 Cédric L. Meunier1*
Cédric L. Meunier1* Maarten Boersma1,2
Maarten Boersma1,2 Rana El-Sabaawi3
Rana El-Sabaawi3 Halvor M. Halvorson4
Halvor M. Halvorson4 Emily M. Herstoff5
Emily M. Herstoff5 Dedmer B. Van de Waal6
Dedmer B. Van de Waal6 Richard J. Vogt7
Richard J. Vogt7 Elena Litchman8
Elena Litchman8- 1Alfred-Wegener-Institut Helmholtz-Zentrum für Polar- und Meeresforschung (AWI), Biologische Anstalt Helgoland, Helgoland, Germany
- 2Faculty Biology/Chemistry (FB2), University of Bremen, Bremen, Germany
- 3Department of Biology, University of Victoria, Victoria, BC, Canada
- 4Department of Biological Sciences, University of Southern Mississippi, Hattiesburg, MS, USA
- 5Department of Ecology and Evolution, Stony Brook University, Stony Brook, NY, USA
- 6Department of Aquatic Ecology, Netherlands Institute of Ecology (NIOO-KNAW), Wageningen, Netherlands
- 7Department of Biological Sciences, Trent University, Peterborough, ON, Canada
- 8W.K. Kellogg Biological Station, Michigan State University, Hickory Corners, MI, USA
The theories developed in ecological stoichiometry (ES) are fundamentally based on traits. Traits directly linked to cell/body stoichiometry, such as nutrient uptake and storage, as well as the associated trade-offs, have the potential to shape ecological interactions such as competition and predation within ecosystems. Further, traits that indirectly influence and are influenced by nutritional requirements, such as cell/body size and growth rate, are tightly linked to organismal stoichiometry. Despite their physiological and ecological relevance, traits are rarely explicitly integrated in the framework of ES and, currently, the major challenge is to more closely inter-connect ES with trait-based ecology (TBE). Here, we highlight four interconnected nutrient trait groups, i.e., acquisition, body stoichiometry, storage, and excretion, which alter interspecific competition in autotrophs and heterotrophs. We also identify key differences between producer-consumer interactions in aquatic and terrestrial ecosystems. For instance, our synthesis shows that, in contrast to aquatic ecosystems, traits directly influencing herbivore stoichiometry in forested ecosystems should play only a minor role in the cycling of nutrients. We furthermore describe how linking ES and TBE can help predict the ecosystem consequences of global change. The concepts we highlight here allow us to predict that increasing N:P ratios in ecosystems should shift trait dominances in communities toward species with higher optimal N:P ratios and higher P uptake affinity, while decreasing N retention and increasing P storage.
Introduction
Ecological stoichiometry (ES) is a framework that links an organism's metabolic demands with the relative supply of elements in the environment (Sterner and Elser, 2002; Hessen et al., 2013). It postulates a crucial relationship between the balance of elements, typically but not limited to carbon (C), nitrogen (N), and phosphorus (P), and their role in determining growth and reproduction of organisms as well as in ecological interactions. The recognition of the importance of stoichiometric constraints between consumer needs and prey nutrient content has substantially increased our understanding of trophic interactions. For example, high C:P food is often of low-quality for a variety of organisms including molluscs (Stelzer and Lamberti, 2002; Fink and Elert, 2006), crustaceans (Boersma and Kreutzer, 2002; Meunier et al., 2012, 2016a), insects (Perkins et al., 2004), fish (Borlongan and Satoh, 2001; Vrede et al., 2011), and birds (Grone et al., 1995). ES has therefore proven to be a highly suitable framework in community ecology, explaining consumer responses to prey food quality (food intake, growth, as well as competition between consumer species, and consumer effects on prey nutrient composition (Sterner, 1990; Sterner et al., 1992; Sterner and Hessen, 1994).
Trait-based ecology (TBE) focuses on functional traits expressed by an organism allowing its growth and survival under distinct environmental conditions (McGill et al., 2006). Functional traits are the morphological, physiological, phenological, and behavioral characteristics of an organism that influence its performance or fitness. As such, TBE couples biological function to the success of species in a food-web (Litchman and Klausmeier, 2008; Litchman et al., 2013; Kremer et al., 2016). More specifically, TBE focuses on traits rather than taxa, providing a functional approach to understanding ecological interactions. For example, it has been shown that trait diversity can provide a better predictor of primary production as compared to taxonomic diversity (Vogt et al., 2010). Numerous studies indicate that abiotic parameters, such as temperature, precipitation, and nutrient concentrations, can directly influence spatial and temporal trait patterns both at the population and at the community levels (Brun et al., 2016). TBE therefore constitutes a powerful tool linking species functional characteristics to their distributions along environmental gradients, as well as to community interactions and ecosystem function.
The framework of ES is essentially, although not explicitly, based on traits such as homeostasis, growth rate, and nutrient uptake. For instance, the relationship between organism size and stoichiometry has been well studied (see below for more detail) but connecting these ES traits with more traditional TBE traits still remains a major challenge. Combining the framework of ES with TBE furthers the coupling of elements to functional traits from subcellular processes to species interactions and ultimately ecosystem dynamics. For example, linking ES with TBE could help studying how variation in traits related to elemental body composition influences organismal fitness (Leal et al., 2016). The focus of this paper is to explore and synthesize existing insights, and to develop novel connections between ES and TBE. We first review key traits and their elemental requirements, highlighting differences between trophic levels as well as ecosystems. Next, we describe trade-offs between traits that directly or indirectly affect the elemental composition or requirements of organisms. We also develop hypotheses on how different traits could be linked to life history trade-offs, and we outline community and ecosystem consequences of variation in ES traits and identify limitations and opportunities for future research further connecting TBE and ES. This framework will foster collaboration between scientists of different disciplines (freshwater, marine, and terrestrial ecologists) and enhance our understanding of fundamental ecological issues.
Traits and Elemental Balances
Four interconnected major trait groups affect the balance of energy and elements in organisms: acquisition, cell/body stoichiometry, storage, and excretion. These traits define how organisms interact with their environment as well as with one another, and are, therefore, among the key determinants of ecological niches. This elemental trait framework has already been successfully used to identify how resource imbalances affect basic physiological processes (Frost et al., 2005). In the following sections, we describe different strategies utilized by autotrophs and heterotrophs to acquire and use C and nutrients, and we explain how these traits directly influence organismal stoichiometry. While traditional stoichiometric approaches are implicitly trait-based, we here aim to fully place life history trade-offs in a stoichiometric context. This conceptual framework should enhance our ability to predict how communities will respond to changes in nutrient conditions in the environment.
Autotrophs
Autotrophs obtain energy (mostly sunlight) and material from different, uncoupled sources within their environment. Therefore, plants needed to develop strategies to store these resources, as the availability of one resource does not guarantee another one. Nutrient uptake strategies are often linked to storage capacity and therefore to plasticity in cellular stoichiometry. When C or nutrients are taken up and fixed by autotrophs, they may be used immediately for growth or accumulate as storage pools for later use (Chapin, 1980; Reynolds, 2006). Due to their need and hence capacity to store nutrients, autotroph C:N:P ratios greatly vary with available nutrient ratios, indicating a high flexibility in chemical composition and a lack of homeostasis (Sterner and Elser, 2002; Meunier et al., 2014). This can result in different competitive strategies between various autotroph species. For instance, for phytoplankton, three major nutrient acquisition strategies have been proposed (Sommer, 1984): (1) velocity-adapted species, or r-strategists, with high maximum nutrient uptake rates and high maximum growth rates that are able to directly utilize nutrient pulses for growth, (2) storage-adapted species, with high rates of nutrient uptake but lower maximum growth rates that have the ability to store excess nutrients, and (3) affinity-adapted species, or K-strategists, with the ability to effectively take up and assimilate growth-limiting nutrients even at low concentrations, a strategy that is advantageous in oligotrophic environments. Comparable nutrient-acquisition strategies exist in terrestrial systems, and nutrient availability strongly influences interspecific plant interactions and community composition (Zemunik et al., 2015). For instance, tall-growing species with high growth rates are generally favored by high nutrient inputs at the expense of species with conservative growth strategies (Diekmann and Falkengren-Grerup, 2002). Physiological traits such as nutrient requirements also alter interspecific competition, giving a competitive advantage to species with high-nutrient affinity (Price and Morgan, 2007).
Such different strategies may affect the stoichiometry of autotrophs. For instance, high growth rates may lead to higher demands for P (see below), storage will increase cellular quota of distinct elements, while high affinities will reduce the minimum nutrient requirements (Figure 1). Stoichiometric plasticity will furthermore be determined by physiological limits, as was shown for phytoplankton N:P ratios that increased non-linearly with increasing N:P supply ratios (Rhee, 1978; Persson et al., 2010). In other words, the upper limit of nutrient quota is determined by storage capabilities, while the lower limits are determined by minimal structural and functional requirements, as well as the affinity for a nutrient. These boundaries within which the organism's stoichiometry can fluctuate represent the “homeostatic capacity” parameter defined by Meunier et al. (2014). This parameter corresponds to the boundaries within which the organism's body stoichiometry can fluctuate and therefore characterizes storage capacity. Consequently, storage- and affinity-adapted species will be generally characterized by variable stoichiometric composition while velocity-adapted species with high P demands will have low cellular N:P ratios (Hillebrand et al., 2013).
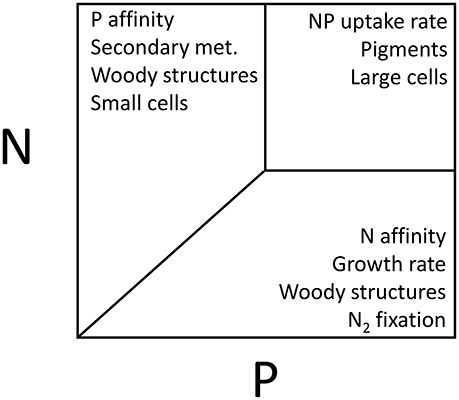
Figure 1. Influence of N and Pavailabilities on trait dominance in autotrophs. Low N and P supplies will favor species with high affinities and may also facilitate synthesis of C-rich woody structures. Furthermore, species with C-rich woody structures may be facilitated by low N and P supplies. Relatively high N supplies may selectively favor synthesis of N-rich secondary metabolites, while relatively high P supplies may selectively favor faster growth, and N2 fixation (in cyanobacteria). Both high N and P supplies may facilitate large species not limited by nutrients, and favor high maximum uptake rates. Furthermore, high N and P supplies will promote autotroph biomass and consequently self-shading, and thus may favor species with high pigment contents and/or species with accessory pigments.
Variation in N:P ratios between organisms often reflect functional differences, such as the ones that have been described for optimal N:P ratios (Güsewell, 2004; Hillebrand et al., 2013), i.e., the N:P ratio at which growth is maximized. In aquatic environment for example, a recent meta-analysis identified a scaling relationship between maximum growth rate and phytoplankton nutrient demand (Hillebrand et al., 2013). Phytoplankton N:P ratios decrease with increasing growth rates and simultaneously display decreasing variance, particularly driven by P limitation, which follows predictions of the Growth Rate Hypothesis (Sterner, 1995a; Elser et al., 1996; Sterner and Elser, 2002). This suggests that fast growing phytoplankton species, or within a species under less severe limiting conditions, phytoplankton are relatively more P-rich. Using a trait-based eco-evolutionary model, Klausmeier et al. (2004) showed that different environmental conditions select for species with different N:P ratios: P-rich conditions select for fast growers with low N:P, while P-limited conditions select for better P competitors with higher N:P ratios (due to their investment in resource acquisition proteins, Figure 1). Not only the N:P ratio itself but the form of N and P available to phytoplankton matters. While we may expect higher biomass of small phytoplankton at lower N:P supply due to their generally fast growth (and thus high P requirements), higher picophytoplankton densities are only observed under high N:P conditions (Figure 1; Glibert, 2016) which is likely caused by changes in N redox and relative proportions of reduced relative to oxidized N (Glibert et al., 2016). A review of interspecific differences in N:P ratios of terrestrial plants concluded that N:P ratios correlate negatively with maximum relative growth rate in herbaceous and woody plants (Güsewell, 2004). Species with inherently low N:P ratios are predicted to dominate N-limited communities and should be favored during P fertilization (Figure 2; Tilman, 1997). Therefore, nutrient supply and composition often shape terrestrial and aquatic autotroph community composition by species sorting as well as by dynamic shifts in species' C:N:P stoichiometry (Sterner and Elser, 2002; Persson et al., 2010; Meunier et al., 2014).
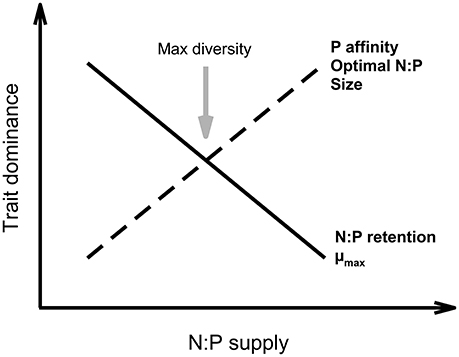
Figure 2. Influence of N:P supply on trait dominance. With increasing N:P supply, an organism's optimal N:P ratio, P uptake affinity, and size are expected to increase, and organisms that selectively retain P over N should have higher maximal growth rates than those which do not, as predicted by the Growth Rate Hypothesis (Sterner, 1995b; Elser et al., 2000c). Further, Tilman's resource-ratio based theory of competition predicts that biodiversity should be maximal (gray arrow) at balanced supply ratios (Tilman, 1982).
Heterotrophs
Autotrophs' stoichiometry also reflects their quality as food for herbivores. Since autotrophs are usually more C-rich than animals, herbivores typically ingest a diet rich in C but deficient in nutrients (Sterner and Elser, 2002), which has implications for herbivore and detritivore performance and subsequent trophic dynamics (Elser et al., 2000b; Hessen et al., 2013). Grazers however possess a large range of adaptations that allow them to minimize the consequences of these nutrient imbalances. Besides selective feeding (e.g., Kagata and Ohgushi, 2011; Meunier et al., 2012, 2016a) and habitat choice (Winder et al., 2004; Reichwaldt, 2008), consumers may handle poor food quality by selectively retaining and excreting nutrients (Elser and Urabe, 1999; Knoll et al., 2009). These physiological adjustments are tightly linked with the stoichiometry of traits such as elemental ratios of recycled materials and body stoichiometric homeostasis.
Nutrient storage capacity is a much more confined trait in heterotrophs than in autotrophs resulting in more constrained body composition (Persson et al., 2010; Meunier et al., 2014). While the lack of homeostasis allows organisms to store nutrients (see above), the advantages of stoichiometric homeostasis remain unclear. It can be argued that the ability to store nutrients, which results in flexible body stoichiometry, is the more advantageous strategy, especially where food supply or food quality fluctuates (Meunier et al., 2014). It has been hypothesized that homeostasis is the optimal response strategy to fluctuations in food quality, as it should minimize the cost of adjusting to new conditions (e.g., selective nutrient retention and excretion) while maximizing growth rate (Giordano, 2013). Indeed, when feeding conditions are relatively stable within an organism's life span, homeostasis minimizes noise in physiological channels and establishes a stable environment for cellular processes (Woods and Wilson, 2013).
Stoichiometric homeostasis also results from adjustments in the elemental ratios of recycled material. The stoichiometry of excreted material is influenced by both the consumer's stoichiometry, e.g., low N:P consumers will have high N:P excretion, as well as by the resource stoichiometry, e.g., the excreted N:P ratio will increase with increasing resource N:P ratio (Vanni, 2002). However, several studies question the link between body stoichiometry and elemental ratios of excreted material (Allgeier et al., 2015; Vanni and McIntyre, 2016). The lack of evidence for the relationship between resource stoichiometry and consumer nutrient excretion is likely due to the fact that, in these studies, bulk stoichiometry was measured instead of the stoichiometry of assimilated materials (e.g., Dodds et al., 2014).
Interspecific variations in recycling traits will also have consequences at the ecosystem scale. For instance, experiments in lakes provided evidence that the replacement of the high body N:P copepods with low N:P Daphnia caused a transition from N to P limitation for primary producers (Sterner, 1990; Elser et al., 1996), likely driven by the higher N:P ratio of recycled material (Elser et al., 1988). In addition, intraspecific variation in nutrient recycling traits during copepod ontogeny has been suggested to influence biogeochemical cycling within marine systems (Meunier et al., 2016a). Like zooplankton, aquatic vertebrates such as fish and amphibians can also have important stoichiometric impacts on nutrient cycling and trophic dynamics (Vanni et al., 2002). Similarly, in terrestrial systems, consumers' body stoichiometry and nutrient recycling traits are expected to shape limitation patterns (Cherif and Loreau, 2009, 2013). However, it is important to consider the intrinsic differences between producer-consumer interactions in aquatic and terrestrial ecosystems. While planktonic herbivores may eat a very large percentage (70–80%) of the daily primary production (Calbet, 2008; Löder et al., 2011), in terrestrial ecosystems, plants are less nutritional and lose lower percentages of production to herbivores, and a higher level of C is channeled as detritus (Cebrian, 1999). Traits directly influencing herbivore stoichiometry should therefore play only a small role in the cycling of nutrients in forested ecosystems, with decomposers and detritivores playing a more important role (Cherif and Loreau, 2009, 2013). Given the importance of homeostasis for biogeochemical cycling in food webs, quantifying the contribution of this trait to ecosystem services related to biogeochemical cycles (i.e., C sequestration, water quality) represents a promising avenue for future research. The consequences of differences in homeostasis between autotrophic and heterotrophic microbes for nutrient cycling have been explored in a number of studies (e.g., Cross et al., 2005; Cotner et al., 2010). Further, exploring the importance of the stoichiometry of traits as both drivers of, and responses to, ecosystem changes will result in datasets that will advance our understanding of how biological communities will perform under different environmental conditions, including the changing climate. This approach could in turn be used to explore traits that both directly and indirectly affect a given ecosystem function or service.
Trait Connections
Correlative Relationships
Several studies have linked elemental stoichiometry to distinct species traits in order to explain ecosystem structure (for an overview see Table 1). As we previously mentioned, the tight relationship between the body N:P ratio and organismal growth rate is formulated in the Growth Rate Hypothesis (Table 1), a central hypothesis in ES. It postulates that species with high growth rates have more ribosomal RNA content for rapid protein synthesis than slow-growing species (Sterner and Elser, 2002). Because ribosomal RNA is rich in P, faster growing species should have a higher P content and lower body N:P ratios (Sterner, 1995a; Elser et al., 1996; Sterner and Elser, 2002). Although the Growth Rate Hypothesis is supported for a large range of organisms, several studies have questioned its generality (Sardans et al., 2012, and reference therein). For instance, mass-balance modeling demonstrated that maintenance costs for high biomass P content can drive the relationship between P content and growth rate (Shimizu and Urabe, 2008). This has been confirmed in experiments showing that consumers release P at a substantial rate, even when fed high C:P food (Demott et al., 1998; Shimizu and Urabe, 2008). Despite these limitations associated with P losses, e.g., through molting (Shimizu and Urabe, 2008), a large body of literature points toward a negative relationship between growth rate and body C:P or N:P (e.g., Carrillo et al., 2001; Meunier et al., 2016a).
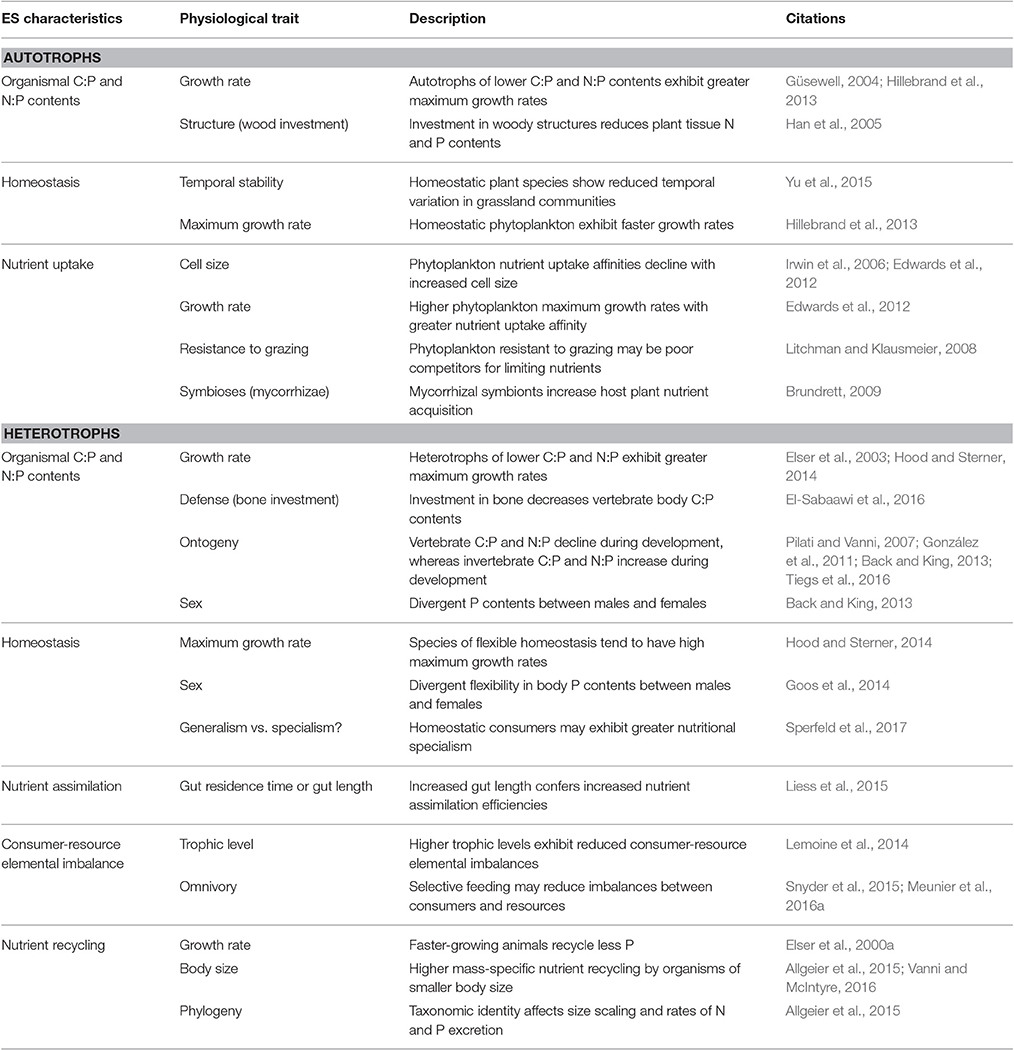
Table 1. Relationships between elemental stoichiometry characteristics and physiological traits among autotrophs and heterotrophs, with abbreviated description of the linkage and associated citations.
Interestingly, organisms' size and growth rate are usually negatively correlated (Table 1), which implies that, based on the growth rate hypothesis, smaller heterotrophic organisms might generally have lower N:P ratios (Figure 2). These explicit connections between body size, ontogeny, and N:P stoichiometry have been well documented (Elser et al., 1996; Gillooly et al., 2005; Méndez and Karlsson, 2005; Meunier et al., 2016a). Further, larger cells have higher biomass-specific storage capacity due to smaller surface/volume ratios and higher minimum cellular metabolic requirement that selectively allows them to provide them with a competitive advantage over smaller cells under higher resource concentrations (Figure 3; Irwin et al., 2006). Experiments in lakes showed that the interaction between top-down and bottom-up controls can lead to the replacement of small zooplankton with high N:P by larger species with low N:P (Sterner et al., 1992; Hall et al., 2004). This community shift also causes a transition from N to P limitation for primary producers, likely caused by the higher N:P ratio of recycled material (Elser et al., 1988). However, the relationship between size and stoichiometry is expected to be opposite in vertebrates (i.e., decreasing N:P ratios with increasing body size) due to relatively higher P needs for bones (Elser et al., 1996; González et al., 2011). The interplay between traits and elemental requirements hence influences the success of an organism in response to different environmental conditions (Table 1), which, in turn, will shape community structure.
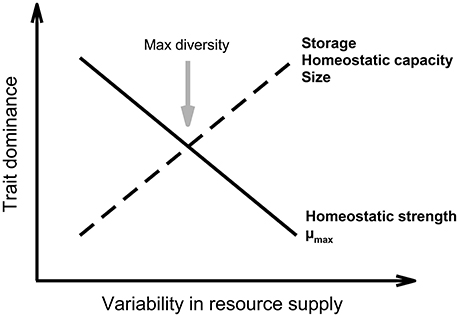
Figure 3. Influence of variability in resource supply on traits. Increasing variability is expected to influence the trade-off between fast growth and investment in storage. Higher variability should therefore yield communities with larger organisms with greater storage and homeostatic capacities, but slower maximal growth rates. Moreover, the intermediate disturbance hypothesis suggests that local species diversity should be maximized when ecological disturbance is neither too rare nor too frequent (Wilkinson, 1999). Biodiversity should therefore be maximal at intermediate resource supply variability (gray arrow).
Changes in environmental conditions can also lead to evolutionary changes in traits. The mutual interactions between the evolved functional traits and environment characteristics therefore have gained increasing interest over the past two decades. In particular, understanding how genome and proteome adaptations are shaped by selection on growth-related traits and the parameters determining the extent of stoichiometrically relevant variation in genomes across taxa are increasingly studied (Kay et al., 2005). For example, genes that respond to changes in P availability have received much attention, and variation in the environmental supply of P has been associated with the expression of highly conserved genes (Jeyasingh and Weider, 2007; Frisch et al., 2014). Rotifer populations with a P limitation selection history yielded higher biomass and reduced food to lower levels when fed with P limited food (Declerck et al., 2015). Although this adaptation of populations did not involve changes in elemental composition, it did alter important population traits, including birth and death rates, population structure and grazing pressure. Both TBE and ES are at the intersection of evolution and ecology, and their integration will thus greatly help understanding eco-evolutionary dynamics (Jeyasingh et al., 2014; Declerck et al., 2015).
Trade-Offs
Natural selection balances traits associated with the three main missions of any organism, i.e., to eat, survive, and reproduce in order to maximize fitness. However, it is generally not possible to maximize all traits simultaneously, particularly as resources are often limiting. Trade-offs are therefore inevitable and different organisms specialize in various aspects of their life history. Such trade-offs are typically enabled by trait specialization and plasticity. For example, consumers with lower P requirements and higher body N:P ratios should be abundant in low P environments despite having reduced maximum growth rates (Sterner, 1995b; Elser et al., 2000c). The quantification of trade-offs associated with key traits will therefore yield a set of comparisons allowing the prediction of physiological, morphological, and behavioral responses of communities to environmental changes (Litchman and Klausmeier, 2008; Litchman et al., 2013). Particularly regarding stoichiometry, Montechiaro and Giordano (2010) discussed the conditions influencing the trade-off between stoichiometric homeostasis and maximal growth rate. Generally, when enduring periods of nutrient limitation, an organism should adopt a strategy that lessens the cost of adjusting to the new conditions (degree of homeostasis) and maximizes growth rate. This trade-off has been observed in experiments where phytoplankton were grown at different growth rates and subjected to a stress (Fanesi et al., 2014). The authors observed that, when the duration of the perturbation was long enough, cells with lower growth rate were more elementally homeostatic after a change in nutrient supply, which indicates that the trade-off between acclimation and homeostasis depends on the duration of the perturbation relative to cellular division rate (Fanesi et al., 2014). Based on these results, we hypothesize that homeostatic strength should decrease with increasing variability in resource supply (Figure 3). Similar patterns are expected in heterotrophs (Meunier et al., 2014) but, to our knowledge, no experiment has yet been conducted to test this prediction.
Similarly, low nutrient availability in the environment forces plants to adopt different strategies. For example, they can invest in storage traits and preferentially accumulate N rather than P (Chapin et al., 1990; Yu et al., 2015). Experimental work in freshwater (e.g., Sommer, 1984; Li and Stevens, 2012), marine (e.g., Smayda, 1997), and terrestrial systems (for review see Chapin et al., 1990) has shown that investing in storage traits provides a competitive advantage under highly variable and unpredictable nutrient availabilities (Figure 3). For example, organisms with high N affinity and storage capacity relative to other nutrients should have a competitive advantage under low N supply, as is often observed in N-limited terrestrial systems (Meunier et al., 2016b). These results indicate that the trade-offs between size, rapid growth, and storage capacity are strongly regulated by nutrient availability as well as the frequency and duration of nutrient pulses relative to organismal growth rates (Figure 3). Thus, if nutrient pulses are regular, small velocity-adapted species should prosper due to their high maximum nutrient uptake rates and high maximum growth rates (Figure 3; Sommer, 1984). Along similar lines of reasoning, the intermediate disturbance hypothesis suggests that local species diversity should be maximized when ecological disturbance is neither too rare nor too frequent (Wilkinson, 1999), although the theoretical underpinnings of this hypothesis have been increasingly questioned (Fox, 2013). This implies that biodiversity is maximized when the resource supply is intermediately variable (Figure 3). Indeed, N pulses of intermediate frequencies may lead to coexistence of different strategies, i.e., velocity and storage specialists (Grover, 1991; Litchman et al., 2009).
In their review of phytoplankton traits and trade-offs, Litchman and Klausmeier (2008) suggest that the trade-off between nutrient competitive abilities and grazer resistance is key among phytoplankton. One strategy to obtain resistance to grazing is through a large cell size, which is associated with lower growth rate and reduced nutrient uptake abilities because of the decreased surface area to volume ratios (Reynolds, 1988) but increases the cell's storage capacity (Litchman et al., 2009; Wirtz, 2013). However, small size also offers several advantages to phytoplankton, including a more efficient acquisition of limiting nutrients (Sherwood et al., 1975; Ploug et al., 1999) and higher maximum growth rates (Banse, 1976), although it can increase susceptibility to grazing (Thingstad et al., 2005). This trade-off is one of the drivers for species succession during phytoplankton blooms as well as annual changes in community composition (Reynolds, 1984; Sommer et al., 1986). Organismal replacement along nutrient gradients can also be driven by this trade-off (Leibold, 1996), which ultimately has consequences for biodiversity within communities (Kneitel and Chase, 2004). The trade-off between cell size and competitive ability for limiting nutrients is therefore a key determinant of species dominance in phytoplankton communities. In addition to pairwise trade-offs, traits can be linked via multidimensional trade-offs. For example, Edwards et al. (2011) identified a three-way trade-off between N vs. P competitive abilities and cell size as a proxy for grazer resistance.
Implications
Understanding the link between nutrient stoichiometry and organismal traits can help predict how human-induced changes in biogeochemical cycles will alter the interaction between producers' stoichiometry and consumer elemental requirements. Human activities have altered the C, N, and P biogeochemical cycles on a global scale (Peňuelas et al., 2012). These changes are apparent from increasing atmospheric CO2 concentrations and the large amounts of nutrients applied on land, subsequently also enriching lakes, rivers, and marine coastal waters. More specifically, the use of agricultural fertilizers has tremendously disturbed global biogeochemical cycles and increased the amount of N and P available to plants and animals (Bobbink et al., 2010). Not only do the C, N, and P supplies increase, also their relative abundances are changing both in aquatic and terrestrial environments (Grizzetti et al., 2012; Peňuelas et al., 2012; Sardans et al., 2012). Such shifts in the relative abundances of elements will alter autotroph stoichiometry and thereby their quality as food for herbivores (Van De Waal et al., 2010). For example, the N:P ratio available to plants has risen over the past decades in both terrestrial and aquatic systems (Sardans et al., 2012). The concepts we highlight here allow us to predict that such changes in nutrient availability should lead to communities dominated by species with higher optimal N:P ratios and higher P uptake affinity, while decreasing N retention and increasing P storage (Figure 2). The Growth Rate Hypothesis allows us to predict slower maximal growth rates at higher N:P supply and decreasing organismal size with increasing N:P supply, because body size is generally negatively correlated with growth rate (Figure 2). Linking TBE with ES therefore enables us to make predictions regarding the impact of human activities on both community structure and functioning. For example, it has been observed that communities dominated by species with higher optimal N:P ratios will substantially influence biogeochemical cycles by preferentially recycling N over P (Vanni et al., 2002; Knoll et al., 2009; El-Sabaawi et al., 2016).
Conclusion
Coupling functional traits to the stoichiometry of organisms allows a more general understanding of ecological interactions. Specifically, optimal body N:P ratios, nutrient uptake and storage traits, as well as their associated trade-offs, have the potential to drive species competition and thereby influence food web interactions and ecosystem dynamics. Quantifying the contribution of these traits to ecosystem services (i.e., C sequestration, water quality) represents a promising avenue for research into changes in biogeochemical cycling associated with global environmental change. At the same time, traits indirectly coupled to elemental demands, such as cell/body size and growth rate have a strong influence on, and are affected by, organismal stoichiometry. Therefore, combining observations and ideas from ES and TBE offers a unified framework that enables answering a wide array of complex ecological questions, for instance how biological communities will perform under changing environmental conditions. Linking and applying multiple ecological frameworks allows crosstalk between the various scientific disciplines, fostering the exchange of comparable efforts in understanding the complexity of ecosystem structure and functioning.
Author Contributions
The manuscript was written by CM. Table 1 was created by HH. Figure 1 was created by DW. Figures 2, 3 were created by CM. All co-authors helped to evaluate and edit the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This article summarizes the work done in the “Workshop on Biological Stoichiometry and Trait-Based Ecology” that took place during the Conference On Biological Stoichiometry (2015, Trent, Canada). This conference was supported by the David Schindler Professorship of Aquatic Sciences at Trent University and by Trent University. The workshop was supported by the Canadian Institute of Ecology and Evolution. We thank all workshop participants for their input into the discussions: Claudia Acquisti, Thomas Anderson, Jeff Back, Stephen Baines, Esteban Balseiro, Gergely Boros, Krista Capps, Sarah Collins, Richard Connon, James Cotner, Michael Danger, Sanatan Das Gupta, Steven Declerck, Lenore Doumas, Dan Durston, Jonathan Ebel, James Elser, Michelle Evans-White, Carolyn Faithfull, Isabel Fernandes, Laura Fidalgo, Michał Filipiak, Stephanie Fong, Paul Frost, Andrea Gall, Pat Glibert, Casey Godwin, Angelica L. Gonzalez, Jared Goos, Jin-Sheng He, Puni Jeyasingh, Susan Kilham, Ryan King, Jude Kong, James Larson, Cecilia Laspoumaderes, Kimberley Lemmen, Shawn Leroux, Patrick Lind, Paloma Lopes, Keeley MacNeill, Adam Martiny, Peter B. McIntyre, Katie Miller, Eric Moody, David Ott, Rachel Paseka, Angela Peace, Amber Rock, Anna Rożen, Amanda Rugenski, Andrew Sanders, Jennifer Schmitt, Kimberly Schulz, Ryan Sherman, Robert Sterner, Maren Striebel, Caroline Turner, Jotaro Urabe, Michael Vanni, Stoycho Velkovsky, Mandy Velthuis, Nicole Wagner, Wei Wang, Tanner Williamson, and Donald Yee.
References
Allgeier, J. E., Wenger, S. J., Rosemond, A. D., Schindler, D. E., and Layman, C. A. (2015). Metabolic theory and taxonomic identity predict nutrient recycling in a diverse food web. Proc. Natl. Acad. Sci. U.S.A. 112, E2640–E2647. doi: 10.1073/pnas.1420819112
Back, J. A., and King, R. S. (2013). Sex and size matter: ontogenetic patterns of nutrient content of aquatic insects. Freshw. Sci. 32, 837–848. doi: 10.1899/12-181.1
Banse, K. (1976). Rates of growth, respiration and photosynthesis of unicellular algae as related to cell size—a review. J. Phycol. 12, 135–140.
Bobbink, R., Hicks, K., Galloway, J., Spranger, T., Alkemade, R., Ashmore, M., et al. (2010). Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol. Appl. 20, 30–59. doi: 10.1890/08-1140.1
Boersma, M., and Kreutzer, C. (2002). Life at the edge: is food quality really of minor importance at low quantities? Ecology 83, 2552–2561. doi: 10.1890/0012-9658(2002)083[2552:LATEIF]2.0.CO;2
Borlongan, I. G., and Satoh, S. (2001). Dietary phosphorus requirement of juvenile milkfish, Chanos chanos (Forsskal). Aquac. Res. 32, 26–32. doi: 10.1046/j.1355-557x.2001.00003.x
Brun, P., Payne, M. R., and Kiørboe, T. (2016). Trait biogeography of marine copepods–an analysis across scales. Ecol. Lett. 19, 1403–1413. doi: 10.1111/ele.12688
Brundrett, M. (2009). Mycorrhizal associations and other means of nutrition of vascular plants: understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 320, 37–77. doi: 10.1007/s11104-008-9877-9
Calbet, A. (2008). The trophic roles of microzooplankton in marine systems. ICES J. Mar. Sci. 65, 325–331. doi: 10.1093/icesjms/fsn013
Carrillo, P., Villar-Argaiz, M., and Medina-Sánchez, J. M. (2001). Relationship between N:P ratio and growth rate during the life cycle of calanoid copepods: an in situ measurement. J. Plankton Res. 23, 537–547. doi: 10.1093/plankt/23.5.537
Cebrian, J. (1999). Patterns in the fate of production in plant communities. Am. Nat. 154, 449–468. doi: 10.1086/303244
Chapin, F. S. (1980). The mineral nutrition of wild plants. Ann. Rev. Ecol. Syst. 11, 233–260. doi: 10.1146/annurev.es.11.110180.001313
Chapin, I. E., Schulze, A., and Mooney, H. A. (1990). The ecology and economics of storage in plants. Annu. Rev. Ecol. Syst. 21, 423–447. doi: 10.1146/annurev.es.21.110190.002231
Cherif, M., and Loreau, M. (2009). When microbes and consumers determine the limiting nutrient of autotrophs: a theoretical analysis. Proc. R. Soc. Lond. B Biol. Sci. 276, 487–497. doi: 10.1098/rspb.2008.0560
Cherif, M., and Loreau, M. (2013). Plant–herbivore–decomposer stoichiometric mismatches and nutrient cycling in ecosystems. Proc. R. Soc. B Biol. Sci. 280:20122453. doi: 10.1098/rspb.2012.2453
Cotner, J. B., Hall, E. K., Scott, T., and Heldal, M. (2010). Freshwater bacteria are stoichiometrically flexible with a nutrient composition similar to seston. Front. Microbiol. 1:132. doi: 10.3389/fmicb.2010.00132
Cross, W. F., Benstead, J. P., Frost, P. C., and Thomas, S. A. (2005). Ecological stoichiometry in freshwater benthic systems: recent progress and perspectives. Freshw. Biol. 50, 1895–1912. doi: 10.1111/j.1365-2427.2005.01458.x
Declerck, S. A., Malo, A. R., Diehl, S., Waasdorp, D., Lemmen, K. D., Proios, K., et al. (2015). Rapid adaptation of herbivore consumers to nutrient limitation: eco-evolutionary feedbacks to population demography and resource control. Ecol. Lett. 18, 553–562. doi: 10.1111/ele.12436
Demott, W. R., Gulati, R. D., and Siewertsen, K. (1998). Effects of phosphorus-deficient diets on the carbon and phosphorus balance of Daphnia magna. Limnol. Oceanogr. 43, 1147–1161. doi: 10.4319/lo.1998.43.6.1147
Diekmann, M., and Falkengren-Grerup, U. (2002). Prediction of species response to atmospheric nitrogen deposition by means of ecological measures and life history traits. J. Ecol. 90, 108–120. doi: 10.1046/j.0022-0477.2001.00639.x
Dodds, W. K., Collins, S., Hamilton, S., Tank, J., Johnson, S., Webster, J., et al. (2014). You are not always what we think you eat: selective assimilation across multiple whole-stream isotopic tracer studies. Ecology 95, 2757–2767. doi: 10.1890/13-2276.1
Edwards, K. F., Klausmeier, C. A., and Litchman, E. (2011). Evidence for a three-way tradeoff between nitrogen and phosphorus competitive abilities and cell size in phytoplankton. Ecology 92, 2085–2095. doi: 10.1890/11-0395.1
Edwards, K. F., Thomas, M. K., Klausmeier, C. A., and Litchman, E. (2012). Allometric scaling and taxonomic variation in nutrient utilization traits and maximum growth rate of phytoplankton. Limnol. Oceanogr. 57, 554–566. doi: 10.4319/lo.2012.57.2.0554
El-Sabaawi, R. W., Warbanski, M. L., Rudman, S. M., Hovel, R., and Matthews, B. (2016). Investment in boney defensive traits alters organismal stoichiometry and excretion in fish. Oecologia 181, 1209–1220. doi: 10.1007/s00442-016-3599-0
Elser, J. J., Acharya, K., Kyle, M., Cotner, J., Makino, W., Markow, T., et al. (2003). Growth rate–stoichiometry couplings in diverse biota. Ecol. Lett. 6, 936–943. doi: 10.1046/j.1461-0248.2003.00518.x
Elser, J. J., Dobberfuhl, D. R., Mackay, N. A., and Schampel, J. H. (1996). Organism size, life history, and N:P stoichiometry. Bioscience 46, 674–684. doi: 10.2307/1312897
Elser, J. J., Elser, M. M., Mackay, N. A., and Carpenter, S. R. (1988). Zooplankton-mediated transitions between N- and P-limited algal growth. Limnol. Oceanogr. 33, 1–14. doi: 10.4319/lo.1988.33.1.0001
Elser, J. J., Fagan, W. F., Denno, R. F., Dobberfuhl, D. R., Folarin, A., Huberty, A., et al. (2000b). Nutritional constraints in terrestrial and freshwater foodwebs. Nature 408, 578–580. doi: 10.1038/35046058
Elser, J. J., Sterner, R. W., Gorokhova, E., Fagan, W. F., Markow, T. A., Cotner, J. B., et al. (2000c). Biological Stoichiometry from genes to ecosystem. Ecol. Lett. 3, 540–550. doi: 10.1046/j.1461-0248.2000.00185.x
Elser, J. J., and Urabe, J. (1999). The Stoichiometry of consumer-driven nutrient recycling: theory, observations, and consequences. Ecology 80, 735–751. doi: 10.1890/0012-9658(1999)080[0735:TSOCDN]2.0.CO;2
Elser, J. J., O'brien, W., Dobberfuhl, D., and Dowling, T. (2000a). The evolution of ecosystem processes: growth rate and elemental stoichiometry of a key herbivore in temperate and arctic habitats. J. Evol. Biol. 13, 845–853. doi: 10.1046/j.1420-9101.2000.00215.x
Fanesi, A., Raven, J. A., and Giordano, M. (2014). Growth rate affects the responses of the green alga Tetraselmis suecica to external perturbations. Plant Cell Environ. 37, 512–519. doi: 10.1111/pce.12176
Fink, P., and Elert, E. V. (2006). Physiological responses to stoichiometric constraints: nutrient limitation and compensatory feeding in a freshwater snail. Oikos 115, 484–494. doi: 10.1111/j.2006.0030-1299.14951.x
Fox, J. W. (2013). The intermediate disturbance hypothesis should be abandoned. Trends Ecol. Evol. 28, 86–92. doi: 10.1016/j.tree.2012.08.014
Frisch, D., Morton, P. K., Chowdhury, P. R., Culver, B. W., Colbourne, J. K., Weider, L. J., et al. (2014). A millennial-scale chronicle of evolutionary responses to cultural eutrophication in Daphnia. Ecol. Lett. 17, 360–368. doi: 10.1111/ele.12237
Frost, P. C., Evans-White, M. A., Finkel, Z. V., Jensen, T. C., and Matzek, V. (2005). Are you what you eat? Physiological constraints on organismal stoichiometry in an elementally imbalanced world. Oikos 109, 18–28. doi: 10.1111/j.0030-1299.2005.14049.x
Gillooly, J. F., Allen, A. P., Brown, J. H., Elser, J. J., Del Rio, C. M., Savage, V. M., et al. (2005). The metabolic basis of whole-organism RNA and phosphorus content. Proc. Natl. Acad. Sci. U.S.A. 102, 11923–11927. doi: 10.1073/pnas.0504756102
Giordano, M. (2013). Homeostasis: an underestimated focal point of ecology and evolution. Plant Sci. 211, 92–101. doi: 10.1016/j.plantsci.2013.07.008
Glibert, P. M. (2016). Margalef revisited: a new phytoplankton mandala incorporating twelve dimensions, including nutritional physiology. Harmful Algae 55, 25–30. doi: 10.1016/j.hal.2016.01.008
Glibert, P. M., Wilkerson, F. P., Dugdale, R. C., Raven, J. A., Dupont, C. L., Leavitt, P. R., et al. (2016). Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr. 61, 165–197. doi: 10.1002/lno.10203
González, A. L., Fariña, J. M., Kay, A. D., Pinto, R., and Marquet, P. A. (2011). Exploring patterns and mechanisms of interspecific and intraspecific variation in body elemental composition of desert consumers. Oikos 120, 1247–1255. doi: 10.1111/j.1600-0706.2010.19151.x
Goos, J. M., French, B. J., Relyea, R. A., Cothran, R. D., and Jeyasingh, P. D. (2014). Sex-specific plasticity in body phosphorus content of Hyalella amphipods. Hydrobiologia 722, 93–102. doi: 10.1007/s10750-013-1682-7
Grizzetti, B., Bouraoui, F., and Aloe, A. (2012). Changes of nitrogen and phosphorus loads to European seas. Glob. Chang. Biol. 18, 769–782. doi: 10.1111/j.1365-2486.2011.02576.x
Grone, A., Swayne, D. E., and Nagode, L. A. (1995). Hypophosphatemic rickets in rheas (Rhea americana). Vet. Pathol. 32, 324–327. doi: 10.1177/030098589503200318
Grover, J. P. (1991). Resource competition in a variable environment: phytoplankton growing according to the variable-internal-stores model. Am. Nat. 138, 811–835. doi: 10.1086/285254
Güsewell, S. (2004). N: P ratios in terrestrial plants: variation and functional significance. New Phytol. 164, 243–266. doi: 10.1111/j.1469-8137.2004.01192.x
Hall, S. R., Leibold, M. A., Lytle, D. A., and Smith, V. H. (2004). Stoichiometry and planktonic grazer composition over gradients of light, nutrients, and predation risk. Ecology 85, 2291–2301. doi: 10.1890/03-0471
Han, W., Fang, J., Guo, D., and Zhang, Y. (2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 168, 377–385. doi: 10.1111/j.1469-8137.2005.01530.x
Hessen, D. O., Elser, J. J., Sterner, R. W., and Urabe, J. (2013). Ecological stoichiometry: an elementary approach using basic principles. Limnol. Oceanogr. 58, 2219–2236. doi: 10.4319/lo.2013.58.6.2219
Hillebrand, H., Steinert, G., Boersma, M., Malzahn, A. M., Meunier, C. L., Plum, C., et al. (2013). Goldman revisited: faster growing phytoplankton has lower N:P and lower stoichiometric flexibility. Limnol. Oceanogr. 58, 2076–2088. doi: 10.4319/lo.2013.58.6.2076
Hood, J. M., and Sterner, R. W. (2014). Carbon and phosphorus linkages in Daphnia growth are determined by growth rate, not species or diet. Funct. Ecol. 28, 1156–1165. doi: 10.1111/1365-2435.12243
Irwin, A. J., Finkel, Z. V., Schofield, O. M., and Falkowski, P. G. (2006). Scaling-up from nutrient physiology to the size-structure of phytoplankton communities. J. Plankton Res. 28, 459–471. doi: 10.1093/plankt/fbi148
Jeyasingh, P. D., Cothran, R. D., and Tobler, M. (2014). Testing the ecological consequences of evolutionary change using elements. Ecol. Evol. 4, 528–538. doi: 10.1002/ece3.950
Jeyasingh, P. D., and Weider, L. J. (2007). Fundamental links between genes and elements: evolutionary implications of ecological stoichiometry. Mol. Ecol. 16, 4649–4661. doi: 10.1111/j.1365-294X.2007.03558.x
Kagata, H., and Ohgushi, T. (2011). Ecosystem consequences of selective feeding of an insect herbivore: palatability–decomposability relationship revisited. Ecol. Entomol. 36, 768–775. doi: 10.1111/j.1365-2311.2011.01327.x
Kay, A. D., Ashton, I. W., Gorokhova, E., Kerkhoff, A. J., Liess, A., and Litchman, E. (2005). Toward a stoichiometric framework for evolutionary biology. Oikos 109, 6–17. doi: 10.1111/j.0030-1299.2005.14048.x
Klausmeier, C. A., Litchman, E., Daufresne, T., and Levin, S. A. (2004). Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton. Nature 429, 171–174. doi: 10.1038/nature02454
Kneitel, J. M., and Chase, J. M. (2004). Trade-offs in community ecology: linking spatial scales and species coexistence. Ecol. Lett. 7, 69–80. doi: 10.1046/j.1461-0248.2003.00551.x
Knoll, L. B., McIntyre, P. B., Vanni, M. J., and Flecker, A. S. (2009). Feedbacks of consumer nutrient recycling on producer biomass and stoichiometry: separating direct and indirect effects. Oikos 118, 1732–1742. doi: 10.1111/j.1600-0706.2009.17367.x
Kremer, C. T., Williams, A. K., Finiguerra, M., Fong, A. A., Kellerman, A., Paver, S. F., et al. (2016). Realizing the potential of trait-based aquatic ecology: new tools and collaborative approaches. Limnol. Oceanogr. 62, 253–271. doi: 10.1002/lno.10392
Leal, M. C., Seehausen, O., and Matthews, B. (2016). The ecology and evolution of stoichiometric phenotypes. Trends Ecol. Evol. 32, 108–117. doi: 10.1016/j.tree.2016.11.006
Leibold, M. A. (1996). A graphical model of keystone predators in food webs: trophic regulation of abundance, incidence, and diversity patterns in communities. Am. Natural. 147, 784–812. doi: 10.1086/285879
Lemoine, N. P., Giery, S. T., and Burkepile, D. E. (2014). Differing nutritional constraints of consumers across ecosystems. Oecologia 174, 1367–1376. doi: 10.1007/s00442-013-2860-z
Li, W., and Stevens, M. H. H. (2012). Fluctuating resource availability increases invasibility in microbial microcosms. Oikos 121, 435–441. doi: 10.1111/j.1600-0706.2011.19762.x
Liess, A., Guo, J., Lind, M. I., and Rowe, O. (2015). Cool tadpoles from Arctic environments waste fewer nutrients–high gross growth efficiencies lead to low consumer-mediated nutrient recycling in the North. J. Anim. Ecol. 84, 1744–1756. doi: 10.1111/1365-2656.12426
Litchman, E., and Klausmeier, C. A. (2008). Trait-based community ecology of phytoplankton. Annu. Rev. Ecol. Evol. Syst. 39, 615–639. doi: 10.1146/annurev.ecolsys.39.110707.173549
Litchman, E., Klausmeier, C. A., and Yoshiyama, K. (2009). Contrasting size evolution in marine and freshwater diatoms. Proc. Natl. Acad. Sci. U.S.A. 106, 2665–2670. doi: 10.1073/pnas.0810891106
Litchman, E., Ohman, M. D., and Kiørboe, T. (2013). Trait-based approaches to zooplankton communities. J. Plankton Res. 35, 473–484. doi: 10.1093/plankt/fbt019
Löder, M. G. J., Meunier, C., Wiltshire, K. H., Boersma, M., and Aberle, N. (2011). The role of ciliates, heterotrophic dinoflagellates and copepods in structuring spring plankton communities at Helgoland Roads, North Sea. Mar. Biol. 158, 1551–1580. doi: 10.1007/s00227-011-1670-2
McGill, B. J., Enquist, B. J., Weiher, E., and Westoby, M. (2006). Rebuilding community ecology from functional traits. Trends Ecol. Evol. 21, 178–185. doi: 10.1016/j.tree.2006.02.002
Méndez, M., and Karlsson, P. S. (2005). Nutrient stoichiometry in Pinguicula vulgaris nutrient availability, plant size, and reproductive status. Ecology 86, 982–991. doi: 10.1890/04-0354
Meunier, C. L., Boersma, M., Wiltshire, K. H., and Malzahn, A. M. (2016a). Zooplankton eat what they need: copepod selective feeding and potential consequences for marine systems Oikos 125, 50–58. doi: 10.1111/oik.02072
Meunier, C. L., Gundale, M. J., Sánchez, I. S., and Liess, A. (2016b). Impact of nitrogen deposition on forest and lake food webs in nitrogen limited environments. Glob. Chang. Biol. 22, 164–179. doi: 10.1111/gcb.12967
Meunier, C. L., Hantzsche, F. M., Cunha-Dupont, A. Ö., Haafke, J., Oppermann, B., Malzahn, A. M., et al. (2012). Intraspecific selectivity, compensatory feeding, and flexible homeostasis in the phagotrophic flagellate Oxyrrhis marina: three ways to handle food quality fluctuations. Hydrobiologia 680, 53–62. doi: 10.1007/s10750-011-0900-4
Meunier, C. L., Malzahn, A. M., and Boersma, M. (2014). A new approach to homeostatic regulation: towards a unified view of physiological and ecological concepts. PLoS ONE 9:e107737. doi: 10.1371/journal.pone.0107737
Montechiaro, F., and Giordano, M. (2010). Compositional homeostasis of the dinoflagellate Protoceratium reticulatum grown at three different pCO2. J. Plant Physiol. 167, 110–113. doi: 10.1016/j.jplph.2009.07.013
Peňuelas, J., Sardans, J., Rivas-Ubach, A., and Janssens, I. A. (2012). The human-induced imbalance between C, N and P in Earth's life system. Glob. Chang. Biol. 18, 3–6. doi: 10.1111/j.1365-2486.2011.02568.x
Perkins, M. C., Woods, H. A., Harrison, J. F., and Elser, J. J. (2004). Dietary phosphorus affects the growth of larval Manduca sexta. Arch. Insect Biochem. Physiol. Behav. 55, 153–168. doi: 10.1002/arch.10133
Persson, J., Fink, P., Goto, A., Hood, J. M., Jonas, J., and Kato, S. (2010). To be or not to be what you eat: regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos 119, 741–751. doi: 10.1111/j.1600-0706.2009.18545.x
Pilati, A., and Vanni, M. J. (2007). Ontogeny, diet shifts, and nutrient stoichiometry in fish. Oikos 116, 1663–1674. doi: 10.1111/j.0030-1299.2007.15970.x
Ploug, H., Stolte, W., Epping, E. H., and Jørgensen, B. B. (1999). Diffusive boundary layers, photosynthesis, and respiration of the colony-forming plankton alga. Phaeocystis sp. Limnol. Oceanogr. 44, 1949–1958. doi: 10.4319/lo.1999.44.8.1949
Price, J. N., and Morgan, J. W. (2007). Vegetation dynamics following resource manipulations in herb-rich woodland. Plant Ecol. 188, 29–37. doi: 10.1007/s11258-006-9145-0
Reichwaldt, E. S. (2008). Food quality influences habitat selection in Daphnia. Freshw. Biol. 53, 872–883. doi: 10.1111/j.1365-2427.2007.01945.x
Reynolds, C. (1988). Functional Morphology and the Adaptive Strategies of Freshwater Phytoplankton. Growth and Reproductive Strategies of Freshwater Phytoplankton. Cambridge: Cambridge University Press.
Reynolds, C. S. (1984). The Ecology of Freshwater Phytoplankton. Cambridge: Cambridge University Press
Reynolds, C. S. (2006). The Ecology of Phytoplankton. Cambridge; New York, NY: Cambridge University Press.
Rhee, G.-Y. (1978). Effects of N:P atomic ratios and nitrate limitation on algal growth, cell composition, and nitrate uptake. Limnol. Oceanogr. 23, 10–25. doi: 10.4319/lo.1978.23.1.0010
Sardans, J., Rivas-Ubach, A., and Peñuelas, J. (2012). The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: a review and perspectives. Biogeochemistry 111, 1–39. doi: 10.1007/s10533-011-9640-9
Shimizu, Y., and Urabe, J. (2008). Regulation of phosphorus stoichiometry and growth rate of consumers: theoretical and experimental analyses with Daphnia. Oecologia 155, 21–31. doi: 10.1007/s00442-007-0896-7
Smayda, T. J. (1997). Harmful algal blooms: their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol. Oceanogr. 42, 1137–1153. doi: 10.4319/lo.1997.42.5_part_2.1137
Snyder, M. N., Small, G. E., and Pringle, C. M. (2015). Diet-switching by omnivorous freshwater shrimp diminishes differences in nutrient recycling rates and body stoichiometry across a food quality gradient. Freshw. Biol. 60, 526–536. doi: 10.1111/fwb.12481
Sommer, U. (1984). The paradox of the plankton: fluctuations of phosphorus availability maintain diversity of phytoplankton in flow-through cultures. Limnol. Oceanogr. 29, 633–636. doi: 10.4319/lo.1984.29.3.0633
Sommer, U., Gliwicz, Z. M., Lampert, W., and Duncan, A. (1986). The PEG-model of seasonal succession of planktonic events in fresh waters. Arch. Hydrobiol. 106, 433–471.
Sperfeld, E., Wagner, N. D., Halvorson, H. M., Malishev, M., and Raubenheimer, D. (2017). Bridging Ecological Stoichiometry and Nutritional Geometry with homeostasis concepts and integrative models of organism nutrition. Funct. Ecol. 31, 286–296. doi: 10.1111/1365-2435.12707
Stelzer, R. S., and Lamberti, G. A. (2002). Ecological stoichiometry in running waters: periphyton chemical composition and snail growth. Ecology 83, 1039–1051. doi: 10.1890/0012-9658(2002)083[1039:ESIRWP]2.0.CO;2
Sterner, R. W. (1990). The ratio of nitrogen to phosphorus resupplied by herbivores: zooplankton and the algal competitive arena. Am. Nat. 136, 209–229. doi: 10.1086/285092
Sterner, R. W. (1995a). “Elemental stoichiometry of species in ecosystems,” in Linking Species and Ecosystems, eds C. G. Jones and J. H. Lawton (New York, NY: Chapman and Hall), 240–252.
Sterner, R. W., and Elser, J. J. (2002). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton; Oxford: Princeton University Press.
Sterner, R. W., Elser, J. J., and Hessen, D. O. (1992). Stoichiometric relationships among producers, consumers and nutrient cycling in pelagic ecosystems. Biogeochemistry 17, 49–67. doi: 10.1007/BF00002759
Sterner, R. W., and Hessen, D. O. (1994). Algal nutrient limitation and the nutrition of aquatic herbivores. Annu. Rev. Ecol. Syst. 25, 1–29. doi: 10.1146/annurev.es.25.110194.000245
Thingstad, T. F., Øvreås, L., Egge, J. K., Løvdal, T., and Heldal, M. (2005). Use of non-limiting substrates to increase size; a generic strategy to simultaneously optimize uptake and minimize predation in pelagic osmotrophs? Ecol. Lett. 8, 675–682. doi: 10.1111/j.1461-0248.2005.00768.x
Tiegs, S. D., Berven, K. A., Carmack, D. J., and Capps, K. A. (2016). Stoichiometric implications of a biphasic life cycle. Oecologia 180, 853–863. doi: 10.1007/s00442-015-3504-2
Tilman, D. (1982). Resource Competition and Community Structure. Princeton, NJ: Princeton University Press.
Van De Waal, D. B., Verschoor, A. M., Verspagen, J. M., Van Donk, E., and Huisman, J. (2010). Climate-driven changes in the ecological stoichiometry of aquatic ecosystems. Front. Ecol. Environ. 8, 145–152. doi: 10.1890/080178
Vanni, M. J. (2002). Nutrient cycling by animals in freshwater ecosystems. Annu. Rev. Ecol. Syst. 33, 341–370. doi: 10.1146/annurev.ecolsys.33.010802.150519
Vanni, M. J., Flecker, A. S., Hood, J. M., and Headworth, J. L. (2002). Stoichiometry of nutrient recycling by vertebrates in a tropical stream: linking species identity and ecosystem processes. Ecol. Lett. 5, 285–293. doi: 10.1046/j.1461-0248.2002.00314.x
Vanni, M. J., and McIntyre, P. B. (2016). Predicting nutrient excretion of aquatic animals with metabolic ecology and ecological stoichiometry: a global synthesis. Ecology 97, 3460–3471. doi: 10.1002/ecy.1582
Vogt, R. J., Beisner, B. E., and Prairie, Y. T. (2010). Functional diversity is positively associated with biomass for lake diatoms. Freshw. Biol. 55, 1636–1646. doi: 10.1111/j.1365-2427.2010.02397.x
Vrede, T., Drakare, S., Eklöv, P., Hein, A., Liess, A., Olsson, J., et al. (2011). Ecological stoichiometry of Eurasian perch – intraspecific variation due to size, habitat and diet. Oikos 120, 886–896. doi: 10.1111/j.1600-0706.2010.18939.x
Wilkinson, D. M. (1999). The disturbing history of intermediate disturbance. Oikos 84, 145–147. doi: 10.2307/3546874
Winder, M., Spaak, P., and Mooij, W. M. (2004). Trade-off in Daphnia habitat selection. Ecology 85, 2027–2036. doi: 10.1890/03-3108
Wirtz, K. (2013). Mechanistic origins of variability in phytoplankton dynamics: Part I: niche formation revealed by a size-based model. Mar. Biol. 160, 2319–2335. doi: 10.1007/s00227-012-2163-7
Woods, H. A., and Wilson, J. K. (2013). An information hypothesis for the evolution of homeostasis. Trends Ecol. Evol. 28, 283–289. doi: 10.1016/j.tree.2012.10.021
Yu, Q., Wilcox, K., Pierre, K. L., Knapp, A. K., Han, X., and Smith, M. D. (2015). Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change. Ecology 96, 2328–2335. doi: 10.1890/14-1897.1
Keywords: food web, biological stoichiometry, functional trait, fitness, trade-off, resource
Citation: Meunier CL, Boersma M, El-Sabaawi R, Halvorson HM, Herstoff EM, Van de Waal DB, Vogt RJ and Litchman E (2017) From Elements to Function: Toward Unifying Ecological Stoichiometry and Trait-Based Ecology. Front. Environ. Sci. 5:18. doi: 10.3389/fenvs.2017.00018
Received: 26 January 2017; Accepted: 19 April 2017;
Published: 08 May 2017.
Edited by:
Michael M. Douglas, University of Western Australia, AustraliaReviewed by:
Wyatt F. Cross, Montana State University, USAJoshua Hamilton, University of Wisconsin-Madison, USA
Copyright © 2017 Meunier, Boersma, El-Sabaawi, Halvorson, Herstoff, Van de Waal, Vogt and Litchman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cédric L. Meunier, Y2VkcmljLm1ldW5pZXJAYXdpLmRl