 Maria Romero-González
Maria Romero-González Bennett C. Nwaobi
Bennett C. Nwaobi Joseph M. Hufton
Joseph M. Hufton Daniel J. Gilmour3
Daniel J. Gilmour3- 1Department of Geography, Kroto Research Institute, The University of Sheffield, Broad Lane, Sheffield, UK
- 2Department of Life Sciences, Imperial College London, South Kensington Campus, London, UK
- 3Department of Molecular Biology and Biotechnology, University of Sheffield, Western Bank, Sheffield, UK
The ex-situ bioremoval of U(VI) from contaminated water using Acidithiobacillus ferrooxidans strain 8455 and 13538 was studied under a range of pH and uranium concentrations. The effect of pH on the growth of bacteria was evaluated across the range 1.5–4.5 pH units. The respiration rate of At. ferrooxidans at different U(VI) concentrations was quantified as a measure of the rate of metabolic activity over time using an oxygen electrode. The biosorption process was quantified using a uranyl nitrate solution, U-spiked growth media, and U-contaminated mine water. The results showed that both strains of At. ferrooxidans are able to remove U(VI) from solution at pH 2.5–4.5, exhibiting a buffering capacity at pH 3.5. The respiration rate of the micro-organism was affected at U(VI) concentration of 30 mg L−1. The kinetics of the sorption fitted a pseudo-first order equation, and depended on the concentration of U(VI). The KD obtained from the biosorption experiments indicated that strain 8455 is more efficient for the removal of U(VI). A bioreactor designed to treat a solution of 100 mg U(VI) L−1 removed at least 50% of the U(VI) in water. The study demonstrated that At. ferrooxidans can be used for the ex-situ bioremediation of U(VI) contaminated mine water.
Introduction
An increase in anthropogenic activity through uranium mining and milling for the nuclear fuel cycle, and other industrial processes, has resulted in elevated levels of uranium within the environment. Low levels of uranium are released during uranium mining and high levels are released during uranium enrichment and reprocessing (Lloyd and Renshaw, 2005). Leaching of these waste materials into nearby ground water and soil sediments, due to ineffective storage mechanisms, results in radionuclides entering the food and water supply, ultimately becoming a hazard to human health (Choy et al., 2006; Gavrilescu et al., 2009). Therefore, safe removal and detoxification of these contaminants is of utmost importance, with current bioremediation technologies showing promise.
Contamination by uranium from mining activities is extensive in surface and ground waters across the world, and the consequences of this are not only detrimental to the health of the natural environment, since they also have an impact on human health. The UNESCO estimates that groundwater provides about 50% of the global drinking water supply (WWAP, 2009). The contamination of groundwater from uranium mining activities is limiting the access to clean water in former mining communities in the US. For example, a 5-year study conducted by the US EPA in abandoned mining areas within the 27,000 square miles of the Navajo Nation showed that about 57,000 people access water from unregulated sources, and some of those sources are contaminated with uranium. From the 240 unregulated sources tested, 29 exceeded the drinking water standard for uranium (USA-EPA, 2014). In the south-west of the Punjab state of India, the levels of uranium in groundwater have been reported to be as high as 100 ppb (Alrakabi et al., 2012), potentially affecting 10 million people. A study of 11,000 public supply wells in California showed that at least 2 million people are exposed to water contaminated with uranium, amongst other trace elements (Belitz et al., 2015). These examples of ground water contamination demonstrate the urgent need to develop water-cleaning technologies for the removal of uranium.
Biologically based remediation techniques, such as bioremediation, provide a cheap and efficient alternative to current remediation technologies. These processes utilize the enzymatic reductive power of microorganisms to produce U(IV) minerals and the biosorbent, and enzymatic precipitation capabilities of bacteria to sequester U(VI) (Merroun and Selenska-Pobell, 2008; Neu et al., 2010; Newsome et al., 2014; Choudhary and Sar, 2015). The enzymatic redox transformations of U(VI) and U(IV) have been studied comprehensively (Wall and Krumholz, 2006). Numerous strains of bacteria are also capable of transforming U(VI) using a similar mechanism (Lloyd et al., 2003; Khijniak et al., 2005; Bernier-Latmani et al., 2010; Orellana et al., 2013). However, the poor stability of these reduced products within aerobic environments results in the reoxidation of U(IV) minerals, hindering the development of large scale bioremediation technologies using this technique (Wall and Krumholz, 2006; Merroun and Selenska-Pobell, 2008). Reoxidation of U(IV) minerals has been observed in the presence of oxygen (Abdelouas et al., 1999; Duff et al., 1999), nitrates/ites (Finneran et al., 2002; Senko et al., 2002, 2005b), carbonates (Choudhary and Sar, 2015), Fe(III) hydroxides (Nevin and Lovley, 2000; Senko et al., 2005a), organic ligands (Luo and Gu, 2011; Singh et al., 2014), and microbial oxidation activity (Dispirito and Tuovinen, 1982; Newsome et al., 2015). Therefore, using indigenous microorganisms' abilities to uptake and precipitate U(VI) in the presence of oxygen may be more efficient in the removal of U(VI) from surface contaminated environments.
Biosorption is a metabolism-independent biological process by which metal species, compounds, and particulate materials can be removed from solution through complexation with biomass (Gadd, 1993). The removal of U(VI) via bacterial biosorption processes has been described as metabolism-independent. Numerous bacterial strains from a diverse origin have displayed binding affinities for the radionuclide species. The process is affected by pH, ionic strength, sorbent and sorbate concentrations, cell physiological state, and the presence of other soluble polymers within solution (Dobson and Burgess, 2007; Merroun and Selenska-Pobell, 2008; Wang and Chen, 2009). Structural differences in cell wall architecture between gram-positive and gram-negative bacteria suggest differences in uranium biosorption to the cell surface. The gram-positive genus Bacillus has been studied extensively for its biosorption capabilities in sequestering uranium from aqueous environments. It is suggested that at very low pH, covalent bonding alongside electrostatic interactions occurs between uranium and the cell surface due to protonation of the bioavailable functional groups (Selenska-Pobell et al., 1999; Fowle et al., 2000; Kelly et al., 2002; Panak et al., 2002; Gorman-Lewis et al., 2005). As the pH increases and deprotonation of the functional groups occurs; electrostatic interactions become more favorable due to the positively charged uranyl species and negatively charged surface. This results in uranyl complexation with neutral phosphate and deprotonated carboxyl functional groups on the bacterial cell wall (Fowle et al., 2000). The composition of gram-positive cell wall suggests that these preferential surface functional groups originate from the peptidoglycan (carboxyl) and secondary teichoic and lipoteichoic acids (phosphate).
The biosorption of uranium to gram-negative bacteria has been demonstrated in a variety of strains (Haas et al., 2001; Haas and Northup, 2004; Lütke et al., 2012). Shewanella putrificiens was shown to biosorb uranium through forming surface complexes with carboxyl and protonated phosphate groups, with sorption being dependent on pH and ionic strength (Haas et al., 2001). A similar U(VI) biosorption mechanism has been described for other Gram negative strains including Idiomarina loihiensis MAH1 (Morcillo et al., 2014), Pseudomonas fluorescens (Lütke et al., 2012), Cupriavidus metallidurans CH34, and Rhodopseudomonas palustris (Llorens et al., 2012). Alongside the phosphate groups within the outer membrane phospholipids, lipopolysaccharides (LPS) have been suggested to be involved in uranyl binding at the gram-negative cell surface (Ferris and Beveridge, 1986; Langley and Beveridge, 1999; Barkleit et al., 2008, 2011). The experiments suggest a binding mechanism of uranyl to phosphate moieties rather than carboxyl groups. This process is governed by the concentration of LPS instead of solution pH. Based on these findings, the treatment of effluents with low to medium concentrations of U(VI) waste using bacterial biosorption may be more favorable due to the attachment of metal cations to the surface (Newsome et al., 2014).
The removal of U(VI) has been demonstrated in situ by underground field studies at the DOE Oak Ridge Field Research Center. Microorganisms indigenous to the site were stimulated using glycerol-3-phosphate. The precipitation of uranium was achieved using small flow-through columns under aerobic conditions and circumneutral pH-values. The precipitates formed were identified as uranium phosphate minerals using X-ray Absorption Spectroscopy (XAS) (Beazley et al., 2011). Studies conducted under similar conditions using glycerol-2-phosphate amended sediments also showed the precipitation of U(VI) phosphate bearing minerals (Salome et al., 2013). Strains isolated from same site displayed varied phosphatase capabilities. The exposure of some of these strains to uranium significantly decreased their culturability, indicating the toxic effects of uranium to bacterial growth (Beazley et al., 2007; Martinez et al., 2007). Further studies from this site suggested that, in the presence of high calcium concentrations, other bacterial strains can hydrolyze inorganic phosphate from glycerol-3-phosphate to form uranium hydroxyapatite precipitates (Shelobolina et al., 2009).
We present here the use of an extremophile microorganism, Acidithiobacillus ferrooxidans, for the ex-situ bioremediation of U(VI) contaminated acidic mine water. Our hypothesis is that the ability of microorganisms to remove U(VI) from solution can be harnessed ex-situ using a bioreactor. The main possible mechanisms of removal proposed are biosorption and biomineralization due to the presence of bacterial biomass.
Materials and Methods
Microorganisms and Growth Conditions
Two bacterial strains of At. ferrooxidans, type strain NCIMB 8455 (ATCC 23270) and strain NCIMB 13538, were obtained from the National Collection of Industrial, Food, and Marine Bacteria (NCIMB), Aberdeen Scotland. At. ferrooxidans was originally isolated from an acid, bituminous coal mine effluent in the United States of America and has been extensively studied (Leduc and Ferroni, 1994), (Amouric et al., 2011). The NCIMB 13538 strain was isolated from an abandoned uranium mine in Dera Ghazi Khan, Pakistan by the National Institute for Biotechnology and Genetic Engineering (Faisalabad, Pakistan). No published reports confirming the identity of the bacterium are available. The prokaryote identity of this species was confirmed using 16S rRNA gene analysis, and the results can be found on Supplementary Table 1. The bacterial strains were cultured using Acidiphilium complex medium, modified by the omission of carbon and peptide sources, and the addition of 10 mM Fe (DSMZ, 2015). This medium consists of 2 mM MgSO4 7H2O, 0.8 mM (NH4)2SO4, 0.7 mM KCl, 0.06 mM Ca(NO3)2 4H2O, 0.4 mM KH2PO4, 0.3 mM NaCl, 0.6 mM K2HPO4, 10 mM FeSO4 7H2O, 1.0 g L−1 mannitol, and 1.7 g L−1 bactotryptone. The effect of temperature on the bacterial growth experiments for both strains can be found in Supplementary Figures 1, 2. The optimum temperature obtained for the growth was 30°C for both strains and therefore all experiments were conducted at this temperature. A Scanning Electron Microscopy (SEM) micrograph showed that the cells are rod-shaped (Supplementary Figure 3).
Effect of pH on the Growth of Acidithiobacillus ferrooxidans
The effect of variation of initial solution pH on the growth of At. ferrooxidans was evaluated by adjusting the starting pH of the modified acidiphilium medium to initial pH-values of 1.5, 2.5, 3.5, and 4.5. The bacteria were then incubated at 30°C for 72 h with agitation. The optical density was measured periodically at 600 nm. The value of pH was monitored constantly for the duration of the experiment using a pH meter (Orion, Thermo Scientific).
Effect of U(VI) Concentration on the Metabolism of Acidithiobacillus ferrooxidans
The effect of U(VI) concentration on the metabolic activity of aerobically grown At. ferrooxidans strains was measured by quantifying respiration rate. An oxygen electrode (Hansatech, Ltd.) calibrated using sodium dithionite (Sigma-Aldrich) was used. A stock solution of U(VI) was prepared using uranyl nitrate hexahydrate, UO2(NO3)2 6H2O (Sigma-Aldrich). Samples of 2 ml containing bacteria culture, incubated with a range of U(VI) concentrations ranging from 1 to 100 mg U(VI) L−1, were analyzed for 5 min. The electrode calibration was matched for the composition of the modified acidiphilium medium by including a non-inoculated medium control. Six replicates of controls and samples were run in alternating pairs. In order to compare the results obtained between strains, cells were diluted to a concentration of 108 cells mL−1.
Kinetics of Biosorption of U(VI) Using Acidithiobacillus ferrooxidans
The bacteria were grown to exponential phase using the modified acidiphilium growth medium. Cells were harvested by centrifugation at 6000 rpm for 20 min to separate the biomass from Fe(III) precipitates and hydroxides produced in the medium during growth. Afterwards, biomass pellets were washed three times with 0.05 M sulfuric acid and then two times with 0.01 M perchloric acid. A sample of 0.015 g wet mass (0.0038 g dry weight approximately) of bacteria was suspended in 10 ml sterile de-ionized water containing 1.08 × 10−3 mmol L−1 U(VI) and incubated at 25°C with agitation. Samples were collected at 1, 2, 4, 6, 12, and 24 h and analyzed using ICP-MS (ELAN 6000, Perkin Elmer). All experiments were performed in triplicate and controls containing uranium without bacteria were also included.
Biosorption of U(VI) Using Acidithiobacillus ferrooxidans
Batch experiments were conducted to quantify the biosorption of U(VI) to At. ferrooxidans using simulated mine water solution at different concentrations of uranium. The bacteria samples were prepared using the same procedure as in the kinetic experiment. Samples of 0.015 g wet mass (0.0038 g dry weight approximately) of bacteria were suspended in 10 ml simulated mine water spiked with U(VI) to a solution concentration of 10, 50, 80, 100, 120, 150, and 200 mg U(VI) L−1. Controls solutions of U(VI) with no bacteria inoculate, and bacteria with no U(VI) added, were also included. Triplicate samples were incubated at 25°C in an orbital shaker for 24 h. The supernatant was separated from the biomass pellet by centrifugation at 3000 rpm, and analyzed for its U(VI) content using ICP-MS. U(VI) biosorption was also quantified using a simulated mine water solution containing 120 mg U(VI) L−1 and a control of acidiphilium growth medium spiked with 120 mg U(VI) L−1. Comparison of mean values was performed using SPSS statistical software (IBM-SPSS, 2011).
Semi-Continuous Bioreactor
A semi-continuous 5 L bioreactor containing growth media and a culture of At. ferrooxidans was designed. The reactor was submerged in a water bath at 30°C to ensure the micro-organism growth. The reactor was kept under continuous agitation using a magnetic stirrer and oxygen was supplied at a rate of 25 mL min−1 using a pump connected and tubing connected to a porous stone to aid the oxygen diffusion through solution. The pH and temperature of the solution were monitored using electrode probes and samples were collected via a sampling port. The reactor was left to equilibrate for 24 h before adding an aliquot of U(VI) solution to a final volume of 100 mg U(VI) L−1. The reactor was operated for 72 h and the concentration of U(VI) was quantified using ICP-MS. Figure 1 shows a sketch of the reactor depicting its components.
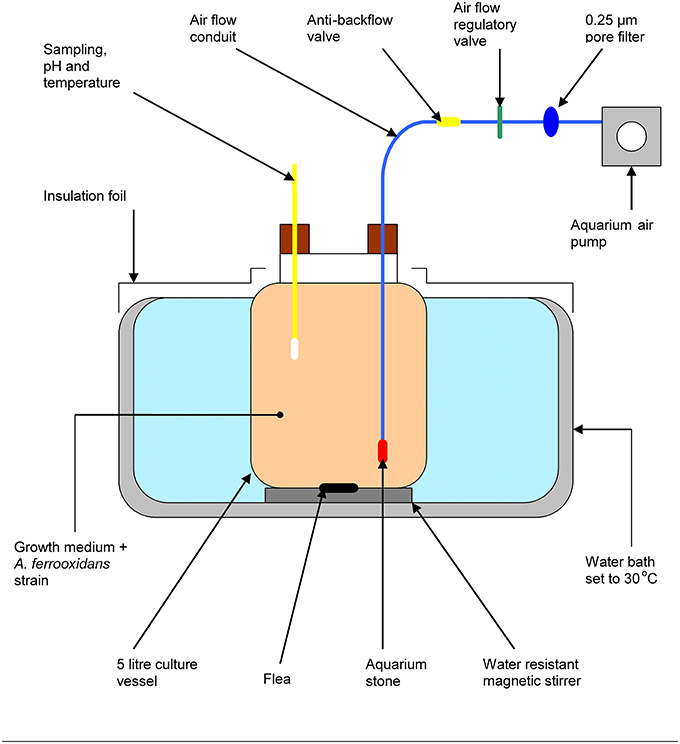
Figure 1. Design sketch of the bioreactor showing its components. The total volume of the vessel was 5 L of a mine water solution containing 100 mg U(VI) L−1. Temperature and pH probes were inserted to monitor the reaction during time. Oxygen was constantly supplied via a aeration pump. The temperature was kept to a constant value of 30°C using a water bath.
Results
Effect of Solution pH
The effect of pH on the growth of At. ferrooxidans was quantified at different initial growth medium pH-values. Figure 2A shows the growth curves obtained at pH 1.5, 2.5, 3.5, and 4.5 for strain 13538 at 30°C. The growth curves obtained for pH 2.5, 3.5, and 4.5 follow a typical exponential curve, with a plateau at OD600-value 1.2. Bacteria did not grow at pH 1.5, indicating that the optimum pH-value is between 2.5 and 4.5 pH units. Figure 2B shows the profile for the pH obtained during the growth experiments. There was no variation in the pH-values during the experiment conducted at pH 1.5. Despite the initial solution pH-values being adjusted to 2.5, 3.5, and 4.5 pH units for the rest of the experiments, the pH measurements returned values of 2.5 pH units after 20 h. The pH remained stable at this value until the end of the experiment at 72 h.
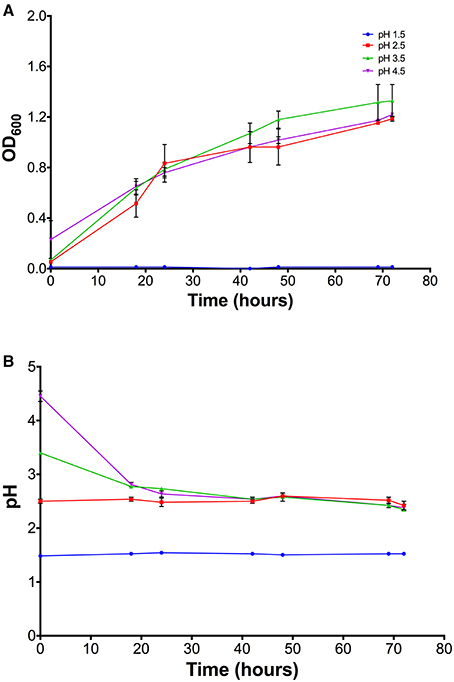
Figure 2. (A) Growth curves of At. ferrooxidans strain 13538 at different pH-values. The optical density at 600 nm was measured over a period of 72 h. Optimal bacteria growth is achieved at pH 2–4.5. (B) Profile of solution pH during growth experiments. Error bars represent 1 standard deviation of the mean.
Effect of U(VI) Concentration on the Respiration Rate of Acidithiobacillus ferrooxidans
The respiration rate of At. ferrooxidans at different U(VI) concentrations was quantified as a measure of the rate of metabolic activity over time by monitoring oxygen consumption. Figure 3A shows the amount of oxygen consumed by At. ferrooxidans strain 8455 as a function of increasing concentration of U(VI) in solution. Control experiments performed without U(VI) were used for baseline measurements of the respiration rate and a mean value of 0.9 nmoles [106 cells]−1 h−1 was calculated for all experiments. Four different uranium concentrations were used to test metabolic oxygen utilization in strain 8455, ranging from 1 to 30 mg L−1. At 1 mg L−1 uranium concentration, strain 8455 averaged 1.29 nmoles [106 cells]−1 h−1, a value higher than the 0.98 nmoles [106 cells]−1h−1 obtained from corresponding controls. Paired t-tests showed that there was a statistically significant difference between the means at a confidence level >99%. Six replicate experiments using strain 8455 in contact with 10 mg U(VI) L−1 uranium solution were performed over a 2 day period to quantify errors due to differences in experimental conditions. Two controls, plus replicate experiments were run on the first day and four on the second, with consistent results. This helped negate experimental anomaly and complemented the statistical tests by confirming that perceived enhanced metabolic performance of strain 8455 at 10 mg L−1 uranium was significant and may be due to a stress response that increases respiration rate in an attempt to detoxify the uranium. At 30 mg L−1 uranium, the bacteria consumed oxygen at rates of 0.86 and 0.87 nmoles [106 cells]−1h−1 for controls, respectively. According to SPSS paired t-test analysis, there was no statistically significant difference between the means. Strain 8455 was hence metabolically uninhibited in terms of oxygen consumption by 30 mg L−1 uranium. Tests at 40 mg L−1 proved different. Average bacterial oxygen consumption was 0.4 nmoles [106 cells]−1h−1, lower than the corresponding control value of 0.87 nmoles [106 cells]−1h−1. Paired t-tests confirmed a statistical difference between the means at a significance level >99%. A uranium concentration of 30 mg L−1 was hence sufficient to inhibit metabolic consumption in strain 8455.
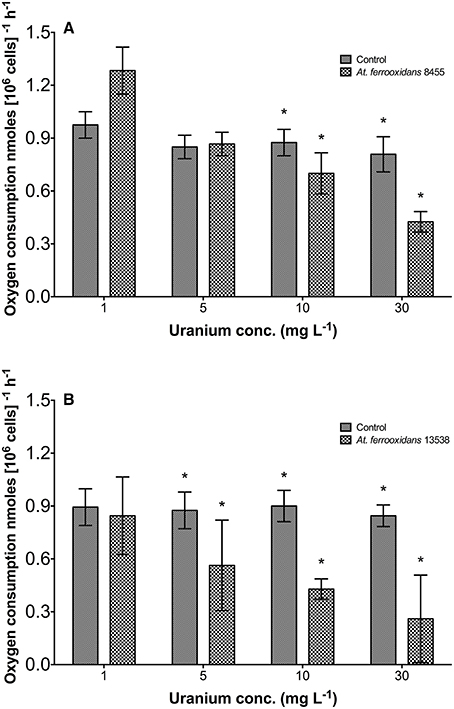
Figure 3. Oxygen consumption rate by At. ferrooxidans 8455 (A) and 13538 (B) as a function of increasing concentration of U(VI) in solution. The respiration rate is inhibited by the increasing concentration of U(VI) in solution up to 30 mg (VI) L−1. Data labeled with * indicate significantly different results.
It was found that strain 13538 was more susceptible to uranium than strain 8455 (Figure 3B). At 1 mg L−1 uranium, strain 13538 recorded an oxygen consumption of 0.85 nmoles [106 cells]−1 h−1 as against 0.89 nmoles [106 cells]−1 h−1 registered by the corresponding controls. Paired t-test analysis using SPSS software showed there was no statistically significant difference between the means. Strain 13538 was metabolically uninhibited by a uranium concentration of 1 mg L−1. Tests conducted at 5 mg L−1 uranium showed strain 13538 consumed oxygen at a rate of 0.56 nmoles [106 cells]−1h−1, lower than the 0.88 nmoles [106 cells]−1 h−1 value recorded by the corresponding controls. The paired t-tests comparing the means showed a statistical difference at the 90% confidence level. As uranium concentrations of 5 mg L−1 were clearly inhibitory to oxygen metabolic activity in strain 13538, it was expected that the results obtained at the higher metal concentrations of 10 and 30 mg L−1 would follow a similar trend, as observed in Figure 3B.
Kinetics of the Biosorption Process
Results from the kinetic experiment performed using batch type reactors with a solution of 1.08 × 10−3 mmol U(VI) L−1 are shown in Figure 4. A reduction of the initial concentration of U(VI) over time was observed, with a typical exponential shape in the curve for the experiments using both At. ferrooxidans strains. Strain 13538 is slightly more efficient in reducing the concentration of U(VI) in solution than strain 8455. The obtained data was fitted to a pseudo-first order model (Supplementary Figure 4). Fitting the data to zero order reactions was not possible since this model is not dependent on the concentrations of the reactants. Assuming that the rate of reaction depends on one reactant, in this case, the concentration of U(VI), a pseudo-first order equation was used to fit the obtained data. The observed decrease of uranium concentration in solution by strain 8455 was from 1.08 × 10−3 to 0.95 × 10−3 mmol L−1 after 24 h, which equates to a total decrease of 0.13 × 10−3 mmol L−1 or 11.9%. Of this amount, nearly half (5.2%) was taken up by the cells after only 4 h. The amount of U(VI) from solution, when in contact with strain 13538, was determined to be from 1.08 × 10−3 to 0.84 × 10−3 mmol L−1 after 24 h, resulting in a decrease of 0.25 × 10−3 mmol L−1 or 22.6%. A 16.5% reduction was observed after 12 h (half of the experiment's duration). The rate of reaction, r, was defined as (Equation 1).
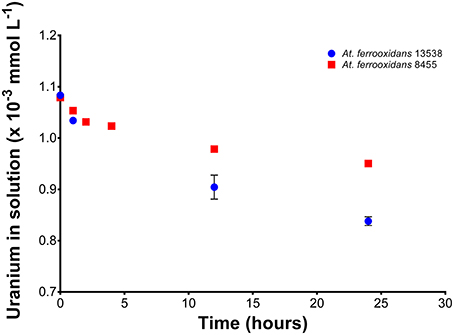
Figure 4. Kinetics of the biosorption removal of U(VI) by At. ferrooxidans strains 8455 and 13538. The data fits a pseudo-first order equation. Bacteria strains are more efficient removing U(VI) from simulated mine water solution compared to growth media or uranyl salt solution.
Bioremediation of U(VI) from Simulated Mine Water
The effect of U(VI) concentration on the growth of At. ferrooxidans was evaluated using a solution of 120 mg U(VI) L−1. Figure 5 shows changes in the concentration of uranium in solutions of U(VI), growth media, and simulated mine water. The concentration of uranium in the uranium salt solution after being in contact with At. ferrooxidans was 57.35 mg L−1 for strain 8455 and 69.50 mg L−1 for strain 13538. The removal of uranium from solution was calculated at 52.6 and 42.6% for strains 8455 and 13538, respectively. This indicated that both strains were efficient in the reduction of uranium concentration from a uranyl salt solution. The efficiency of At. ferrooxidans in removing uranium from a solution of modified acidiphilium media was also evaluated. The mean concentrations of uranium in solution were 106.06 and 105.77 mg L−1 for strain 8455 and strain 13538, respectively. These values represent a removal of uranium from growth media solution of only 12% for strain 8455 and 13% for strain 13538 when in the presence of nutrients. The removal of uranium from a simulated mine water solution by strain 8455 was 48.81 mg L−1 and 50.15 mg L−1 for strain 13538. These represent an efficiency of removal of 59.7 and 58.5%, respectively. T-test results showed that the removal of uranium from the three solutions by each one of the strains was significantly different at 99% confidence. The statistical comparison also showed that strain 8455 was more efficient compared to strain 13538 in removing uranium from the uranyl salt solution. However, no significant differences were found when comparing the efficiency of both strains in removing uranium from the growth media or simulated mine water solution.
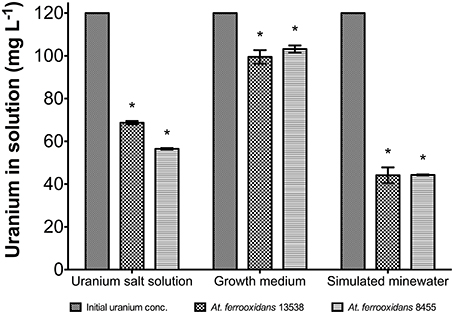
Figure 5. Amount of uranium removed from uranyl nitrate, growth media spiked with U(VI) and simulated mine water spiked with U(VI) solutions by At. ferrooxidans strains 8455 and 13538. A higher concentration of U(VI) was removed from simulated mine water solution. Data labeled with * indicate significant different results.
The uptake of U(VI) by At. ferrooxidans from simulated mine water solution was quantified using batch sorption experiments. Figure 6 shows the sorption isotherm obtained for the removal of uranium from simulated mine water solution at pH 3.5 for strain 8455 and strain 13538. The sorption isotherms exhibited a typical curve shape with a plateau, indicating that the sorption capacity of uranium by At. ferrooxidans is limited. The maximum sorption obtained was 53% for strain 8455 and 43% for strain 13538. The isotherm data was fitted to a Langmuir equation as follows:
where q represents a relationship between the initial and final concentration of U(VI) in solution as a function of the biomass in a given volume:
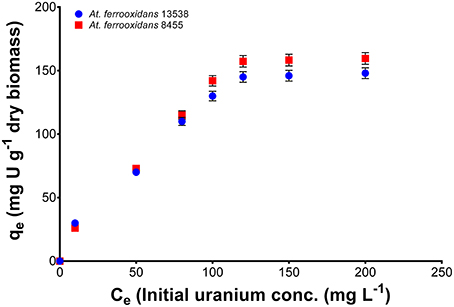
Figure 6. Sorption isotherms for At. ferrooxidans strains 8455 and 13538 obtained at pH 3.5. The data fitted a Langmuir equation. Error bars correspond to 1 standard deviation of the mean.
A plot of the inverse of q vs. C produces a straight line (Supplementary Figure 5) with a slope of and an intercept of , where qm is the maximum sorption capacity and KL is the Langmuir constant. The sorption parameters obtained for strain 8455 were KL = 0.013 and qm = 227.27 mg g dry biomass−1. For strain 13538 the calculated sorption parameters were KL = 0.019 and qm = 178.57 mg g dry biomass−1. The dissociation constant, KD was calculated by:
Using the Langmuir isotherm model, the concentration in the biomass sorbent is quantified using:
Assuming a concentration in solution, Cw of 100 mg L−1, the estimated KD-values were 1.3 L g −1for strain 8455 and 1.2 L g −1 for strain 13538. The KD-value obtained suggests that both strains possess similar ability to remove uranium.
The bioremoval of U(VI) using At. ferrooxidans was also quantified using a semi-continuous reactor. The reactor containing growth media and bacteria biomass was left to equilibrate for 24 h to half of the exponential growth phase, ensuring the availability of bacterial cells in the reactor. A spike of U(VI) solution was added to make a final concentration of 100 mg U(VI) L−1 in the reactor. The pH throughout the bioremoval process was constant, at a value of 3.5, and no pH amendments were required during the reaction time. The oxygen concentration in solution was also found to be constant at 8.1 mg O2 L−1. The concentration of U(VI) was reduced from 100 mg U(VI) L−1 to 51.2 mg U(VI) L−1 in the first 24 h. The amounts of U(VI) quantified at 48 and 72 h were 22.7 mg L−1 and 15.4 mg L−1, respectively. The removal efficiency was then calculated to be 51% at 24 h reducing to 32% at 72 h.
Discussion
Effect of Solution pH and Oxygen on the Activity of Acidithiobacillus ferrooxidans
Acidithiobacillus ferroxidans has great potential for use in biotechnology processes as it grows well under acidic conditions typical of acid mine drainage and tolerates high concentrations of metals such as Cu(II) (Dew et al., 1999) and U(VI) (Merroun and Selenska-Pobell, 2001). These characteristics make this organism an ideal candidate for use in bioleaching or bioremediation technology. Studies conducted with solution pH in the range 1.5–4.5 show that At. ferrooxidans growth is best at pH 3.5 and hindered at pH 1.5. The optimum range of pH obtained (2.5–4.5) is consistent with previously reported growth conditions for At. ferrooxidans (Merroun and Selenska-Pobell, 2001; Mazuelos et al., 2010). Therefore, any successful biotechnology process using this organism requires an influent pH-value within this range. However, experiments conducted using At. ferrooxidans showed a buffering capacity around pH 2.5 (Figure 2B), indicating that the system readily equilibrates. The stabilization of the pH profile across the experiment is comparable to that obtained during an iron bio-oxidation experiment using At. ferrooxidans (Mazuelos et al., 2010) where pH fluctuated around a value of 2.3 under constant oxygen conditions. The observed pH profile supports the hypothesis that pH stabilization in solution is due to limitation of oxygen in solution and the formation of Fe3+ via oxidation, as follows:
This process is limited by oxygen and hence indicates that the abiotic oxidation of Fe2+ to Fe3+ in solution contributes to maintenance of the buffering conditions that favor the healthy activity of the At. ferrooxidans culture. Since pH plays such an important role in the survival of At. ferrooxidans, any potential reactor needs to be conceptualized using pH as a master variable.
The metabolic activity of At. ferrooxidans strains 8455 and 13538 was also measured as a function of U(VI) concentration in order to evaluate the limits of resistance of the bacterial cells to the presence of U(VI). The inhibition of oxygen uptake by aerobic microorganisms may be considered a suitable method to evaluate the toxicity of chemicals. There is a marked difference in the behavior observed for strain 8455 compared to 13538 at 30 mg L−1. Strain 13538 is more affected by the presence of U(VI) in solution, exhibiting a reduction in its respiration rate of ~70% compared to only 50% reduction for strain 8455. This behavior is comparable with results obtained for three A. ferrooxidans eco-types that are capable of removing 25 mg U(VI) L−1 from solution (Merroun and Selenska-Pobell, 2001). Tolerance to concentration of U(VI) varies between strains. A study of a large number of A. ferrooxidans isolates found that the 16 rRNA genes have specific signatures distinguishing strains within the species (Selenska-Pobell, 2002), and these different types exhibited differences in their ability to accumulate uranium (Merroun and Selenska-Pobell, 2001). These findings have been reported for other types of micro-organisms, and lead to the conclusion that accumulation of U(VI) is type specific.
In addressing practical considerations of a bioreactor technology, strain 8455 adapted best to the conditions of the study, and so it may be more suitable for use in bioremediation. Designing a reactor will need to consider the limitations imposed by the pH of the solution, the content of oxygen supplied to the reactor and the concentration of U(VI) in solution, although this is not as critical as the first two parameters.
Bioremoval Process of U(VI) by Acidithiobacillus ferrooxidans
Immobilization of U(VI) by bacterial biomass has been reported for a wide range of micro-organisms including natural isolates from U-contaminated sites such as At. ferrooxidans, D. radiodurans, Bacillus sp., Arthrobacter sp., and P. fluorescens (Merroun and Selenska-Pobell, 2001; Bencheikh-Latmani and Leckie, 2003; Suzuki and Banfield, 2004). Similar behavior was observed for At. ferrooxidans strains 8455 and 13538, where U(VI) was efficiently removed from solution despite its nutrient composition. Interestingly, both strains seem to remove U(VI) from solution more efficiently when in the presence of other ions typical of mine waters. This result highlights the importance of the presence of Fe2+ and the role as electron donor that this ion plays in the metabolism of At. ferrooxidans. Also interesting was that the initial supply of nutrients provided did not result in enhanced removal of U(VI) from solution (Figure 5). This has an impact on the cost associated with culture maintenance. At. ferrooxidans seems to adapt to the nutrient and electrolyte conditions supplied by mine water solution and efficiently remove U(VI) without any added growth medium components in small reactor systems. Compared to other bacteria species strains 8455 and 13538 demonstrated a good removal performance. For example, Bacillus thuringiensis exhibited a superior efficiency of U(VI) removal of to 420 mg g−1 dry biomass, compared to 227 and 179 mg g−1 dry biomass for strains 8455 and 13538, respectively. However, the efficiency of B. thuringiensis was quantified using a uranyl acetate solution, where acetate contributed to the maintenance of the bacterial culture (Table 1). The ability of strain 8455 for the removal of U(VI) is comparable to Microbacterium and Pseudomonas aeruginosa and exceeds the efficiency of Stenotrophomonas sp. and Acinetobacter sp. (Table 1). The behavior of strain 13538 is more similar to the one reported for Bacillus pumilus at pH 4. Most of the reported studies were conducted using U(VI) solutions only in contrast to the mine water solution used in this study. The obtained efficiency confirms that At. ferrooxidans is a good candidate organism to be used in bioremediation systems.
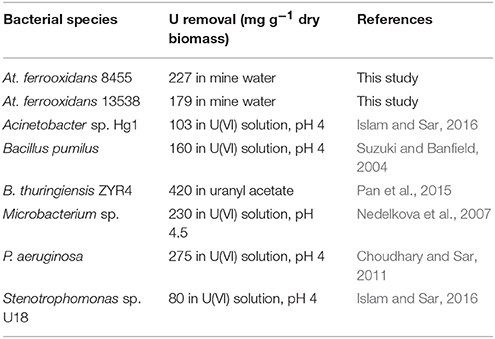
Table 1. Efficiency of different bacteria species biomass (mg g−1 dry biomass) for the removal of U(VI) compared to At. ferrooxidans strains.
The biosorption process followed a typical Langmuir isotherm shape and the distribution constants (KD) derived from the Langmuir model fitting indicated that At. ferrooxidans strain 8455 is more efficient in the removal and sorption of U(VI) than strain 1358. These results are consistent with previously observed behavior of At. ferrooxidans in mine water solution. For example, the difference in efficiency of U(VI) removal from solution between the different strains has been previously reported (Merroun and Selenska-Pobell, 2001), where binding and complexation of U(VI) was different between three At. ferrooxidans eco-types. Although, we conducted genetic characterization of the strains reported here, a comparison of the strains to determine if they belong to different eco-type groups was not conducted and hence comparison to previous studies is only speculative. The biosorption process for strains 8455 and 1358 is described as a monolayer sorption process, where ions in solution bind to the surface of At. ferrooxidans at specific binding sites. These binding sites are energetically similar and possess similar chemical characteristics. Since this study was focused on quantifying the efficiency of removal for a bioprocessing technology, a full characterization of the chemical nature of binding was not conducted. However, the binding and removal of ions by bacterial biomass extensively reported in the literature indicates that the main bacteria binding sites have been identified as phosphate containing sites. Merroun and Selenska-Pobell (2001) described the complexes formed by three eco-types of At. ferrooxidans studied by fluorescence and infrared spectroscopy as uranyl organic-phosphate compounds. Other bacterial strains have shown the ability to precipitate U(VI) from solution as a phosphate type of compound, which suggests that the mechanism of sorption and precipitation is not unique to the bacteria type but to the chemical nature of bacterial cell wall components. Beazley et al. (2007) reported the formation of organophosphate compounds by three heterotrophic bacteria isolated from soils when in the presence of U(VI). The compounds were identified and characterized using EXAFS and reported as organophosphate of the autinite/met-autinite group mineral. Choudhary and Sar (2011) observed a similar organophosphate compound formed by the interaction of P. aeruginosa. The precipitates formed on the cell envelope had a needle shape and were identified as uranium organophosphate complexes by X-ray diffraction analysis and infrared spectroscopy. Suzuki and Banfield (2004) reported the formation of uranyl organophosphate complexes by D. radiodurans and Arthrobacter spp. as observed from TEM and EDX analysis. More recently, studies conducted by the authors on Idiomarina loihiensis (Morcillo et al., 2014) showed that this bacterium forms uranyl organophosphate compounds when in the presence of U(VI) at higher salinity concentrations. These findings indicate that the preferred sorption mechanism of U(VI) by bacterial biomass is via complexation and precipitation on phosphate rich binding sites. The evidence hence suggests that At. ferroxidans would remove U(VI) as uranyl organophosphate as a preferred sorption and removal mechanism.
Implications for Ex-situ Mine Water Treatment—toward the Design of Bioreactors
Very few technologies have been proposed for the removal of uranium from water using treatment plant technologies and most of the literature focuses on description of in situ remediation of uranium mill tailings sites. Therefore, there is an opportunity for advances by coupling removal of uranium with high-density sludge systems (HDS; Campbell et al., 2015). A small scale ion exchange system was developed by the US EPA as early as 1989 (Jelinek and Sorg, 1988). The removal of uranium was achieved using anion exchange resins based on chloride ion and a water softener tank. Uranium was reduced form 23.8 mg L−1 to 1 μg L−1. The treatment was efficient in the removal of uranium, but the re-use of the ion exchange cartridges was limited and additional uranium waste was generated.
Harnessing the power of At. ferroooxidans and the formation of Fe-containing sludge may provide a feasible biotechnology for the removal of uranium from contaminated waters. Using the partition parameters obtained from batch experiments, a bioreactor was designed to test the limitations of the technology. The bioreactor used in this study contained At. ferrooxidans culture, spiked with 100 mg L−1 of U(VI), under constant oxygen supply to ensure the survival of the micro-organism. Care had to be taken to ensure that the aeration process did not oxidize Fe2+ at a faster rate than At. ferrooxidans, otherwise the depletion of Fe2+ would hinder the metabolism of the micro-organism. Therefore, an oxygen supply of 25 ml/min was used. The reactor was operated for 72 h and samples of the solution were analyzed for their U(VI) content. The efficiency of U(VI) removal obtained was of 51% at 24 h for strain 8455. This efficiency was reduced to 32% at 72 h of reaction. A reactor tested using strain 13538 did not operate successfully. The reduction in the efficiency of removal of U(VI) from solution indicates that there is a decay in the ability of the micro-organisms to perform the biosorption process, possibly due to the consumption of nutrient resources that sustain the microbial biomass over time. A large amount of Fe-containing sludge was formed at the bottom of the reactor, limiting the diffusion of oxygen through solution and impeding the bacteria respiration process. Therefore, removal of the sludge during the water cleaning process may be required to maximize the reactor performance. In mine water treatment, HDS are a common feature of treatment plants. Sludge formed from precipitation using lime is separated using a clarifier and recycled into a tank until density is increased and the volume is reduced by 90%. The resulting sludge is of high density producing a high quality of effluent water. The HDS formed is easier to remove from the plant and dispose of as waste. The combination of the bioreactor with a HDS system would provide an optimal solution for remediating U(VI).
The efficiency of removal achieved using the reactor was close to the batch experiments, despite the reactor limitations of nutrient and oxygen availability. Reactors packed using At. ferrooxidans have been previously reported for ferrous ion bio-oxidation and the removal of H2S (Mazuelos et al., 2010) and (Giro et al., 2006). However, this is the first time a bioreactor for the removal of U(VI) using At. ferrooxidans has been described. The results presented here demonstrate an efficient ex-situ treatment of mine water contaminated with U(VI) using At. ferrooxidans. The ability of this microorganism to operate under acidic pH and low nutrient concentration makes it ideal for use in bioreactor technology. Compared to the performance of bacteria indigenous to uranium ore (Islam and Sar, 2016), At. ferrooxidans ability to remove U(VI) is higher, indicating its suitability for use in bioremediation. The efficiency of a reactor such as the one described here may be improved by integration with HDS.
Author Contributions
MR designed the hypothesis and experimental work presented here and contributed to the writing of all sections of the manuscript. BN conducted the laboratory work and contributed to the writing of the manuscript. JH conducted the literature review, contributed to the writing of the literature review, and experimental section and the preparation of the manuscript. DG contributed to the design of the experimental work and the preparation of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to the EPSRC project “Nanometre-scale observation of biodegradation in soil and groundwater” (GR/S72467/01) and the EPSRC Programme grant “Hard-soft matter interfaces: from understanding to engineering” (EP/I001514/1) for financial support. BN and JH thank EPSRC for financial support via The University of Sheffield DTA scholarships.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fenvs.2016.00039
References
Abdelouas, A., Lutze, W., and Nuttall, H. E. (1999). Oxidative dissolution of uraninite precipitated on Navajo sandstone. J. Contam. Hydrol. 36, 353–375. doi: 10.1016/S0169-7722(98)00151-X
Alrakabi, M., Singh, G., Bhalla, A., Kumar, S., Srivastava, A., Rai, B., et al. (2012). Study of uranium contamination of ground water in Punjab state in India using X-ray fluorescence technique. J. Radioanal. Nucl. Chem. 294, 221–227. doi: 10.1007/s10967-011-1585-x
Amouric, A., Brochier-Armanet, C., Johnson, D. B., Bonnefoy, V., and Hallberg, K. B. (2011). Phylogenetic and genetic variation among Fe(II)-oxidizing acidithiobacilli supports the view that these comprise multiple species with different ferrous iron oxidation pathways. Microbiology 157, 111–122. doi: 10.1099/mic.0.044537-0
Barkleit, A., Foerstendorf, H., Li, B., Rossberg, A., Moll, H., and Bernhard, G. (2011). Coordination of uranium(VI) with functional groups of bacterial lipopolysaccharide studied by EXAFS and FT-IR spectroscopy. Dalton Trans. 40, 9868–9876. doi: 10.1039/c1dt10546a
Barkleit, A., Moll, H., and Bernhard, G. (2008). Interaction of Uranium(VI) with lipolysaccharide. Dalton Trans. 21, 2879–2886. doi: 10.1039/b715669c
Beazley, M. J., Martinez, R. J., Sobecky, P. A., Webb, S. M., and Taillefert, M. (2007). Uranium biomineralization as a result of bacterial phosphatase activity: insights from bacterial isolates from a contaminated subsurface. Environ. Sci. Technol. 41, 5701–5707. doi: 10.1021/es070567g
Beazley, M. J., Martinez, R. J., Webb, S. M., Sobecky, P. A., and Taillefert, M. (2011). The effect of pH and natural microbial phosphatase activity on the speciation of uranium in subsurface soils. Geochim. Cosmochim. Acta 75, 5648–5663. doi: 10.1016/j.gca.2011.07.006
Belitz, K., Fram, M. S., and Johnson, T. D. (2015). Metrics for assessing the quality of groundwater used for public supply, CA, USA: equivalent-population and area. Environ. Sci. Technol. 49, 8330–8338. doi: 10.1021/acs.est.5b00265
Bencheikh-Latmani, R., and Leckie, J. O. (2003). Association of uranyl with the cell wall of Pseudomonas fluorescens inhibits metabolism. Geochim. Cosmochim. Acta 67, 4057–4066. doi: 10.1016/S0016-7037(03)00214-X
Bernier-Latmani, R., Vecchia, E. D., Junier, P., Lezama-Pacheco, J. S., Veeramani, H., Suvorova, E. I., et al. (2010). Non-uraninite products of microbial U(VI) reduction. Environ. Sci. Technol. 44, 9456–9462. doi: 10.1021/es101675a
Campbell, K. M., Gallegos, T. J., and Landa, E. R. (2015). Biogeochemical aspects of uranium mineralization, mining, milling, and remediation. Appl. Geochem. 57, 206–235. doi: 10.1016/j.apgeochem.2014.07.022
Choudhary, S., and Sar, P. (2011). Uranium biomineralization by a metal resistant Pseudomonas aeruginosa strain isolated from contaminated mine waste. J. Hazard. Mater. 186, 336–343. doi: 10.1016/j.jhazmat.2010.11.004
Choudhary, S., and Sar, P. (2015). Interaction of uranium (VI) with bacteria: potential applications in bioremediation of U contaminated oxic environments. Rev. Environ. Sci. Biotechnol. 14, 347–355. doi: 10.1007/s11157-015-9366-6
Choy, C. C., Korfiatis, G. P., and Meng, X. (2006). Removal of depleted uranium from contaminated soils. J. Hazard. Mater. 136, 53–60. doi: 10.1016/j.jhazmat.2005.11.011
Dew, D. W., van Buuren, C., McEwan, K., and Bowker, C. (1999). “Bioleaching of base metal sulphide concentrates: a comparison of mesophile and thermophile bacterial cultures,” in Biohydrometallurgy and the Environment Toward the Mining of the 21st Century, Part A, Vol. 9, eds A. Ballester and R. Amils (Madrid: Elsevier Science), 229–238. doi: 10.1016/s1572-4409(99)80022-4
Dispirito, A. A., and Tuovinen, O. H. (1982). Uranous ion oxidation and carbon-dioxide fixation by Thiobacillus ferrooxidans. Arch. Microbiol. 133, 28–32. doi: 10.1007/BF00943765
Dobson, R. S., and Burgess, J. E. (2007). Biological treatment of precious metal refinery wastewater: a review. Miner. Eng. 20, 519–532. doi: 10.1016/j.mineng.2006.10.011
DSMZ (2015). Acidiphilium Medium. Available online at: https://www.dsmz.de/ (Accessed 2016).
Duff, M. C., Hunter, D. B., Bertsch, P. M., and Amrhein, C. (1999). Factors influencing uranium reduction and solubility in evaporation pond sediments. Biogeochemistry 45, 95–114. doi: 10.1007/BF00992875
Ferris, F. G., and Beveridge, T. J. (1986). Site specificity of metallic ion binding in Escherichia coli k-12 lipopolysaccharide. Can. J. Microbiol. 32, 52–55. doi: 10.1139/m86-010
Finneran, K. T., Housewright, M. E., and Lovley, D. R. (2002). Multiple influences of nitrate on uranium solubility during bioremediation of uranium-contaminated subsurface sediments. Environ. Microbiol. 4, 510–516. doi: 10.1046/j.1462-2920.2002.00317.x
Fowle, D. A., Fein, J. B., and Martin, A. M. (2000). Experimental study of uranyl adsorption onto Bacillus subtilis. Environ. Sci. Technol. 34, 3737–3741. doi: 10.1021/es991356h
Gadd, G. M. (1993). Interactions of fungi with toxic metals. New Phytol. 124, 25–60. doi: 10.1111/j.1469-8137.1993.tb03796.x
Gavrilescu, M., Pavel, L. V., and Cretescu, I. (2009). Characterization and remediation of soils contaminated with uranium. J. Hazard. Mater. 163, 475–510. doi: 10.1016/j.jhazmat.2008.07.103
Giro, M. E. A., Garcia, O., and Zaiat, M. (2006). Immobilized cells of Acidithiobacillus ferrooxidans in PVC strands and sultite removal in a pilot-scale bioreactor. Biochem. Eng. J. 28, 201–207. doi: 10.1016/j.bej.2005.11.005
Gorman-Lewis, D., Elias, P. E., and Fein, J. B. (2005). Adsorption of aqueous uranyl complexes onto Bacillus subtilis cells. Environ. Sci. Technol. 39, 4906–4912. doi: 10.1021/es047957c
Haas, J. R., Dichristina, T. J., and Wade, R. (2001). Thermodynamics of U(VI) sorption onto Shewanella putrefaciens. Chem. Geol. 180, 33–54. doi: 10.1016/S0009-2541(01)00304-7
Haas, J. R., and Northup, A. (2004). Effects of aqueous complexation on reductive precipitation of uranium by Shewanella putrefaciens. Geochem. Trans. 5, 41–48. doi: 10.1186/1467-4866-5-41
Islam, E., and Sar, P. (2016). Diversity, metal resistance and uranium sequestration abilities of bacteria from uranium ore deposit in deep earth stratum. Ecotoxicol. Environ. Saf. 127, 12–21. doi: 10.1016/j.ecoenv.2016.01.001
Jelinek, R. T., and Sorg, T. J. (1988). Operating a small full-scale ion exchange system for uranium removal. Am. Water Works Assoc. J. 80, 79–83.
Kelly, S. D., Kemner, K. M., Fein, J. B., Fowle, D. A., Boyanov, M. I., Bunker, B. A., et al. (2002). X-ray absorption fine structure determination of pH-dependent U-bacterial cell wall interactions. Geochim. Cosmochim. Acta 66, 3855–3871. doi: 10.1016/S0016-7037(02)00947-X
Khijniak, T. V., Slobodkin, A. I., Coker, V., Renshaw, J. C., Livens, F. R. Bonch-Osmolovskaya, E. A., et al. (2005). Reduction of Uranium (VI) phosphate during growth of thermophilic bacterium Thermoterrabacterium ferrireducens. Appl. Environ. Microbiol. 71, 6423–6426. doi: 10.1128/AEM.71.10.6423-6426.2005
Langley, S., and Beveridge, T. J. (1999). Effect of O-side-chain-lipopolysaccharide chemistry on metal binding. Appl. Environ. Microbiol. 65, 489–498.
Leduc, L. G., and Ferroni, G. D. (1994). The chemolithotrophic bacterium Thiobacillus ferrooxidans. FEMS Microbiol. Rev. 14, 103–119. doi: 10.1111/j.1574-6976.1994.tb00082.x
Llorens, I., Untereiner, G., Jaillard, D., Gouget, B., Chapon, V., and Carriere, M. (2012). Uranium interaction with two multi-resistant environmental bacteria: Cupriavidus metallidurans CH34 and Rhodopseudomonas palustris. PLoS One 7:e51783. doi: 10.1371/journal.pone.0051783
Lloyd, J. R., Leang, C., Myerson, A. L. H., Coppi, M. V., Cuifo, S., Methe, B., et al. (2003). Biochemical and genetic characterization of PpcA, a periplasmic c-type cytochrome in Geobacter sulfurreducens. Biochem. J. 369, 153–161. doi: 10.1042/bj20020597
Lloyd, J. R., and Renshaw, J. C. (2005). Microbial transformations of radionuclides: fundamental mechanisms and biogeochemical implications. Metal Ions Biol. Syst. 44, 205–240.
Lütke, L., Moll, H., and Bernhard, G. (2012). Insights into the uranium(VI) speciation with Pseudomonas fluorescens on a molecular level. Dalton Trans. 41, 13370–13378. doi: 10.1039/c2dt31080e
Luo, W., and Gu, B. (2011). Dissolution of uranium-bearing minerals and mobilization of uranium by organic ligands in a biologically reduced sediment. Environ. Sci. Technol. 45, 2994–2999. doi: 10.1021/es103073u
Martinez, R. J., Beazley, M. J., Taillefert, M., Arakaki, A. K., Skolnick, J., and Sobecky, P. A. (2007). Aerobic uranium (VI) bioprecipitation by metal-resistant bacteria isolated from radionuclide- and metal-contaminated subsurface soils. Environ. Microbiol. 12:9. doi: 10.1111/j.1462-2920.2007.01422.x
Mazuelos, A., Carranza, F., Romero, R., Iglesias-Gonzalez, N., and Villalobo, E. (2010). Operational pH in packed-bed reactors for ferrous ion bio-oxidation. Hydrometallurgy 104, 186–192. doi: 10.1016/j.hydromet.2010.06.002
Merroun, M. L., and Selenska-Pobell, S. (2001). Interactions of three eco-types of Acidithiobacillus ferrooxidans with U(VI). Biometals 14, 171–179. doi: 10.1023/A:1016658209397
Merroun, M. L., and Selenska-Pobell, S. (2008). Bacterial interactions with uranium: an environmental perspective. J. Contam. Hydrol. 102, 285–295. doi: 10.1016/j.jconhyd.2008.09.019
Morcillo, F., González-Muñoz, M. T., Reitz, T., Romero-González, M. E., Arias, J. M., and Merroun, M. L. (2014). Biosorption and biomineralization of U(VI) by the marine bacterium Idiomarina loihiensis MAH1: effect of background electrolyte and pH. PLoS ONE 9:e91305. doi: 10.1371/journal.pone.0091305
Nedelkova, M., Merroun, M. L., Rossberg, A., Hennig, C., and Selenska-Pobell, S. (2007). Microbacterium isolates from the vicinity of a radioactive waste depository and their interactions with uranium. FEMS Microbiol. Ecol. 59, 694–705. doi: 10.1111/j.1574-6941.2006.00261.x
Neu, M. P., Boukhalfa, H., and Merroun, M. L. (2010). Biomineralization and biotransformations of actinide materials. MRS Bull. 35, 849–857. doi: 10.1557/mrs2010.711
Nevin, K. P., and Lovley, D. R. (2000). Potential for nonenzymatic reduction of Fe(III) via electron shuttling in subsurface sediments. Environ. Sci. Technol. 34, 2472–2478. doi: 10.1021/es991181b
Newsome, L., Morris, K., and Lloyd, J. R. (2014). The biogeochemistry and bioremediation of uranium and other priority radionuclides. Chem. Geol. 363, 164–184. doi: 10.1016/j.chemgeo.2013.10.034
Newsome, L., Morris, K., Shaw, S., Trivedi, D., and Lloyd, J. R. (2015). The stability of microbially reduced U(IV); impact of residual electron donor and sediment ageing. Chem. Geol. 409, 125–135. doi: 10.1016/j.chemgeo.2015.05.016
Orellana, R., Leavitt, J. J., Comolli, L. R., Csencsits, R., Janot, N., Flanagan, K. A., et al. (2013). U(VI) reduction by diverse outer surface c-type cytochromes of Geobacter sulfurreducens. Appl. Environ. Microbiol. 79, 6369–6374. doi: 10.1128/AEM.02551-13
Pan, X. H., Chen, Z., Chen, F. B., Cheng, Y. J., Lin, Z., and Guan, X. (2015). The mechanism of uranium transformation from U(VI) into nano-uramphite by two indigenous Bacillus thuringiensis strains. J. Hazard. Mater. 297, 313–319. doi: 10.1016/j.jhazmat.2015.05.019
Panak, P. J., Knopp, R., Booth, C. H., and Nitsche, H. (2002). Spectroscopic studies on the interaction of U(VI) with Bacillus sphaericus. Radiochim. Acta 90, 779–783. doi: 10.1524/ract.2002.90.9-11_2002.779
Salome, K. R., Green, S. J., Beazley, M. J., Webb, S. M., Kostka, J. E., and Taillefert, M. (2013). The role of anaerobic respiration in the immobilization of uranium through biomineralization of phosphate minerals. Geochim. Cosmochim. Acta 106, 344–363. doi: 10.1016/j.gca.2012.12.037
Selenska-Pobell, S. (2002). “Diversity and activity of bacteria in uranium waste piles,” in Interactions of Microorganisms with Radionuclides, Vol. 2, eds M. J. Keith-Roach and F. R. Livens (Amsterdam: Elsevier Science), 225–254.
Selenska-Pobell, S., Panak, P., Miteva, V., Boudakov, I., Bernhard, G., and Nitsche, H. (1999). Selective accumulation of heavy metals by three indigenous Bacillus strains, B. cereus, B. megaterium and B. sphaericus, from drain waters of a uranium waste pile. FEMS Microbiol. Ecol. 29, 59–67. doi: 10.1111/j.1574-6941.1999.tb00598.x
Senko, J. M., Istok, J. D., Suflita, J. M., and Krumholz, L. R. (2002). In-situ evidence for uranium immobilization and remobilization. Environ. Sci. Technol. 36, 1491–1496. doi: 10.1021/es011240x
Senko, J. M., Mohamed, Y., Dewers, T. A., and Krumholz, L. R. (2005a). Role for Fe(III) minerals in nitrate-dependent microbial U(IV) oxidation. Environ. Sci. Technol. 39, 2529–2536. doi: 10.1021/es048906i
Senko, J. M., Suflita, J. M., and Krumholz, L. R. (2005b). Geochemical controls on microbial nitrate-dependent U(IV) oxidation. Geomicrobiol. J. 22, 371–378. doi: 10.1080/01490450500248911
Shelobolina, E. S., Konishi, H., Xu, H., and Roden, E. E. (2009). U(VI) sequestration in hydroxyapatite produced by microbial glycerol 3-phosphate metabolism. Appl. Environ. Microbiol. 75, 5773–5778. doi: 10.1128/AEM.00628-09
Singh, G., Sengoer, S. S., Bhalla, A., Kumar, S., De, J., Stewart, B., et al. (2014). Reoxidation of biogenic reduced uranium: a challenge toward bioremediation. Crit. Rev. Environ. Sci. Technol. 44, 391–415. doi: 10.1080/10643389.2012.728522
Suzuki, Y., and Banfield, J. F. (2004). Resistance to, and accumulation of, uranium by bacteria from a uranium-contaminated site. Geomicrobiol. J. 21, 113–121. doi: 10.1080/01490450490266361
USA-EPA (2014). Federal Actions to Address the Impact of Uranium Contamination in the Navajo Nation. San Francisco, CA.
Wall, J. D., and Krumholz, L. R. (2006). Uranium reduction. Annu. Rev. Microbiol. 60, 149–166. doi: 10.1146/annurev.micro.59.030804.121357
Keywords: bioremediation of uranium, At. ferrooxidans, bioreactor, mine water, ex-situ process
Citation: Romero-González M, Nwaobi BC, Hufton JM and Gilmour DJ (2016) Ex-situ Bioremediation of U(VI) from Contaminated Mine Water Using Acidithiobacillus ferrooxidans Strains. Front. Environ. Sci. 4:39. doi: 10.3389/fenvs.2016.00039
Received: 05 February 2016; Accepted: 06 May 2016;
Published: 20 May 2016.
Edited by:
Kartik Chandran, Columbia University, USAReviewed by:
Jun-Jie Zhang, Indiana University School of Medicine, USAYinjie Tang, Washington University, USA
Copyright © 2016 Romero-González, Nwaobi, Hufton and Gilmour. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Romero-González, bS5lLnJvbWVyby1nb256YWxlekBzaGVmZmllbGQuYWMudWs=