 Nesrine Rouini
Nesrine Rouini Alyssa Salazar1,2
Alyssa Salazar1,2 Kai Lepley
Kai Lepley Greg A. Barron-Gafford
Greg A. Barron-Gafford- 1School of Geography, Development and Environment, Tucson, AZ, United States
- 2Biosphere 2, University of Arizona, Tucson, AZ, United States
- 3Strategic Energy Analysis Center, National Renewable Energy Laboratory, Golden, CO, United States
Introduction: The increasing global demand for food and energy is intensifying land-use competition. Agrivoltaic systems are a multifunctional land-use approach that vertically integrates the production of agricultural crops and solar power on the same land area. Most food crops are adapted to full-sun conditions, and the physiological responses of these crops to the novel microclimate under solar panels remain poorly understood. We hypothesized that the microclimate beneath the high-density photovoltaic system would influence carbon uptake, water use, and yield outcomes of zucchini summer squash.
Methods: We conducted a field experiment in a hot, semi-arid climate on zucchini (Cucurbita pepo). Plants were grown under an agrivoltaic system with a 75% ground cover ratio (GCR) and in a full-sun control plot, each with two irrigation regimes (100 and 50%). We measured leaf-level photosynthesis, microclimate variables, and fruit yield at plant maturity and throughout the growing season.
Results: The agrivoltaic array reduced photosynthetically active radiation (PAR) by ~79%, resulting in a cooler (−1.1 °C), more humid environment with higher soil moisture. These microclimatic conditions enhanced midday photosynthesis and daily cumulative carbon uptake. However, fruit yield was consistently lower under the panels, indicating a shift in carbon allocation toward vegetative growth. Photosynthesis was primarily driven by PAR across treatments, while soil moisture significantly influenced photosynthesis only in the control plots, suggesting water limitation was alleviated under the panels.
Discussion: These findings highlight a trade-off between improved physiological performance and reduced yield under high-density agrivoltaics. While the system buffered heat and drought stress and improved overall plant function, excessive shade reduced reproductive output. Optimizing panel density or selecting crops cultivated for non-fruit yields will be essential for balancing food production and energy generation in dryland agrivoltaic settings.
Introduction
Providing food and energy to a growing population is increasingly challenging under climate change and as suitable land declines (IPCC, 2019). In the Southwest US, projected warming and drying trends pose a serious threat to food crop production through water deficits and heat stress. At the same time, consumer demand continues to increase (University of Arizona, 2023; US Global Change Research Program, 2018). Strategies for agricultural climate adaptation are increasingly recognized as critical for sustaining productivity under a changing climate. Within the United States, the USDA Climate Hubs encourage adoption of agronomic practices including crop diversification, improved water-use efficiency, and soil health management through cover cropping and reduced tillage (United States Department of Agriculture (USDA), 2023). However, most of these strategies referred to as climate-smart agricultural practices, have been shown to lose effectiveness under a warming climate (IPCC, 2022). Ecosystem approaches such as agroecology and agroforestry on the other hand are considered some of the most effective strategies that help ensure long term crop productivity while acting as carbon sinks, lowering agricultural greenhouse gas emissions and mitigating temperature extremes (Dittmer et al., 2023; Schoeneberger et al., 2012; IPCC, 2022).
Agrivoltaics is another climate-smart technology that offers a solution to adaptation challenges and to land competition between agriculture and the energy production sector by combining food production and electric power generation to optimize land-use efficiency (Dupraz et al., 2010). Similar to the vertical stratification created by agroforestry (Dupraz et al., 2010), these systems have also been shown to have other environmental and socio-economic benefits related to water saving (Adeh et al., 2018; TSE Energy, 2025) reducing plant stress (TSE Energy, 2025; Barron-Gafford et al., 2019), and supporting energy and food security (Dupraz et al., 2010; Barron-Gafford et al., 2019).
The installation of a photovoltaic array within an agricultural setting reduces photosynthetically active radiation (PAR) throughout the day. However, the degree and timing of this reduction varies across agrivoltaic designs. Even within overhead agrivoltaic systems, which create an ‘overstory’ of PV panels and an understory of agriculture, some designs utilize a full-density of panels, and some intentionally create gaps between the panels to allow for light transmission. Because of this reduction in direct incoming radiation, agrivoltaics can alter the microclimate beneath the solar panels relative to the surrounding environment. Under solar panels in agrivoltaic systems, daytime temperatures have been found to be over 1 °C cooler on average (Barron-Gafford et al., 2025; Weselek et al., 2021; Marrou et al., 2013) or not vary between agrivoltaics and full-sun treatments (Marrou et al., 2013). While insufficient PAR can limit photosynthesis and yield, excessive PAR can also damage the photosynthetic machinery of plants (Chen et al., 2023; Barron-Gafford et al., 2025). Understanding the ways that different crops respond physiologically to these varying light levels under solar panels is essential for optimizing agrivoltaic designs, but these responses remain understudied.
Here we quantify these dynamic shade-plant interactions and the ways that the agrivoltaic microclimate can affect the growth, ecophysiology, and water-use of the summer squash zucchini (Cucurbita pepo) as a major vegetable crop around the world. We test whether a high-density agrivoltaic system in a dryland environment improve physiological performance of zucchini while evaluating associated yield trade-offs. Our results provide important insights for enhancing zucchini production within agrivoltaic system in drylands and informing agrivoltaic design strategies and modeling for this crop.
Materials and methods
To understand the influence of microclimate parameters on leaf-level photosynthesis of zucchini in a dryland agrivoltaic setting, we ran an open-air field experiment at the Biosphere 2 Agrivoltaics Learning Lab in Oracle, Arizona, USA (32.578989°N, 110.851103°W; elevation 1,381 m above sea level). In the summer of 2022, we planted zucchini ‘Black Beauty’ (Cucurbita pepo) seedlings under a 15 m by 20 m photovoltaic array with a 75% ground cover ratio (GCR), and in a control full-sun treatment plot located 15 m to the west of the agrivoltaics system. Plants in the agrivoltaic and control plots received two irrigation regimes as a secondary treatment: full-water irrigation (100%) and half-water irrigation (50%). We tested four treatments: (1) Control + 100% irrigation, (2) Control + 50% irrigation, (3) Agrivoltaic + 100% irrigation, and (4) Agrivoltaic + 50% irrigation.
Leaf-level photosynthesis
Once plants reached reproductive maturity, we measured leaf-level net photosynthesis using LI-6400XT infrared gas analyzers (LI-COR Inc., NE, USA). We used the red–blue light source (LI-6400-02b) attached to the leaf cuvette to provide constant irradiance of ambient light levels for each measurement area (open-field sun versus the shade under the PV panels). Following best practices developed in previous work (Barron-Gafford et al., 2019; Barron-Gafford et al., 2025), cuvette air temperature was set to match that of ambient conditions at every measurement time point, and the reference CO2 was held constant at 400 ppm across all measurements. We took hourly diurnal photosynthesis measurements throughout the day from 05:00 to 19:00—for a total of 14 measurement periods to capture a diurnal cycle of carbon uptake. For each of the four treatments, we measured five plant replicates for a total of 20 individuals. During each hour of measurement throughout the day we measured five leaves per plant replicate. Due to a chamber configuration error, the leaf in the control 50% irrigation treatment was exposed to a non-representative PAR level during the 7:00 measurement. This value was excluded from the analysis and figures.
Light-saturation point was derived from morning diurnal photosynthesis data (07:00–12:00 local time) filtered to treatment-specific 10th–75th percentiles of air temperature to avoid midday depression. For each treatment, we fit leaf light–response curves (net CO₂ assimilation A, versus PAR) using a non-rectangular hyperbola (NRH). The fitted light-saturated value was defined as A∞ = Pmax − Rd from the NRH parameters (Pmax: gross light-saturated photosynthesis; Rd: day respiration). We report an apparent light-saturation point defined as the PAR (μmol m−2 s−1) at which modeled net photosynthesis reaches 90% of A∞ (Niinemets et al., 2015; Hieke et al., 2002; Stangl et al., 2022; Gómez-Gallego et al., 2025).
Microclimate
The microclimatic instrumentation system used for this study was described by Barron-Gafford et al. (2019). In the agrivoltaic and control plots, ambient air temperature (°C) and relative humidity (%) were measured with a shaded, aspirated temperature probe (Vaisala HMP60, Vaisala, Helsinki, Finland), and PAR was measured using LI-COR quantum flux sensor (PAR; LI-190R, LI-COR, Lincoln, NE, USA). Both probes were mounted on a post placed within the center of each installation at 2.5 m above the soil surface. In addition, on the day of measurements, we used the LICOR LI-6400XT sensors to spot check air temperature and PAR at the start of every hour. PAR is reported as photon flux density (μmol m−2 s−1). We monitored volumetric water content (VWC) and soil temperature at 5 cm depth (ECH2O 5TM, METER Group, Pullman, WA, USA) at six points across each of the control and agrivoltaic system sites. Data was averaged across the six points per treatment to give a single representative value for each time period for each site. All measurements were recorded at 30-min intervals throughout a 24-h day (CR1000, Campbell Scientific, Logan, UT, USA).
Statistical analysis
We used Python to perform our data processing and statistical analysis. We adopted a mixed statistical analysis approach combining independent t-tests and multiple linear regression modeling. This dual approach allowed us to identify micrometeorological variables that exhibited significant differences between the two primary treatments (the agrivoltaic system and the full-sun control) and then to explore the potential relationships between these variables and photosynthetic activity. The micrometeorological variables investigated were air temperature (T-air), PAR, soil temperature (T-soil), soil moisture (VWC), and vapor pressure deficit (VPD). To achieve a normal distribution our microclimate data was log transformed prior to analysis.
The independent t-tests were conducted to compare the means of each micrometeorological variable between the agrivoltaic system and the full-sun control, allowing us to determine whether the differences observed were statistically significant. Following this, multiple linear regression modeling was employed to investigate the relationships between the elected micrometeorological variables and photosynthesis. This approach enabled us to quantify the extent to which variations in T-air, PAR, T-soil, VWC, and VPD could explain changes in photosynthetic rates, providing deeper insights into the interplay between microclimate and plant physiology in the context of agrivoltaic systems.
The t-tests indicated that PAR, soil temperature, and soil moisture were significantly different between treatments. These three variables were then used as predictors in multiple linear regression models to quantify their individual and combined effects on photosynthetic rates within each treatment. This approach allowed us to determine not only if the microclimate differed, but the ways that these differences affected plant physiology.
When constructing multiple linear regression models we encountered high multicollinearity between T-soil and VWC, as indicated by their variance inflation factors (VIFs). T-soil consistently demonstrated the highest VIFs across all models, suggesting it contributed the most to multicollinearity. To address this issue, we removed T-soil from the models and retained PAR and VWC as predictors.
Results and discussion
Microclimate
The dense solar panel array in our experimental dryland agrivoltaic system effectively attenuated incoming solar radiation, leading to a significant reduction in PAR of 78.6% under the solar panels (Figure 1a). Mean daytime air temperature was reduced by 1.1 °C compared to the full-sun control plot, with this temperature mitigation observed between 6:00 a.m. and 6:00 p.m. (Figure 1b).
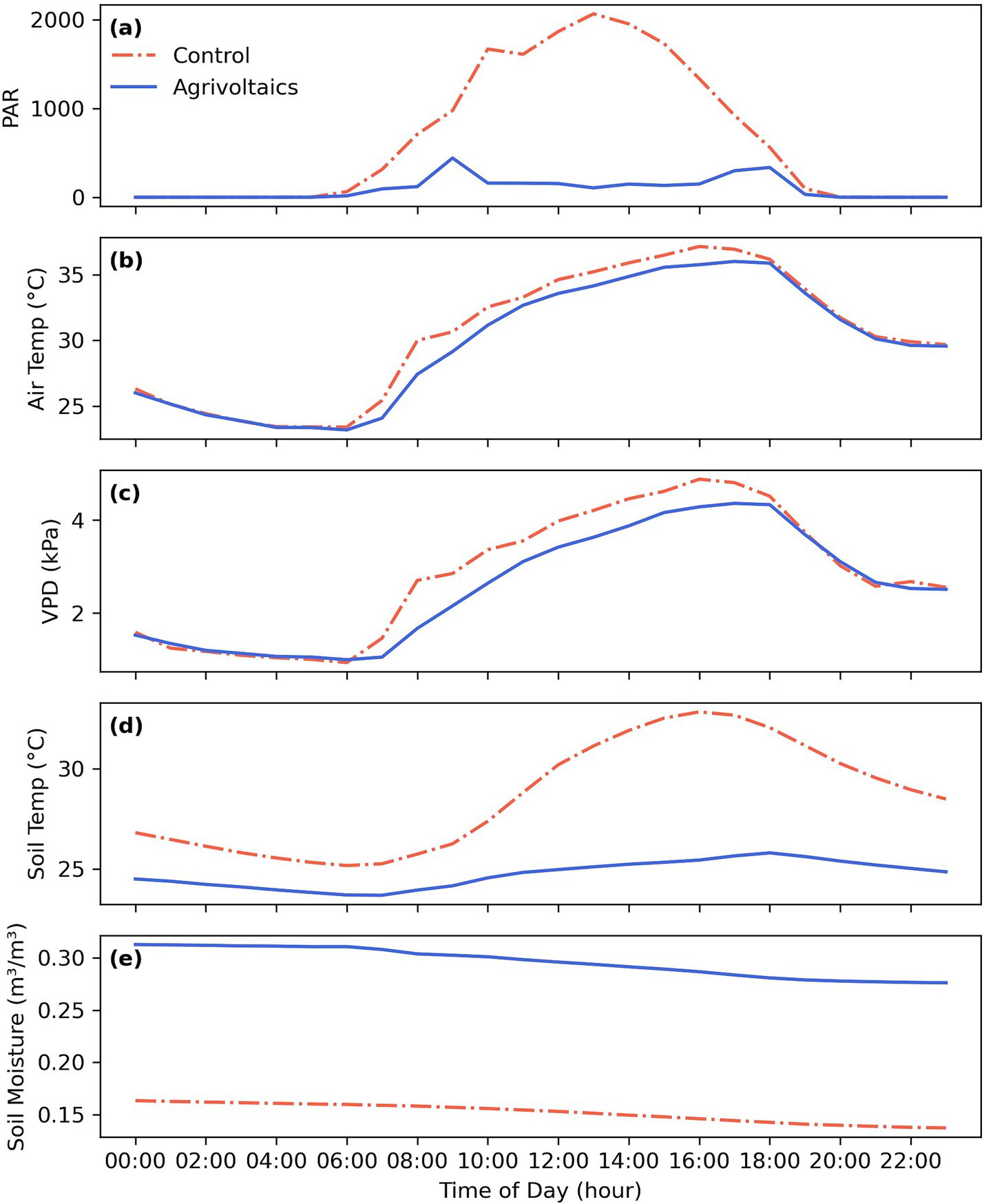
Figure 1. Representative diurnal microclimatic conditions in terms of (a) photosynthetically active radiation PAR, (b) ambient air temperatures, (c) vapor pressure deficit (VPD) as a measure of atmospheric dryness, and (d) soil temperature and (e) soil moisture content in a dryland agrivoltaic system (blue) vs. full-sun conditions in the control plot (red) during the 2022 growing season in Oracle, Arizona, USA.
Concurrently, the agrivoltaic system exhibited lower VPD by 0.5 kPa, indicating a more humid microclimate compared to the control throughout the daytime hours (Figure 1c). Notably, soil temperature in the agrivoltaic plot remained relatively stable between 24 and 25.5 °C, while the control plot soil experienced a pronounced increase from 25.5 °C to 32.5 °C between 9:00 a.m. and 4:00 p.m. (Figure 1d). In addition, the reduced temperature and PAR under the solar panels, coupled with the lower VPD, led to higher average soil volumetric water content (VWC). VWC averaged 0.30 m3/m3 under agrivoltaics compared to control 0.15 m3/m3 in the control (Figure 1e). These results underscore pronounced microclimatic modifications in the agrivoltaic system. The attenuated solar radiation resulted in a cooler and more humid environment within the crop canopy and soil. These microclimatic modifications have implications for plant photosynthesis and productivity, as they are the primary aboveground parameters that limit plant function.
Across irrigation and shading treatments, PAR was consistently statistically significant and had a positive relationship with photosynthesis (p < 0.001; Table 1). This underscores the fundamental role of light as a primary driver of photosynthetic carbon assimilation in C₃ crops, in line with theoretical models (Farquhar et al., 1980) and empirical evidence (Evans and Poorter, 2001). Zucchini’s positive response to PAR, even under shaded conditions, highlights the plant’s heliophilic nature and the importance of optimizing light management in agrivoltaic systems. Light saturation point estimated from morning diurnal photosynthesis was around 800 μmol m−2 s−1 under full-irrigation treatments in both AV and control. It increased to 1,100 μmol m−2 s−1 under control half-irrigation and 2,500 μmol m−2 s−1 under AV half-irrigation. C₃ leaves often approach saturation by around 800–1,200 μmol m−2 s−1 PAR under non-stress conditions (Huang et al., 2021; Huber et al., 2021; Gavhane et al., 2023). The increase of light-saturation point under half-irrigation indicates that photosynthesis is responding less efficiently to light (lower quantum yield) and only approaches its light-saturated rate at much higher PAR. Increasing irradiance only yields minimal gains under heat and water stress. We emphasize that these are apparent, field-condition light saturation points derived from diurnal fits rather than dedicated light response curves and should be approached as indicative thresholds. While PAR is the primary driver of net photosynthesis across treatments, the treatment-specific light saturation point values show where gains become insignificant. Increases in PAR will boost photosynthesis when the canopy operates below the light saturation point but provide little benefit once light saturation point is elevated under water and heat stress (Flexas and Medrano, 2002).
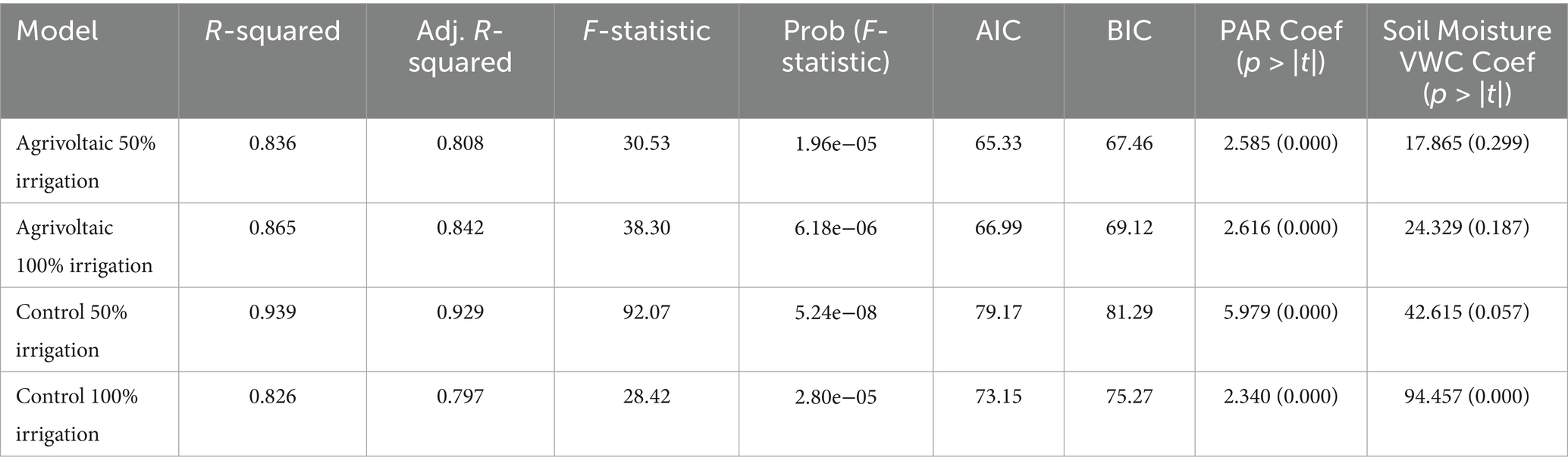
Table 1. Performance metrics for photosynthesis prediction regression models across treatments: agrivoltaics half irrigation water, agrivoltaics full water, control low water, and control full water.
The influence of soil moisture on photosynthesis varied markedly between the two shade treatments. In the full-sun control treatment, soil moisture was positively associated with photosynthesis in both 50 and 100% irrigation treatments, reaching statistical significance in the latter (p = 0.057 and p < 0.001, respectively). These results suggest that under full sun, water availability was a limiting factor for photosynthesis—likely due to stomatal closure under high VPD conditions, which reduces intercellular CO₂ concentration and suppresses carbon assimilation (Chaves et al., 2003, 2009; Flexas et al., 2004).
In the agrivoltaic treatment, no statistically significant relationship between soil moisture and photosynthesis was detected, regardless of irrigation regime. This might indicate that zucchini plants grown under shade were not water limited. The reduction in solar radiation and ambient temperature under the panels likely decreased evaporative demand and transpiration, thereby conserving soil moisture and maintaining plant water status (Marrou et al., 2013; Barron-Gafford et al., 2019). These findings align with previous studies reporting that agrivoltaic systems reduce evapotranspiration and prolong soil water availability in arid and semi-arid regions (Valle et al., 2017; Sekiyama and Nagashima, 2019; Warmann et al., 2024). By stabilizing leaf water potential and supporting continuous stomatal conductance (Montanaro et al., 2009), agrivoltaic shading allows for sustained photosynthetic function even under limited irrigation. An important outcome of this finding is illustrating that as dryland regions become warmer and experience increased periods of atmospheric and hydrologic drought, agrivoltaics can serve as a tool to allow for continued food production despite reduced access to water for irrigation.
Photosynthesis
We observed differences in the photosynthetic rates of zucchini through time across treatments (Figure 2). In the morning, through measurements at 08:00, rates of zucchini photosynthesis increased through time as PAR increased in both the agrivoltaic and control treatments at similar rates. By 10:00, morning light was no longer directly hitting agrivoltaic zucchini crops, and photosynthetic rates stabilized in the agrivoltaic treatment around 13.5 ± 0.65 μmol m−2 s−1 for the remainder of the day. However, photosynthetic rates continued to increase in the control treatment as light continued to intensify, reaching a peak by 10:00 with a rate of around 16.4 ± 1.3 μmol m−2 s−1. After 10:00, zucchini photosynthetic rates declined within the control treatment despite light levels increasing. After 14:00, zucchini photosynthetic rates increased in the agrivoltaic treatments while in the control treatment photosynthetic rates rapidly declined for the reminder of the day below the stable photosynthetic rate of zucchini in the agrivoltaic treatment in both irrigation treatments.
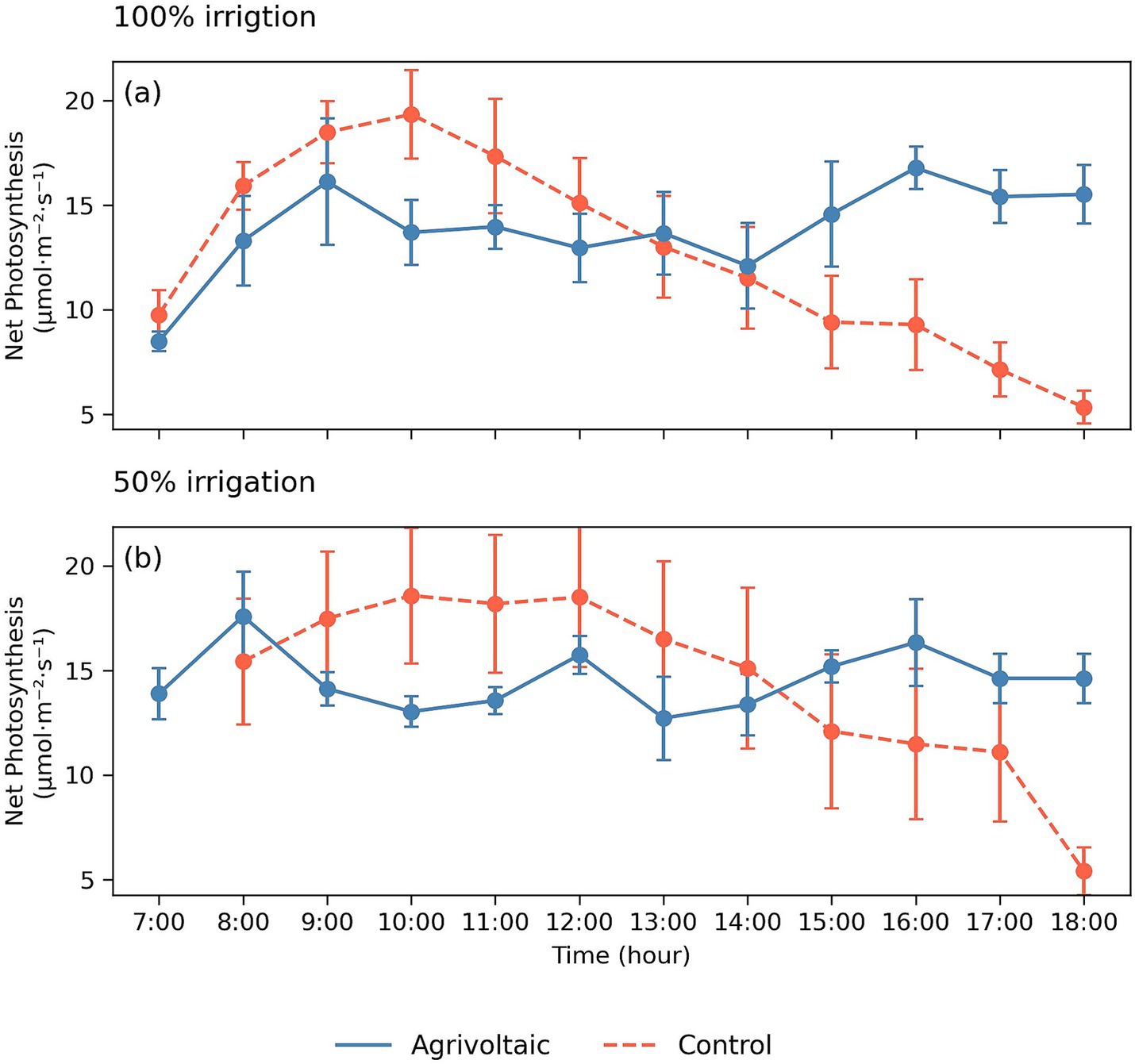
Figure 2. Diurnal photosynthetic rates of zucchini under agrivoltaic and control conditions. (a) 100% irrigation sub-treatments. (b) 50% irrigation sub-treatments.
The higher PAR intensity in the control treatment coupled with higher air temperature and VPD, representative of many dryland environments, likely induced photoinhibition and caused these control plants’ photosynthetic rates to crash by 12:00 p.m. Photoinhibition is a light-induced reduction of plant photosynthetic capacity that can cause crop yield losses by reducing plant carbon accumulation (Scheller and Haldrup, 2005; Barth et al., 2001). This specific form of photoinhibition that impacts diurnal patterns of carbon assimilation is referred to as midday depression of photosynthesis (Roessler et al., 1985; Barron-Gafford et al., 2025) and has been well documented across natural vegetation and cropping systems. The shading provided by the agrivoltaic system helped reduce PAR exposure of the crops as well as the rates of evaporation, preventing excessive light and drought stress while allowing sufficient light for consistent photosynthesis to occur. This protective effect is especially prominent in water-limited environments, where shaded plants experience less heat and drought stress (Kannenberg et al., 2023), further diminishing the risk of mid-day depression of photosynthesis (Barron-Gafford et al., 2025). Here, the reduced air temperature and increased humidity in the agrivoltaic plot enhanced stomatal conductance, which increased daily CO2 uptake of zucchini plants. Similar empirical observations have been shown related to kiwi leaves (another C3 plant), where a 50% reduction in incident light on kiwifruit leaves under drought conditions resulted in enhanced water use efficiency and higher photosystem II efficiency (Montanaro et al., 2009).
Cumulative carbon uptake and fruit yield
Cumulative carbon uptake was higher in the agrivoltaic system and highest under the 50% irrigation regime (Figure 3a). However, this increased carbon uptake did not translate into higher fruit yields in the agrivoltaic setting. In fact, regardless of water treatment, the control plants consistently produced higher fruit yields compared to the agrivoltaic system (Figure 3b). While moderate shade can protect plant function and intentionally reduce evaporative loss of soil moisture, excessive shade can negatively impact yields (Warmann et al., 2024). Determining the balance between stress reduction and light limitation is species and context dependent, and creating a tool to best estimate the ground cover ratio (GCR) of solar energy sharing remains a top priority in agrivoltaics research. In temperate climates where sunlight is not a limiting factor, field trials have shown that a lightly shaded crop can yield as well or better than full-sun grown crops (Weselek et al., 2021). However, as panel coverage increases, agricultural yields begin to decline: in one wheat experiment, ~23% shade caused only a 7% fruit yield reduction in a shade-tolerant variety, but ~31% shade led to a 50% fruit yield reduction in a shade sensitive variety (Li et al., 2010; Dufour et al., 2013). Ultimately, this leads to a lower harvest index (the ratio of agricultural yield to total biomass) in shaded crops, which is what we observed here with zucchini plants.
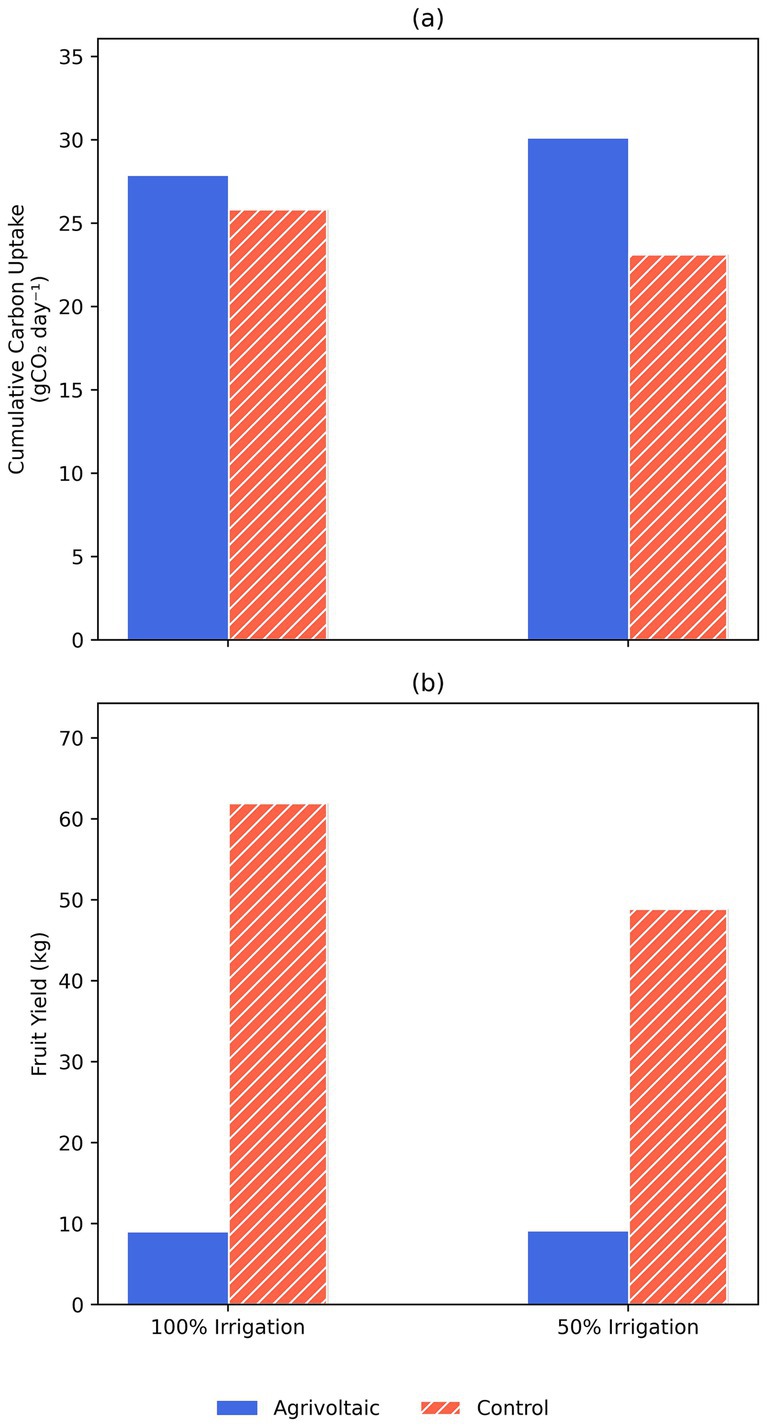
Figure 3. Cumulative daily carbon uptake (a) and fruit yield (b) of zucchini under control vs. agrivoltaic conditions with 100 and 50% irrigation.
Shade modifies the morphological and anatomical characteristics of leaves (Boardman, 1977; Araus et al., 1986; Andersen et al., 1991). Zucchini plants in these agrivoltaic conditions exhibited shade-acclimation and shade-avoidance responses in their morphology and leaf anatomy which influences carbon allocation. The higher cumulative carbon uptake in shaded zucchini may have been accompanied by increased investment in leaf and stem biomass, a hallmark of shade avoidance, which can lead to a lower harvest index and ultimately reduce reproductive allocation (Smith and Whitelam, 1997). This suggests that even when total photosynthetic carbon gain increases under shade, reproductive output can suffer if assimilates are preferentially allocated to vegetative structures rather than fruits. In shade-grown C3 plants, shade-induced elongation and increased allocation to stems can carry substantial costs, reducing whole-plant biomass and photosynthesis when elongation fails to improve light capture (Weinig, 2000).
Along with size and stature adjustments, shaded foliage tends to have higher specific leaf area (SLA). Leaves growing in shade conditions typically contain more chlorophyll per unit area, appearing darker green, which enhances light absorption in low light (Zhang et al., 2022; Gotoh et al., 2018). They are also often characterized by a reduced palisade mesophyll layer and a greater proportion of spongy mesophyll, resulting in overall thinner leaf blades (Gotoh et al., 2018). This anatomy enhances light interception and internal CO₂ diffusion efficiency by increasing the surface area per unit mass, manifested as higher SLA. Increased SLA in leaves growing in the shade correlates with higher mesophyll conductance, facilitating improved CO₂ diffusion to the chloroplasts and thereby enhancing photosynthetic efficiency under limited light conditions (Niinemets, 1999; Flexas et al., 2008).
Differences in leaf area between the leaves of zucchini plants grown in the agrivoltaic plot and the ones in the control plot were visually noticeable in the field. This aligns with earlier empirical observations by different authors: lettuce grown under an AV system compensated for 50% light reduction by producing increased total leaf area per plant, thereby maintaining radiation interception and yield (Marrou et al., 2013). Shaded winter wheat had a taller canopy (+8–20% height) and periodically higher leaf area index reflecting an elongation and leaf-expansion response to reduced light (Weselek et al., 2021; McMaster et al., 1987). Such morphological changes are a well-known shade-avoidance strategy in C3 plants, driven by a lower red:far-red light ratio, and are thought to reflect an effort to outcompete neighbors for light (Ballaré and Pierik, 2017).
The design of our agrivoltaic installation induced excessive shade, resulting in reduced zucchini fruit yield, despite greater and more consistent rates of carbon assimilation. Maintaining a panel density of 75% GCR, as was done here for optimized energy production, would require any colocation with agriculture to use crops that are either shade tolerant or have a high degree of shade acclimation potential. Such a system may be more economically advantageous to crop types such as leafy greens and herbs that are cultivated for their leaves. The increased leaf area resulting from mitigated environmental stresses and morphological adjustments, could enhance leafy greens agricultural productivity and economic value while reducing their water usage. Crops cultivated for their reproductive organs such as zucchini may be a suitable crop for agrivoltaic farms in high irradiance, hot semi-arid regions at a panel density that should not exceed a 25–50% GCR where microclimate benefits can be maximized without excessively reducing the sunlight needed for a normal fruit yield. Finally, it should be noted that these are the results from one growing season, and agricultural yields are inherently variable from year to year. Thus, these findings and conclusions might not reflect long-term crop outcomes in high-shade agrivoltaic environments.
Conclusion
This study investigated the interactions between microclimate, plant physiology, and yield of zucchini under a 75% GCR dryland agrivoltaic system. The array reduced PAR and created a cooler, more humid microclimate, which enhanced midday photosynthesis and increased daily cumulative carbon uptake. However, these gains were largely allocated to leaf growth and shade-acclimation processes rather than fruit production, resulting in lower fruit yield under the array. Under high-density agrivoltaics the microclimate alleviates heat and water constraints and sustains photosynthesis but shifts carbon allocation from reproduction.
A 75% GCR configuration is unlikely to be optimal for the agricultural output of fruiting crops in hot semi-arid regions such as zucchini. System design should balance panel density and crop selection. Using lower GCR or greater panel spacing and prioritizing shade-tolerant or leaf vegetables to leverage the water-saving benefits without excessively limiting light for fruit set. In water-limited drylands, the buffering of temperature and VPD under arrays can reduce irrigation requirements; optimizing array density together with irrigation management should maximize both agricultural and energy yields.
Future research should explore a wider range of crops, including leafy greens and other vegetables, under varying agrivoltaic array densities to determine optimal configurations for maximizing both crop productivity and renewable energy generation in water-stressed environments. Specifically, quantifying leaf functional traits and investigating the physiological mechanisms underlying the observed shift in carbon allocation from fruit to vegetative growth in zucchini under shade conditions will be critical for informing optimized agrivoltaic systems. Intelligent agrivoltaic design will require a nuanced understanding of crop-specific responses to the modified microclimate to achieve the desired balance between food production, water conservation, and renewable energy generation.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: DOI: 10.25422/azu.data.29922692.
Author contributions
NR: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. AS: Investigation, Writing – review & editing. PM: Data curation, Writing – review & editing. KL: Formal analysis, Writing – review & editing. GB-G: Conceptualization, Funding acquisition, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research and data were supported by (1) the National Science Foundation Geography and Spatial Sciences through Award 2025727; (2) the Accelerate For Success Grants Program from the Research, Innovation & Impact office at the University of Arizona; (3) the National Renewable Energy Laboratory’s InSPIRE project, through the U.S. Department of Energy Office of Energy Efficiency and Renewable Energy Solar Energy Technologies Office under award DE-EE00034165, and (4) U.S. Department of Agriculture’s National Institute of Food and Agriculture (NIFA) Sustainable Agricultural Systems project entitled ‘Sustainably Co-locating Agricultural and Photovoltaic Electricity Systems (SCAPES),’ led by the University of Illinois Urbana-Champaign, Grant No. 2021-68012-35898, 2021–2025.
Acknowledgments
The authors thank John Adams and the Biosphere 2 team for their assistance in the maintenance of the Biosphere 2 Agrivoltaics Learning Lab.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adeh, E. H., Good, S. P., Calaf, M., and Higgins, C. W. (2018). Remarkable agrivoltaic influence on soil moisture, micrometeorology and water-use efficiency. PLoS One 13:e0203256. doi: 10.1371/journal.pone.0203256
Andersen, P. C., Knox, G. W., and Norcini, J. G. (1991). Influence of irradiance on leaf physiology and growth characteristics of Rhododendron x ‘pink ruffles’. J. Am. Soc. Hort. Sci. 116, 881–887. doi: 10.21273/JASHS.116.5.881
Araus, J. L., Alegre, L., Tapia, L., Calafell, R., and Serret, M. D. (1986). Relationship between photosynthetic capacity and leaf structure in several shade plants. Am. J. Bot. 73, 1760–1770. doi: 10.1002/j.1537-2197.1986.tb09708.x
Ballaré, C. L., and Pierik, R. (2017). The shade-avoidance syndrome: multiple signals and ecological consequences. Plant Cell Environ. 40, 2530–2543. doi: 10.1111/pce.12914
Barron-Gafford, G. A., Murphy, P., Salazar, A., Lepley, K., Rouini, N., Barnett-Moreno, I., et al. (2025). Agrivoltaics as a climate-smart and resilient solution for midday depression in photosynthesis in dryland regions. npj Sustain. Agric. 3:32. doi: 10.1038/s44264-025-00073-1
Barron-Gafford, G. A., Pavao-Zuckerman, M. A., Minor, R. L., Sutter, L. F., Barnett-Moreno, I., Blackett, D. T., et al. (2019). Agrivoltaics provide mutual benefits across the food–energy–water nexus in drylands. Nat. Sustain. 2, 848–855. doi: 10.1038/s41893-019-0364-5
Barth, C., Krause, G. H., and Winter, K. (2001). Responses of photosystem I compared with photosystem II to high-light stress in tropical shade and sun leaves. Plant Cell Environ. 24, 163–176. doi: 10.1046/j.1365-3040.2001.00673.x
Boardman, N. K. (1977). Comparative photosynthesis of sun and shade plants. Ann. Rev. Plant Physiol. 28, 355–377. doi: 10.1146/annurev.pp.28.060177.002035
Chaves, M. M., Flexas, J., and Pinheiro, C. (2009). Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Ann. Bot. 103, 551–560. doi: 10.1093/aob/mcn125
Chaves, M. M., Maroco, J. P., and Pereira, J. S. (2003). Understanding plant responses to drought—from genes to community ecology. Funct. Plant Biol. 36, 1096–1103. doi: 10.1071/FP02076
Chen, L. X., Mao, H. T., Lin, S., Din, A. M. U., Yin, X. Y., Yuan, M., et al. (2023). Different photosynthetic response to high light in four Triticeae crops. Int. J. Mol. Sci. 24:1569. doi: 10.3390/ijms24021569
Dittmer, K. M., Rose, S., Snapp, S. S., Kebede, Y., Brickman, S., Shelton, S., et al. (2023). Agroecology can promote climate change adaptation outcomes without compromising yield in smallholder systems. Environ. Manag. 72, 333–342. doi: 10.1007/s00267-023-01816-x
Dufour, L., Metay, A., Talbot, G., and Dupraz, C. (2013). Assessing light competition for cereal production in temperate agroforestry systems using experimentation and crop modelling. J Agron Crop Sci. 199, 217–227.
Dupraz, C., Marrou, H., Talbot, G., Dufour, L., Nogier, A., and Ferard, Y. (2010). Combining solar PV panels and food crops for optimising land use: towards new agrivoltaic schemes. Renew. Energy 36, 2725–2732. doi: 10.1016/j.renene.2011.03.005
Evans, J. R., and Poorter, H. (2001). Photosynthetic acclimation of plants to growth irradiance: the relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ. 24, 755–767. doi: 10.1046/j.1365-3040.2001.00724.x
Farquhar, G. D., von Caemmerer, S., and Berry, J. A. (1980). A biochemical model of photosynthetic CO₂ assimilation in leaves of C₃ species. Planta 149, 78–90. doi: 10.1007/BF00386231
Flexas, J., Bota, J., Loreto, F., Cornic, G., and Sharkey, T. D. (2004). Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 6, 269–279. doi: 10.1055/s-2004-820867
Flexas, J., and Medrano, H. (2002). Drought-inhibition of photosynthesis in C3 plants: stomatal and non-stomatal limitations revisited. Ann. Bot. 89, 183–189. doi: 10.1093/aob/mcf027
Flexas, J., Ribas-Carbó, M., Diaz-Espejo, A., Galmés, J., and Medrano, H. (2008). Mesophyll conductance to CO₂: current knowledge and future prospects. Plant Cell Environ. 31, 602–621. doi: 10.1111/j.1365-3040.2007.01757.x
Gavhane, K. P., Hasan, M., Singh, D. K., Kumar, S. N., Sahoo, R. N., and Alam, W. (2023). Determination of optimal daily light integral (DLI) for indoor cultivation of iceberg lettuce in an indigenous vertical hydroponic system. Sci. Rep. 13:10923. doi: 10.1038/s41598-023-36997-2
Gómez-Gallego, M., Sellier, D., Dickson, A., Bader, M. K., Leuzinger, S., and Williams, N. (2025). Light limitation and foliar pathogenic infection impact phloem anatomy and function in Pinus radiata D. Don. Plant Cell Environ. 48, 6356–6372. doi: 10.1111/pce.15610
Gotoh, E., Suetsugu, N., Higa, T., Matsushita, T., Tsukaya, H., and Wada, M. (2018). Palisade cell shape affects the light-induced chloroplast movements and leaf photosynthesis. Sci. Rep. 8:1472. doi: 10.1038/s41598-018-19896-9
Hieke, S., Menzel, C. M., and Lüdders, P. (2002). Effects of light availability on leaf gas exchange and expansion in lychee (Litchi chinensis). Tree Physiol. 22, 1249–1256. doi: 10.1093/treephys/22.17.1249
Huang, M. Y., Wong, S. L., and Weng, J. H. (2021). Rapid light-response curve of chlorophyll fluorescence in terrestrial plants: relationship to CO2 exchange among five Woody and Four Fern species adapted to different light and water regimes. Plants (Basel) 10:445. doi: 10.3390/plants10030445
Huber, B. M., Louws, F. J., and Hernández, R. (2021). Impact of different daily light integrals and carbon dioxide concentrations on the growth, morphology, and production efficiency of tomato seedlings. Front. Plant Sci. 12:615853. doi: 10.3389/fpls.2021.615853
IPCC (2019). Climate Change and Land: An IPCC Special Report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems s (P.R. Shukla et al., Eds.). Available online at: https://www.ipcc.ch/srccl/
IPCC. (2022). Climate Change 2022: impacts, adaptation and vulnerability. Contribution of working group II to the sixth assessment report of the intergovernmental panel on climate change. Available online at: https://doi-org.ezproxy3.library.arizona.edu/10.1017/9781009325844
Jiang, Y., Wang, Y., Zhang, Y., and Liu, Y. (2022). Effects of different photovoltaic shading levels on kiwifruit growth, yield and water productivity under agrivoltaic system in Southwest China. Agric. Water Manag. 265:107572. doi: 10.1016/j.agwat.2022.107572
Kannenberg, S. A., Sturchio, M. A., Venturas, M. D., and Knapp, A. K. (2023). Grassland carbon-water cycling is minimally impacted by a photovoltaic array. doi: 10.1038/s43247-023-00904-4
Li, H., Jiang, D., Wollenweber, B., Dai, T., and Cao, W. (2010). Effects of shading on morphology, physiology and grain yield of winter wheat. Eur J Agron. 33, 267–275. doi: 10.1016/j.eja.2010.07.002
Marrou, H., Guilioni, L., Dufour, L., Dupraz, C., and Wery, J. (2013). Microclimate under agrivoltaic systems: is crop growth rate affected in the partial shade of solar panels? Agric. For. Meteorol. 177, 117–132. doi: 10.1016/j.agwat.2018.07.001
McMaster, G. S., Morgan, J. A., and Willis, W. O. (1987). Effects of shading on winter wheat yield, spike characteristics, and carbohydrate allocation1. Crop Science 27, 967–973.
Montanaro, G., Dichio, B., and Xiloyannis, C. (2009). Shade mitigates photoinhibition and enhances water use efficiency in kiwifruit under drought. Photosynthetica 47, 340–348. doi: 10.1007/s11099-009-0057-9
Niinemets, Ü. (1999). Components of leaf dry mass per area—thickness and density—alter leaf photosynthetic capacity in reverse directions in woody plants. New Phytol. 144, 35–47. doi: 10.1046/j.1469-8137.1999.00466.x
Niinemets, Ü., Sun, Z., and Talts, E. (2015). Controls of the quantum yield and saturation light of isoprene emission in different-aged aspen leaves. Plant Cell Environ. 38, 2707–2720. doi: 10.1111/pce.12582
Roessler, P. G., and Monson, R. K. (1985). Midday depression in net photosynthesis and stomatal conductance in Yucca glauca. Oecologia 67, 380–387. doi: 10.1007/BF00384944
Scheller, H. V., and Haldrup, A. (2005). Photoinhibition of photosystem I. Planta 221, 5–8. doi: 10.1007/s00425-005-1507-7
Schoeneberger, M. M., Bentrup, G., De Gooijer, H., Soolanayakanahally, R., Sauer, T., Brandle, J., et al. (2012). Branching out: agroforestry as a climate change mitigation and adaptation tool for agriculture. J. Soil Water Conserv. 67, 128A–136A. doi: 10.2489/jswc.67.5.128A
Sekiyama, T., and Nagashima, A. (2019). Solar sharing for both food and clean energy production: performance of agrivoltaic systems for corn, a typical shade-intolerant crop. Environments 6:65. doi: 10.3390/environments6060065
Smith, H., and Whitelam, G. C. (1997). The shade avoidance syndrome: multiple responses mediated by multiple phytochromes. Plant Cell Environ. 20, 840–844. doi: 10.1046/j.1365-3040.1997.d01-104.x
Stangl, Z. R., Tarvainen, L., Wallin, G., and Marshall, J. D. (2022). Limits to photosynthesis: seasonal shifts in supply and demand for CO2 in scots pine. New Phytol. 233, 1108–1120. doi: 10.1111/nph.17856
TSE Energy (2025). Reducing crop water stress through agrivoltaics. Available online at: https://www.tse.energy/en/articles/reduire-le-deficit-hydrique-des-cultures-grace-a-lagrivoltaisme
United States Department of Agriculture (USDA) (2023). Oklahoma State Agriculture Crop overview 2022. Available online at: https://www.nass.usda.gov/Quick_Stats/Ag_Overview/stateOverview.php?state=OKLAHOMA
US Global Change Research Program (2018) in Fourth National Climate Assessment, volume II: impacts, risks, and adaptation in the United States (chapter 25: Southwest). eds. D. R. Reidmiller, C. W. Avery, D. R. Easterling, K. E. Kunkel, K. L. M. Lewis, and T. K. Maycock, et al. (Washington, DC, USA: U.S. Global Change Research Program), 1506.
Valle, B., Simonneau, T., Sourd, F., Pechier, P., Hamard, P., Frisson, T., et al. (2017). Increasing the total productivity of a land by combining mobile photovoltaic panels and food crops. Appl. Energy 206, 1495–1507. doi: 10.1016/j.apenergy.2017.09.113
Warmann, E., Jenerette, G. D., and Barron-Gafford, G. A. (2024). Agrivoltaic system design tools for managing trade-offs between energy production, crop productivity and water consumption. Environ. Res. Lett. 19:034046. doi: 10.1088/1748-9326/ad2ab8
Weinig, C. (2000). Plasticity versus canalization: population differences in the timing of shade-avoidance responses. Evolution 54, 441–451. doi: 10.1111/j.0014-3820.2000.tb00047.x
Weselek, A., Bauerle, A., Hartung, J., Zikeli, S., Lewandowski, I., and Högy, P. (2021). Agrivoltaic system impacts on microclimate and yield of different crops within an organic crop rotation in a temperate climate. Agron. Sustain. Dev. 41:59. doi: 10.1007/s13593-021-00714-y
Keywords: dryland agrivoltaics, midday depression, food–energy–water nexus, crop carbon uptake, crop ecophysiology, microclimate
Citation: Rouini N, Salazar A, Murphy P, Lepley K and Barron-Gafford GA (2025) High-shade dryland agrivoltaic conditions enhanced carbon uptake and water-use efficiency in zucchini (Cucurbita pepo). Front. Sustain. Food Syst. 9:1686773. doi: 10.3389/fsufs.2025.1686773
Edited by:
Vijay Singh Meena, Indian Agricultural Research Institute (ICAR), IndiaReviewed by:
Shubham Durgude, International Maize and Wheat Improvement Center, MexicoNaveenkumar Patil, ICAR - Mahatma Gandhi Integrated Farming Research Institute, India
Copyright © 2025 Rouini, Salazar, Murphy, Lepley and Barron-Gafford. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nesrine Rouini, bmVzcmluZXJvdWluaUBhcml6b25hLmVkdQ==