 Shatha I. Alqurashi
Shatha I. Alqurashi- Department of Biological Science, College of Science, University of Jeddah, Jeddah, Saudi Arabia
Introduction: Organic waste presents a significant environmental challenge, causing environmental issues, such as landfill accumulation, greenhouse gas emissions, and water pollution. Food leftovers from restaurants, which are rich in fats and other elements, are among the most threatening organic waste materials in Saudi Arabia. This study evaluated the potential of black soldier fly (BSF) larvae, Hermetia illucens, to bioactive post-consumer food leftovers from restaurants into valuable protein-rich feed and organic fertilizer.
Methods: Three substrates were tested: 100% poultry feed (PF) as a control, 100% food leftovers from restaurants (LF 100%), and a 50:50 mixture of PF and LF (LF 50%). Larval growth performance, survival rate, proximate chemical composition, amino acid content, mineral profile, and toxic elements were assessed, alongside the safety of residual frass.
Results: Larvae fed the LF 50% achieved the highest growth and survival rates. Both larvae and frass contained essential amino acids and key macro- and micro-minerals, with heavy metal concentrations remaining below hazardous thresholds. The residual frass also showed potential as a plant growth-promoting fertilizer due to its amino acid composition. Therefore, these findings demonstrate that BSF bioconversion is an effective and sustainable approach for valorizing high-fat restaurant waste, an underexplored substrate in the region.
Conclusion: This study presents insights into the benefits of producing insect protein for feed and nutrient-rich frass, which can support agriculture from high-fat restaurant waste, an underexplored substrate in the region, contribute to food security, and align with environmental objectives under the Saudi Green Initiative. Further research should focus on optimizing the substrates, exploring the long-term applications of BSF larvae frass in agriculture, and scaling up the BSF larvae systems using various waste streams.
1 Introduction
Due to the expected increase in the world population, insect production has recently been considered a novel approach for addressing the shortage of protein resources for both animals and humans (Secci et al., 2022). In Saudi Arabia, an estimated four million tons of organic waste are generated annually, presenting significant concerns but also offering an opportunity for both economic and environmental benefits, in particular considering recent global crises, such as the Russia-Ukraine war, the Israeli-Palestinian conflict, and the COVID-19 pandemic, which have raised concerns regarding energy supply, food/feed availability, and dependence on imported food/feed and fertilizer (Attia et al., 2022). Recent research focused on bio-converting insects, particularly the black soldier fly (BSF), Hermetia illucens, (Diptera: Stratiomyidae), because of the larvae's ability to efficiently convert various organic wastes into valuable biomass, which is rich in both lipids and proteins (Scieuzo et al., 2023). Moreover, BSF produces bioactive compounds, including lauric acid, chitin, and antibacterial peptides, which can positively influence the intestinal and overall health of both humans and animals (Saviane et al., 2021; Candian et al., 2023). Recent studies have also shown that replacing poultry meal with BSF larvae in animal diets preserved nutrient digestibility and supported gut health, thereby confirming their safety and potential as a sustainable protein source for animal nutrition (Bosch et al., 2024). In particular, incorporating BSF larvae reared on animal-based substrates at a 3% replacement level for poultry meal in cat diets proved to be the most effective strategy, reinforcing their potential as a practical and efficient feed ingredient (Kim et al., 2025). Thus, after these larvae are harvested, the resulting residual frass, consisting of exuviae, excrement, and unused substrate, can be utilized as fertilizer or feedstock for biogas and biofuel production (Carroll et al., 2023; Wedwitschka et al., 2023; Mohan et al., 2023).
Several previous studies investigated the effects of various diets and organic waste types on the growth and development of BSF larvae, under both commercial and laboratory rearing conditions (St-Hilaire et al., 2007; Nguyen et al., 2015; Tinder et al., 2017). The life history of BSF larvae including the maturity, development rate, reproduction, and survival, is affected significantly by nutrition and temperature (Clissold and Simpson, 2015). The structure of the feed, including many factors, such as particle size, thickness, and moisture content, plays an important role in influencing its spreadability and ease of insect consumption. Numerous experiments have been conducted to explore various substrates for rearing BSF larvae, including different types of vegetable residue; kitchen waste; municipal organic waste, such as poultry, pig, and cattle manure; straw; dried distillers' grain with soluble content (DDGS); and fish waste (Van Huis, 2020; Meneguz et al., 2018; Oonincx et al., 2015). However, limited research has been conducted on the mineral content and chemical and amino acid profiles of larvae and their frass in relation to their substrate (Proc et al., 2020). Therefore, the composition of the larval diet plays a crucial role in the economics of BSF larval production, because of the trade-offs between the cost of the substrate, its nutritional quality, and its effect on larval development and body composition, including the lipid and protein content (Onsongo et al., 2018; Barragan-Fonseca et al., 2017). These trade-offs extend to the safety of the final insect product (Purschke et al., 2017).
Nowadays, considering the introduction of insects into the food chain, it is essential to determine the potential risks associated with their consumption, especially when different types of growth substrates are utilized. To advance the principles of the circular economy, many studies have examined the utilization of organic waste as a substrate for rearing BSF larvae. Regulation (EC) No. 767/2009 restricts the use of certain ‘unsafe' substrates, such as manure, digestive tract contents, catering waste, and processed animal protein. Although these materials are often preferred for oviposition by BSF (Boafo et al., 2023), they are prohibited from mitigating microbiological risks. However, additional potential risks have largely been overlooked. Organic waste often contains persistent pollutants, such as heavy metals that can accumulate in both larvae and frass, subsequently entering the food chain and the environment (Addeo et al., 2024). These heavy metals can enter waste streams in several ways, including via atmospheric emissions or the contamination of materials. Consequently, terrestrial organisms may ingest these pollutants, causing the escalation of environmental hazards (Diener et al., 2015).
Therefore, BSF demonstrates the most promising potential for industrial production. In particular, BSF larvae validate the efficient conversion of organic waste and produce valuable nutrients that are suitable for pet, fish, and poultry diets, along with residual fertilizer for soil improvement. This study aims to address several environmental issues. The primary aim is to develop alternative protein-rich feed and high-quality organic fertilizers from food leftovers in restaurants. It also seeks to assess the bioaccumulation of toxic elements in the larvae, in order to evaluate the risks to animal and human health. To achieve these objectives, this study examines the leftovers food from restaurant as the primary substrate. This substrate comprises a variety of organic components, including chicken, meat, rice, and diverse vegetables, such as arugula, onions, lemons, and lettuce. By investigating these elements, this study not only addresses food waste management, but also aligns with the Saudi Green Initiative (SGI) by reducing carbon emissions, mitigating disease-carrying organisms, preserving the environment, and promoting environmental sustainability.
2 Materials and methods
2.1 Rearing BSF
The BSF larvae used in this study were obtained from Egypt (Agro BIO Production Corporation). Subsequent generations were reared in colonies. These larvae were aged 6 days and had an average individual weight of approximately 0.088 mg (±0.009). They were transferred into rectangular rearing boxes (60 × 40 × 10 cm) at a density of two larvae/cm2. Each group was fed at an estimated rate of 0.93 kg of substrate per larvae per day in increasing quantities, resulting in a total average quantity of 13 kg for the entire period of approximately 14 days. The poultry feed used as a control substrate was formulated commercially (Arasco Feedmill, Broiler starter crumbs, SAP code: 20104013), and its chemical composition was analyzed. It included moisture content (13%), crude protein (20.5%), crude fat (2.5%), crude fiber (3%), methionine (0.50%), and lysine (1.20%).
The BSF pupae were placed in a mesh cage (36 × 36 × 36 cm) in a cooled incubator with a temperature range of 27 °C to 30 °C, a relative humidity of 65%, and a photoperiod of 12:12 L:D (light: dark) under undirected light. The rearing cages were supplied with water using a water-filled plastic cup (284 ml), which was inverted onto a 90 mm diameter filter paper placed inside a Petri dish lid of the same size. Eggs were collected using egg traps made of corrugated cardboard, cut into 10 × 6 cm pieces, and taped together to form four bundles (Booth and Sheppard, 1984). These traps were positioned 3 cm above the oviposition substrate and contained organic food waste. The eggs collected were then transferred to the laboratory and placed in rectangular plastic rearing boxes (60 cm × 40 cm × 10 cm) containing poultry feed. The rearing process was conducted under controlled laboratory conditions, with temperatures ranging from 27 °C to 30 °C and with a relative humidity of 65%. These steps were repeated to obtain overlapping generations.
The eggs hatched approximately 3 days after oviposition. The larval duration was approximately 22 days, processing through six instars. The larvae subsequently transformed into the pre-pupal stage and converted to the pupal stage (a non-feeding phase) over 14 days. The entire life cycle of the BSF was completed in approximately 45 days.
2.2 Studying area and substrates
This study was conducted using organic leftover food (LF) from Bukhari restaurants, which are particularly popular among Hajj and Umrah pilgrims and are widely known in the western region of Saudi Arabia. These restaurants are highly favored for their accessible and readily available meals. The leftovers were collected from six Bukhari restaurants in Jeddah and Makkah over three consecutive days. The leftovers collected were sorted carefully to ensure that they were free from solid, non-food waste, such as plastic and metal cans.
The organic food waste consisted of various of components, such as cooked chicken and meat, bones, raw chicken and meat pieces, rice, oils, roquette, onions, lemon, lettuce, various vegetables, and bread. The preparation of the LF involved placing it initially in a sieve to allow the excess liquid to drain in order to reduce the moisture content. The food waste was then ground using a chopping machine (JINFUDA) and mixed thoroughly to create a homogenous substrate suitable for the BSF larvae. The grinding process was repeated multiple times to ensure the consistency of the mixture. This approach enabled the effective processing of the moist food waste, while maintaining the required texture and composition for the larval rearing. The prepared mixture was then left at room temperature for approximately 36–48 h. This step was important for facilitating the digestion process and for improving the metabolic activity of the BSF larvae.
In this study, three different types of substrates were prepared. The first meal was prepared as a control substrate, with 100% of poultry feed (100% poultry feed [PF]), the second meal was 100% of the LF from the Bukhari restaurants (100% leftover food [LF]), and the third meal was a mixture of 50% LF and 50% PF. All of the substrates were mixed with water to obtain a moisture content of approximately 65% to ensure that there were optimal conditions for rearing the BSF larvae. The samples of each substrate were collected and refrigerated at 4 °C until further analysis. Nutritional profiles, such as crude protein, crude fiber, crude fat, ash, and moisture percentage, were determined, using the standard procedures outlined by the Association of Official Analytical Chemists [Association of Official Analytical Chemists (AOAC), 2005].
2.3 Experimental design
Eggs were collected from the egg traps that were set for 10 h from 3 am to 1 pm, to standardize the oviposition timing. The egg masses weighing 1 g each were placed on stainless steel mesh plates (6 × 6 cm) and positioned above 250 g of poultry feed within three small breeding boxes (39.5 × 29.5 × 24 cm) made of thick polypropylene with ventilation holes.
The experiment was designed to evaluate the effects of the three different substrates: (1) 100% poultry feed (100% PF) as a control; (2) 100% LF from Bukhari restaurants (100% LF); and (3) 50% poultry feed and 50% Bukhari restaurant LF (50% PF and 50% LF). Each substrate treatment included two types of containers, each with three replicates to ensure statistical robustness: (i) plastic rearing boxes (60 × 40 × 10 cm), which were used for chemical composition and amino acids analyses, and (ii) plastic measurement bowls (14 L; approximately 42 cm diameter × 10 cm depth), which were used to monitor survival and growth parameters. Both container types were provided with sufficient ventilation to prevent overheating.
On day five after hatching, larvae from the three small breeding boxes were transferred into the rearing boxes at a density of 10 larvae/cm2 (approximately 24,000 larvae per box). On day six, about 600 larvae (6-day-old) of the same size were collected from the rearing boxes and gently sieved through a stainless-steel laboratory sieve (4 mm mesh size) to ensure consistency across three replicates. A group of 200 larvae was then transferred into measurement bowls filled with 200 g of each respective substrate. The bowls of all replicates were covered with muslin cloth to prevent predation by other insects. Throughout the experiment, both the moisture content and the quantity in each substrate were monitored continuously.
In order to evaluate the larval growth, 10 larvae were randomly selected from each measurement bowl replicate (total 30 larvae per substrate) and subjected to measurements in terms of their length, width, and weight at four different larval ages throughout the experiment, on days six, 12, and 18, and on the day when pigmentation appeared on day 21. The larvae were then returned to their experimental group after each measurement. At the same time points (days 12, 18, and 21), the survival rate was determined by counting the number of live larvae remaining in each replicate and dividing this by the initial number of larvae introduced on day 6 (200 larvae), expressed as a percentage. The mean value of the three replicates was calculated to represent the survival rate for each treatment.
On day 12, 100 g of substrate was only added to the measurement bowls, each containing 200 larvae, to sustain sufficient food quantities for all of the experimental durations. For the rearing boxes, 200 g of substrate was added on day 12, followed by 250 g on both days 14 and 16. These increments corresponded to an estimated feeding rate of 0.93 g/larvae/day, with a total average of 13 g per larvae provided over the 21-day period, adjusted according to the number of larvae in each replicate type. The substrate humidity was set initially at 65% and gradually reduced to approximately 50% by day 18, aiming for fertilizer production by the end of the experiment.
By the 20th day, pigmentation indicative of pre-pupal development began to appear in some of the larvae, prompting their harvest. Before collection, the larvae were subjected to a 24 h starvation period to allow for gut clearance. Following starvation, the by-products of both molting and digestion were separated from the larvae using a standard stainless-steel laboratory sieve with small holes (4 mm diameter) and they were subsequently classified as organic fertilizer.
The larvae were sterilized by immersing them in hot water at a temperature of 70–75 °C with 5% salt to remove any surface impurities. They were then placed in perforated stainless-steel pan trays with holes (60 × 80 × 4.5 cm, 4 mm diameter) and dried in an oven at 70 °C for 18 h, producing golden larvae. The dried larvae and residual frass were collected and analyzed to determine their nutritional profile, including the crude protein, crude fiber, crude fat, ash, and moisture percentage, as well as the amino acid concentrations.
2.4 Larvae measurements
For purposes of larval development, 10 BSF larvae were selected randomly from each measurement bowl replicate at four different larval ages on days six, 12, 18, and 20, before the starvation stage at the end of the experiment. These were weighed using the scale balance (EAB 124e, ae ADAM) to determine their mean weight. The length and width were measured using digital calipers to calculate the means. The body length/width ratio was calculated using the formula . The body mass index (BMI) was calculated by BMI . The randomly selected larvae were then returned to their respective experimental groups after each measurement.
2.5 Chemical profile analysis
The approximate chemical composition of the substrates, larvae, and frass were analyzed in triplicate using the standard methods outlined by Association of Official Analytical Chemists (AOAC), 2005. Samples for chemical composition analysis were taken from the rearing box replicates, which were maintained to provide sufficient biomass for these analyses. All of the analyses were conducted as part of a collaborative service that provided a specialized laboratory in Egypt (Regional Center for Food & Feed [RCFF], Cairo University, Al Giza, Giza Governorate and Soil, Water & Environment Research Institute [SWERI], Cairo University, Al Giza, Giza Governorate).
2.6 Mineral profile analysis
The mineral profile analysis was conducted using ICP-OES techniques in the same abovementioned laboratory. Samples for mineral analysis were taken from the rearing box replicates, which were maintained to provide sufficient biomass for these analyses. The samples were prepared via microwave-assisted digestion, following established protocols, and the trace element concentrations were measured using CETAC U5000AT and Perkin Elmer Optima 2100 DV instruments.
2.7 Amino acid profile analysis
The amino acid profiles of the samples were analyzed at in the same abovementioned laboratory, using High-Performance Liquid Chromatography (HPLC). Samples for amino acid analysis were taken from the rearing box replicates, which were maintained to provide sufficient biomass for these analyses. The analysis involved hydrolyzing the samples with 6N HCI to release the free amino acids, followed by derivatization with phenyl isothiocyanate (PITC) for HPLC detection. The HPLC system was calibrated with certified standards to ensure accuracy, and each sample was analyzed in triplicate to maintain precision. The results provided a detailed profile of the essential and non-essential amino acids in the larvae and the frass, providing valuable insights into their nutritional quality.
2.8 Statistical analysis
All analyses were performed using IBM Corporation's SPSS version 22 (Armonk, NY, USA). The data was subjected to Shapiro–Wilk test to determine the normality of value distributions. Then, Kruskal Wallis test followed by Mann Whitney tests utilized between groups as data were abnormally distributed, with data presented as the mean ± standard deviation (SD). This involved comparing the average values from three replicates for larvae and frass with the corresponding mineral contents in the substrate (n = 3). Statistical significance was set at P <0.05, while the p-values between 0.05 and 0.1 were considered.
3 Results
3.1 Chemical composition, mineral profile and heavy metals of substrates
3.1.1 Chemical composition
The data revealed that there were significant variations in the chemical composition and mineral profile of the three substrates (Table 1). The average moisture content was significantly highest in the LF 100% and followed by the LF 50% substrate group, with the lowest value recorded in the PF 100% (Table 1). The highest cured protein content was observed in both the control group (PF 100%) and the LF 50%, followed by the LF 100% substrate group (P = 0.050; Table 1). The lipid and crude fiber contents were significantly higher in the LF 100% substrate and followed by the LF 50% substrate group, with the lowest value recorded in the PF 100% (Table 1). While no significant differences found in the carbohydrate among the three substrate groups (P = 0.296; Table 1). The ash content was highest in the PF 100 and the LF 50%, which were significantly higher than that of the LF 100% substrate group (P = 0.011; Table 1).
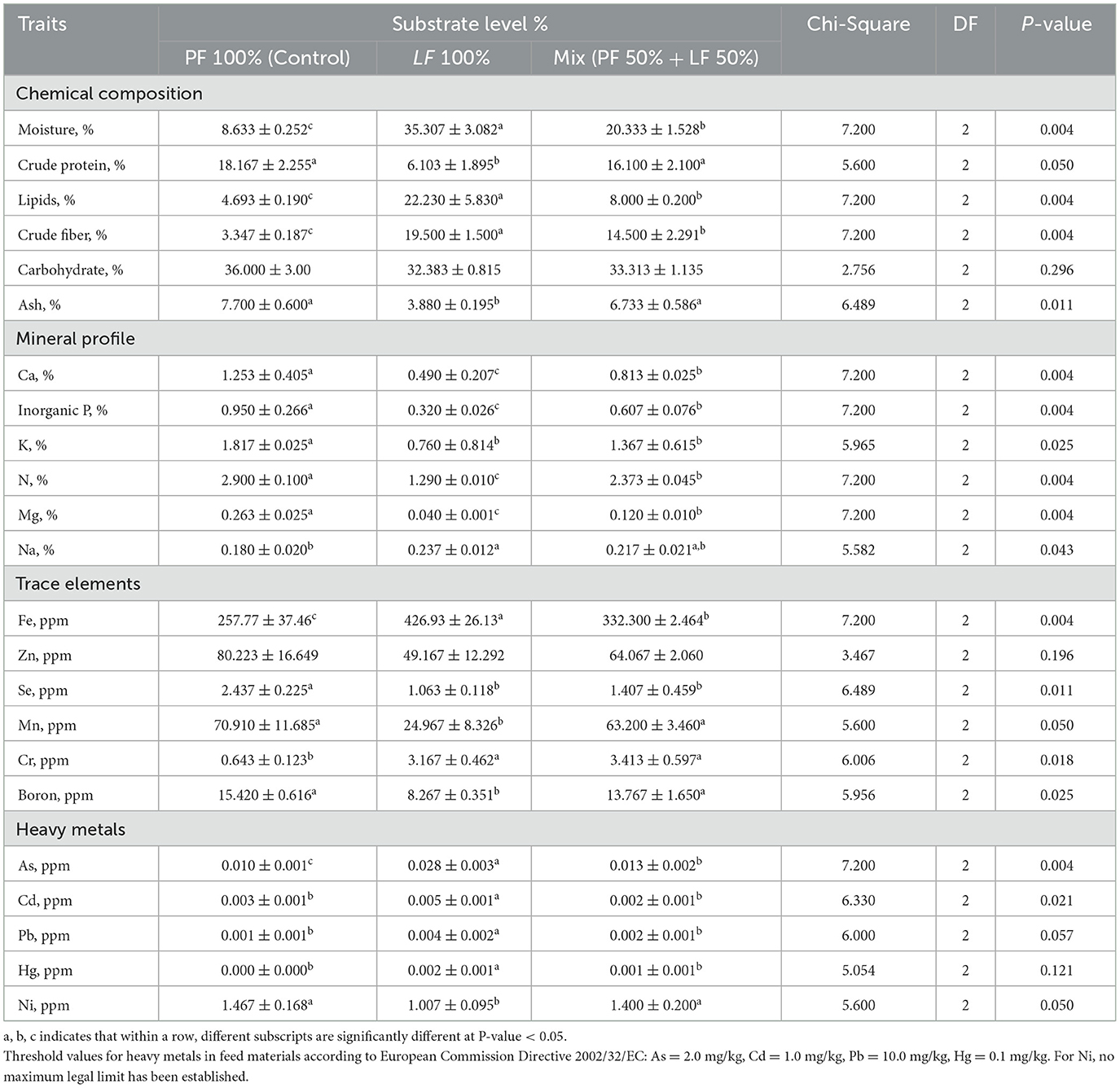
Table 1. Chemical composition and mineral profiles of three substrates used for rearing black soldier fly larvae during the experimental period, all values are expressed on dray matter (DM) basis.
3.1.2 Mineral profile
The mineral composition showed significant differences among the substrates. The calcium (Ca), the inorganic phosphorus (P), the nitrogen (N) and the magnesium (Mg) levels were significantly higher in the control group (PF 100%) and followed by the LF 50% substrate group, with the lowest value recorded in the LF 100% substrate group (P = 0.004, Table 1). The Potassium (K) levels were significantly higher in the PF 100% substrate group, with significant reductions in the LF 100% and the LF 50% substrates (P = 0.025; Table 1). The Sodium (Na) value was significantly higher in both substrates LF 100% group (0.237% ± 0.012) and in the LF 50% group (0.217% ± 0.021) than in the PF 100% substrate group (0.180% ± 0.020), with no significant difference observed between the LF 100% and the LF 50% groups (P = 0.043, Table 1).
3.1.3 Trace elements
Regarding trace elements, significant differences were observed across different substrates. The ferrous (Fe) values were significantly higher in the LF 100% substrate (426 ppm ± 26.127), compared with both the PF 100% substrate and LF 50% substrate (257.767 ppm ± 37.464 and 332.300 ppm ± 2.464, respectively; P = 0.004, Table 1), with significant difference observed between them. The zinc (Zn) levels were not significant difference observed across three different substrates (P = 0.196, Table 1). Manganese (Mn) and Boron (B) concentrations were significantly lowest in the LF 100% substrate group (24.967 ppm ± 8.326 and 8.267 ppm ± 0.351; P = 0.050 and 0.025, respectively; Table 1), compared with the other substrates, with no significant differences between the PF 100% and LF 50% substrates. The selenium (Se) content was significant lower in both the LF 100% substrate and the LF 50% substrate than the PF 100% substrate (P = 0.011, Table 1). Whereas the chromium (Cr) levels were significant higher in the LF 100% substrate and the LF 50% substrate than the PF 100% substrate (P = 0.018, Table 1).
3.1.4 Heavy metals
The heavy metal concentrations varied significantly among the different substrates. The levels of arsenic (As) observed were the highest in the LF 100% substrate, followed by the LF 50% substrate, with the lowest value recorded in the PF 100% substrate (P = 0.004, Table 1). The lead (Pb), the mercury (Hg), and the cadmium (Cd) were the highest in the LF 100% substrate, whereas no significant differences were observed between the PF 100% and LF 50% substrates. The nickel (Ni) value was the highest in both the PF 100% and the LF 50% (1.467 ppm ± 0.168 and 1.400 ppm ±0.200), followed by the LF 100% substrate (1.007 ppm ±0.095; P = 0.050; Table 1).
3.2 Amino acid composition of substrates
The essential and non-essential amino acid compositions showed significant differences across the different substrates. Their ratio was significantly higher in the PF 100% substrate (0.625 ± 0.006) than in the other substrates (P = 0.050, Table 2). The essential amino acid values as methionine, phenylalanine, isoleucine, and leucine were significantly higher in the PF 100%, followed by the LF 50% substrate, with the lowest value in the LF 100% substrate (P < 0.05). While histidine content was the highest in the LF 100%, followed by the LF 50% substrate, with the lowest content in the PF 100% substrate (P = 0.004, Table 2). The total essential amino acid values were significantly higher in the control group (PF 100%) at 7.743 ± 0.620 and followed by the LF 50% substrate (3.057 ± 0.160), with the lowest value recorded at 2.813 ± 0.059 in the LF 100% substrate (P = 0.004, Table 2). The highest levels of total non-essential amino acids were observed in the PF 100% substrate (12.403 ± 1.080), compared to the LF 100% and LF 50% substrates (5.253 ± 0.296 and 5.543 ± 0.047, respectively; P = 0.025, Table 2). However, glycine levels did not show any significant differences across other substrates (P = 0.100, Table 2). While the PF 100% substrate had the highest percentage of essential amino acids, the LF 100% and LF 50% substrates had high the percentages of non-essential amino acids.
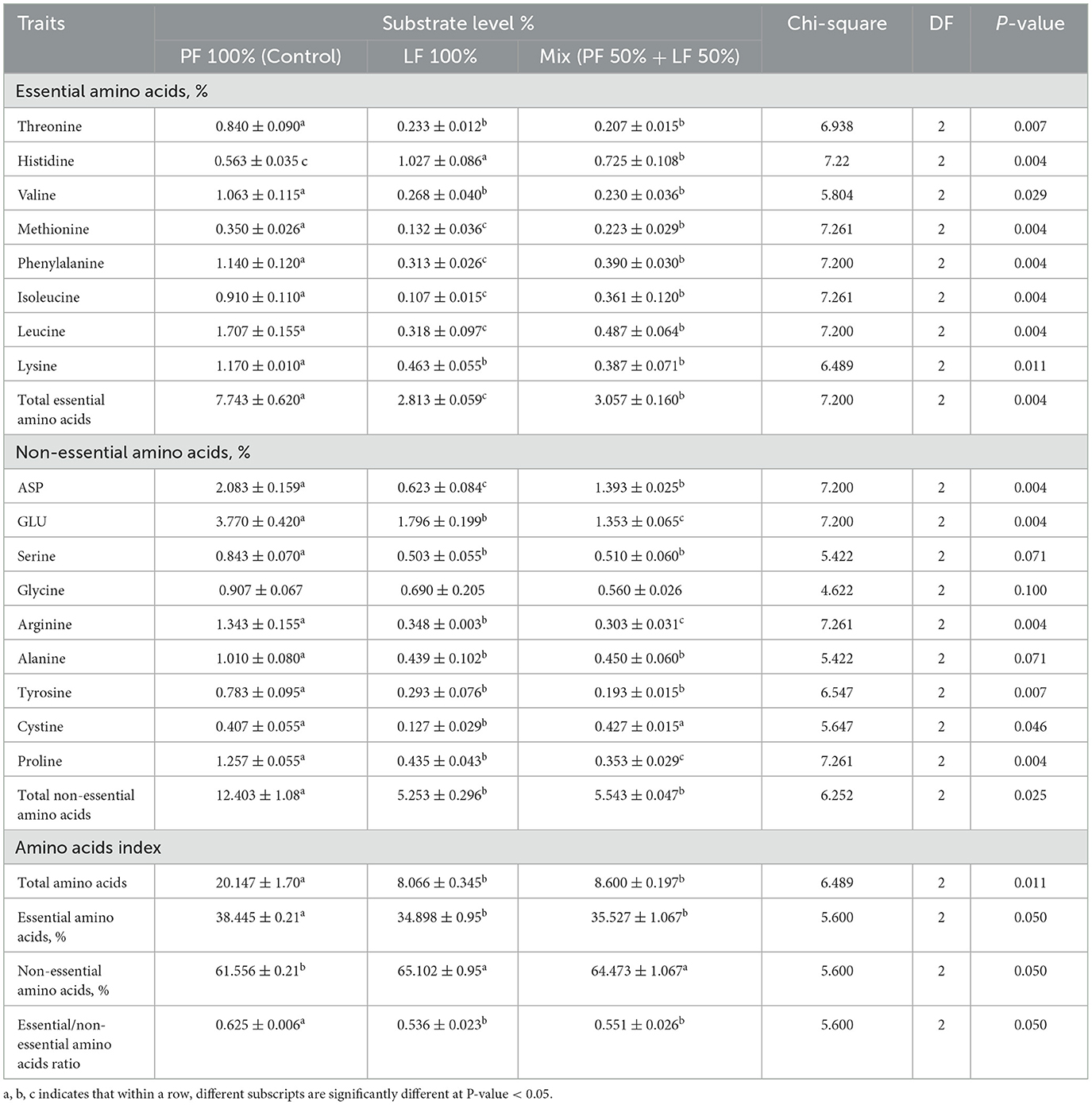
Table 2. Amino acid composition of substrates used for rearing black soldier fly larvae during the experimental period, all values are expressed on dray matter (DM) basis.
3.3 Growth performance and survival rate
While no significant differences were observed among the groups in terms of body length, width, weight, mass index, and survival rate across all the substrate types on the first day of the experiment (day six of larval age), the data revealed that substrate type had a significant impact on the growth performance and survival rate of the BSF larvae at 12, 18, and 21 days of age (Table 3).
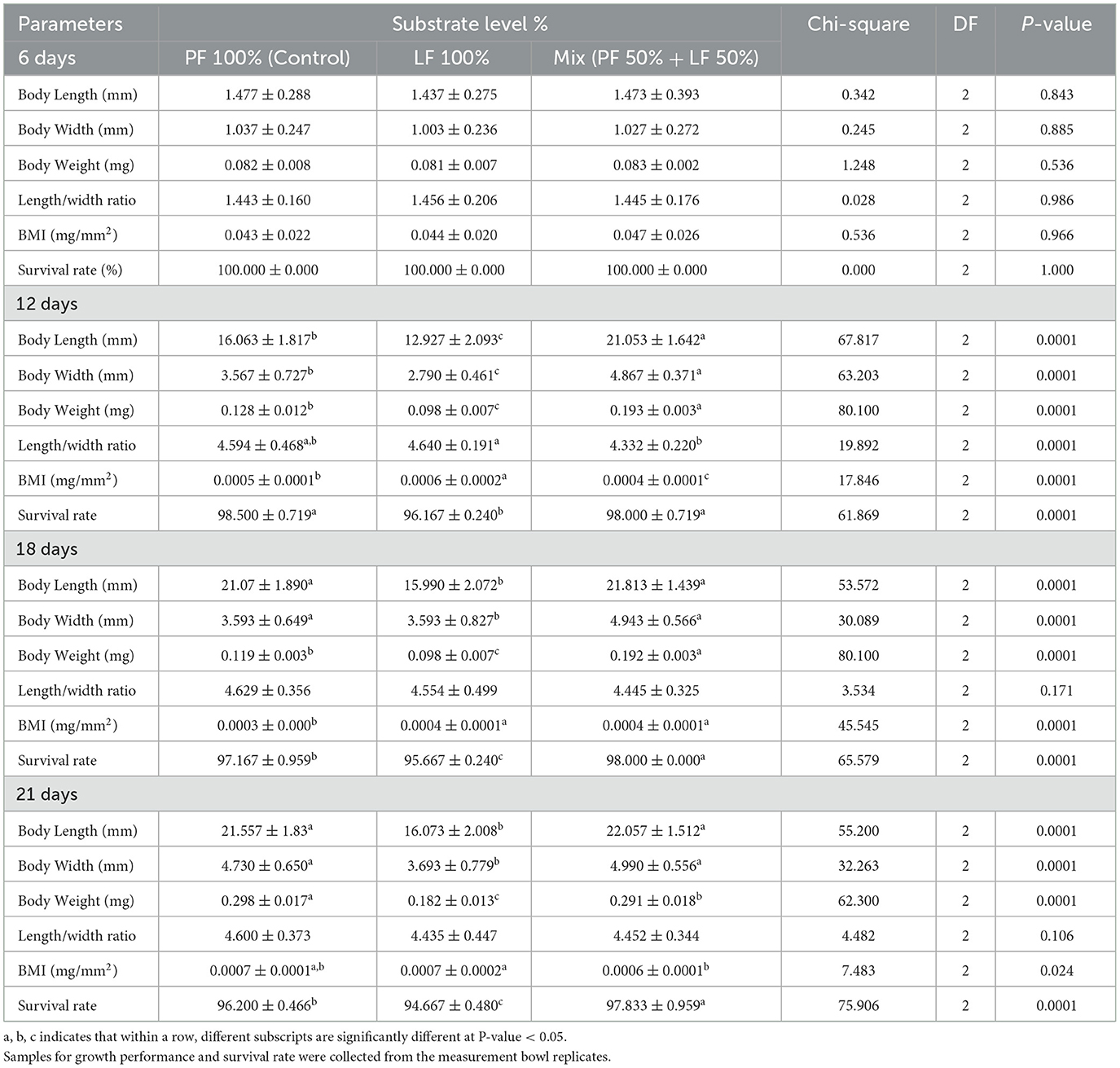
Table 3. Effect of different substrates on growth performance and survival rate of black soldier fly larvae of different ages, all values are expressed on dray matter (DM) basis.
On day 12, the mean larval length, width, weight, and body mass index across all groups provided with the different substrates differed significantly (P = 0.0001, Table 3). Larvae fed on the LF 100% showed significantly lower mean length, width, and weight than those fed on the LF 50% and the PF 100%. Whereas larvae that fed on the LF 50% recorded a significantly higher mean than those fed on the PF 100%. The ratios of length to width were significantly decrease in the LF 50% group, compared to both LF 100% and LF 50% groups (P < 0.050).
On days 18 and 21, the larvae fed on the LF 50% showed a significantly greater mean length and width than those fed on the LF 100%, with no significant differences in the larvae fed on the PF 100%. However, there were no significant differences observed in the length/weight ratios across three different groups at day 18 and 21 (P = 0.171 and 0.106, respectively; Table 3). The mean body weight was significantly lower in the LF 100% group at day 18 and 21. The BMI was significantly higher in both the LF 100% and the LF 50% groups than the PF 100% at day 18, while there were slight significant differences found in BMI between the LF 100% and LF 50% groups at day 21 (P = 0.024, Table 3). Overall, the larvae reared on LF 50% demonstrated a significantly higher growth rate than those fed on the LF 100% across all ages (P < 0.0001, Table 3).
The survival rate also differed significantly among the groups at 18 and 21 days of age, with the larvae reared on the LF 50% substrate reaching the highest survival rate of 97.83% ± 0.959 by day 21 (P < 0.0001, Table 3). In contrast, the larval group fed on the LF 100% substrate exhibited significantly lower survival rates among the different groups at 18 and 21 days of age, which may reflect the effect of lower nutritional availability in this substrate.
3.4 Chemical composition and mineral profile of BSF larvae
3.4.1 Chemical composition
The chemical composition of the BSF larvae differed significantly among the different substrates (Table 4). The moisture values were 4.667% ± 0.252, significantly higher in the larvae reared on the PF 100% group than in the LF 50% group, with no significant differences in the LF 100% group (P = 0.096; Table 4). However, no significant differences were observed in the protein content of the larvae across the three substrates (P = 0.629, Table 4). The larvae fed on the LF 100% substrate showed the highest lipid content 42.480 ± 2.035, but it was not significant in the larvae fed on the LF 50% (P = 0.086, Table 4). In contrast, the fiber and ash levels were significantly higher in the LF 100% substrate group compared with the other groups (9.510 ± 0.506 and 10.930 ± 1.309, P = 0.0029 and P = 0.004, respectively; Table 4).
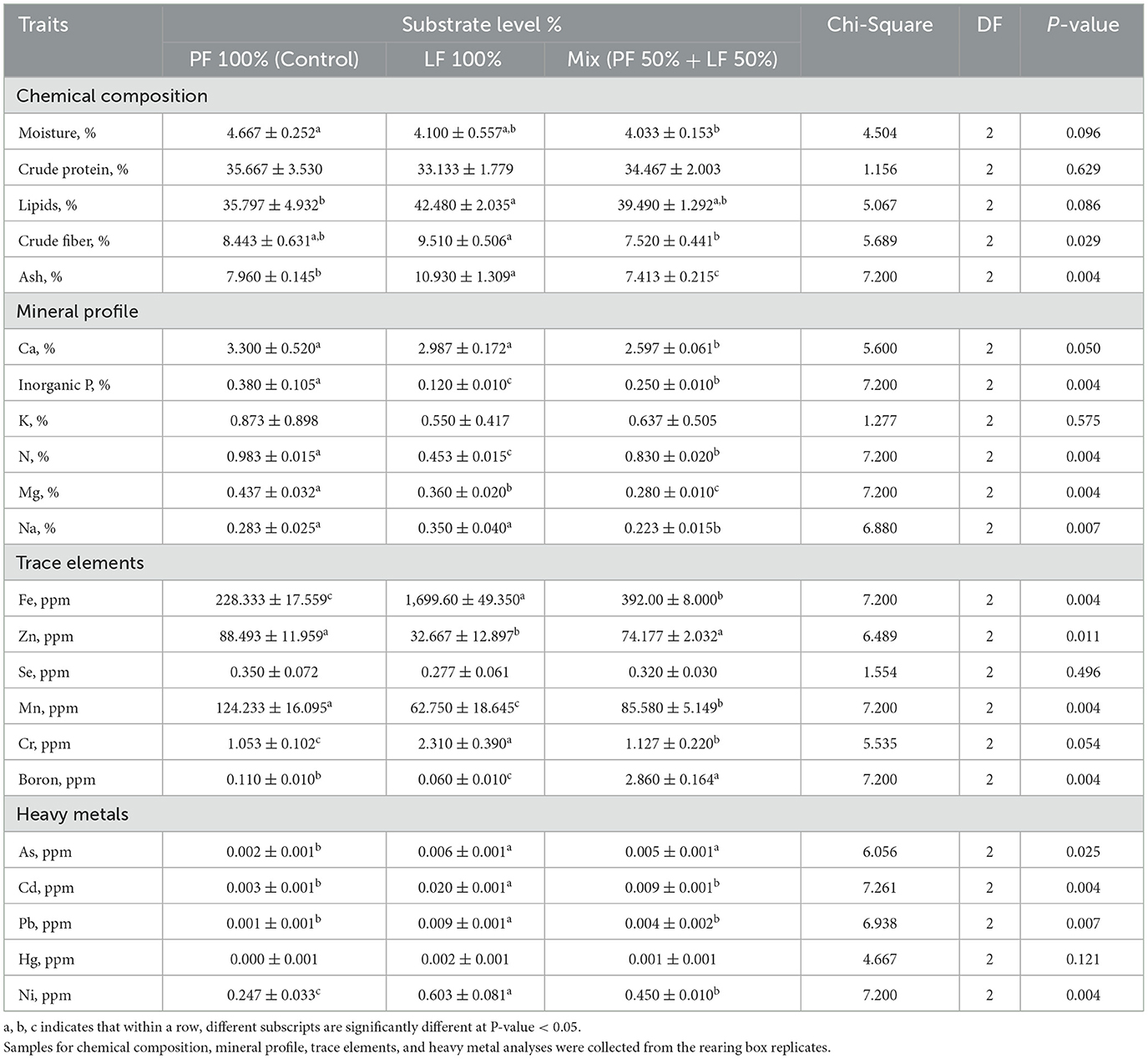
Table 4. Effect of different substrates on the chemical composition and mineral profile of black soldier fly larvae at 21 days of age, all values are expressed on dray matter (DM) basis.
3.4.2 Mineral profile
As for the mineral profile, no significant differences were observed in the K in the larvae fed different substrates (P = 0.575; Table 4). However, there were differences found in the inorganic P and N levels, with the highest concentration recorded in the larvae reared on the PF 100% substrate, followed by the LF 50% group, and the lowest in the LF 100% substrate group. Meanwhile, the Ca and Na values in the larvae fed on the PF 100% and the LF 100% substrates were affected significantly, showing the highest values, compared with the LF 50% substrate group (P = 0.05 and 0.007, respectively; Table 4). The level of Mg was significantly lowest in the LF 50% substrate (P < 0.004, Table 4).
3.4.3 Trace elements
Regarding trace elements, the Fe and Cr concentrations in larvae that fed on the LF 100% substrate were 1,699.60 ppm ± 49.350 and 2.310 ppm ± 0.390, respectively, which were significantly higher than those of the LF 50% and the LF 100% substrate groups (P = 0.004 and 0.054, respectively; Table 4). There were significant differences found in Zn levels, where the highest concentrations were significantly recorded in larvae reared on both the PF 100% and the LF 100% substrates (88.493 ppm ± 11.959 and 74.177 ppm ± 2.032, respectively), followed by the LF 50% (32.667 ppm ± 12.897, P = 0.011, Table 4). No significant differences were recorded in the levels of Se in the larvae among all substrate groups (P = 0.496, Table 4). However, significant differences were found in the Mn and B levels, with the highest Mn value in the larvae that fed on PF 100% substrate, whereas Cr was the highest in the group fed on LF 50% (P = 0.004, Table 4).
3.4.4 Heavy metals
The heavy metal accumulation in the larvae differed significantly only and clearly in Ni levels, which were the highest at 0.603 ppm ± 0.081 in the fed group (LF 100%), followed by the LF 50% group (0.450 ppm ± 0.010), and the lowest (0.247 ppm ± 0.033) in the PF 100% group (P = 0.004, Table 4). However, no significant difference was detected in the levels of Hg in the larvae across the different substrates (P = 0.121, Table 4). The values of As, Cd, and Pb in the larvae fed on the LF 100% substrate were affected significantly, showing the highest values compared with the other groups (Table 4).
3.5 Amino acid composition of BSF larvae
The percentage of essential non-essential amino acid compositions and their ratios showed no significant differences in larvae reared among the different groups. However, the type of substrate significantly influenced the total essential amino acids of BSF larvae (Table 5). The concentrations of threonine, isoleucine, and lysine were the highest in the larvae fed the PF 100% and the LF 100% substrate groups, whereas methionine levels were the highest in the larvae reared on the LF 100% substrate, reaching 0.487% ± 0.021 (P = 0.004, Table 5). No significant differences were observed in the valine levels among the larvae reared on the different substrates (P = 0.121, Table 5).
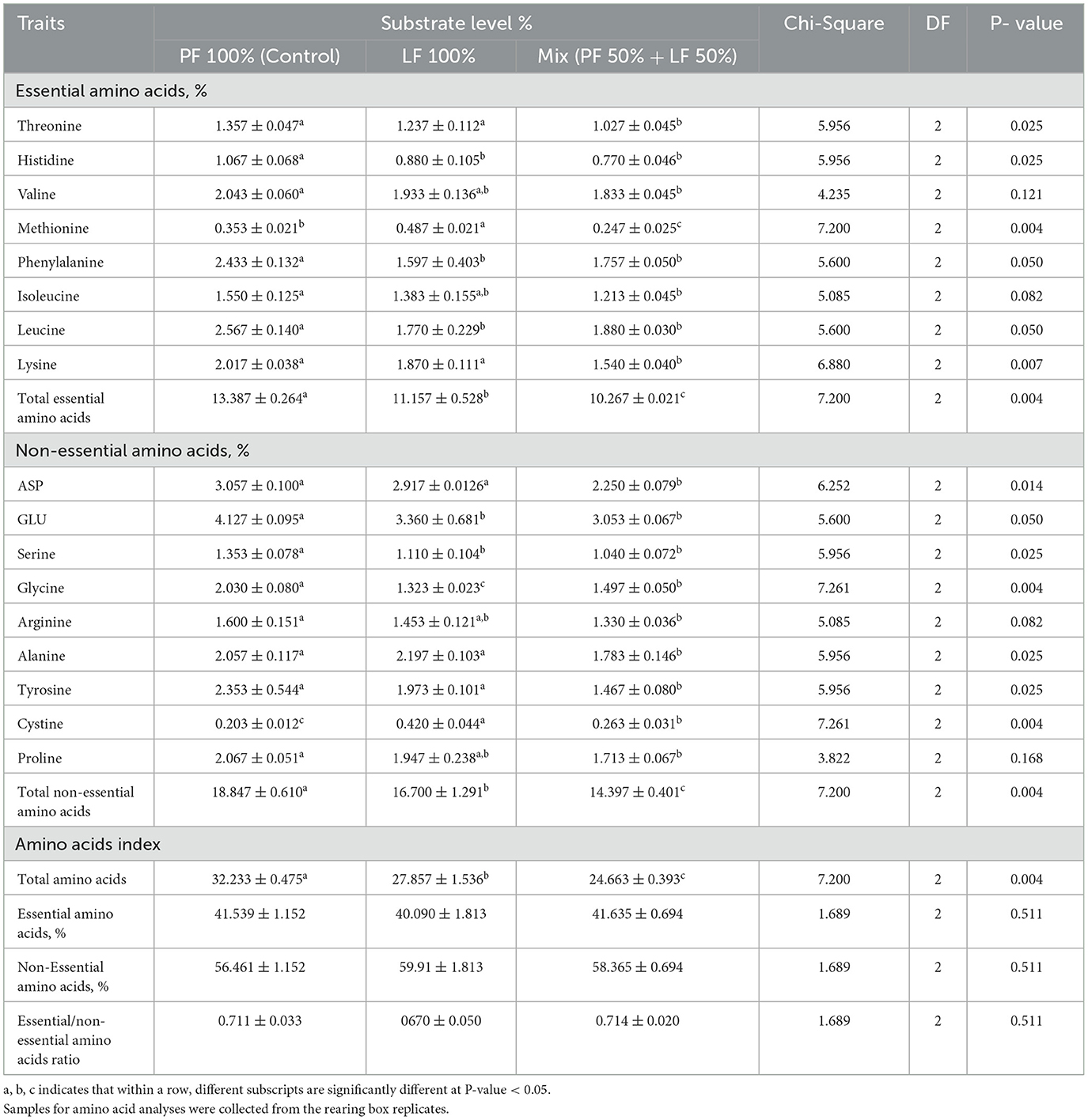
Table 5. Effect of different substrates on the amino acid composition of black soldier fly larvae at 21 days of age, all values are expressed on dray matter (DM) basis.
The concentrations of non-essential amino acids in the BSF larvae varied significantly among the different substrates (Table 5). The highest concentrations were observed in the larvae fed on the PF 100% substrate, except for cystine, which showed the significantly lowest value of 0.203 ± 0.012 in larvae fed on PF 100% (Table 5). The highest total amino acid content was found in the larvae reared on the control substrate (PF 100%), whereas the lowest content was recorded in the larvae fed on the LF 50% substrate (32.233 ± 0.475 and 24.663 ± 0.393, respectively; P = 0.004; Table 5).
3.6 Chemical composition and mineral profile in residual frass
3.6.1 Chemical composition
The data in Table 6 indicates that there were significant variations in the chemical composition and mineral profile of the residual the frass across the different substrates. The moisture value in the residual frass was significantly higher in the PF 100% substrate, which reacted at 11.80% ± 0.265 than in the LF 100% and LF 50% substrates, with values of 9.733% ± 1.002 and 8.800% ± 0.265, respectively (P = 0.003, Table 6). The lipid content in the residual frass was affected significantly across the different substrates (P < 0.0001, Table 6), reaching the highest level of 22.397% ± 1.480 in the LF 100% group, followed by the LF 50% group (9.607% ± 0.546), with the lowest levels of 3.663% ± 0.126 in the PF 100% group. The fiber values in the residual frass were significantly higher in the PF 100% and LF 100% substrate groups (P < 0.0001, Table 6). Meanwhile, ash values changed significantly in the residual frass across all of the groups, reaching the highest at 12.163% ± 0.071 the PF 100% substrate group (P < 0.0001, Table 6).
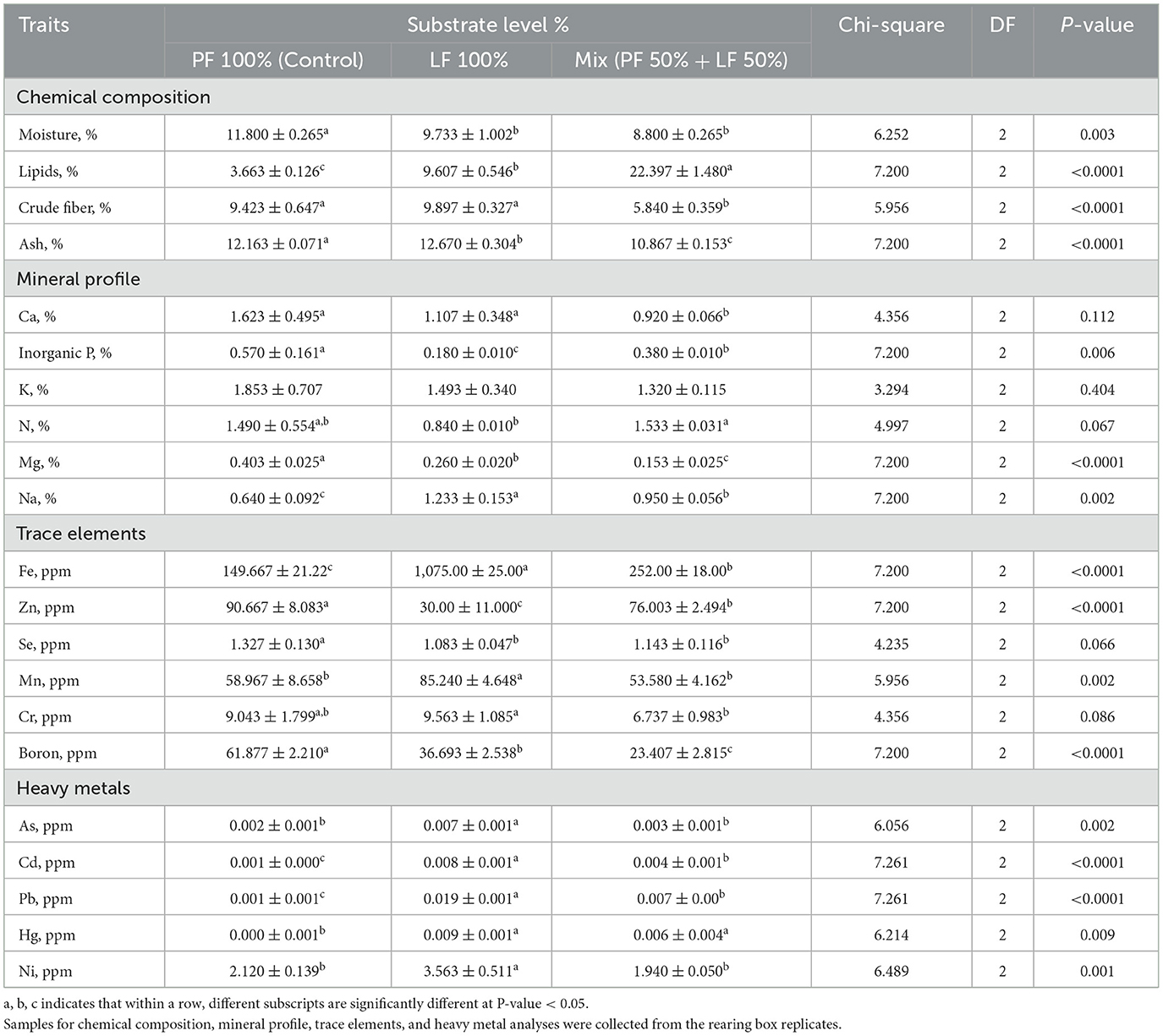
Table 6. Effect of different substrates on chemical composition and mineral profile in residual frass obtained at the end of black soldier fly larvae at 21 days of age, all values are expressed on dray matter (DM) basis.
3.6.2 Mineral profile
The mineral concentration in the residual frass differed significantly among the substrates. Although the K levels were not significantly different in the residual frass among all of the substrates (P = 0.404, Table 6), the Ca, inorganic P, and Mg levels were significantly higher in the control group (PF 100%) (Table 6). Meanwhile, the Na content in the residual frass was significantly higher in the LF 100% substrate, followed by the LF 50% substrate, with the lowest content recorded in the PF 100% substrate (P = 0.002; Table 6). The nitrogen value in the residual frass was the highest in the LF 50% and the PF 100% groups.
3.6.3 Trace elements
The trace element analysis showed that the Fe, Mn, and Cr contents in the residual frass were significantly higher in the LF 100% substrate compared with the other substrates. However, the Zn content showed the lowest value of 30.00% ± 11.00 in the LF 100% substrate (Table 6). There were no significant differences in the Se level in the residual frass between the LF 100% and the LF 50% substrates but reaching the highest level in the PF 100% group (P = 0.066, Table 6). Meanwhile, the B levels in the residual frass were the highest in the PF 100% substrate, followed by the LF 100% substrate, with the lowest levels in the LF 50% substrate.
3.6.4 Heavy metals
Finally, the heavy metal values in the residual frass differ significantly among the different substrates, with the highest values in the LF 100% substrate, followed by the LF 50% substrate (Table 6). Meanwhile the Ni was significantly higher in the LF 100% substrate (3.563 ppm ± 0.511), compared with the other groups (P = 0.001; Table 6).
3.7 Amino acid composition in residual frass
The findings in Table 7 indicate the variations in the residual frass of the BSF larvae in terms of both the essential and non-essential amino acid content across the different substrate types. The levels of essential amino acids such as histidine and valine did not differ significantly in the residual frass obtained from the larvae reared on the three different substrates. However, the methionine content was significantly higher in the frass from the larvae fed on both the LF 100% and the LF 50% substrates, whereas isoleucine, leucine, and lysine were the highest in the PF 100% substrate (Table 7). The highest levels of threonine were observed in the LF 100% substrate, compared with the PF 100% and LF 50% substrates. Phenylalanine concentration was significantly highest in the residual frass obtained from larvae fed on both the PF 100% and the LF 100% substrates, followed by the LF 50% substrate. The highest total amino acid content was found in the residual frass gained from the LF 100% and the PF 100% substrate groups (Table 7).
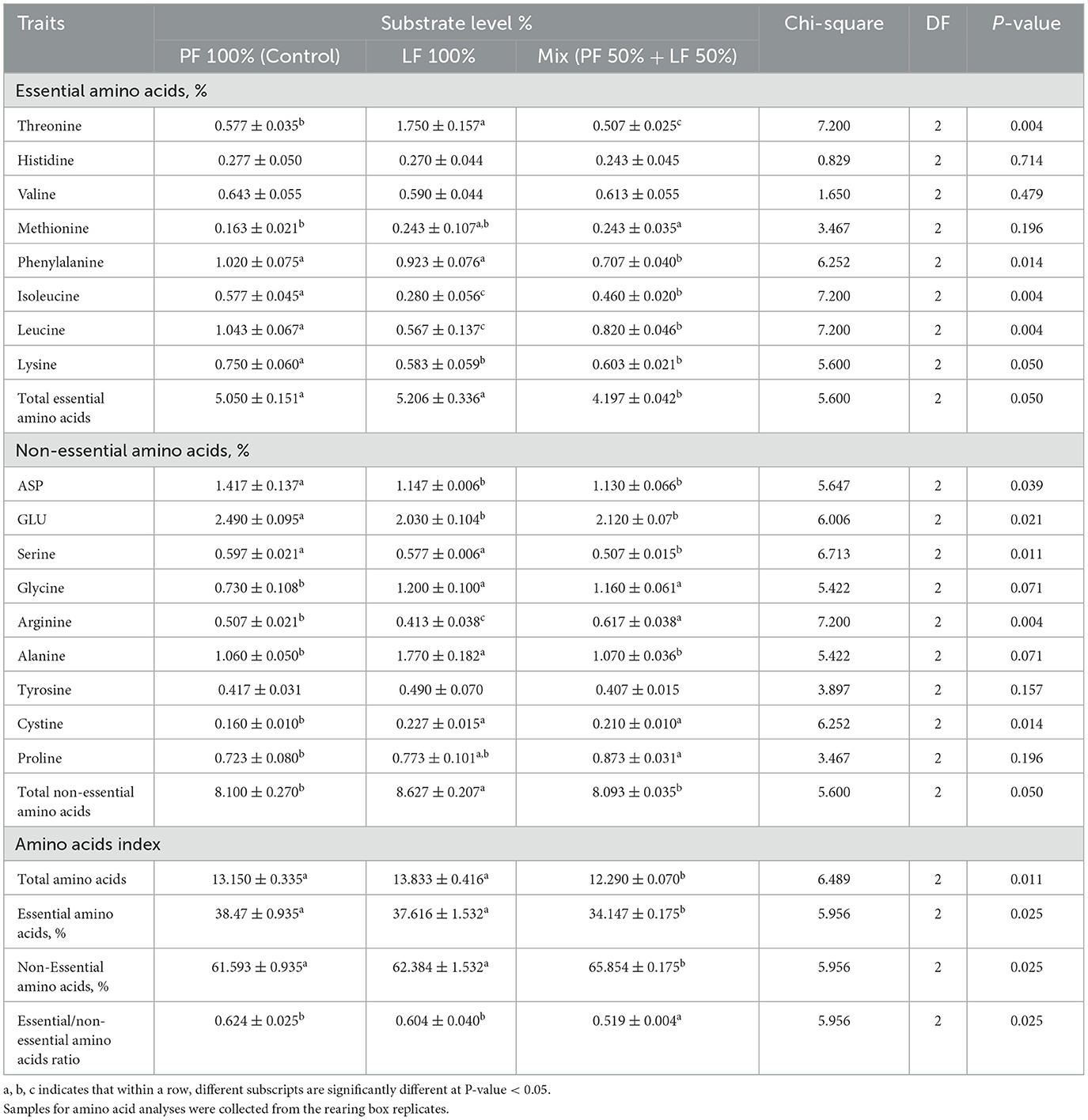
Table 7. Effect of different substrates on amino acid composition and mineral profile in residual frass obtained at the end of black soldier fly larvae at 21 days of age, all values are expressed on dray matter (DM) basis.
Among the non-essential amino acids in the residual frass obtained from the BSF larvae, significant differences were observed across the different substrates (Table 7). The composition of the ASP, GLU, and serine levels was significantly higher in the residual frass obtained from the PF 100% substrate than the other substrate groups. Meanwhile, the concentrations of glycine and cystine were significantly higher in the LF 100% group (Table 7). The proline composition was 0.773% ± 0.101 and 0.873 ± 0.031 in the residual frass from the BSF larvae fed the LF 100% and LF 50% substrate groups, respectively, with no significant differences between both of them (P = 0.196; Table 7). No significant differences were observed in tyrosine levels among the residual frass obtained from the BSF larvae fed on all of the different substrates (P = 0.157, Table 7). The highest concentrations of alanine were observed in the residual frass from the LF 100% group, except for arginine, which showed a significantly lower value of 0.413 ± 0.038 in the same group (LF 100%, Table 5).
The total amino acid content in the residual frass differed significantly across the three substrates, with the highest value of 13.833 ± 0.416 in the LF 100% and 13.150 ± 0.335 in the PF 100% substrate groups, compared with 12.290 ± 0.070 in the LF 50% substrate group (P = 0.011; Table 7). The percentage of essential amino acids was the highest in both the PF 100% and the LF 100% groups, while the percentage of non-essential amino acids was the highest in the LF 50% group. Thus, the ratio of essential to non-essential amino acids was significantly different between the LF 100% and the LF 50% substrate groups (P = 0.025, Table 7).
4 Discussion
This study demonstrates that use of BSF larvae is a promising and efficient strategy for managing food waste. In Saudi Arabia, Bukhari Restaurants are a significant source of food waste due to their substantial volume of rice-based meals. Preliminary estimates indicate that these restaurants may generate up to 406,000 tons of waste annually. The method proposed by this study offers both environmental and economic benefits due to the ease of BSF cultivation and the absence of any known pathogen transmission associated with these larvae (Choi et al., 2009). The present study, therefore provided a sustainable solution for managing the high-fat food waste generated by these restaurants. The findings illustrated the presence of a significant growth rate in BSF larvae fed with 100% LF. Furthermore, the results revealed that substrate variation affects the growth performance, survival rate, chemical composition, mineral profile, amino acid concentration, and toxic elements in BSF larvae and their residual frass. Thus, the substrate type employed plays a crucial role in larval development, nutrient accumulation, and residual frass composition, demonstrating the importance of substrate optimization for improving BSF productivity and for enhancing by-product quality. This supports the potential for efficient waste management solutions that can convert organic waste into valuable resources, such as feed and organic fertilizers (Fischer et al., 2021).
This study found that the composition of the substrate influenced the growth performance and survival rate of the BSF larvae significantly. By day 21, the larvae reared on the 50% LF substrate demonstrated the highest survival rates and optimal growth, emphasizing the importance of selecting an appropriate substrate in order to optimize BSF biomass production. This suggests that mixed substrates may enhance larval growth, potentially due to greater nutrient diversity. This growth rate may be attributed to a moderate level of lipids, proteins, and carbohydrates in the 50% LF substrate, which likely contribute to increased body weight and size. It was noted by previous researchers that an increase in body size is associated with higher amounts of carbohydrate and especially a higher protein content in the larval diet (Sentinella et al., 2013). Moreover, lipid content plays a crucial role in larval growth, with moderate lipid levels supporting maximum weight gain, whereas a significant decline in larval weight gain occurs when the lipid content of the substrate is too low (Kieβling et al., 2023; Addeo et al., 2021). These findings provided valuable insights for enhancing the efficiency of waste-to-protein conversion in insect farming systems. Therefore, the selection of a nutrient-rich substrate is fundamental to BSF larval production, as higher protein feed engenders a greater biomass yield and impacts larval development significantly (Addeo et al., 2021; Somarny et al., 2023). In the current study, it was noticed that the fat content of BSF larvae was influenced by the nutrient composition of the substrate utilized. Previous studies have revealed that larvae reared on lipid-rich substrates tend to exhibit fatty acid profiles that reflect the composition of the ingested feed, further confirming the strong link between substrate quality and larval fat (Ewald, 2019). According to (Barragan-Fonseca et al., 2018), larval crude fat content increases with higher nutrient concentrations in substrate and larval densities, and the findings of their study suggested that while larval protein levels remain relatively stable within narrow limits, crude fat content is more susceptible to variations in nutrient concentration and population density. These observations were consistent with the findings of the present study, in which the substrate composition and lipid content played a significant role in determining the fat accumulation of the BSF larvae. In particular, the larvae reared on the LF 100% accumulated higher fat levels, which can be explained by the fact that foods rich in fatty foods and containing fat may reduce the efficiency of protein yield for feed purposes, it also highlights the potential of such larvae as a source of high-energy feed ingredients or as raw material for biodiesel production. However, several studies have reported that too much dietary fat may negatively affect larval performance. For instance, (Knudsen et al., 2021; Kröncke et al., 2023) observed that high-fat substrates reduced specific growth rates and either increased mortality or lowered weight gain, indicating a trade-off between fat accumulation and overall larval fitness. These reports align with the results of the present study, where larvae reared on the LF %100 showed significantly lower survival rates and body weight by day 21. Thus, the implications of high fat content extend beyond larval growth performance, offering alternative valorization pathways in bath sectors of feed and bioenergy. The fatty acid composition is critical for assessing nutritional quality, future studies should include detailed about a fatty acid profile of the larvae to better evaluate the suitability of BSF larvae as a feed ingredient for animals. Similarly, proteins and fats are important for meeting the requirements to support the metabolic processes and to promote growth, whereas minerals are essential for bone development, enzyme activation, and eggshell formation in laying hens (Attia et al., 2022).
In terms of amino acids, the total amino acid composition was higher in the larvae than in their frass, reflecting the fact that the protein content in the larvae ranged from 31.50% to 35%. The total amino acid content of the larvae harvested varied significantly across the different substrates. When formulating diets for aquatic and terrestrial animals, it is crucial to consider not only the amount but also the quality of the protein in the BSF larvae. This is significant because methionine and lysine often limit the nutritional value of animal diets (Attia et al., 2022; Scieuzo et al., 2023; Saviane et al., 2021; Candian et al., 2023; Bosch et al., 2024; Kim et al., 2025; Carroll et al., 2023; Wedwitschka et al., 2023; Mohan et al., 2023; St-Hilaire et al., 2007; Nguyen et al., 2015; Tinder et al., 2017; Clissold and Simpson, 2015; Van Huis, 2020; Meneguz et al., 2018; Oonincx et al., 2015; Proc et al., 2020; Onsongo et al., 2018; Barragan-Fonseca et al., 2017; Purschke et al., 2017; Boafo et al., 2023; Addeo et al., 2024; Diener et al., 2015; Booth and Sheppard, 1984; Association of Official Analytical Chemists (AOAC), 2005; Choi et al., 2009; Fischer et al., 2021). Previous research indicated that substrate could affect these limiting amino acids, although the variations are typically minor but significant. For instance, slight differences were observed in the amino acid profiles of BSF larvae fed chicken feed, vegetable waste, biogas digestate, or restaurant waste (Spranghers et al., 2017). Similarly, certain notable differences were identified in BSF larvae fed various waste products, although these differences were also numerically small (Fischer et al., 2021; Lalander et al., 2019; Fischer and Romano, 2021). Amino acids play crucial roles in the metabolic processes, and are essential for the health, development, and reproduction of organisms (Pagliaro et al., 2022). They optimize nutrient absorption and transport, contribute to vitamin biosynthesis, stimulate growth, and increase tolerance to environmental stress (Mosa et al., 2021).
Regarding the residual frass in the present study, the amino acid composition presented alanine with a concentration ranging from 1.060 to 1.770%, with the highest concentration in the LF 100% group and no significant differences recorded between the other groups, whereas it was observed at 0.88% in the by-product of the fish oil extraction, which is called AnchoisFert (Pagliaro et al., 2022), a novel organic fertilizer produced as a by-product in a single process that combines fish oil extraction with limonene from leftover anchovy filets (Muscolo et al., 2022). Alanine constitutes a vital amino acid and accumulates as a general stress response molecule that protects plants from extreme temperatures, hypoxia, drought, and chemical and biological stresses (Parthasarathy et al., 2019). Moreover, proline plays a crucial role in aiding plant recovery from stress, thus accumulating proline in various plant species in response to environmental stress (Szabados and Savouré, 2010). The present study revealed excellent results in terms of high concentrations of threonine, aspartic acid, methionine, and phenylalanine. These findings highlighted the significance of threonine, as it plays an essential role in plant growth, seed development, metabolism, and gene expression (Ghazanfari et al., 2024), and aspartic acid also helps plants to endure adverse environmental conditions, such as drought, salinity, and heavy metal toxicity (Alfosea-Simón et al., 2021). Methionine also regulates plant evolutionary processes and plays a pivotal role in metabolic ecosystem responses, helping plants to adapt to different environmental conditions (Alfosea-Simón et al., 2020). Meanwhile, phenylalanine plays an important role in plant growth and development, especially in roots (Alfosea-Simón et al., 2020). Black soldier fly frass has also been shown to improve soil fertility and crop productivity in diverse settings, from field and greenhouse to soilless systems (Anedo et al., 2025). When applied frass in limited and known amounts, leading to increase nitrogen availability, improve both levels of phosphorus and potassium, and enhance biological activity (Boudabbous et al., 2023). In addition to serving as a direct source of organic nitrogen, it is important for crop production. For example, potatoes increased by 34–72%, while tomatoes, kale, and beans yielded 2.4–5.4 times more than with standard fertilizers (Anedo et al., 2025). Also, lettuce maintained higher CO2 assimilation and achieved yields up to 30.4 g under drought stress (Sawińska et al., 2024). Although the amino acid effect was not isolated, the natural amino acid content of frass is inferred to enhance nutrient efficiency, crude protein in crops, and overall plant nutrition (Anyega et al., 2021). This suggests that BSF frass could be applied not only as a nutrient input but also as a functional soil amendment capable of improving crop resilience and productivity under challenging environmental conditions. Hence, the amino acid composition found in the residual frass of larvae emphasizes their critical role in plant health, growth, and adaptation to environmental stresses. The results of the current study highlighted the potential of BSF larvae as a valuable organic fertilizer for enhancing agricultural sustainability and promoting plant resilience under various environmental conditions.
It is vital to consider macro- and micro-minerals for utilizing insect products in humans, animals, and plants to meet their requirements. Additionally, heavy metals are considered the main source of contamination for food supplements and have been shown to be a serious environmental issue (Korish and Attia, 2020; Mehar et al., 2023). Chronic exposure to toxic elements poses serious health risks to both humans and animals, potentially causing nephrotoxicity, neurotoxicity, hepatotoxicity, and dermatological and cardiovascular disorders (Mitra et al., 2022). Furthermore, the accumulation of toxic elements in residual frass used as a soil amendment may be facilitated. Their entry into the food chain increases the risk of prolonged exposure in humans and exerts direct adverse effects on plant physiology. Heavy metal toxicity is recognized as a critical abiotic stressor in plants that affects their growth, development, and metabolic processes negatively (Saxena and Shekhawat, 2013).
In the current study, the levels of Cd in the residual frass and larvae were observed to be lower than those observed in other studies concerning edible insects (Truzzi et al., 2019; Bessa et al., 2021). This was probably due to its low concentration in larval substrates. Cadmium is recognized as an element that is highly toxic to humans and animals, with prolonged oral exposure linked to several adverse health effects on many organs, including the liver, kidney, lungs, bones, and blood, as well as being linked to the induction of cancerous growth (Huff et al., 2007). In the present study, elevated concentrations of Pb were detected in the harvested BSF larvae, despite the absence or low levels of lead in the corresponding substrates. This observation was consistent with the findings reported by (Proc et al., 2020) but raised important questions regarding the potential sources of contamination. The presence of Pb in insects is likely due to external contamination, as insects cannot biosynthesize heavy metals. This contamination can potentially occur from various sources, from the rearing equipment or process, environmental exposure, or the breeding facility itself (Meyer et al., 2021). Thus, the results of the present study underscore the necessity for accurate traceability and control of all input and materials used in BSF production systems to ensure contamination-free biomass.
The Hg measured in the harvested BSF larvae was below 1 mg/kg, indicating a negligible amount and no associated toxicological risk. Although the Hg levels were minimal in the present trial, future research should account for the substrate origin, particularly when marine- or fish-based materials are used, due to their higher propensity for Hg contamination (Bespalova and Mousavi, 2013). Moreover, arsenic accumulation was observed in the harvested larvae in all of the groups, which was related to their substrates and was consistent with the study conducted by (Biancarosa et al., 2018). Although the Ni levels were less than 0.5 mg/kg in the larvae of all three groups, their level in the frass was higher than 1.88 mg/kg. This meant that the Ni did not accumulate in the larvae (Truzzi et al., 2020). Therefore, the Ni concentration in the frass varied significantly in the range 1.94–4.42 ppm. Thus, the levels of heavy metals reported in the present study were below the maximum permissible limits established by the European Commission for animal feed materials (Directive 2002/32/EC), which specify thresholds of 2.5 mg/kg for Pb, 1.0 mg/kg for Cd, 0.1 mg/kg for Hg and 2.0 mg/kg for As. The findings indicated that the BSF larvae and frass produced by this study were within the safety limits for potential use in feed and agricultural applications.
However, macro- and micro-minerals have structural, physiological, and regulatory functions in the bodies of animals (Andrieu, 2008; Suttle, 2010). The levels of Ca and P are the most abundant in the body and are functionally related because of their roles in eggshell and bone formation (Li et al., 2017; Zotte et al., 2019). In the current study, the calcium levels in the larvae across the different groups were generally higher than those in their frass. Ca is a vital element in the body of animals (Andrieu, 2008; Suttle, 2010). Moreover, Ca, P, K, Na, and Mg varied in the larval accumulation across the three groups, depending on the substrate composition, which was consistent with the findings of (Addeo et al., 2024; Diener et al., 2015; Booth and Sheppard, 1984; Association of Official Analytical Chemists (AOAC), 2005; Choi et al., 2009; Fischer et al., 2021; Sentinella et al., 2013; Kieβling et al., 2023; Addeo et al., 2021; Somarny et al., 2023; Ewald, 2019; Barragan-Fonseca et al., 2018; Knudsen et al., 2021; Kröncke et al., 2023; Spranghers et al., 2017; Lalander et al., 2019; Fischer and Romano, 2021; Pagliaro et al., 2022; Mosa et al., 2021; Muscolo et al., 2022; Parthasarathy et al., 2019; Szabados and Savouré, 2010; Ghazanfari et al., 2024; Alfosea-Simón et al., 2021, 2020; Jiao et al., 2017; Anedo et al., 2025; Boudabbous et al., 2023; Sawińska et al., 2024; Anyega et al., 2021; Korish and Attia, 2020; Mehar et al., 2023; Mitra et al., 2022; Saxena and Shekhawat, 2013; Truzzi et al., 2019; Bessa et al., 2021; Huff et al., 2007; Meyer et al., 2021; Bespalova and Mousavi, 2013; Biancarosa et al., 2018; Truzzi et al., 2020; Andrieu, 2008; Suttle, 2010; Li et al., 2017; Zotte et al., 2019; Chia et al., 2020). However, the P, Na, Mg, and K concentrations in the larvae were lower than those in their frass. Currently, there are various commercially available BSF larvae frass products used as organic fertilizers and soil amendments because of their high nitrogen-phosphorous-potassium (NPK) content, which is similar to, or even surpasses, that of other organic fertilizers (Choi et al., 2009). The NPK balance of BSF larval frass can be manipulated by adjusting the composition of the feed and amending it with additional carbon. Optimal NPK ratios can be achieved by using a standardized diet for BSF larvae, which includes 50% wheat bran, 30% alfalfa meal, and 20% corn meal (Setti et al., 2019). Additionally, increasing the carbon-to-nitrogen ratio to 15 via sawdust addition enhances N and P retention and accelerates composting (Beesigamukama et al., 2021). The amount of phosphorus in frass promotes the development and growth of roots and improves the formation of flowers, fruits, and seeds (Vance et al., 2003). Potassium is essential for numerous biochemical and physiological functions that contribute to plant growth and development (Hasanuzzaman et al., 2018). It also plays a role in protein synthesis, carbohydrate metabolism, and enzyme activation (Wang et al., 2013). Moreover, the Mg in frass is crucial because it contributes to increasing chlorophyll production and facilitates the metabolism of phosphorus and potassium (Marschner, 2012).
In addition, some elements are essential nutrients, albeit in limited amounts, because they can act as toxins in high concentrations. For instance, Fe is stored in insect bodies via ferritin, and contributes to larval pigmentation (Janssen et al., 2019). As (Nairuti et al., 2021) revealed, BSF larvae can accumulate high levels of Fe, depending on its concentration in the substrate. They observed high Fe concentrations in larvae in the LF 100% and LF 50% groups in their study, but the frass in these groups contained more than double the amount of Fe found in the larvae, suggesting limited Fe ingestion. These findings suggested that the accumulation of Fe in frass is influenced by the substrate source, which affects the bioavailability of Fe in BSF larvae. Meanwhile, Zn was found in elevated concentrations in the BSF larvae in their study, despite its relatively low levels in their substrate. This observation was consistent with the findings of (Diener et al., 2015), who reported that Zn uptake in insects is subject to active regulatory mechanisms. As an essential mineral for insect metabolism, Zn is tightly regulated and BSF larvae are capable of increasing Zn absorption under conditions of limited dietary availability (Böhm et al., 2022). Additionally, the presence of Zn in frass plays a crucial role in enhancing seedling growth and promoting their resistance to diseases (Alloway, 2008). Therefore, nutrients essential for fortifying plants, such as Mg, Fe, Mn, and Zn, can potentially improve their nutritional value for human consumption (Jiao et al., 2017). Meanwhile, Se plays a critical role in enzymatic function in animals and humans (Attia et al., 2010; Adadi et al., 2019), although there are limited studies concerning its accumulation in BSF larvae and their frass; therefore, further research is required to determine whether Se bioavailability varies across different substrates.
In all, three active processes can be identified that influence the concentration of certain minerals in insect frass: first, the concentration of a specific mineral, due to the reduction of water and other nutrients; second, the decrease of the mineral as it accumulates in the insect's body; and third, the increase of a specific mineral, due to its expulsion through insect excrement and exuviae (Addeo et al., 2024). The interplay of these three factors can elucidate the varying behaviors of a specific mineral in relation to its concentration in feed, larvae, and their frass.
Furthermore, several previous studies demonstrated the benefits of chitin content in BSF larvae frass for soil, highlighting its effectiveness for mitigating erosion, preventing plant diseases, and treating root sepsis (Ahmed et al., 2021). Chitin not only influences the carbon-to-nitrogen (C:N) ratio, but its degradation product, chitosan, can also offer overlooked advantages for plant health, including enhanced pathogen resistance, pest reduction, and improved plant growth (Tharanathan and Kittur, 2003; Sharp, 2013). The C:N ratio is a vital consideration when deciding whether to use frass as a soil amendment or co-substrate in processes such as anaerobic digestion or composting (Dioha et al., 2013; Wang et al., 2019). The role of chitin in insects is often associated with chitinolytic gut symbionts (Borkott and Insam, 1990), a topic that requires further exploration, particularly in the context of BSF larvae. Fertilizers containing chitin have been identified as an excellent source of nitrogen (Wang et al., 2019). Therefore, the chitin content of BSF larvae frass shows promise for pest management and for enhancing plant growth and health (Fischer et al., 2021), underscoring the need for further research in these areas to promote sustainable agricultural practices.
5 Conclusions
This study highlighted the potential of using BSF larvae as an effective and sustainable solution for managing food waste, particularly for addressing the substantial waste generated by Bukhari restaurants in Saudi Arabia. The BSF larvae efficiently transform organic waste into high-value biomass and nutrient-rich residual frass, supporting environmental sustainability and circular economic principles. The composition of the rearing substrate influences the key biological parameters of the BSF larvae significantly, including their growth performance, survival rate, proximate composition, mineral profile, amino acid content, and toxic elements, accumulation. Notably, the LF 50% substrate produced the highest larval body weight and survival rate, which was attributed to its rich lipid and protein content. These nutrients are important for supporting larval metabolism and development, with downstream implications for feed and fertilizer quality. Both larvae and residual frass contained high levels of essential amino acids and beneficial minerals, whereas the toxic elements remained below hazardous thresholds, confirming the safety of the resulting products. These findings highlighted the novel evidence on the valorization of high-fat restaurant leftover food waste, a largely underexplored substrate in the region. By demonstrating that BSF larvae can safely and efficiently process such organic waste into protein-rich feed and organic fertilizer, this work contributes new insights into sustainable waste management strategies under the Saudi Green Initiative. These results have broader implications for policy, as supporting the integration of insect bioconversion into national food waste reduction targets, and for agricultural practice, where BSF-derived products can serve as cost-effective feed alternatives and biofertilizer to enhance soil health. substrates to enhance the nutritional value and safety of BSF-derived feed and fertilizer. Overall, this study demonstrated that BSF larvae bioconversion is a viable and sustainable approach for organic waste management, with strong potential to support food security and environmental goals under the SGI. Future research should focus on the long-term field applications of BSF frass, the scalability of BSF systems using various waste streams, and the refining of feed formulations to optimize insect nutrient profiles for specific agricultural uses.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
SA: Writing – original draft, Writing – review & editing.
Funding
The author declares that financial support was received for the research and/or publication of this article. This work was funded by University of Jeddah, Jeddah, Saudi Arabia under Grand No (UJ-24-DR-22-1037-1). Therefore, the author thanks the University of Jeddah for its technical and financial support.
Acknowledgments
The author extends their appreciation to the University of Jeddah for its technical and financial support.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author declares that Gen AI was used in the creation of this manuscript. I have used AI only to check grammar, improve language and polish writing.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
BSF, Black Soldier Fly; PF 100%, Poultry Feed 100%; LF 100%, Leftovers Food from restaurants 100%; LF 50%, Leftovers Food from restaurants 50% and Poultry Feed 50%.
References
Adadi, P., Barakova, N. V., Muravyov, K. Y., and Krivoshapkina, E. F. (2019). Designing selenium functional foods and beverages: a review. Food Res. Int. 120, 708–725. doi: 10.1016/j.foodres.2018.11.029
Addeo, N. F., Scivicco, M., Vozzo, S., Bovera, F., Asiry, K. A., Alqurashi, S., et al. (2024). Mineral profile and heavy metals bioaccumulation in black soldier fly (Hermetia illucens, L.) larvae and frass across diverse organic substrates. Ital. J. Anim. Sci. 23, 179–188. doi: 10.1080/1828051X.2024.2302845
Addeo, N. F., Vozzo, S., Secci, G., Mastellone, V., Piccolo, G., Lombardi, P., et al. (2021). Different combinations of butchery and vegetable wastes on growth performance, chemical-nutritional characteristics and oxidative status of black soldier fly growing larvae. Animals 11:3515. doi: 10.3390/ani11123515
Ahmed, N. M., Al-Assie, A. H. A., and Hassan, A. A. (2021). Detection of Mn—dependent chitinase for wheat root rot disease control by real time PCR. Indian J. Forensic Med. Toxicol. 15, 2024–2035. doi: 10.37506/ijfmt.v15i4.16999
Alfosea-Simón, M., Simón-Grao, S., Zavala-Gonzalez, E. A., Cámara-Zapata, J. M., Simón, I., Martínez-Nicolás, J. J., et al. (2020). Application of biostimulants containing amino acids to tomatoes could favor sustainable cultivation: implications for tyrosine, lysine, and methionine. Sustainability 12:9729. doi: 10.3390/su12229729
Alfosea-Simón, M., Simón-Grao, S., Zavala-Gonzalez, E. A., Cámara-Zapata, J. M., Simón, I., Martínez-Nicolás, J. J., et al. (2021). Physiological, nutritional and metabolomic responses of tomato plants after the foliar application of amino acids aspartic acid, glutamic acid and alanine. Front. Plant Sci. 11:581234. doi: 10.3389/fpls.2020.581234
Andrieu, S. (2008). Is there a role for organic trace element supplements in transition cow health?. Vet. J. 176, 77–83. doi: 10.1016/j.tvjl.2007.12.022
Anedo, E. O., Beesigamukama, D., Mochoge, B., Korir, N. K., Haukeland, S., Cheseto, X., et al. (2025). Unpacking the benefits of black soldier fly frass fertilizer towards nematode suppression and potato production. Front. Plant 16:1509643. doi: 10.3389/fpls.2025.1509643
Anyega, A. O., Korir, N. K., Beesigamukama, D., Changeh, G. J., Nkoba, K., Subramanian, S., et al. (2021). Black soldier fly-composted organic fertilizer enhances growth, yield, and nutrient quality of three key vegetable crops in sub-Saharan Africa. Front. Plant Sci. 12:680312. doi: 10.3389/fpls.2021.680312
Association of Official Analytical Chemists (AOAC) (2005). Association of Official Analytical Chemists, Official Methods of Analysis, 18th Edn. Gaithersburg, MD: Association of Official Analytical Chemists.
Attia, Y. A., Abdalah, A. A., Zeweil, H. S., Bovera, F., Tag El-Din, A. A., Araft, M. A., et al. (2010). Effect of inorganic or organic selenium supplementation on productive performance, egg quality and some physiological traits of dual-purpose breeding hens. Czech J. Anim. Sci. 55, 505–519. doi: 10.17221/1715-CJAS
Attia, Y. A., Rahman, M. T., Hossain, M. J., Basiouni, S., Khafaga, A. F., Shehata, A. A., et al. (2022). Poultry production and sustainability in developing countries under the COVID-19 crisis: lessons learned. Animals. 12:644. doi: 10.3390/ani12050644
Barragan-Fonseca, K. B., Dicke, M., and van Loon, J. J. A. (2018). Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia illucens). Entomol. Exp. Appl. 166, 761–770. doi: 10.1111/eea.12716
Barragan-Fonseca, K. B., Dicke, M., van Loon, J. J. A., Purschke, B., Scheibelberger, R., Axmann, S., et al. (2017). Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed-a review. J Insects Food Feed. 34, 105–120. doi: 10.3920/JIFF2016.0055
Beesigamukama, D., Mochoge, B., Korir, N. K., Fiaboe, K. M. K., Nakimbugwe, D., Khamis, F. M., et al. (2021). Low-cost technology for recycling agro-industrial waste into nutrient-rich organic fertilizer using black soldier fly. Waste Manag. 119, 183–194. doi: 10.1016/j.wasman.2020.09.043
Bespalova, Y. A., and Mousavi, A. (2013). Mercury (Hg) levels in the Caspian Sea Kilka fish: a communication. Environ. Forensics. 14, 179–181. doi: 10.1080/15275922.2013.814183
Bessa, L. W., Pieterse, E., Marais, J., Dhanani, K., and Hoffman, L. C. (2021). Food safety of consuming black soldier fly (Hermetia illucens) larvae: microbial, heavy metal and cross-reactive allergen risks. Foods 10:1934. doi: 10.3390/foods10081934
Biancarosa, I., Liland, N. S., Biemans, D., Araujo, P., Bruckner, C. G., Waagbø, R., et al. (2018). Uptake of heavy metals and arsenic in black soldier fly (Hermetia illucens) larvae grown on seaweed-enriched media. J. Sci. Food Agric. 98, 2176–2183. doi: 10.1002/jsfa.8702
Boafo, H. A., Gbemavo, D. S. J. C., Timpong-Jones, E. C., Eziah, V., Billah, M., and Chia, S. Y. Substrates most preferred for black soldier fly Hermetia illucens (L.) oviposition are not the most suitable for their larval development. J Insects Food Feed. (2023) 9, 183–192. doi: 10.3920/JIFF2022.0034.
Böhm, M., Anker, S., Mahfoud, F., Lauder, L., Filippatos, G., Ferreira, J. P., et al. (2022). Empagliflozin, irrespective of blood pressure, improves outcomes in heart failure with preserved ejection fraction: the EMPEROR-Preserved trial. Eur. Heart J. 44, 396–407. doi: 10.1093/eurheartj/ehac693
Booth, D. C., and Sheppard, C. (1984). Oviposition of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae): eggs, masses, timing, and site characteristics. Environ. Entomol. 13, 421–423. doi: 10.1093/ee/13.2.421
Borkott, H., and Insam, H. (1990). Symbiosis with bacteria enhances the use of chitin by the springtail, Folsomia candida (Collembola). Biol. Fertil. Soils. 9, 126–129. doi: 10.1007/BF00335794
Bosch, G., Loureiro, B. A., Schokker, D., Kar, S. K., Paul, A., Sluczanowski, N., et al. (2024). Black soldier fly larvae meal in an extruded food: effects on nutritional quality and health parameters in healthy adult cats. J. Insects Food Feed. 10, 1595–1606. doi: 10.1163/23524588-00001093
Boudabbous, K., Hammami, S. B. M., Toukabri, W., Bouhaouel, I., Ayed, S., Fraihi, W., et al. (2023). Black soldier fly (Hermetia illucens) larvae frass organic fertilizer improves soil quality and the productivity of durum wheat. Commun. Soil Sci. Plant Anal. 54, 2491–2507. doi: 10.1080/00103624.2023.2227208
Candian, V., Meneguz, M., and Tedeschi, R. (2023). Immune responses of the black soldier fly Hermetia illucens (L.) (Diptera: Stratiomyidae) reared on catering waste. Life. 13:213. doi: 10.3390/life13010213
Carroll, A., Fitzpatrick, M., and Hodge, S. (2023). The effects of two organic soil amendments, biochar and insect frass fertilizer, on shoot growth of cereal seedlings. Plants. 12:1071. doi: 10.3390/plants12051071
Chia, S. Y., Tanga, C. M., Osuga, I. M., Cheseto, X., Ekesi, S., Dicke, M., et al. (2020). Nutritional composition of black soldier fly larvae feeding on agro-industrial by-products. Entomol. Exp. Appl. 168, 472–481. doi: 10.1111/eea.12940
Choi, Y-. C., Choi, J-. Y., Kim, J-. G., Kim, M-. S., Kim, W-. T., Park, K-. H., et al. (2009). Potential usage of food waste as a natural fertilizer after digestion by Hermetia illucens (Diptera: Stratiomyidae). Int. J. Indus. Entomol. Biomat. 19, 171–4.
Clissold, F. J., and Simpson, S. J. (2015). Temperature, food quality and life history traits of herbivorous insects. Curr. Opin. Insect Sci. 11, 63–70. doi: 10.1016/j.cois.2015.10.011
Diener, S., Zurbrügg, C., and Tockner, K. (2015). Bioaccumulation of heavy metals in the black soldier fly, Hermetia illucens and effects on its life cycle. J. Insects Food Feed. 1, 261–70. doi: 10.3920/JIFF2015.0030
Dioha, I. J., Ikeme, C. H., Nafi'u, T., Soba, N. I., and Yusuf, M. B. S. (2013). Effect of carbon to nitrogen ratio on biogas production. Int. Res. J. Nat. Sci. 1, 1–10.
Fischer, H., and Romano, N. (2021). Fruit, vegetable, and starch mixtures on the nutritional quality of black soldier fly (Hermetia illucens) larvae and resulting frass. J. Insects Food Feed. 7, 319–28. doi: 10.3920/JIFF2020.0100
Fischer, H., Romano, N., and Sinha, A. K. (2021). Conversion of spent coffee and donuts by black soldier fly (Hermetia illucens) larvae into potential resources for animal and plant farming. Insects. 12:332. doi: 10.3390/insects12040332
Ghazanfari, S., Pakkish, Z., and Mohammadreazakhani, S. (2024). Enhancing shelf life and antioxidant capacity in nectarine fruit with threonine under low temperature. Plant Process Funct. 12, 21–28. doi: 10.22034/12.58.21
Hasanuzzaman, M., Bhuyan, M. H. M. B., Nahar, K., Hossain, M. S., Mahmud, J., Al Hossen, M. S., et al. (2018). Potassium: a vital regulator of plant responses and tolerance to abiotic stresses. Agronomy. 8:31. doi: 10.3390/agronomy8030031
Huff, J., Lunn, R. M., Waalkes, M. P., Tomatis, L., and Infante, P. F. (2007). Cadmium-induced cancers in animals and in humans. Int. J. Occup. Environ. Health. 13, 202–212. doi: 10.1179/oeh.2007.13.2.202
Janssen, R. H., Canelli, G., Sanders, M. G., Bakx, E. J., Lakemond, C. M. M., Fogliano, V., et al. (2019). Iron-polyphenol complexes cause blackening upon grinding Hermetia illucens (black soldier fly) larvae. Sci. Rep. 9:2967. doi: 10.1038/s41598-019-38923-x
Jiao, Y., Chen, Y., Ma, C., Qin, J., Nguyen, T. H. N., Liu, D., et al. (2017). Phenylalanine as a nitrogen source induces root growth and nitrogen-use efficiency in Populus × canescens. Tree Physiol. 38, 66–82. doi: 10.1093/treephys/tpx109
Kieβling, M., Franke, K., Heinz, V., and Aganovic, K. (2023). Relationship between substrate composition and larval weight: a simple growth model for black soldier fly larvae. J Insects Food Feed. 9, 1027–1036. doi: 10.3920/JIFF2022.0096
Kim, H., Song, M., Lee, J., Chang, S., Song, D. C., Jeon, K.-D., et al. (2025). Evaluation of black soldier fly larvae reared on different organic substrates on nutrient digestibility and palatability in cats. J Anim Sci Technol. 67, 477–488. doi: 10.5187/jast.2025.e13
Knudsen, A. K. S., Jespersen, E. E., Markwardt, M. J., Johansen, A. S., Ortind, A. P., Terp, M., et al. (2021). Effects of fish oil on growth kinetics and lipid accumulation in black soldier fly larvae. J Insects Food Feed. 8, 237-244 doi: 10.3920/JIFF2021.0083
Korish, M. A., and Attia, Y. A. (2020). Evaluation of heavy metal content in feed, litter, meat, meat products, liver, and table eggs of chickens. Animals. 10:727. doi: 10.3390/ani10040727
Kröncke, N., Neumeister, M., and Benning, R. M. (2023). Near-infrared reflectance spectroscopy for quantitative analysis of fat and fatty acid content in living tenebrio molitor larvae to detect the influence of substrate on larval composition. Insects. 14:114. doi: 10.3390/insects14020114
Lalander, C., Diener, S., Zurbrügg, C., and Vinnerås, B. (2019). Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens). J. Clean. Prod. 208, 211–219. doi: 10.1016/j.jclepro.2018.10.017
Li, X., Zhang, D., and Bryden, W. L. (2017). Calcium and phosphorus metabolism and nutrition of poultry: are current diets formulated in excess?. Anim Prod Sci. 57, 2304–2310. doi: 10.1071/AN17389
Marschner, H. (2012). Marschner's Mineral Nutrition of Higher Plants. Cambridge, MA: Academic press.
Mehar, S., Anam, I., Masood, Z., Alvi, S., Khan, W., Kabir, M., et al. (2023). Bioaccumulation of heavy metals in the different tissues of Mackerel scad, Decapterus macarellus (Cuvier, 1833) collected from Karachi and Gwadar Coasts of Pakistan. Saudi J. Biol. Sci. 30:103540. doi: 10.1016/j.sjbs.2022.103540
Meneguz, M., Schiavone, A., Gai, F., Dama, A., Lussiana, C., Renna, M., et al. (2018). Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 98, 5776–5784. doi: 10.1002/jsfa.9127
Meyer, A. M., Meijer, N., den Hil, E. F. H., and van der Fels-Klerx, H. J. (2021). Chemical food safety hazards of insects reared for food and feed. J. Insects Food Feed. 7, 823–832. doi: 10.3920/JIFF2020.0085
Mitra, S., Chakraborty, A. J., Tareq, A. M., Emran, T. B., Nainu, F., Khusro, A., et al. (2022). Impact of heavy metals on the environment and human health: Novel therapeutic insights to counter the toxicity. J. King Saud Univ. Sci. 34:101865. doi: 10.1016/j.jksus.2022.101865
Mohan, K., Sathishkumar, P., Rajan, D. K., Rajarajeswaran, J., and Ganesan, A. R. (2023). Black soldier fly (Hermetia illucens) larvae as potential feedstock for the biodiesel production: recent advances and challenges. Sci. Total Env. 859:160235. doi: 10.1016/j.scitotenv.2022.160235
Mosa, W. F. A., Ali, H. M., and Abdelsalam, N. R. (2021). The utilization of tryptophan and glycine amino acids as safe alternatives to chemical fertilizers in apple orchards. Environ. Sci. Pollut. Res. 28, 1983–1991. doi: 10.1007/s11356-020-10658-7
Muscolo, A., Mauriello, F., Marra, F., Calabrò, P. S., Russo, M., Ciriminna, R., et al. (2022). AnchoisFert: a new organic fertilizer from fish processing waste for sustainable agriculture. Global Challenges. 6:2100141. doi: 10.1002/gch2.202100141
Nairuti, R. N., Musyoka, S. N., Yegon, M. J., and Opiyo, M. A. (2021). Utilization of black soldier fly (Hermetia illucens Linnaeus) larvae as a protein source for fish feed—a review. Aquac Stud 22:AQUAST697. doi: 10.4194/AQUAST697
Nguyen, T. T. X., Tomberlin, J. K., and Vanlaerhoven, S. (2015). Ability of black soldier fly (Diptera: Stratiomyidae) larvae to recycle food waste. Environ. Entomol. 44, 406–410. doi: 10.1093/ee/nvv002
Onsongo, V. O., Osuga, I. M., Gachuiri, C. K., Wachira, A. M., Miano, D. M., Tanga, C. M., et al. (2018). Insects for income generation through animal feed: effect of dietary replacement of soybean and fish meal with black soldier fly meal on broiler growth and economic performance. J. Econ. Entomol. 111, 1966–1973. doi: 10.1093/jee/toy118
Oonincx, D. G. A. B., van Huis, A., and van Loon, J. J. A. (2015). Nutrient utilisation by black soldier flies fed with chicken, pig, or cow manure. J. Sci. Food Agric. 1, 131–139. doi: 10.3920/JIFF2014.0023
Pagliaro, M., Lino, C., Pizzone, D. M., Mauriello, F., Russo, M., Muscolo, A., et al. (2022). Amino acids in new organic fertilizer anchoisfert. Chem. Sel. 7:e202203665. doi: 10.1002/slct.202203665
Parthasarathy, A., Savka, M. A., and Hudson, A. O. (2019). The synthesis and role of β-alanine in plants. Front. Plant Sci. 10:921. doi: 10.3389/fpls.2019.00921
Proc, K., Bulak, P., Wiacek, D., and Bieganowski, A. (2020). Hermetia illucens exhibits bioaccumulative potential for 15 different elements—Implications for feed and food production. Sci. Total Env. 723:138125. doi: 10.1016/j.scitotenv.2020.138125
Purschke, B., Scheibelberger, R., Axmann, S., Adler, A., and Jäger, H. (2017). Impact of substrate contamination with mycotoxins, heavy metals and pesticides on the growth performance and composition of black soldier fly larvae (Hermetia illucens) for use in the feed and food value chain. Food Addit. Contam. Part A. 34, 1410–1420. doi: 10.1080/19440049.2017.1299946
Saviane, A., Tassoni, L., Naviglio, D., Lupi, D., Savoldelli, S., Bianchi, G., et al. (2021). Mechanical processing of Hermetia illucens larvae and bombyx mori pupae produces oils with antimicrobial activity. Animals 11:783. doi: 10.3390/ani11030783
Sawińska, Z., Radzikowska-Kujawska, D., Kowalczewski, P. Ł., Grzanka, M., Sobiech, Ł., Skrzypczak, G., et al. (2024). Hermetia illucens frass fertilization: a novel approach for enhancing lettuce resilience and photosynthetic efficiency under drought stress conditions. Appl Sci. 14:2386. doi: 10.3390/app14062386
Saxena, I., and Shekhawat, G. S. (2013). Nitric oxide (NO) in alleviation of heavy metal induced phytotoxicity and its role in protein nitration. Nitric Oxide. 32, 13–20. doi: 10.1016/j.niox.2013.03.004
Scieuzo, C., Franco, A., Salvia, R., Triunfo, M., Addeo, N. F., Vozzo, S., et al. (2023). Enhancement of fruit byproducts through bioconversion by Hermetia illucens (Diptera: Stratiomyidae). Insect Sci. 30, 991–1010. doi: 10.1111/1744-7917.13155
Secci, G., Dabbou, S., de Medeiros, A. C., Addeo, N. F., Atallah, E., Parisi, G., et al. (2022). Low dietary inclusion levels of Tenebrio molitor larva meal slightly modify growth performance, carcass and meat traits of Japanese quail (Coturnix japonica). J. Sci. Food Agric. 102, 6578–6585. doi: 10.1002/jsfa.12023
Sentinella, A. T., Crean, A. J., and Bonduriansky, R. (2013). Dietary protein mediates a trade-off between larval survival and the development of male secondary sexual traits. Funct. Ecol. 27, 1134–44. doi: 10.1111/1365-2435.12104
Setti, L., Francia, E., Pulvirenti, A., Gigliano, S., Zaccardelli, M., Pane, C., et al. (2019). Use of black soldier fly (Hermetia illucens (L.), Diptera: Stratiomyidae) larvae processing residue in peat-based growing media. Waste Manag. 95, 278–288. doi: 10.1016/j.wasman.2019.06.017
Sharp, R. G. A. (2013). Review of the applications of chitin and its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy 3, 757–793. doi: 10.3390/agronomy3040757
Somarny, W., Kuppusamy, G., Samat, N., and Azam-Ali, S. (2023). Performance of black soldier fly larvae (Hermetia illucens, L.) on different feed substrates. J. Insects Food Feed. 10, 345–358. doi: 10.1163/23524588-20230078
Spranghers, T., Ottoboni, M., Klootwijk, C., Ovyn, A., Deboosere, S., De Meulenaer, B., et al. (2017). Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 97, 2594–2600. doi: 10.1002/jsfa.8081
St-Hilaire, S., Cranfill, K., McGuire, M. A., Mosley, E. E., Tomberlin, J. K., Newton, L., et al. (2007). Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J. World Aquac. Soc. 38, 309–313. doi: 10.1111/j.1749-7345.2007.00101.x
Szabados, L., and Savouré, A. (2010). Proline: a multifunctional amino acid. Trends Plant Sci. 15, 89–97. doi: 10.1016/j.tplants.2009.11.009
Tharanathan, R. N., and Kittur, F. S. (2003). Chitin—the undisputed biomolecule of great potential. Crit. Rev. Food Sci. Nutr. 43, 61–87. doi: 10.1080/10408690390826455
Tinder, A. C., Puckett, R. T., Turner, N. D., Cammack, J. A., and Tomberlin, J. K. (2017). Bioconversion of sorghum and cowpea by black soldier fly (Hermetia illucens (L.)) larvae for alternative protein production. J. Insects Food Feed. 3, 121–130. doi: 10.3920/JIFF2016.0048
Truzzi, C., Annibaldi, A., Girolametti, F., Giovannini, L., Riolo, P., Ruschioni, S., et al. (2020). A chemically safe way to produce insect biomass for possible application in feed and food production. Int. J. Environ. Res. Public Health. 17:2121. doi: 10.3390/ijerph17062121
Truzzi, C., Illuminati, S., Girolametti, F., Antonucci, M., Scarponi, G., Ruschioni, S., et al. (2019). Influence of feeding substrates on the presence of toxic metals (Cd, Pb, Ni, As, Hg) in larvae of tenebrio molitor: risk assessment for human consumption. Int. J. Environ. Res. Public Health 16:4815. doi: 10.3390/ijerph16234815
Van Huis, A. (2020). Insects as food and feed, a new emerging agricultural sector: a review. J Insects Food Feed. 6, 27–44. doi: 10.3920/JIFF2019.0017
Vance, C. P., Uhde-Stone, C., and Allan, D. L. (2003). Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource. New Phytol. 157, 423–447. doi: 10.1046/j.1469-8137.2003.00695.x
Wang, L., Li, Y., Prasher, S. O., Yan, B., Ou, Y., Cui, H., et al. (2019). Organic matter, a critical factor to immobilize phosphorus, copper, and zinc during composting under various initial C/N ratios. Bioresour. Technol. 289:121745. doi: 10.1016/j.biortech.2019.121745
Wang, M., Zheng, Q., Shen, Q., and Guo, S. (2013). The critical role of potassium in plant stress response. Int. J. Mol. Sci. 14, 7370–7390. doi: 10.3390/ijms14047370
Wedwitschka, H., Gallegos Ibanez, D., and Jáquez, D. R. (2023). Biogas production from residues of industrial insect protein production from black soldier fly larvae Hermetia illucens (L.): an evaluation of different insect frass samples. Processes 11:362. doi: 10.3390/pr11020362
Keywords: bioconversion, waste valorization, black soldier fly larvae, organic fertilizer, insect protein biomass, larval frass, sustainable agriculture
Citation: Alqurashi SI (2025) Valorization of post-consumer food leftovers by black soldier fly (Hermetia illucens) bioconversion into protein feed and organic fertilizer. Front. Sustain. Food Syst. 9:1689902. doi: 10.3389/fsufs.2025.1689902
Received: 22 August 2025; Accepted: 15 October 2025;
Published: 03 November 2025.
Edited by:
Payel Ghosh, School of Bio Sciences and Technology, IndiaReviewed by:
Vincenzo Tufarelli, University of Bari Aldo Moro, ItalyFleurdeliz Maglangit, University of the Philippines Cebu College, Philippines
Copyright © 2025 Alqurashi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shatha I. Alqurashi, c2FhcXVyc2hpQHVqLmVkdS5zYQ==