 Berna Erdal1
Berna Erdal1 Bahar Yilmaz2
Bahar Yilmaz2 Esra Bozgeyik3
Esra Bozgeyik3 Seydi Yıkmış4*
Seydi Yıkmış4* Isam A. Mohamed Ahmed5
Isam A. Mohamed Ahmed5 Moneera O. Aljobair6*
Moneera O. Aljobair6* Monica Trif7
Monica Trif7- 1Department of Medical Microbiology, Tekirdağ Namık Kemal University Faculty of Medicine, Tekirdağ, Türkiye
- 2Department of Tumor Biology and Immunology, Institute of Health Sciences, Tekirdağ Namık Kemal Üniversitesi, Tekirdağ, Türkiye
- 3Department of Medical Biology, Adiyaman University Faculty of Medicine, Adıyaman, Türkiye
- 4Department of Food Technology, Tekirdağ Namık Kemal Üniversitesi, Tekirdağ, Türkiye
- 5Department of Food Science and Nutrition, College of Food and Agricultural Sciences, King Saud University, Riyadh, Saudi Arabia
- 6Department of Sports Health, College of Sports Sciences and Physical Activity, Princess Nourah Bint Abdulrahman University, Riyadh, Saudi Arabia
- 7Centre for Innovative Process Engineering (CENTIV) GmbH, Syke, Germany
Sustainability in food systems increasingly involves the integration of natural bioactive compounds that support human health while promoting eco-friendly production processes. Probiotic microorganisms, particularly those derived from food systems, have gained growing attention due to their dual role in health promotion and sustainable biotechnological applications. This study investigates both the anticancer and antifungal properties of live and thermally inactivated cell-free supernatants (CFSs) of Lactobacillus paracasei subsp. paracasei, a food-grade probiotic, thereby addressing a significant research gap in the dual-functional potential of probiotic metabolites. Human glioma (U-87) and non-cancerous kidney epithelial (HEK293T) cells were treated with live and inactivated CFSs. A dose-dependent cytotoxic effect was observed, with IC₅₀ values indicating greater selective efficacy of the inactivated CFSs against glioma cells, while sparing normal cells. Flow cytometry and fluorescence microscopy revealed that both CFSs induced apoptosis and cell cycle arrest at the sub-G1 phase, findings further supported by DNA fragmentation analysis. In parallel, antifungal activity against Candida albicans and Candida parapsilosis was evaluated using the broth microdilution method. Notably, the inactivated CFSs exhibited potent fungistatic and fungicidal effects at lower concentrations than the live form, highlighting a novel, stable postbiotic alternative with therapeutic relevance. The present study is the first study to simultaneously demonstrate selective anticancer activity against U-87 glioma cells and broad-spectrum antifungal potential of inactivated L. paracasei CFSs. These findings represent a promising strategy for integrating probiotic-derived bioactives into sustainable food systems and functional food development.
Introduction
Brain and other central nervous system (CNS) tumors, both malignant and non-malignant, represent a diverse group of diseases with distinct histopathological subtypes, descriptive epidemiology, clinical features, and treatment strategies (Miller et al., 2021). Although primary brain tumors are less common than other cancer types, their mortality rates are high, and the 5-year survival rate remains relatively low (Miller et al., 2021; Siegel et al., 2022). Gliomas are the most prevalent form of primary malignant brain tumors in adults, with glioblastoma multiforme (GBM) representing the most aggressive and lethal subtype. Despite advances in neuroimaging, surgical resection techniques, radiotherapy, and chemotherapeutic regimens such as temozolomide, the overall prognosis for patients remains dismal, with a median survival time of approximately 12–15 months and a 5-year survival rate of less than 10%. This is largely attributed to the highly invasive nature of glioma cells, their resistance to conventional therapies, and the presence of the blood–brain barrier, which limits drug delivery to the tumor site. In recent years, immunotherapy has emerged as a promising frontier in glioma treatment, aiming to overcome the limitations of traditional approaches. Strategies such as immune checkpoint inhibitors, cancer vaccines, chimeric antigen receptor (CAR) T-cell therapy, and oncolytic viruses are being actively investigated for their potential to stimulate robust, tumor-specific immune responses. Although clinical results in glioma remain modest compared to other cancer types, ongoing research continues to explore the optimization of these modalities, often in combination with standard therapies, to improve therapeutic outcomes. The integration of immunotherapy into glioma treatment reflects a broader shift toward personalized and immune-based oncology approaches (de Robles et al., 2015). Due to the limited success of current treatments, there is growing interest in complementary strategies that harness the body’s own biological systems—such as the gut-brain axis and microbiota-mediated immune modulation—to support glioma therapy. In this context, food-derived probiotics like Lactobacillus paracasei have emerged as promising candidates due to their potential to influence tumor progression and enhance host immune responses.
Moreover, there has been a growing recognition of the intricate interplay between the gut microbiota and cancer progression in numerous studies, and recent research suggests that polymorphic microbiomes may represent a novel hallmark of cancer (Hanahan, 2022). Probiotics are beneficial microorganisms that influence the microbial flora and help maintain host health. They have been shown to possess potential in the prevention of cancer and neurodegenerative diseases. Various strains of Lactobacillus, in particular, have demonstrated anti-tumorigenic properties through multiple mechanisms, including modulation of oxidative stress, apoptosis induction, and immune system enhancement, the activation of multiple pathways, including the secretion of antioxidant enzymes, stimulation of apoptosis, and regulation of the cell cycle (Mohseni et al., 2021). Accumulating evidence also suggests that lactobacilli possess notable anticancer activity against gliomas (D’Alessandro et al., 2021; Fatahi et al., 2021; Li et al., 2021).
Lactobacillus paracasei subsp. paracasei (L. paracasei) is a gram-positive, homofermentative lactic acid bacterium widely utilized in fermented dairy products and probiotic cultures. Previous studies have shown that the total secreted metabolites of L. paracasei significantly reduce the viability of HT-29 and Caco-2 colorectal cancer cells (Wang et al., 2023). Additionally, this probiotic bacterium has been shown to reduce tumor volume in a rat colorectal cancer model induced by dimethylhydrazine, alone or in combination with 5-FU, while promoting apoptosis (Jam et al., 2021). These findings highlight the significant potential of food-derived probiotics as sustainable and biologically compatible agents in cancer management.
Opportunistic fungal pathogens such as Candida albicans (C. albicans) are among the essential causes of life-threatening infections, especially in immunosuppressed individuals (Fidan et al., 2014). The antifungal activities of L. paracasei against Candida species are remarkable. Probiotic-derived metabolites and postbiotic compounds from L. paracasei have demonstrated significant antifungal effects, including inhibition of biofilm formation, suppression of hyphal development, and disruption of fungal cell wall integrity through medium acidification (Elbaz et al., 2022; Rossoni et al., 2020; Spaggiari et al., 2022). A study has shown that L. paracasei live cells and postbiotic components exhibit antifungal activity by reducing cell viability and biofilm formation of Candida auris (C. auris) resistant to antifungal agents (Rossoni et al., 2020). It has also been reported that L. paracasei postbiotics exhibit antifungal activity by disrupting C. albicans cell wall integrity by reducing the pH of the medium (Divyashree et al., 2023; García-Gamboa et al., 2024). These in vivo and in vitro studies have reinforced the potential of L. paracasei to be an essential therapeutic aid in treating glioma and fungal infections. Accordingly, this study aims to determine the anticancer effects of L. paracasei in U-87 glioma cells in vitro, alongside its antifungal activity against clinically relevant Candida spp. Such properties highlight the multifunctional potential of L. paracasei as a biotherapeutic agent with relevance to both oncology and infectious disease contexts. As the field of sustainable food systems increasingly explores the role of microbiome-based interventions in health promotion and disease mitigation, the integration of such probiotic strategies into therapeutic frameworks becomes particularly compelling. These findings may contribute to a broader understanding of how sustainable, food-derived microbial solutions can support human health in complex disease landscapes.
Materials and methods
Probiotic bacteria/probiotic bacterial isolates
The probiotic bacterium L. paracasei used in this study was first inoculated into DeMan, Rogosa, and Sharpe (MRS) broth (Cat. No: 4017282 Biolife, Milano, Italy), followed by incubation at 37°C for 24–48 h. Subsequently, to obtain a single colony from the activated bacterial culture, inoculation was performed on MRS agar (Cat. No: 401722, Biolife, Milano, Italy), followed by incubation at 37°C for 24–48 h. After incubation, the purified cultures of the activated L. paracasei isolate were examined.
Inactivation of probiotic Bacteria
A single colony obtained from the purified L. paracasei isolate was inoculated into MRS broth and incubated at 37°C for 24–48 h. Following the observation of growth, the culture was subjected to inactivation in a water bath at 100°C for 30 min. Subsequently, the cultures were centrifuged at 4500 rpm for 20 min to obtain the CFSs from the live and inactivated bacteria. The CFSs obtained after centrifugation were sterilized using 0.22 μm PES membrane filters (Cat. No: 1.094.07.006.050, Isolab, Eschau, Germany). In addition, the pellet was resuspended in MRS broth medium to control for heat inactivation. Subsequently, the resuspended culture was re-inoculated onto MRS agar to check any bacterial growth (Karimi Ardestani et al., 2019; Sadeghi-Aliabadi et al., 2014).
Cell culture
Glioblastoma (U-87 MG, ATCC® HTB-14™) and non-cancerous human embryonic kidney (HEK-293, ATCC® CRL-1573™) cell lines were used in this study. The cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Cat. No: ECM0103L, Euroclone, Pero (MI), Italy) supplemented with 10% fetal bovine serum (FBS, Cat. No: P30-3306, Pan Biotech, Aidenbach, Germany), 2 mM L-glutamine (Cat. No: 25030–024, Gibco, Paisley, UK), and 1% penicillin/streptomycin (Cat. No: 15140–148, Gibco, New York, United States) at 37°C in a 5% CO2 incubator.
MTT cell viability assay
When cell confluency reached approximately 80–90%, the cells were detached using 0.05% trypsin (Cat. No: 25200056, ThermoFisher, Paisley, UK) and centrifuged at 1250 rpm for 3 min. After centrifugation, the supernatant was carefully removed. The pellet was then resuspended, and 100 μL of the cell suspension containing 1 × 104 cells was added to each 96-well flat-bottom cell culture plate. For the positive and negative controls, fresh medium was added, while various percentage concentrations (0.39, 0.78, 1.56, 3.125, 6.25, 12.5, 25, 50, and 100) of live and inactivated CFSs, prepared through serial dilution, were applied to the other wells. The plates were then incubated for 24 h. Following treatments, supernatants were discarded, and cells were incubated with 5 mg/mL MTT solution (Cat. No: 475989-1GM Merck Darmstadt, Germany) for 4 h. After MTT incubations, the MTT solution was removed, and the plates were air dried. Then, dimethyl sulfoxide was used to dissolve MTT formazan particles, and plates were read at 570 nm wavelength using a microplate reader (BioTek-800-TS absorbance reader, Agilent, Santa Clara, United States). The percentage of cell viability was calculated as previously described in Karimi Ardestani et al., 2019. All experiments were performed in triplicate.
Flow cytometric analysis of the cell cycle
Flow cytometric analysis of cell cycle was performed as mentioned earlier (Chen et al., 2023b). Briefly, U-87 and HEK293T cells were seeded in 6-well flat-bottom cell culture plates at 2.5 × 105 cells/well. The cells were incubated at 37°C with 5% CO₂ for 24 h to allow for attachment. Fresh medium was added to the positive and negative controls. At the same time, the IC50 concentrations of live and inactivated CFSs, prepared through serial dilution, were applied to the other wells and left for incubation for 24 h. Following incubation, excess medium was removed, and the wells were washed with Dulbecco’s phosphate-buffered saline (DPBS) (Cat. No: ECM0103L, Euroclone, Pero (MI), Italy). The cells were detached and resuspended, and 5 mL ice-cold 70% ethanol was added while vortexing, then incubated overnight at +4°C. After incubation, the cells were centrifuged at 1200 rpm for 5 min, and the supernatant was discarded. The cells were then rewashed with DPBS. Subsequently, 1 mL of DPBS and 5 μL of 20% Triton X detergent were added to the samples and gently mixed by shaking for 3 min, and then, the cells were centrifuged at 1700 rpm for 5 min, and the supernatant was discarded. A mixture of 500 μL of DPBS and 3.3 μL of RNAse (Cat. No: 1263MG050, Neofroxx, Einhausen, Germany) was added to the pellet, and the samples were incubated at 37°C for 30 min. After incubation, 10 μL of 10 mg/mL propidium iodide (PI) (Cat. No. P4170-10MG, Sigma, Saint Louis, United States) was added to each sample, and the samples were incubated in the dark for 1 h. Sub G refers to the Sub G phase, which is associated with apoptotic cells; G0/G1 indicates the resting (G0) and growth (G1) phases of the cell cycle; S represents the DNA synthesis phase; and G2/M refers to the G2 phase, where cells prepare for mitosis, and the M phase, during which mitosis occurs. Finally, the stained cells were transferred to flow cytometry tubes, and cell cycle distribution (Sub G, G0/G1, S, and G2/M phases) was analyzed using the DxFLEX B4-R2-V3 Flow Cytometer Autoloader. Data acquisition and analysis were performed using CytExpert DxFLEX software. All experiments were performed in triplicate.
Fluorescent imaging
A dual staining method using acridine orange (AO) and propidium iodide (PI) was employed to assess the viability and morphology of U-87 and HEK293T cells treated with live and heat-inactivated CFSs of L. paracasei isolates (Seber et al., 2020). Briefly, cells were seeded in 96-well plates at a concentration of 1 × 104 cells/well. Fresh culture medium was added to the positive and negative controls, and IC50 concentration of the live and inactive CFSs, prepared through serial dilution, was applied to the other wells, which were then incubated for 24 h. Cells were incubated with propidium iodine and acridine orange solution and washed with DPBS to remove excess dye. Cells were visualized under fluorescent microscope and images captured through Cytovision Capture Station (version 7.5; Genetix; Leica Microsystems).
DNA fragmentation analysis
DNA fragmentation analysis was previously described (Öğüt et al., 2025). For DNA fragmentation analysis, U-87 and HEK293T cells were seeded into six-well culture plates at a density of 2.5 × 105 cells per well and incubated overnight at 37°C to allow for attachment. Then, cells were incubated with IC50 concentration of the live and inactive CFSs prepared through serial dilution. The positive and negative controls were incubated with fresh culture medium. Following 24 h of incubation, cells were rinsed with DPBS and collected with the help of a scraper. Then, the collected cells were transferred into Eppendorf tubes and centrifuged at 12000 rpm for 1 min. Supernatants were then discarded, and the pellet was washed with DPBS. Cell pellets were later resuspended in 30 μL nuclease-free water and gently mixed. 200 μL InstaGene Matrix (Cat. No: 7326030, Bio-Rad, United States) was then homogenized with the cell suspension. The mixture was incubated at 56°C for 15 to 30 min. Subsequently, the suspension was vortexed at high speed for 10 s and then incubated in a water bath at 100°C for 8 min. The suspension was then centrifuged at 12,000 rpm for 2–3 min. The concentration and purity of the isolated DNA samples were measured using a Nano Drop spectrophotometer (Nano-100, Allsheng, Hangzhou, China). DNA samples were run on a 1.0% agarose gel (Cat. No: 35–1,010 Peqlab, Erlangen, Germany) supplemented with 1 μg/mL ethidium bromide (Thermo Scientific, Germany) and visualized using ultraviolet transillumination (Bio-Rad, United States).
Antifungal susceptibility assay
n this study, the antifungal susceptibility of yeast strains was evaluated using the broth microdilution method in accordance with the guidelines of the Clinical and Laboratory Standard strains C.albicans ATCC 10231 and Candida parapsilosis (C. parapsilosis) ATCC 22019 were used in the assay. Serial dilutions of both live and heat-inactivated CFSs were prepared, with final concentrations ranging from 0.39 to 100%. A yeast suspension was standardized to 1.5 × 103 CFU/mL, and 10 μL of this suspension was inoculated into microplate wells containing different CFS sample concentrations. Each assay included positive and negative control groups. The microplates were incubated at 35°C for 24 h, after which microbial growth or inhibition was assessed visually. The minimum inhibitory concentration (MIC) and minimum fungicidal concentration (MFC) values were determined for each fungal strain. To ascertain the MFC, 10 μL aliquots from wells at MIC, and two- and three-fold MIC concentrations were subcultured onto Sabouraud dextrose agar (SDA) plates. The plates were incubated at 35°C for 48 h, and the absence of fungal colonies was interpreted as the MFC value. To benchmark the antifungal activity of the supernatants, fluconazole (FLC; 128 μg/mL) and voriconazole (VOR; 8 μg/mL) were used as standard reference agents. All experiments were performed in triplicate.
Statistical analysis
The data were analyzed using GraphPad Prism 9. All experiments were performed in triplicate, and the results are presented as the mean ± standard deviation (SD). Two-way ANOVA, followed by Tukey’s multiple comparison test was conducted to determine statistically significant differences between the control and treatment groups. A p-value of <0.05 was considered statistically significant.
Results
Cytotoxic effect of live and inactivated CFSs of Lactobacillus paracasei subsp. paracasei isolate on U-87 and HEK293T cells
To investigate the cytotoxic potential of L. paracasei, both live and heat-inactivated CFSs were applied to U-87 human glioblastoma cells and HEK293T human embryonic kidney epithelial cells across a concentration gradient ranging from 0.39 to 100%. Cell viability was assessed using the MTT assay after 24 h of treatment. The results demonstrated a clear dose-dependent decrease in cell viability in U-87 cells for both forms of the supernatant. Notably, the inactivated supernatant exhibited stronger cytotoxic activity than the live supernatant. The calculated half-maximal inhibitory concentration (IC₅₀) values in U-87 cells were 15.73% ± 0.50 for the live supernatant and 10.40% ± 0.38 for the inactivated supernatant (Figure 1). A similar trend was observed in HEK293T cells, although the overall cytotoxicity was lower than in the glioma cells. The IC₅₀ values in HEK293T cells were 21.71% ± 0.45 and 13.81% ± 0.30 for the live and inactivated supernatants, respectively. These findings indicate that the inactivated form of L. paracasei supernatant (Figure 1) was more effective in reducing cell viability in both cancerous and non-cancerous cell lines. However, the more pronounced cytotoxic effects in U-87 glioma cells compared to normal HEK293T cells suggest a degree of selective anticancer activity. Therefore, there is the need for further studies to identify the specific bioactive compounds present in the inactivated supernatant and to elucidate how they contribute to the observed cytotoxic effects.
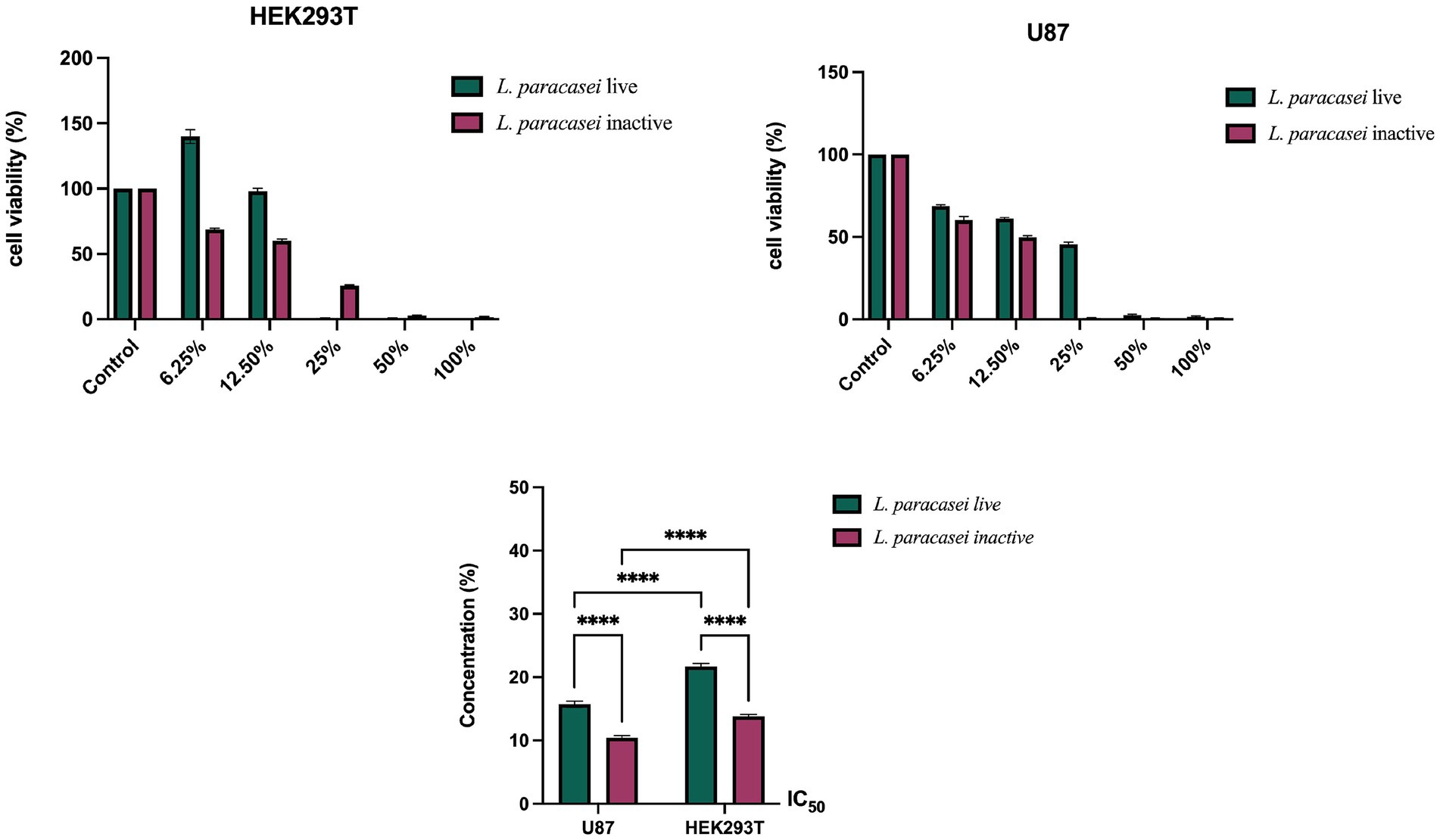
Figure 1. Cell viability percentages and calculated IC50 values for HEK293T and U-87 cells after 24-h treatment with live and inactivated L. paracasei CFSs.
Analysis of apoptosis and morphological changes
Cell cycle analyses were conducted using flow cytometry on U-87 and HEK293T cells treated for 24 h with IC50 concentrations of live and inactive L. paracasei isolate supernatants. The percentage distribution of cells in the Sub G, G0/G1, S, and G2/M phases of the cell cycle was assessed (Figure 2). In U-87 and HEK293T cells treated with live and inactive supernatants of the L. paracasei isolate, the cell population in the G0/G1 and S phases decreased compared to the control group. In contrast, the population in the Sub G and G2/M phases increased relative to the control (Figure 2). These findings indicate that both live and inactive forms of the L. paracasei isolate were found to induce cell death by arresting cells in the Sub-G phase.
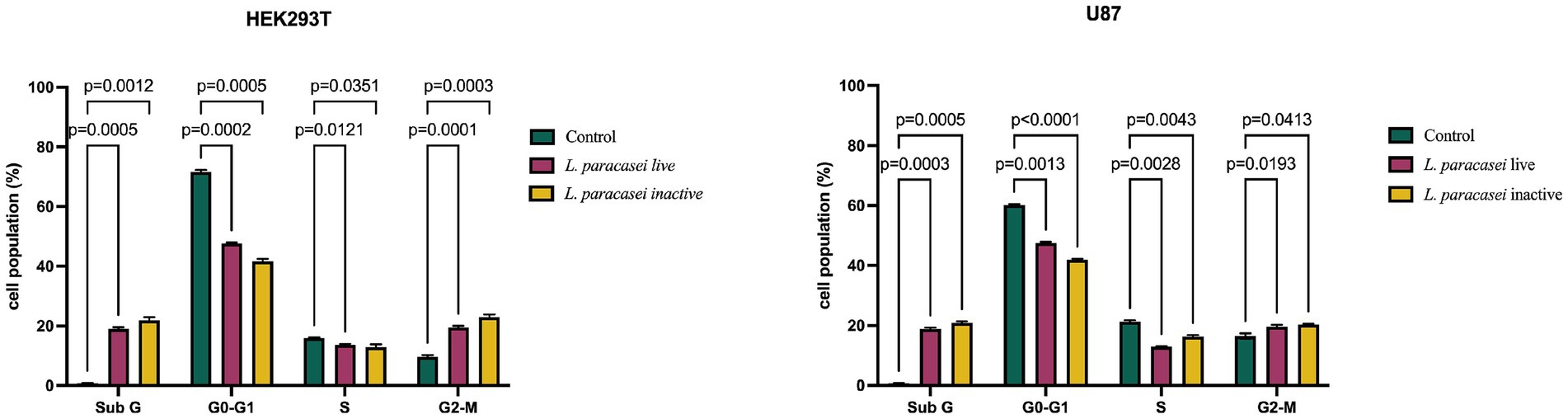
Figure 2. Cell cycle distribution of HEK293T and U-87 cells treated with IC50 concentrations of live and inactive supernatants from L. paracasei, showing changes in the Sub G, G0/G1, S, and G2/M phases.
The morphological changes in U-87 and HEK293T cells treated for 24 h with the IC50 concentrations of live and inactive L. paracasei CFSs were examined using a fluorescence microscope. Nuclear changes such as chromatin condensation and shrinkage, along with cytological features like necrotic and apoptotic cell death, were assessed. AO/PI dual staining was used to distinguish viable and apoptotic cells: green, fluorescent cells indicate viable cells stained with AO (white arrows), whereas orange/red fluorescent cells represent apoptotic cells stained with PI (yellow arrows). A higher number of apoptotic cells was observed in both U-87 and HEK293T cultures treated with live and inactive supernatants compared to untreated controls (Figure 3A).
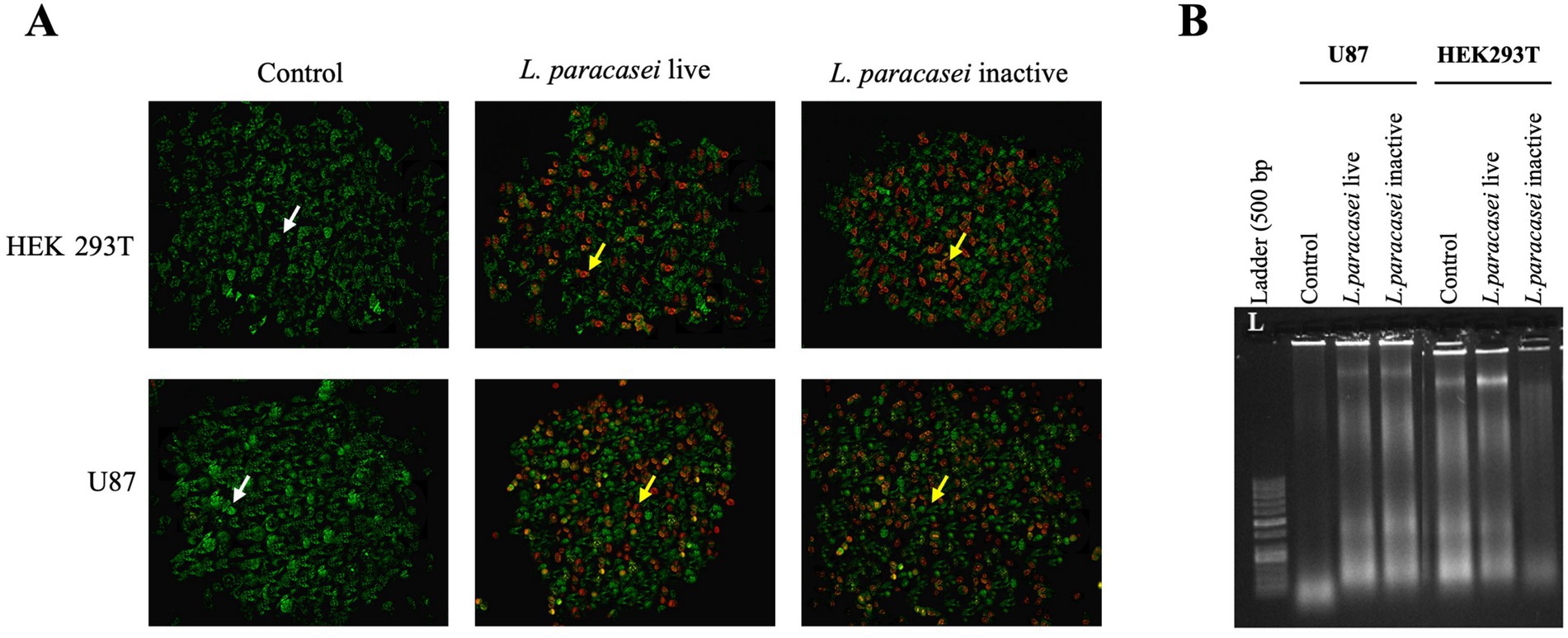
Figure 3. (A) Fluorescence microscope image (20X) showing U-87 and HEK293T cells treated with IC50 concentrations of live and inactive CFSs of L. paracasei. White arrow indicates live cells stained with acridine orange (green fluorescence, AO), and yellow arrow points to apoptotic cells stained with propidium iodide (orange/red fluorescencei PI). (B) Effects of DNA fragmentation in U-87 and HEK293T cells following treatment with IC50 concentrations of live and inactive CFSs from the L. paracasei isolate.
Furthermore, the high population of apoptotic cells following treatment with live and inactive supernatants from U-87 and HEK293T cells supports the MTT findings.
To validate and complement the findings of the AO/PI (Acridine Orange/Propidium Iodide) dual staining assay, a DNA fragmentation assay was performed to assess the induction of apoptosis at the molecular level in both U-87 and HEK293T cells. The assay was conducted after 24-h exposure of the cells to the IC₅₀ concentrations of both live and inactivated CFSs. DNA fragmentation is a hallmark of late-stage apoptosis and serves as a reliable indicator of programmed cell death, and represents the confirmation of evidence for cytotoxic effects observed in viability and morphological assays. The results of the DNA fragmentation assay revealed clear patterns of nucleosomal DNA laddering in both U-87 glioblastoma cells and non-tumorigenic HEK293T cells treated with either form of the supernatant. The presence of fragmented DNA bands in treated samples, compared to the intact genomic DNA in untreated control cells, indicates that apoptosis was actively induced following exposure to the bacterial supernatants (Figure 3B). Notably, the fragmentation pattern was more pronounced in U-87 cells, and this suggests a higher susceptibility of the cancer cell line to apoptosis triggered by the supernatant treatments. These findings reinforce the results obtained from the AO/PI staining, which already suggested apoptotic cell death based on nuclear condensation and membrane integrity changes. The convergence of results from both assays provides robust evidence that the cytotoxic effect of L. paracasei supernatants operates, at least in part, through the induction of apoptotic mechanisms. Moreover, the fact that both live and inactivated supernatants elicited comparable DNA fragmentation supports the hypothesis that bioactive compounds responsible for apoptosis may be heat-stable and retained after inactivation. These results strengthen that the L. paracasei isolate supernatants—particularly in their inactivated form—hold promise as sources of apoptosis-inducing agents with potential applications in cancer therapy research.
Fungistatic and fungicidal effects of live and inactivated supernatants of Lactobacillus paracasei subsp. paracasei isolate
In this study, the antifungal properties of both live and heat-inactivated CFSs derived from L. paracasei were assessed against two clinically relevant fungal pathogens: C. albicans and C. parapsilosis (Table 1; Figure 4). The broth microdilution method was employed to determine the minimum inhibitory concentration (MIC) and minimum fungicidal concentration (MFC) values, which are essential parameters in evaluating antifungal efficacy. Fluconazole (FLC; 128 μg/mL) and voriconazole (VOR; 8 μg/mL) were included as standard reference antifungal agents to benchmark the activity of the supernatants. The results revealed distinct antifungal profiles for the live and inactivated forms of the L. paracasei supernatant. For C. albicans, the MIC and MFC of the live supernatant were both 50%, indicating that only relatively high concentrations exerted inhibitory and fungicidal effects. In contrast, the inactivated supernatant displayed potent antifungal activity at much lower concentrations, with MIC and MFC values of just 12.5%. This suggests that the inactivated supernatant retains or possibly enhances its antifungal components after heat treatment, pointing to the presence of thermostable bioactive metabolites. A similar trend was observed for C. parapsilosis. The live supernatant exhibited antifungal effects only at the highest tested concentration of 100%, highlighting its comparatively weaker activity against this strain. On the other hand, the inactivated supernatant showed fungistatic and fungicidal activity at a concentration of 25%, underscoring its significantly higher potency. These findings suggest that the process of inactivation may concentrate or stabilize antifungal components, enhancing their effectiveness against fungal pathogens. When comparing the MIC and MFC values between live and inactivated supernatants across both fungal strains, the inactivated form consistently demonstrated greater antifungal activity. The superior efficacy of the inactivated supernatant against C. albicans, particularly at a low concentration of 12.5%, is of special interest. This highlights the potential of postbiotic compounds—metabolites derived from non-viable probiotics—as a promising alternative or adjunct to traditional antifungal therapies. Compared to standard antifungal drugs, fluconazole exhibited an MIC of 4 μg/mL against C. albicans, while voriconazole demonstrated a potent MIC of 0.0625 μg/mL against C. parapsilosis. Although the MIC values for the inactivated probiotic supernatants are higher than those of conventional antifungals, the natural origin, low toxicity, and potential synergistic use with existing treatments make them attractive candidates for further development. Overall, the data support that L. paracasei supernatants, particularly in their inactivated form, possess notable antifungal properties. These effects may be attributed to secreted bioactive molecules such as organic acids, peptides, or other antimicrobial compounds. Demonstrating significant activity against common and clinically important Candida species, these findings lay the groundwork for exploring probiotic-derived or postbiotic products as natural antifungal agents.
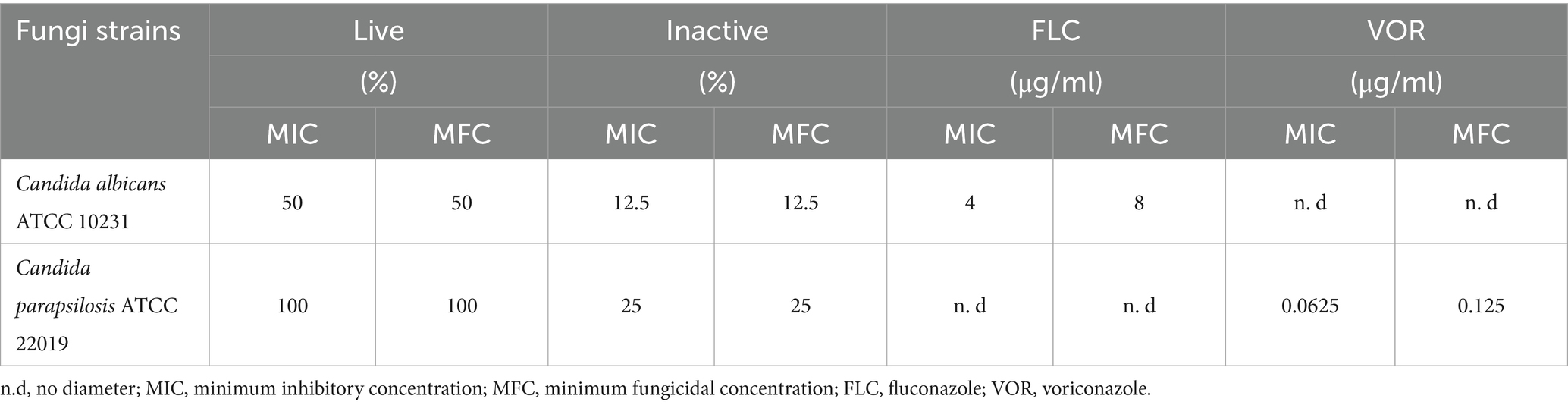
Table 1. Antifungal activity of live and inactive L. paracasei CFSs against Candida spp.

Figure 4. MFC petri dish images of live and inactive L. paracasei CFSs. (A) Candida albicans. (B) Candida parapsilosis; NC: negative control; PC: positive control.
Discussion
Extensive research has demonstrated that probiotics possess protective and preventive effects against various forms of cancer. The gut microbiota plays a crucial role in modulating cancer risk, with well-established impacts on gastrointestinal malignancies such as colon cancer. More recently, emerging studies have begun to unravel the intricate connections between the gut microbiome and neurological disorders, including brain tumors like glioma, highlighting a promising and rapidly evolving area of investigation. The ability of probiotics to modulate and restore a healthy gut microbial balance underscores their potential as adjunctive agents in cancer prevention and therapy. Preclinical studies involving both murine models and human cell lines have further explored the therapeutic potential of probiotics, providing encouraging evidence for their role in regulating tumor progression and enhancing host immune responses.
Research has emphasized the anti-carcinogenic effects of Lactobacillus species, particularly about colon cancer. For example, the probiotic bacterium Lactobacillus rhamnosus has been shown to exert cytotoxic and anticarcinogenic effects by inhibiting cell proliferation and inducing apoptosis in HT-29 cells (Dehghani et al., 2021). On the other hand, Khosrovan et al. (2020) demonstrated in their study that the probiotics, Lactobacillus acidophilus and Bifidobacterium bifidum, exhibit antitumor effects in colon and breast cancer cells, enhancing immune surveillance by stimulating inflammatory cytokines such as INF-γ and TNF. Propionibacterium freudenreichii has been shown to inhibit cell proliferation in the gastric cancer cell line HGT-1 in a time- and dose-dependent manner (Cousin et al., 2012). In a study conducted by Zhang et al. (2022) a new commensal L. paracasei strain, SH2020, was identified, and it was shown to prevent tumor growth and stimulate anti-tumor immunity in mice. Moreover, the combination of L. paracasei SH2020 with anti-PD-1 enhanced tumor suppression significantly, yielding better results compared to either anti-PD-1 or L. paracasei SH2020 monotherapy (Zhang et al., 2022). Moreover, while research has primarily focused on gastrointestinal cancers, studies have also demonstrated that probiotics exert significant cytotoxic effects in brain tumors such as glioma. In a study conducted by Wang et al. (2023) using a glioma mouse model, it was demonstrated that Bifidobacterium lactis (B. lactis) and Lactobacillus plantarum (L. plantarum) can regulate the composition of the gut microbiota and inhibit tumor growth via the PI3K/AKT pathway. Additionally, it has been shown that B. lactis and L. plantarum possess potential protective effects against glioma (Wang et al., 2022). Furthermore, it has been demonstrated that Lactobacillus casei mediates the regulation of oxidative stress and the p53 pathway, suppressing the growth of U-87 glioma cells (Chen et al., 2023a). Fan et al. (2022) have shown a relationship between the gut microbiome and glioma. Their study on mice showed that a high abundance of Bacteroidia in the gut microbiota accelerates glioma progression, while a low abundance reduces it; conversely, Firmicutes had the opposite effect (Fan et al., 2022). Our study demonstrated that both live and inactive forms of the L. paracasei isolate suppress cell viability in U-87 glioma cells, halt cell cycle progression by retaining cells in the sub-G1 phase and induce apoptosis. Additionally, it was found that the live and inactive forms of the L. paracasei isolate had effects at lower doses in U-87 cells compared to HEK293T normal kidney epithelial cells. Moreover, it has been demonstrated that the inactive form of L. paracasei is more effective than the live form in both cell viability and cell cycle assays. These findings suggest that the inactive form of L. paracasei exhibits greater anti-tumor properties in U-87 glioma cells than in normal cells. However, further detailed studies are needed to confirm these findings.
Probiotic bacteria, especially Lactobacillus species, have gained attention recently with their antimicrobial activities against a wide range of pathogenic microorganisms. In particular, investigating the antifungal potential of CFSs of these bacteria is vital for alternative treatment approaches. Spaggiari et al. (2022) reported that CFSs belonging to Lactobacillus acidophilus, Lactobacillus plantarum, Lactobacillus rhamnosus and Lactobacillus reuteri suppressed the adhesion of C. parapsilosis to vaginal epithelial cells and its capacity to cause infections. In experimental analyses using both single-layer and transwell coculture systems, it was shown that these CFSs significantly reduced the pathogenicity of C. parapsilosis. The study results revealed that probiotic-derived metabolites provide direct antifungal effects and play a role in strengthening host defense mechanisms by improving epithelial resistance to fungi. Similarly, Rossoni et al. (2020) reported that postbiotic products belonging to L. paracasei 28.4 isolate were effective on Candida auris resistant to antifungal drugs and had fungistatic and fungicidal properties. The study emphasizes that postbiotics may offer a new and effective treatment approach against resistant fungal species. On the other hand, another study conducted by Rossoni et al. (2020) revealed that various clinical Lactobacillus strains inhibited the biofilm formation capacity of C. albicans. Biofilm structures play a critical role in developing resistance to antifungal treatments. Therefore, these findings indicate that probiotics may be effective on planktonic cells and biofilm-forming C. albicans species. This study determined that live and inactivated supernatants of L. paracasei isolates showed antifungal effects on both C. albicans and C. parapsilosis strains. In particular, the inactivated supernatant was fungicidal and fungistatic at low concentrations against C. albicans and C. parapsilosis strains, suggesting that inactivated supernatants may have higher antifungal potential than live supernatants.
While this study presents compelling evidence of the anticancer and antifungal potential of L. paracasei, it opens certain areas that offer exciting opportunities for further exploration. The current investigation focused on the U-87 glioma cell line, and expanding this work to include a broader range of glioma cell models with diverse genetic backgrounds would enrich the understanding of L. paracasei’s therapeutic versatility. Additionally, extending these findings through in vivo studies will be instrumental in validating the probiotic’s efficacy and safety within complex biological systems, thereby advancing its translational potential toward clinical applications. Importantly, these study findings demonstrate a strong foundation for using L. paracasei as a supportive agent in glioma treatment, encouraging the integration of such probiotic-based interventions alongside conventional therapies to enhance treatment outcomes potentially. Regarding the antifungal activity, the study successfully highlights the promising effects of L. paracasei supernatants against common Candida strains. Building upon this study’s promising findings, future research should focus on exploring the anticancer effects of L. paracasei across multiple glioma cell lines and in vivo models, while also investigating the broad-spectrum applicability and clinical relevance of probiotic-derived metabolites in combating fungal pathogens. Such studies will help elucidate the mechanisms of action and pave the way for innovative, sustainable therapies. Additionally, investigating the antifungal efficacy of L. paracasei supernatants against clinical Candida isolates will further provide critical insights into their clinical relevance and support the development of probiotic-based antifungal therapies. Such studies will be essential to translate these initial results into practical applications, ultimately advancing the role of probiotics in sustainable and integrative treatment approaches for cancer and fungal infections.
Conclusion
This study demonstrates that probiotic-derived bioactive compounds can be a sustainable health approach in food systems by revealing both anticancer and antifungal effects of L. paracasei CFSs. The findings show that L. paracasei supernatants suppress the growth of cancer cells by inducing apoptosis and causing cell cycle arrest in U-87 glioma cells; and also have antifungal potential by showing fungistatic and fungicidal effects on C. albicans and C. parapsilosis. These results show that probiotic microorganisms and their metabolites support human health and can provide sustainable health solutions through functional foods. Integrating probiotics into food systems has great potential in reducing food waste, promoting the use of natural biological agents, and developing preventive strategies against diseases. However, this study was carried out under in vitro conditions. The findings obtained need to be supported by in vivo studies, and their applicability in food matrices should be investigated. In further studies, the potential of L. paracasei supernatants to be integrated into sustainable food systems can be evaluated more comprehensively by addressing aspects such as formulation with different food products, stability, sensory properties, and consumer acceptance. This study highlights the importance of probiotics in the development of functional foods and offers a new perspective on strengthening sustainable food systems by emphasizing their potential to replace synthetic additives and contribute to long-term health and environmental sustainability.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
BE: Writing – original draft, Conceptualization, Methodology, Writing – review & editing. BY: Writing – review & editing, Conceptualization, Writing – original draft. EB: Writing – review & editing, Writing – original draft, Data curation. SY: Data curation, Writing – review & editing, Writing – original draft. IAMA: Writing – review & editing, Writing – original draft. MOA: Writing – original draft, Funding acquisition, Writing – review & editing. MT: Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors are grateful for the support from Princess Nourah Bint Abdulrahman University Researchers Supporting Project Number (PNURSP2025R251), Princess Nourah Bint Abdulrahman University, Riyadh, Saudi Arabia.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Chen, Y., Miao, X., Xiang, Y., Kuai, L., Ding, X., Ma, T., et al. (2023b). Qinzhu Liangxue inhibits IL-6-induced hyperproliferation and inflammation in HaCaT cells by regulating METTL14/SOCS3/STAT3 axis. J. Ethnopharmacol. 317:116809. doi: 10.1016/j.jep.2023.116809
Chen, F., Teng, Z., Peng, X., Wu, H., Wan, W., and Long, H. (2023a). Lactobacillus casei enhances the apoptosis inducing effect of geniposide on U87 human glioma cells in vitro. Food Sci. Technol. 43:e112822. doi: 10.1590/fst.112822
Cousin, F. J., Jouan-Lanhouet, S., Dimanche-Boitrel, M.-T., Corcos, L., and Jan, G. (2012). Milk fermented by Propionibacterium freudenreichii induces apoptosis of HGT-1 human gastric cancer cells. PLoS One 7:e31892. doi: 10.1371/journal.pone.0031892
D’alessandro, G., Lauro, C., Quaglio, D., Ghirga, F., Botta, B., Trettel, F., et al. (2021). Neuro-signals from gut microbiota: perspectives for brain glioma. Cancer 13:2810. doi: 10.3390/cancers13112810
De Robles, P., Fiest, K. M., Frolkis, A. D., Pringsheim, T., Atta, C., St Germaine-Smith, C., et al. (2015). The worldwide incidence and prevalence of primary brain tumors: a systematic review and meta-analysis. Neuro-Oncology 17, 776–783. doi: 10.1093/neuonc/nou283
Dehghani, N., Tafvizi, F., and Jafari, P. (2021). Cell cycle arrest and anti-cancer potential of probiotic Lactobacillus rhamnosus against HT-29 cancer cells. Bioimpacts 11:245. doi: 10.34172/bi.2021.32
Divyashree, S., Shruthi, B., Vanitha, P., and Sreenivasa, M. (2023). Probiotics and their postbiotics for the control of opportunistic fungal pathogens: a review. Biotechnol. Rep. 38:e00800. doi: 10.1016/j.btre.2023.e00800
Elbaz, M., Chikly, A., Meilik, R., and Ben-Ami, R. (2022). Frequency and clinical features of Candida bloodstream infection originating in the urinary tract. J. Fungi 8:123. doi: 10.3390/jof8020123
Fan, Y., Su, Q., Chen, J., Wang, Y., and He, S. (2022). Gut microbiome alterations affect glioma development and Foxp3 expression in tumor microenvironment in mice. Front. Oncol. 12:836953. doi: 10.3389/fonc.2022.836953
Fatahi, A., Soleimani, N., and Afrough, P. (2021). Anticancer activity of kefir on glioblastoma cancer cell as a new treatment. Int. J. Food Sci. 2021:8180742. doi: 10.1155/2021/8180742
Fidan, I., Kalkanci, A., Yesilyurt, E., and Erdal, B. (2014). In vitro effects of Candida albicans and Aspergillus fumigatus on dendritic cells and the role of beta glucan in this effect. Adv. Clin. Exp. Med. 23, 17–24. doi: 10.17219/acem/37016
García-Gamboa, R., Perfecto-Avalos, Y., Gonzalez-Garcia, J., Alvarez-Calderon, M. J., Gutierrez-Vilchis, A., and Garcia-Gonzalez, A. (2024). In vitro analysis of postbiotic antimicrobial activity against Candida species in a minimal synthetic model simulating the gut mycobiota in obesity. Sci. Rep. 14:16760. doi: 10.1038/s41598-024-66806-3
Hanahan, D. (2022). Hallmarks of Cancer: new dimensions. Cancer Discov. 12, 31–46. doi: 10.1158/2159-8290.CD-21-1059
Jam, S. A. M., Talebi, M., Alipour, B., and Khosroushahi, A. Y. (2021). The therapeutic effect of potentially probiotic Lactobacillus paracasei on dimethylhydrazine induced colorectal cancer in rats. Food Biosci. 41:101097. doi: 10.1016/j.fbio.2021.101097
Karimi Ardestani, S., Tafvizi, F., and Tajabadi Ebrahimi, M. (2019). Heat-killed probiotic bacteria induce apoptosis of HT-29 human colon adenocarcinoma cell line via the regulation of Bax/Bcl2 and caspases pathway. Hum. Exp. Toxicol. 38, 1069–1081. doi: 10.1177/0960327119851255
Khosrovan, Z., Haghighat, S., and Mahdavi, M. (2020). The probiotic bacteria induce apoptosis in breast and colon cancer cells: an immunostimulatory effect. Immunoregulation 3, 37–50. doi: 10.32598/IMMUNOREGULATION.3.1.5
Li, X.-C., Wu, B.-S., Jiang, Y., Li, J., Wang, Z.-F., Ma, C., et al. (2021). Temozolomide-induced changes in gut microbial composition in a mouse model of brain glioma. Drug Des. Devel. Ther. 15, 1641–1652. doi: 10.2147/DDDT.S298261
Miller, K. D., Ostrom, Q. T., Kruchko, C., Patil, N., Tihan, T., Cioffi, G., et al. (2021). Brain and other central nervous system tumor statistics, 2021. CA Cancer J. Clin. 71, 381–406. doi: 10.3322/caac.21693
Mohseni, A. H., Casolaro, V., Bermúdez-Humarán, L. G., Keyvani, H., and Taghinezhad-S, S. (2021). Modulation of the PI3K/Akt/mTOR signaling pathway by probiotics as a fruitful target for orchestrating the immune response. Gut Microbes 13, 1–17. doi: 10.1080/19490976.2021.1886844
Öğüt, S., Türkol, M., Yikmiş, S., Bozgeyik, E., Abdi, G., Kocyigit, E., et al. (2025). Ultrasound-assisted enhancement of bioactive compounds in hawthorn vinegar: a functional approach to anticancer and antidiabetic effects. Ultrason. Sonochem. 114:107245. doi: 10.1016/j.ultsonch.2025.107245
Rossoni, R. D., De Barros, P. P., Mendonça, I. D. C., Medina, R. P., Silva, D. H. S., Fuchs, B. B., et al. (2020). The postbiotic activity of Lactobacillus paracasei 28.4 against Candida auris. Front. Cell. Infect. Microbiol. 10:397. doi: 10.3389/fcimb.2020.00397
Sadeghi-Aliabadi, H., Mohammadi, F., Fazeli, H., and Mirlohi, M. (2014). Effects of Lactobacillus plantarum A7 with probiotic potential on colon cancer and normal cells proliferation in comparison with a commercial strain. Iran. J. Basic Med. Sci. 17:815.
Seber, S., Sirin, D., Yetisyigit, T., and Bilgen, T. (2020). Piperlongumine increases the apoptotic effect of doxorubicin and paclitaxel in a cervical cancer cell line. Niger. J. Clin. Pract. 23, 386–391. doi: 10.4103/njcp.njcp_80_19
Siegel, R. L., Miller, K. D., Fuchs, H. E., and Jemal, A. (2022). Cancer statistics, 2022. CA Cancer J. Clin. 72, 7–33. doi: 10.3322/caac.21708
Spaggiari, L., Sala, A., Ardizzoni, A., De Seta, F., Singh, D. K., Gacser, A., et al. (2022). Lactobacillus acidophilus, L. plantarum, L. rhamnosus, and L. reuteri cell-free supernatants inhibit Candida parapsilosis pathogenic potential upon infection of vaginal epithelial cells monolayer and in a transwell coculture system in vitro. Microbiol. Spectr. 10:e02696-21. doi: 10.1128/spectrum.02696-21
Wang, L., Li, S., Fan, H., Han, M., Xie, J., Du, J., et al. (2022). Bifidobacterium lactis combined with Lactobacillus plantarum inhibit glioma growth in mice through modulating PI3K/AKT pathway and gut microbiota. Front. Microbiol. 13:986837. doi: 10.3389/fmicb.2022.986837
Wang, S., Shan, Y., Zhang, S., Zhang, L., Jiao, Y., Xue, D., et al. (2023). Lactobacillus paracasei subsp. paracasei X12 strain induces apoptosis in HT-29 cells through activation of the mitochondrial pathway. Nutrients 15:2123. doi: 10.3390/nu15092123
Keywords: Lactobacillus paracasei, anticancer, antifungal, apoptosis, sustainable food systems, postbiotics, probiotics
Citation: Erdal B, Yilmaz B, Bozgeyik E, Yıkmış S, Mohamed Ahmed IA, Aljobair MO and Trif M (2025) In vitro anticancer and antifungal effects of Lactobacillus paracasei supernatants: a step toward sustainable food systems. Front. Sustain. Food Syst. 9:1645521. doi: 10.3389/fsufs.2025.1645521
Edited by:
Saiful Irwan Zubairi, National University of Malaysia, MalaysiaReviewed by:
Iahtisham-Ul-Haq, Forman Christian College, PakistanEda Şensu, Gelisim University, Türkiye
Copyright © 2025 Erdal, Yilmaz, Bozgeyik, Yıkmış, Mohamed Ahmed, Aljobair and Trif. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Seydi Yıkmış, c3lpa21pc0Bua3UuZWR1LnRy; Moneera O. Aljobair, bW9hbGpvYmFpckBwbnUuZWR1LnNh