 Ali Ahmad1†Zubair Aslam1*Muhammad Awais Arshad1
Ali Ahmad1†Zubair Aslam1*Muhammad Awais Arshad1 Usman Zulfiqar2*
Usman Zulfiqar2* Hossam S. El-Beltagi3*Mohammed O. Alshaharni4
Hossam S. El-Beltagi3*Mohammed O. Alshaharni4 P. V. Vara Prasad5
P. V. Vara Prasad5- 1Department of Agronomy, University of Agriculture Faisalabad, Faisalabad, Pakistan
- 2Department of Agronomy, Faculty of Agriculture and Environment, The Islamia University of Bahawalpur, Bahawalpur, Pakistan
- 3Agricultural Biotechnology Department, College of Agriculture and Food Sciences, King Faisal University, Al-Ahsa, Saudi Arabia
- 4Biology Department, College of Science, King Khalid University, Abha, Saudi Arabia
- 5Department of Agronomy, Kansas State University, Manhattan, KS, United States
Wheat is a major cereal crop whose growth and productivity are highly influenced by soil moisture availability. Vermicompost application has been proposed as a sustainable approach to mitigate the adverse impacts of drought stress. This study, conducted at the University of Agriculture, Faisalabad, during 2020–2021 and 2021–2022, investigated the effects of different vermicompost sources on agronomic, physiological, and nutrient traits of wheat under varying soil moisture regimes. The experiment included three soil moisture levels (no drought, mild drought, and severe drought), four vermicompost (VT) levels (control, 8 t ha−1 wheat straw vermicompost, 6 t ha−1 rice straw vermicompost, and 6 t ha−1 cow dung vermicompost), and two wheat cultivars: Faisalabad-08 (drought-tolerant) and Galaxy-13 (drought-sensitive). Results revealed that drought stress significantly reduced growth, yield, and physiological attributes. However, vermicompost application alleviated these effects, with cow dung vermicompost showing the greatest improvement, followed by rice straw and wheat straw vermicompost. Furthermore, Faisalabad-08 performed better than Galaxy-13 under both mild and severe drought conditions. Overall, the findings highlight that applying vermicompost, particularly from cow dung, to the soil is an effective strategy to improve wheat resilience and productivity under drought stress.
1 Introduction
Wheat (Triticum aestivum L.) belongs to the family Poaceae, subfamily Pooideae, and tribe Triticeae (Talaat and Abdel-Salam, 2024; Shahbazi et al., 2015; West et al., 1988). It is one of the most widely adapted cereal crops, capable of growing under diverse environmental conditions across the globe. Wheat is primarily a cool-season crop, requiring a minimum of 3–4 °C for growth, with an optimum around 25 °C and a maximum temperature tolerance of approximately 32 °C. Its cultivation spans a wide range of agro-climatic zones, mainly concentrated between latitudes 30°N–60°N and 27°S–40°S. Wheat thrives best on well-drained soils and can be grown from sea level up to elevations of about 4,500 m. Although optimal production is achieved in regions receiving 375–875 mm of annual precipitation, wheat is also growing in areas with rainfall ranging from 250 to 1,750 (Kheir et al., 2025; Briggle and Curtis, 1987; Kimber and Sears, 1987).
Wheat provides a wide range of essential nutrients, including starch, gluten proteins, dietary fiber, and B-group vitamins. Owing to its gluten content, wheat is preferred over other cereals for bread making. The whole grain, with its rich composition of nutrients and vitamins, helps protect against several chronic diseases such as ischemic disorders, diverticulitis, constipation, appendicitis, diabetes, and obesity (Rashtbari et al., 2020; Seimrizade et al., 2021; Kumar et al., 2011). In addition to being a major source of carbohydrates, wheat also supplies vitamin E, niacin, riboflavin, thiamine, protein, and important minerals such as Zn, Cu, Fe, Mg, and P (Siddiqui and Sarwar, 2002; Sarwar and Sattar, 2007; Ali et al., 2009).
Plant morphological, physiological, and biochemical traits are adversely affected by various stress factors, including extreme climatic conditions (elevated temperatures and drought), erratic weather, inappropriate sowing time, salinity, heavy metals, pest infestation, and limited irrigation water. Among these, drought is considered the most critical constraint, strongly associated with yield reduction (Anjum and Tanveer, 2016; Anjum et al., 2017; Hussain et al., 2018). Drought stress disrupts physiological processes and nutrient uptake efficiency. The incorporation of vermicompost (VT), however, can alleviate such effects by improving soil structure, enhancing microbial activity, and increasing nutrient availability. This, in turn, facilitates greater uptake of essential nutrients such as nitrogen, phosphorus, and potassium, which play vital roles in sustaining plant metabolism, enhancing water-use efficiency, and maintaining cellular and physiological functions under stress. Higher nutrient concentrations in leaves consequently improve plant adaptation to drought, mitigating its adverse effects on growth and yield (Bellitürk et al., 2020; Aboelsoud and Ahmed, 2020).
Water scarcity further reduces crop productivity by altering metabolic and physiological processes, including enhanced oxidative damage, reduced leaf gaseous exchange, and decreased carbon assimilation rates (Ramazanoglu, 2024; Hussain et al., 2018). It also suppresses enzymatic activities, ion uptake, and leaf development, thereby severely limiting crop productivity (Prassad et al., 2008; Farooq et al., 2009; Anjum et al., 2017; Todaka et al., 2017). Overall, drought stress is a major challenge as it negatively impacts multiple physiological processes, ultimately reducing crop growth, production, and grain yield (GY) (Prassad et al., 2008). To address this problem, the use of organic amendments such as vermicompost, along with cultivation of drought-tolerant genotypes, has been strongly recommended (Farooq et al., 2009; Ji et al., 2010). Notably, wheat genotypes exhibit differential responses to water scarcity, indicating the importance of varietal selection for drought-prone environments (Akram, 2011; Khakwani et al., 2011; Joshi et al., 2013).
Previous studies have demonstrated that vermicompost application can effectively mitigate water stress owing to high porosity, aeration, water retention, and drainage capacity (Hosseinzadeh et al., 2016; Hussain et al., 2018; Jahan et al., 2023). Rich in beneficial microorganisms, vermicompost enhances root moisture absorption, improves soil water holding capacity, and increases the availability of nutrients in the soil solution (Hosseinzadeh et al., 2016; Hrčka et al., 2025; Hussain et al., 2016). In addition, it promotes the accumulation of soluble sugars, sorbitol, betaine, organic acids, and amino acids in plants, while serving as a source of essential nutrients such as N, P, K, Ca, Zn, B, Mg, S, and Fe (Hosseinzadeh et al., 2016; Farooqi et al., 2024; Hafez et al., 2020). Although biofertilizers can also be produced through thermophilic composting, vermicomposting is considered a faster and more efficient method for producing nutrient-rich amendments (Atiyeh et al., 2000). The steady release of organic nutrients from vermicompost supports the beneficial soil microorganisms that contribute to nutrient cycling, nitrogen fixation, and disease suppression (Aslam et al., 2021). Over time, this reduces reliance on chemical fertilizers by providing a sustainable, slow-release source of essential nutrients. Complementary practices such as crop rotation, incorporation of green manures, application of biofertilizers (Ahmad et al., 2024; Aslam et al., 2025), and conservation tillage further enrich soil fertility and improve soil health. Collectively, these practices improve soil water-holding capacity, aeration, and root development, thereby increasing plants’ resilience to abiotic stresses such as drought, salinity, and temperature extremes. In wheat and other crops, integrating vermicompost with such sustainable management not only boosts productivity but also fosters a resilient agroecosystem capable of sustaining yields under changing climatic conditions (Farooqi et al., 2024; Hafez et al., 2020).
Several studies on cereal crops such as wheat and maize (Zea mays L.) have reported the positive effects of vermicompost, applied alone or in combination with organic amendments, under non-drought, moderate drought, and severe drought conditions. These studies demonstrated variable outcomes, including reductions in morpho-physiological and yield attributes under drought, as well as improvements in biochemical characteristics that support stress tolerance (Aboelsoud and Ahmed, 2020; Hafez et al., 2020; Anee et al., 2022). The beneficial effects of humic substances present in vermicompost have been widely recognized. For instance, the application of vermicompost extracts in rice has been shown to mitigate the harmful effects of drought-induced reactive oxygen species (ROS) by enhancing the activity of antioxidant enzymes such as superoxide dismutase (SOD) and catalase (CAT) (García et al., 2012; Kiran, 2019). Positive effects of vermicompost have also been documented in a range of legume crops including chickpea (Gholipoor et al., 2011; Hosseinzadeh et al., 2018), mung-bean (Vigna radiata L.; Mahmoudi et al., 2016) and lentil (Lens culinaris Medik.; Ahmadpour and Hosseinzadeh, 2017), as well as in other crops such as canola (Brassica sp.; Rashtbari et al., 2020), wheat (Ahmad et al., 2024), and other plant families (Akhzari and Pessarakli, 2017; Kiran, 2019). However, the comparative effectiveness of wheat straw vermicompost (WSv), rice straw vermicompost (RSv), and cow dung-derived vermicompost (CDv) in alleviating drought stress under field conditions remains largely unexplored. The VTs selected for this study were based on our earlier pot experiments (Ahmad et al., 2024), which examined the effects of plant-based (wheat straw, rice straw) and animal-based (cow dung) vermicompost on the physiological and biochemical traits of wheat seedlings under water deficit conditions. These preliminary studies identified optimal vermicompost application rates that enhanced plant performance.
Building on these findings, the present field experiment was designed to test the hypothesis that wheat straw, rice straw, and cow dung vermicompost can improve wheat yield, physiology, growth, and nutrient uptake by mitigating the adverse impacts of drought stress. Accordingly, a 2-year field trial was conducted to evaluate the effects of different vermicompost types on agronomic, biochemical, physiological, and growth traits of two wheat cultivars—Faisalabad-08 (drought-tolerant) and Galaxy-13 (drought-sensitive)—under varying soil moisture regimes.
2 Materials and methods
The field experiments were carried out at the Student Research Area, University of Agriculture Faisalabad, during the winter season 2020–2021 and repeated in 2021–2022. The experimental site is situated 184.4 m above mean sea level, at 73° 06′E longitude and 31° 04′N latitude. A randomized complete block design (RCBD) with a split–split plot arrangement and three replications was employed. Vermicompost was prepared from wheat straw, rice straw, and cow dung using the earthworm Eisenia fetida. Optimized doses of these substrates were applied at sowing time and uniformly incorporated into the soil at a depth of 10–15 cm using a cultivator to ensure proper nutrient distribution and accessibility for wheat plants. The seedbed was prepared according to standard crop requirements, and wheat was sown at a rate of 120 kg ha−1 seed rate. Fertilizers were applied at a recommended rate of 110:100 kg ha−1 N: P, while all other management practices were kept uniform across treatments. Each experimental plot measured 4.5 m × 2.25 m. The treatments consisted of (a) three soil moisture regimes: D0 = 70% field capacity (FC) (no drought), D1 = 45% field capacity (mild drought), and D2 = 30% field capacity (severe drought); (b) four vermicompost levels: VT0 = control, VT1 = 8 t ha−1 wheat straw vermicompost (WSv), VT2 = 6 t ha−1 rice straw vermicompost (RSv), and VT3 = 6 t ha−1 cow dung vermicompost (CDv); and (c) two wheat cultivars: Faisalabad-08 (drought-tolerant) and Galaxy-13 (drought-sensitive).
2.1 Irrigation water analysis
Canal water was used for irrigation in this experiment, and its quality was analyzed prior to wheat sowing. Water samples were collected in rubber bottles, properly labeled, and immediately transported to the “Soil and water testing laboratory, Ayub Agricultural Research Institute, Faisalabad” for examination. The analyses were performed following the guidelines of the International Center for Agricultural Research in the Dry Areas (ICARDA) manual for soil, plant, and water analyses. The detailed physicochemical properties of the irrigation water are provided in Supplementary Table S1.
2.2 Soil analysis
Soil samples were collected in polythene bags, properly tagged, and immediately transported to the “soil and water testing laboratory in Ayub Agricultural Research Institute, Faisalabad” for analysis. Physical properties (sand, silt, and clay) were determined following the ICARDA manual for soil, plant, and water analysis. Chemical parameters, including pH, EC, organic matter, total nitrogen, available phosphorus, available potassium, calcium, magnesium, iron, zinc, copper, and nickel, were analyzed according to the methods described by Bellitürk et al. (2020) and Anee et al. (2022). Microbial biomass, including microbial activity and bacterial and fungal populations, was assessed using the method of Aslam et al. (2021). A summary of physicochemical analysis from this study is presented in Supplementary Table S2.
2.3 Data to be recorded
Standardized methods were used to record data on agronomic, biochemical, and physiological traits of wheat plants grown in the field, as described below.
2.3.1 Agronomic parameters
Plant height (cm) was measured from the soil surface to the top of the plant using a meter rod, with ten plants randomly selected from each sub-plot. The number of tillers per square meter was counted using a 1 m2 quadrat (Aslam et al., 2023). Spike length (cm) was determined from ten plants per plot using a 1-foot scale, while the number of spikelets per spike and the number of grains per spike were counted from ten randomly selected plants in each plot. At threshing, a 1,000-grain weight (g) was obtained by counting and weighing one thousand grains on an electronic balance. Biological yield (BY) (t ha−1) was measured by harvesting above-ground biomass from 1 m2, weighing it, and converting the value to t ha−1. Grain yield (t ha−1) was determined by hand-threshing the grains from 1 m2, weighing them, and converting to t ha−1. Straw yield (t ha−1) was calculated as the difference between total biomass and grain yield and expressed in t ha−1. Finally, the harvest index (HI, %) was calculated as the ratio of grain yield to biological yield using the formula (Aslam et al., 2023).
2.3.2 Biochemical parameters
Leaf nitrogen contents (mg g−1 dw) were determined by digesting 0.1 g of dried, ground leaves in digestion tubes containing 5 mL of concentrated H2SO4. After incubation at room temperature, 1 mL of 35% H2O2 was added along the tube walls. The tubes were placed in a digestion block and heated to 350 °C until fumes appeared, then removed, cooled, and treated again with H2O2. This process was repeated until the digest became colorless. The final extract was diluted to 50 mL in a volumetric flask, and nitrogen concentration was measured using Kjeldahl’s apparatus (Model: KJELTEC™ 8,400, Foss Analytical A/S, Hillerød, Denmark). Leaf phosphorus content (mg g−1 dw) was determined by reacting 5 mL of sample solution with 10 mL of Barton’s reagent in a volumetric flask, then diluting to volume with distilled water. KH2PO4 was used as a standard, and color development was measured at 420 nm using a spectrophotometer (Model: UV-1800, Shimadzu Corporation, Kyoto, Japan). Leaf potassium content (mg g−1 dw) was determined by digesting 0.1 g of dried, ground leaves with 5 mL concentrated H2SO4 in digestion tubes, followed by the gradual addition of H2O2 until the digest became clear and colorless. The final volume was adjusted to 50 mL in volumetric flasks, and potassium concentration was measured using a flame photometer (Model: Jenway PFP7, Bibby Scientific Ltd., Staffordshire, UK). Potassium contents were determined from the extract (Aslam et al., 2023). Leaf iron concentration was analyzed using atomic absorption spectrophotometry. A stock Fe solution (100 mg L−1) was prepared by dissolving 0.1 g of clean, untarnished iron in 10 mL of 10% slightly heated H2SO4, and diluting to 1,000 mL. Working standards (0–20 mg/L) were prepared from the stock, and Fe concentration in the samples was obtained by comparing absorbance values with the standard calibration curve. Leaf zinc content (ppm) was determined using the wet digestion method described by Bellitürk et al. (2020). Briefly, 0.5 g of dried, ground leaf samples (0.5 g) were digested with a mixture of nitric acid (HNO3) and perchloric acid (HClO4) in a 2:1 ratio on a block digester until a clear solution was obtained. After cooling, the digested samples were diluted with distilled water, filtered, and analyzed for Zn concentration using an atomic absorption spectrophotometer (AAS).
2.3.3 Physiological parameters
Total chlorophyll (Chl) contents (a + b) [mg g−1 fwt] were determined using Arnon’s (1949) method. Briefly, 0.2 g of freshly cut leaves was extracted in 80% acetone and kept overnight at 0–4 °C. The samples were then centrifuged at 10,000 g for 5 min, and the absorbance of the supernatant was recorded at 645 and 663 nm using a spectrophotometer. Chl a and b concentrations were calculated using the following equations:
where OD represents to optical density, V represents the extract volume (in mL), and W represents the fresh weight of leaf tissue (in g). Leaf chlorophyll index was further assessed using a portable Soil Plant Analysis Development (SPAD) meter (Model SPAD-502; Minolta Corporation, Ramsey, NJ, USA).
2.4 Statistical analysis
Data on agronomic, biochemical, and physiological attributes were subjected to analysis of variance (ANOVA) using Fisher’s method. Treatments were compared using Tukey’s Honestly Significant Difference (HSD) test at p ≤ 0.05, following the procedure of Steel et al. (1997). All statistical analyses were performed using Statistix version 10.0 (Analytical Software, Tallahassee, FL, USA). Graphical illustrations were prepared with SigmaPlot 10.0 and Origin 2024b.
3 Results
3.1 Agronomic parameters
The interaction between moisture regime and VTs was significant for plant height, number of total tillers, and spike length in both 2020–2021 and 2021–2022 (Supplementary Table S3; Figure 1). Irrigation levels, vermicompost (VT), and their interaction exerted significant effects on these agronomic traits across both years. Plant height, tiller number, and spike length were highest under well-watered conditions, whereas drought stress significantly reduced these attributes, with the greatest decline observed under severe drought stress (30% FC). Application of VT mitigated the adverse effects of drought with CDv (6 t ha−1), producing the greatest improvement, followed by RSv (6 t ha−1) and WSv VT (8 t ha−1). This positive effect of vermicompost was consistent across both years and under all moisture regimes. Even under non-stressed conditions, VT application at sowing enhanced agronomic traits. The maximum values for plant height, tiller number, and spike length were obtained with CDv (6 t ha−1) across 70% FC (well-watered), 45% FC (moderate drought), and 30% FC (severe drought). Between cultivars, Faisalabad-08 (drought-tolerant) outperformed Galaxy-13 (drought-sensitive), showing higher plant height, spike length, and tiller number under both moderate and severe drought conditions.
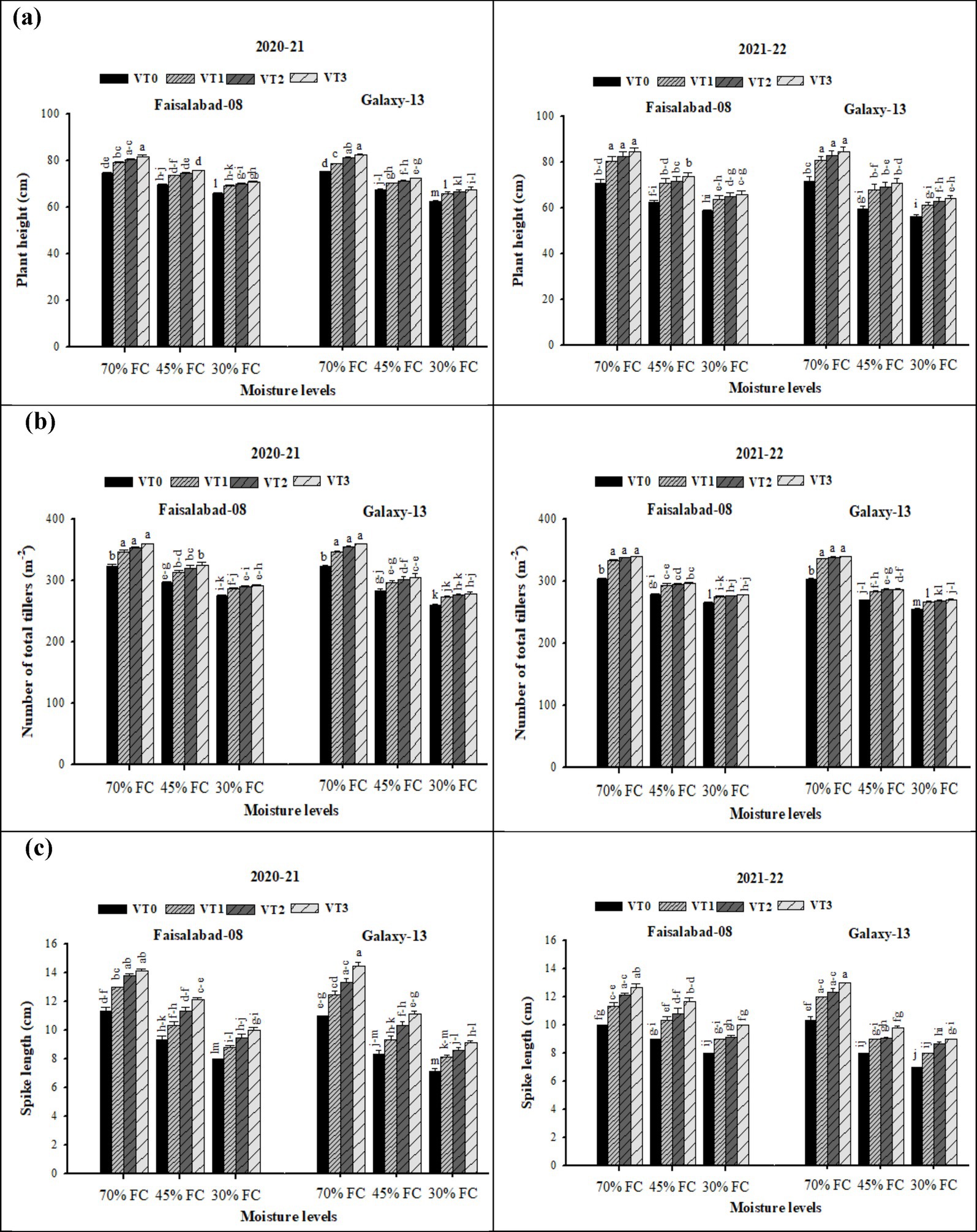
Figure 1. (a–c) Effects of soil applied with wheat straw, rice straw, and cow dung vermicompost on plant height, number of total tillers, and spike length of both wheat varieties under three water scarcity levels. Tukey’s HSD test shows that bars with changed alphabetical letters differ significantly at a 5% probability level.
Spikelets per spike, grains per spike, and thousand-grain weight (TGW) were significantly affected by both drought stress and vermicompost application (Supplementary Table S4 and Figure 2). Drought stress markedly (p ≤ 0.05) reduced these traits under 45% FC (moderate drought) and 30% FC (severe drought) compared with the well-watered condition (70% FC) in both years. Vermicompost application enhanced spikelets per spike, grains per spike, and TGW under both drought and well-watered conditions, with the highest values recorded for CDv (6 t ha−1) and the lowest in the control. Under moderate and severe drought stress, Faisalabad-08 produced more spikelets per spike, grains per spike, and also higher TGW than Galaxy-13. However, under well-watered conditions, both cultivars exhibited comparable values for these traits across both years.
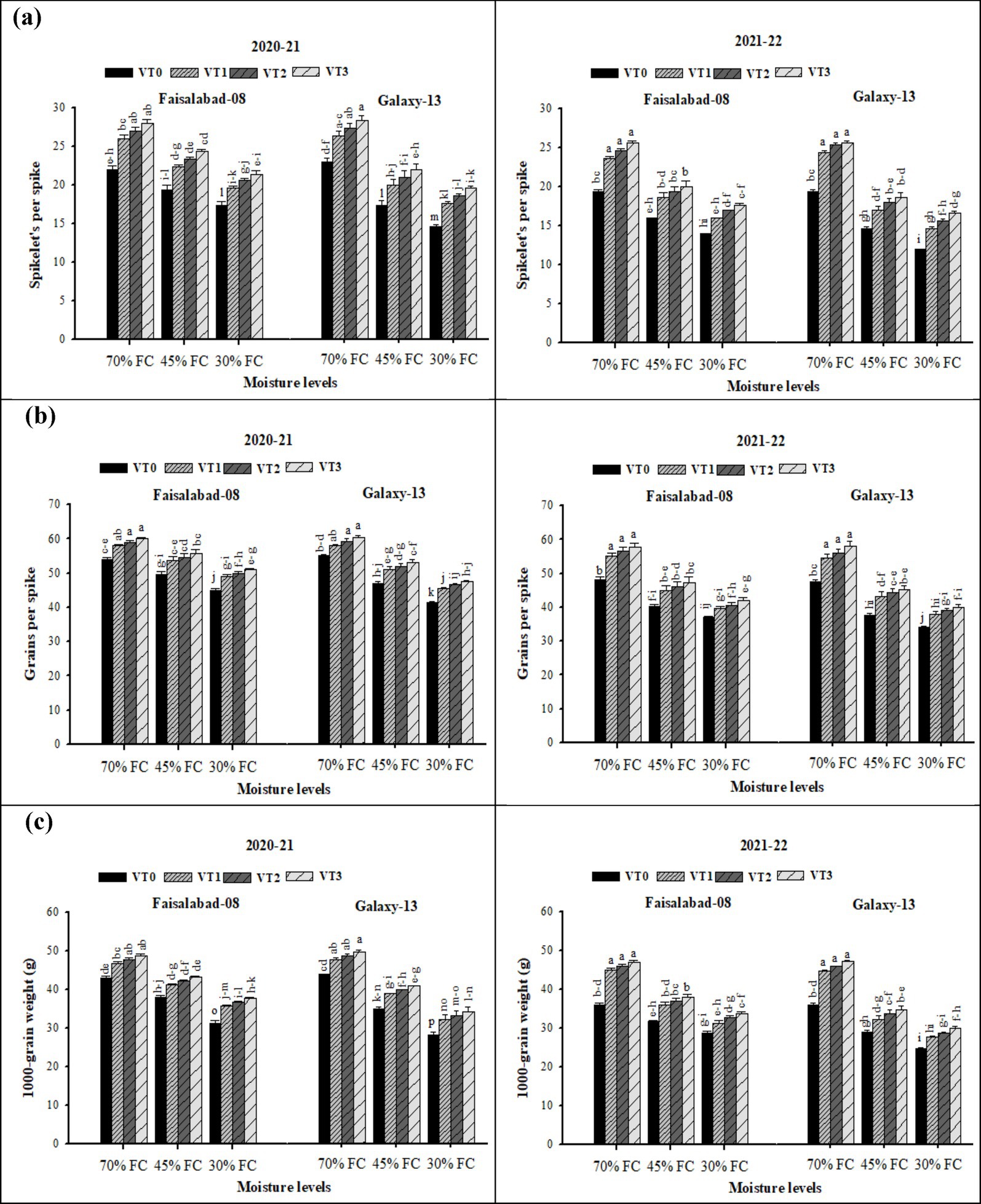
Figure 2. (a–c) Effects of soil applied with cow dung, wheat, and rice straw vermicompost on the spikelets per spike, grains per spike, and thousand-grain weight of both wheat varieties under various drought levels. Tukey’s HSD test shows that bars with varying alphabetical letters differ significantly at a 5% probability level.
The interaction between moisture regime and vermicompost application had a significant effect on grain yield, straw weight, BY, and also HI of wheat in both 2020–2021 and 2021–2022, with HI showing a relatively greater increase in 2021–2022 than in 2020–2021. Both drought stress and VT significantly influenced these traits across both years (Supplementary Table S5 and Figure 3). Compared with well-watered conditions (70% FC), drought stress markedly reduced GY, straw weight, BY, and HI, with reductions being more pronounced under severe drought stress (30% FC) than under moderate drought stress (45% FC). Application of vermicompost mitigated the adverse effects of water stress, significantly improving BY, GY, straw weight, and HI in both cultivars across both years (Supplementary Table S5 and Figure 3). Among treatments, CDv (6 t ha−1) produced the highest values, followed by RSv (6 t ha−1) and WSv (8 t ha−1), all of which were significantly superior to the control under both drought and well-watered conditions. When comparing cultivars, Faisalabad-08 and Galaxy-13 formed statistically similar under well-watered conditions. However, under drought stress, Faisalabad-08 consistently outperformed Galaxy-13 across all measured parameters.
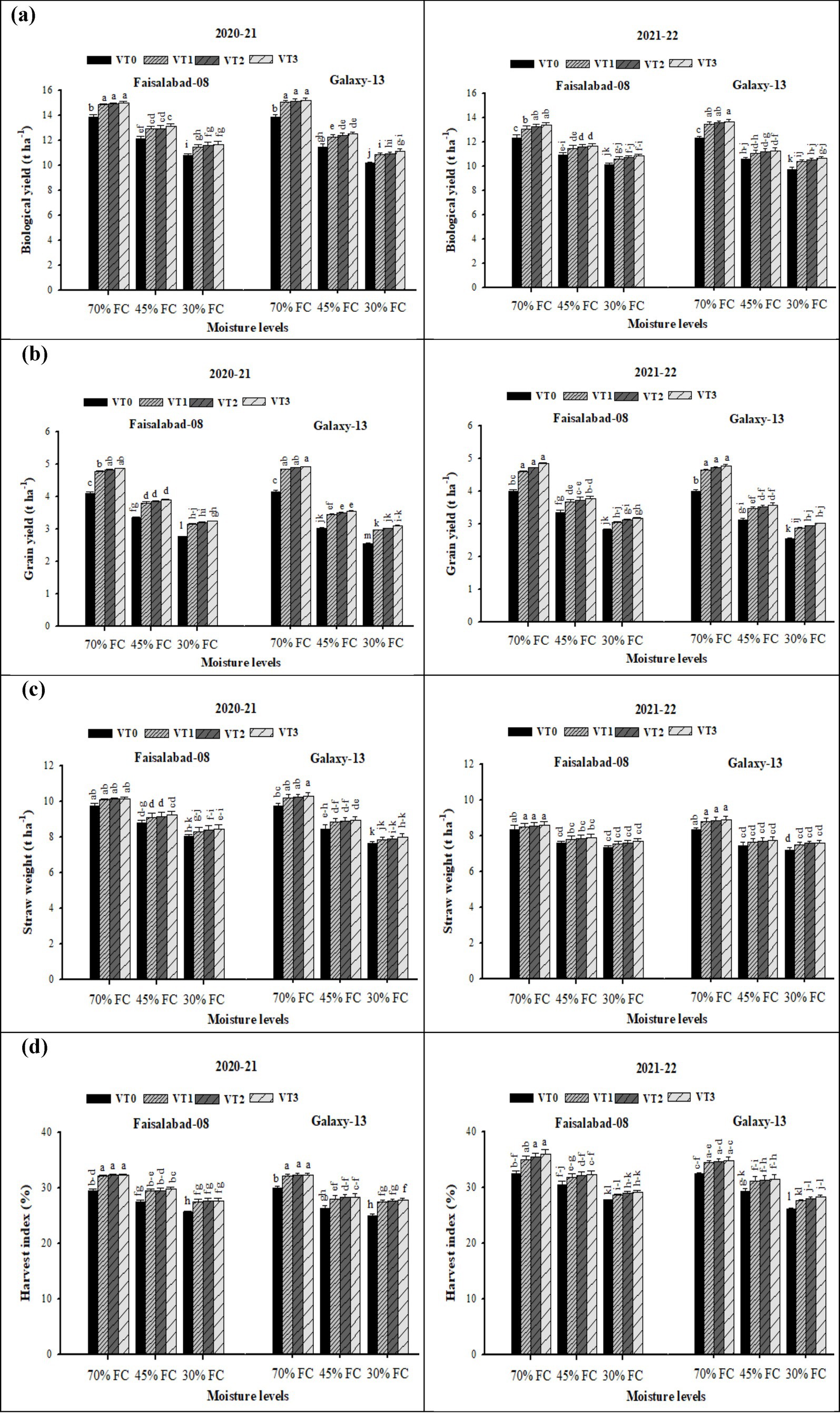
Figure 3. (a–d) Effects of soil applied with wheat straw, rice straw, and cow dung vermicompost on biological yield, grain yield, straw weight, and harvest index of two wheat cultivars under different drought levels. Tukey’s HSD test shows that bars with different alphabetical letters differ significantly at a 5% probability level.
3.2 Nutrient traits
Leaf nitrogen (N), phosphorus (P), and potassium (K) (NPK) contents (Supplementary Table S6 and Figure 4) were significantly (p ≤ 0.05) reduced under moderate drought (45% FC) and severe drought (30% FC) conditions compared with well-watered (70% FC) conditions in both wheat varieties. The main effects of drought stress, vermicompost application, and their interaction were significant for leaf NPK contents in both 2020–2021 and 2021–2022. Water scarcity markedly reduced the leaf NPK contents, with the greatest declines observed under severe stress. However, the adverse impact of drought in both years was effectively alleviated by vermicompost application (Supplementary Table S6 and Figure 4). Among treatments, CDv (6 t ha−1) resulted in the highest leaf NPK contents, followed by RSv (6 t ha−1) and WSv (8 t ha−1), while the lowest values were recorded in the control. Between the two cultivars, Faisalabad-08 consistently exhibited higher leaf NPK contents than Galaxy-13.
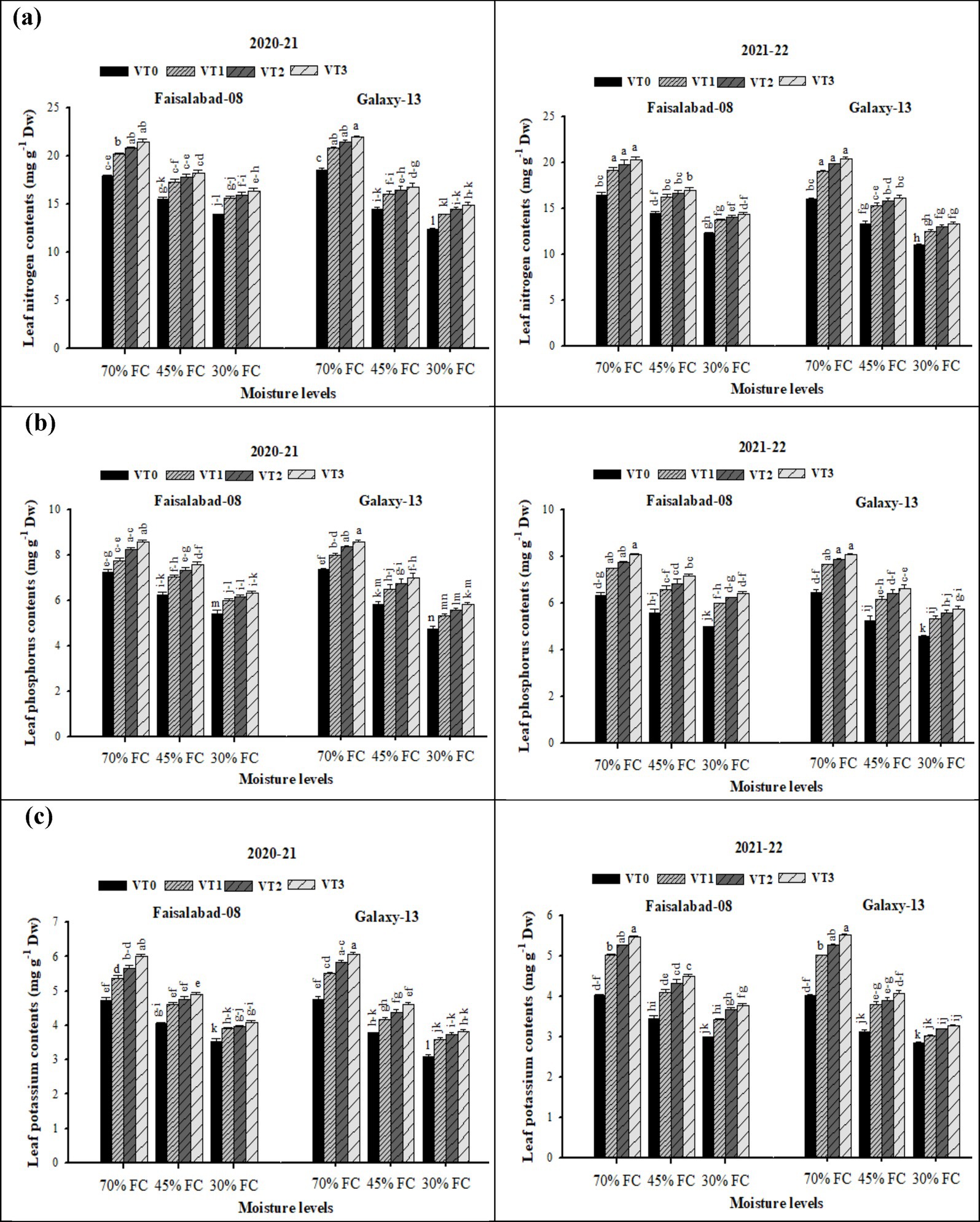
Figure 4. (a–c) Effects of soil applied with wheat straw, rice straw, and cow dung vermicompost on leaf NPK concentration of both varieties under different drought levels. Tukey’s HSD test shows that bars with changed alphabetical letters differ significantly at a 5% probability level.
Similarly, leaf Fe and Zn contents (Supplementary Table S7 and Figure 5) were strongly affected by moderate and severe drought stress conditions in both years. Vermicompost application significantly enhanced leaf Fe and Zn contents under drought stress, with the greatest response observed to CDv (6 t ha−1), followed by RSv (6 t ha−1), and WSv (8 t ha−1). The lowest values were recorded in the control across both cultivars. Consistent with NPK results, Faisalabad-08 maintained higher leaf Fe and Zn contents than Galaxy-13 under moderate and severe drought conditions.
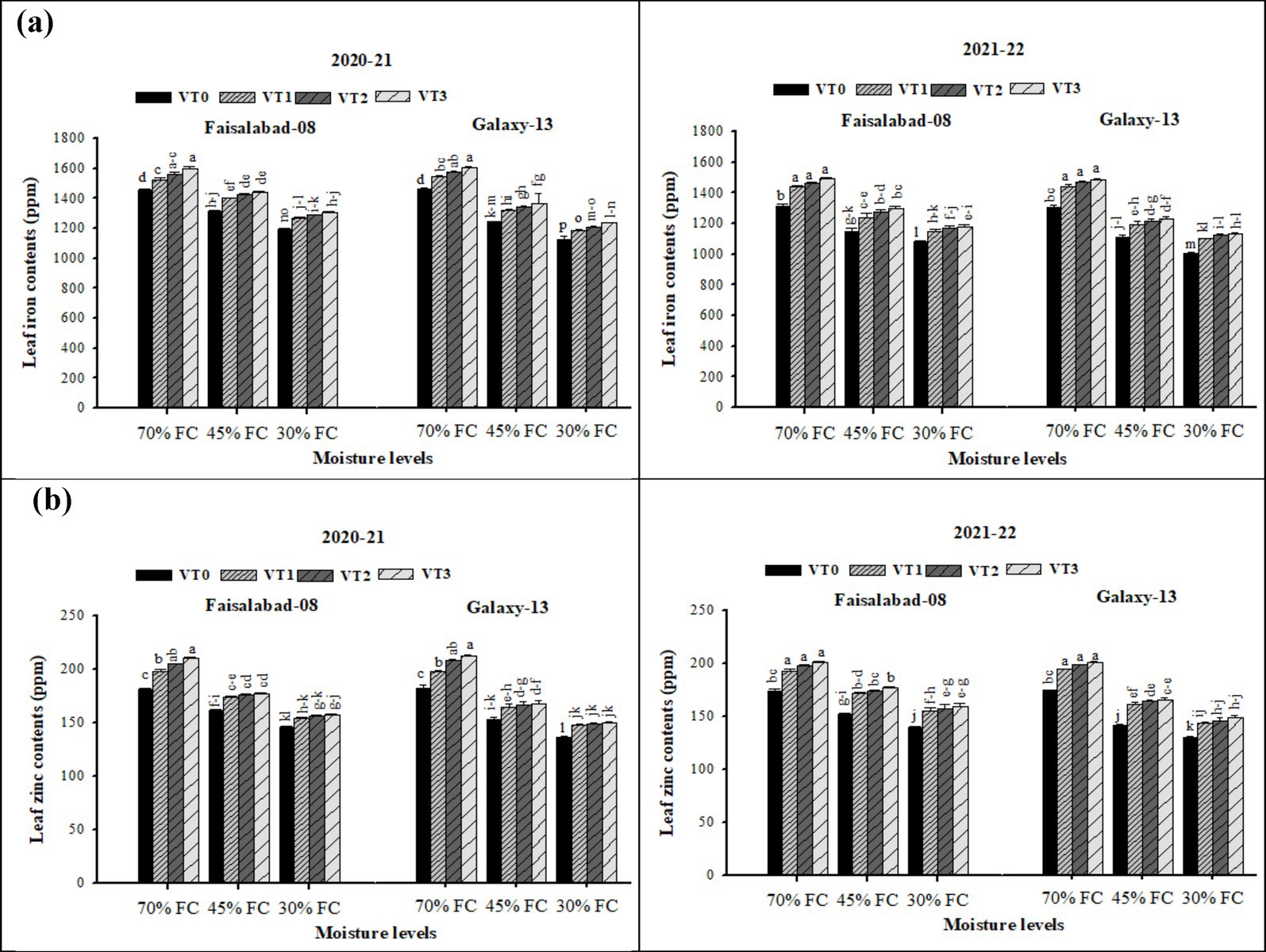
Figure 5. (a,b) Effects of soil applied with cow dung, wheat, and rice straw vermicompost on leaf iron and zinc contents on both wheat varieties under different drought levels. Tukey’s HSD test shows that bars with different alphabetical letters differ significantly at a 5% probability level.
3.3 Physiological parameters
Chlorophyll content and index were significantly influenced by drought stress levels, vermicompost application, and their interaction (Supplementary Table S8 and Figure 6). Under well-watered conditions, total chlorophyll contents and chlorophyll index were significantly higher compared with moderate and severe drought stress. Application of CDv (6 t ha−1) resulted in the greatest increase in total chlorophyll contents and SPAD values, followed by RSv (6 t ha−1) and WSv (8 t ha−1) across both years. Under well-watered conditions, the drought-sensitive and drought-tolerant cultivars exhibited statistically similar chlorophyll content and SPAD values. However, under drought stress, Faisalabad-08 maintained significantly higher chlorophyll content and chlorophyll index than Galaxy-13.
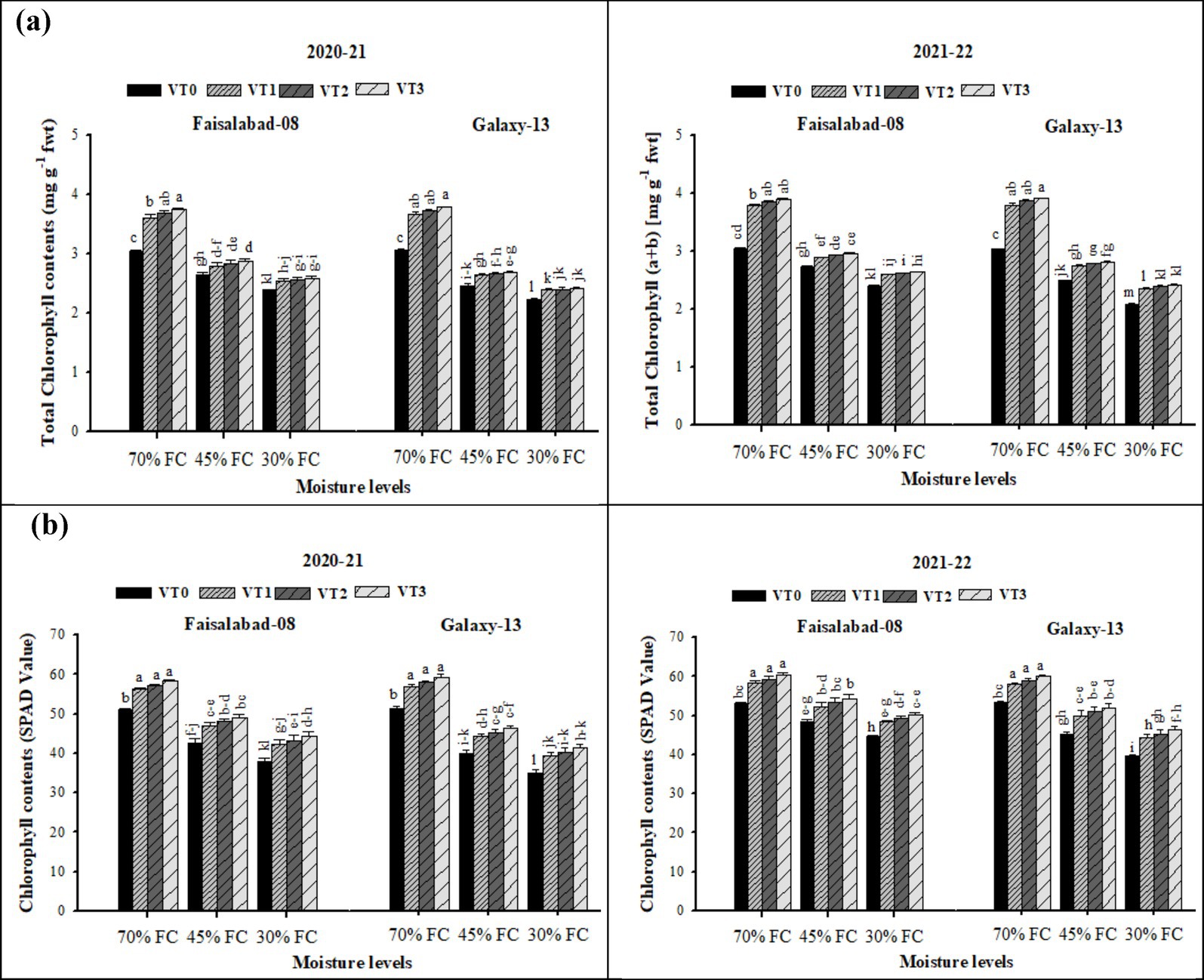
Figure 6. (a,b) Effects of soil applied with cow dung, wheat, and rice straw vermicompost on total chlorophyll contents (a + b) and chlorophyll contents (SPAD value) of both wheat varieties in various drought levels. Tukey’s HSD test shows that bars with different alphabetical letters differ significantly at a 5% probability level.
3.4 Chord diagrams
The chord diagram illustrates the interrelationships among agronomic, physiological, and nutrient traits of wheat cultivars under different soil moisture regimes during the 2020–2021 and 2021–2022 growing seasons (Figure 7). Strong associations were observed between GY and key traits such as BY, HI, and TGW, emphasizing their pivotal role in yield determination. Similarly, nutrient traits such as leaf N, P, and K showed significant interconnections, reflecting their combined influence on plant growth and productivity. Architectural characteristics, such as plant height (PH), spike length (SL), and number of total tillers (TT), also displayed notable associations, indicating their importance in cultivar adaptability under contrasting soil moisture conditions. Similarly, physiological parameters, particularly chlorophyll contents (CP) and total chlorophyll contents (TC), were closely linked with nutrient traits, indicating their role in maintaining photosynthetic efficiency and nutrient uptake under varying water availability. Additionally, traits such as grains per spike (GS), spikelets per spike (SS), and straw weight (SW) demonstrated moderate associations with yield-related parameters, revealing their indirect contributions to productivity.
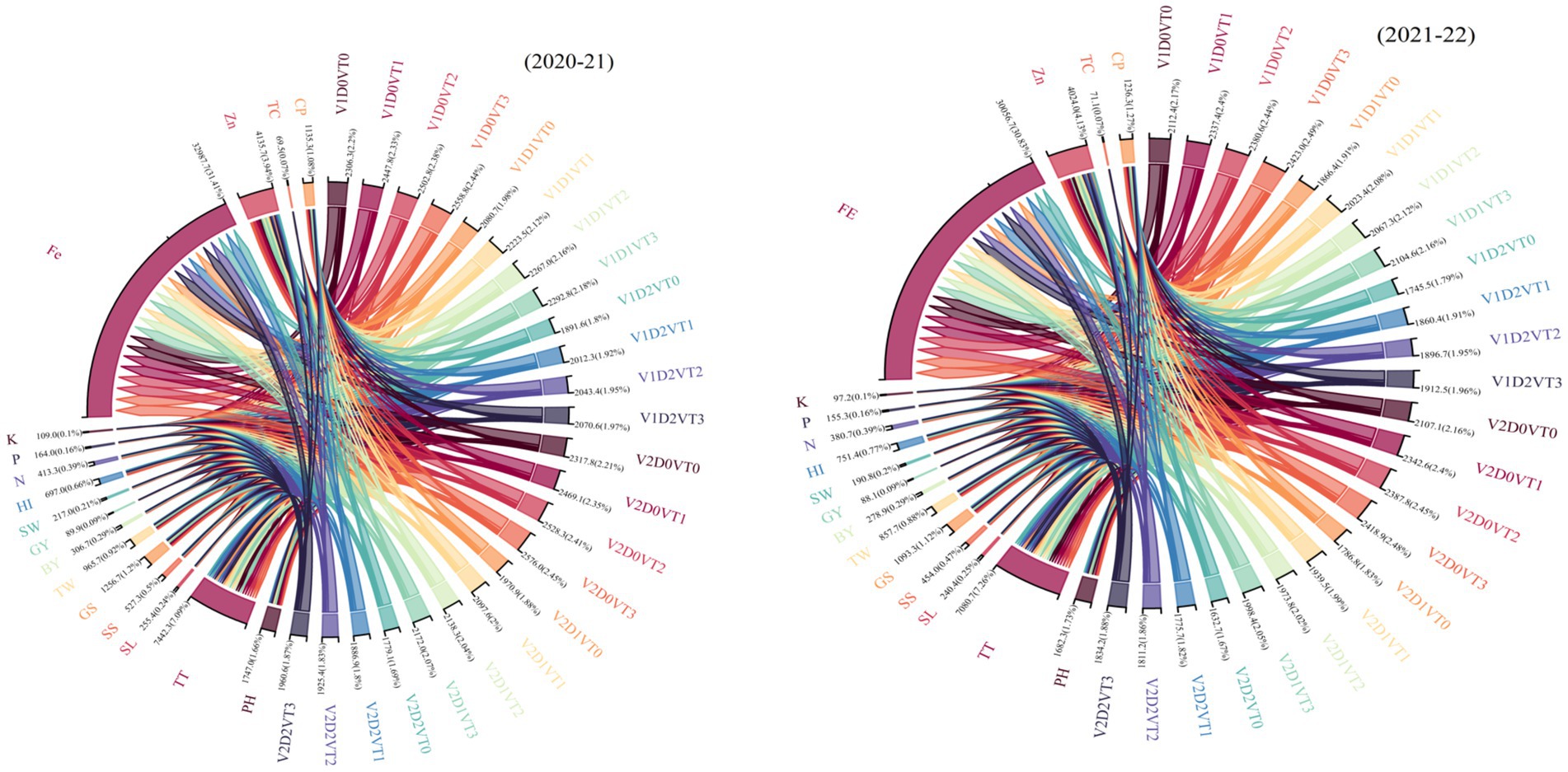
Figure 7. The chord diagram for the study provides a visual representation of the interrelationships among agronomic, physiological, and nutrient traits in wheat cultivars under different soil moisture regimes for the years 2020–2021 and 2021–2022. CP, chlorophyll contents; TC, total chlorophyll contents; Zn, leaf zinc contents; Fe, leaf iron contents; K, leaf potassium contents; P, leaf phosphorus contents; N, leaf nitrogen contents; HI, harvest index; SW, straw weight; GY, grain yield; BY, biological yield; TW, thousand-grain weight; GS, grains per spike; SS, spikelets per spike; SL, spike length; TT, number of total tillers; PH. plant height.
3.5 Correlation matrix
The correlation matrix provided a detailed overview of the interrelationships among agronomic, physiological, and nutrient traits in wheat cultivars under contrasting soil moisture regimes during 2020–2021 and 2021–2022 growing seasons (Figure 8). Each cell in the matrix represents the strength and direction of the association between two traits, quantified using Pearson’s correlation coefficient. Positive correlations indicated traits that tended to increase together, such as GY and BY, or HI and TGW, highlighting their synergistic roles in yield formation. Nutrient traits, including leaf N, P, K, Zn, and Fe, were also positively correlated with one another, reflecting their collective contribution to plant health, metabolism, and resilience under stress. Similarly, plant architectural traits, such as plant height (PH), spike length (SL), and the number of total tillers (TT), showed strong positive associations with yield components, underscoring their importance in maintaining productivity under water-deficit conditions. Physiological traits, particularly chlorophyll index and total chlorophyll contents (TC) exhibited significant correlations with both nutrient and yield-related traits, suggesting their role in sustaining photosynthetic efficiency and resource allocation under varying soil moisture regimes.
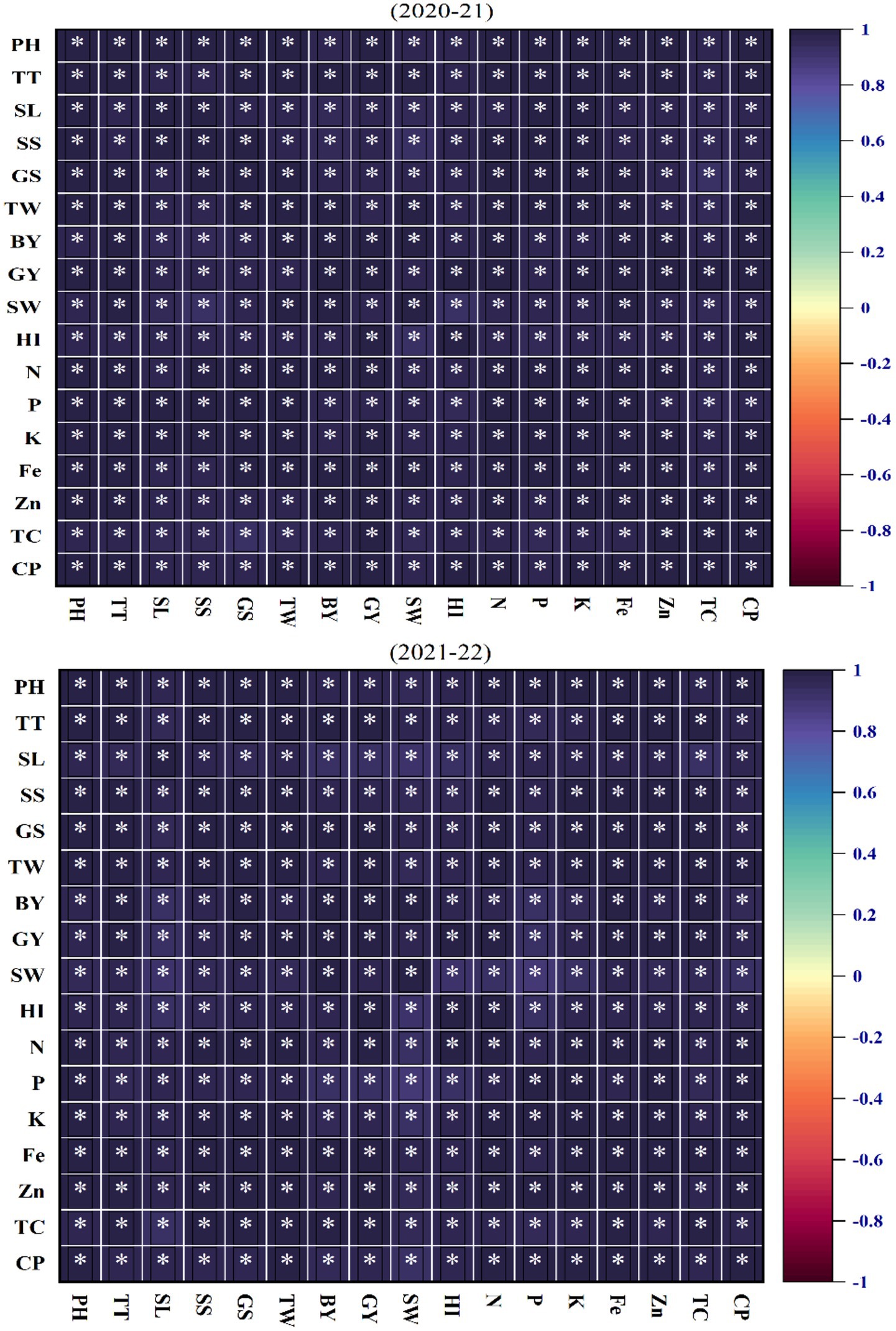
Figure 8. The correlation analysis for the 2-year study (2020–2021 and 2021–2022) highlights the relationship between physiological, agronomic, and nutrient traits in wheat cultivars under varying moisture conditions. CP, chlorophyll contents; TC, total chlorophyll contents; Zn, leaf zinc contents; Fe, leaf iron contents;, K, leaf potassium contents; P, leaf phosphorus contents; N, leaf nitrogen contents; HI, harvest index; SW, straw weight; GY, grain yield; BY, biological yield; TW, thousand-grain weight; GS, grains per spike; SS, spikelets per spike; SL, spike length; TT, number of total tillers; PH, plant height.
3.6 Heatmap analysis
The heatmap analysis provided a visual representation of the magnitude and direction of associations among agronomic, physiological, and nutrient traits in wheat cultivars across the 2020–2021 and 2021–2022 growing seasons (Figure 9). The color gradient in the heatmap, ranging from warm tones (e.g., red and orange) indicating strong positive correlations to neutral tones (e.g., white or light gray), indicating weak or negligible relationships, allowed for intuitive pattern recognition. Key patterns observed in the heatmap highlight that GY shares strong positive associations with BY, HI, and TW, signifying their collective importance in determining wheat productivity. Similarly, nutrient traits such as leaf N, P, K, Zn, and Fe are clustered together, exhibiting positive interrelationships that underscore their integrated role in supporting plant growth and resilience. The heatmap also emphasizes the interplay between physiological traits like CP and TC with yield-related parameters, indicating the contribution of photosynthetic efficiency to productivity. Agronomic traits, including PH, SL, and TT, showed notable correlations with both nutrient and yield traits, further highlighting their significance in adapting to moisture stress conditions.
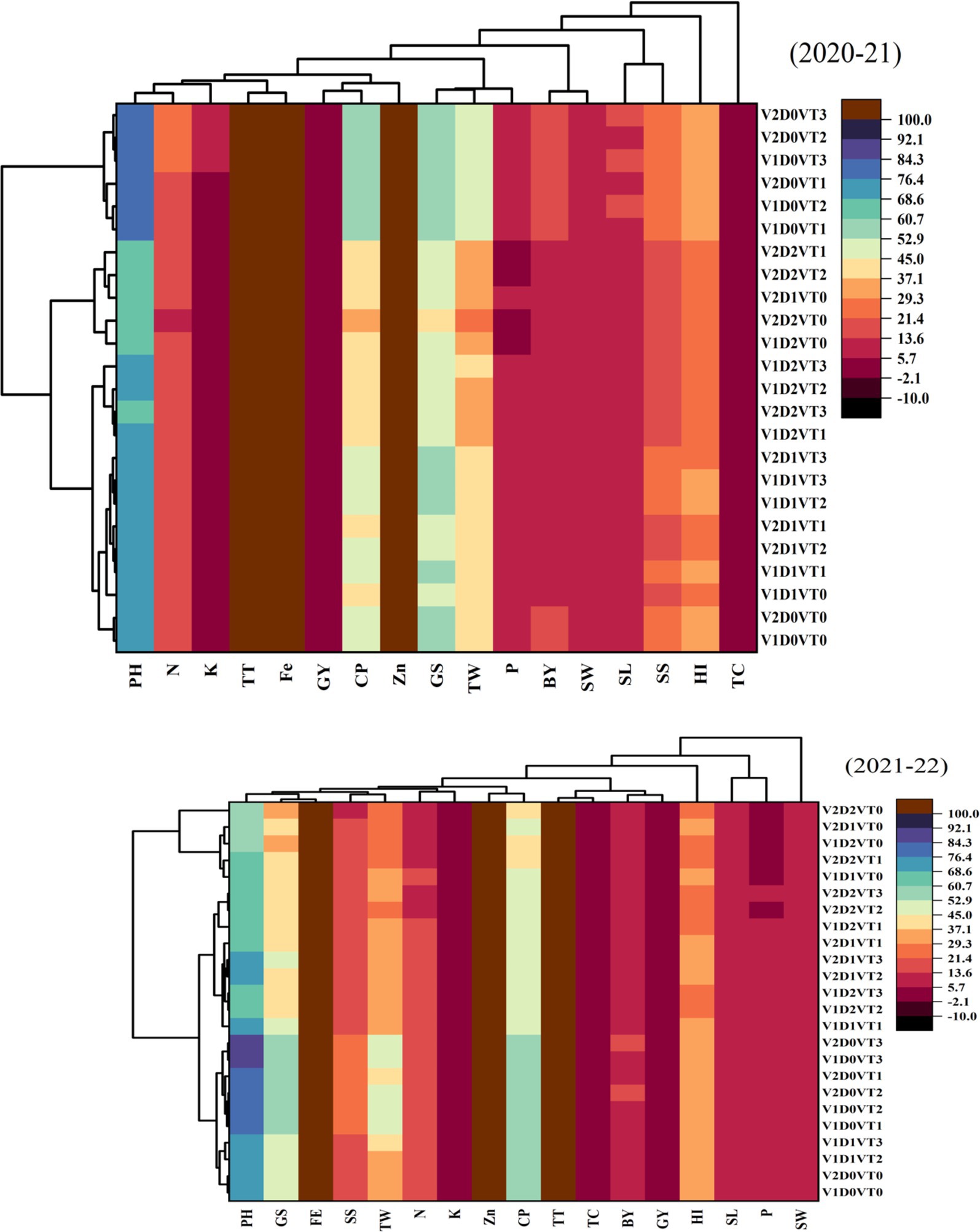
Figure 9. The heatmap analysis for this 2-year study (2020–2021 and 2021–2022) offers a clear visualization of the strength and direction of relationships among physiological, agronomic, and nutrient traits in wheat cultivars under different moisture regimes. CP, chlorophyll contents; TC, total chlorophyll contents; Zn, leaf zinc contents; Fe, leaf iron contents; K, leaf potassium contents; P, leaf phosphorus contents; N, leaf nitrogen contents; HI, harvest index; SW, straw weight; GY, grain yield; BY, biological yield; TW, thousand-grain weight; GS, grains per spike; SS, spikelets per spike; SL, spike length; TT, number of total tillers; PH, plant height.
3.7 Principal component analysis
The principal component analysis (PCA) plots revealed the relationships among different traits across the 2-year study (2020–2021 and 2021–2022). In both years, principal component 1 (PC1) accounted for the majority of variation (98.3% in 2020–2021 and 97.2% in 2021–2022), while principal component (PC2) explained a much smaller proportion (0.8 and 1.6%, respectively) (Figure 10). Key traits such as leaf Zn, Fe, K, P, HI, and TGW contributed strongly to PC1 and played a critical role in separating the two groups: Galaxy-13 and Faisalabad-08. The 95% confidence ellipses indicated some overlap between the two groups, though they were partially distinct. In 2020–2021, traits like HI, Fe, and Zn were highly associated with positive PC1 values, differentiating the groups. PC2 contributed minimally, suggesting that variables along this axis have a smaller influence. In 2021–2022, the importance of SW (straw weight) increased slightly, and PC2 explained slightly more variation compared to the previous year. This year also showed slightly more overlap between Galaxy-13 and Faisalabad-08, indicating reduced differentiation. Traits like grain yield (GY), biological yield (BY), and leaf N remained less influential compared to those associated with PC1. Overall, HI, Zn, Fe, and P consistently emerged as the most discriminating traits across both years, while minor shifts in PC2 indicated year-to-year variability in trait associations. These findings provide useful insights for identifying key traits in the evaluation of wheat performance under varying conditions.
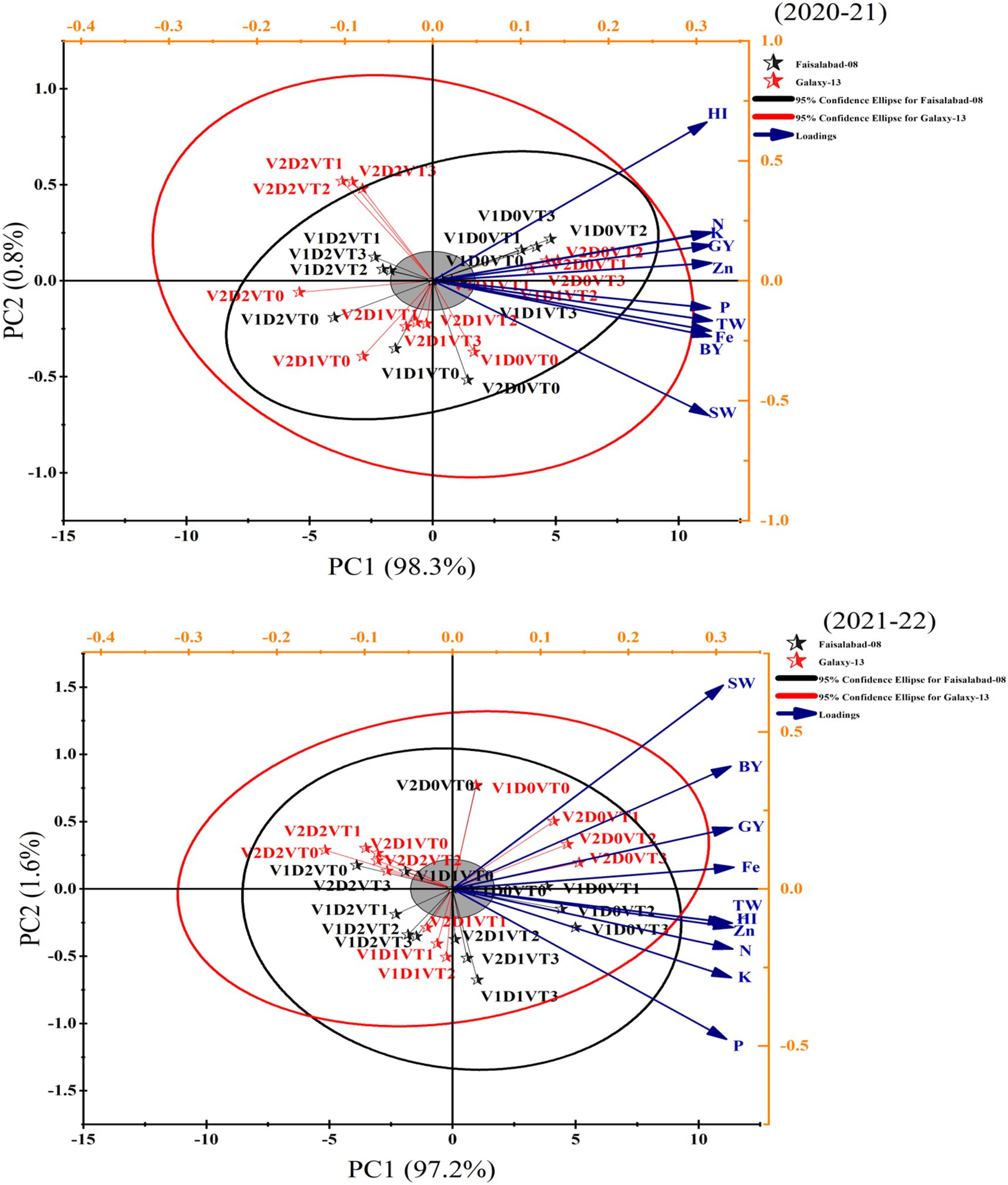
Figure 10. The principal component analysis (PCA) for this 2-year study (2020–2021 and 2021–2022) was conducted to reduce the dimensionality of the dataset and identify key traits that explain most of the variation in wheat cultivars under different moisture regimes. Zn, leaf zinc contents; Fe, leaf iron contents; K, leaf potassium contents; P; leaf phosphorus contents; N, leaf nitrogen contents; HI, harvest index; SW, straw weight; GY, grain yield; BY, biological yield; TW, thousand-grain weight.
4 Discussion
Plant’s response to extreme water stress is multifaceted, leading to reduced growth and yield (Hussain et al., 2018) by impairing physiological and metabolic processes (Jan et al., 2019). Water scarcity significantly limits productivity (Fathi et al., 2025). In the present study, key agronomic traits such as plant height, number of total tillers, and spike length were markedly reduced under moderate (45% FC), and severe drought stress (30% FC) in both cultivars (Faisalabad-08 and Galaxy-13) with or without vermicompost application, compared to well-watered (70% FC) conditions across both years. Similarly, drought stress significantly decreased grains per spike, spikelets per spike, and TGW, ultimately leading to declines in GY, straw weight, BY, and HI. Comparable reductions in wheat performance under field drought conditions have also been reported by Iqbal et al. (2018). Vermicompost, being a rich source of macro- and micronutrients, plays a protective role against drought stress (Ahmad et al., 2021; Aslam and Ahmad, 2020). Enhanced nutrient uptake and increased chlorophyll concentration under vermicompost application can be attributed to improved soil health, microbial activity, and nutrient mineralization, as observed in earlier studies (Aslam et al., 2023). Vermicompost enhances microbial enzymatic activities, thereby improving the bioavailability of essential nutrients (Anee et al., 2022). The earthworm species E. fetida, used in vermicompost production, plays a critical role in organic matter decomposition and nutrient cycling, which contributes to improved plant resilience under water deficit. Our results are in line with these findings, showing that CDv (6 t ha−1) produced by E. fetida was the most effective under both drought stress and non-stress conditions. Wheat plants grown with vermicompost showed significantly higher leaf N, P, K, Fe, and Zn contents under water-limited conditions, consistent with the findings of Akhzari and Pessarakli (2017) and Aslam et al. (2025). In our experiment, CDv at 6 t ha−1 significantly improved biochemical traits, followed by RSv (6 t ha−1) and WSv (8 t ha−1), whereas control plots without vermicompost showed the lowest values. Vermicompost not only enriches mineral nutrition but also improves the biological and structural properties of soil (Bellitürk et al., 2020; Aboelsoud and Ahmed, 2020). Its high porosity ensures optimal aeration and drainage, while its excellent moisture retention capacity provides resilience under drought stress (Hosseinzadeh et al., 2016). Vermicompost also contains higher concentrations of N, P, K, Ca, Mg, Fe, Zn, Cu, and Mn compared to other organic amendments (Huerta et al., 2010). Positive effects of vermicompost on yield under drought conditions have been reported in cereals and legumes (Lazcano et al., 2010), where it also improved growth attributes (Lu et al., 2002). Continuous application of vermicompost enhances soil organic matter, stimulates microbial activity, and increases nutrient availability, leading to better soil structure, water-holding capacity, and, ultimately, improved nutrient uptake and physiological performance in wheat. Over time, these improvements sustain agronomic productivity and reduce reliance on synthetic fertilizers, thereby promoting long-term soil fertility and agricultural sustainability (Ahmad et al., 2024; Hafez et al., 2020). Taken together, our findings demonstrate that vermicompost application significantly enhances biochemical and physiological traits under drought stress, making it a promising strategy for improving wheat resilience and productivity in water-limited environments.
Chlorophyll plays a vital role in photosynthesis, crop growth, and yield (Lawlor, 2002). In the present study, water scarcity significantly reduced chlorophyll content in both wheat cultivars. However, the drought-tolerant cultivar Faisalabad-08 maintained relatively higher chlorophyll under moderate and severe water stress compared to drought-sensitive wheat cultivar Galaxy-13 (Ibrahim et al., 2021). Plants with higher chlorophyll typically accumulate more photosynthates, leading to stronger roots, shoots, and leaves. Vermicompost has been widely recognized for improving plant physiological functions and mitigating stress conditions (Jat and Ahlawat, 2006; Hosseinzadeh et al., 2016). Recent studies indicate that optimized VT applications reduce the adverse effects of water stress in wheat by enhancing and sustaining chlorophyll a, total chlorophyll, and SPAD values. Flexibility in growth parameters such as leaf area index (LAI), leaf area duration (LAD), and crop growth rate (CGR) of wheat under different VTs was linked to variable soil moisture levels. These growth parameters were highest under CDv (6 t ha−1), followed by RSv (6 t ha−1) and WSv (8 t ha−1) across both cultivars under moderate drought (45% FC), severe drought (30% FC), and well-watered (70% FC) conditions. The lowest growth responses were observed under severe drought without VT application. Similar results were also reported by Ahmad et al. (2021), who found significant changes in LAI, LAD, and CGR. Beyond improving growth indices, vermicompost also enriched the soil with nutrients, organic matter, humic acid, beneficial microbes, actinomycetes, and various growth-promoting regulators (Aslam et al., 2021).
However, WSv at 8 t ha−1 showed comparatively lower improvements, likely due to its slower decomposition rate and limited availability in the region. Previous research has indicated that cereal straw-based vermicomposs generally have lower nutrient concentrations and slower mineralization compared to those derived from animal dung or leguminous residues (Aslam and Ahmad, 2020; Aslam et al., 2023; Ahmad et al., 2024).
Overall, the findings suggest that CDv at 6 t ha−1 should be prioritized for wheat production in water-scarce regions due to its superior nutrient profile, better availability, and positive impact on yield and physiological traits. RSv at 6 t ha−1 can serve as a viable alternative where cow dung is less accessible, while WSv at 8 t ha−1 appears less efficient and economically less feasible, particularly in Punjab, due to limited availability.
5 Conclusion
This study demonstrates that applying cow dung vermicompost at 6 t ha−1 is the most effective strategy for enhancing wheat growth, physiology, and yield under drought stress. The drought-tolerant cultivar Faisalabad-08 consistently outperformed the sensitive cultivar Galaxy-13 under both moderate and severe drought, primarily due to superior adaptive mechanisms. Its improved performance was associated with enhanced nutrient uptake (K, N, P, Fe, Zn), higher chlorophyll content, and better water relations, which together sustained photosynthetic efficiency and cellular turgor. Among vermicompost sources, cow dung was the most effective, followed by rice straw, whereas wheat straw vermicompost proved to be the least effective. These findings underscore the importance of selecting suitable vermicompost materials to maximize wheat productivity under water-limited conditions. For a practical perspective, the use of cow dung vermicompost at 6 t ha−1, prepared using E. fetida is recommended for farmers in drought-prone areas to improve wheat resilience and yield. However, this study was limited to two field seasons and may not fully capture long-term soil–plant-microbe interactions, climatic variability, or broader ecosystem effects. Future research should focus on multi-year, multi-location trials to validate these results and investigate vermicompost’s role in soil microbial dynamics, carbon sequestration, and ecosystem services. Such studies will be crucial for developing sustainable and climate-resilient wheat production systems.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
AA: Conceptualization, Software, Data curation, Formal analysis, Methodology, Writing – original draft. ZA: Conceptualization, Data curation, Writing – original draft, Project administration, Supervision, Visualization. MAA: Formal analysis, Methodology, Software, Validation, Writing – review & editing. UZ: Software, Validation, Writing – review & editing, Conceptualization, Investigation, Visualization. HE-B: Funding acquisition, Investigation, Software, Validation, Visualization, Writing – review & editing. MOA: Formal analysis, Resources, Software, Validation, Visualization, Writing – review & editing. PVP: Formal analysis, Funding acquisition, Investigation, Project administration, Resources, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by the Deanship of Scientific Research, Vice Presidency for Graduate Studies and Scientific Research, King Faisal University, Saudi Arabia (KFU253517). The authors extend their appreciation to the Deanship of Research and Graduate Studies at King Khalid University for funding this work through Large Research Project under grant number RGP2/351/46. The authors gratefully acknowledge HEC for providing financial support to carry out this research work under HEC project “NRPU-HEC project no. 7527/Punjab/NRPU/R&D/ HEC/2017_Vermicomposting: A resourceful organic fertilizer to improve agriculture production and soil health.” and PARB project# 18-550, “Developing agricultural waste management system to produce different kinds of organic fertilizers for sustainable agriculture.”
Acknowledgments
The results presented in this paper are a part of Ali Ahmad’s PhD studies. “Contribution Number 26-049-J from the Kansas Agricultural Experiment Station”. The authors acknowledge the support provided by the Deanship of Scientific Research, Vice Presidency for Graduate Studies and Scientific Research, King Faisal University, Saudi Arabia. Moreover, the authors extend their appreciation to the Deanship of Research and Graduate Studies at King Khalid University for funding this work through Large Research Project under grant number RGP2/351/46.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2025.1695055/full#supplementary-material
References
Aboelsoud, H. M., and Ahmed, A. A. (2020). Effect of biochar, vermicompost and polymer on wheat and maize productivity in sandy soils under drought stress. Environ. Biodivers. Soil Secur. 4, 85–102. doi: 10.21608/jenvbs.2020.29442.1095
Ahmad, A., Aslam, Z., Ahmad, M., Zulfiqar, U., Yaqoob, S., Hussain, S., et al. (2024). Vermicompost application upregulates morpho-physiological and antioxidant defense to conferring drought tolerance in wheat. Plant Stress 11:100360. doi: 10.1016/j.stress.2024.100360
Ahmad, A., Aslam, Z., Bellitürk, K., Iqbal, N., Idrees, M., Nawaz, M., et al. (2021). Earth worms and vermicomposting: a review on the story of black gold. J. Innov. Sci. 7, 167–173. doi: 10.17582/journal.jis/2021/7.1.167.173
Ahmadpour, R., and Hosseinzadeh, S. R. (2017). Effect of vermicompost fertilizer on morphological traits of lentil under water stress. Conference: 3rd. International Conference on agricultural Engineering and Natural Resources At Iran.
Akhzari, D., and Pessarakli, M. (2017). Effects of vermicompost and urea fertilizers on qualitative and quantitative characteristics of Vetiveria zizanioides stapf. Grown under drought stress conditions. J. Plant Nutr. 40, 2063–2075. doi: 10.1080/01904167.2017.1346126
Akram, M. (2011). Growth and yield components of wheat under water stress of different growth stages. Bangladesh J. Agric. Res. 36, 455–468. doi: 10.3329/bjar.v36i3.9264
Ali, A., Sarwar, M., Khanzada, S., and Abro, G. H. (2009). Reaction of certain wheat varieties to the action of red flour beetle, Tribolium castaneum (Herbst) (Coleoptera) under insectary conditions. Pak. J. Zool. 41, 51–56.
Anee, T. I., Islam, M. N. N., Hassan, M. M., Masud, A. A. C., Alam, M. M., and Hasanuzzaman, M. (2022). Organic amendments improve plant morpho-physiology and antioxidant metabolism in mitigating drought stress in bread wheat (Triticum aestivum L.). Python Int. J. Exp. Bot. 91, 1959–1972. doi: 10.32604/phyton.2022.021137
Anjum, S. A., Ashraf, U., Zohaib, A., Tanveer, M., Naeem, M., Iftikhar, A. L. I., et al. (2017). Growth and developmental responses of crop plants under drought stress: a review. Zemdirbyste Agric. 104, 267–276. doi: 10.13080/z-a.2017.104.034
Anjum, S. A., and Tanveer, M. (2016). Effect of progressive drought stress on growth, leaf gas exchange, and antioxidant production in two maize cultivars. Environ. Sci. Pollut. Res. 23, 17132–17141. doi: 10.1007/s11356-016-6894-8
Arnon, D. I. (1949) Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 24, 1–15. doi: 10.1104/pp.24.1.1
Aslam, Z., and Ahmad, A. (2020). Effects of vermicompost, vermi-tea and chemical fertilizer on morpho-physiological characteristics of maize (Zea mays L.) in Suleymanpasa District, Tekirdag of Turkey. J. Innov. Sci. 6, 41–46. doi: 10.17582/journal.jis/2020/6.1.41.46
Aslam, Z., Ahmad, A., Abbas, R. N., Sarwar, M., and Bashir, S. (2023). Morpho-physiological, biochemical and yield responses of wheat (Triticum aestivum L.) to vermicompost, simple compost and NP fertilizer applications. Pak. J. Bot. 55, 2143–2154. doi: 10.30848/PJB2023-6(24)
Aslam, Z., Ahmad, A., Aziz, M. M., Mushtaq, Z., Zuhra, N., and Alasmarid, A. (2025). Impact of organic and inorganic sources of nutrition on the morpho-physiological, biochemical, yield traits and uptake of nutrients by mung bean (Vigna radiata L.). New Zeal. J. Crop Hort. Sci. 53, 2260–2284. doi: 10.1080/01140671.2025.2458816
Aslam, Z., Ahmad, A., Ibrahim, M., Iqbal, N., Idrees, M., Ali, A., et al. (2021). Microbial enrichment of vermicompost through earthworm Eisenia fetida (Savigny, 1926) for agricultural waste management and development of useful organic fertilizer. Pak. J. Agric. Sci. 58, 851–861. doi: 10.21162/PAKJAS/21.1378
Atiyeh, R. M., Edwards, C. A., Subler, S., and Metzger, J. D. (2000). Earthworm processed organic wastes as components of horticultural potting media for growing marigolds and vegetable seedlings. Compost Sci. Util. 8, 215–223. doi: 10.1080/1065657X.2000.10701994
Bellitürk, K., Aslam, Z., Ahmad, A., and Rehman, S. U. (2020). Alteration of physical and chemical properties of livestock manures by Eisenia fetida (Savigny, 1926) and developing valuable organic fertilizer. J. Innov. Sci. 6, 47–53. doi: 10.17582/journal.jis/2020/6.1.47.53
Briggle, L. W., and Curtis, B. C. (1987). “Wheat worldwide” in Wheat and wheat improvement. ed. E. G. Heyne (Madison, WI: American Society of Agronomy), 1–31.
Farooqi, Z. U. R., Qadir, A. A., Khalid, S., Murtaza, G., Ashraf, M. N., Javed, W., et al. (2024). Greenhouse gas emissions, carbon stocks and wheat productivity following biochar, compost and vermicompost amendments: comparison of non-saline and salt-affected soils. Sci. Rep. 14:7752. doi: 10.1038/s41598-024-56381-y
Farooq, M., Wahid, A., Kobayashi, N., Fujita, D., and Basra, S. M. A. (2009). Plant drought stress: effects, mechanisms and management. Agron. Sustain. Dev. 29, 153–188. doi: 10.1007/978-90-481-2666-8_12
Fathi, A., Shiade, S. R. G., Saleem, A., Shohani, F., Fazeli, A., Riaz, A., et al. (2025). Reactive oxygen species (ROS) and antioxidant systems in enhancing plant resilience against abiotic stress. Int. J. Agron. 2025:8834883. doi: 10.1155/ioa/8834883
García, A. C., Santos, L. A., Izquierdo, F. G., Sperandioa, M. V. L., Castroc, R. N., and Berbara, R. L. L. (2012). Vermicompost humic acids as an ecological pathway to protect rice plant against oxidative stress. Ecol. Eng. 47, 203–208. doi: 10.1016/j.ecoleng.2012.06.011
Gholipoor, M., Karamzadeh, A., and Gholami, A. (2011). Vermicompost as a soil supplement to relieve the effects of low-intensity drought stress on chickpea yield. Int Symp. Organ. Matter Manag. Comp. Use Horticult. 1018, 219–226. doi: 10.17660/ActaHortic.2014.1018.22
Hafez, E. M., Omara, A. E. D., Alhumaydhi, F. A., and El-Esawi, M. A. (2020). Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant. 172, 587–602. doi: 10.1111/ppl.13261
Hosseinzadeh, S. R., Amiri, H., and Ismaili, A. (2016). Effect of vermicompost fertilizer on photosynthetic characteristics of chickpea (Cicer arietinum L.) under drought stress. Photosynthetica 54, 87–92. doi: 10.1007/s11099-015-0162-x
Hosseinzadeh, S. R., Amiri, H., and Ismaili, A. (2018). Evaluation of photosynthesis, physiological, and biochemical responses of chickpea (Cicer arietinum L. cv. Pirouz) under water deficit stress and use of vermicompost fertilizer. J. Integr. Agric. 17, 2426–2437. doi: 10.1016/S2095-3119(17)61874-4
Hrčka, M., Kraus, K., Hřebečková, T., Tunklová, B., Kubeš, J., and Hanč, A. (2025). Effects of sewage sludge compost and vermicompost on wheat yield and vitality. Agriculture 15:551. doi: 10.3390/agriculture15050551
Huerta, E., Vidal, O., Jarquin, A., Geissen, V., and Gomez, R. (2010). Effect of vermicompost on the growth and production of amashito pepper, interactions with earthworms and rhizobacteria. Compost Sci. Util. 18, 282–288. doi: 10.1080/1065657X.2010.10736967
Hussain, M., Farooq, S., Hasan, W., Ul-Allah, S., Tanveer, M., Farooq, M., et al. (2018). Drought stress in sunflower: physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manag. 201, 152–167. doi: 10.1016/j.agwat.2018.01.028
Hussain, S., Sharif, M., Khan, S., Wahid, F., Nihar, H., Ahmad, W., et al. (2016). Vermicompost and mycorrhiza effect on yield and phosphorus uptake of wheat crop. Sarhad J. Agric. 32, 372–381. doi: 10.17582/journal.sja/2016.32.4.372.381
Ibrahim, M. U., Khaliq, A., Hussain, S., and Murtaza, G. (2021). Sorghum water extract application mediates antioxidant defense and confers drought stress tolerance in wheat. J. Plant Growth Regul. 41, 863–874. doi: 10.1007/s00344-021-10345-y
Iqbal, M., Asif, S., Ilyas, N., Raja, N. I., Hussain, M., Ejaz, M., et al. (2018). Smoke produced from plants waste material elicits growth of wheat (Triticum aestivum L.) by improving morphological, physiological and biochemical activity. Biotechnol. Rep. 17, 35–44. doi: 10.1016/j.btre.2017.12.001
Jahan, S., Ahmad, F., Rasul, F., Amir, R., and Shahzad, S. (2023). Physicochemical analysis of vermicompost-perlite based activated biochar and its influence on wheat (Triticum aestivum L.) growth under water stress. J. Soil Sci. Plant Nutr. 23, 3034–3050. doi: 10.1007/s42729-023-01258-8
Jan, S., Abbas, N., Ashraf, M., and Ahmad, P. (2019). Roles of potential plant hormones and transcription factors in controlling leaf senescence and drought tolerance. Protoplasma 256, 313–329. doi: 10.1007/s00709-018-1310-5
Jat, R. S., and Ahlawat, I. P. S. (2006). Direct and residual effect of vermicompost, biofertilizers and phosphorus on soil nutrient dynamics and productivity of chickpea-fodder maize sequence. J. Sustain. Agric. 28, 41–54. doi: 10.1300/J064v28n01_05
Ji, X., Shiran, B., Wan, J., Lewis, D. C., Jenkins, C. L. D., Condon, A. G., et al. (2010). Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant Cell Environ. 33, 926–942. doi: 10.1111/j.1365-3040.2010.02130.x
Joshi, R., Vig, A. P., and Singh, J. (2013). Vermicompost as soil supplement to enhance growth, yield and quality of Triticum aestivum L.: a field study. Int. J. Recycl. Org. Waste Agric. 2, 1–7. doi: 10.1186/2251-7715-2-16
Khakwani, A. A., Dennet, M. D., and Munir, M. (2011). Drought tolerance screening of wheat varieties by inducing water stress conditions. Songklanakarin J. Sci. Technol. 33, 135–142.
Kheir, A. M. S., Govind, A., Nangia, V., El-Maghraby, M. A., Elnashar, A., Ahmed, M., et al. (2025). Hybridization of process-based models, remote sensing, and machine learning for enhanced spatial predictions of wheat yield and quality. Comput. Electron. Agric. 234:110317. doi: 10.1016/j.compag.2025.110317
Kimber, G., and Sears, E. R. (1987). “Evolution in the genus Triticum and the origin of cultivated wheat” in Wheat and wheat improvement. ed. E. G. Heyne (Madison, WI: American Society of Agronomy), 31.
Kiran, S. (2019). Effects of vermicompost on some morphological, physiological and biochemical parameters of lettuce (Lactuca sativa var. crispa) under drought stress. Not. Bot. Horti. Agrobo. 47, 352–358. doi: 10.15835/nbha47111260
Kumar, P., Yadav, R. K., Gollen, B., Kumar, S., Verma, R. K., and Yadav, S. (2011). Nutritional contents and the medicinal properties of wheat: a review. Life Sci. Med. Res. 22, 1–10.
Lawlor, D. W. (2002). Limitation to photosynthesis in water stressed leaves: stomata versus metabolism and the role of ATP. Ann. Bot. 89, 871–885. doi: 10.1093/aob/mcf110
Lazcano, C., Sampedro, L., Zas, R., and Dominguez, J. (2010). Assessment of plant growth promotion by vermicompost in different progenies of maritime pine (Pinus pinaster Ait.). Compost Sci. Util. 18, 111–118. doi: 10.1080/1065657X.2010.10736943
Lu, Q., Lu, C., Zhang, J., and Kuang, T. (2002). Photosynthesis and chlorophyll a fluorescence during flag leaf senescence of field-grown wheat plants. J. Plant Physiol. 159, 1173–1178. doi: 10.1078/0176-1617-00727
Mahmoudi, S. V. R., Nasri, M., and Azizi, P. (2016). The effect of different type of vermicompost organic fertilizer litter on quantitative, qualitative and biochemical characteristics of green mung bean (Vigna radiata L.) in drought stress conditions in Varamin. Int. J. Adv. Biotechnol. Res. 7, 205–215.
Prassad, P. V. V., Staggeborg, S. A., and Ristic, Z. (2008). Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plant. In:L. Ahuja, V Reddy, S. Saseendran, and Q. Yu, (Eds.), Response of crops to limited water: understanding and modeling water stress effects on plant growth processes, American society of agronomy, Inc., Madison, 1, 301–355. doi: 10.2134/advagricsystmodel1.c11
Ramazanoglu, E. (2024). Effects of vermicompost application on plant growth and soil enzyme activity in wheat (Triticum aestivum L.) monitored by thermal imaging. Cogent Food Agric. 10:2373872. doi: 10.1080/23311932.2024.2373872
Rashtbari, M., Hossein, A. A., and Ghorchiani, M. (2020). Effect of vermicompost and municipal solid waste compost on growth and yield of canola under drought stress conditions. Commun. Soil Sci. Plant Anal. 51, 2215–2222. doi: 10.1080/00103624.2020.1820023
Sarwar, M., and Sattar, M. (2007). Varietals assessment of different wheat varieties for their resistance response to Khapra beetle Trogoderma granarium. Pak. J. Seed Technol. 1, 1–7.
Seimrizade, S., Moshatati, A., Bakhshandeh, A. A., Khodaei, J. A., and Koochekzadeh, A. (2021). The effect of vermicompost on yield and yield components of wheat under terminal heat stress conditions in Ahwaz. Environ. Stresses Crop Sci. 14, 1139–1145. doi: 10.22077/escs.2020.3384.1853
Shahbazi, S., Fateh, E., and Aynehband, A. (2015). Evaluation of the effect of humic acid and vermicompost on yield and yield components of three wheat cultivars in tropical regions. Plant Prod. 38, 99–110. doi: 10.22055/ppd.2015.11323
Siddiqui, Q. H., and Sarwar, M. (2002). Pre and post harvest losses in wheat. Pak. Gulf Econ. 6, 30–32.
Steel, R. G. D., Torrie, J. H., and Dickey, D. (1997). Principles and procedures of statistics, a biometrical approach. 3rd Edn. New York: McGraw Hill, Inc. Book Co., 352–358.
Talaat, N. B., and Abdel-Salam, S. A. (2024). A novel eco-friendly approach of combining vermicompost and effective microorganisms sustains wheat (Triticum aestivum L.) drought tolerance by modulating photosynthetic performance and nutrient acquisition. Acta Physiol. Plant. 46:76. doi: 10.1007/s11738-024-03698-w
Todaka, D., Zhao, Y., Yoshida, T., Kudo, M., Kidokoro, S., and Mizoi, M. (2017). Temporal and spatial changes in gene expression, metabolite accumulation and phytohormone content in rice seedlings grown under drought stress conditions. Plant J. 90, 61–78. doi: 10.1111/tpj.13468
Keywords: drought levels, field experiment, straw, vermicompost, wheat cultivars
Citation: Ahmad A, Aslam Z, Arshad MA, Zulfiqar U, El-Beltagi HS, Alshaharni MO and Prasad PVV (2025) Vermicompost-mediated modulation of agronomic and physiological traits enhances wheat performance under variable water regimes. Front. Sustain. Food Syst. 9:1695055. doi: 10.3389/fsufs.2025.1695055
Edited by:
Mohamed Ait-El-Mokhtar, University of Hassan II Casablanca, MoroccoReviewed by:
Ashraf E. Hamdy, Al-Azhar University, EgyptMuhammad Nadeem Shah, Government College University, Lahore, Pakistan
Copyright © 2025 Ahmad, Aslam, Arshad, Zulfiqar, El-Beltagi, Alshaharni and Prasad. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Usman Zulfiqar, dXNtYW4uenVsZmlxYXJAaXViLmVkdS5waw==; Zubair Aslam, emF1YWZAZ21haWwuY29t; Hossam S. El-Beltagi, aGVsYmVsdGFnaUBrZnUuZWR1LnNh
†Present address: Ali Ahmad, Arid Zone Research Institute (AZRI), Bhakkar, Pakistan