microbial inoculants on potato performance under combined stress (Papp et al., 2021b). The results of the following years (2020–2021) are presented in this study.
2 Materials and methods
2.1 Study site and field experiment
The research was carried out in Soroksár (47° 39’ N; 19° 15′E), a district of Budapest (Hungary) at the Organic Educational Farm of the Hungarian University of Agriculture and Life Sciences (MATE) during the 2020–2021 seasons. The farm has been organically cultivated for more than a decade. The site is situated at an altitude of less than 200 m and has a moderately hot and dry continental climate. The average annual precipitation is approximately 500–550 mm (Kocsis et al., 2022). Meteorological data were recorded continuously during the trial, and the means of monthly precipitation, temperature, relative air humidity, soil temperature, and leaf wetness values for the growing seasons of 2021 and 2021, are shown in Tables 1, 2, respectively.
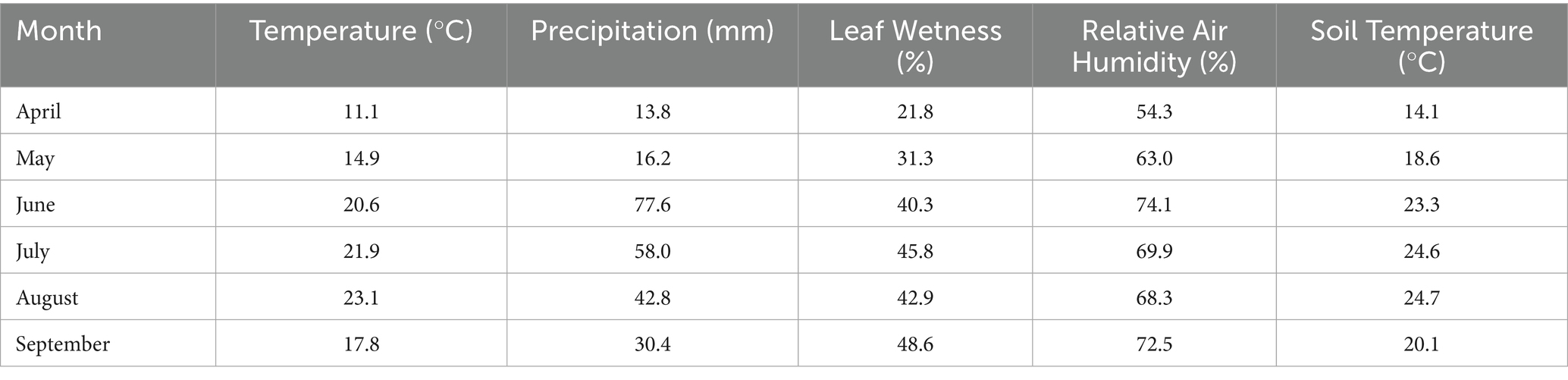
Table 1. Monthly means of temperature (°C), precipitation (mm), leaf wetness (%), relative air humidity (%), and soil temperature (°C) during the vegetation period in the experimental site in 2020 (Soroksár, Hungary).
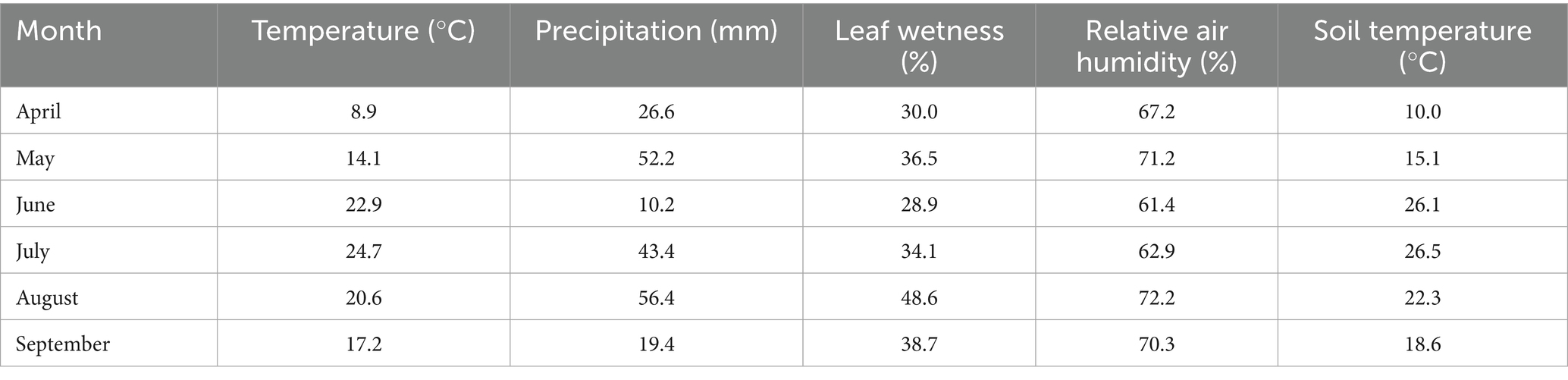
Table 2. Monthly means of temperature (°C), precipitation (mm), leaf wetness (%), relative air humidity (%), and soil temperature (°C) during vegetation period in the experimental site in 2021 (Soroksár, Hungary).
The soil-forming rock under the trial site consists of glacial and alluvial sediments, and the type of soil is sandy, with a low humus content (Arenosol) (MePAR, 2013). The key soil characteristics are as follows: pH (KCl): 7.9, humus content 2.33 m/m%, CaCO₃ 9.17 m/m%, NO₃-N 10.3 mg kg−1, AL-P₂O₅ 428 mg kg−1, AL-K₂O 121 mg kg−1, and Cu 17 mg kg−1.
Rye was used as the pre-crop in both years. The potato tubers were pre-sprouted for 5 weeks, and the average temperature in the sprouting room was 10.4°C in 2020 and 15.8°C in 2021. The pre-sprouted tubers were planted on 22.03.2020 and 26.04.2021. Organically certified fertilizers were used at the time of planting, consisting of 120 kg ha−1 nitrogen (N), 36 kg ha−1 phosphorus (P), 372 kg ha−1 potassium (K), and 20 kg ha−1 magnesium (Mg). Ridges measuring 20–30 in height were formed after the shoots emerged. The potato field was managed in accordance with the EU legislation on organic production (834/2007/EK). Weeds were controlled manually, and only permitted substances were used for plant protection (For example, copper was used against Phytophthora infestans, and Bacillus thuringiensis and spinosad were used to combat Leptinotarsa decemlineata). The amount of irrigation water only in the irrigated treatments was 120 mm (2020) and 182 mm (2021). The harvests were carried out on 08.09.2020 and 09.09.2021.
2.2 Study design and the method of potato inoculation
The trial was designed as a three-factor split-split-plot experiment with irrigation, inoculation, and variety as plot, sub-plot, and sub-subplot factors (respectively). This setup included a total of 64 plots. Each plot was 13.5 m2 (2.25 m x 6 m) in size and consisted of three rows of potatoes: one row of Pentland Dell and two rows of Desirée cultivars (one row for measuring yield and one row for destructive sampling). A total of 24 tubers were planted in a row, and the spacing was 0.75 cm (interrow) x 0.25 cm (intra-row). To avoid border effects, all plots as well as the entire field were surrounded by buffer potato rows. All combinations of variety, inoculation, and irrigation were present with four replications in the experiment.
The application of irrigation water was based on the calculations from the Metagro crop production software. This software calculated the required amount of irrigation water using potato crop models and the local climatic data collected in a meteorological station installed at the center of the trial site.1 The plots without regular irrigation relied solely on rainfall, except during serious drought periods when minimal irrigation was necessary to ensure the survival of the plants.
To investigate how different strains affect the stress tolerance of potato cultivars, seven types of treatments were applied in comparison to an untreated control. The four individual strains and their mixtures are listed in Table 3.
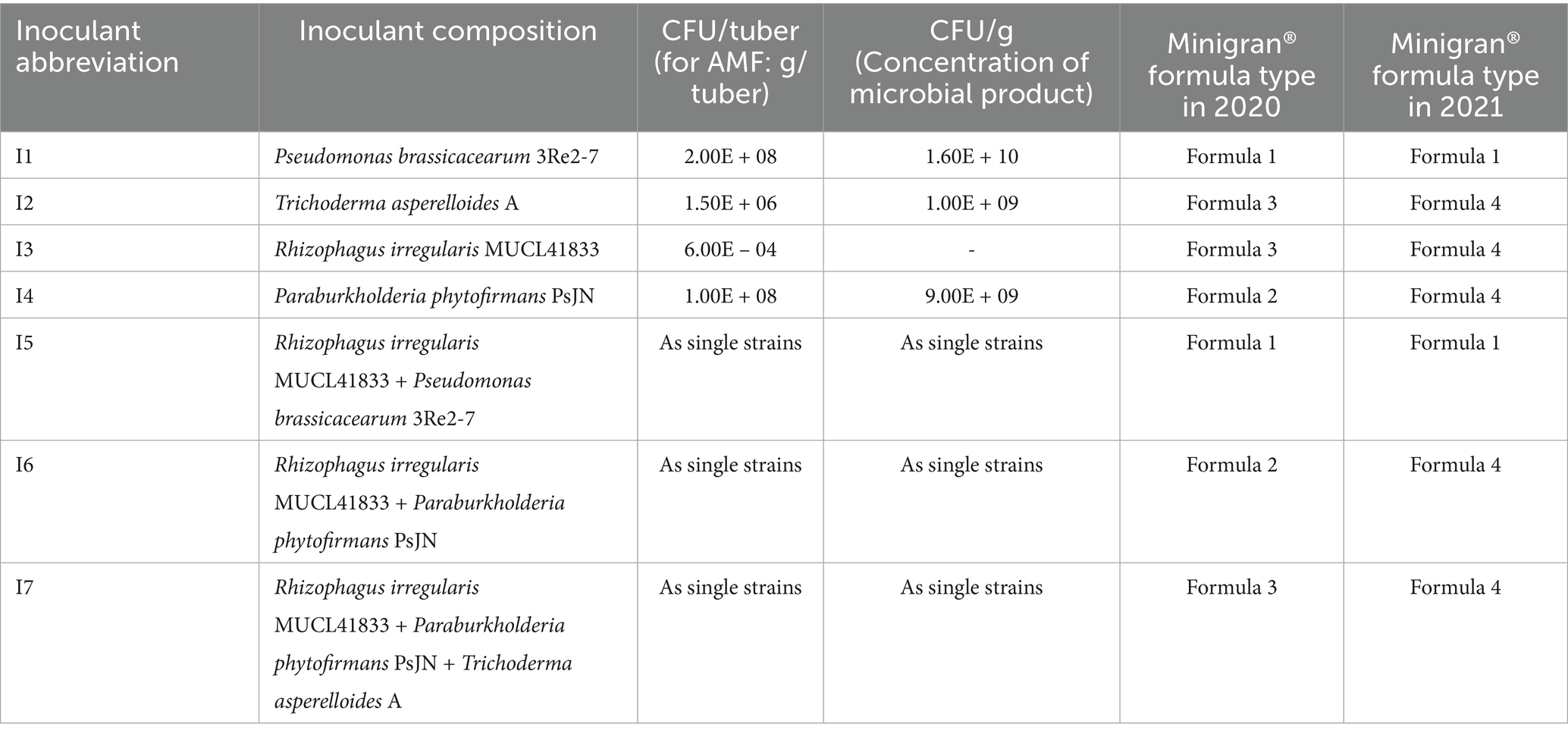
Table 3. Composition, concentration, and formulation types of the microbial inoculants applied in the experiment in 2020–2021.
The microbial strains were selected based on previous research: 6 strains and their 10 combinations were tested and evaluated in greenhouse pot experiments, and the best-performing strains were proposed for on-field research (Mamun et al., 2024). The selected inoculants were formulated with the Minigran® formula developed by De Ceuster Meststoffen (DCM, Belgium). All variations of the Minigran® formula contained soy, vinasse, tobacco, and peat-based materials; however, four different formulations were used in 2020 and 2021 with minor differences in their composition, in order to improve the survival of the strains: Minigran® 1 (NPK 3–0.6 – 2.7), Minigran® 2 (4–0.3 – 2), Minigran® 3 (4.4–0.4 – 1.9), and Minigran® 4 (5–0.5 – 2) (Darbon et al., 2024). The inoculation was performed at planting by placing 0.75 g of the Minigran® formula per tuber on pre-sprouted tubers, after which the tubers were directly covered with soil.
2.3 Pre-harvest plant measurements
SPAD values were measured by a chlorophyll meter SPAD-502Plus (Konica Minolta, Osaka, Japan). The measurements were taken on matured, healthy, green flag leaves of 5 plants/row, on 30.06.2020, 22.07.2020, 24.06.2021, and 04.08.2021.
Stomatal conductivity was measured using an AP4 Porometer (AT Delta-T Devices, Cambridge, United Kingdom) on 30.06.2020, 22.07.2020, 24.06.2021, and 29.07.2021. Measurements were taken between 11:00 and 14:00 under a cloudless sky, with one leaf taken from each of five plants per row.
Aerial data collection was performed using a Parrot Bluegrass Fields agricultural monitoring drone (Parrot SA, France) equipped with a Sequoia+ 4-band multispectral camera and a FLIR One Pro TIR camera (FLIR Systems Inc., USA) mounted on the top of the UAV at flight altitude of 15 m, resulting in a GSD of 1.4 cm/pixel GSD for the multispectral data and an effective GSD of 10 cm/pixel for the thermal data. The orthomosaics were created based on photogrammetry and location data in Pix4Dmapper software. On the field, eight 33 × 33 cm VIS-LWIR GCPs were placed for precise georeferencing, and their position was recorded using a Qpad X8 GNSS RTK Rover. The TIR emissivity index was 95%, and the reference target for multispectral reflectance calibration was grey (Green: 18.6%, Red: 19.9%, RE: 22.9%, and NIR: 26.3%).
The soil was filtered out of the orthomosaics by OSAVI binarized at 0.4, determined by the bimodal peaks on their histograms. This mask was applied to both the multispectral reflectance and TIR orthomosaics; thus, the vegetation index (NDVI) and canopy temperature (°C) calculations only included pixels of the vegetation, providing information about soil coverage or vegetation area (expressed as m2/row). The parcels were vectorized with a positive buffer, and then zonal statistics were calculated for each row in QGIS 3.10 LTR depending on coverage. Aerial data were collected on 25.06.2020, 21.07.2020, 29.06.2021, 13.07.2021, and 28.07.2021.
2.4 Post-harvest plant measurements
After harvest, the total yield (kg fresh weight/row) was measured in each row. Then, the tubers were sorted into five classes based on their diameter: <35 mm, 35 < 42 mm, 42 < 55 mm, 55 < 70 mm, and >70 mm. The tubers within each fraction were counted and weighed.
The quality of the potato tubers was assessed on 25 tubers per row, representing one sample. Tubers were washed in clean water and symptoms of common scab (CS—caused by Streptomyces spp., several types of lesions), black scurf (BS—caused by Rhizoctonia solani, raised black spots), silver scurf (SS—caused by Helminthosporium solani, Colletotrichum coccodes, small light grayish leathery spots), and dry rot (DR—caused by Fusarium spp., dark depressions on the surface of tuber) were visually assessed and infected tubers were counted. These pathological features were assessed directly after harvest, during the days: 2020.09.14–18. and 2021.09.13–17.
The starch content of tubers was also measured after harvesting, by weighing 5 kg of potatoes in air and water, using the so-called gravity method (Miernik and Jakubowski, 2021). The starch content was calculated using the following formula: starch content (%) = ((Wa/(Wa – Ww)) - 1.01506)/0.0046051, where Wa (g) = weight in air, Ww (g) = weight in water, and Wa/(Wa – Ww) = tuber density. The phosphorus content (%) of the potato tubers was determined on samples of five tubers from each row, according to the Hungarian MSZ 21470–50:2006 standard.
2.5 Data analysis
Regarding the SPAD and stomatal conductance data, the arithmetic (yearly) means of the multiple measurements made in each experimental row were used for the analyses. In addition to the incidence of the four pathogens, another ordinal variable (hereafter cumulative infection) was created to indicate the cumulative presence of the different pathogens in a potato tuber sample (containing 25 tubers). For this variable, zero indicates no infection, and four indicates that the tuber sample was affected by all four pathogens.
All statistical analyses were performed within the R (v.4.2.2.) statistical environment.2 The putative effects of the different predictors on the response variables were tested using various linear mixed models (LMMs) and generalized linear mixed models (GLMMs) using the “lme4” R package.3 For the cases where the response variable was counts (e.g., pathogen incidence, count of potato tubers affected by the given pathogen in a sample of 25 tubers), GLMMs assuming a Poisson error structure (GLMM-p) were used. In other cases, we ran linear models assuming a normal distribution. The initial model structures consisted of the following predictors: Inoculant (an eight-level factor), Variety, Irrigation, and Year (the latter three were two-level factors). Besides the above-mentioned four predictors, the models included the main interaction terms, Inoculant x Variety and Inoculant x Irrigation (only these two interaction terms to avoid over-parameterization of the models), and plot IDs as a random effect. The “Anova” function (Type II) from the R package “car”4 was used to obtain the effects of the predictors on the response variable. To avoid misleading results based on statistical artifacts, numerous model diagnostic statistics were performed using the R package “DHARMa.”5 In our analysis, we paid particular attention to ensuring that the assumptions of the models were not violated. The R package “emmeans”6 was used to infer pairwise contrasts. In the case of multiple comparisons, the “eff_size” function of the package was used to obtain the effect sizes (Cohen’s D) and related confidence intervals for every pairwise difference. For making the plots, various functions of the R package “ggplot2”7 were used.
3 Results
Inoculation did not affect potato yield or the distribution of tuber sizes (Table 4). However, variety (in six out of seven cases) and irrigation (in all cases) significantly affected yield: cv. Desirée produced more than cv. Pentland Dell, regardless of variety, showed irrigation increased both the total and net weight of potatoes (kg/row) (Figure 1, Table 4, Supplementary Tables 1, 2). The year only affected the net weight of potatoes (Table 4, Supplementary Tables 1, 2).
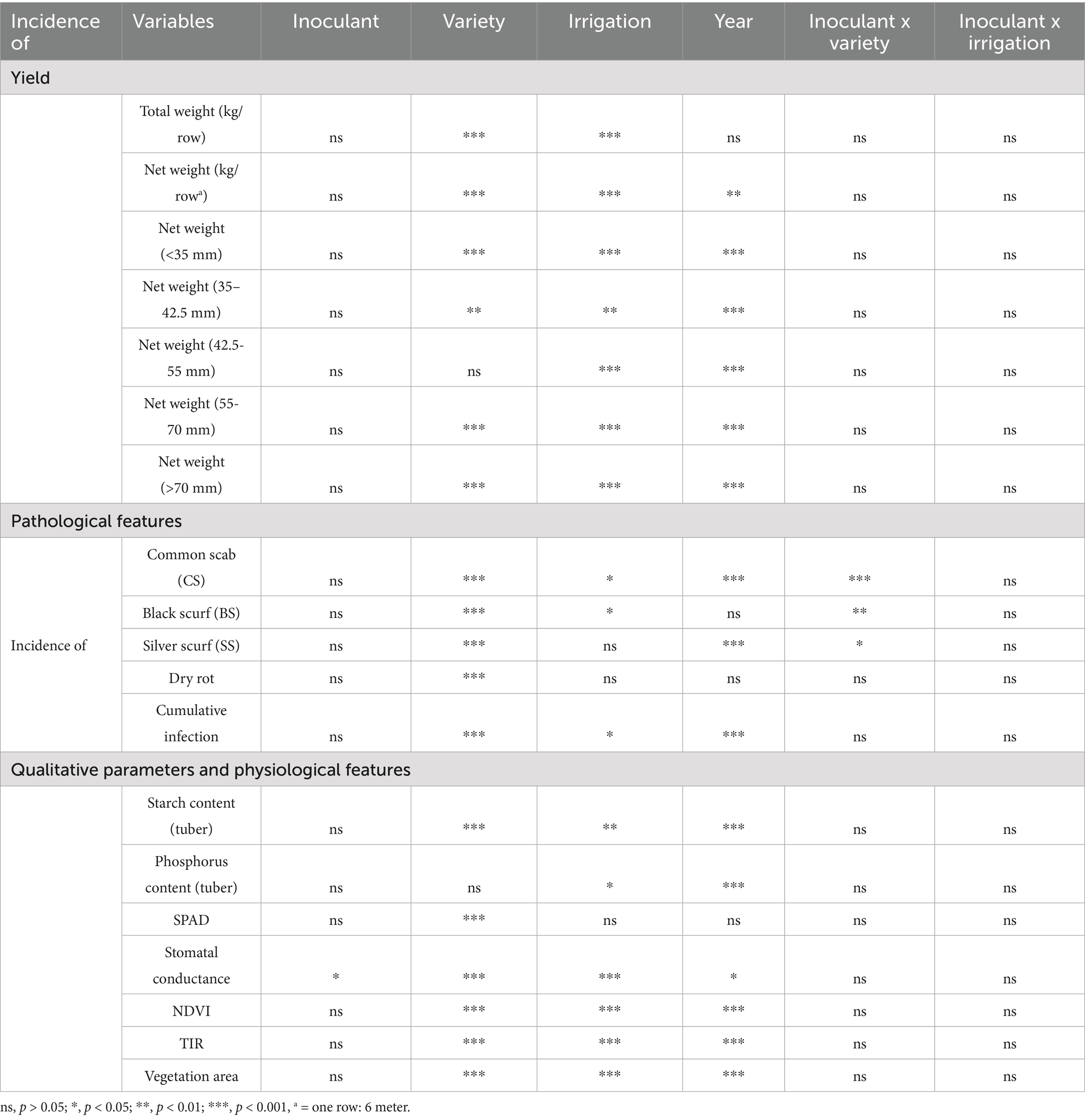
Table 4. Summary of significance levels for the effects of inoculant, variety, irrigation, and year on potato yield, tuber quality, and physiological traits, based on linear and generalized linear mixed models (LMMs and GLMMs).
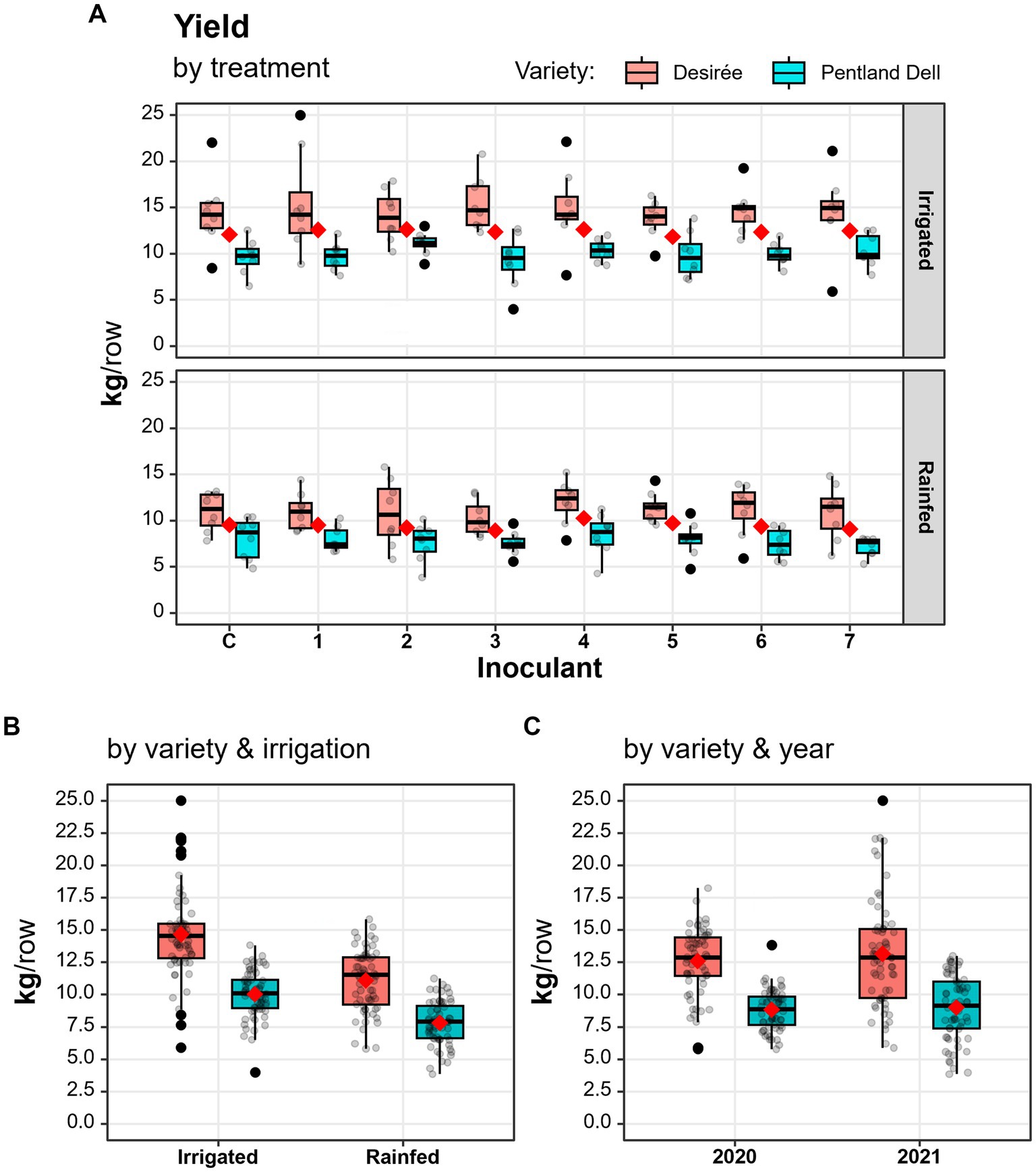
Figure 1. Yield (kg/row) of the two potato varieties by inoculant treatment (A), water supply (B), and year (C). C: Control plots, Inoculant1—Inoculant7: Inoculated plots (see Table 3). The distance between the box bottom (first quartile) and top (third quartile) corresponds to the interquartile, while the whisker shows the non-outlier range. The red diamond and the bold horizontal line indicate mean and median values, respectively. Grey dots indicate the yield of individual experimental rows. Data points were jittered horizontally. (A,B) Data were pooled across years.
Inoculation did not affect the relative incidence of pathogens or cumulative infection of potato samples (Table 4, Supplementary Tables 1, 3). However, susceptibility to pathogens differed significantly between the two cultivars: cv. Pentland Dell was more susceptible to common scab, black scurf, and dry rot, whereas cv. Desirée was more affected by silver scurf (Table 4, Supplementary Tables 1, 3, Supplementary Figures 1, 2). Furthermore, in general, cv. Pentland Dell samples were more likely to be infected by multiple pathogens than the samples of cv. Desirée (see Cumulative infection, Table 4, Supplementary Tables 1, 3). Irrigation increased the incidence of symptoms associated with common scab and black scurf (regarding black scurf, see Supplementary Figure 1). In addition, under irrigated conditions, potato samples were more likely to be infected with multiple pathogens, regardless of variety (Table 4, Supplementary Tables 1, 3). The year also influenced the incidence of common scab, silver scurf, and the cumulative infection (Table 4, Supplementary Tables 1, 3). Significant interaction between Inoculant and Variety was detected for the following response variables: incidence of common scab, black scurf, and silver scurf (Table 4, Supplementary Table 3). Therefore, in these cases, the varieties were analyzed separately. However, inoculation did not affect the concerned pathological variables in either case (not shown).
Inoculation had no impact on the qualitative parameters of potato tubers such as starch and phosphorus content (Table 4, Supplementary Tables 1, 4). On average, the highest starch content was detected in plots inoculated with T. asperelloides single (Inoculant2) under irrigated conditions (Supplementary Figure 3). The two potato varieties differed in starch content (cv. Pentland Dell had a higher starch content) but not in phosphorus content (Table 4, Supplementary Tables 1, 4, Supplementary Figure 3). Irrigation increased both the starch and phosphorus content of the tubers, and year influenced both parameters of potato samples (Table 4, Supplementary Tables 1, 4).
While most physiological features were not affected by microbial inoculations, stomatal conductance differed significantly between some treatments (Table 4, Supplementary Tables 1, 4, Figure 2). Compared with the control, inoculation of T. asperelloides (Inoculant2) significantly decreased the stomatal conductance of potato plants (Figures 2, 3). Besides this, T. asperelloides treatment differed significantly from Rh. irregularis treatment (Inoculant3) and from the treatment comprising Rh. irregularis, P. phytofirmans, and T. asperelloides strains together (Inoculant7) (Figures 2, 3).
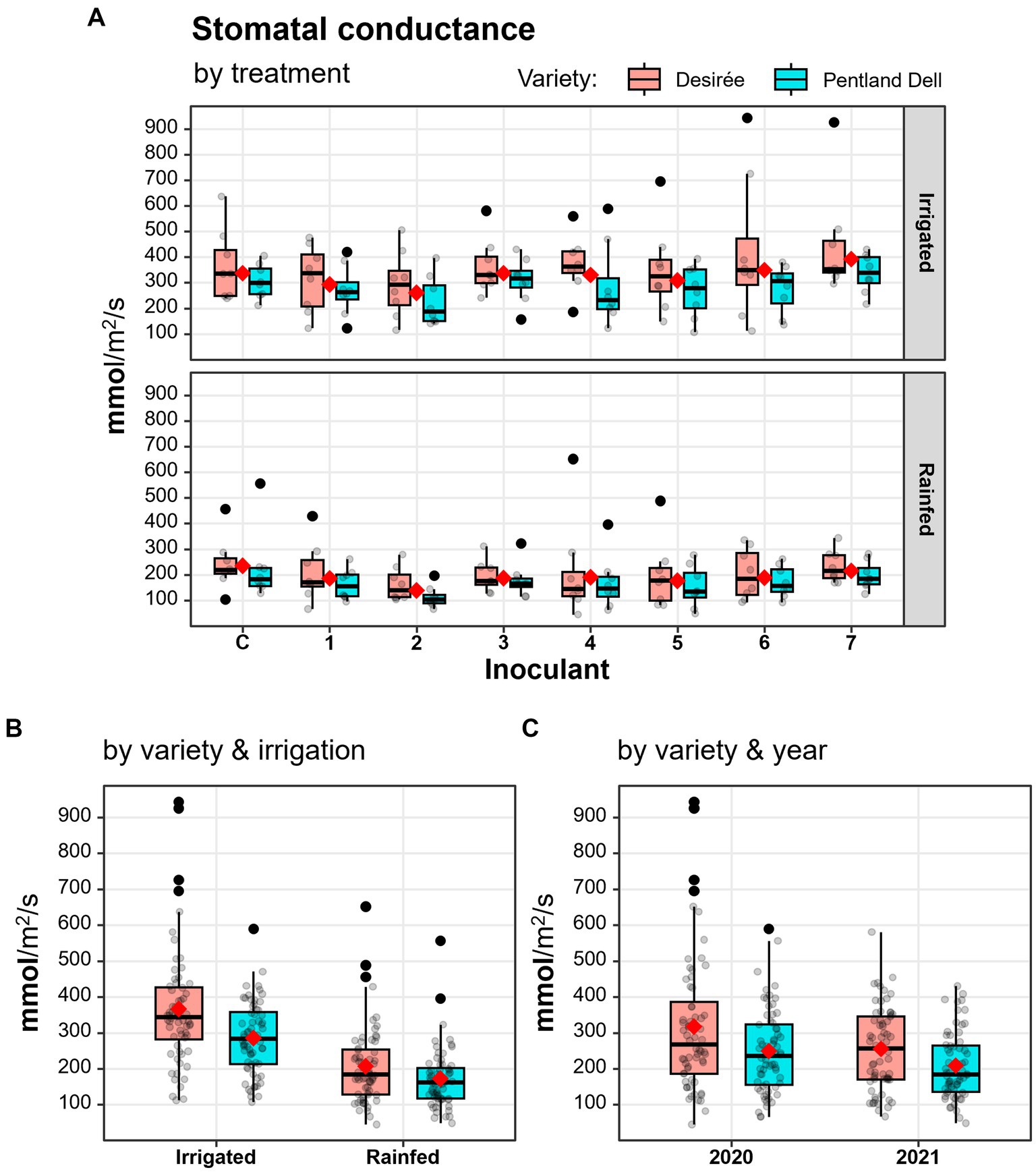
Figure 2. Stomatal conductance (mmol/m2/s) of the two potato varieties by inoculant treatment (A), water supply (B), and year (C). C: Control plots, Inoculant1—Inoculant7: Inoculated plots (see Table 3). The distance between the box bottom (first quartile) and top (third quartile) corresponds to the interquartile, while the whisker shows the non-outlier range. The red diamond and the bold horizontal line indicate mean and median values, respectively. Grey dots indicate the mean stomatal conductance values of the potato plants measured within a given experimental row. Data points were jittered horizontally. (A,B) Data were pooled across years.
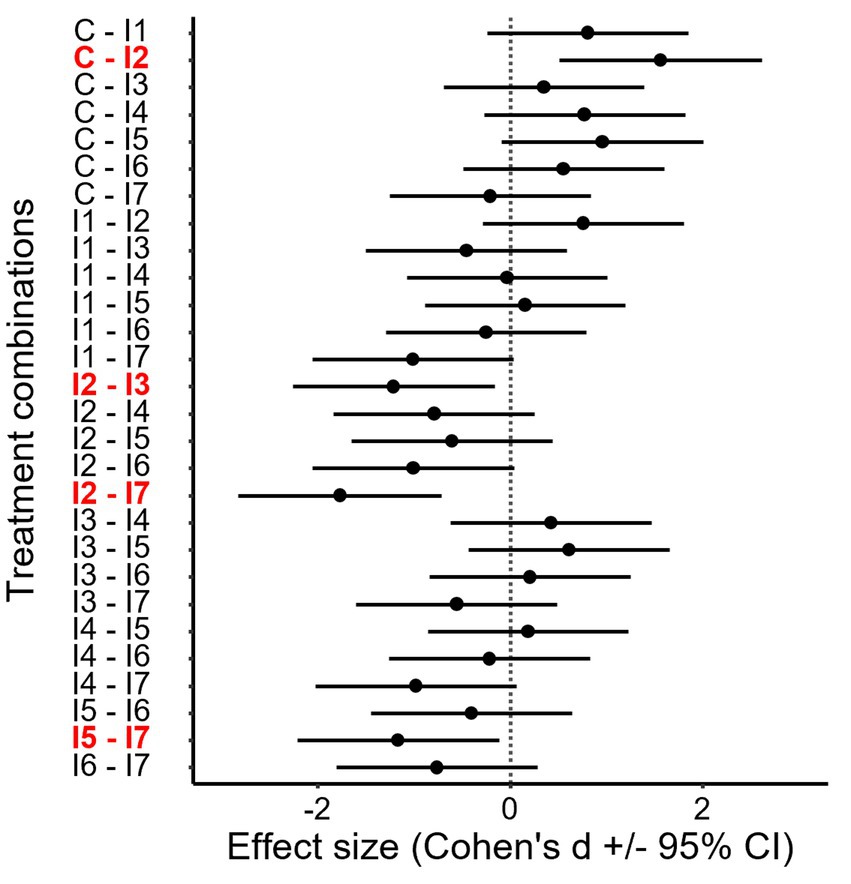
Figure 3. Pairwise comparisons of treatment mean for the stomatal conductance of potato plants. The effect size and corresponding confidence interval (CI) show the difference between the two group means. The difference is considered significant if the given 95% CI does not overlap with zero (indicated by bold red text). For the applied inoculants (Inoculant1—Inoculant7), see Table 3.
The varieties differed significantly in relative chlorophyll content (based on SPAD values), stomatal conductance, vegetation index (NDVI), canopy temperature (TIR), and vegetation area (seen from above), with cv. Desirée showed higher values for all variables except the canopy temperature (Table 4, Supplementary Tables 1, 4). Irrigation increased stomatal conductance, vegetation index, and vegetation area, but decreased canopy temperature (Table 4, Supplementary Tables 1, 4). The 2 years also differed in terms of the four physiological variables mentioned previously (Table 4, Supplementary Tables 1, 4).
4 Discussion
Regarding the effect of PGPR treatments, significant differences were only found in the stomatal conductance of all the tested parameters in our research. Compared to the control, inoculation of T. asperelloides (Inoculant2) significantly decreased the stomatal conductance of potato plants. Moreover, the treatment of T. asperelloides differed significantly from the treatment with the consortium containing Rh. irregularis, P. phytofirmans, and T. asperelloides strains (Inoculant7).
Stomatal conductance in potato plants plays a key role in regulating water loss through transpiration and influencing photosynthetic rates under different environmental conditions. Under water stress, the closing of stomata reduces the rate of transpiration, which limits water loss in plants. This action can be seen as part of the plant’s adaptive strategy to cope with stress (Boguszewska-Mańkowska et al., 2017). Factors such as irrigation, nitrogen management, and mycorrhization have also been shown to influence the stomatal conductance of potatoes, emphasizing the importance of agronomic practices in modulating plant physiological responses (Kingori et al., 2016; Lombardo et al., 2020; Shrestha et al., 2024).
Although the primary area of action of T. asperelloides is biocontrol activity, some studies report effectiveness in improving drought tolerance in rice and tomato by enhancing the expression of specific genes (Quazi et al., 2024; Rawal et al., 2023). For example, if a strain or product causes the stomata to either close more frequently or open less regularly, it reduces the rate of gas exchange (Harrison et al., 2020). By controlling water loss, the plant can use available water more efficiently, potentially helping it to survive in water-limited environments. The results of our experiment suggest that T. asperelloides is worth further investigation as a potential tool for mitigating the negative effects of drought stress.
For all other parameters studied (yield, tuber infection, starch and phosphorus content of tubers, SPAD values), the effects of variety, irrigation, and year were more apparent than those of inoculation, applied either alone or in consortium. The lack of significant effects of inoculation can be attributed to several factors related to the specific interactions between these microorganisms, soil microbiota, and the potato plant, as well as environmental conditions. The complexity of soil ecosystems, meaning the complexity of interactions between soil and plant microbiota, may influence the efficacy of application, leading to inconsistent results of Plant Growth-Promoting Microbes (PGPM) application (Hart et al., 2017; Hoeksema et al., 2010; Kaminsky et al., 2019; Malgioglio et al., 2022; Mayer et al., 2010). The physical, chemical, and physicochemical properties of soil, as well as its indigenous microorganisms, can influence the microbial population (Khare and Arora, 2015; Malusà et al., 2016; Nkebiwe et al., 2024; Rivett et al., 2018).
For example, Nkebiwe et al. (2024) and Schütz et al. (2018) observed declining bioeffector responses at both the lowest phosphorus (P) availability and higher P levels. In addition, for Rh. irregularis, many studies have shown that elevated levels of phosphorus can inhibit AMF colonization in plant roots, primarily due to the ability of plants to meet their phosphorus needs independently, thereby reducing their reliance on AMF for nutrient acquisition (Breuillin et al., 2010; Ilyas et al., 2024; Jansa et al., 2006; Lombardo et al., 2020; Lekberg and Koide, 2005; Verbruggen et al., 2013; Zhang et al., 2021). Therefore, the high phosphorus content of the soil of the study site (see Materials and Methods section) may have prevented the strain’s establishment.
Recent findings also suggest that the success of AMF inoculation is influenced not only by nutrient levels but also by the structure and composition of the indigenous soil microbiome. For instance, Lutz et al. (2023) demonstrated that the presence of specific microbial indicators, particularly pathogenic fungi, was more predictive of plant growth response to AMF inoculation than soil nutrient status alone. In a similar vein, Rog et al. (2025) discovered that AMF inoculation tends to be more effective in soils with reduced health and productivity. In such cases, the mutualistic fungi can compensate for limitations in nutrient availability and microbial community function. The findings of this study support the hypothesis that, in addition to soil P levels, broader biotic interactions play a crucial role in determining the establishment and efficacy of arbuscular mycorrhizal fungi (AMF).
Moreover, Schütz et al. (2018) reported a decrease in yield response not only for P-solubilizing microbes but also for AMF with increasing levels of soil organic matter. The soil at the study site, with approximately 2.3% humus, lies near the upper threshold of organic matter content associated with active yield responses. In line with this, Nkebiwe et al. (2024) also observed a reduced bioeffector response in yield with increasing soil organic carbon content. A higher organic matter content is generally associated with enhanced native microbial activity (Francioli et al., 2016; Lori et al., 2020), which could intensify competition with the applied inoculants, potentially diminishing their establishment and efficacy.
Trichoderma species, including T. asperelloides, have been shown to thrive in slightly acidic conditions, with optimal growth typically occurring at pH levels ranging from 5 to 7. It was also reported that Trichoderma species produced more biomass in acidic environments, which is favorable for their growth compared to alkaline conditions (Ghazanfar et al., 2018; Singh et al., 2013). The slightly alkaline pH of the study site (7.9) may have negatively affected the survival of these microorganisms. This finding aligns with the meta-study by Herrmann et al. (2022), which reported higher responsiveness to bioeffector treatments in more acidic environments.
On the other hand, experiencing no significant differences could also be caused by microbial inoculants sometimes failing to establish or survive in their target environment, which could explain the absence of yield effects. Survival is probable if bacteria reach the roots and can utilize root exudates as substrate (Malgioglio et al., 2022). The native microbial community of the soil also has a significant influence on inoculant survival, which can be recessive or restrictive, depending on microbial growth rate and the availability of necessary nutrients in the soil (Malgioglio et al., 2022). The rich and diverse microbial life in soils under organic farming systems is a well-documented phenomenon, characterized by enhanced microbial abundance, diversity, and functional capacity compared to conventional farming practices. Lori et al.’s (2017) meta-analysis found that organic farming has a positive impact on microbial communities, significantly enhancing six of seven key indicators of microbial abundance and activity compared to conventional systems. Higher microbial community abundance and activity could mean a more competitive environment for inoculants. Altogether, confirming inoculum colonization and survival in planta (via RNA-sequencing during the vegetation period) would provide additional support for interpreting inoculation outcomes.
5 Conclusion
Microbial inoculation of agricultural crops is an exciting and dynamically evolving field of science. The application of beneficial bacteria and fungi to plants enhance understanding and contributes knowledge about the composition and function of the soil microbial community, especially the rhizosphere. Successful inoculants and their combinations have been formulated into products in previous years, whereas testing of new strains continues to address the challenges we face, such as increasing water and nutrient deficiencies, as well as the combined stresses caused by climate change.
The tested T. asperelloides strain was successful in reducing stomatal conductance in potato, which suggests it could be a useful tool for enhancing potato drought tolerance. In order to investigate the reasons why no significant changes were observed in the other parameters studied, further research is necessary. It is known that soils in long-term organic farming exhibit greater suppressiveness against external plant pathogens. Additionally, soil health may affect the efficacy of beneficial microorganism applications. For this reason, preliminary studies of local soil microbiota are recommended before applying PGPMs to define interventions based on local soil conditions, environmental factors, crops, and cultivated genotypes. Future studies should examine the optimal timing and composition effectiveness of inoculation. If research indicates that bulk soil microbial community composition changes significantly across potato growth stages (Overbeek et al., 2021), then a detailed study of the microorganisms associated with each phenophase of potato is necessary for a more targeted approach.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material; further inquiries can be directed to the corresponding authors.
Author contributions
OP: Investigation, Project administration, Supervision, Writing – original draft, Writing – review & editing. TK: Supervision, Writing – review & editing. DG: Investigation, Project administration, Supervision, Writing – review & editing. ÉH: Investigation, Methodology, Supervision, Writing – review & editing. LM: Data curation, Formal analysis, Software, Visualization, Writing – review & editing. BL: Data curation, Formal analysis, Software, Writing – review & editing. NSz: Data curation, Formal analysis, Writing – original draft. DS: Writing – original draft. FT: Conceptualization, Supervision, Validation, Writing – review & editing. GD: Methodology, Writing – review & editing. DD: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was carried out as part of the SolACE - Solutions for Improving Agroecosystem and Crop Efficiency for Water and Nutrient Use project, funded by the European Union’s Horizon 2020 research program under Grant Agreement No. 727247.
Acknowledgments
The authors owe their sincere appreciation to Michelle Van Dyck, Hervé Dupré de Boulois, Annelies Justé, and Inge Hanssen from De Ceuster Meststoffen (DCM) SA for preparing the Minigran® formulas required to perform field inoculations and for accepting to develop and share a tracing tool for their strain T. asperelloides A. We also thank Karin Mai from Sourcon Padena for providing P. brassicacearum 3Re2-7 colonies and primer pairs, Angela Sessitsch and Hanna Faist from the Austrian Institute of Technology for providing P. phytofirmans PsJN DNA extracts and detection protocols. Many thanks to Günter Neumann (Universität Hohenheim) for his advice and useful references, to Philippe Hinsinger for his helpful and effective coordinating of SolACE project, to Gábor Balázs (the head of the training farm of Hungarian University of Agriculture and Life Sciences Institute of Horticultural Sciences) for using the study site, to Barbara Ferschl (farm division manager) for her continuous support in fieldworks.
Conflict of interest
DG was employed by the Geonardo Environmental Technologies Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2025.1641878/full#supplementary-material
Footnotes
2. ^https://www.R-project.org/
3. ^https://CRAN.R-project.org/package=lme4
4. ^https://CRAN.R-project.org/package=car
5. ^https://CRAN.R-project.org/package=DHARMa
References
Alaux, P.-L., César, V., Naveau, F., Cranenbrouck, S., and Declerck, S. (2018). Impact of Rhizophagus irregularis MUCL 41833 on disease symptoms caused by Phytophthora infestans in potato grown under field conditions. Crop Prot. 107, 26–33. doi: 10.1016/j.cropro.2018.01.003
Boguszewska-Mańkowska, D., Pieczyński, M., Wyrzykowska, A., Kalaji, H. M., Sieczko, L., Szweykowska-Kulińska, Z., et al. (2017). Divergent strategies displayed by potato (Solanum tuberosum L.) cultivars to cope with soil drought. J. Agron. Crop Sci. 204, 13–30. doi: 10.1111/jac.12245
Breuillin, F., Schramm, J., Hajirezaei, M., Ahkami, A. H., Favre, P., Druege, U., et al. (2010). Phosphate systemically inhibits development of arbuscular mycorrhiza in Petunia hybrida and represses genes involved in mycorrhizal functioning. Plant J. 64, 1002–1017. doi: 10.1111/j.1365-313x.2010.04385.x
Dahal, K., Li, X.-Q., Tai, H., Creelman, A., and Bizimungu, B. (2019). Improving potato stress tolerance and tuber yield under a climate change scenario – a current overview. Front. Plant Sci. 10:563. doi: 10.3389/fpls.2019.00563
Darbon, G., Declerck, S., Riot, G., Doubell, M., and Dupuis, B. (2024). Inoculation and tracking of beneficial microbes reveal they can establish in field-grown potato roots and decrease blemish diseases. Biol. Fertil. Soils 60, 699–712. doi: 10.1007/s00374-024-01822-z
Diallo, S., Crépin, A., Barbey, C., Orange, N., Burini, J. F., and Latour, X. (2011). Mechanisms and recent advances in biological control mediated through the potato rhizosphere. FEMS Microbiol. Ecol. 75, 351–364. doi: 10.1111/j.1574-6941.2010.01023.x
FiBL Statistics Forschungsinstitut für Biologischen Landbau (FiBL): Data on organic agriculture in Europe. Area data for selected crops (2024). Available online at: https://statistics.fibl.org/europe/selected-crops-europe.html (Accessed September 10, 2024).
Francioli, D., Schulz, E., Lentendu, G., Wubet, T., Buscot, F., and Reitz, T. (2016). Mineral vs. organic amendments: microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 7:1446. doi: 10.3389/fmicb.2016.01446
Ghazanfar, M. U., Raza, M., and Raza, W. (2018). Effect of physiological parameters on mass production of Trichoderma species. Pakistan J Phytopathol 30:61. doi: 10.33866/phytopathol.030.01.0447
Hamid, B., Zaman, M., Farooq, S., Fatima, S., Sayyed, R. Z., Baba, Z. A., et al. (2021). Bacterial plant biostimulants: a sustainable way towards improving growth, productivity, and health of crops. Sustainability 13:2856. doi: 10.3390/su13052856
Harrison, E. L., Arce Cubas, L., Gray, J. E., and Hepworth, C. (2020). The influence of stomatal morphology and distribution on photosynthetic gas exchange. Plant J. 101, 768–779. doi: 10.1111/tpj.14560
Hart, M. M., Antunes, P. M., Chaudhary, V. B., and Abbott, L. K. (2017). Fungal inoculants in the field: is the reward greater than the risk? Funct. Ecol. 32, 126–135. doi: 10.1111/1365-2435.12976
Herrmann, M. N., Wang, Y., Hartung, J., Hartmann, T., Zhang, W., Nkebiwe, P. M., et al. (2022). A global network meta-analysis of the promotion of crop growth, yield, and quality by bioeffectors. Front. Plant Sci. 13:816438. doi: 10.3389/fpls.2022.816438
Hijri, M. (2016). Analysis of a large dataset of mycorrhiza inoculation field trials on potato shows highly significant increases in yield. Mycorrhiza 26, 209–214. doi: 10.1007/s00572-015-0661-4
Hoeksema, J. D., Chaudhary, V. B., Gehring, C. A., Johnson, N. C., Karst, J., Koide, R. T., et al. (2010). A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecol. Lett. 13, 394–407. doi: 10.1111/j.1461-0248.2009.01430.x
Ilyas, U., du Toit, L. J., Hajibabaei, M., and McDonald, M. R. (2024). Influence of plant species, mycorrhizal inoculant, and soil phosphorus level on arbuscular mycorrhizal communities in onion and carrot roots. Front. Plant Sci. 14:1324626. doi: 10.3389/fpls.2023.1324626
Jansa, J., Wiemken, A., and Frossard, E. (2006). The effects of agricultural practices on arbuscular mycorrhizal fungi. Geol. Soc. Lond., Spec. Publ. 266, 89–115. doi: 10.1144/GSL.SP.2006.266.01.08
Kaminsky, L. M., Trexler, R. V., Malik, R. J., Hockett, K. L., and Bell, T. H. (2019). The inherent conflicts in developing soil microbial inoculants. Trends Biotechnol. 37, 140–151. doi: 10.1016/j.tibtech.2018.11.011
Khare, E., and Arora, N. K. (2015). “Effects of soil environment on field efficacy of microbial inoculants” in Plant microbes Symbiosis: Applied facets. ed. N. Arora (New Delhi: Springer).
Kingori, G. G., Nyamori, A. J., and Khasungu, I. D. (2016). Improving seed potato leaf area index, stomatal conductance and chlorophyll accumulation efficiency through irrigation water, nitrogen and phosphorus nutrient management. J. Agric. Stud. 4:127. doi: 10.5296/jas.v4i1.8908
Knutson, C. L., Haigh, T., Hayes, M. J., Widhalm, M., Nothwehr, J., Kleinschmidt, M., et al. (2011). Farmer perceptions of sustainable agriculture practices and drought risk reduction in Nebraska, USA. Renew. Agric. Food Syst. 26, 255–266. doi: 10.1017/s174217051100010x
Kocsis, T., Kotroczó, Z., Juhos, K., Ferschl, B., Rozmann, V., Brückner, A., et al. (2022). Opposite tendency between yield and taste of organic tomato by increasing biochar doses in a slightly humous arenosol. Agron. Res. 20, 200–214. doi: 10.15159/AR.22.024
Kumar, A., and Verma, J. P. (2018). Does plant-microbe interaction confer stress tolerance in plants: a review? Microbiol. Res. 207, 41–52. doi: 10.1016/j.micres.2017.11.004
Larkin, R. P. (2016). Impacts of biocontrol products on Rhizoctonia disease of potato and soil microbial communities, and their persistence in soil. Crop Prot. 90, 96–105. doi: 10.1016/j.cropro.2016.08.012
Lekberg, Y., and Koide, R. T. (2005). Is plant performance limited by abundance of arbuscular mycorrhizal fungi? A meta-analysis of studies published between 1988 and 2003. New Phytol. 168, 189–204. doi: 10.1111/j.1469-8137.2005.01490.x
Li, J., Wang, J., Liu, H., Macdonald, C. A., and Singh, B. K. (2022). Application of microbial inoculants significantly enhances crop productivity: a meta-analysis of studies from 2010 to 2020. J. Sustain. Agric. Environ. 1, 216–225. doi: 10.1002/sae2.12028
Lombardo, S., Abbate, C., Pandino, G., Parisi, B., Scavo, A., and Mauromicale, G. (2020). Productive and physiological response of organic potato grown under highly calcareous soils to fertilization and mycorrhization management. Agronomy 10:1200. doi: 10.3390/agronomy10081200
Lombardo, S., Scavo, A., Abbate, C., Pandino, G., Parisi, B., and Mauromicale, G. (2021). Mycorrhizal inoculation improves the mineral content of organic potatoes grown under calcareous soil. Agriculture 11:333. doi: 10.3390/agriculture11040333
Lori, M., Piton, G., Symanczik, S., Legay, N., Brussaard, L., Jaenicke, S., et al. (2020). (2020): compared to conventional, ecological intensive management promotes beneficial proteolytic soil microbial communities for agro-ecosystem functioning under climate change-induced rain regimes. Sci. Rep. 10:7296. doi: 10.1038/s41598-020-64279-8
Lori, M., Symnaczik, S., Mäder, P., Deyn, G. B. D., and Gattinger, A. (2017). Organic farming enhances soil microbial abundance and activity - a meta-analysis and meta-regression. PLoS One 12:e0180442. doi: 10.1371/journal.pone.0180442
Lugtenberg, B. J. J., de Weger, L. A., and Bennett, J. W. (1991). Microbial stimulation of plant growth and protection from disease. Curr. Opin. Biotechnol. 2, 457–464. doi: 10.1016/S0958-1669(05)80156-9
Lutz, S., Bodenhausen, N., Hess, J., Valzano-Held, A., Waelchli, J., Deslandes-Hérold, G., et al. (2023). Soil microbiome indicators can predict crop growth response to large-scale inoculation with arbuscular mycorrhizal fungi. Nat. Microbiol. 8, 2277–2289. doi: 10.1038/s41564-023-01520-w
Malgioglio, G., Rizzo, G. F., Nigro, S., Lefebvre du Prey, V., Herforth-Rahmé, J., Catara, V., et al. (2022). Plant-microbe interaction in sustainable agriculture: the factors that may influence the efficacy of PGPM application. Sustainability 14:2253. doi: 10.3390/su14042253
Malusà, E., Pinzari, F., and Canfora, L. (2016). “Efficacy of biofertilizers: challenges to improve crop production” in Microbial inoculants in sustainable agricultural productivity: Vol 2: Functional applications. eds. D. P. Singh, H. B. Singh, and R. Prabha (New Delhi: Springer), 17–40.
Mamun, A. A., Neumann, G., Moradtalab, N., Ahmed, A., Dupuis, B., Darbon, G., et al. (2024). Microbial consortia versus single-strain inoculants as drought stress protectants in potato affected by the form of N supply. Horticulturae 10:102. doi: 10.3390/horticulturae10010102
Martins, B. R., Siani, R., Treder, K., Michałowska, D., Radl, V., Pritsch, K., et al. (2023) Low soil moisture induces recruitment of actinobacteria in the rhizosphere of a drought-sensitive and rhizobiales in a drought-tolerant potato cultivar.
Mayer, J., Scheid, S., Widmer, F., Fließbach, A., and Oberholzer, H.-R. (2010). How effective are ‘effective microorganisms®(EM)’? Results from a field study in temperate climate. Appl. Soil Ecol. 46, 230–239. doi: 10.1016/j.apsoil.2010.08.007
MePAR (2013) MePAR - Agricultural Field Soil Identification System. Available online at: www.mepar.hu (Accessed April 21, 2024)
Miernik, A., and Jakubowski, T. (2021). Selected methods for starch content determination in plant materials. J. Phys. Conf. Ser. 1782:012019. doi: 10.1088/1742-6596/1782/1/012019
Nkebiwe, P. M., Stevens Lekfeldt, J. D., Symanczik, S., Thonar, C., Mäder, P., Bar-Tal, A., et al. (2024). Effectiveness of bio-effectors on maize, wheat and tomato performance and phosphorus acquisition from greenhouse to field scales in Europe and Israel: a meta-analysis. Front. Plant Sci. 15:1333249. doi: 10.3389/fpls.2024.1333249
Overbeek, W., Jeanne, T., Hogue, R., and Smith, D. L. (2021). Effects of microbial consortia, applied as fertilizer coating, on soil and rhizosphere microbial communities and potato yield. Front. Agron. 3:700. doi: 10.3389/fagro.2021.714700
Papp, O., Bencze, S. Z., Barta, L., and Drexler, D.. (2021b) Testing microbial inoculants and precrop effect on organic potato in Hungary. Organic World Congress 2021, Science Forum: 6th ISOFAR Conference co-organised with INRA, FiBL, Agroecology Europe, TP Organics and ITAB, Rennes, France, 8 - 10 September, 2021.
Papp, O., Kocsis, T., Biró, B., Jung, T., Ganszky, D., Abod, É., et al. (2021a). Co-inoculation of organic potato with fungi and bacteria at high disease severity of Rhizoctonia solani and Streptomyces spp. increases beneficial effects. Microorganisms 9:28. doi: 10.3390/microorganisms9102028
Peralta, J. M., and Stockle, C. O. (2002). Dynamics of nitrate leaching under irrigated potato rotation in Washington state: a long-term simulation study. Agric. Ecosyst. Environ. 88, 23–34. doi: 10.1016/S0167-8809(01)00157-8
Quazi, S. A. J., Ferdous, J., Shozib, H. B., Khaton, A., and Zaidi, N. W. (2024). Role of Trichoderma asperelloides and Trichoderma brevicompactum in improving drought tolerance in rice, plant stress. ISSN 12:100457. doi: 10.1016/j.stress.2024.100457
Raklami, A., Bechtaoui, N., Tahiri, A., Anli, M., Meddich, A., and Oufdou, K. (2019). Use of rhizobacteria and mycorrhizae consortium in the open field as a strategy for improving crop nutrition, productivity and soil fertility. Front. Microbiol. 10:1106. doi: 10.3389/fmicb.2019.01106
Rawal, R., Poelstra, J. W., Scheerens, J. C., and Benitez, M.-S. (2023). Inoculation of Trichoderma asperelloides NT33 causes minor transcriptional changes in tomato plants under water deficit stress conditions. CABI Int. Agri. agriRxiv20230014023. doi: 10.31220/agriRxiv.2023.00168
Rivett, D. W., Jones, M. L., Ramoneda, J., Mombrikotb, S. B., Ransome, E., and Bell, T. (2018). Elevated success of multispecies bacterial invasions impacts community composition during ecological succession. Ecol. Lett. 21, 516–524. doi: 10.1111/ele.12916
Rog, I., van der Heijden, M. G., Bender, S. F., Boussageon, R., Lambach, A., Schlaeppi, K., et al. (2025). Mycorrhizal inoculation success depends on soil health and crop productivity. FEMS Microbiol. Lett. 372:31. doi: 10.1093/femsle/fnaf031
Sagili, V. S., Chakrabarti, P., Jayanty, S. S., Kardile, H. B., and Sathuvalli, V. (2022). The glycemic index and human health with an emphasis on potatoes. Foods 11:2302. doi: 10.3390/foods11152302
Santos, M. S., Nogueira, M. A., and Hungria, M. (2019). Microbial inoculants: reviewing the past, discussing the present and previewing an outstanding future for the use of beneficial bacteria in agriculture. AMB Express 9:205. doi: 10.1186/s13568-019-0932-0
Schütz, L., Gattinger, A., Meier, M., Müller, A., Boller, T., Mäder, P., et al. (2018). Improving crop yield and nutrient use efficiency via biofertilization - a global meta-analysis. Front. Plant Sci. 8, 1–13. doi: 10.3389/fpls.2017.02204
Shaffique, S., Khan, M. A., Imran, M., Kang, S.-M., Park, Y.-S., Wani, S. H., et al. (2022). Research progress in the field of microbial mitigation of drought stress in plants. Front. Plant Sci. 13:870626. doi: 10.3389/fpls.2022.870626
Shrestha, B., Stringam, B. L., Darapuneni, M. K., Lombard, K. A., Sanogo, S., Higgins, C., et al. (2024). Effect of irrigation and nitrogen management on potato growth, yield, and water and nitrogen use efficiencies. Agronomy 14:560. doi: 10.3390/agronomy14030560
Singh, A., Shahid, M., Srivastava, M., Sonika, P., Sharma, A., and Kumar, V. (2013). Optimal physical parameters for growth of Trichoderma species at varying pH, temperature and agitation. Virol. Mycol. 3:127. doi: 10.4172/2161-0517.1000127
Tsror, L., Barak, R., and Sneh, B. (2001). Biological control of black scurf on potato under organic management. Crop Prot. 20, 145–150. doi: 10.1016/S0261-2194(00)00124-1
Verbruggen, E., van der Heijden, M. G., Rillig, M. C., and Kiers, E. T. (2013). Mycorrhizal fungal establishment in agricultural soils: factors determining inoculation success. New Phytol. 197, 1104–1109. doi: 10.1111/j.1469-8137.2012.04348.x
Zaki, H. E., and Radwan, K. S. (2022). Response of potato (Solanum tuberosum L.) cultivars to drought stress under in vitro and field conditions. Chem. Biol. Technol. Agric. 9, 1–19. doi: 10.1186/s40538-021-00266-z
Keywords: potato, organic farming, microbial inoculation, Pseudomonas, Trichoderma, Rhizophagus, Paraburkholderia, stomatal conductance
Citation: Papp O, Kocsis T, Ganszky D, Hunyadi ÉB, Mezőfi L, Labus B, Szigeti N, Selmeczi D, Tóth F, Darbon G and Drexler D (2025) Irrigation, variety, and year exert a greater influence on organic potato production than microbial inoculation: case study results of a 2-year on-farm experiment. Front. Sustain. Food Syst. 9:1641878. doi: 10.3389/fsufs.2025.1641878
Edited by:
Shaowu Hu, Yangzhou University, ChinaReviewed by:
Jiyi G. E. Zhang, Harpe Bioherbicide Solutions INC., United StatesBorbala Biro, Hungarian University of Agricultural and Life Sciences, Hungary
Copyright © 2025 Papp, Kocsis, Ganszky, Hunyadi, Mezőfi, Labus, Szigeti, Selmeczi, Tóth, Darbon and Drexler. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Orsolya Papp, b3Jzb2x5YS5wYXBwQGJpb2t1dGF0YXMuaHU=; Tamás Kocsis, a29jc2lzLnRhbWFzLmpvenNlZkB1bmktbWF0ZS5odQ==
†These authors have contributed equally to this work