 Cuihua Huang
Cuihua Huang Fei Peng
Fei Peng Quangang You
Quangang You Jie Liao
Jie Liao Hanchen Duan
Hanchen Duan Tao Wang
Tao Wang Xian Xue
Xian Xue- Drylands Salinization Research Station, Key Laboratory of Desert and Desertification, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou, China
Currently, grazing exclosure is one of the most important grassland management measures for restoring all types of degraded alpine grassland in the Qinghai–Tibetan Plateau (QTP). The most widely distributed grassland ecosystems across the northeastern QTP are the alpine meadow (AM), alpine meadow steppe (AMS), and alpine steppe (AS). However, whether the impacts of fencing on vegetation characteristics and soil properties vary among different grassland types remains poorly understood despite that numerous individual studies have been conducted. This study investigated the vegetation characteristics and soil properties in fenced and grazed AM, AMS, and AS in the northeastern QTP. Grazing exclosure significantly increased the vegetation coverage and Shannon–Wiener diversity index in all the three grasslands. Plant species richness was significantly increased in AM, but there were no significant changes in AMS and AS. Aboveground biomass was significantly increased in AMS and AS but not significant in AM. Increase in the percentage of high-quality forage grasses was only observed in AMS. Fencing significantly decreased the soil bulk density (BD) and significantly increased soil organic carbon (SOC) and total nitrogen at a depth of 0–50 cm in AMS and AS but had no effect in AM. Our results indicate that the use of fencing for restoring degraded AM might not achieve the same expected results as in AS and AMS on the QTP.
Introduction
Land degradation has been increasing at an annual rate of 5–10 million ha and affecting about 1.5 billion people globally (Gisladottir and Stocking, 2005; Ilan and Rattan, 2015). The alpine grassland on the Qinghai–Tibetan Plateau (QTP), occupying over 60% total area of the QTP, is proved to be sensitive to climate change and human activities (Wu et al., 2012; Xue et al., 2015; Bakhshi et al., 2019) and has important functions in protecting the headwaters of major rivers in Asia (Yan and Lu, 2015). However, it has severely degraded since the 1980s (Saito et al., 2009; Li et al., 2013; Zhao et al., 2015; Xue et al., 2017) because of intensification of human activities and climate change (Harris, 2010; Xue et al., 2015; Zeng et al., 2015). The grassland degradation induced 73% reduction in the aboveground biomass, but the amount of poisonous plants almost doubled, which suggests more severe reduction in the palatable grasses. The dramatic reduction in edible biomass accompanied by the sharp increase in livestock number leads to the overgrazing, hence the severe vegetation cover reduction (Wu et al., 2012; Zhao et al., 2015; Lu et al., 2017; Yu et al., 2019; Wang et al., 2020), plant species diversity loss (Chillo et al., 2015), percentage of palatable forage species attenuation (Li et al., 2016), and productivity decrease (Wu et al., 2009). Vegetation degradation will interact with the soil and impose positive feedback to soil degradation (Miao et al., 2015; Tang et al., 2016).
Degraded grassland ecosystems have the capacity for self-recovery if the disturbance ceases for an extended length of time allowing for natural succession (Cheng et al., 2011; Deng et al., 2014). Exclosure is a worldwide management practice, which significantly influences vegetation characteristics (Wu et al., 2009; Deng et al., 2014) and soil properties (Wu et al., 2010a, b; Zhao et al., 2015; Hu et al., 2016), then probably allows for self-recovery of the degradation grassland ecosystem. Realizing the severe alpine grassland degradation and its destructive consequence, the central and local government advocated the use of metal fences in family ranch scale to protect grassland degradation since 2004 in QTP (Yan and Lu, 2015).
The most widely distributed grassland ecosystems across the northeastern QTP are the alpine meadow (AM), alpine meadow steppe (AMS), and alpine steppe (AS) in the northeastern QTP (Wu et al., 2012). AM is a good natural pasture with low layer, soft quality, rich nutrition, strong palatability, and resistance to grazing and trampling. The AM community is simple in structure, not obvious in hierarchy, with dense growth and low plants, sometimes forming a flat planting mat (Wang et al., 2020). An AMS-type rangeland is developed in alpine (or plateau) sub-frigid zones and cold sub-humid regions, with an annual precipitation of 300–400 mm. It is a grassland type mainly composed of hardy perennial arid medium or medium xerophytic herbaceous plants. AS plants are low clustered, with reduced leaf area and shallow roots (Li et al., 2019). The species composition of grassland determines ecosystem stability and resistance to disturbance (Wardle et al., 2000). Different vegetation types have their unique structure composition and stability. Under different environmental conditions, vegetation types with their own unique structural characteristics have different responses to environmental changes (Zhao et al., 2016). So, in different regions or different types of grassland, fencing might result in a wide range of effects on vegetation characteristics and soil properties (Wu et al., 2012; Jing et al., 2014; Cheng et al., 2016). Quantifying the changes in vegetation characteristics and soil properties of different grasslands can help us understand how to carry out land management regimes (Li et al., 2013). Previous studies show different responses of different alpine grasslands to climate warming (Ganjurjav et al., 2016), N addition (Li et al., 2019), and changes in soil properties (Peng et al., 2020a). For example, the plant community of AS shows a stronger association with soil properties than AM alongside degradation (Peng et al., 2020a). Warming did not significantly change the plant composition and species diversity in the AM, but it did cause rapid changes in species diversity (Ganjurjav et al., 2016). Whether the effects of fencing on vegetation and soil properties are consistent among different biomes remains still unclear. To address this scientific gap, we studied the effects of fencing on vegetation and soil properties selected in three typical vegetation types. Each study’s grassland type was regarded as a single data point in paired comparisons of grazed vs. fenced sites and then for analysis on the difference in those responses among three alpine grasslands with the vegetation and soil indicators. The results will inform alpine grassland conservation and sustainable management in the future.
Materials and Methods
Experimental Design
The climate is characterized by strong solar radiation with short, cool summers and long, cold winters in QTP. The growing season of alpine grasslands lasts from May to September (Gao et al., 2013). We carried out the surveys from July to September 2014 on the QTP. In order to avoid the impact of different fencing durations on the ecosystem, we selected three alpine grassland types (AM, AMS, AS), which was fenced off grazing at the same year. Location, climate, and vegetation information of the three sites can be seen in Table 1.

Table 1. Location, climate, and vegetational information of the three sampling sites.
Sampling and Measurements
Vegetation Characteristics
In each site, three plots (30 cm × 30 cm) were selected for vegetation characteristic measurements and soil sampling inside and outside of the fence. A photo was taken for each plot vertically downward by a camera, and the photo was processed by the software CAN-EYE-V6313, developed at the French National Institute of Agricultural Research (INRA) to get the plant coverage (Peng et al., 2018). Plant species identification was done in situ. Unidentified specimens were collected and later identified by plant taxonomists. The total species in each plot were counted after the identification of species, and the frequency of each species and coverage were obtained by using a frame with 100 small quadrats (Peng et al., 2017). At the same time, the height of every species was measured in situ with a ruler. Species richness is the number of species in each plot (Stirling and Wilsey, 2001). The importance value (IV) of each species was derived by averaging the values of relative frequency, relative coverage, and relative height, which is the ratio of the average value of that species to the summed value of all the species in the plot. The richness index (R) and Shannon–Wiener diversity index (H) of the communities were calculated as follows:
where S is the total species numbers of the grassland community, IVi is the IV of a specific species i, and IVtotal is the sum of the IVi values of all the species.
Plant Biomass Measurement
The aboveground parts of plants, including all litter, for each species, were cut, collected, and put into envelops and tagged for each plot. After the dry-up in the air, the plants were separated into high-quality forage grasses (sedge and grass) and forbs (forbs and shrub). After separation, the biomass was dried in an oven at 65°C for 48 h to a constant weight to obtain the aboveground biomass.
The soil cores were extracted at depths of 0–10, 10–20, 20–30, and 30–50 cm in the center of each plot. The samples were immediately placed in a cooler and then transported to the laboratory. In the laboratory, the soil samples were air-dried and crumbled to pass through a 2-mm diameter sieve to remove large particles from the finer soil. Subsequently, fine living roots were hand-picked based on their color and consistency in a distilled water bath (Peng et al., 2018). The picked fine roots were dried at 65°C for 48 h to a constant weight, and the belowground biomass was obtained. The remaining soil was used for soil organic carbon (SOC) and total N measurement in the lab.
Soil Properties
Soil organic carbon was measured by the potassium dichromate oxidation titration method (Walkley, 1947), and total N was measured by the Kjeldahl method (Bremner, 1996) in the Key Laboratory of Desert and Desertification, Chinese Academy of Sciences (CAS). Other soil samples also were taken at depths of 0–10, 10–20, 20–30, and 30–50 cm to measure soil moisture and bulk density (BD). Soil samples were collected and then put in aluminum boxes (volume, 100 cm3), then the weight of boxes and wet soil was measured in situ. The collected samples were transported and then were dried at 105°C for 48 h in an oven to a constant weight. Then, the BD and soil moisture were calculated as follows:
Soil gravimetrical water content was expressed as a percentage of soil water to dry soil weight, and soil moisture was the product of soil gravimetrical water content and BD.
Statistical Analysis
Three-way ANOVA was performed to test the fencing, grassland type, depth, and their interaction effects on vegetation and soil variables. Significant differences were evaluated at the 0.05 level. All statistical analyses were performed using the software program SPSS 19.0 (IBM Corp, 2010). Figures were made by Origin 8.0 (OriginLab Corp, 2007).
Results
Coverage, Biomass, and Diversity
Long-term (8–9 years) fencing significantly increased plant coverage by 32.53, 17.10, and 46.63% in AM, AMS, and AS, respectively (Table 2). There was no significant increase in species richness (R) and Shannon–Wiener diversity index (H) after long-term fencing in all three grasslands (Table 2). The R and H of AM and AMS were significantly higher than that of AS in either enclosed or grazed sites (Tables 2, 3). Fencing only led to a significant increase in aboveground biomass in AMS and AS (Table 2 and Figure 1). The aboveground biomass of AM in fenced plot was not significantly higher than in grazing plot. Increase in percentage of palatable forage was only observed in AMS (Table 2).
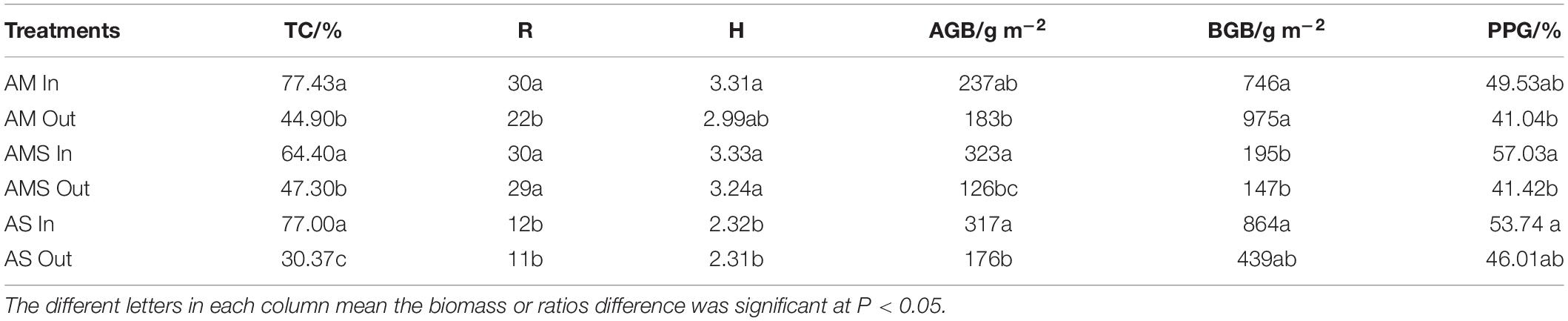
Table 2. Comparisons of total coverage (TC), species richness (R), Shannon–Wiener diversity index (H), aboveground biomass (AGB), belowground biomass (BGB), percentage of palatable grasses (PPG), between enclosed (In), and grazed (Out) sites with the three vegetation types: alpine meadow (AM), alpine meadow steppe (AMS), and alpine steppe (AS).
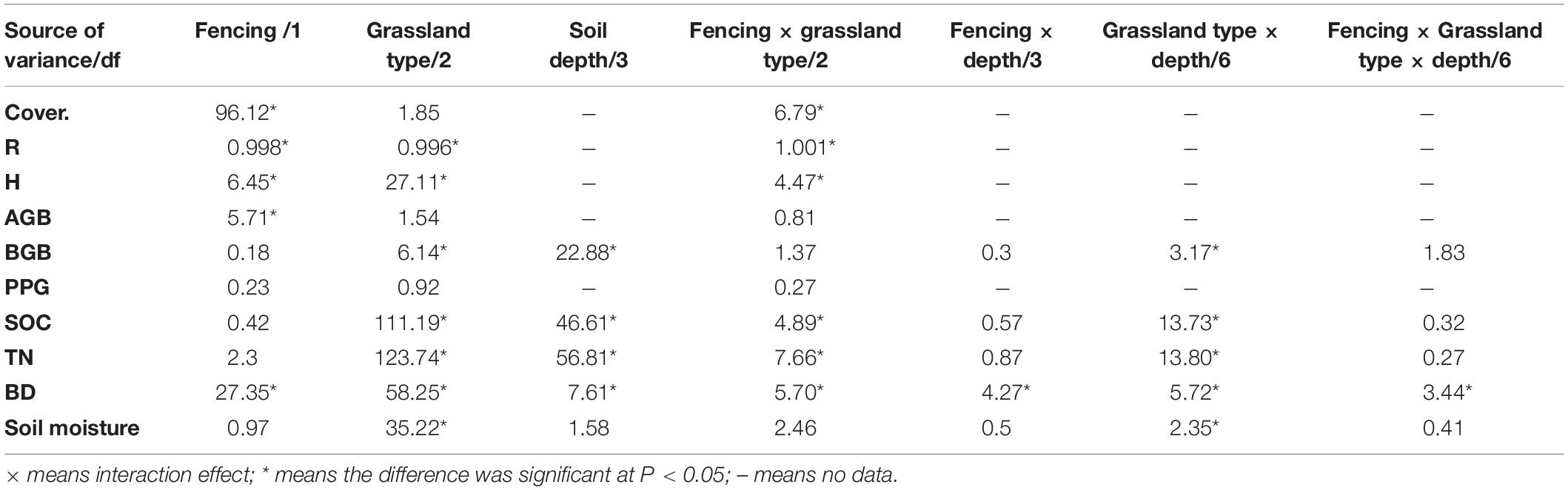
Table 3. Results (F value) of three-way ANOVA analysis about the effect of fencing, depth, grassland type, and interaction of fencing and grassland type, fencing and depth, grassland types and depth, fencing with grassland type and depth on total coverage (Cover.), plant species richness (R), Shannon–Wiener diversity index (H), aboveground biomass (AGB), belowground biomass (BGB), proportion of palatable grasses (PPG), soil organic C (SOC), total nitrogen (TN), soil bulk density (BD), and soil moisture.
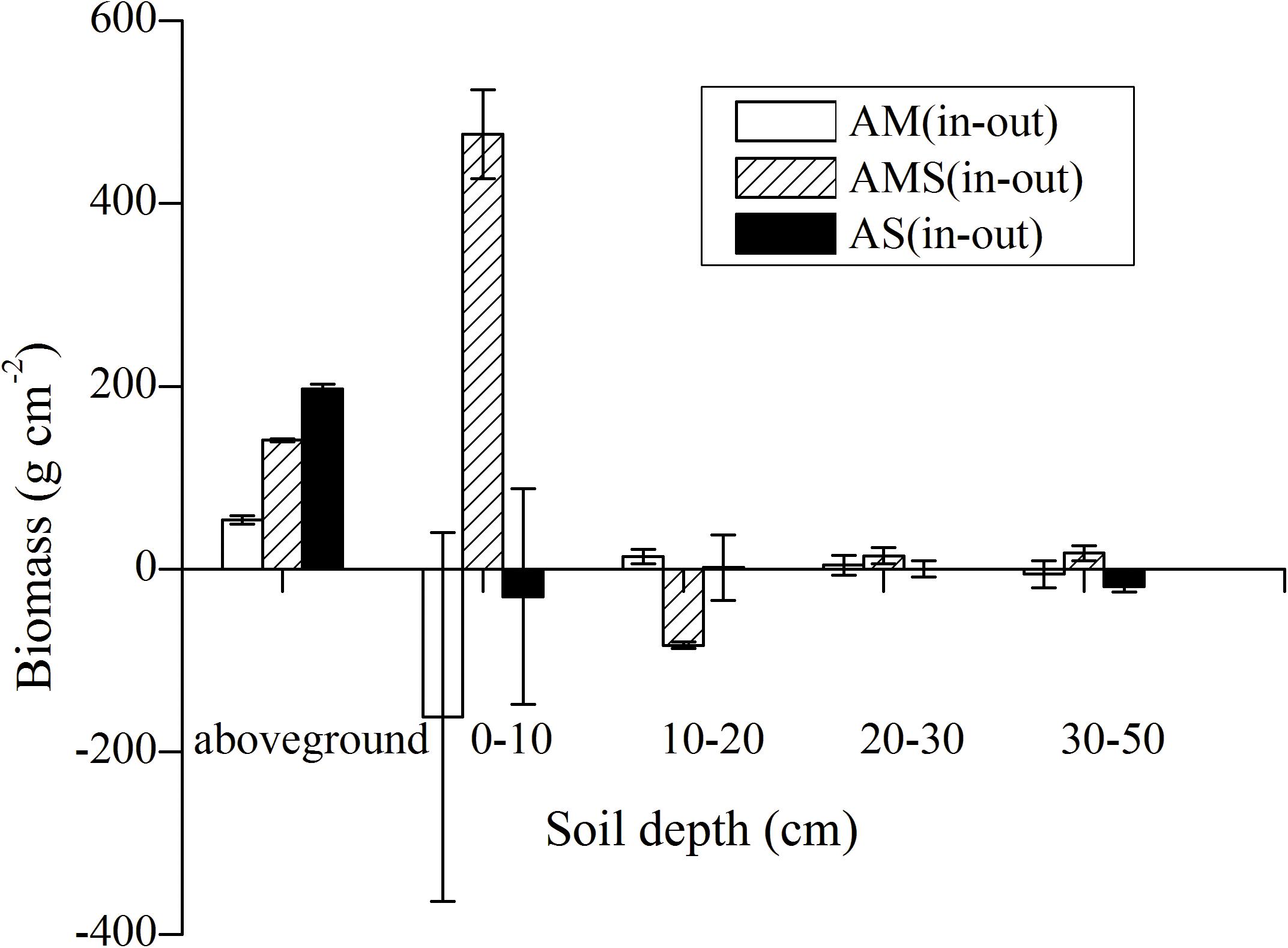
Figure 1. Biomass distribution in the vertical direction between enclosed and grazed sites with three vegetation types: alpine meadow (AM), alpine meadow steppe (AMS), and alpine steppe (AS). Error bars were the standard error of three replicates’ biomass difference between enclosed and grazed area at corresponding layers.
Figure 1 shows the biomass distribution in the vertical direction between enclosed and grazed sites in AM, AMS, and AS after long-term fencing. Aboveground biomass was significantly increased after long-term fencing in the three grassland types (Table 3 and Figure 1). Grassland type and soil depth and the interaction between them significantly affected BGB (Table 3). Statistical results showed that with the increase in altitude, from AS to AMS and AM, the percentage of BGB at 0–10 cm to the total belowground biomass (0–50 cm) was decreased in the enclosed site, which was 84.5, 81.3, and 73.5%, respectively. The trend of the grazed site was opposite that of the enclosed site at 0–10 cm; it was 57.8, 76.9, and 80.7%, in AS to AMS and AM, respectively.
Soil Bulk Density and Moisture
Bulk density (BD) was significantly affected by fencing, grassland type, soil depth, and their interactions (Table 3). With the increase in soil depth, BD was increased in AM both at enclosed and grazed sites (Figure 2), but there was no regular change after long-term fencing in AMS and AS (Figure 2). The BD in grazed sites was significantly higher than that in enclosed sites in AMS and AS at the soil depths of 0–30 cm, but there were no obvious differences at all the three layers in AM (Figure 2).
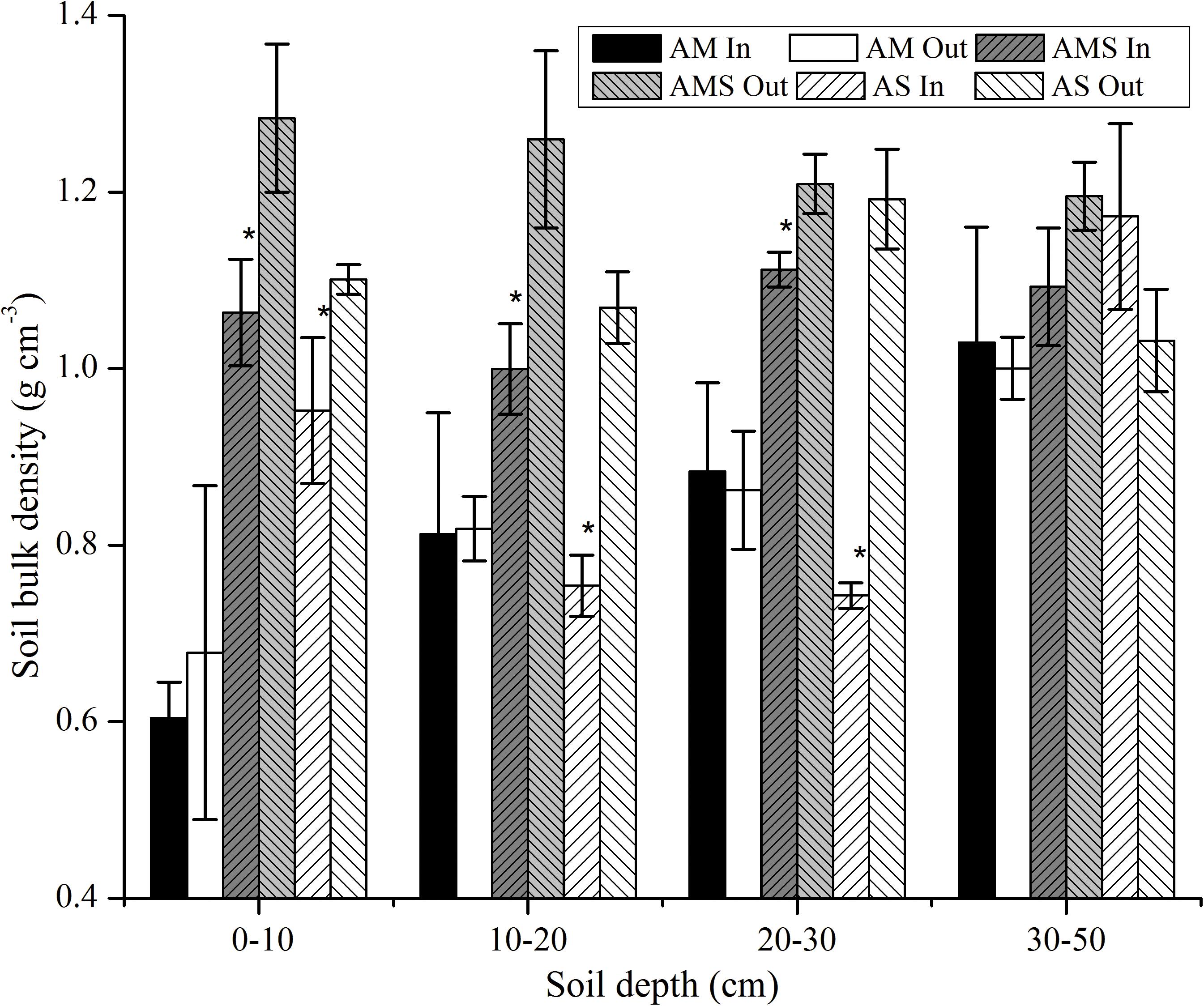
Figure 2. Soil bulk density (BD) in the vertical direction between enclosed (In) and grazed (Out) sites with three vegetation types: alpine meadow (AM), alpine meadow steppe (AMS), and alpine steppe (AS). Error bars were the standard error of three replicates BD between enclosed and grazed sites. *Mean of the change of BD was significant at P < 0.05.
Grassland type, soil depth, and their interactions significantly affected soil moisture (Table 3). Soil moisture was higher in AM and AMS than in AS at all the soil depths (Figure 3). Fencing increased the soil moisture at the depth 0–20 cm in AM, and it showed no obvious effects at the depth 20–50 cm (Figure 3). Soil moisture was decreased at all depths in AMS and AS after long-term fencing, but it was only significant at the depth 0–10 and 20–30 cm in AS (Figure 3).
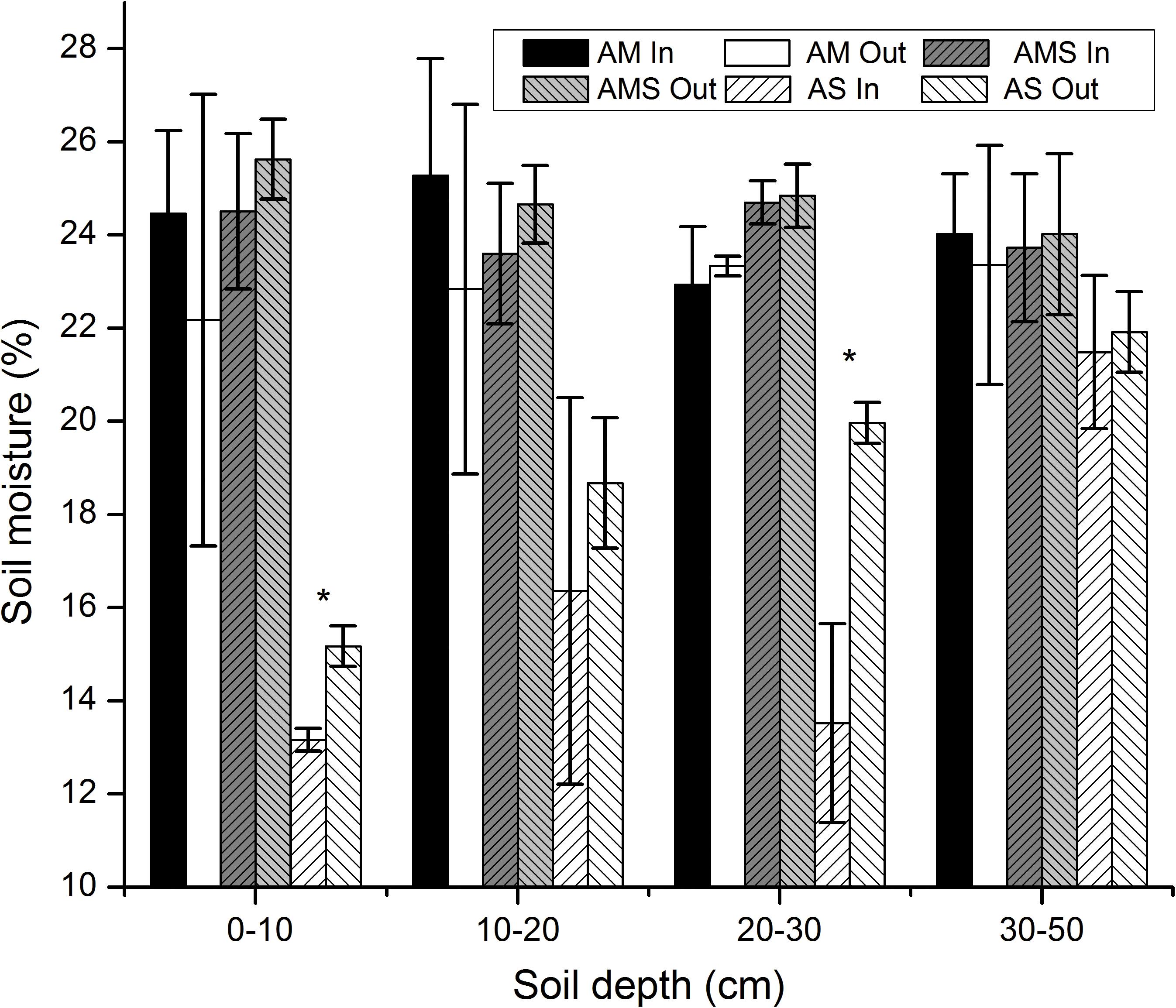
Figure 3. Soil moisture in the vertical direction between enclosed (In) and grazed (Out) sites with three vegetation types: alpine meadow (AM), alpine meadow steppe (AMS), and alpine steppe (AS). Error bars were the standard error of three replicates soil moisture between enclosed and grazed sites; *Mean of the change of soil moisture was significant at P < 0.05.
Soil Organic C and Total Nitrogen
With the increase in soil depth, SOC was decreased in all alpine grassland types in either enclosed sites or grazed sites (Figure 4). The SOC content varied remarkably among different grassland types (Table 3 and Figure 4). The SOC was higher in AM than in AMS and AS in either enclosed or grazed sites at the depth 0–30 cm (Figure 4). Long-term fencing did not have a positive impact on SOC in AM (Figure 4). The SOC increased at all depths in AMS and AS after long-term fencing. The increase was only significant in AMS (Figure 4). The results of three-way ANOVA analysis showed that grassland type, soil depth, fencing, and their interactions significantly affected SOC. The total soil nitrogen (TN) has a similar pattern with SOC, and their correlation coefficient was 0.99.
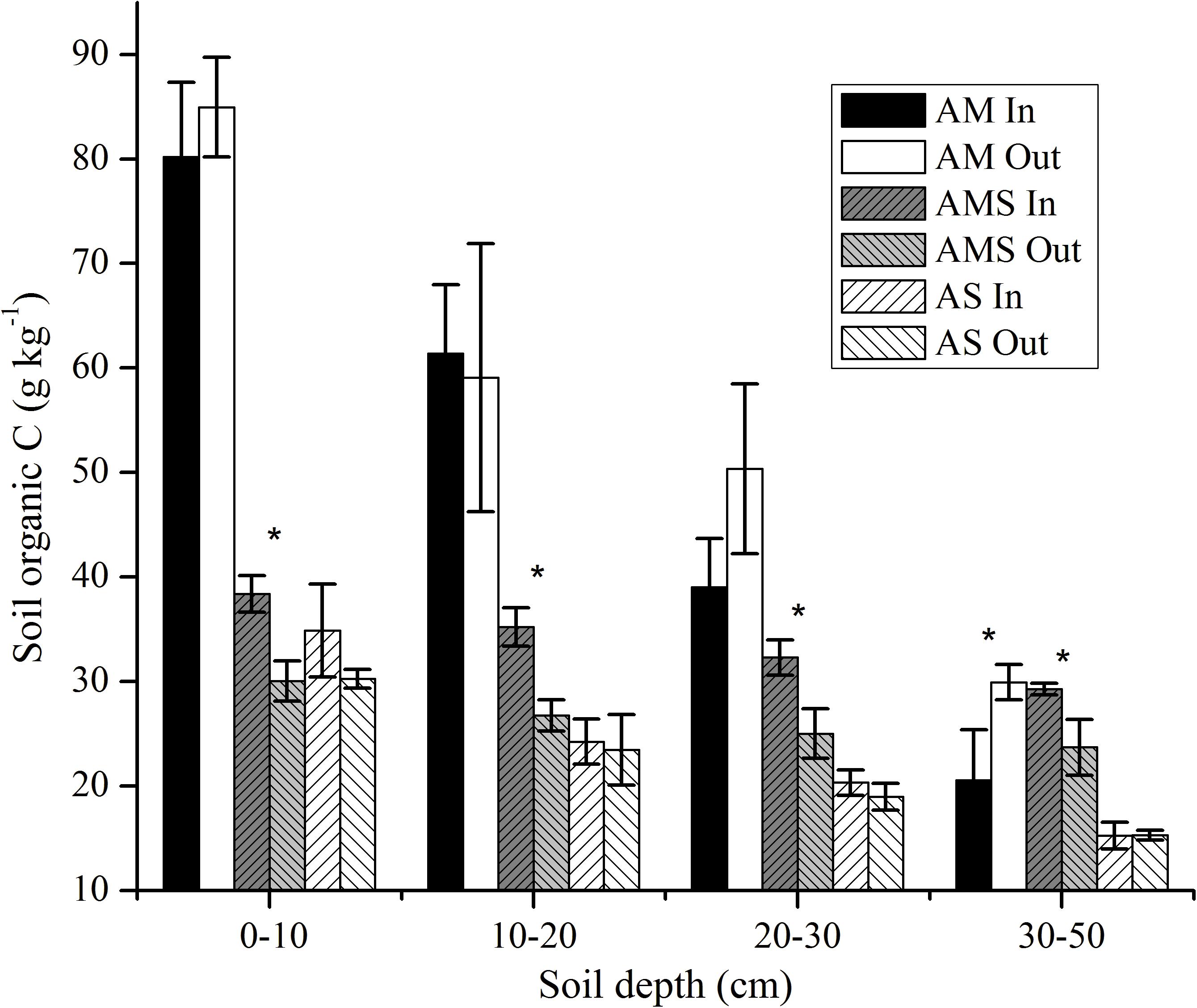
Figure 4. Soil organic C in the vertical direction between enclosed (In) and grazed (Out) sites with three vegetation types: alpine meadow (AM), alpine meadow steppe (AMS), and alpine steppe (AS). Error bars were the standard error of three replicates soil organic C between enclosed and grazed sites. *Mean the change of SOC difference was significant at P < 0.05.
Discussion
Vegetation Characteristics
Fencing has a significant effect on plant community structure and composition, which will have feedbacks on ecosystem productivity (Polley et al., 2014). Due to the complexity of the growing environment, plants are subjected to different degrees and types of external interference in each stage of plant growth, but plants always adjust their growth strategies through certain traits, such as plant height, leaf size, leaf shape, tiller quantity, biomass allocation (Wu et al., 2009; Li et al., 2019), etc., to ensure the successful completion of their life history. Exclosure is a disturbance to plant growth, and plant community characteristics may change accordingly (Wu et al., 2009; Miao et al., 2015).
Biomass is the basis of the flow of matter and energy in an ecosystem (Yan and Lu, 2015), which is an important quantitative feature of the plant community and can directly reflect the material production of producers in the ecosystem (Spring et al., 1996; Zeng et al., 2015). The AGB of different types of grassland increased to a great extent (Table 2) after fencing, which agrees with the results of many studies (Wu et al., 2009; Deng et al., 2014; Zeng et al., 2015; Cheng et al., 2016; Li et al., 2017). The main reason was that foraging was prohibited inside the fences. AGB was significantly increased after long-term exclosure in AMS and AS but not significant in AM. Belowground biomass showed complex responses (Table 2) (Milchunas and Lauenroth, 1993; Frank et al., 2002; Miehe et al., 2019). The BGB at 0–10 cm inside fences was lower than that of the corresponding grazed area in AMS and AS, but in AM it is higher (Figure 1). Because of the rapid increase of AGB in AS after fencing (Liu et al., 2018), most of the soil nutrients are consumed aboveground, resulting in relatively slow root growth. With the increase in altitude, from AS to AMS and AM, the percentages of 0–10 cm to the total BGB (0–50 cm) were decreased in enclosed sites. The trend of grazed sites was opposite with that in enclosed sites at 0–10 cm. This is because AS recovered at the fastest speed, followed by AMA, and AM at the slowest speed. More than 80% of the species in AM are grasses and sedges (Peng et al., 2020b). Sedge and graminoid species have highly branched fibrous root systems that are mainly distributed near the soil surface, and this leads to a rapid increase near the soil surface biomass of the grasslands during vegetation restoration (Wang et al., 2014). The vegetation composition of AM is dominated by forbs with a deep root system (Liu et al., 2018), and then the AGB increase after fencing is dominated by deep layer (Peng et al., 2020a). In high altitude, the constructive species were rhizome grass which the roots distribute mainly in the upper layer of soil in the enclosed site. With the decrease in altitude, from AM to AMS and AS, the dense cluster type grass was increased, which has more distribution of roots at the depth below 10 cm in the enclosed site (Figure 1). Long-term fencing increased AGB and improved the grassland quality especially in AS with lower altitude (Table 3). These results support the viewpoint that with the fencing the distribution of biomass in the vertical direction was changed: a part of biomass “transfer” from belowground to aboveground.
Communities responded to environmental change by altering the functional characters of some dominant species (Macgillivray et al., 1995; Bakhshi et al., 2019); for example, palatable grasses have greater competitive ability than unpalatable grasses and show an increase (Table 2) after long-term fencing (Wu et al., 2009; Deng et al., 2014). Vegetation coverage is an important indicator of land surface vegetation and ecosystem environmental change. The total coverage was significantly increased after long-term fencing in three different alpine grasslands (Tables 2, 3), which were consistent with many other fencing experiment results (Cheng et al., 2011; Mekuria and Veldkamp, 2012). The results support the viewpoint that fencing is a simple and effective measure for restoring degraded grasslands (Wu et al., 2009; Golodets et al., 2010; Deng et al., 2014; Zhao et al., 2015). The species richness (R) in AM (30) and AMS (30) was than twice in AS (12) in an enclosed area, and there was similarity in the grazed area (Table 2). It is similar with the results of Wu et al. (2012), which showed that the mean S (R = S) in AM (23.30) was nearly twice as much as in AS (11.80). Previous research has suggested that fencing may improve species richness and diversity index across the QTP (Wu et al., 2012), and our results are similar (Table 2). Increasing ranges of species richness and diversity index were bigger in AM than in AMS and AS (Table 2). Some experiment results also indicated that fencing might not necessarily increase species richness and diversity index even to decline in species richness and diversity index (Shi et al., 2010; Schultz et al., 2011; Bakhshi et al., 2019) due to differences in the competitive dominance of species (Tilman et al., 1997) and different fencing times (Deng et al., 2014). There is still no consensus on how long-term fencing affects plant species richness and diversity (Zervas, 1998; Schultz et al., 2011; Wu et al., 2012).
The high diversity makes AM more stable and resistant to perturbations than steppes (Kuhsel and Bluthgen, 2015). Species-rich plant communities are relatively more resistant to change in management regimes (Klimes et al., 2013). The sparser canopy of AS relative to that of AM (Zhu et al., 2015) also suggests much easier recruitment and settlement capabilities for grasses. Thus, long-term fencing likely promoted the recruitment and settlement of Stipa species and allowed AS communities to cumulatively benefit from natural conditions. While AM was less sensitive to fencing than AMS and AS (Figure 2), a more nuanced management regime should thus be considered in the rehabilitation of AM.
Soil Properties
Soil properties, such as BD, moisture SOC, and TN, directly regulate plant growth (Ganjurjav et al., 2016). BD was found to be lower in fenced grassland compared to grazing grassland. The reasons included the elimination of soil trampling by livestock, an increase in root biomass accumulation (Yuan et al., 2012), high soil silt and clay content, and presence of extensive shallow root systems in fenced areas (Su et al., 2005). Our results have a similar trend in AMS and AS, but there was almost no change in AM after long-term fencing (Table 3 and Figure 2). The variation of BD of different alpine grassland types was inconsistent, which was mainly caused by the inconsistent increase in BGB of different alpine grassland types after the fencing. The variation of BGB and SOC confirmed this opinion (Figures 1, 4). Long-time fencing leads to the increase in vegetation coverage, which can improve topsoil microhabitats and moisture then prevent the grassland from degradation in alpine grassland (Wu et al., 2011; Deng et al., 2014). On the other hand, increasing coverage also has stronger transpiration, which leads to a decrease in soil moisture. Our results show that soil moisture at the depth of 0–20 cm in AM was increased but in AMS and AS it was decreased at all soil depths (Figure 3). Although the coverage of all the three grasslands was increased after fencing (Table 2), the transpiration of AMS and AS was stronger than that of AM because of the large leaf area (Li et al., 2020). The stronger transpiration leads to higher water consumption, which decreased the soil moisture in AMS and AS (Figure 3).
Soil organic matter is widely recognized to be an important aspect of soil fertility, fulfilling various functions such as improving soil structure, aggregate stability, and water-holding capacity. The SOC at the 0–50 cm depth after long-term fencing in the AS and AMS was increased, but there was no obvious regular change pattern in AM (Figure 4). These were consistent with the results of other researches (Wu et al., 2010a; Zhao et al., 2015; Deng and Shangguan, 2017). Vesterdal et al. (2002) thought that the change in land use from grazing to fencing means that the annual cycle of plants was changed by the much longer grassland cycle. Consequently, this enables the development of high net primary productivity and reduces the degree of soil disturbance, leading to increased SOC fractions. TN has a similar, varying pattern with that of SOC (Yu et al., 2019).
As shown throughout this study, the community characteristics and soil properties have different changes after long-time fencing among three different alpine grasslands in northeast QTP. The vegetation coverage, AGB, plant species richness, Shannon–Wiener Diversity index, and proportion of palatable forage all improved in three alpine grasslands after long-term fencing. With the increase in altitude, from AS to AMS and AM, the percentage BGB at 0–10 cm to the total at 0–50 cm was decreased in the enclosed site, and the change in trend of the grazed site was opposite with that in the enclosed site. BD was obviously decreased in AMS and AS, but there was almost no change in AM after long-term fencing. Fencing increased the SOC and TN at the depth of 0–50 cm in the AS and AMS, but there was no obvious regular change pattern in AM. According to the results, AM was less sensitive to fencing among the three alpine grasslands, so a more nuanced management regime should be considered in the rehabilitation of AM. The similar countermeasures to prevent further degradation and restoration activities of all three alpine grasslands would lead to the failure of some ecological projects.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
CH contributed to conceptualization, methodology, data curation, and writing – original draft preparation. FP contributed to data curation, investigation, and editing. QY, JL, and HD contributed to data curation and investigation. TW contributed to conceptualization. XX contributed to supervision, funding acquisition, and writing – reviewing and editing. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by funds from the National Key R&D Program of China (2016YFC0501803) and the National Natural Science Foundation of China (41771233).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer XL declared a past co-authorship with one of the authors FP to the handling editor.
Acknowledgments
The authors wish to express their gratitude to the reviewers and editor for their time and effort.
References
Bakhshi, J., Javadi, S. A., Tavili, A., and Arzani, H. (2019). Study on the effects of different levels of grazing and exclosure on vegetation and soil properties in semi-arid rangelands of Iran. Acta Ecol. Sin. doi: 10.1016/j.chnaes.2019.07.003
Bremner, J. M. (1996). “Nitrogen-total,” in Methods of Soil Analysis, Part 3, SSSA Book Series: 5, ed. D. L. Sparks (Madison: America Society of Agronomy), 1085–1121.
Cheng, J., Wu, G. L., Zhao, L. P., Li, Y., Li, W., and Cheng, J. M. (2011). Cumulative effects of 20-year exclusion of livestock grazing on above- and belowground biomass of typical steppe communities in arid areas of the Loess Plateau, China. Plant Soil Environ. 57, 40–44. doi: 10.17221/153/2010-PSE
Cheng, J. M., Jing, G. H., Wei, L., and Jing, Z. B. (2016). Long-term grazing exclusion effects on vegetation characteristics, soil properties and bacterial communities in the semi-arid grasslands of China. Ecol. Eng. 97, 170–178. doi: 10.1016/j.ecoleng.2016.09.003
Chillo, V., Ojeda, R. A., Anand, M., and Reynolds, J. F. (2015). A novel approach to assess livestock management effects on biodiversity of drylands. Ecol. Indic. 50, 69–78. doi: 10.1016/j.ecolind.2014.10.009
Deng, L., and Shangguan, Z. P. (2017). Afforestation drives soil carbon, and nitrogen changes in China. Land Degrad. Dev. 28, 151–165. doi: 10.1002/ldr.2537
Deng, L., Zhang, Z. N., and Shangguan, Z. P. (2014). Long-term fencing effects on plant diversity and soil properties in China. Soil Till. Res. 137, 7–15. doi: 10.1016/j.still.2013.11.002
Frank, D. A., Kuns, M. M., and Guido, D. R. (2002). Consumer control of grassland plant production. Ecology 83, 602–606. doi: 10.1890/0012-96582002083
Ganjurjav, H., Gao, Q., Gornish, E. S., Schwartz, M. W., Liang, Y., Cao, X., et al. (2016). Differential response of alpine steppe and alpine meadow to climate warming in the central Qinghai-Tibetan Plateau. Agric. For. Meteorol. 223, 233–240. doi: 10.1016/j.agrformet.2016.03.017
Gao, Y. H., Zhou, X., Wang, Q., Wang, C. Z., Zhan, Z. M., Chen, L. F., et al. (2013). Vegetation net primary productivity and its response to climate change during 2001-2008 in the Tibetan Plateau. Sci. Total Environ. 444, 356–362. doi: 10.1016/j.scitotenv.2012.12.014
Gisladottir, G., and Stocking, M. (2005). Land degradation control and its global environmental benefits. Land Degrad. Dev. 16, 99–112. doi: 10.1002/ldr.687
Golodets, C., Kigel, J., and Sternberg, M. (2010). Recovery of plant species composition and ecosystem function after cessation of grazing in a Mediterranean grassland. Plant Soil 329, 365–378. doi: 10.1007/s11104-009-0164-1
Harris, R. B. (2010). Range land degradation on the Qinghai-Tibetan plateau: a review of the evidence of its magnitude and causes. J. Arid Environ. 74, 1–12. doi: 10.1016/j.jaridenv.2009.06.014
Hu, Z. M., Li, S. G., Guo, Q., Niu, S. L., He, N. P., Li, L. H., et al. (2016). A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China. Glob. Chang. Biol. 22, 1385–1393. doi: 10.1111/gcb.13133
Ilan, S., and Rattan, L. (2015). Achieving zero net land degradation: challenges and opportunities. J. Arid Environ. 112, 45–51. doi: 10.1016/j.jaridenv.2014.01.016
Jing, Z. B., Cheng, J. M., Su, J. S., Bai, Y., and Jin, J. W. (2014). Changes in plant community composition and soil properties under 3-decade grazing exclusion in semiarid grassland. Ecol. Eng. 64, 171–178. doi: 10.1016/j.ecoleng.2013.12.023
Klimes, L., Hajek, M., Mudrak, O., Dancak, M., Preislerova, Z., Hajkova, P., et al. (2013). Effects of changes in management on resistance and resilience in three grassland communities. Appl. Veg. Sci. 16, 640–649. doi: 10.1111/avsc.12032
Kuhsel, S., and Bluthgen, N. (2015). High diversity stabilizes the thermal resilience of pollinator communities in intensively managed grasslands. Nat. Commun. 6:7989. doi: 10.1038/Ncomms8989
Li, F., Zeng, Y., Luo, J. H., Ma, R. H., and Wu, B. F. (2016). Modeling grassland aboveground biomass using a pure vegetation index. Ecol. Indic. 62, 279–288. doi: 10.1016/j.ecolind.2015.11.005
Li, L. J., Song, X. Y., Xia, L., Fu, N., Feng, D., Li, H. Y., et al. (2020). Modelling the effects of climate change on transpiration and evaporation in natural and constructed grasslands in the semi-arid Loess Plateau, China. Agric. Ecosyst. Environ. 302:107077. doi: 10.1016/j.agee.2020.107077
Li, S., Dong, S., Shen, H., Han, Y., Zhang, J., Xu, Y., et al. (2019). Different responses of multifaceted plant diversities of alpine meadow and alpine steppe to nitrogen addition gradients on Qinghai-Tibetan Plateau. Sci. Total Environ. 688, 1405–1412. doi: 10.1016/j.scitotenv.2019.06.211
Li, W., Cao, W. X., Wang, J. L., Li, X. L., Xu, C. L., and Shi, S. L. (2017). Effects of grazing regime on vegetation structure, productivity, soil quality, carbon and nitrogen storage of alpine meadow on the Qinghai-Tibetan Plateau. Ecol. Eng. 98, 123–133. doi: 10.1016/j.ecoleng.2016.10.026
Li, X. L., Gao, J., Brierley, G., Qiao, Y. M., Zhang, J., and Yang, Y. W. (2013). Rangeland degradation on the Qinghai-Tibet plateau: implications for rehabilitation. Land Degrad. Dev. 24, 72–80. doi: 10.1002/ldr.1108
Liu, H., Mi, Z., Lin, L., Wang, Y., Zhang, Z., Zhang, F., et al. (2018). Shifting plant species composition in response to climate change stabilizes grassland primary production. Proc. Natl. Acad. Sci. U.S.A. 115, 4051–4056.
Lu, X. Y., Kelsey, K. C., Yan, Y., Sun, J., Wang, X. D., Cheng, G. W., et al. (2017). Effects of grazing on ecosystem structure and function of alpine grasslands in Qinghai-Tibetan Plateau: a synthesis. Ecosphere 8:e0165610. doi: 10.1002/ecs2.1656
Macgillivray, C. W., Grime, J. P., Band, S. R., Booth, R. E., Campbell, B., Hendry, G. A. F., et al. (1995). Testing predictions of the resistance and resilience of vegetation subjected to extreme events. Funct. Ecol. 9, 640–649. doi: 10.2307/2390156
Mekuria, W., and Veldkamp, E. (2012). Restoration of native vegetation following exclosure establishment on communal grazing lands in Tigray, Ethiopia. Appl. Veg. Sci. 15, 71–83. doi: 10.1111/j.1654-109X.2011.01145.x
Miao, R. H., Jiang, D. M., Musa, A., Zhou, Q. L., Guo, M. X., and Wang, Y. C. (2015). Effectiveness of shrub planting and grazing exclusion on degraded sandy grassland restoration in Horqin sandy land in Inner Mongolia. Ecol. Eng. 74, 164–173. doi: 10.1016/j.ecoleng.2014.10.004
Miehe, G., Schleuss, P. M., Seeber, E., Babel, W., Biermann, T., Braendle, M., et al. (2019). The Kobresia pygmaea ecosystem of the Tibetan highlands - Origin, functioning and degradation of the world’s largest pastoral alpine ecosystem Kobresia pastures of Tibet. SCI Total Environ. 648, 754–771. doi: 10.1016/j.scitotenv.2018.08.164
Milchunas, D. G., and Lauenroth, W. K. (1993). Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol. Monogr. 63, 327–366. doi: 10.2307/2937150
OriginLab Corp (2007). The Data Analysis and Graphing Workspace. Northampton, MA: OriginLab Corporation.
Peng, F., Xue, X., Li, C., Lai, Z., Sun, J., Tsubo, L., et al. (2020a). Plant community of alpine steppe shows stronger association with soil properties than alpine meadow alongside degradation. Sci. Total Environ. 733:139048. doi: 10.1016/j.scitotenv.2020.139048
Peng, F., Xue, X., You, Q., Sun, J., Zhou, J., Wang, T., et al. (2020b). Change in the trade-off between aboveground and belowground biomass of alpine grassland: implications for the land degradation process. Land Degrad. Dev. 31, 105–117. doi: 10.1002/ldr.3432
Peng, F., Xue, X., Xu, M. H., You, Q. G., Jian, G., and Ma, S. X. (2017). Warming-induced shift towards forbs and grasses and its relation to the carbon sequestration in an alpine meadow. Environ. Res. Lett. 12:aa6508. doi: 10.1088/1748-9326/Aa6508
Peng, F., Xue, X., You, Q. G., Huang, C. H., Dong, S. Y., Liao, J., et al. (2018). Changes of soil properties regulate the soil organic carbon loss with grassland degradation on the Qinghai-Tibet Plateau. Ecol. Indic. 93, 572–580. doi: 10.1016/j.ecolind.2018.05.047
Polley, H. W., Derner, J. D., Jackson, R. B., Wilsey, B. J., and Fay, P. A. (2014). Impacts of climate change drivers on C4 grassland productivity: scaling driver effects through the plant community. J. Exp. Bot. 65, 3415–3424. doi: 10.1093/jxb/eru009
Saito, M., Kato, T., and Tang, Y. (2009). Temperature controls ecosystem CO2 exchange of an alpine meadow on the northeastern Tibetan Plateau. Glob. Chang. Biol. 15, 221–228. doi: 10.1111/j.1365-2486.2008.01713.x
Schultz, N. L., Morgan, J. W., and Lunt, I. D. (2011). Effects of grazing exclusion on plant species richness and phytomass accumulation vary across a regional productivity gradient. J. Veg. Sci. 22, 130–142. doi: 10.1111/j.1654-1103.2010.01235.x
Shi, F. S., Chen, H., Wu, Y., and Wu, N. (2010). Effects of livestock exclusion on vegetation and soil properties under two topographic habitats in an alpine meadow on the Eastern Qinghai-Tibetan Plateau. Pol. J. Ecol. 58, 125–133. doi: 10.1017/S0032247409008626
Spring, G. M., Priestman, G. H., and Grime, J. P. (1996). A new field technique for elevating carbon dioxide levels in climate change experiments. Funct. Ecol. 10, 541–545. doi: 10.2307/2389948
Stirling, G., and Wilsey, B. (2001). Empirical relationships between species richness, evenness, and proportional diversity. Am. Nat. 158, 286–299. doi: 10.1086/321317
Su, Y. Z., Li, Y. L., Cui, H. Y., and Zhao, W. Z. (2005). Influences of continuous grazing and livestock exclusion on soil properties in a degraded sandy grassland, Inner Mongolia, northern China. Catena 59, 267–278. doi: 10.1016/j.catena.2004.09.00
Tang, J., Davy, A. J., Jiang, D. M., Musa, A., Wu, D. F., Wang, Y. C., et al. (2016). Effects of excluding grazing on the vegetation and soils of degraded sparse-elm grassland in the Horqin Sandy Land, China. Agric. Ecosyst. Environ. 235, 340–348. doi: 10.1016/j.agee.2016.11.005
Tilman, D., Knops, J., Wedin, D., Reich, P., Ritchie, M., and Siemann, E. (1997). The influence of functional diversity and composition on ecosystem processes. Science 277, 1300–1302. doi: 10.1126/science.277.5330.1300
Vesterdal, L., Ritter, E., and Gundersen, P. (2002). Change in soil organic carbon following afforestation of former arable land. For. Ecol. Manag. 169, 137–147. doi: 10.1016/S0378-1127(02)00304-3
Walkley, A. (1947). A critical examination of a rapid method for determining organic carbon in soils - effect of variations in digestion conditions and of inorganic soil constituents. Soil Sci. 63, 251–264. doi: 10.1097/00010694-194704000-00001
Wang, J., Wang, X. T., Liu, G. B., Wang, G. L., Wu, Y., and Zhang, C. (2020). Fencing as an effective approach for restoration of alpine meadows: evidence from nutrient limitation of soil microbes. Geoderma 363:148. doi: 10.1016/j.geoderma.2019.114148
Wang, X., Dong, S., Yang, B., Li, Y., and Su, X. (2014). The effects of grassland degradation on plant diversity, primary productivity, and soil fertility in the alpine region of Asia’s headwaters. Environ. Monit. Assess. 186, 6903– 6917.
Wardle, D. A., Bonner, K. I., and Barker, G. M. (2000). Stability of ecosystem properties inresponse to above-ground functional group richness and composition. Oikos 89, 11–23. doi: 10.1034/j.1600-0706.2000.890102.x
Wu, G. L., Du, G. Z., Liu, Z. H., and Thirgood, S. (2009). Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau. Plant Soil 319, 115–126. doi: 10.1007/s11104-008-9854-3
Wu, G. L., Li, W., Zhao, L. P., and Shi, Z. H. (2011). Artificial management improves soil moisture, C, N and Pin an alpine sandy meadow of western China. Pedosphere 21, 407–412. doi: 10.1016/S1002-0160(11)60142-2
Wu, G. L., Liu, Z. H., Zhang, L., Chen, J. M., and Hu, T. M. (2010a). Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China. Plant Soil 332, 331–337. doi: 10.1007/s11104-010-0299-0
Wu, G. L., Liu, Z. H., Zhang, L., Hu, T. M., and Chen, J. M. (2010b). Effects of artificial grassland establishment on soil nutrients and carbon properties in a black-soil-type degraded grassland. Plant Soil 333, 469–479. doi: 10.1007/s11104-010-0363-9
Wu, J. S., Zhang, X. Z., Shen, Z. X., Shi, P. L., Yu, C. Q., Song, M. H., et al. (2012). Species richness and diversity of alpine grasslands on the Northern Tibetan Plateau: effects of grazing exclusion and growing season precipitation. J. Resour. Ecol. 3, 236–242. doi: 10.5814/j.issn.1674-764x
Xue, X., Peng, F., You, Q. G., Xu, M. H., and Dong, S. Y. (2015). Belowground carbon responses to experimental warming regulated by soil moisture change in an alpine ecosystem of the Qinghai-Tibet Plateau. Ecol. Evol. 5, 4063–4078. doi: 10.1002/ece3.1685
Xue, X., You, Q. G., Peng, F., Dong, S. Y., and Duan, H. C. (2017). Experimental warming aggravates degradation-induced topsoil drought in alpine meadows of the Qinghai-Tibetan Plateau. Land Degrad. Dev. 28, 2343–2353. doi: 10.1002/ldr.2763
Yan, Y., and Lu, X. Y. (2015). Is grazing exclusion effective in restoring vegetation in degraded alpine grasslands in Tibet, China? Peer J. 3:e1020. doi: 10.7717/peerj.1020
Yu, L. F., Chen, Y., Sun, W. J., and Huang, Y. (2019). Effects of grazing exclusion on soil carbon dynamics in alpine grasslands of the Tibetan Plateau. Geoderma 353, 133–143. doi: 10.1016/j.geoderma.2019.06.036
Yuan, J. Y., Ouyang, Z. Y., Zheng, H., and Xu, W. H. (2012). Effects of different grassland restoration approaches on soil properties in the southeastern Horqin sandy land, northern China. Appl. Soil Ecol. 61, 34–39. doi: 10.1016/j.apsoil.2012.04.003
Zeng, C. X., Wu, J. S., and Zhang, X. Z. (2015). Effects of grazing on above- vs. below-ground biomass allocation of alpine Grasslands on the Northern Tibetan Plateau. PLoS One 10:e0135173. doi: 10.1371/journal.pone.0135173
Zervas, G. (1998). Quantifying and optimizing grazing regimes in Greek mountain systems. J. Appl. Ecol. 35, 983–986. doi: 10.1111/j.1365-2664.1998.tb00019.x
Zhao, F. Z., Kang, D., Han, X. H., Yang, G. H., and Feng, Y. Z. (2015). Soil stoichiometry and carbon storage in long-term afforestation soil affected by understory vegetation diversity. Ecol. Eng. 74, 415–422. doi: 10.1016/j.ecoleng.2014.11.010
Zhao, J. X., Li, X., Li, R. C., Tian, L. H., and Zhang, T. (2016). Effect of grazing exclusion on ecosystem respiration among three different alpine grasslands on the central Tibetan Plateau. Ecol. Eng. 94, 599–607. doi: 10.1016/j.ecoleng.2016.06.112
Keywords: alpine grassland, degradation, fencing, Qinghai–Tibetan plateau, soil organic carbon, species richness
Citation: Huang C, Peng F, You Q, Liao J, Duan H, Wang T and Xue X (2020) The Response of Plant and Soil Properties of Alpine Grassland to Long-Term Exclosure in the Northeastern Qinghai–Tibetan Plateau. Front. Environ. Sci. 8:589104. doi: 10.3389/fenvs.2020.589104
Received: 31 July 2020; Accepted: 13 October 2020;
Published: 12 November 2020.
Edited by:
Lu-Jun Li, Northeast Institute of Geography and Agroecology (CAS), ChinaReviewed by:
Yangong Du, Northwest Institute of Plateau Biology (CAS), ChinaXuyang Lu, Institute of Mountain Hazards and Environment (CAS), China
Copyright © 2020 Huang, Peng, You, Liao, Duan, Wang and Xue. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xian Xue, eGlhbnh1ZUBsemIuYWMuY24=