 Shumin Liu
Shumin Liu Zhiheng Li
Zhiheng Li Alida M. Bailleul
Alida M. Bailleul Min Wang
Min Wang Jingmai O’Connor
Jingmai O’Connor- 1Key Laboratory of Vertebrate Evolution and Human Origins, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China
- 2University of the Chinese Academy of Sciences, Beijing, China
- 3CAS Center for Excellence in Life and Paleoenvironment, Beijing, China
- 4Field Museum of Natural History, Chicago, IL, United States
Gastroliths, where preserved, can provide indirect evidence regarding diet in extinct avian and non-avian dinosaurs. Masses of gastroliths consistent with the presence of a gastric mill are preserved in many Early Cretaceous Jehol birds mostly belonging to the Ornithuromorpha. Gastroliths are also present in basal birds Sapeornis and Jeholornis in which herbivory is supported by direct evidence these taxa consumed seeds in the form of crop or stomach contents. Although gastroliths have been correlated with herbivory in non-avian dinosaurs, the presence of gastroliths and bone together in Ambopteryx calls this association in to question. Despite being known from greater numbers of specimens than other avian lineages, no unequivocal direct or indirect evidence of diet has been recovered from Jehol deposits for the Enantiornithes. A referred specimen of Bohaiornis guoi IVPP V17963 was described as preserving a small number of gastroliths interpreted as rangle, gastroliths whose function is cleaning the stomach in extant raptorial birds. However, based on comparison with gastroliths in other Jehol birds, it has alternatively been suggested that the identified structures are not ingested stones at all but some unusual mineral precipitate. Considering the limited evidence regarding diet in Enantiornithes and the importance of accurately identifying the traces in Bohaiornis in order to understand the enantiornithine digestive system, we extracted two samples of these purported gastroliths and explored these traces using computerized laminography scanning, scanning electron microscopy, energy dispersive x-ray spectroscopy, ground sections, and body size to gastral mass regressions. Similar analyses were conducted on gastroliths extracted from undisputed gastral masses of two Jehol ornithuromorphs and the non-avian pennaraptoran Caudipteryx. The combined results contradict the hypothesis that these traces are gastroliths and supports the interpretation they are mineral precipitate, most likely authigenic quartz (chalcedony). Although authigenesis is commonly responsible for the preservation of soft tissues, it is unclear if these traces record part of the tissues of this Bohaiornis. This study highlights the importance of a multidisciplinary approach in understanding unusual traces in the fossil record and reveal a previously unidentified taphonomic phenomenon in fossils from Jehol deposits.
Introduction
The digestive system of living birds is highly modified to meet the high caloric demand of flight within an aerodynamically light weight structure (Gill, 2007). Highly efficient food processing capabilities provide birds with the necessary calories to support powered flight, the most energetically costly form of vertebrate locomotion. The neornithine digestive system is characterized by several unique features not found in crocodilians, the other extant clade of archosaurs: many modern birds possess a crop, an expansion of the esophagus used to temporarily store excess food; and the stomach is divided into two chambers (Gill, 2007). The cranial chamber, the proventriculus, produces acid to dissolve ingested items, and is especially large in carnivorous raptorial birds who chemically digest their food. The caudal chamber, the ventriculus or gizzard, in many taxa has a specialized grinding function in which a large number of stones are intentionally ingested to aid in the digestion of hard food items (e.g., seeds, insects) forming a structure referred to as a gastric mill (Duke, 1986). Notably, although raptorial birds lack a gastric mill, they have also been observed intentionally ingesting stones (Albuquerque, 1982). This small number of stones is referred to as rangle and is thought to help clean the digestive system (Fox, 1976).
Most data currently available concerning the diversity and physiology of Cretaceous birds comes from the Lower Cretaceous volcanolacustrine Jehol deposits in north-eastern China (Xu et al., 2020). These Lagerstätten have produced an abundance of fossil birds whose small and hollow bones only rarely survive diagenesis in most other depositional environments. Most reported specimens are largely complete and articulately and many preserve rare traces such as soft tissues (e.g., feathers, lungs, ovarian remains) (Ji et al., 1998; O'Connor et al., 2013; Wang et al., 2018) and ingested remains (e.g., stomach and crop contents) (Zheng et al., 2011; O'Connor, 2019).
Large numbers of gastroliths preserved in the abdominal cavity of some dinosaur groups suggests these taxa had evolved a gastric mill (Ji et al., 1998; Zhou and Wang, 2000; Ji et al., 2003; Wings, 2007; Cerda, 2008; Varricchio et al., 2008; Xu et al., 2009; Makovicky et al., 2011; Choiniere et al., 2012; Wang et al., 2019). The phylogenetic distribution of these groups suggests that the two-part stomach evolved early in the Dinosauria. Large clusters of gastroliths consistent with their identification as gizzard stones forming a gastric mill have been described in the long boney tailed bird Jeholornis (O'Connor et al., 2018), the basal pygostylian Sapeornis (Zhou and Zhang, 2003; Zheng et al., 2011), and numerous Jehol ornithuromorphs (e.g., Archaeorhynchus, Eogranivora, Bellulornis, Changzuiornis and Iteravis) (Zhou and Zhang, 2006; Zhou et al., 2013; Zhou et al., 2014; Wang et al., 2016b; Huang et al., 2016; Wang and Zhou, 2016; Wang et al., 2018; Zheng et al., 2018). Interpretations that gastral masses were utilized to digest hard plant material is supported by direct evidence of ingested seeds in three lineages (Sapeornis, Jeholornis, and Eogranivora) (Zhou and Zhang, 2002; Zheng et al., 2011; O'Connor et al., 2018; Zheng et al., 2018). However, ornithuromorphs preserve a diversity of gastral mass morphotypes and the presence of gastroliths and an herbivorous diet may not strictly correlate in this clade, as is true of Neornithes (O'Connor, 2019). This is somewhat supported by a pellet apparently containing bone found associated with the holotype of Iteravis (O'Connor and Zhou, 2019).
Enantiornithines are considered the dominant clade of Cretaceous land birds (O'Connor, 2009; O’Connor et al., 2011; Wang and Zhou, 2017a). Specimens referable to this group account for approximately half of all known Cretaceous birds. Although they have a global distribution being recovered from every continent with the exception of Antarctica (O’Connor et al., 2011), the largest source of enantiornithine fossils is the Jehol Biota, where hundreds if not thousands of specimens have been collected (Zhou, 2006). The remains of enantiornithines in the Huajiying Formation, the oldest geologic unit preserving the Jehol Biota, represents their earliest known record (Wang et al., 2017); the diversity of enantiornithines increases in younger deposits of the Jehol Biota, being most diverse in the Jiufotang Formation (Wang and Zhou, 2017b).
The most diverse clade of enantiornithines is the Bohaiornithidae, which includes six taxa: Bohaiornis guoi, Parabohaiornis martini, Shenqiornis mengi, Sulcavis geeorum, Longusunguis kurochkini, and Zhouornis hani (Wang et al., 2010; Wang et al., 2014; Zhang et al., 2014; Li et al., 2015). This group is characterized by their fairly large size, robust dentition, and large pedal claws.
Exceptional specimens from the Jehol Biota have allowed scientists to begin to reconstruct the structure of the digestive system in basal birds. These specimens indicate that a digestive system similar to Neornithes (modern birds) in terms of structure and digestive abilities is present in the Ornithuromorpha (O'Connor, 2019). With the exception of the fact teeth are retained by some taxa, the digestive system of Jehol ornithuromorphs appears similar to that of modern birds in the presence of crop and grinding gizzard and the capacity for bidirectional peristalsis including the regurgitation of pellets (Wang et al., 2016a; Zheng et al., 2018).
No unequivocal record of gastrolith use in the Enantiornithes exists so far. The only described specimen purportedly preserving this feature is a referred specimen of Bohaiornis IVPP V17963 (Li et al., 2015). This specimen preserves two small masses of rounded objects that are superficially similar to gastroliths (Figures 1A,C). Based on their small size and low number, Li et al. (2015) interpreted these traces as gastroliths and likened them to rangle, the small numbers of stones intentionally ingested by extant raptorial birds (Fox, 1976; Li et al., 2015). However, based on comparison with gastroliths preserved in other Jehol birds, O’Connor (2019) argued that these traces did not represent stones but rather some strange form of mineral precipitate (O'Connor, 2019). Understanding these traces will lead to a better understanding of diet, behavior, and digestive function in enantiornithines. In order to test these two competing hypotheses, we utilized scanning electron microscopy (SEM), energy dispersive spectroscopy (EDS), traditional histology, computed laminography (CL) and the relationship between body mass and gastrolith mass in birds to further investigate the identity of the purported gastroliths in Bohaiornis IVPP V17963.
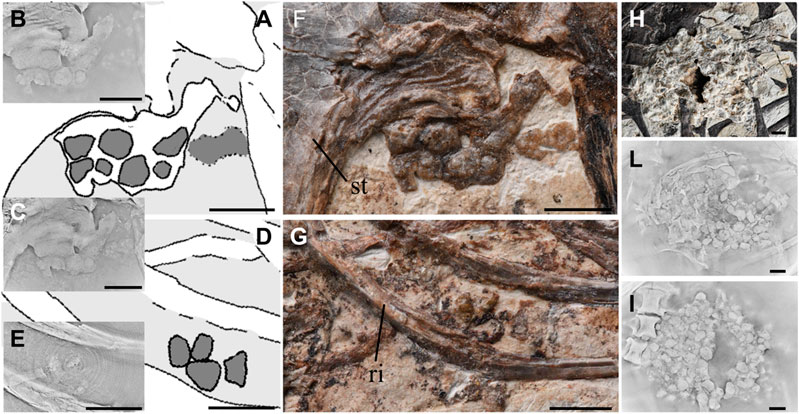
FIGURE 1. Description and comparison of purported gastroliths and true gizzard stones further illuminated with the aid of CL. (A,D). Line drawing from Li et al. (2015) of the purported gastroliths of Bohaiornis IVPP V17963 near the sternum (A) and between the thoracic ribs (D) (Li et al., 2015); (B,C). CL scans results of gastroliths depicted in (A) from two different levels showing the interconnectedness of the purported gastroliths; (E). CL scans result corresponding to (D) depicting the weak impressions of the purported gastroliths; (F). Photograph of purported gastroliths of Bohaiornis IVPP V17963 near the sternum; (G). Photograph of purported gastroliths of Bohaiornis IVPP V17963 between two thoracic ribs; (H). Photograph of the gastral mass preserved Archaeorhynchus IVPP V17075; (L,I). Two location of CL scans corresponding to (H), both revealing distinctly separated individual gastroliths. Anatomical abbreviations: st, sternum; ri, rib. Scale bar equals five milimeters.
Methods
Bohaiornis IVPP V17963 was studied first under direct observation and then CL scanned at 7.83 μm resolution using the Xm-Tracer-CL-160 at the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP). Two samples of the purported gastroliths were then extracted for additional analysis. Definitive gastroliths from Archaeorhynchus IVPP V17075 were also CL scanned at 18.71 μm resolution. Gastroliths were extracted from Archaeorhynchus IVPP V17075, Iteravis IVPP V23346 and Caudipteryx STM (Shandong Tianyu Museum of Nature) 4–5 for detailed comparative analysis.
GROUND-SECTIONING Ground sections were made from the extracted gastroliths of all four specimens.
All extracted gastrolith samples were embedded in EXAKT Technovit 7,200 1-component resin, placed in a vacuum for 1 h to remove bubbles, and then cured for a minimum of 8 h in the EXAKT Technovit 7,200. The resin blocks were cut using an EXAKT 300CP accurate circular saw. The samples were then ground down using 500# sand paper until the sample was exposed and then polish with 4,000# sand paper, both using the EXAKT 400CS grinding system.
The blocks were placed back in the vacuum for 5 min after which they were adhered to slides and placed under UV light for 10 min. The slides were further polished until the desired optical contrast was reached at approximately 45 μm thickness.
The slides were observed using a Leica DM-RX polarized light microscope under both normal and polarized light. Pictures are taken with a DS-Fi3 camera using the software NIS-Element v4.60.
SEM-EDS SEM images were taken using the Merlin Compact Ultra-high resolution field scanning electron microscope at the Chinese Academy of Geological Sciences (Beijing, China) using FEI Quanta 450 (FEG) at 20 kv. Samples were coated in gold prior to imaging and EDS line scanning and mapping scanning.
Body:gastrolith mass correlation Measurements of Bohaiornis IVPP V17963 were taken from Li et al. (2015). The regression formula for estimating birds body mass was adopted from Serrano et al. (2015). Correlations between body mass and gastrolith mass in birds were taken from Wings (2007). Numerous methods for estimating body mass of stem birds have been developed (Butler and Goseami, 2008; Liu et al., 2012; Field et al., 2013; Serrano et al., 2015; Serrano et al., 2016). The methods by Serrano et al. (2015) utilized data acquired from modern birds to build multiple regression functions for each of the major stem avian lineages. This model has greater predictive strength and applicability (Serrano et al., 2015). We used the regressions of Serrano et al. (2015) to estimate the body mass of Bohaiornis IVPP V17963 in order to determine if the purported gastroliths are proportionate to the bird’s body mass (Table 1).
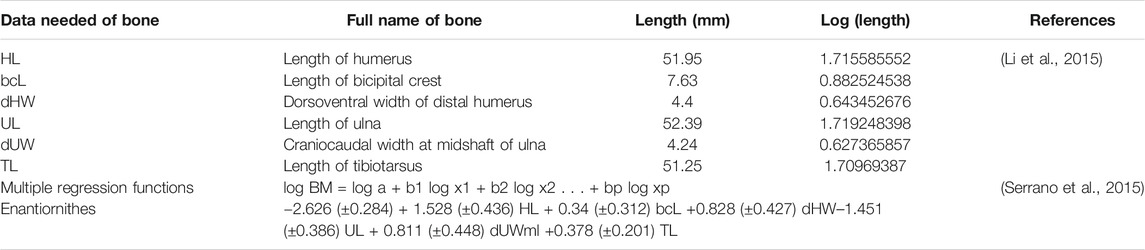
TABLE 1. Data and formula to calculate the body mass of Bohaiornis guoi IVPP V17963.
Results
Raw Morphology
As described by Li et al. (2015), Bohaiornis IVPP V17963 preserves a small number of stone-like objects in two clusters: one located near the sternum and the other between two thoracic ribs approximately level with the cranial end of the synsacrum (Figure 1) (Li et al., 2015). The color of these purported gastroliths is a dark brown (Figures 1F,G) that is very similar to the bone preserved in Bohaiornis, which is unusual when compared to other gastral masses found in Jehol birds in which the gastroliths are often a range of colors and are typically clearly distinct from the color of the fossilized bone (Figure 2). The purported gastroliths near the sternum superficially appear to be very rounded and circular.

FIGURE 2. Comparison of purported gastral masses in Early Cretaceous Jehol paravians and birds. (A). Caudipteryx STM4-5 (Oviraptorosauria); (B). Jeholornis STM2-15 (Jeholornithiformes); (C). Archaeorhynchus IVPP V17075 (Ornithuromorpha); (D). Bohaiornis IVPP V17963 (Enantiornithes); (E). Iteravis IVPP V23346 (Ornithuromorpha); (F). Archaeorhynchus IVPP V14287 (Ornithuromorpha); (G). Dingavis IVPP V20284 (Ornithuromorpha); (H). Bellulornis IVPP V17970 (Ornithuromorpha); Anatomical abbreviations: fe, femur; ga, gastralia; gs, gastroliths; hu, humerus; il, ilium; is, ischium; pb, pubes; pu, pubis; py, pygostyle; ri, rib; sc, scapula; sr, sternal rib; st, sternum; sy, synsacrum; tb, tibiotarsus; tr, thoracic rib; tv, thoracic vertebra. Scale bar equals 1 cm.
When observing the mass of purported gastroliths adjacent to the sternum, two spherical structures are visible just below the purported caudal margin of the sternum. These are clearly visible under direct observation (Figure 1A) and in some CL scan slices (Figure 1B). An additional two slightly smaller spherical structures are visible below the two larger structures (Figure 1A) but again are not visible in all CL slices (Figure 1C). The CL data reveals that all of the purported gastroliths are connected, in some cases even with areas adjacent that were previously interpreted as bone continuous with the sternum. Although superficially circular structures are visible, these represent areas of higher relief and upon closer examination the purported gastroliths from a large amorphous structure that varies in thickness, without distinct boundaries (Figures 1A,C). This includes areas located adjacent to the purported gastroliths that were not identified as gastroliths by Li et al. (2015), but exhibit a similar morphology both through direct observation and in the CL scans (Figures 1A,C) suggesting they have a similar taphonomic origin.
The group of purported gastroliths located between two thoracic ribs are smaller and ill defined (Figure 1D). They are only faintly visible in the CL scans (Figure 1E). These more distally located purported gastroliths are much thinner (visible in 55 CL slices) compared to those near the sternum (visible in 164 slices).
In comparison, gastroliths preserved in Archaeorhynchus IVPP V17075 (Figures 1F–H,2C) form a tight circular aggregate and are embedded in some matrix that has not been prepared away to completely reveal the surfaces of the gastroliths, as in some other specimens (Archaeorhynchus IVPP V17091). The individual stones can be seen readily distinguished in the CL scans. The white material surrounding the gastroliths has a lower density compared to the gastroliths themselves.
The morphology of the gastroliths in Caudipteryx STM4-5 (Figure 2A) are similar to those preserved in Archaeorhynchus IVPP V17075, although much of the fine-grained white material in which the Archaeorhynchus gastroliths are embedded has been removed, presumably during mechanical preparation in order to expose the surface of the gastral mass. The mass is crescent-shaped with the convex margin facing the ventral surface of the body and a mineralized reddish layer covers parts of the exposed surface. Gastroliths in Iteravis IVPP V23346 (Figure 2E) are scattered–not preserved in close association with each other forming an aggregate. The individual gastroliths are black, polished, and mostly subrounded to rounded with moderate sphericity, although some gastroliths exhibit a subrectangular morphology. The same morphology is observed in Dingavis IVPP V20284 (Zhou et al., 2014).
Analyses of Purported Gastroliths of Bohaiornis guoi IVPP V17963
Two purported gastroliths, revealed by CL scans to be connected, were extracted and cut into ground sections (Figures 3A,B,F). The ground sections reveal that the two circular structures, with diameter of 0.683 mm and 0.793 mm respectively, each consist of fibral-like chalcedony (Figures 3H,K) in a spherulitic arrangement which are embedded in a microcrystalline matrix (Figures 3A,B). The fibers radiate from the central dark region, forming four to five petal-shaped crystals (Figure 3C). The circular structures exhibit order Ⅰ aurantium of the highest interference color and the fibers exhibit radiated extinction under cross polarized light (Figures 3A,B). The outer region lining one margin of the quartz spherulites consists of a fine-grained matrix (Figure 3C).
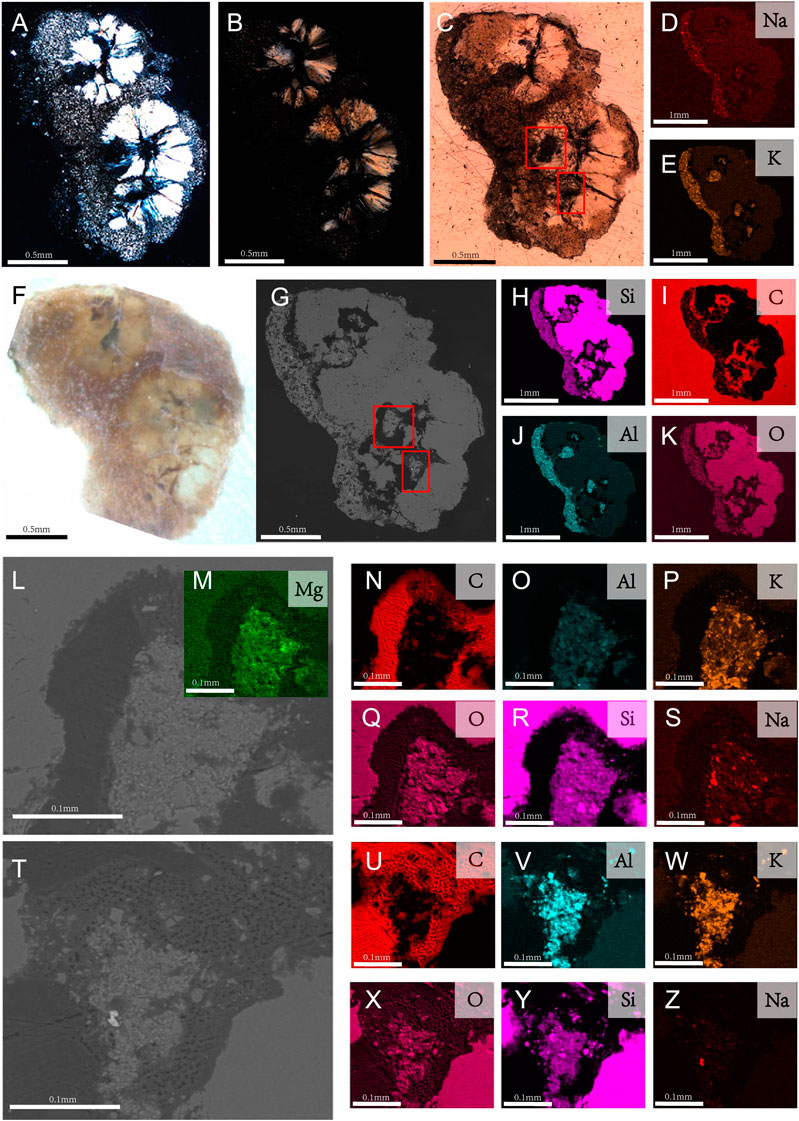
FIGURE 3. Lithology results and EDS element mapping of the ground-sections of the purported gastroliths of Bohaiornis guoi IVPP V17963. (A,B). Photographs under polarized light through microscope; (A). In high brightness; (B). In low brightness; (C). Photograph under natural light; (D,E,H–K). Elemental mapping results of the whole slide. Its major elements include sodium (Na), potassium (K), silicon (Si), carbon (C), aluminum (Al), oxygen (O); (F). Photograph under Stereo Microscope; (G). SEM photograph. (I–Z). Close up photographs of intermediate region of purported gastroliths; Close up regions (L,T) are the regions boxed in (C,G). (M–S). Elements mapping of (L); (U–Z). Elements mapping of (T).
The purported gastroliths are embedded in carbon (C) resin (Figure 3I). Carbon can also be observed within the purported gastroliths which may suggest some original organic material formed the nucleus on which the quartz fibers precipitated (Figures 3I,N,U). However, the possibility that the C in the intermediate region is resin that permeated into the sample during ground-sectioning can’t be excluded (Figure 3). The EDS profile of the purported gastroliths may support interpretations that the C in the internal region is from the resin because it shows the same high content of C as the resin surrounding the sample (Figure 4). However, the fact that light does not penetrate these regions as it does in the resin surrounding the sample may support the alternative hypothesis that the C is original organic material. This internal C surrounds small pockets that contain Magnesium (Mg), Aluminium (Al), Potassium (K), and traces of Sodium (Na) in addition to Silica (Si) and Oxygen (O) (Figures 3M–Z). The microcrystalline region that forms the left outer margin of the two purported gastroliths (as pictured in Figure 3) similarly contains Aluminum (Al), Potassium (K), and traces of Sodium (Na) in addition to Silica (Si) and Oxygen (O) (Figures 3D,E,H,J,K). Despite their similar element content, the morphology of the inner pockets and the outer left margin under the microscope are completely different with the inner pockets being black in thin section, without light passing through, whereas the external margin has a microcrystalline morphology.
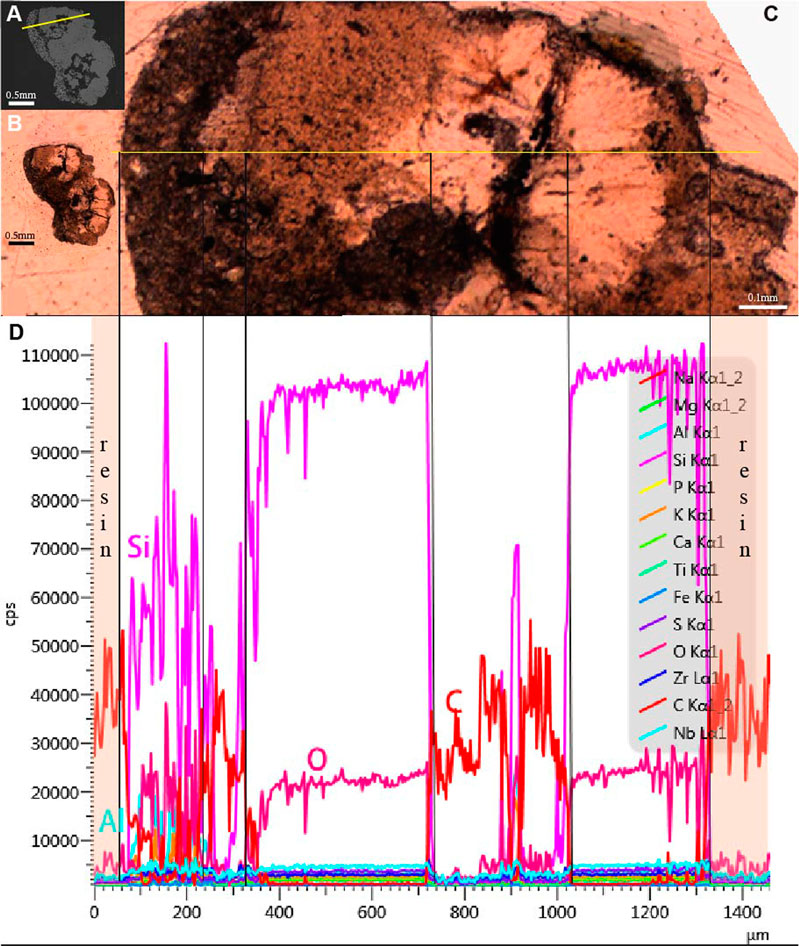
FIGURE 4. EDS profile of the purported gastroliths extracted from Bohaiornis guoi IVPP V17963. (A). SEM photograph with yellow line indicating where the EDS profile was taken; (B). Ground section under microscope with natural light; (C). Enlargement of the cross-section marked in A with the cross-section leveled horizontal; (D). The EDS profile of the line marked in A and C revealing the chemical composition of major elements of the sample in counts per second (cps). Content of Si, O in the interior part are higher than that in the exterior part and in the intermediate region. Content of C in the intermediate region are the same as that of the resin.
Analyses and Comparison of Paravian Gastroliths
The gastrolith ground section from Caudipteryx STM4-5 was identified under the microscope as fine-grained sandstone, which consists of small quartz particles and a minor portion of mica (Figure 5A). The long axis of mineral particle size ranges from 0.1 mm to 0.4 mm. The quartz particles are characterized by an order Ⅰ yellowish white interference color and parallel extinction in cross polarized light. The gastrolith extracted from Iteravis IVPP V23346 was identified as consisting entirely of quartzite, with the main minerals being quartz and the main component being cement (Figure 5B); the long axis of the mineral particle size measures approximately 0.025 mm, and minerals exhibit an even distribution with quartz accounting for approximately 50%. The gastrolith extracted from Archaeorhynchus IVPP V17075 was identified as an igneous rock containing feldspar and biotite, with a porphyritic structure (Figure 5C); the main body of gastrolith is groundmass with serious weathering traces and some brownish substances. Minerals show order Ⅰ white of the highest interference color and oblique extinction in cross polarized light. These features indicate that their source is probably weathered basalt.
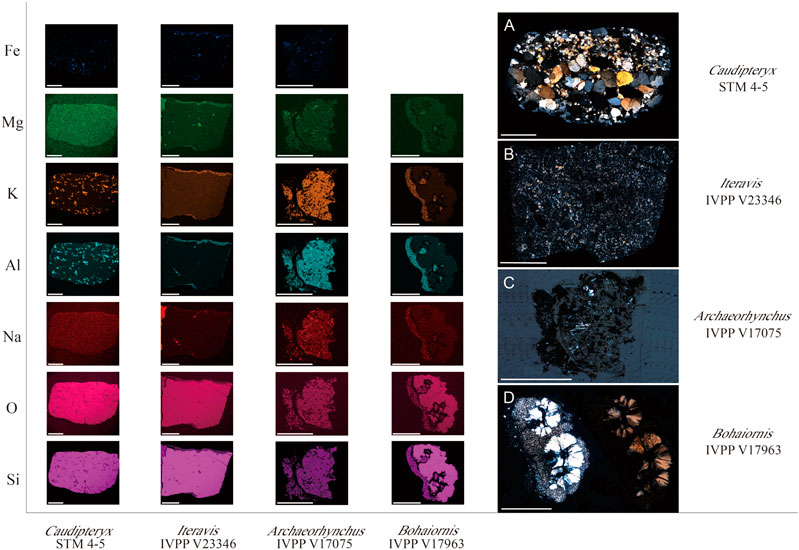
FIGURE 5. EDS element mapping and ground-sectioning slides of four extracted gastrolith samples. Photographs on the right side are ground sections of samples under polarized microscope. Scale bar equals 1 mm.
The elemental composition of the gastrolith from Caudipteryx STM4-5 has an even distribution from the interior to exterior. The gastrolith primarily consists of Si, O and Na, with smaller amounts of Al, K, Mg and Fe (Figure 5A). The gastrolith extracted from Iteravis IVPP V23346 mainly contains Si and O, with small amounts of Na, Al, Mg and Fe. Some regions along the edge form a thin layer that contains less Si and O and more Al, Mg and Na compared to the main body of the gastrolith. The Archaeorhynchus IVPP V17075 gastrolith is also composed primarily of Si and O, with Al, Mg, Na and K content higher than that of the other gastroliths studied here (Figure 5C).
EDS indicates the most common elements among the four samples analyzed here are Si and O, which have the highest content of all the identified elements in all the studied samples. Gastroliths of Caudipteryx STM4-5 and Iteravis IVPP V23346 are rock types in which quartz is the main component. The gastrolith of Archaeorhynchus IVPP V17075 is an igneous rock composed of Si and O. All samples also contain Na, Al, K and Mg. The samples from Iteravis IVPP V23346 and Bohaiornis IVPP V17963 both exhibit an outer layer on some margins that has a higher content of Mg, K, Al, and Na compared to the main body of the gastrolith. None of the true gastroliths show similar internal structures high in C and other minerals like that observed in the purported gastroliths extracted from Bohaiornis.
Size Correlation
The body mass of Bohaiornis IVPP V17963 is estimated to be approximately 308 g based on measurements in Li et al. (2015). Published data indicates that gastroliths mass account for 1% of the body mass, so the gastroliths of Bohaiornis should measure 3.08 g (Wings, 2004). Reports of rangle in New Zealand falcons (Falco novaeseelandiae) indicate the stones account for about 3.3% of the body mass in females and 3.0% in males (Fox, 1976). This would suggest that if the purported gastroliths in Bohaiornis IVPP V17963 represent rangle they should have a mass of approximately 10.14 g. Since the entire purported gastral mass of Bohaiornis could not be extracted, we compared the mass in the fossils to a piece of chert (Figure 6). The chemical composition of chert is mainly Si and O, which has a density of 2.53 g/cm3 which is higher than that of pure quartz. The chert sample weighing 3.09 g is much bigger than the combined volume of the purported gastroliths, which suggests the weight of purported gastroliths are far less than 3.08 g. Therefore, the purported gastroliths do not satisfy the relationship ratio between gastrolith mass and body mass, either as rangle or as gizzard stones.

FIGURE 6. Volume comparison between the purported gastroliths of Bohaiornis guoi IVPP V17963 and a 3.09 g piece of chert. Red rectangle indicates areas where the purported gastroliths occur. (A) close up photograph; (B) photograph of the entire specimen. Scale bar equals 1 cm.
It is possible that the preserved mass of “purported gastroliths” is incomplete and that some gastroliths were taphonomically lost. However, even to meet 1% of the body mass (one third the expected mass if these traces are indeed rangle), the purported gastral mass would have to be several orders of magnitude greater. Such a loss of gastrolith mass is highly unlikely in light of the fact that Bohaiornis IVPP V17963 is largely complete, even preserving delicate bones like gastralia (Li et al., 2015), and articulated, suggested minimal disruption of the carcass after death, consistent with a majority of described specimens from the Jehol, which are preserved in a static (low energy) lacustrine environment (Zhang and Miman, 2003).
Discussion and Comparison
Despite the large number of recovered specimens, traces of diet in enantiornithines (Aves: Ornithothoraces) are very rare (Sanz et al., 1996; Dala Vecchia and Chiappe, 2002; Cau and Arduini, 2008; Li et al., 2015; Wang et al., 2016a). Such traces have only been reported in two Jehol specimens both of which have subsequently been questioned in the literature (O'Connor, 2019; O'Connor and Zhou, 2019). Structures in Bohaiornis IVPP V17963 identified as gastroliths ingested for use as rangle, a specific form of gastral mass utilized by raptorial birds to clean the digestive system (Li et al., 2015), were alternatively interpreted as some form of mineral precipitate (O'Connor, 2019). In order to shed new light on this academic debate, we utilized numerous methods including CL scans, ground-sections, lithology analysis, and SEM-EDS on samples extracted from IVPP V17963 and two other Jehol birds (Iteravis IVPP V23346 and Archaeorhynchus IVPP V17075) and one non-avian pennaraptoran (oviraptorosaur Caudipteryx STM4-5) with distinct masses of gastroliths interpreted without controversy as forming a gastric mill. We consider the various lines of evidence independently below.
Anatomical Location
The preserved location of the purported gastral mass in Bohaiornis IVPP V17963 is consistent with their original identification that they are ingested stones. The purported gastroliths occur in two places, one patch along the caudal margin of the sternum and a second patch more distally located between two thoracic ribs, approximately level with the cranial margin of the synsacrum. This is generally consistent with the position of the stomachs. If these two patches are mineral precipitate, the mechanism responsible for the silicification of only these two small patches of soft tissue and the tissues they may represent are equally unknown.
Comparison of Gastral Mass and Macroscopic Gastrolith Morphology
Superficially, gastral mass morphology in Jehol paravians can be roughly divided into three types: 1) a large number of proportionately small gastroliths that are tightly aggregated together, presumably held together by stomach contents due to the tightly packed nature of their association in vivo–this morphology is observed in Caudipteryx (Figure 2A) (Ji et al., 1998; Zhou and Wang, 2000; Zhou et al., 2000), Jeholornis (Figure 2B) (O'Connor et al., 2018), and Archaeorhynchus (STM7-11, IVPP V17075, IVPP V17091) (Zhou et al., 2013; Wang et al., 2018); 2) a small number of proportionately larger stones found in loose association–this morphology is observed in Iteravis IVPP V23346 (Figure 2E), Dingavis IVPP V20284 (Figure 2G) (O'Connor et al., 2016) and Hongshanornis DNHM D2945 (Chiappe et al., 2014); and 3) a small number of proportionately larger stones preserved tightly compacted together, protruding from the surface of the slab–this morphology is observed in Bellulornis IVPP V17970 (Figure 2H) (Wang et al., 2016b). In addition, this gastral mass morphology is also found in Gansus yumenensis (Wang et al., 2015) from the Early Cretaceous Xiagou Formation in Gansus Province, northwestern China.
The purported gastroliths in Bohaiornis IVPP V17963 are few in number and proportionately smaller, not fitting any of the gastral mass morphology types described above. As observed by O’Connor (2019), they are morphologically distinct from gastral masses preserved in other Early Cretaceous birds (O'Connor, 2019). However, the purported gastral mass in Bohaiornis IVPP V17963 was interpreted as rangle, which is morphologically distinct from gastral masses in herbivorous and omnivorous birds. Thus the, unusual size of the gastral mass does not exclude the original interpretation from being a viable hypothesis. However, although reported by Li et al. (2015) that the amount of stones ingested is typically very small, literature suggests otherwise; rangle reportedly forms a greater percentage of body mass (3–3.3%) (Fox, 1976) than normal gizzard stones (1%) (Wings, 2004). Thus, if these percentages are correct, the size of the mass of purported gastroliths is inconsistent with their interpretation as rangle.
However, upon closer inspection the shape of the purported gastroliths is unusual for gizzard stones. Some of the purported gastroliths superficially appear spherical in shape, similar to the rounded to subrounded gastroliths found in Caudipteryx, Mesozoic birds like Iteravis and Archaeorhynchus, and neornithines. However, closer examination reveals that the purported gastroliths are actually more similar to circular discs, being very thin, and the two superficially separated circular structures in fact form one amorphous structure connected by thinner regions of mineral that lack the spherulitic structure of the silica fibers and contain higher elemental diversity. This connection is visible both in the CL data and in the ground sections. In contrast, although the gastroliths in Archaeorhynchus IVPP V17075 are bound together by matrix, the individual gastroliths can be readily distinguished as separate structures in the CL data (Figure 1B). The purported gastroliths located between the two thoracic ribs form only weak impressions (Figures 1D,E) and it is unclear if these are indeed similar structures to the purported gastroliths near the sternum.
Among the purported gastroliths indicated by Li et al. (2015), closer inspection of Bohaiornis IVPP V17963 reveals that one is very thin and amorphous, two are thicker and still amorphous, and two, including the sample extracted for analysis, contain within their margins two circular areas of higher relief connected by thinner areas, one forming an overall amorphous shape and the other together forming a subspherical shape (Figure 1).
Gastrolith Lithology Comparison
EDS of the extracted purported gastrolith ground section reveals that the circular structures are silica (SiO2) fibers in a spherulitc arrangement (Figure 3). The thinner areas connecting these spherules show a more diverse elemental composition, additionally contain Al, K, Na, and small amounts of Mg. Small regions near the center of the quartz spherules show similar elemental compositions (Figures 3, 5). In addition, regions within the spherules appear to contain organic remains as indicated by the presence of C, suggesting the quartz spherules are authigenic. However, at this time we cannot rule out the possibility that the C is contaminant originating from the resin (Figure 4).
Three gastrolith samples were extracted from undisputed gastral masses belonging to both birds (Iteravis IVPP V23346 and Archaeorhynchus IVPP V17075) and a non-avian dinosaur (Caudipteryx STM4-5) for comparative analysis. Elemental and lithological analysis indicate they are also siliceous rocks composed mainly of quartz. Quartz is a hard, relatively insoluble mineral that retains its angularity until completely ground to a powder (Meinertzhagen, 1954). The fact quartz predominates in the gizzard of extant birds reflects the requirement of gizzard stones that they be insoluble to survive the acid environment of digestive system and hard to resist gastrolith on gastrolith abrasion and thus serve to better grind ingested remains for longer periods of time (Gionfriddo and Best, 1999). These results indicate that the preference for siliceous rocks for ingestion as gastroliths is extended to Mesozoic birds.
Detailed chemical EDS analysis revealed compositional zonation in the purported gastroliths of Bohaiornis. Zonation is also somewhat present in the gastrolith of Iteravis IVPP V23346 in that along the exterior of one margin of the gastrolith there is a thin layer that shows greater elemental diversity. However, only the sample from Bohaiornis has internal zones as well. Observed under the microscope the internal region of Bohaiornis IVPP V17963 that contains higher elemental diversity as well as the similarly elementally diverse region also containing C along one margin of the sample both appear much darker than the remainder of the sample. The cause of this zonal structure in Bohaiornis IVPP V17963 is unknown. Notably, despite similar results in the EDS elemental mapping, the internal black region of the purported gastroliths in Bohaiornis IVPP V17963 appears completely different under the microscope compared to the area external to the quartz spherules that shows similar chemical composition (Figure 3).
Although chemical analyses indicate that the samples from Bohaiornis IVPP V17963 and the comparative samples all primarily consist of Si and O, the lithology varies between specimens and the distribution of elements and structure in the purported gastroliths are distinct from the other three samples.
Gastral Mass Proportions
Modern raptors often ingest stones to clean the stomach, which is referred to as rangle (Albuquerque, 1982). The smallest reported diameter of these stones is 7 mm (Fox, 1976). This is much larger than the purported gastroliths in Bohaiornis IVPP V17963 which measure 0.683 mm and 0.793 mm, even accounting for differences in body size between bohaiornithids and extant raptors. Rangle mass accounts for approximately 3.3% of the body mass in female New Zealand falcons and 3.0% in males (Fox, 1976). Gastral masses, which consist of gizzard stones (stones used for grinding food as opposed to rangle which is used to clean the stomach), are reportedly proportionately much smaller, forming only 1% predicted of body mass in neornithines (Wings, 2004). The body mass of Bohaiornis IVPP V17963 is estimated to be 308 g (Serrano et al., 2015). Compared to both rangle and gastral masses, the purported gastral mass in Bohaiornis is very small, much less than the 3.08–10.14 g expected if these indeed represented rangle or gizzard stones. Based on reports from the literature, the diameter of the individual purported gastroliths and their total mass is not comparable to rangle reported in New Zealand falcons. Due to the excellent preservation of Bohaiornis IVPP V17963, which is largely complete and articulated, it is unlikely that a large number of gastroliths were originally present and have been subsequently lost during taphonomy.
Mineral Precipitate Origins?
If the structures are not gastroliths ingested for use in the digestive system and indeed some form of mineral precipitate, this begs the question of what conditions produced these structures. Authigenic minerals can be associated with the fossilization of soft tissues (Tomescu et al., 2016). The radially fibrous morphology of the spherulites is consistent with chalcedony. The purported gastroliths record no microstructures identified thus far that would clearly indicate they are silicified soft tissue. However, chalcedony has been reported to replace fossils and to preserve inclusions of original organic material in carbonate rocks (Noble and Stempvoort, 1989) and authigenic quartz is common in organic rich shales (Zhao et al., 2017). The presence of C within the spherules may be indicative of original organic matter, which can reportedly serve as centers of nucleation for authigenic quartz (Noble and Stempvoort, 1989). However, at this time it cannot be ruled out that the C is resin that has somehow infiltrated into the interior of the spherules (Bailleul et al., 2020). If this is indeed the case, it would indicate that the spherules were not solid, which further suggests that these stones would be unsuitable for use in the digestive system being prone to breaking.
We consider it more likely that this C is original material of Bohaiornis IVPP V17963 that has served as a center of nucleation for the precipitation of authigenic quartz in the form of chalcedony. In the absence of histochemical or microstructural data there are numerous candidates for what this tissue may have been based on their anatomical position. It is possible it originated from the m. rectus adominis that stretched caudally from the sternum, stomach tissue, or epithelium.
Conclusion
Further analysis of the purported gastroliths in Bohaiornis IVPP V17963 confirms that the traces are unusual when compared to gastroliths found in gastral masses of other Jehol pennraptorans (Table 2). Morphological observation and CL data indicate that the purported gastroliths are interconnected, amorphous in shape with no consistent structure between themselves, and very thin, altogether inconsistent with the morphology of gizzard stones. In addition, the estimated mass of the purported gastroliths is far below that consistent with interpretation they are either gizzard stones or rangle.
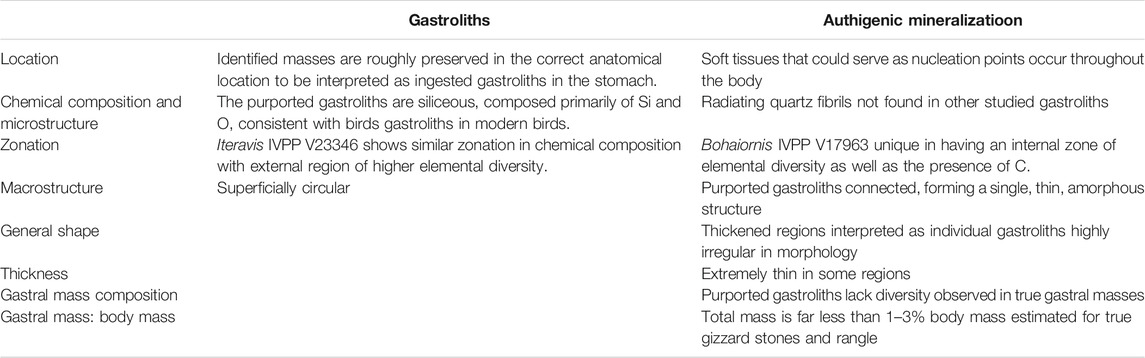
TABLE 2. Evidence in support of the two competing hypothesis.
Ground sections and EDS reveal that the superficially circular structures of the purported gastroliths are spherulites composed of Si and O. The internal regions of the spherulites and the thin regions connecting them have more diverse elemental compositions forming a zonal morphology in ground section. These structures resemble authigenic quartz. The gastrolith extracted from the oviraptorosaur Caudipteryx is fine-grained sandstone; that of Archaeorhynchus is volcanic; and that of Iteravis is silty mudstone–all stones with high silica content. Zonation similar to that observed in Bohaiornis IVPP V17963 is not present in any of the gastroliths with the minor exception of Iteravis, in which one margin of the gastrolith preserves a thin layer with similar higher elemental diversity.
Only the chemical composition of the purported gastroliths is consistent with their identification as ingested stones. Consideration of all the new evidence generated from these analyses taken together favors the hypothesis that the structures in question are not gastroliths. However, as of yet it is unclear what these structures represent or how they formed. Based on the available data we conclude that there is no strong evidence that Bohaiornis IVPP V17963 ingested gastroliths to use as rangle similar to extant raptorial birds. However, these unusual interconnected spherulitic quartz structures themselves represent a new structure that deserves further study. The conditions under which these structures formed is as yet unknown. Similar structures may have been overlooked in other specimens. Identification of similar structures in other Jehol fossils and further analysis of these spherulites may shed light on the taphonomic conditions that produced them in the future. However, the fact that such structures have not been widely reported suggests that this is not a common phenomenon. This research represents the first indepth analysis of gastroliths in Jehol birds using a variety of methods not previously applied to investigate early avian digestive function. This research highlights the importance of a multidisciplinary approach when explore the soft tissue preservation.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
SL: experiment design; data collection and analysis; manuscript writing. JO’C: designed research; writing and revising the paper. ZL: designed research; provided guidance. AB: data collection. MW: provided specimens.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Wu Qian, Wang Shiying for useful comments on paper; Cai Jiachen, Zhu Xufeng, Liu Biying for help with mineral identification under microscope; Hou Hongxiang, Zhang Shukang, Fang Gengyu, Wang Xiaomin for help and guidance on experiments; Gao Wei for photographs assistance; Liu Xinzheng for extracting samples.
References
Albuquerque, J. L. B. (1982). Observations on the use of rangle by the Peregrine falcon (Falco peregrinus tundrius) wintering in southern brasil. Raptor Res. 16 (3), 91–92.
Bailleul, A. M., O'Connor, J., Li, Z., Wu, Q., Zhao, T., Martinez Monleon, M. A., et al. (2020). Confirmation of ovarian follicles in an enantiornithine (Aves) from the Jehol Biota using soft tissue analyses. Commun. Biol. 3 (1), 399. doi:10.1038/s42003-020-01131-9 |
Butler, R. J., and Goswami, A. (2008). Body size evolution in Mesozoic birds: little evidence for Cope's rule. J. Evol. Biol. 21 (6), 1673–1682. doi:10.1111/j.1420-9101.2008.01594.x
Cau, A., and Arduini, P. (2008). Enantiophoenix electrophyla gen. et sp. nov. (Aves, Enantiornithes) from the Upper Cretaceous (Cenomanian) of Lebanon and its phylogenetic relationships. Atti Soc. it. Sci. nat. Museo civ. Stor. nat. Milano 149 (2), 293–324.
Cerda, I. A. (2008). Gastroliths in an ornithopod dinosaur. Acta Palaeontol. Pol. 53 (2), 351–355. doi:10.4202/app.2008.0213
Chiappe, L. M., Zhao, B., O'Connor, J. K., Chunling, G., Wang, X., Habib, M., et al. (2014). A new specimen of the Early Cretaceous bird Hongshanornis longicresta: insights into the aerodynamics and diet of a basal ornithuromorph. PeerJ 2, e234. doi:10.7717/peerj.234 |
Choiniere, J. N., Forster, C. A., and Klerk, W. J. d. (2012). New information on Nqwebasaurus thwazi, a coelurosaurian theropod from the early cretaceous kirkwood Formation in South Africa. J. Afr. Earth Sci. 71-72, 1–17. doi:10.1016/j.jafrearsci.2012.05.005
Dala Vecchia, F. M. D., and Chiappe, L. M. (2002). First avian skeleton from the Mesozoic of northern Gondwana. J. Vertebr. Paleontol. 22 (4), 856–860. doi:10.1671/0272-4634(2002)022[0856:Fasftm]2.0.Co;2
Duke, G. E. (1986). “Alimentary canal: anatomy, regulation of feeding, and motility,” in Avian physiology. Editor P.D. Sturkie (New York, NY: Springer-Verlag), 269–288.
Field, D. J., Lynner, C., Brown, C., and Darroch, S. A. (2013). Skeletal correlates for body mass estimation in modern and fossil flying birds. PLoS One. 8 (11), e82000. doi:10.1371/journal.pone.0082000 |
Gionfriddo, J. P., and Best, L. B. (1999). “Grit use by birds: a review,” in Current ornithology. Editors V. N. JR, E. D. Ketterson, and C. F. Thompson (New York, NY: Springer Science+Business), 89–148.
Huang, J., Wang, X., Hu, Y., Liu, J., Peteya, J. A., and Clarke, J. A. (2016). A new ornithurine from the Early Cretaceous of China sheds light on the evolution of early ecological and cranial diversity in birds, PeerJ 4, e1765. doi:10.7717/peerj.1765 |
Ji, Q., Currie, P. J., Norell, M. A., and Ji, S. A. (1998). Two feathered dinosaurs from northeastern China. Nature 393 (25), 753–761. doi:10.1038/31635
Ji, Q., Norell, M. A., Makovicky, P. J., Gao, K., Ji, S. A., and Yuan, C. (2003). An early ostrich dinosaur and implications for Ornithomimosaur phylogeny. Am. Mus. Novit. 3420 (1), 1–19. doi:10.1206/0003-0082(2003)420<0001:Aeodai>2.0.Co;2
Li, Z., Zhou, Z., Wang, M., and Clarke, J. A. (2015). A new specimen of large-bodied basal enantiornithine Bohaiornis from the Early Cretaceous of China and the inference of feeding ecology in mesozoic birds. J. Paleontol. 88 (1), 99–108. doi:10.1666/13-052
Liu, D., Zhou, Z., and Zhang, Y. (2012). Mass estimate and evolutionary trend in Chinese Mesozoic fossil birds. Vertebr. Palasiat. 50 (1), 39–52. doi:10.19615/j.cnki.1000-3118.2012.01.006
Makovicky, P. J., Kilbourne, B. M., Sadleir, R. W., and Norell, M. A. (2011). A new basal ornithopod (Dinosauria, Ornithischia) from the late cretaceous of Mongolia. J. Vertebr. Paleontol. 31 (3), 626–640. doi:10.1080/02724634.2011.557114
Noble, J. P. A., and Stempvoort, D. R. V. (1989). Early burial quartz authigenesis in Silurian platform carbonates, New Brunswick, Canada. J. Sediment. Petrol. 59 (1), 65–76. doi:10.1306/212F8F1C-2B24-11D7-8648000102C1865D
O'Connor, J. K. (2009). A systematic review of enantiornithes (aves: Ornithothoraces). PhD thesis. Los Angeles (CA): University of Southern California.
O'Connor, J. K. (2019). The trophic habits of early birds. Palaeogeogr. Palaeoclimatol. Palaeoecol. 513, 178–195. doi:10.1016/j.palaeo.2018.03.006
O'Connor, J. K., Wang, M., and Hu, H. (2016). A new ornithuromorph (Aves) with an elongate rostrum from the Jehol Biota, and the early evolution of rostralization in birds. J. Syst. Palaeontol. 14 (11), 939–948. doi:10.1080/14772019.2015.1129518
O'Connor, J. K., Wang, X., Sullivan, C., Wang, Y., Zheng, X., Hu, H., et al. (2018). First report of gastroliths in the Early Cretaceous basal bird Jeholornis. Cretac. Res. 84, 200–208. doi:10.1016/j.cretres.2017.10.031
O'Connor, J. K., Zheng, X., Wang, X., Wang, Y., and Zhou, Z. (2013). Ovarian follicles shed new light on dinosaur reproduction during the transition towards birds. Natl. Sci. Rev. 1 (1), 15–17. doi:10.1093/nsr/nwt012
O'Connor, J. K., and Zhou, Z. (2019). The evolution of the modern avian digestive system: insights from paravian fossils from the Yanliao and Jehol Biotas. Palaeontology 63 (1), 13–27. doi:10.1111/pala.12453
O’Connor, J. K., Chiappe, L. M., and Bell, A. (2011). “Pre-modern birds: avian divergences in the mesozoic,” in Living Dinosaurs: the evolution history of modern birds. 1st Edn, Editors G. Dyke, and G. Kaiser (Hoboken, NJ: John Wiley & Sons), 40–114.
Sanz, J. L., Chiappe, L. M., Pérez-Moreno, B. P., Buscalioni, A. D., Moratalla, J. J., Ortega, F., et al. (1996). An Early Cretaceous bird from Spain and its implictions for the evolution of avian flight. Nature 382 (1), 442–445. doi:10.1038/382442a0
Serrano, F. J., Palmqvist, P., Chiappe, L. M., and Sanz, J. L. (2016). Inferring flight parameters of Mesozoic avians through multivariate analyses of forelimb elements in their living relatives. Paleobiology 43 (1), 144–169. doi:10.1017/pab.2016.35
Serrano, F. J., Palmqvist, P., and Sanz, J. L. (2015). Multivariate analysis of neognath skeletal measurements: implications for body mass estimation in Mesozoic birds. Zool. J. Linn. Soc. 173 (4), 929–955. doi:10.1111/zoj.12215
Tomescu, A. M. F., Klymiuk, A. A., Matsunaga, K. K. S., Bippus, A. C., and Shelton, G. W. K. (2016). “Microbes and the fossil record: selected topics in paleomicrobiology,” in Their world: a diversity of microbial environments. Editor A. A. Klymiuk (New York, NY: Springer International Publishing), 69–169.
Varricchio, D. J., Sereno, P. C., Zhao, X., Lin, T., Wilson, J. A., and Lyon, G. H. (2008). Mud-trapped herd captures evidence of distinctive dinosaur sociality. Acta Palaeontol. Pol. 53 (4), 567–578. doi:10.4202/app.2008.0402
Wang, M., O'Connor, J. K., Pan, Y., and Zhou, Z. (2017). A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nat. Commun. 8, 14141. doi:10.1038/ncomms14141 |
Wang, M., O'Connor, J. K., Xu, X., and Zhou, Z. (2019). A new Jurassic scansoriopterygid and the loss of membranous wings in theropod dinosaurs. Nature 569 (7755), 256–259. doi:10.1038/s41586-019-1137-z |
Wang, M., Zhou, Z., and Sullivan, C. (2016a). A fish-eating enantiornithine bird from the early cretaceous of China provides evidence of modern avian digestive features. Curr. Biol. 26 (9), 1170–1176. doi:10.1016/j.cub.2016.02.055 |
Wang, M., Zhou, Z., and Zhou, S. (2016b). A new basal ornithuromorph bird (Aves: Ornithothoraces) from the Early Cretaceous of China with implication for morphology of early ornithuromorpha. Zool. J. Linn. Soc. 176 (1), 207–223. doi:10.1111/zoj.12302
Wang, M., and Zhou, Z.-H. (2016). A new adult specimen of the basalmost ornithuromorph bird Archaeorhynchus spathula (Aves: ornithuromorpha) and its implications for early avian ontogeny. J. Syst. Palaeontol. 15, 1–18. doi:10.1080/14772019.2015.1136968
Wang, M., Zhou, Z., O’Connor, J. K., and Zelenkov, N. V. (2014). A new diverse enantiornithine family (Bohaiornithidae fam. nov.) from the Lower Cretaceous of China with information from two new species. Vertebr. Palasiat. 52 (1), 31–76. doi:10.19615/j.cnki.1000-3118.2014.01.004
Wang, M., and Zhou, Z. (2017a). “The evolution of birds with implications from new fossil evidences,” in The biology of the avian respiratory system. Editor J. N. Maina (Newyork, NY: Springer International Publishing), 1–26.
Wang, M., and Zhou, Z. (2017b). A morphological study of the first known piscivorous enantiornithine bird from the Early Cretaceous of China. J. Vertebr. Paleontol. 37 (2), e1278702. doi:10.1080/02724634.2017.1278702
Wang, X., O'Connor, J. K., Maina, J. N., Pan, Y., Wang, M., Wang, Y., et al. (2018). Archaeorhynchus preserving significant soft tissue including probable fossilized lungs. Proc. Natl. Acad. Sci. Unit. States Am. 115 (45), 11555–11560. doi:10.1073/pnas.1805803115
Wang, X., O'Connor, J. K., Zhao, B., Chiappe, L. M., Gao, C., and Cheng, X. (2010). New species of enantiornithes (aves: Ornithothoraces) from the qiaotou Formation in northern hebei, China. Acta Geol. Sin. 84 (2), 247–256. doi:10.1111/j.1755-6724.2010.00156.x
Wang, Y., O'Connor, J. K., Li, D., and You, H. (2015). New information on postcranial skeleton of the early cretaceous Gansus yumenensis (aves: ornithuromorpha). Hist. Biol. 28 (5), 666–679. doi:10.1080/08912963.2015.1006217
Wings, O. (2007). A review of gastrolith function with implications for fossil vertebrates and a revised classification. Acta Palaeontol. Pol. 52 (1), 1–16. doi:10.1038/sj.onc.1207250
Wings, O. (2004). Identification, distribution, and function of gastroliths in dinosaurs and extant birds with emphasis on ostriches (Struthio camelus). PhD thesis. Bonn (Germany): Rheinische Friedrich-Wilhelms-Universität.
Xu, X., Clark, J. M., Mo, J., Choiniere, J., Forster, C. A., Erickson, G. M., et al. (2009). A Jurassic ceratosaur from China helps clarify avian digital homologies. Nature 459 (7249), 940–944. doi:10.1038/nature08124 |
Xu, X., Zhou, Z., Wang, Y., and Wang, M. (2020). Study on the Jehol Biota: recent advances and future prospects. Sci. China Earth Sci. 63, 1–17. doi:10.1007/s11430-019-9509-3
Zhang, , and Miman, (2003). The Jehol biota. Shanghai, China: Shanghai Scientific and Technical Publishers.
Zhang, Y., O'Connor, J., Di, L., Qingjin, M., Sigurdsen, T., and Chiappe, L. M. (2014). New information on the anatomy of the Chinese early cretaceous Bohaiornithidae (aves: enantiornithes) from a subadult specimen of Zhouornis hani. PeerJ 2, e407. doi:10.7717/peerj.407 |
Zhao, J., Jin, Z., Jin, Z., Wen, X., and Geng, Y. (2017). Origin of authigenic quartz in organic-rich shales of the Wufeng and longmaxi formations in the sichuan basin, south China: implications for pore evolution. J. Nat. Gas Sci. Eng. 38, 21–38. doi:10.1016/j.jngse.2016.11.037
Zheng, X., Martin, L. D., Zhou, Z., Burnham, D. A., Zhang, F., and Miao, D. (2011). Fossil evidence of avian crops from the Early Cretaceous of China. Proc. Natl. Acad. Sci. U.S.A. 108 (38), 15904–15907. doi:10.1073/pnas.1112694108 |
Zheng, X., O'Connor, J. K., Wang, X., Wang, Y., and Zhou, Z. (2018). Reinterpretation of a previously described Jehol bird clarifies early trophic evolution in the ornithuromorpha. Proc. Biol. Sci. 285, 20172494. doi:10.1098/rspb.2017.2494 |
Zhou, S., O’Connor, J. K., and Wang, M. (2014). A new species from an ornithuromorph (Aves: Ornithothoraces) dominated locality of the Jehol Biota. Chin. Sci. Bull. 59 (36), 5366–5378. doi:10.1007/s11434-014-0669-8
Zhou, S., Zhou, Z., and O’Connor, J. (2013). Anatomy of the basal ornithuromorph bird Archaeorhynchus spathulafrom the early cretaceous of liaoning, China. J. Vertebr. Paleontol. 33, 141–152. doi:10.2307/23361078
Zhou, Z., and Zhang, F. (2002). A long‐tailed, seed‐eating bird from the Early Cretaceous of China. Nature 418, 405. doi:10.1038/nature00930 |
Zhou, Z. (2006). Evolutionary radiation of the Jehol Biota: chronological and ecological perspectives. Geol. J. 41 (3-4), 377–393. doi:10.1002/gj.1045
Zhou, Z., and Wang, X. (2000). A new species of Caudipteryx from the yixian formation of liaoning northeast China. Vertebr. Palasiat. 38 (2), 111–127. doi:10.19615/j.cnki.1000-3118.2000.02.005
Zhou, Z., Wang, X., Zhang, F., and Xu, X. (2000). Important features of Caudipteryx-evidence from two nearly complete new specimens. Vertebr. Palasiat. 38 (4), 241–254. doi:10.19615/j.cnki.1000-3118.2000.04.001
Zhou, Z., and Zhang, F. (2006). A beaked basal ornithurine bird (Aves, Ornithurae) from the Lower Cretaceous of China. Zool. Scripta. 35, 363–373. doi:10.1111/j.1463-6409.2006.00234.x
Keywords: gastrolith, Enantiornithes, SEM, rangle, authigeneic mineral, siliceous mineral, EDS
Citation: Liu S, Li Z, Bailleul AM, Wang M and O’Connor J (2021) Investigating Possible Gastroliths in a Referred Specimen of Bohaiornis guoi (Aves: Enantiornithes). Front. Earth Sci. 9:635727. doi: 10.3389/feart.2021.635727
Received: 30 November 2020; Accepted: 25 January 2021;
Published: 19 February 2021.
Edited by:
Martin Daniel Ezcurra, Museo Argentino de Ciencias Naturales Bernardino Rivadavia, ArgentinaReviewed by:
Ignacio Cerda, Consejo Nacional de Investigaciones Científicas y Técnicas, ArgentinaFederico Agnolin, Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Argentina
Copyright © 2021 Liu, Li, Bailleul, Wang and O’Connor. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shumin Liu, bGl1c2h1bWluQGl2cHAuYWMuY24=; Jingmai O’Connor, amluZ21haS5vY29ubm9yQGdtYWlsLmNvbQ==