 Yanan Zhang
Yanan Zhang Yunhuan Liu
Yunhuan Liu- School of Earth Science and Resources, Chang’an University, Xi’an, China
Microscopic medusozoans from the Cambrian Fortunian stage of South China are well known for their exceptionally preserved embryos and elongated tubes. However, additional details of their morphology remain unclear. This paper describes new medusozoan fossils showing the whole apical complement and covering the morphological integrity of Qinscyphus. The apical part of Qinscyphus is considered to be soft during early ontogeny, and the inverted pentagonal pyramid may gradually form with growth and development. This discovery of the apical complement of Qinscyphus is novel and essential to complement the external morphology of early medusozoan fossils. More importantly, the new specimens have different annuli with triangular thickenings, providing a more comprehensive view on the developmental sequence of Qinscyphus. Therefore, this work allows a better understanding of early medusozoans ecology and evolution.
1 Introduction
The origin and evolution of medusa are the focus of both paleontologists and modern biologists (Park et al., 2012; Wang D et al., 2020). The most distinctive characteristics of medusozoans are their radial symmetry and their ability to swim (Han et al., 2020). Paleontologists have found numerous medusozoans in the early Cambrian (531.8–536.4 Ma) (Dong et al., 2013; Han et al., 2013; Liu et al., 2014a; Steiner et al., 2014; Dong et al., 2016; Liu et al., 2017; Wang et al., 2017; Shao et al., 2018b; Han et al., 2018; Shao et al., 2019a; Wang X et al., 2020). In the Zhangjiagou Section, South China, various Fortunian radial animals were found such as Olivooides, Quadrapyrgites, and Hexaconularia (Conway Morris and Chen, 1992; Van Iten et al., 2006; Van Iten et al., 2010; Liu et al., 2014b; Van Iten et al., 2014; Liu et al., 2017). These taxa were compared to cnidarians as they show a radial body plan with an apical–abapical differentiation (Conway Morris and Chen, 1992; Van Iten et al., 2006; Van Iten et al., 2010; Liu et al., 2014b; Van Iten et al., 2014; Liu et al., 2017). However, most olivooids structures are lacking from living cnidarians. Olivooid tubes had prominent annulations and fine longitudinal striations with the oral aperture being the only opening of these tubes (Liu et al., 2014a; Steiner et al., 2014). Therefore, olivooids were considered as coronate scyphozoans, and their elongate tubes were considered as polyp stages that develop after hatching (Conway Morris and Chen, 1992; Bengtson and Yue, 1997; Van Iten et al., 2006; Van Iten et al., 2010; Dong et al., 2013; Han et al., 2013; Liu et al., 2014a; Liu et al., 2017; Wang et al., 2017).
The microscopic animal, Qinscyphus, was first reported from the early Cambrian Kuanchuanpu Formation at the Zhangjiagou section in South China (Liu et al., 2017). Qinscyphus necopinus is a cup-shaped animal with an embryonic tissue (apical part) and a post-embryonic tissue (abapical part). The embryonic tissue formed an inverted pentagonal pyramid with a smooth surface (Liu et al., 2017; Shao et al., 2018a). Post-embryonic tissue was tube-shaped with densely spaced and slightly raised annuli (Liu et al., 2017). Several circlets of triangular thickenings formed five longitudinal rows, pentaradially arranged on post-embryonic tissues, each straddling several annuli (Shao et al., 2018a). Moreover, Qinscyphus might be a direct developer due to the comparable morphology of the oral part in both embryos and adults (Shao et al., 2018a; Qin et al., 2020). Qinscyphus was interpreted as a possible coronate scyphozoan for its pentagonal symmetry, its tube (comparable to a periderm), and its oral part (its only opening) (Liu et al., 2017). This paper reports five new three-dimensionally phosphatized specimens of Q. necopinus from South China; from which four are complete, unlike the fragmentary nature of previously described material, including partially the holotype.
2 Materials and methods
The studied specimens are deposited at the University Museum of Chang’an University (UMCU), Xi’an, China. They were extracted from the phosphatic limestone of the lower part of the second member of the Kuanchuanpu Formation, Zhangjiagou section, Xixiang County, southern Shaanxi Province, South China (Figure 1). The lithostratigraphy and biostratigraphy of the Zhangjiagou section were studied by Li (1984) and Steiner et al. (2004a), Steiner et al. (2007). The same locality and horizon have previously yielded abundant and beautifully preserved microfossils, such as specimens of Olivooides (Liu et al., 2014a; Steiner et al., 2014; Liu et al., 2017; Shao et al., 2018b; Shao et al., 2019a), Pseudooides, the oldest known priapulid–like cycloneuralians as well as other scalidophorans (Liu et al., 2014b; Shao et al., 2016; Shao et al., 2018b; Shao et al., 2019b; Wang et al., 2019; Wang D et al., 2020) and possibe Saccorhytus (Han et al., 2017; Liu et al., 2022). The fossil-bearing horizon correlates with the Anabarites trisulcatus–Protohertzina anabarica assemblage zone (Steiner et al., 2007; Steiner et al., 2014), which has an estimated age of 531.8–536.4 Ma and falls within the Fortunian Stage (Peng et al., 2020).
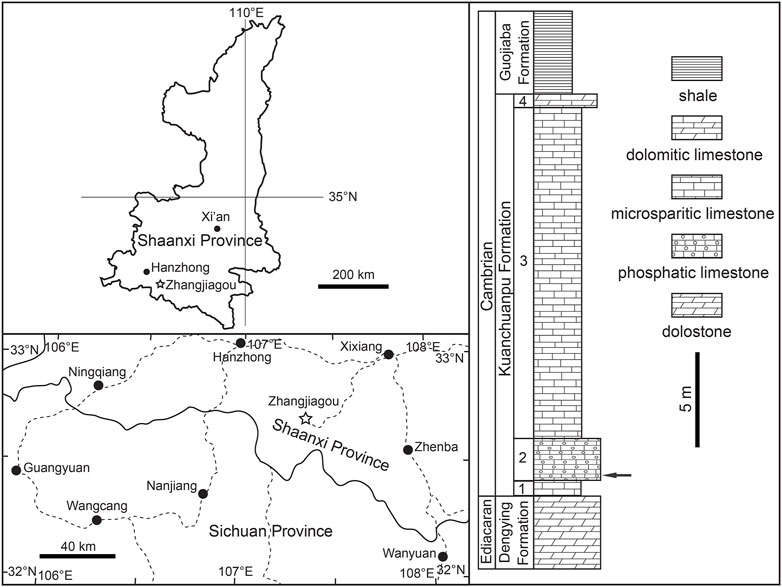
FIGURE 1. Location map and stratigraphic column of the Zhangjiagou Section in southern Shaanxi Province, South China. The key horizon yielding the current specimens is denoted by an arrow.
Nearly 300 kg of rock samples from the key horizon at the Zhangjiagou section (Figure 1) were cracked into football-sized pieces. Each sample was put in a plastic pail or basin with a capacity of 20 L macerated using diluted acetic acid (∼8%). The sieving and changing acid cycle requires three to 7 days, depending on the temperature (ambient temperature around 40°C in summer and 20°C in winter). Undissolvable residues dried naturally and were then handpicked under a binocular microscope. The microfossils were picked out and mounted on aluminium stubs for observation under a Hitachi S4800 environmental scanning electron microscope at Chang’an University, Xi’an. The figures were processed using Adobe Photoshop CS5.
3 Results
Qinscyphus is rarely found in the Kuanchuanpu Formation, and most specimens in the collection are fragmented. The newly discovered specimens are three-dimensionally preserved and only four intact tubes with a preserved apical complement have been found (Figures 2A,H and Figures 3A,F).

FIGURE 2. SEM images of Qinscyphus necopinus respectively with one to three circlets of triangular thickenings. (A,E,H) lateral views of 2021XQB102, 2021XQB211, 2021XQB107; (B,F,I) oral views; (C,G,J) close-up oral views of (B,H,I); (D) the enlargement of the apical part of (A); In (I), white arrows denote the perradial belts with triangular thickenings, and black arrows denote the interradial belts. The scale bar represents 50 μm in (F), and 100 μm in other images.
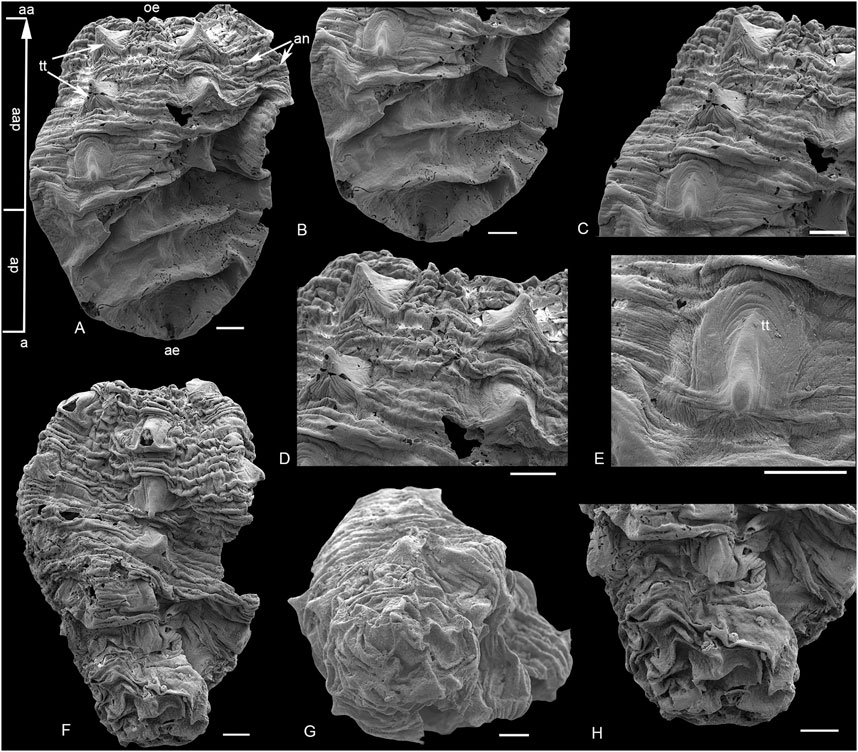
FIGURE 3. SEM images of Qinscyphus necopinus respectively with three and six circlets of triangular thickenings. (A–E) 2021XQB105; (B) apical part; (C,D) enlargement of postembryonic crests with triangular thickenings; (E) enlargement of triangular thickening; (F–H) 2021XQB106; (G) apical view; (H) enlargement of the apical part. Scale bars: 100 μm. a = apical; aa = abapical; ap = apical part; aap = abapical part; ae = apical end; an = annuli; e = edge; oe = oral end; tt = triangular thickening.
Only one specimen (Figure 2A) reveals five longitudinal edges by slight folds corresponding to five groups of triangular thickenings (Figure 2D). These edges merge at the apical end (Figure 2A), with both length and width of 950 μm. The apical end is similar to the shape of the hatchlings of Olivooides (Figure 4), but has a smooth surface (Figure 3B). The apical end represents a small percentage of the entire organism (Figure 2D). However, for the remaining specimens, the apical part shows multiple distinct transverse folds, with no apparent longitudinal edges (Figures 3A,B,G,H).

FIGURE 4. SEM images of Olivooides multisulcatus. (A,D) XX21CHD01-1; (B,E) XX19-b05; (C,F) XX21CHD01-1; (D,E) enlargement of triangular thickening; (F) close-up oral views. Scale bars: 100 μm. a = apical; aa = abapical; ap = apical part; aap = abapical part; ae = apical end; oe = oral end; ss = stellate structure; tt = triangular thickening.
The post-embryonic tissue (abapical part) is tube-shaped with densely spaced and slightly raised annuli (Figures 3A,F). Five groups of triangular thickenings are developed radially on the abapical part (Figure 2I). In each group, the triangular thickening occurs repetitively every seven annuli, and straddles the next four to five annuli (Figures 3C,D). These annuli are only slightly raised, rather than strongly extended outwards to form ridges (Fig. 2C, 3D). The largest complete specimen with six circlets of triangular thickenings is 1.4 mm long and 1.1 mm wide (Figure 3F). Two oral views indicate that the new crest with five distinct triangular thickenings formed at the oral part (Figures 2F,I). Around the oral aperture, perradial belts and interradial belting are demarcated by marginal furrows, with a median furrow bisecting each interradial belt (Figures 2F,G,I,J). The morphology and number of triangular thickenings at the oral part are comparable to Olivooides multisulcatus (Figure 4F), while the new crest with triangular thickenings did not occur continuously but spaced several annuli to the next crest (Figure 3C). The new specimens are well preserved and have different circlets of triangular thickenings: one (Figure 2A), two (Figure 2E), three (Figure 2H, Figure 3A), and six circlets (Figure 3F). The surface of these triangular thickenings shows peristaltic deformation and distortion (Figure 3E), suggesting that the triangular tickenings were soft.
4 Discussion
The Cambrian fossil embryos are represented by taxa of Markuelia, Pseudooides, Olivooides, Quadrapyrgites and Qinscyphus (Dong et al., 2013; Steiner et al., 2014; Dong et al., 2016; Duan et al., 2017; Liu et al., 2017; Shao et al., 2018a). However, the affinity of many Fortunian radial animals remains debatable. For example, Pseudooides was previously regarded as the embryonic stage of some co-occurring arthropod-like fossil (Steiner et al., 2004b). But it was challenged because the features were unusual for any arthropod or even bilaterian (Donoghue et al., 2006). Olivooides was proposed as a coronate scyphozoan (Bengtson and Yue, 1997; Yue and Bengtson, 1999; Dong et al., 2013; Dong et al., 2016) or an echinoderm (Chen, 2004), a cubozoan (Han et al., 2016a; Han et al., 2016b), a stem-group cycloneuralian (Steiner et al., 2014) or a diploblastic stem-group eumetazoan (Yasui et al., 2013). Quadrapyrgites might be the sister group of Olivooides (Steiner et al., 2014), and has been proposed as a stem-group cycloneuralian (Steiner et al., 2014), a coronate scyphozoan (Liu et al., 2014b), or a cubozoan (Han et al., 2016a). Qinscyphus might also be a coronate scyphozoan (Liu et al., 2017; Shao et al., 2018a). The coronate scyphozoan hypothesis for Olivooides, Quadrapyrgites and Qinscyphus is favoured here.
The possible polypoid cnidarians from the Fortunian stage correspond to Anabaritidae (Conway Morris and Chen, 1989; Kouchinsky et al., 2009), Hexangulaconulariidae (Qian and Bengtson, 1989; Van Iten et al., 2010; Steiner et al., 2014), Carinachitidae (Qian, 1977; Conway Morris and Chen, 1992; Yue and Bengtson, 1999; Liu et al., 2005; Liu et al., 2011; Han et al., 2018), Olivooidae (Steiner et al., 2010; Steiner et al., 2014; Han et al., 2016a; Han et al., 2016b) and Family uncertain (Steiner et al., 2004a; Dong et al., 2013; Shao et al., 2015). They had comparable radial symmetry, tube morphology, and developmental cycles. Furthermore, Olivooidae might include several genera with similar morphologies and developmental modes: Olivooides (Qian, 1977; Yasui et al., 2013; Steiner et al., 2014), Quadrapyrgites (Li, 1984; Li et al., 2007; Liu et al., 2009; Liu et al., 2014a; Steiner et al., 2014; Shao et al., 2019a), Qinscyphus (Liu et al., 2017; Shao et al., 2018a), Octapyrgites (Guo et al., 2020).
Olivooides multisulcatus (Figure 4) represents the closest analogue to Q. necopinus. Both are cup-shaped with an apical part and an abapical part (Figure 5). They had a similar body of pentaradial symmetry, five longitudinal rows of triangular thickenings and were found in coeval strata from South China. However, Q. necopinus and O. multisulcatus still had significant differences (Table 1). The apical part of Q. necopinus lacked tiny stellate protuberances on the external surface (Figures 4D,E). The newly described material shows multiple distinct transverse folds (Figure 2A) rather than an apparent longitudinal edge. Moreover, the triangular thickennings of O. multisulcatus occurred continuously and independently on each annulus (Figures 4A–C), whereas for Q. necopinus they appeared every four to seven annuli and straddled the next four to five annuli (Figures 3A,F and Figures 5C,D). Qinscyphus necopinus annuli were slightly raised rather than enormously extended ridged ones (Figures 3C,D). Those annuli might have contributed to a better stretching, contraction, and buffering in the living environment. Qinscyphus necopinus and O. multisulcatus also differed from each other by the absence/presence of stellate structures, and the shape of the perradial lobes and interradial belts in their embryonic stages. The perradial lobes of Q. necopinus had an elliptical end. The interradial belts had median and marginal furrows (Figure 5B). However, the interradial lobes of O. multisulcatus were strongly squeezed by the perradial lobes (Figure 5A) (see Shao et al., 2018a).
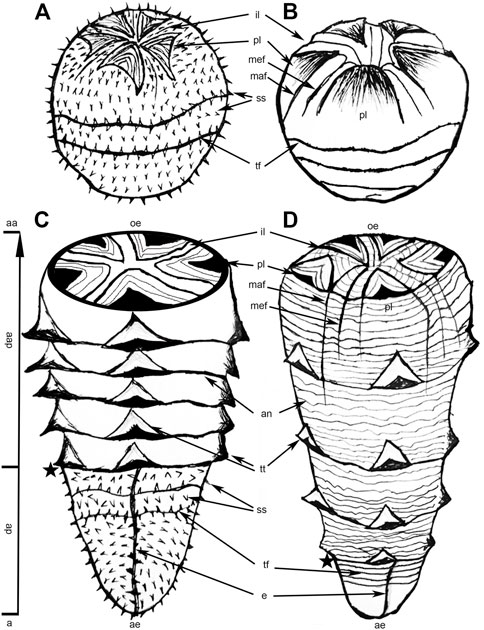
FIGURE 5. A comparative sketch of Olivooides multisulcatus and Qinscyphus necopinus. (A) Embryo of Olivooides multisulcatus; (B) embryo of Qinscyphus necopinus; (C) hatchling of Olivooides multisulcatus; (D) hatchling of Qinscyphus necopinus; Asterisks denote the boundary between the apical and abapical parts. (B) and (D) modified from Shao et al., 2018a. a = apical; aa = abapical; ap = apical part; aap = abapical part; ae = apical end; an = annuli; e = edge; il = interradial lobe; maf = marginal furrow; mef = median furrow; oe = oral end; pl = perradial lobe; ss = stellate structure; tf = transverse fold; tt = triangular thickening.
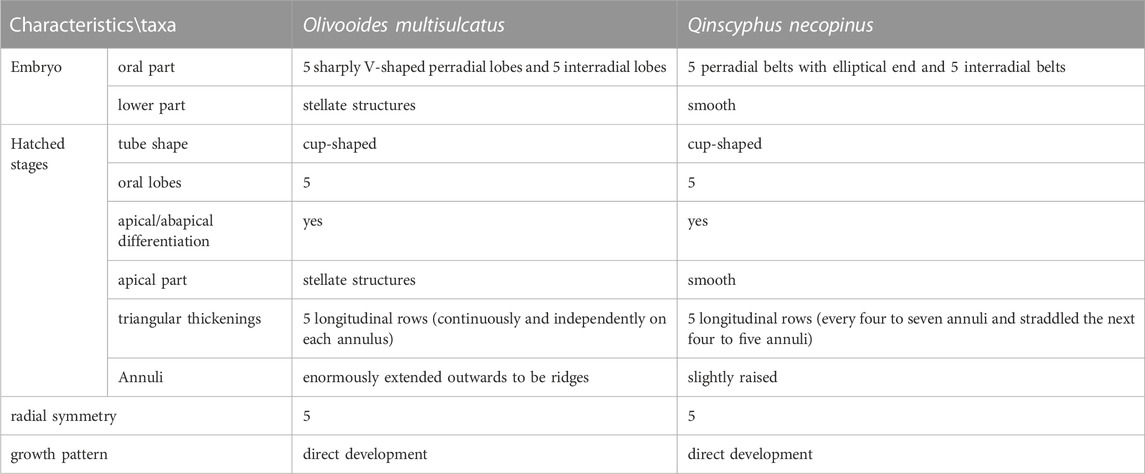
TABLE 1. Morphological comparisons of Olivooides multisulcatus and Qinscyphus necopinus.
Previous work suggested that the apical part of Q. necopinus is an inverted pentagonal pyramid with five prominent longitudinal edges (Liu et al., 2017; Shao et al., 2018a; Qin et al., 2020). However, the new specimens did not show this feature (Figure 2; Figure 3). The apical part of the new specimens had multiple distinct transverse folds (Figures 3A,F), but no apparent longitudinal edges. Thus, there are two possible forms of the apical part of Qinscyphus: an inverted pentagonal cone or a circular one. This contrast might be resulting from intraspecific variations. However, considering that the new fossils show a circular cone in early ontogenetic stages (Liu et al., 2017), we suggest that the shape of the apical part might gradually become an inverted pentagonal pyramid with growth and development.
We consider that the embryonic tissue (apical part) of Qinscyphus necopinus was soft during early ontogeny (fossils with fewer circlets of triangular thickenings) (Figure 2A). As such, the transverse folds on the embryonic tissue might be taphonomic artifacts due to the soft nature of the apical part. And it is probable that this part was smooth during life. Even in adult individuals, the embryonic tissues could still show transverse folds in taxa such as Qinscyphus, Olivooides, and Quadrapygites (Steiner et al., 2014; Dong et al., 2016). These soft transverse folds were absent in previous reconstructions of Olivooides and Quadrapygites (Steiner et al., 2014), with transverse folds only occurring on the upper part of the embryonic tissue. Therefore, we modified the reconstruction of Q. necopinus based on the newly described material, by adding transverse folds on the entire embryonic tissue (Figures 5A,D).
Previously reported Qinscyphus were mostly fragmented. The new specimens preserved intact tubes with apical and abapical parts. They also show one (Figure 2A), two (Figure 2E), three (Figure 2H and Figure 3A), and six (Figure 3F) circlets with triangular thickenings respectively. Combined with previous studies, the developmental sequence of Qinscyphus is reconstructed (Figure 6). The oral part of the embryo had a morphology that is comparable to that of the hatched stages (Shao et al., 2018a), demonstrating that Qinscyphus underwent direct development. After hatching, the tube with transverse annuli and pentaradially arranged triangular thickenings began to develop. New annuli and triangular thickenings might have regenerated from within the abapical end. The circlet of triangular thickenings at the abapical end would have evaginated to form a new circlet of triangular thickenings. Qinscyphus would have elongated its length through the addition of annuli and triangular thickenings at the abapical end (Liu et al., 2017; Shao et al., 2018a).
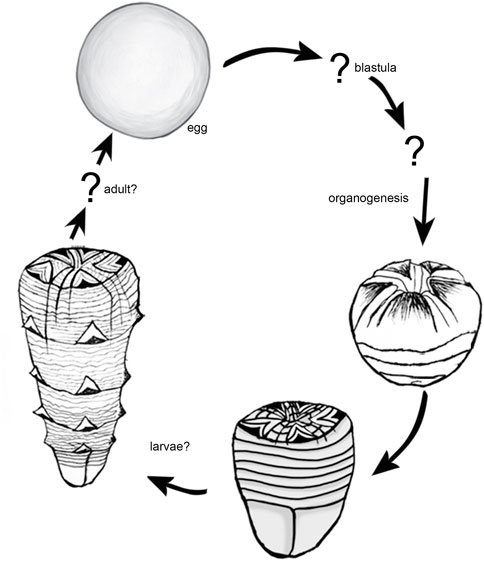
FIGURE 6. Reconstruction of the developmental cycle of Qinscyphus necopinus.
5 Conclusion
The newly discovered fossils with the whole apical part complete the morphology of Qinscyphus. The apical portion of Q. necopinus is assumed soft during early ontogenetic stages. And, it does not always manifest as an inverted pentagonal pyramid. More importantly, the new specimens have different annuli with triangular thickenings, completing the developmental sequence and expanding the morphological disparity of Q. necopinus.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
YZ coordinated the field work, preparation and curation of fossils, wrote the manuscript, and processed the images. YL edited the manuscript. TS and JQ assisted in collecting the rock samples in the field. All authors contributed to the article and approved the submitted version.
Funding
The National Natural Science Foundation of China (grant numbers 41872014, 41572007 and 41572009), the Strategic Priority Research Program of Chinese Academy of Sciences (grant number XDB26000000), the State Key Laboratory of Paleobiology and Stratigraphy, Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences (grant number 193123), the Basic Research Plan of Natural Science of Shaanxi Province (grant number 2018JM4002).
Acknowledgments
We thank Xiaoke Sun and Mingjin Liu, as they helped revise some of the manuscript’s images and language issues.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bengtson, S., and Yue, Z. (1997). Fossilized metazoan embryos from the earliest Cambrian. Science 277, 1645–1648. doi:10.1126/science.277.5332.1645
Conway Morris, S., and Chen, M. G. (1989). Lower cambrian anabaritids from SouthSouth China. Geol. Mag. 126, 615–632. doi:10.1017/s0016756800006920
Conway Morris, S., and Chen, M. E. (1992). Carinachitids, hexaconulariids, and punctatus, problematic metazoans from the early cambrian of SouthSouth China. J. Paleontol. 66, 384–406. doi:10.1017/s0022336000033953
Dong, X., Cunningham, J. A., Bengtson, S., Thomas, C. W., Liu, J., Stampanoni, M., et al. (2013). Embryos, polyps and medusae of the early Cambrian scyphozoan Olivooides. P. Roy. Soc. B–Biol. Sci. 280, 20130071. doi:10.1098/rspb.2013.0071
Dong, X., Vargas, K., Cunningham, J. A., Zhang, H., Liu, T., Chen, F., et al. (2016). Developmental biology of the early Cambrian cnidarianOlivooides. Olivooides, Palaeontol. 59, 387–407. doi:10.1111/pala.12231
Donoghue, P. C. J., Bengtson, S., Dong, X., Gostling, N. J., Huldtgren, H., Cunningham, J. A., et al. (2006). Synchrotron x-ray tomographic microscopy of fossil embryos. Nature 442, 680–683. doi:10.1038/nature04890
Duan, B., Dong, X., Porras, L., Vargas, K., Cunnigham, J. A., and Donoghue, P. C. J. (2017). The early Cambrian fossil embryo Pseudooides is a direct - developing cnidarian, not an early ecdysozoan. Proc. R. Soc. B 284, 20172188. doi:10.1098/rspb.2017.2188
Guo, J., Han, J., Van Iten, H., Song, Z., Sun, J., Wang, W., et al. (2020). A new tetraradial olivooid (medusozoa) from the lower cambrian (stage 2) yanjiahe formation, south China. J. Paleontol. 94, 457–466. doi:10.1017/jpa.2019.101
Han, J., Kubota, S., Li, G., Yao, X., Yang, X., Shu, D., et al. (2013). Early Cambrian pentamerous cubozoan embryos from South China. PloS One 8, e70741. doi:10.1371/journal.pone.0070741
Han, J., Kubota, S., Li, G., Ou, Q., Wang, X., Yao, X., et al. (2016a). Divergent evolution of medusozoan symmetric patterns: Evidence from the microanatomy of Cambrian tetramerous cubozoans from South China. Gondwana Res. 31, 150–163. doi:10.1016/j.gr.2015.01.003
Han, J., Li, G. X., Kubota, S., Ou, Q., Toshino, S., Eang, X., et al. (2016b). Internal microanatomy and zoological affinity of the early CambrianOlivooides. Olivooides, Acta Geol. Sin–Engl. 90, 38–65. doi:10.1111/1755-6724.12641
Han, J., Morris, S. C., Ou, Q., Shu, D., and Huang, H. (2017). Meiofaunal deuterostomes from the basal cambrian of Shaanxi (China). Nature 542, 228–231. doi:10.1038/nature21072
Han, J., Li, G., Wang, X., Yang, X., Guo, J., Sasaki, O., et al. (2018). Olivooides–like tube aperture in early Cambrian carinachitids (Medusozoa, Cnidaria). J. Paleontol. 92, 3–13. doi:10.1017/jpa.2017.10
Han, J., Guo, J., Ou, Q., Song, Z., Liu, P., Hao, W., et al. (2020). Evolutionary framework of early Cambrian cnidarians from South China. Earth Sci. Front. 27, 067–078. doi:10.13745/j.esf.sf.2020.6.3
Kouchinsky, A., Bengtson, S., Feng, W., Kutygin, R., and Val'kov, A. (2009). The lower cambrian fossil anabaritids: Affinities, occurrences and systematics. J. Syst. Palaeontol. 7, 241–298. doi:10.1017/S1477201909002715
Li, P., Hua, H., Zhang, L., Zhang, D., Jin, X., and Liu, Z. (2007). Lower Cambrian phosphatized Punctatus from southern Shaanxi and their ontogeny sequence. Chin. Sci. Bull. 52, 2820–2828. doi:10.1007/s11434-007-0447-y
Li, Z. (1984). The discovery and its significance of small shelly fossils in Hexi area, Xixiang. Geol. Shaanxi 2, 73–77.
Liu, Y., Li, Y., Shao, T., Wang, Y., Yu, B., Han, H., et al. (2005). Two new species of protoconulariids from the early Cambrian in South Shaanxi, China. Acta micropalaeontol. Sin. 22, 311–321. doi:10.1360/gs050302
Liu, Y., Li, Y., Gong, H., Zhang, Z., and Yan, T. (2009). New date on Quadrapyrgites from the early cambrian of South Shaanxi. Acta Palaeontol. Sin. 48, 688–694. doi:10.1360/972009-495
Liu, Y., Li, Y., Shao, T., Zheng, X., and Wang, K. (2011). A new genus and specie of protoconulariids from the early Cambrian in the south Shaanxi, China. Acta micropalaeontol. Sin. 28, 245–249. doi:10.1631/jzus.B1000197
Liu, Y., Li, Y., Shao, T., Zhang, H., Wang, Q., and Qiao, J. (2014a). Quadrapyrgites from the lower Cambrian of South China: Growth pattern, post–embryonic development, and affinity. Chin. Sci. Bull. 59, 4086–4095. doi:10.1007/s11434-014-0481-5
Liu, Y., Xiao, S., Shao, T., Broce, J., and Zhang, H. (2014b). The oldest known priapulid–like scalidophoran animal and its implications for the early evolution of cycloneuralians and ecdysozoans. Evol. Dev. 16, 155–165. doi:10.1111/ede.12076
Liu, Y., Shao, T., Zhang, H., Wang, Q., Zhang, Y., Chen, C., et al. (2017). A new scyphozoan from the cambrian fortunian stage of SouthSouth China. Palaeontology 60, 511–518. doi:10.1111/pala.12306
Liu, Y., Carlisle, E., Zhang, H., Yang, B., Steiner, M., Shao, T., et al. (2022). Saccorhytus is an early ecdysozoan and not the earliest deuterostome. Nature 609, 541–546. doi:10.1038/s41586-022-05107-z
Park, E., Hwang, D., Lee, J., Song, J., Seo, T., and Won, Y. (2012). Estimation of divergence times in cnidarian evolution based on mitochondrial protein-coding genes and the fossil record. Mol. Phylogenetics Evol. 62, 329–345. doi:10.1016/j.ympev.2011.10.008
Peng, S. C., Babcock, L. E., and Ahlberg, P. (2020). “The Cambrian Period,” in Geological Time Scale 2020. Editors F. M. Gradstein, J. G. Ogg, M. Schmitz, and G. M. Ogg (Oxford: Elsevier), 565–629.
Qian, Y., and Bengtson, S. (1989). Palaeontology and biostratigraphy of the early cambrian meishucunian stage in yunnan Province, south China. Fossils Strata 24, 293–294. doi:10.1016/0012-8252(91)90034-D
Qian, Y. (1977). Hyolitha and some problematica from the lower Cambrian Meishucun Stage in central and SW China. Acta Palaeontol. Sin. 16, 255–275. doi:10.19800/j.cnki.aps.1977.02.007
Qin, J., Liu, Y., Wang, Q., Zhang, H., Zhang, Y., Liu, M., et al. (2020). A new fortunian scyphozoa and its development. Acta Geol. sin–engl. 94, 1303–1304. doi:10.1111/1755-6724.13838
Shao, T., Liu, Y., Wang, Q., Zheng, Y., Tang, H., Cao, X., et al. (2015). Early Cambrian gastrozooid of polyp and problematic tubular microfossils from the southern Shaanxi, China. Acta micropalaeontol. Sin. 32, 194–199. doi:10.16087/j.cnki.1000-0674.2015.02.008
Shao, T., Liu, Y., Wang, Q., Zhang, H., Tang, H., and Li, Y. (2016). New material of the oldest known scalidophoran animal Eopriapulites sphinx. Palaeoworld 25, 1–11. doi:10.1016/j.palwor.2015.07.003
Shao, T., Liu, Y., Duan, B., Zhang, H., Zhang, H., Wang, Q., et al. (2018a). The Fortunian (lowermost Cambrian) Qinscyphus necopinus (Cnidaria, Scyphozoa, Coronatae) underwent direct development. Neues Jahrb. Geol. P.–A. 289, 149–159. doi:10.1127/njgpa/2018/0755
Shao, T., Tang, H., Liu, Y., Waloszek, D., Maas, A., and Zhang, H. (2018b). Diversity of cnidarians and cycloneuralians in the fortunian (early cambrian) Kuanchuanpu Formation at Zhangjiagou, south China. Paleontology 92, 115–129. doi:10.1017/jpa.2017.94
Shao, T., Hu, B., Shao, Y., Zhang, Y., Zhang, H., Qin, J., et al. (2019a). Intraspecific variation of radial symmetry number of a 535 million–year–old jellyfish. Precambrian Res. 349, 105412. doi:10.1016/j.precamres.2019.105412
Shao, T., Qin, J., Shao, Y., Liu, Y., and Zhang, H. (2019b). New macrobenthic cycloneuralians from the fortunian (lowermost cambrian) of SouthSouth China. Precambrian Res. 2019, 105413. doi:10.1016/j.precamres.2019.105413
Steiner, M., Li, G. X., Qian, Y., and Zhu, M. (2004a). Lower Cambrian small shelly fossils of northern Sichuan and southern Shaanxi (China), and their biostratigraphic importance. Geobios 37, 259–275. doi:10.1016/j.geobios2003.08.001
Steiner, M., Zhu, M., Li, G., Qian, Y., and Erdtmann, B. D. (2004b). New Early Cambrian bilaterian embryos and larvae from China. Geology 32, 833–836. doi:10.1130/g20567.1
Steiner, M., Li, G. X., Qian, Y., Zhu, M., and Erdtmann, B. D. (2007). Neoproterozoic to early Cambrian small shelly fossil assemblages and a revised biostratigraphic correlation of the Yangtze Platform (China). Palaeogeogr. Palaeoclimatol. Palaeoecol. 254, 67–99. doi:10.1016/j.palaeo.2007.03.046
Steiner, M., Li, G., Hu, S., and Keupp, H. (2010). Soft–tissue preservation in small shelly faunas. Geol. Soc. Am. Annu. Meet. 42, 539.
Steiner, M., Qian, Y., Li, G., Hagadorn, J. W., and Zhu, M. (2014). The developmental cycles of early cambrian Olivooidae fam. Novol. (? Cycloneuralia) from the yangtze platform (China). Palaeogeogr. Palaeoclimatol. Palaeoecol. 398, 97–124. doi:10.1016/j.palaeo.2013.08.016
Van Iten, H., de Moraes Leme, J., Simões, M. G., Marques, A. C., and Collins, A. G. (2006). Reassessment of the phylogenetic position of conulariids (? Ediacaran–Triassic) within the subphylum medusozoa (phylum Cnidaria). J. Syst. Palaeontol. 4, 109–118. doi:10.1017/S1477201905001793
Van Iten, H., Zhu, M., and Li, G. (2010). Redescription of Hexaconularia He and Yang, 1986 (lower Cambrian, South China): Implications for the affinities of conulariid–like small shelly fossils. Palaeontology 53, 191–199. doi:10.1111/j.1475-4983.2009.00925.x
Van Iten, H., Marques, A. C., Leme, J. d. M., Forancelli Pacheco, M. L. A., and Simões, M. G. (2014). Origin and early diversification of the phylum Cnidaria Verrill: Major developments in the analysis of the taxon’s Proterozoic–Cambrian history. Palaeontology 57, 677–690. doi:10.1111/pala.12116
Wang, X., Han, J., Vannier, J., Ou, Q., Yang, X., Uesugi, K., et al. (2017). Anatomy and affinities of a new 535–million–year–old medusozoan from the Kuanchuanpu Formation, South China. Palaeontology 60, 853–867. doi:10.1111/pala.12320
Wang, D., Vannier, J., Schumann, I., Wang, X., Han, S. J., Komiya, T., et al. (2019). Origin of ecdysis: Fossil evidence from 535–million–year–old scalidophoran worms. Proc. R. Soc. B 286, 20190791. doi:10.1098/rspb.2019.0791
Wang, D., Vannier, J., Yang, X., Sun, J., Han, J., Hao, W. j., et al. (2020). Cuticular reticulation replicates the pattern of epidermal cells in lowermost Cambrian scalidophoran worms. Proc. R. Soc. B 287, 20200470. doi:10.1098/rspb.2020.0470
Wang X, X., Kubota, S., Ou, Q., Vannier, J., Han, J., Yao, X., et al. (2020). An intermediate type of medusa from the early Cambrian Kuanchuanpu Formation, South China. Palaeontology 4, 775–789. doi:10.1111/pala.12483
Yasui, K., Reimer, J., D., Liu, Y. H., Yang, X. G., Daisuke, K., Degan, S., et al. (2013). A diploblastic radiate animal at the dawn of cambrian diversification with a simple body plan: Distinct from Cnidaria? PLoS ONE 8, e65890. doi:10.1371/journal.pone.0065890
Keywords: medusozoan, olivooids, apical complement, Zhangjiagou section, developmental sequence
Citation: Zhang Y, Liu Y, Shao T and Qin J (2023) New Qinscyphus material from the Fortunian of South China. Front. Earth Sci. 11:1038686. doi: 10.3389/feart.2023.1038686
Received: 07 September 2022; Accepted: 28 February 2023;
Published: 17 March 2023.
Edited by:
Farid Saleh, Yunnan University, ChinaReviewed by:
Jean Vannier, Université Claude Bernard Lyon 1, FrancePeiyun Cong, Yunnan University, China
Jonathan Antcliffe, Université de Lausanne, Switzerland
Copyright © 2023 Zhang, Liu, Shao and Qin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yunhuan Liu, eXVuaHVhbmxAY2hkLmVkdS5jbg==