 Alexey M. Klementiev1,2*
Alexey M. Klementiev1,2* Arina M. Khatsenovich1*Yadmaa Tserendagva3
Arina M. Khatsenovich1*Yadmaa Tserendagva3 Evgeny P. Rybin1
Evgeny P. Rybin1 Dashzeveg Bazargur3Daria V. Marchenko1Byambaa Gunchinsuren3Anatoly P. Derevianko1†John W. Olsen1,4†
Dashzeveg Bazargur3Daria V. Marchenko1Byambaa Gunchinsuren3Anatoly P. Derevianko1†John W. Olsen1,4†- 1Institute of Archaeology and Ethnography SB RAS, Novosibirsk, Russia
- 2Institute of the Earth’s Crust SB RAS, Irkutsk, Russia
- 3Institute of Archaeology MAS, Ulaanbaatar, Mongolia
- 4School of Anthropology, University of Arizona, Tucson, AZ, United States
Throughout the arid lands of Africa and Eurasia, camelids facilitated the expansion of human populations into areas that would not likely have been habitable without the transportation abilities of this animal along with the organic resources it provides, including dung, meat, milk, leather, wool, and bones. The two-humped, Bactrian, species of Camelus, C. ferus in its wild state and C. bactrianus when domesticated, is much more poorly known in prehistoric archaeological contexts than its single-humped congeneric, C. dromedarius. Our research uses a convergence of evidence approach to analyze reports and remains of Plio-Pleistocene camelids in Central and Northern Asia and trace the latest-known fossil Bactrian relative, Camelus knoblochi, that seems to have survived in the Gobi Desert until the Last Glacial Maximum (ca. 26.5–19 ka). Rock art depictions, some of which may be of Pleistocene age, record the complexity of nascent human-camel interactions and provide the impetus for further archaeological studies of both the origins of C. bactrianus and its increasingly complex relationships with the highly mobile prehistoric peoples of Central and Northern Asia.
1 Introduction
Camelus knoblochi was the largest of the Pleistocene Eurasian Bactrian (two-humped) members of the genus Camelus. The broad zoogeographic range of their remains has made this extinct tylopod the fossile directeur of Middle Pleistocene Khazar and Singil mammalian complexes in the northern Caucasus, Volga and Caspian basins, Tajikistan, Kazakhstan, and southern Siberia, including Transbaikalia (the mountainous region east of Lake Baikal) (Titov 2008). C. knoblochi remains are also known from loess deposits in the Ordos Region in northern China (Boule and de Chardin 1928; Qi 1975; Yamanaka 1993; Dong et al., 2014). Although the Ordos Region and the Altai Mountains in Kazakhstan and Russia have yielded multiple fossils of Camelus knoblochi (Kozhamkulova 1963; Vasiliev 2016), this camelid has not been previously reported from Mongolia. Joint Mongolian-Soviet and Mongolian-Russian-American expeditions ongoing since 1985 have recovered Middle and Late Pleistocene camelid fossils in Mongolia, but most such remains have not yet been published.
A brief history of Camelus finds reflects the growing data on Quaternary fossil faunas in Mongolia. The earliest and most complete description of a Mongolian camelid fossil is the Camelus sivalensis astragalus from the Öoshiin-Bor-Uzuur-Uul Member in the Öshiin Formation of the Dzeregen Valley in southwestern Mongolia (Belyeva 1937). C. sivalensis was first described by Falconer and Cautley (1836) based on remains from the Sivalik Hills in northern India. They concluded that this camel was similar to both modern Bactrian and dromedary species (Colbert 1935; Bibi and Métais 2016). The astragalus from Öoshiin is the only C. sivalensis fossil yet reported outside of India.
In the 1960s, a camelid metatarsal was found at the base of colluvial deposits at Bulan-Khujir near Ulaanbaatar (Okladnikov and Troitsky 1967), but further details are lacking. Similarly, E.V. Devyatkin published the discovery of Pliocene camel remains from Mongolia without accompanying detailed descriptions (Devyatkin 1981). These specimens were found at three localities: Paracamelus sp. from Khyargas Nuur 2, P. gigas from Yavar 1, situated in the Great Lakes Depression (Mong. Ikh Nuuruudyn Khotgor), in northwestern Mongolia, and Camelus? sp. from Orog Nuur (Orok Nor) in the Valley of the Lakes (Mong. Nuuruudyn Khöndii) in south-central Mongolia.
Khyargas Nuur 2 is a paleontological locality on the north shore of the eponymous lake in the Uvs Province, western Mongolia. The stratigraphic section includes three units; fossils of Paracamelus sp. were found in the upper part of Unit A (Layers 9–11, 37–61 m), together with several species of Hipparion, Sinomegaceros sp., Antilospira sp., Olonbulukia sp. (a stem-caprine species), Canis sp., and members of the Chalicotheriidae, among others (Devyatkin 1981). The Yavar 1 locality is situated in the Dzabkhan River Valley, near Khyargas Nuur. A variety of fossil remains were found there, including those of Paracamelus gigas, various species of Hipparion, and members of the Felidae, Bovidae, Cervidae, Giraffidae, and Mustelidae among others. Finally, a locality yielding Camelus sp. remains was recorded near Orog Nuur, in the Valley of the Lakes, in association with remains of Hipparion spp., Gazella sp., Castor sp., and Struthiolithus sp. along with members of the Rhinocerotidae and Mastodontidae (Berkey and Morris 1927; Devyatkin 1981). All these fossils are associated with Pliocene deposits. Some researchers consider these faunal assemblages to be Middle Pliocene, pre-Villafranchian (Vangengeim 1977; Devyatkin 1970, Devyatkin 1981) and are distributed throughout Mongolia, northeastern China, and northeastern Kazakhstan (Devyatkin et al., 1968; Vangengeim et al., 1972; Devyatkin 1981; but see; Ravsky et al., 1964).
From the Middle Pliocene through the late Holocene when domesticated Bactrian camels spread throughout Central Asia, especially during the Bronze Age, there is a gap in camelid evolutionary history in Mongolia. Obviously, the Camelidae occupied this vast region during the Pleistocene, but their remains have not previously been reported or published. In 1985, V.T. Petrin found fossil remains of large mammals at Tugrug Shireet in Ömnögovi Province, southern Mongolia. Our analysis of these remains revealed both fossil horses and camels. Remains of fossil camels were also mentioned in a description of deposits in the Uvs Nuur Basin in northwest Mongolia. The Chusutuin Gol section is situated in the Khustyn Gol Valley and includes deposits with mammalian remains going back to the beginning of Marine Isotope Stage 3 (48.3–45.3 ka BP) (Grunert et al., 2000), corresponding to a rise in the level of Uvs Nuur. These two camelid specimens have only been recently reported and never taxonomically identified or described in detail. A single find of Camelus ferus has been reported from Otson Tsokhio in the Gobi Desert, also without complete description (Janz et al., 2021).
2 Material and Methods
Here, we consider camelid remains recovered from two archaeological sites in Mongolia: Tsagaan Agui Cave and the Tugrug Shireet open-air site. Tsagaan Agui Cave is in Bayankhongor Province (44°42′53.3″ N, 101°10′13.4″ E; 2000 m asl) south-central Mongolia, in the Tsagaan Tsakhir limestone massif–part of the eastern Gobi Altai Mountains (Figure 1). The modern climate of the Gobi Altai Mountains is arid, and the Tsagaan Tsakhir inselberg is in a desert to semi-desert zone. There is no readily accessible surface fresh water; local pastoral nomads exploit a network of wells. Wet years bring abnormally high amounts of rain that result in short-term flooding, turning the Gobi Desert green. Flora and fauna are dominated by desert and semi-desert adapted species. Although vegetation cover is poor (ca. 15–20 cm high for therophytes and a bit higher for perennials), various reptiles and rodents thrive there. Lagomorphs are represented by the pikas (Ochotona spp.) and tolai hare (Lepus tolai). Herbivores and predators also inhabit the Gobi Altai region, including argali (Ovis ammon) and Siberian ibex (Capra sibirica), Mongolian gazelle or dzeren (Procapra gutturosa), red fox (Vulpes), corsac or steppe fox (Vulpes corsac), dhole (Cuon alpinus), and grey wolf (Canis lupus). Modern herding activity focuses on sheep, goats, camels and, to a lesser extent, cattle and horses. At higher altitudes near Ikh Bogd Uul Mountain, yaks are herded.
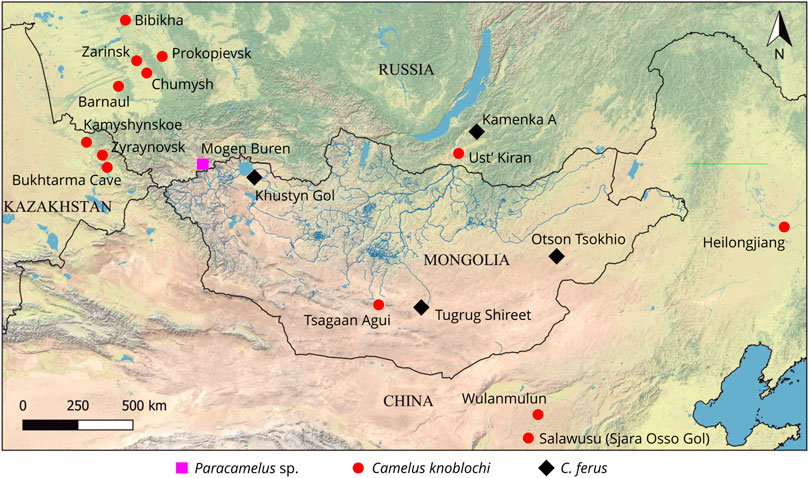
FIGURE 1. Principal finds of fossil camels in eastern Eurasia.
The ecology of the of Gobi Altai is characterized not only by sparse vegetation cover and a lack of large herds of ungulates, but also by significant annual and daily temperature differentials, extremely arid conditions, and very strong winds–all at altitudes above 2000 m a.s.l. in the Tsagaan Agui area. Hypoxia generally does not negatively impact human health in dry continental climates below 3,000–3,500 m a.s.l., in comparison with wetter regions like the mountainous interior of the Kamchatka Peninsula in Russia’s Far East. However, modern mountain medicine regards 2000 m a.s.l. as falling within the high-altitude rubric (defined as beginning at 1,500 m a.s.l.) (Young and Reeves 2002). Climatic conditions are harsh in the Gobi and even xerocoles can be negatively impacted by dehydration and excessive solar radiation as well as a dearth of potable water and strong winds that transport sand, injuring the eyes. Paleolithic humans also needed to adapt to harsh environmental conditions, but one of our goals here is to explore whether the Pleistocene Gobi Altai region was characterized by milder climatic and environmental conditions that were not significantly different from Mongolia’s forest and steppe zones.
Tsagaan Agui Cave has been known as an archaeological site since 1972 when the Mongolian archaeologist, D. Dorj first collected artifacts there. A subsequent intensive excavation campaign in 1987–1989 led by A.P. Derevianko and Dorj investigated the cave’s Entrance and Entrance Grotto (Vkhodnoy grot), where abundant faunal remains and archaeological materials were found (Derevianko and Petrin 1985; Martynovich 2002). From 1995 to 2000, the Joint Mongolian-Russian-American Archaeological Expedition carried out excavations in the Lower and Inner Grottos and Main Chamber of the cave (Derevianko et al., 2000a, Derevianko et al., 2000b), yielding data on paleoclimate, accumulation of cave deposits, chronology and lithic technology. The cave’s stratigraphy comprises 14 layers; archaeological material has been found in Layers 1–13. The chronology of the cultural horizons was determined by radiocarbon and then-experimental ESR dating approaches. Layer 2 was dated to the Last Glacial Maximum–(QT?) 23,000–25,000 BP; for Layer 3 there is a series of radiocarbon dates (AA-23159; 26588; 26587; 23158; 26589) falling between 31–34,000 BP; Layer 4 is dated to (QT40 & 41) 44–59,000 BP (Derevianko et al., 2000a, Derevianko et al., 2000b, Derevianko et al., 2004).
We recommenced excavations at Tsagaan Agui Cave in 2021 to obtain samples for additional radiocarbon and OSL dates and other analyses. A one square meter excavation pit, designated 2021/2, was dug in the northern part of the Main Chamber (Khatsenovich et al., 2021), in which Layers 1-6, according to the stratigraphic sequence revealed in the 2000 sondage, were uncovered. The upper part of a large camelid metacarpal was found in Layer 4 (Figure 2). The floor of the cave also contains Pleistocene material, both bones and artifacts, exposed by the erosion of underlying layers and sometimes transported by human agency (primarily tourists and Buddhist pilgrims) from the Inner Grotto and cave meanders. A large mammalian astragalus was found in such context. Three more camelid bones were subsequently rediscovered in the curated 1988–1989 faunal collections, including a carpal IV + V from Layer 2B near a combustion feature, a tibia fragment from Layer 1 (apparently redeposited), and a calcaneus fragment from an undetermined layer.

FIGURE 2. Proximal metacarpal (MC III + IV) fragment of Camelus knoblochi from Tsagaan Agui Cave, Mongolia: 1 - proximal view, 2 - dorsal view, 3 - plantar view.
A camelid second phalanx was found at Tugrug Shireet (N 44°13′49.6″, 103°16′24.2″ E; 2000 m asl) in the northern Gobi Desert (Figure 1). This bone, together with the other mammalian remains, are likely not associated with the mostly Neolithic archaeological remains at the site. Taxonomic identification of the Tugrug Shireet faunal assemblage and preservation of the bone indicate a Pleistocene age for this phalanx.
We analyzed five camelid bones from Tsagaan Agui Cave and one from Tugrug Shireet. Comparable samples of modern Camelus bactrianus were collected in the vicinity of Tsagaan Agui Cave. We reviewed published data on fossil camels from China, Kazakhstan and Siberia and employed logarithmic curves, applied earlier in the study of horses (Eisenmann et al., 1986) and bison (Vasiliev 2008), to better visualize these data. Phalanges of a modern domestic llama (Lama glama) sourced from the Irkutsk Zoo served as samples to create the baseline curves.
3 Systematic Paleontology
Suborder Selenodontia
Infraorder Tylopoda
Family Camelidae Gray, 1821
Subfamily Camelinae Gray, 1821
Tribe Camelini Gray, 1821
Genus Camelus Linnaeus, 1758
Camelus knoblochi Nehring, 1901
Material: Os carpal IV + V (Tsagaan Agui, 1989; Unit А16, Layer 2B), proximal left metacarpal (Tsagaan Agui, Pit 2021/2, Layer 4), distal epiphysis of right tibia (Tsagaan Agui, 1988; Unit B9, Layer 1), distal portion of right calcaneus (Tsagaan Agui, 1988; unit A5), right astragalus (Tsagaan Agui, 2021; cave floor).
3.1 Description and Comparison
The os carpal IV + V (Tsagaan Agui, 1989; Unit А16, Layer 2B) is very large with a transverse diameter of 41.5 mm, a width of 49.5 mm, and an anterior height at the dorsal portion of 28.6 mm (Figure 3: 1–2). Equivalent bones of modern domesticated camels in the Gobi Altai (n = 5) are significantly smaller with a transverse diameter ranging between 37.4–38.7 mm, widths falling between 35.6–44.7 mm, and anterior heights ranging from 22.1–24.8 mm. The facet for the scaphoid bone on the fossil specimen porrects (extends forward) more than in modern specimens and the posterior scrobiculus is much deeper. Based upon these parameters, these bones are assigned to C. knoblochi (Boule and de Chardin 1928) rather than P. gigas. C. knoblochi is distinguished from the latter by the shape of the upper articular surfaces for MCIII and MCIV (Zdansky 1926). These differences, as well as the assumed Pliocene age of known P. gigas, excluded the latter from further comparative analysis.
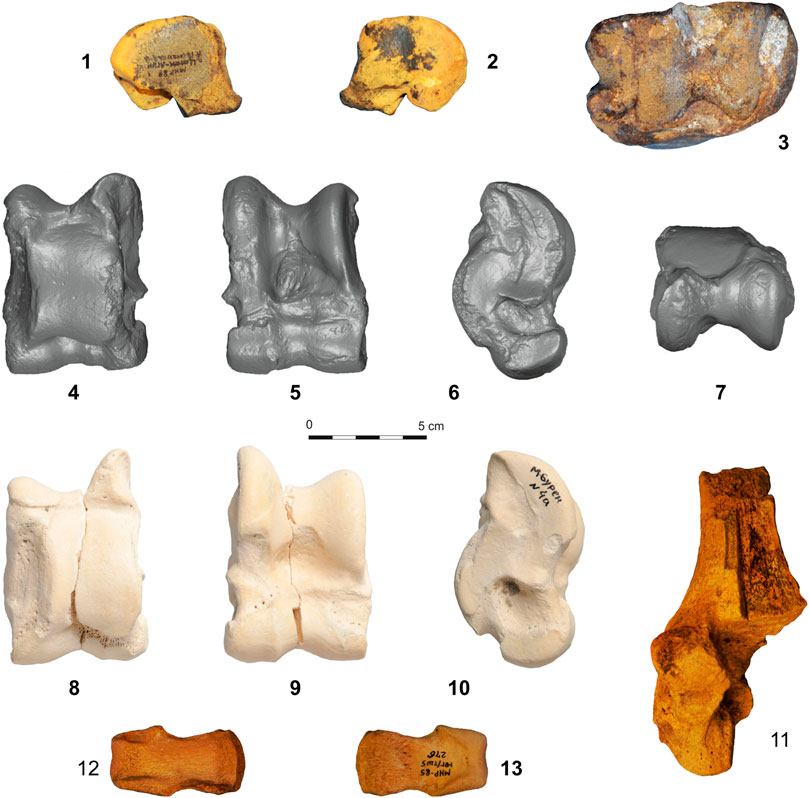
FIGURE 3. Fossil camel bones from Central Asia: 1, 2–Camelus knoblochi carpal IV + V (distal and proximal view); 3–Camelus knoblochi distal tibia fragment (distal view); 4–7–C. knoblochi astragalus (dorsal, plantar, lateral and proximal views); 8–9–Paracamelus sp. astragalus (dorsal, plantar and lateral views); 11–fragmentary C. knoblochi calcaneus (view from the dorsal side); 12–13–C. ferus second phalanx of the hind leg (Sites: 1–7, 11–Tsagaan Agui Cave; 8–10–Mogen Buren; 12–13–Tugrug Shireet).
A proximo-dorsal metacarpal fragment (Tsagaan Agui, Pit 2021/2, Layer 4) derives from a large individual (Table 1; Figure 2). The width and transverse diameter of the proximal epiphysis is much larger than the metacarpal of modern Bactrian camels in the Gobi Altai region (Table 1). Based on these morphometric parameters, this bone correlates well with both C. knoblochi (Khaveson 1954; Vangengeim and Gerbova 1962) and P. gigas, but the latter differs in the shape of its upper articular surface for metacarpals III and IV (Zdansky 1926). This fact and its assumed Pliocene age exclude P. gigas from conclusive correlation.
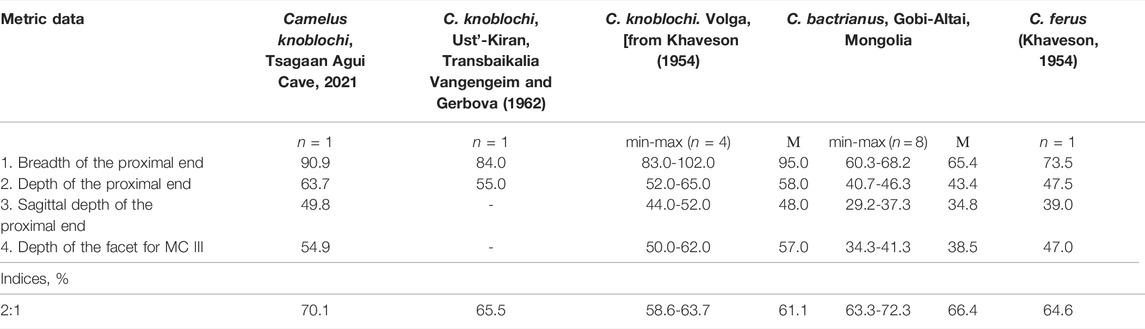
TABLE 1. Measurements of camel metacarpals III+IV.
The distal epiphysis of a camelid right tibia from Tsagaan Agui Cave (1988, Unit B9, Layer 1) exhibits a parallelepiped articular surface (Figure 3: 3); facets for the astragalus are located oblique to the primary axis of the bone. The facet for os malleolare is large and its anterior face is larger than the posterior. The width and transverse diameter of the distal epiphysis are 90.0 and 56.2 mm respectively; much larger than in the domesticated Camelus bactrianus (73.3–86.4 and 44.6–50.8 mm, respectively). The equivalent bone from Tsagaan Agui is slightly smaller than the C. knoblochi tibias from Sjara Osso Gol or Salawusu (Boule and Teilhard 1928), Zyryanovsk, Kazakhstan (Kozhamkulova 1963) and the Volga River Valley (specimen PIN 2348; Khaveson 1954).
The Tsagaan Agui astragalus (2021, cave floor) is massive and wide, with a width:length ratio of 73.6% (Table 2; Figure 3: 4–7). The bone’s dorsal face is asymmetric, and the external crista is much higher than the internal. The cuboidal facet is large with a deep, rough pit. This astragalus is much larger than similar bones of modern camels, falling within the range of variability of C. knoblochi.

TABLE 2. Morphometrics of Central Asian camel astragali.
Metric data on a calcaneus from Tsagaan Agui (1988, Unit A5) are incomplete, because the bone is fragmented (Figure 3: 11). The width of the bone body is 30.6 mm; much larger than that of the modern Mongolian domesticated camel (19.0–24.4 mm), but less than that of the specimen of C. knoblochi from Zyryanovsk, Kazakhstan (Kozhamkulova 1963). The calcaneal facet for os malleolare is 23.5° oblique to the bone’s major axis and the length and width of the facet are 40.2 and 26.8 mm, respectively. The width of the facet of the C. knoblochi calcaneus from Sjara Osso Gol is 26 mm (Boule and de Chardin 1928). The lateral bone surface exhibits a wide pit.
Camelus ferus Przewalski, 1878
Material: second phalanx of the hind leg (Tugrug Shireet 5, 1985, specimen 276) (Figure 3: 12–13).
3.2 Description and Comparison
The rugose surface for attaching major ligaments on the caudal surface of the bone’s distal margin is significant and occupies roughly half of the diaphyseal length. Logarithmic curves, based on Brigg’s logarithm for the ratio of measurements for the camelid second phalanx to phalanges of modern Lama glama yield similar curves for domesticated Bactrian and fossil camels from Tugrug Shireet (Figure 4). We included data on the first phalanx of C. knoblochi (Khaveson 1954; Vasiliev et al., 2015), and fossil Camelus remains from the Kamenka site in Transbaikalia (for which the authors determined genus, though not the species, of this camelid (Germonpre and Lbova 1996)), because equivalent data for the second phalanx of C. knoblochi are lacking. The diagram indicates the difference in phalanx length between C. knoblochi and Bactrian camels (Figure 5). The significant width of the fossil camelid first phalanx from the Kamenka site in Siberia allows it to be classified as C. ferus, which suggests that wild two-humped camels were latitudinally distributed from the Gobi Desert, north to western Transbaikalia in southern Siberia during the Late Pleistocene.
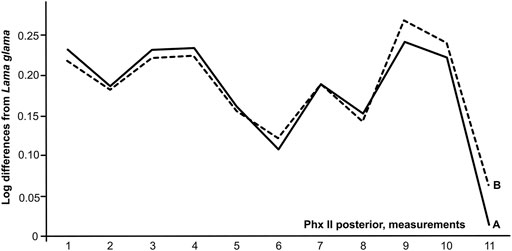
FIGURE 4. Ratio diagrams of posterior second phalanges of Camelus from Mongolia: 1–Tugrug Shireet; 2–C. bactrianus (n = 6). Measurements: 1 - anterior length; 2 - posterior length; 3 - middle articular length; 4 - minimal articular length; 5 - breadth of the upper epiphysis; 6 - depth of the upper epiphysis; 7 - breadth of the diaphysis; 8 - depth of the diaphysis; 9 - greatest lower breadth; 10 - minimum breadth of the lower epiphysis; 11 - depth of the lower epiphysis.
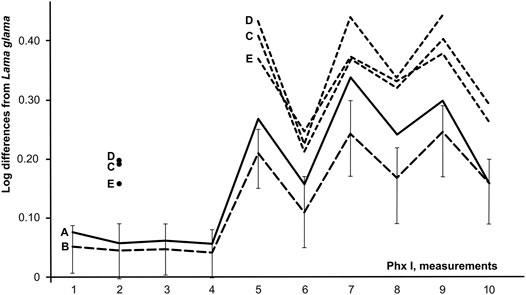
FIGURE 5. Ratio diagrams of first phalanges of Camelus spp.: 1–Kamenka A; 2–C. bactrianus (n = 18; vertical lines indicate range of variability); 3, 4–C. knoblochi (Khaveson, 1954); 5–C. knoblochi (Vasiliev, 2011). Measurements: 1 - anterior length, 2 - posterior length, 3 - middle articular length, 4 - minimal articular length, 5 - breadth of the upper epiphysis, 6 - depth of the upper epiphysis, 7 - breadth of the diaphysis, 8 - depth of the diaphysis, 9 - greatest lower breadth, 10 - minimum breadth of the lower epiphysis, 11 - depth of the lower epiphysis.
4 Camelid Evolution and Zoogeography in Eastern Central Asia
Situated near the geographical center of Asia, Mongolia has played a pivotal role in the evolution and dispersal of many continental mammals, and various ungulate and carnivore groups appear to have originated there (Lopatin 2019). Although fossil camelid remains are scarce in Mongolia, we can conclude that they have existed there since the Pliocene. The oldest known camelid remains in Mongolia were found in the Pliocene Öoshiin Formation (Belyeva 1937) and further investigations carried out by the Mongolian-Soviet Complex Paleontological Expedition correlated this formation with the Altan-Teli Pliocene stratigraphic assise (Devyatkin 1981). The initial identification of these remains as C. sivalensis needs to be revised. It is likely that giant camelids, such as those described in China (Zdansky 1926) and Kazakhstan (Aubekerova 1974) also inhabited Mongolia during the Pliocene.
A subsequent camelid evolutionary link has been found in the Tyva Republic on the Russian-Mongolian border: a Paracamelus sp. astragalus was recovered from Early Pleistocene deposits at Mogen Buren (Klementiev et al., 2021). The morphological features of this bone do not allow assigning it to the genus Camelus (Figure 3: 8–10), but its absolute size is characteristic of a smaller individual C. knoblochi (Table 2). This camelid specimen from Mogen Buren may represent a transitional stage of Pliocene Paracamelus spp. evolving into Pleistocene Camelus knoblochi. It is likely that camelid remains from the Kuruksai and Lakhuti deposits in the Afghan-Tajik Depression also represent intermediary forms, but there is a notable discrepancy in species identification. Sharapov defined a new species, P. trofimovi, based on all remains available at the time (Sharapov 1986). Alekseeva, considering the different ages of the Kuruksai and Kairubak stratigraphic assises, assigned camelid remains from Lakhuti 2 to C. knoblochi and from Lagernaya and Kuruksai to P. praebacrianus (Vangengeim et al., 1988).
The timing of the peak distribution of C. knoblochi and its prochoresis cannot be reconstructed directly from fossil remains in Mongolia, but the history of this species is known in eastern Europe and Kazakhstan as well (Titov 2008). The demonstrated presence of C. knoblochi in the Late Pleistocene of the Gobi Altai and Ordos regions and in China’s Heilongjiang Province (“Manchuria”) raises an important question regarding the age of sandy deposits in Transbaikalia and Mongolia. The chronology of the Ust’ Kiran faunal complex in western Transbaikalia, including a bone of C. knoblochi, is based on long-held assumptions about the Early Pleistocene age of the sandy deposits there, based on faunal remains (Vangengeim and Gerbova 1962). According to chronometric dates for this formation, that have been skeptically received, sand deposits in the Selenga River Basin accumulated up to the end of the Pleistocene (Perevalov and Rezanov 1997). Finds of C. knoblochi in Late Pleistocene deposits in Tsagaan Agui Cave support these dates.
Additional finds are associated with the persistence of C. knoblochi in arid landscapes of the Gobi Altai region at the end of the Middle Pleistocene and in the Late Pleistocene. Camel remains were first identified at Tsagaan Agui Cave by N.D. Ovodov, but never published, although other faunal remains recovered from the cave were reported (Ovodov 2001). The nearest localities to Tsagaan Agui yielding Late Pleistocene C. knoblochi remains are in the Ordos Region (Yamanaka 1993) and Heilongjiang Province in North China (Yuan et al., in press). The wild Bactrian camel, Camelus ferus, also appeared in Central Asia during the Late Pleistocene at Tugrug Shireet in Mongolia and the Kamenka site in Transbaikalia. Limited data do not illuminate its phyletic links with ancestral species and its possible co-existence with C. knoblochi. Recent genetic data suggesting a ca. 300 ka origin for C. knoblochi (Yuan et al., in press) are at odds with its presence at Lakhuti 2, which is dated to the Early Pleistocene (i.e., >773 ka).
In the Middle Pleistocene, C. knoblochi co-existed with Canis lupus, Mammuthus chosaricus, Stephanorhinus kirchbergensis, Elasmotherium sibiricum, Bison priscus longicornis, Saiga tatarica, Megaloceros giganteus and Cervus elaphus, a faunal association reflecting predominantly steppic conditions, and identifying C. knoblochi as an inhabitant of a steppe zone in the Volga River Basin (Titov 2008). Faunal remains from Late Pleistocene layers in Tsagaan Agui Cave include cave hyaena (Crocuta spelaea), khulan (Equus hemionus), argali (Ovis ammon), Siberian ibex (Capra sibirica), Mongolian gazelle or dzeren (Procapra gutturosa), Tibetan antelope or chiru (Pantholops hodgsonii), and tolai hare (Lepus tolai). These species, which did not become extinct at the end of the Pleistocene, are typical of modern steppe zones in Mongolia and high-altitude regions of Tibet. This faunal association indicates a dry steppe environment in the Gobi Altai during its occupation by both Camelus knoblochi and Paleolithic humans in the Late Pleistocene. The large body-size of this camel is evidence of abundant forage resources that are not characteristic of desert biomes.
Two finds of this fossil species in southwestern Siberia indicate its occasional association with forest-steppe landscapes during the Late Pleistocene (Vasiliev et al., 2015; Vasiliev 2018), perhaps indicating that, just prior to its extinction, C. knoblochi spread into unfamiliar and less salubrious ecological niches. Apparently, this species was poorly adapted to desert biomes, primarily because such landscapes could not support such large animals, but perhaps there were other reasons as well, related to the availability of fresh water and the ability of camels to store water within the body, poorly adapted mechanisms of thermoregulation, and competition from other members of the faunal community occupying the same trophic niche. Both wild and domesticated modern Bactrian and dromedary camels are xerocole species highly adapted to desert conditions based on genomic studies of insulin resistance, salt tolerance (Wang et al., 2012; Burger and Palmieri, 2013), fat metabolism, stress responses to arid environments, thermo- and osmoregulation, changes in respiratory systems and other biological factors (Wu et al., 2014). But it is unknown whether C. knoblochi expressed any of those adaptive mechanisms. Tsagaan Agui Cave contains some of the latest evidence of the presence of C. knoblochi in a steppe biome just prior to its extinction. Considering the age of Tsagaan Agui Layer 4, where a camelid cannon bone was found in an undisturbed deposit (59,000–44,000 BP; Derevianko et al., 2000b) and characteristics of the associated archaeological complex and the presence of a bone tool industry (Khatsenovich et al., 2021) that implicitly support this age, camels occupied the Tsagaan Agui region during MIS 3. Pollen analysis indicates increasing aridification and cooling throughout this period, in comparison with underlying layers, with predominant grasses and shrubs in the pollen spectra as well as small quantities of spruce (Picea), pine (Pinus), and birch (Betula) (Derevianko et al., 2000b). The last C. knoblochi were present at Tsagaan Agui probably during the Last Glacial Maximum (ca. 26.5–19 ka), the presumptive age of Layer 2, determined by ESR in the late 1990s (Derevianko et al., 2004). This was a period of increasing aridification when semi-desert landscapes likely replaced steppe biomes.
5 Camel-Human Interactions
Domesticated camels have played a pivotal role in human societies inhabiting arid zones in both prehistoric and historic times. Camels can carry significant burdens over long distances and are the source of organic resources including dung, meat, milk, leather, wool, and bones (Figure 6). The significance of this domesticated animal is documented by numerous petroglyphs occurring throughout arid zones of Mongolia since the Bronze Age. One example includes camels depicted on dark rocks at Ikher Gashuur, in the southern Gobi Desert, that have very tall and thick humps, sometimes higher than the length of the animals’ legs (Figure 7). They are often depicted in herds, sometimes accompanied by people riding horses or other camels (Figure 7: 1–2). Another common rock art motif is the image of a human leading a camel (Figure 7: 3–4) and, less commonly, people protecting camels from their natural enemies (Figure 7: 5). All such petroglyphs reflect close relationships between humans and domesticated camels, but there is also rarer petroglyphic evidence of human interactions with wild camels. In such rock art panels, C. ferus is portrayed with small, often triangular humps, compared with their domesticated counterparts. One such depiction occurs at Zamyn Baruun Barsgai in Noyon County, Ömnögovi Province in which a person on horseback fends off a morphologically wild camel with a hatchet to protect an apparently domesticated camel (Figure 7: 6).

FIGURE 6. Herds of modern domesticated C. bactrianus near Tsagaan Agui Cave in the Gobi Altai Region and Holboljin Nuur Lake in the Valley of the Lakes Mongolia (photos by A. M. Khatsenovich, 2021).

FIGURE 7. Bronze Age camel petroglyphs in the Gobi Desert: 1–4–Ikher Gashuun, Noyon County, Ömnögovi Province; 5–Bichigtiyn-am, Bayanlig sum, Bayankhongor Province; 6–Zamyn Baruun Barsgai, Noyon County, Ömnögovi Province.
The only known presumed Paleolithic image of camel is a pictograph in Khoid Tsenkheriin Agui Cave in the Mongolian Altai Mountains of Mankhan County, Khovd Province (Okladnikov 1972; Novgorodova 1984). There, several animals, including mammoths and ostriches, were painted as realistic contour images on walls inside of the cave with red and brownish red organic pigments. These realistic images are regarded as having been painted in the so-called “Kalgutinsky style,” distinct from images typical of later Neolithic and Bronze Age rock art (Molodin et al., 2019; but see; Miklashevich 2020). This fact, and the depiction of Pleistocene animal species, suggest that these associated images of mammoths, camels and ostriches are of Pleistocene age. One widely known image was redrawn and published by Okladnikov (1972): a walking camel appears full-faced with distinct ears and a thickly outlined body (Figure 8: 1). Landscape is depicted under its feet. A recent photo, taken by one of us (Y.Ts.) in Khoid Tsenkheriin Agui Cave, and enhanced in iDStretch (version 2.2 created by Jon Harmon) (Figure 8: 3–4), suggests that the camel’s head is depicted in profile, exhibiting only a small right ear (Figure 8: 2). The landscape depicted under the animal’s legs more closely resembles a hill.
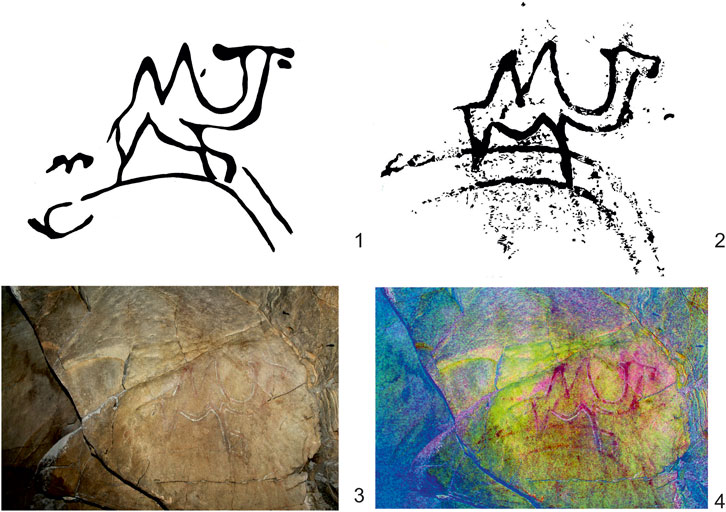
FIGURE 8. Purportedly Paleolithic image of a camel in Khoid Tsenkheriin Agui Cave, Mongolian Altai: 1–widely published drawing by Academician A.P. Okladnikov in the 1970s (Okladnikov 1972); 2, 3, 4–drawing made from a photograph taken for this study and enhanced in iDStretch (version 2.2., created by Jon Harmon).
Evidence of human-camel interactions can also be detected in analyzed bone samples from archaeological contexts, including the Camelus metacarpal from Tsagaan Agui Cave Layer 4. This large bone was flaked by human agency, probably to access the bone’s marrow cavity: it was intentionally split with negative conchoidal fractures apparent on the proximal epiphysis (Figure 2: 2). The other end of the bone was utilized after having been split; it exhibits negative scars of spall removals, most likely unintentional, on the narrow side of the fracture (Figure 9A). It is difficult to determine if this animal was hunted or scavenged by humans, but we know that cave hyaenas (Crocuta spelaea), notorious scavengers whose abundant coprolites were found in Tsagaan Agui Cave Layers 2-5, occupied the grotto concurrently with or closely following the human presence there (Figure 10: 1–2). Traces of hyaena gnawing are visible on this metacarpal above the anthropogenic damage apparent on the bone (Figure 9B).
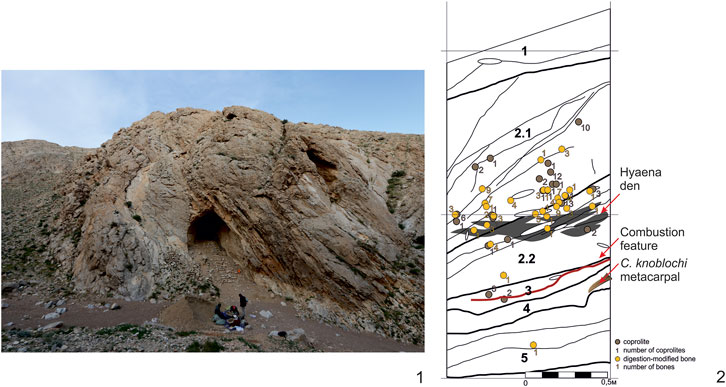
FIGURE 9. Evidence of human-animal interaction at Tsagaan Agui Cave, Gobi Altai Region, Mongolia: 1 – view of the cave; 2 – cross-section of Pit 2021/2 indicating hyaena dens, coprolites and the position of C. knoblochi metacarpal in Layer 4.

FIGURE 10. C. knoblochi metacarpal from Tsagaan Agui Cave, Layer 4 exhibiting traces of anthropogenic splitting with hyaena gnawing above.
Conclusion
The results of our study help fill the chronological gap between Pliocene Paracamelus spp. and Holocene wild camels in Mongolia. They indicate that Camelus knoblochi occupied eastern Central Asia to the end of the Pleistocene and inhabited mainly steppe biomes, expanding to forest-steppe zones near the time of its extinction, which was most likely the consequence of increasing aridification of its natural habitat. The latest known regional evidence of C. knoblochi, as well as Crocuta spelaea, occurs in Tsagaan Agui Cave, Layer 2, presumably dating no earlier than the Last Glacial Maximum (ca. 26.5 ka). The wild Bactrian camel, Camelus ferus, appeared in Mongolia during the Late Pleistocene, and it is possible this species was already adapted to harsh desert environments which may have further reduced the geographic range of Camelus knoblochi through competitive exclusion. Camels played a pivotal role in human adaptations in historic and, likely, Paleolithic times as well. Because modern Camelus ferus exhibits genetic differences from modern domesticated C. bactrianus in Mongolia, the latter of whose ancestry is still poorly understood, intriguing possibilities for the study of Camelus knoblochi and its relationship to both C. ferus and, ultimately, C. bactrianus emerge.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
Ethics Statement
Ethical review and approval was not required for the animal study because We studied fossil remains of extinct Pleistocene species.
Author Contributions
AKh, JO, AD, ER, BG designed research; AKh, AK, JO, ER, BG, DB, and YT performed research; AK, AKh, JO, YT, and DM analyzed data; AK, AKh, JO, and YT wrote the paper.
Funding
Faunal studies were supported by the Russian Scientific Foundation, Project 19-78-10112. Field investigations were undertaken with support from the Leakey Foundation and the Je Tsongkhapa Endowment for Central and Inner Asian Archaeology. Our fieldwork permit (#1–-14) was kindly provided to B. Gunchinsuren and D. Bazargur by the Mongolian Government.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank A.E. Murzintseva of the Museum of the Buryat Scientific Center in Ulan-Ude for granting access to faunal remains currently on exhibit. We appreciate the reviewers’ thoughtful comments on our manuscript.
References
Aubekerova, P. A. (1974). “A New Giant Camel from Pliocene Deposits of the Tekesskaya Depression (Northern Tien Shan),” in Mesocenozoic Flora and Fauna of Southern Kazakhstan. Editor B. S. Kozhamkulova (Almaty: Nauka), 6, 74–83. (In Russian).
Belyeva, E. I. (1937). Materials on Characteristic of Late Tertiary north-western Mongolia (In Russian). Proc. Mongolian Comm. 33, 5–52.
Berkey, C. P., and Morris, F. K. (1927). Geology of Mongolia: A Reconnaissance Report Based on the Investigations of the Years 1922-1923, 2. New York City, New York: American Museum of Natural History.
Bibi, F., and Métais, G. (2016). “Evolutionary History of the Large Herbivores of South and Southeast Asia (Indomalayan Realm),” in The Ecology of Large Herbivores in South and Southeast Asia. Editors F. S. Ashrestani, and M. Sankaran (Berlin/Heidelberg, Germany: Springer), 15–88. doi:10.1007/978-94-017-7570-0_2
Boule, M., and de Chardin, T. (1928). Paleontologie” in La Paléolithique de la Chine Archives d. Memoire, 4. Paris: L’Institut de Paleontologie Humaine, Fondation Albert 1-er Prince de Monaco.
Burger, P. A., and Palmieri, N. (2013). Estimating the Population Mutation Rate from Ade novoAssembled Bactrian Camel Genome and Cross-Species Comparison with Dromedary ESTs. Jhered 105, 933–940. doi:10.1093/jhered/est005
Colbert, E. H. (1935). Siwalik Mammals in the American Museum of Natural History. Trans. Am. Philos. Soc. 26, 278–294. doi:10.2307/1005467
Derevianko, A. P., Brantingham, P. J., Olsen, J. W., and Tseveendorj, D. (2004). “Initial Upper Paleolithic Blade Industries from the North-central Gobi Desert, Mongolia,” in The Early Upper Paleolithic beyond Western Europe. Editors P. J. Brantingham, S. L. Kuhn, and K. W. Kerry (Berkeley, California, United States: University of California Press), 207–222.
Derevianko, A. P., Olsen, J. W., Tseveendorzh, D., Petrin, V. T., Gladyshev, S. A., Zenin, A. N., et al. (2000a). Archaeological Studies Carried Out by the Joint Russian-Mongolian-American Expedition in Mongolia in 1997 and 1998. Novosibirsk: IAET SB RAS. (In Russian).
Derevianko, A. P., Olsen, J. W., Tsevendorzh, D., Krivoshapkin, A. I., Petrin, V. T., and Brantingham, P. D. (2000b). Stratified Cave Site of Tsagaan Agui in the Gobi Altai (Mongolia). (In Russian). Archaeology, Ethnology Anthropol. Eurasia 1, 23–36.
Derevianko, A. P., and Petrin, V. T. (1995). Investigation of the Tsagaan Agui Cave in the Southern Side of Gobi Altai in Mongolia. Novosibirsk: IAET SO RAN Publ. (In Russian).
Devyatkin, E. V. (1970). “Cenozoic Geology of Western Mongolia,” in Mesozoic and Cenozoic Geology of western Mongolia: Proceedings of the Joint Soviet-Mongolian Research Geological Expedition, Moscow, 1970, 44–102. (In Russian).
Devyatkin, E. V. (1981). The Cenozoic in Inner Asia: Stratigraphy, Geochronology, Correlation. Moscow: Nauka. (In Russian).
Devyatkin, E. V., Zazhigin, V. S., and Liskun, I. G. (1968). The First Findings of Faunal Remains Belonging to Small Mammals in Pliocene of Tyva and Western Mongolia. (In Russian). Doklady AS SSSR 2 (183), 404.
Dong, W., Hou, Y.-M., Yang, Z.-M., Zhang, L.-M., Zhang, S.-Q., and Liu, Y. (2014). Late Pleistocene Mammalian Fauna from Wulanmulan Paleolithic Site, Nei Mongol, China. Quat. Int. 347, 139–147. doi:10.1016/j.quaint.2014.05.051
Eisenmann, V., and Beckouche, S. (1986). “Identification and Discrimination of Metapodials from Pleistocene and Modern Equus, Wild and Domestic,” in Equids in the Ancient World. Editors H. Meadow, and H.-P. Uerpmann (Wiesbaden: L. Reichert Verlag), 116–163.
Falconer, H., and Cautley, P. T. (1836). Note on the Fossil Camel of the Sivalik Hills. Asiatic Researches 19 (1), 115–134.
Germonpré, M., and Lbova, L. (1996). Mammalian Remains from the Upper Palaeolithic Site of Kamenka, Buryatia (Siberia). J. Archaeological Sci. 23, 35–57. doi:10.1006/jasc.1996.0004
Grunert, J., Lehmkuhl, F., and Walther, M. (2000). Paleoclimatic Evolution of the Uvs Nuur basin and Adjacent Areas (Western Mongolia). Quat. Int. 65-66 (66), 171–192. doi:10.1016/s1040-6182(99)00043-9
Janz, L., Rosen, A. M., Bukhchuluun, D., and Odsuren, D. (2021). Zaraa Uul: An Archaeological Record of Pleistocene-Holocene Palaeoecology in the Gobi Desert. PLOS ONE 16 (4), e0249848. doi:10.1371/journal.pone.0249848
Khatsenovich, A. M., Tserendagva, Ya., Tserendagva, Y., Bazargur, D., Marchenko, D. V., Rybin, E. P., et al. (2021). Preliminary Results of the 2021 Excavation Campaign at Tsagaan Agui Cave in the Gobi Altai Region, Mongolia. Paeasnt 27, 321–329. doi:10.17746/2658-6193.2021.27.0321-0329
Khaveson, Ya. I. (1954). Tertiary Camels of Eastern Hemisphere (Genus Paracamelus) (In Russian). Proc. Paleontological Inst. RAS 47, 100–161.
Klementiev, A. M., Ondar, S. O., and Dargyn-ool, D. V. (2021). “Villafranchian Fauna in Tyva,” in Dynamics and Interaction of Earth Geospheres. Editors T. N. Zhilina, N. M. Semyonova, S. L. Budaev, and N. S. Evseeva (Tomsk: Tomsk CNTI), 1, 182–184. In Russian.
Kozhamkulova, B. S. (1963). “Additional Data on Fossil Camels of Kazakhstan,” in Materials on History of Fauna and Flora in Kazakhstan (Nauka), 4, 22–33. (In Russian).Almaty
Lopatin, A. V. (2019). “The Main Results of Research on Fossil Mammals of Mongolia,” in 50 Years of the Joint Russian-Mongolian Paleontological Expedition, Results. Editor S. V. Rozhnov (Moscow: RAS), 136–149. (In Russian).
Martynovich, N. (2002). Pleistocene Birds from Tsagaan Agui Cave (Gobian Altai). Acta Zoologica Cracoviensia 45, 283–292.
Miklashevich, E. A. (2020). Similarity of the Earliest Rock Art in South Siberia and Central Asia and the Issue of Their Attribution. Brief Commun. Inst. Archaeology 261, 82–98. doi:10.25681/IARAS.0130-2620.261.82-98
Molodin, V. I., Geneste, J.-M., Zotkina, L. V., Cheremisin, D. V., and Cretin, C. (2019). The “Kalgutinsky” Style in the Rock Art of Central Asia. Archaeology, Ethnology Anthropol. Eurasia 3 (47), 12–26. doi:10.17746/1563-0110.2019.47.3.012-026
Okladnikov, A. P., and Troitsky, S. L. (1967). Study of Quaternary Deposits and Paleolithic of Mongolia. Bull. Comm. Study Quat. 33, 3–30. (In Russian).
Ovodov, N. D. (2001). “Pleistocene Fauna of Tsagaan Agui Cave (Mongolia),” in Environmental Conditions, History and Culture of Western Mongolia and Neighboring Regions. Editor A. S. Revushkin (Tomsk: Tomsk CNTI), 146–147. (In Russian).
Perevalov, A. V., and Rezanov, I. N. (1997). First Attempt of Radiothermoluminescent Dating of Anthropogen Deposits in Southwestern Transbaikalia. Russ. Geology. Geophys. 7 (38), 12–82.
Qi, G. Q. (1975). Quaternary Mammalian Fossils from Salawusu River District, Nei Mongol. Vertebrata PalAsiatica 13, 239–249.
Ravsky, E. I., Aleksandrova, L. P., Vangengeim, E. A., Gerbova, V. G., and Golubeva, L. V. (1964). Anthropogenic Deposits in the South of Eastern Siberia. Moscow: AS SSSR Press.
Sharapov, S. H. (1986). Kuruksai Complex of the Late Pliocene Mammals in Afghan-Tajik Depression. Dushanbe: Donish. (In Russian).
Titov, V. V. (2008). Habitat Conditions for Camelus Knoblochi and Factors in its Extinction. Quat. Int. 179, 120–125. doi:10.1016/j.quaint.2007.10.022
Vangengeim, E. A., and Gerbova, V. G. (1962). Some Data on Time and Conditions of Transbaikalian Sand Accumulation. Bull. Comm. Study Quat. 19, 268–274. (In Russian).
Vangengeim, E. A. (1977). Paleontological Basis for Stratigraphy of Anthropogen in Northern Asia (Based on Mammals). Moscow: Nauka.
Vangengeim, E. A., Sotnikova, M. V., Alexeeva, L. I., Vislobokova, I. A., Zhegallo, V. I., Zazhigin, V. S., et al. (1988). Biostratigraphy of Late Pliocene – Early Pleistocene in Tadjikistan (Based on Mammalian Fauna). Moscow: Nauka. (In Russian).
Vangengeim, E. A., Zhegallo, V. I., and Zazhigin, V. S. (1972). The Stages of Evolution of the Mammalian Fauna in the Late Neogene and Early Anthropogen in Northern Asia. Geology. Geophys. 6, 58–65.
Vasiliev, S. K. (2008). Late Pleistocene bison (bison P. Priscus Bojanis, 1827) from the southeastern Part of Western Siberia. Archaeology, Ethnology Anthropol. Eurasia 34, 34–56. doi:10.1016/j.aeae.2008.07.004
Vasiliev, S. K. (2016). “New Findings of Camelus Knoblochi (Nehring, 1901) on the Altai Plain,” in Theriofauna in Russia and Neighboring Territories (Moscow: KMK), 16. (In Russian).
Vasiliev, S. K., Serednyov, M. A., and Milutin, K. I. (2018). The Pleistocene Megafauna Fossil Remains from the Alluvial Sites of Bibikha, Taradanovo on the Ob River (Novosibirsk Region) and Chumysh River (Altai Region). Paeasnt 24, 47–51. doi:10.17746/2658-6193.2018.24.047-051
Vasiliev, S. K., Serednyov, M. A., Milyutin, K. I., Slyusarenko, I. Yu., Kozlikin, M. B., and Chekha, A. M. (2015). Collecting of the Theriofaunal Materials at the Rivers Chumysh (Altai Region) and Ob Near Bibikha Village (Novosibirsk Region) in 2015. Probl. Archaeology, Ethnography, Anthropol. Siberia Neighboring Territories 21, 36–40.
Vereshchagin, N. K. (1956). About Previous Distribution of Some Ungulates in the Region between European-Kazakh and Central Asian Steppes. Zoolog. J. 35 (10), 1541–1553. (In Russian).
Wang, Z., Jirimutu, G., Wang, Z., Ding, G., Chen, G., Sun, Y., et al. (2012). Genome Sequences of Wild and Domestic Bactrian Camels. Nat. Commun. 3, 1202. doi:10.1038/ncomms2192
Wu, H., Guang, X., Al-Fageeh, M. B., Cao, J., Pan, S., Zhou, H., et al. (2014). Camelid Genomes Reveal Evolution and Adaptation to Desert Environments. Nat. Commun. 5, 5188. doi:10.1038/ncomms6188
Yamanaka, I. (1993). L'Industrie lithique du site de Shuidonggou (Choei-Tong-Keou) dans L'Ordos en Chine, 32. Kyoto, Japan: Bulletin of Faculty of Arts and Sciences, Kyoto University, 105–154.
Young, A. J., and Reeves, J. T. (2002). “Human Adaptation to High Terrestrial Altitude,” in Medical Aspects of Harsh Environments. Editors K. B. Pandolf, and R. E. Burr (Virginia: Office of the Surgeon General), 2, 645–688.
Yuan, J., Westbury, M. V., Chen, S., Hu, J., Zhang, F., Wang, S., et al. Palaeogenomics Reveal a Hybrid Origin of the World's Largest Camelus Species. BioRxiv. (in press). doi:10.1101/2021.10.14.464381
Keywords: paleontology, taxonomy, eastern Central Asia, Mongolia, pleistocene, Camelus knoblochi, human-animal interactions
Citation: Klementiev AM, Khatsenovich AM, Tserendagva Y, Rybin EP, Bazargur D, Marchenko DV, Gunchinsuren B, Derevianko AP and Olsen JW (2022) First Documented Camelus knoblochi Nehring (1901) and Fossil Camelus ferus Przewalski (1878) From Late Pleistocene Archaeological Contexts in Mongolia. Front. Earth Sci. 10:861163. doi: 10.3389/feart.2022.861163
Received: 24 January 2022; Accepted: 22 February 2022;
Published: 14 March 2022.
Edited by:
Shuangquan Zhang, Institute of Vertebrate Paleontology and Paleoanthropology (CAS), ChinaReviewed by:
Vadim Titov, Southern Scientific Center (RAS), RussiaWei Dong, Institute of Vertebrate Paleontology and Paleoanthropology (CAS), China
Copyright © 2022 Klementiev, Khatsenovich, Tserendagva, Rybin, Bazargur, Marchenko, Gunchinsuren, Derevianko and Olsen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexey M. Klementiev, a2xlbS1hbEB5YW5kZXgucnU=; Arina M. Khatsenovich, YXJjaGVvbW9uZ29saWFAZ21haWwuY29t
†These authors have contributed equally to this work and share senior authorship