 Mel McGregor
Mel McGregor Kelly Matthews
Kelly Matthews Darryl Jones
Darryl Jones- 1School of Environment, Environmental Futures Research Institute, Griffith University, Nathan, QLD, Australia
- 2Green Tape Solutions – Environmental Consulting, Moggill, QLD, Australia
The effects of urbanization on flying vertebrates historically have been underestimated, with roads in particular impacting bat populations through vehicle collisions, habitat fragmentation and unwillingness to disperse across roads. Although vegetated fauna overpasses have not traditionally been considered as beneficial for microbats, these structures have recently been identified as a potential solution for assisting microbat communities. In Brisbane, Queensland, Australia, the Compton Road fauna overpass traverses a four–lane road, facilitating connectivity for mammals, birds, reptiles and amphibians between two forest reserves. This study examined patterns of activity and species richness of microbat assemblages in relation to the Compton Road fauna overpass. Call recordings made over 7 months identified nine species and two species groups (groups of species unable to be differentiated), all of which were recorded using the overpass. Bat activity on the overpass was higher than in the adjoining forest, with higher detectability. Species richness and activity at the roadside where the overpass was present was also higher than the unvegetated roadside. Activity patterns based on feeding behaviors suggest that forest species such as Nyctophilus and Myotis spp. were using the overpass, alongside more generalist species. The vegetation on the overpass is likely to be the key factor encouraging microbat activity. The Compton Road overpass appears to lessen the impact of road presence and may facilitate permeability of a major road, suggesting potential for enhancing habitat connectivity and facilitating crossings. This study is the first indication that vegetated fauna overpasses can facilitate activity and provide natural habitat continuity for diverse Australian microbat communities.
Introduction
The effects of urbanization on highly mobile flying vertebrates, such as bats, have been historically underestimated (Threlfall et al., 2013), in part from the assumption that they are less influenced by habitat fragmentation than are non-volant species (Gilbert, 1989). Realization of this misconception has facilitated increased research efforts on volant species (e.g., birds: Pell and Jones, 2015; bats: Lesinski, 2008; Bennett and Zurcher, 2013), ultimately showing that urbanization, and the subsequent decrease in habitat connectivity, reduction of remnant habitat and increased anthropogenic disturbance, can reduce species richness, abundance and activity in volant species including bats (see review by Russo and Ancillotto, 2014). Roads in particular, have been shown to directly influence bat mortality (Medinas et al., 2013) by posing significant barriers to movement (Abbott et al., 2012; Bennett and Zurcher, 2013) and restricting dispersal for roosting and foraging (Abbott et al., 2012). Bat density and diversity have also been shown to decrease with proximity to major roads (Berthinussen and Altringham, 2012) while the presence of vehicles themselves can restrict foraging and commuting behaviors (Kerth and Melber, 2009; Zurcher et al., 2010; Bennett et al., 2013). Direct effects such as vehicle strikes (Gaisler et al., 2009; Lesinski et al., 2011), street lighting and traffic noise (Longcore and Rich, 2004; Stone et al., 2009; Bennett and Zurcher, 2013) are also known to impact urban bat communities by altering foraging, commuting, roosting and activity times (Stone et al., 2015). For example, increased artificial light is known to dramatically reduce foraging activity and delay commuting behavior in lesser horseshoe bats (Rhinolophus hipposideros) (Stone et al., 2009), while road noise greatly impairs the passive listening abilities of greater mouse–eared bats (Myotis myotis) (Schaub et al., 2008). Vegetated fauna overpasses have the potential to mitigate these impacts for bats by reintroducing structural habitat complexity, providing safe passage across roads, as has been shown for other taxa (Bond and Jones, 2008; Mata et al., 2008; Jones and Pickvance, 2013; McGregor et al., 2015).
Fauna overpasses are gaining recognition as a key mitigation strategy for increasing or restoring landscape connectivity and permeability affected by roads (van der Ree et al., 2015). Overpasses have traditionally targeted medium to large mammals (Forman et al., 2003; Glista et al., 2009; Clevenger and Ford, 2010); however, the presence of vegetation has been found to enhance utility by non-target taxa such as birds, reptiles and bats, by establishing a continuation of the natural landscape (Bond and Jones, 2008; Pell and Jones, 2015). Fully vegetated overpasses, particularly those which closely resemble the surrounding natural habitat, have been shown to encourage diverse overpass use by non-target species (McGregor et al., 2015) including small mammals (Bond and Jones, 2008), herpetofauna (Bond and Jones, 2008; Mata et al., 2008), birds (Pell and Jones, 2015) and potentially bats (Bach and Muller-Steiss, 2005; Abbott et al., 2015). Although some studies have observed the use of underpasses by bats (Abson and Lawrence, 2003; Bach et al., 2004; Abbott et al., 2012; Berthinussen and Altringham, 2012), overpasses have only recently been suggested as a mitigation tool for potentially improving road permeability for bats (Bach and Muller-Steiss, 2005; Berthinussen and Altringham, 2012; Abbott et al., 2015). The current understanding of overpass use by bats is extremely limited (Altringham and Kerth, 2015), with only one study, to our knowledge, specifically addressing vegetated overpass use by bats (Bach and Muller-Steiss, 2005). A relatively small number of studies have documented bat interactions with over–road structures (e.g., foot bridges) or unvegetated overpasses (Abbott et al., 2012; Berthinussen and Altringham, 2012; Bhardwaj et al., 2017). These studies noted that features such as strategic location, size, connectivity of tree lines and mature vegetation encouraged use of overpasses by bats, whereas road flyovers (high level road bridges) or footbridges, did not facilitate effective road crossings for bats (Abbott et al., 2015; Altringham and Kerth, 2015).
Roadside vegetation directly affects bat behavior (Zurcher et al., 2010). For example, vegetation along road verges may facilitate commuting, while gaps in vegetation such as tree canopies can prevent the establishment of bat commuting routes (Bennett and Zurcher, 2013). Vegetation near roads can also provide protection as well as foraging habitat (Verboom and Huitema, 1997) and increased insect prey (Avila-Flores and Fenton, 2005). Considering the importance of vegetation to the movement (Russo and Ancillotto, 2014) and behavior (Bennett and Zurcher, 2013) of many bat species, it is likely that a key determinant of overpass use by bats is the presence and structure of appropriate vegetation. The Compton Road fauna overpass is a component of a diverse array of fauna crossing structures in south Brisbane, Queensland, Australia, which were constructed specifically to facilitate the movement of medium to large mammals. The overpass has since been shown to provide road permeability for a range of target and non-target taxa including mammals (Bond and Jones, 2008; Taylor and Goldingay, 2012), herpetofauna (McGregor et al., 2015) and birds (Pell and Jones, 2015). The success of this overpass for non-target species in particular, has been attributed to the established vegetation that integrates the overpass into the surrounding forest (McGregor et al., 2015; Pell and Jones, 2015), facilitating habitat continuity across Compton Road. The aim of the present study was to explore whether the Compton Road overpass, as a fully vegetated fauna crossing, facilitates road permeability for microbat species. It was expected that the road would influence bat activity and species richness, the impacts of which may be lessened where the overpass is present.
Study Site
Compton Road is a major urban arterial consisting of two dual lanes, located in southern Brisbane, Queensland, Australia. The speed limit of Compton Road is 70 km/h and current traffic volume is estimated at approximately 10,000 cars daily (Brisbane City Council, 2014). The road bisects Karawatha Forest Reserve (940 ha) to the south and Kuraby Bushland (140 ha) to the north, two of Brisbane's most significant urban bushland remnants. Both reserves provide natural bushland habitat for a range of native flora and fauna, including numerous significant or threatened species (Mack, 2005; Veage and Jones, 2007). A creek line runs throughout the study area, from Kuraby, underneath Compton Road and parallel with the road within Karawatha forest. Some areas of the forest have been cleared for service roads, fire brakes and power lines (see Figure 2).
During 2004–2005, the widening of Compton Road from two to four lanes threatened to permanently isolate the remnant forests, as well as increase collisions between wildlife and vehicles (Veage and Jones, 2007). To mitigate these impacts, the road upgrade incorporated the Compton Road fauna array (27°36′53.11″S, 153°05′03.12″E) which included a vegetated overpass (Figure 1) and two underpasses. The overpass is hourglass shaped, 70 m long, 15 m wide at the midpoint and 20 m wide at the ends. Roadside exclusion fencing (2.5 m high) extends the full length of the overpass and along the length of each forest at the roadside (Bond and Jones, 2008; Jones, 2010). The natural vegetation of Karawatha and Kuraby is classified as dry eucalypt woodland with native heath understories (Veage and Jones, 2007; Jones, 2010). The vegetation structure and composition on the overpass is similar to the surrounding forests and was planted at a rate of 70 shrubs and six trees per 100 m2 shortly after construction was complete (2005). This planting regime, primarily consisting of mature saplings, was maintained to facilitate the movement of macropods (Jones, 2010). Surveys undertaken in 2009 confirmed that 95% of the planted trees and shrubs had survived (Jones et al., 2011), maintaining compositional similarity with the surrounding forest.

Figure 1. Compton Road fauna overpass in 2015 (Photo: M. McGregor).
Materials and Methods
Bat Detection and Capture
Bat presence and activity were recorded over two consecutive nights each month for 7 months between December 2014 and July 2015 (February was excluded due to adverse weather). Two EchoMeter Touch (EMT) devices (30 m diameter maximum range, Wildlife Acoustics) and corresponding iPad 4s (Apple Technologies) were used to monitor eight walking transects and two stationary points, beginning approximately 1 h after sunset (in varying order). Transect layout was mirrored on each side of the road, while stationary points were located on the overpass apex, approximately 30 m apart, all of which were monitored each night (Figure 2). In each forest, two “road transects” (RT) (defined as transects beginning at the managed, and therefore unvegetated, roadsides) were located to the west, one to the east of the overpass (see Figure 2). A single eastern transect was monitored due to the impassable nature of nearby terrain/creek line. Additionally, two overpass transects (OT) (defined as transects beginning level with the road side, but located on the overpass) began level with the fence line and extended into the forest (see Figure 2). Each walking transect extended 75 m into the forest, at least 75 m from neighboring transects, with each one being monitored for 20 min. Stationary points on the apex were also monitored simultaneously for 20 min each. Stationary points were monitored by personnel (holding the EMT) in order to maintain human presence at all sites. On each survey night, a single EMT was used to monitor each transect by a maximum of two personnel, always beginning from the roadside, but in no regimented order.
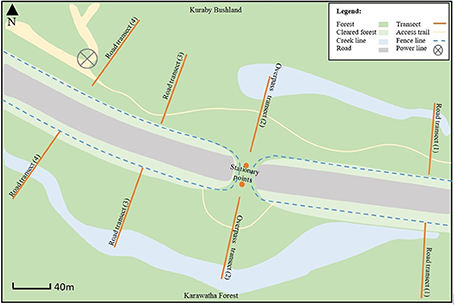
Figure 2. Layout of Eight 75 m walking transects and two stationary points, used to record bat call activity throughout Karawatha Forest and Kuraby Bushland.
Road transects started from the unvegetated roadside fence line, extending into the forest, while the overpass transects began in the constructed habitat on the overpass (in line with the fence) and continued into the forest. The important distinction between road and overpass transects was the lack of vegetation at the roadside, meaning while the overpass transects remained within vegetated habitat for the length of the transect, the road transects did not. This difference allowed for two important comparisons. First, comparing the overpass and road transects at the roadside allowed an assessment of variation in bat activity at the roadside in both the presence and absence of vegetation at the roadside. Second, monitoring from the roadside into the forest determined whether there was variation in bat activity on the roadside compared with in the forest.
To ensure that each walking transect occupied 20 min, six stops were made along the length of the transect of approximately 3 min each; data recorded between the stops was also included. Monitoring did not occur under the overpass for safety reasons. Ethical clearance was approved by the Griffith University Animal Ethics Committee (ENV 08/14 AEC).
Data Analysis
Call analysis was completed using Kaleidoscope V3.1 and SongScope V4.1.3 (Wildlife Acoustics). Identification was based on local call libraries (south east Queensland and northern New South Wales; from Pennay et al., 2004; Churchill, 2009). Search phase calls, consisting of a series of regular sound pulses (Pennay et al., 2004), were used in this study for species identification, while feeding buzzes (a series of often erratic pulses), were used to identify active hunting behavior if occurring within a clearly identifiable call (Figure 3).
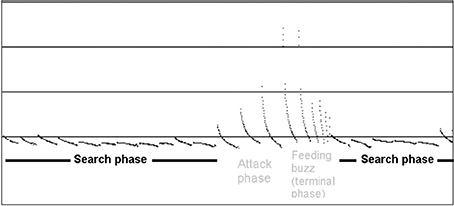
Figure 3. Example diagram shows search phase and feeding buzz call features of a Mormopterus species, used for identification during data analysis (figure reproduced from Pennay et al., 2004).
A complete call was identified by the presence of multiple (three or more), clearly identifiable pulses. Pulse characteristics such as initial sweep shape, body shape and terminal sweep shape (e.g., presence of tail) were used to distinguish between similar species (Pennay et al., 2004). Recordings were identified to species level by M.M. and K.M. independently, while calls that could not be confidently identified were analyzed by a third party for verification. All remaining unidentified calls (less than 5%) were removed from the data set. Calls from species that could not be reliably distinguished from each other, for example Scotoropens sp., were grouped together as species groups (Hourigan et al., 2010) (see Table 1).
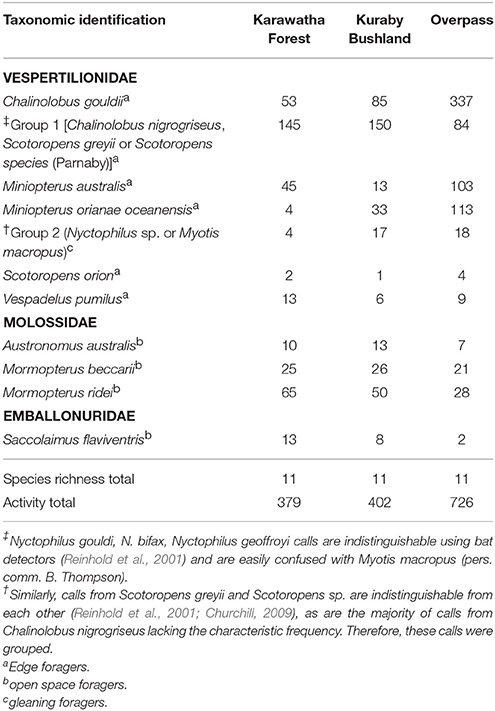
Table 1. Bat species identified from the recorded echolocation calls, including total calls recorded for Karawatha, Kuraby and the overpass.
Vocalization analyses focussed on exploring three major trends over the 7 months: microbat activity and richness between the three study areas (Karawatha, Kuraby and the overpass); whether microbat activity and richness varied at the roadside compared with the forest; and finally, whether microbat activity and richness was altered at the roadside by the presence of the overpass. SIMPER analysis (PRIMER 7, PRIMER–E Ltd.) was used to determine the similarity of species activity with distance from the roadside. SIMPER was also used to compare overpass transects to the other transects to determine the impact of vegetation on species similarity at the roadside. Species accumulation curves (SPSS 24, IBM Statistics1; Microsoft Excel, 20162) based on richness detected at the beginning and end of the transects were used to explore the likelihood of methodological bias, as well as variation in detectability (likelihood of detecting representative species richness), particularly between the overpass and the forest, and the roadside and the forest. The latter was also used to examine if species richness varied considerably between the roadside and the forest. QGIS 2.10.1 geoprocessing software was used to produce heat maps of total call activity using Google Earth base layers (Google Earth version 7.1.5, 2015) based on Kernel Density Estimation (using night activity as a single data point to determine mean activity over 7 months). Heat map values used call activity numbers and are shown at a radius of 15 m to reflect the maximum potential range of the EMT devices. Transect points were used to display activity within the transect in its entirety and were not treated as discrete data points for analysis, except when compared with each other, to avoid sample bias.
The cryptic nature and high mobility of the study species presented methodological challenges during monitoring. Bat call detection was the only feasible way to monitor sufficient quantities of bats; harp traps were trialed and deemed ineffective due to the lack of captures relative to what was a highly demanding procedure. However, using detectors restricted analysis and application, primarily due to the inability to identify individuals. In this respect, “activity” was explored, rather than abundance, removing any requirement to identify individuals, but was considered a reasonable basis for determining general patterns of bat richness and activity around Compton Road and the overpass. This study is limited to a single study site. Therefore, it relies on replication of transects within the area to determine patterns of activity and species richness, while replication for monitoring the overpass apex uses two monitoring points for comparison. Due to these limitations, analyses were limited to exploratory and descriptive statistical methods, with a view to helping to construct viable hypotheses that can be tested in future studies.
Results
Nine bat species were reliably identified within the study area from 1,550 clear echolocation call recordings. Clear calls that could not reliably distinguished between species were grouped (Group 1 and Group 2; see Table 1). All nine species and both species groups were recorded in Karawatha, Kuraby and on the overpass. Call activity in the two forests were similar, with a total of 387 recorded calls in Karawatha ( = 27.07 ± 23.34 SD) and 402 in Kuraby ( = 28.71 ± 27.46). Call activity recorded on the overpass (apex points) was almost double (726 calls; = 51.86 ± 59.97) that of the surrounding forest (Table 1).
Species Richness and Similarity
Mean overpass calls (stationary points only), averaged throughout the monitoring period, were predominantly comprised of Chalinolobus gouldii (24.07 ± 45.28 SD), Miniopterus orianae oceanensis (12.56 ± 11.81), and Miniopterus australis (8.07 ± 16.55 SD) which were consistently present on every occasion throughout the study. Mean within group species similarity was lowest at the roadside (17.42% similar) and increased in the forest (to a maximum of 57.61% similarity). Dissimilarity was higher between the roadside and the forest (X = 66.21) compared with dissimilarity within the forest (X = 59.376). Species contribution to dissimilarity between the roadside and the forest was dominated by three species, which were present in all comparisons: Group 1 (mean contribution 30.45%), Mormopterus beccarii (mean contribution 25.27%) and C. gouldii (mean contribution 13.19%). Calls from M. beccarii (34.6%), Group 1 (25.64%) and C. gouldii (16.67%) comprised the majority of roadside call activity.
Mean dissimilarity was high between the overpass transects (level with the roadside) and the roadside recordings at all other transects (RT1 = 93.22, RT3 = 77.55, RT4 = 65.85). C. gouldii and Mormopterus ridei contributed to dissimilarity in all cases (mean contributions 31.06, 24.79% respectively) due to high calls of both species on the overpass transects.
Species Accumulation
Species accumulation quickly reached asymptote within each of the study areas (11 species/species groups) (Figure 4). The overpass reached an asymptote sooner than either forest, reaching 11 recorded species on the fourth site visit (January 2015), while Karawatha and Kuraby reached asymptote on the seventh (March 2015) and 10th visits, respectively; however, all three sites displayed similar detection rates.
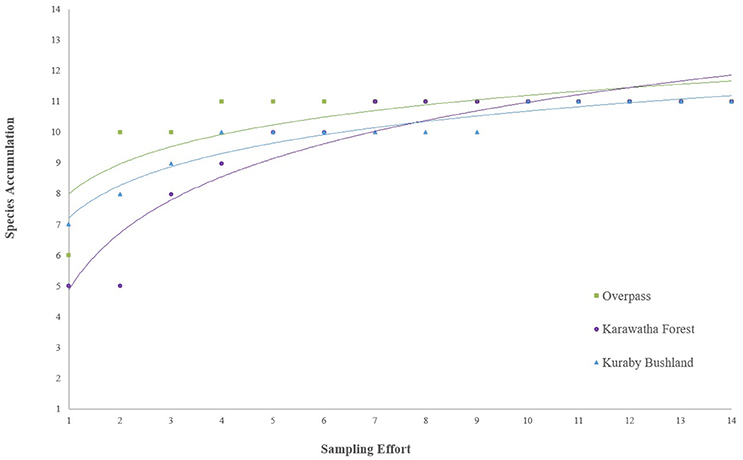
Figure 4. Species accumulation graph comparing detectability and species richness within Karawatha Forest, Kuraby Bushland and on the overpass.
In comparing species accumulation at the roadside end of each transect to the inner forest end, there was little difference in species richness and species detectability, with the exception of the T2 transect at the roadside (which began on the overpass). The “road” end of T2 transects, in both Karawatha and Kuraby, displayed higher species richness at a faster detection rate than any other forest or roadside point (Figure 5).
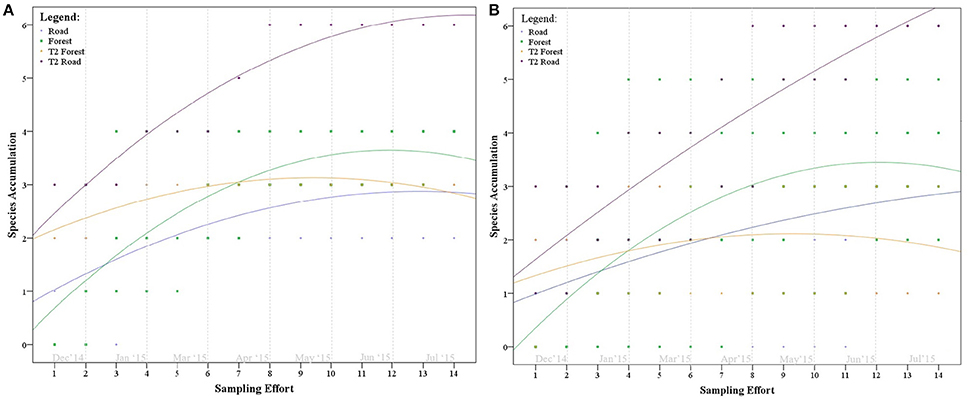
Figure 5. Species accumulation graph comparing detectability and species richness at the roadside and forest ends of each transect (15 m radius) in (A) Karawatha Forest and (B) Kuraby Bushland.
Call Activity
Call activity was divided into three feeding behavior categories: open foragers (Austronomus australis, M. beccarii, M. ridei, Saccolaimus flaviventris); edge foragers (C. gouldii, Group 1, M. australis, M. orianae oceanensis, S. orion, Vespadelus pumilus); and gleaning foragers (Group 2) (Table 1), to determine whether a visual trend could be identified, based on species behavior. All three behavior categories were strongly concentrated on the overpass. Open foragers showed higher activity at the roadside, the creek lines, as well as on the overpass (Figure 6). Edge foragers appeared to be spread evenly throughout most transects, while also showing high concentrations on the overpass (Figure 6). Gleaning species were rarely recorded within the transects but appeared to be concentrated on the Karawatha side of the overpass and on one Kuraby transect (Figure 6).
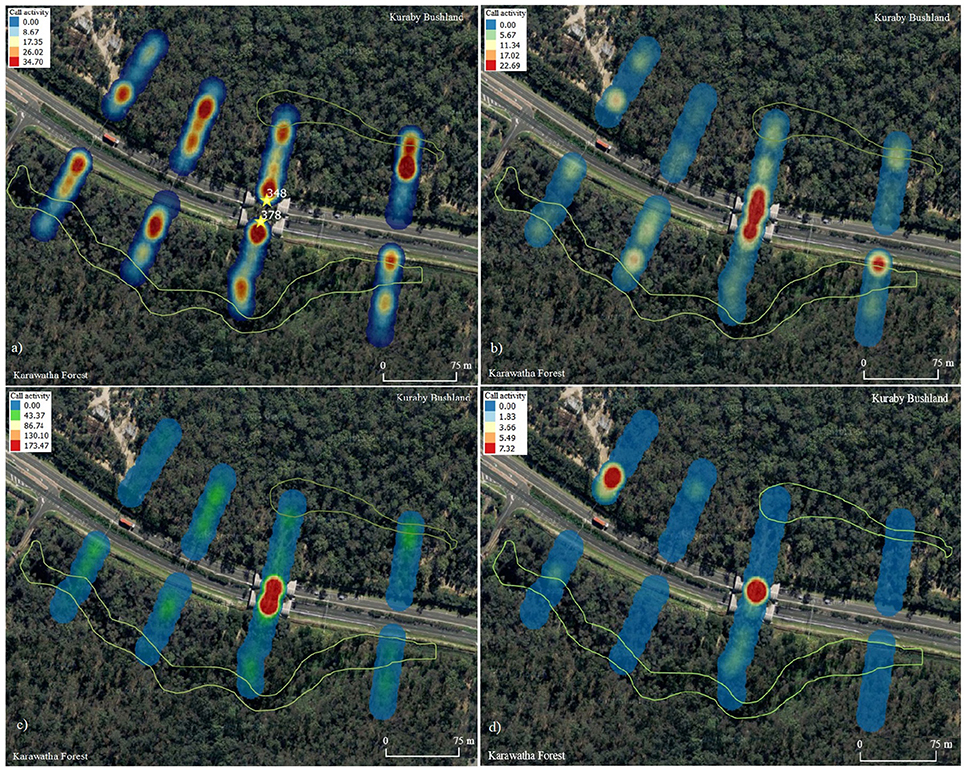
Figure 6. Call activity along the transects for (a) all species, (b) open foraging species only, (c) edge foraging species, only and (d) gleaning species only (mean total calls). Call activity increases from blue to red with intensity (creek and marshland areas outlined in green). *Overpass calls for total species activity are noted separately.
Calls pertaining to specific behaviors varied slightly between the three areas, with the highest percentage of feeding buzzes (indicating active hunting) being found on the overpass at 4.57%, compared with Karawatha (3.31%) and Kuraby (2.95%). Search phases (indicating no active hunting or interactions between individuals) were the least common call recorded on the overpass, with 11.69%, compared with Karawatha (40%) and Kuraby (20.64%).
Discussion
South east Queensland supports high microbat diversity, with at least 24 known species (Churchill, 1998), 18 of which occur around Brisbane (Churchill, 2009; Hourigan et al., 2010). Previous studies conducted in Karawatha Forest detected at least nine species (Hourigan, 2011), all of which were detected in the present study (although total species richness may be as high as 14, considering indistinguishable call groupings). The presence of microbats on the overpass was substantial, with a high level of activity compared with either forest, and a species richness equal to that found in Karawatha and Kuraby (Table 1). Comparisons of species accumulation between the three areas showed detected species richness on the overpass reaching asymptote before either of the forest areas (Figure 4). However, both forests did achieve asymptote, suggesting the use of apex point counts on the overpass did not contribute to sample bias, irrespective of using different observation methods. In comparing the overpass transects to the forest transects directly, where the overpass intersects the road had comparatively higher species richness, and high detectability (Figures 5A,B), compared to both the unvegetated roadsides and the inner forest. Although the “road” end of the overpass transects had higher species richness and greater detectability than any other roadside or forest records, neither side reached asymptote, suggesting that more species may be using the sides of the overpass, but have not yet been detected. High species detectability, equal species richness and high activity observed throughout the study suggest that diverse microbat communities were using the Compton Road overpass.
Contrary to initial predictions, call activity did not appear to vary substantially with proximity to the roadside. Detectability was similar between the cleared roadside and the forest, as was species richness, while the total activity density map shows activity hotspots that did not appear to be correlated with road presence (Figure 6a). Typical foraging behaviors were used to explore patterns of activity throughout the study area, based on differences between microbats that may not be affected by road presence, compared with those that are more likely to be susceptible. These groups form the basis for determining how foraging behaviors influence activity patterns, and although some bats such as Mormopterus sp. can alter feeding behaviors as required, these groups represent the primary feeding behavior typically displayed by each species. When the data were explored in this way, patterns of activity between bats with typically varied behaviors appear apparent (Figure 5A). Call activity along the roadside varied from activity observed in the forest, with most roadside activity dominated by open foragers (M. beccarii, C. gouldii and Group 1). Forest specialists such as Nyctophilus (Group 2) were not specifically correlated with road presence or dense forest. Instead, two hotspots of Group 2 detections were present: transect 4 in Kuraby, and on the overpass. Landscape variation is likely to have contributed to activity patterns observed during the study, a recognized aspect of bat behavior (Bennett and Zurcher, 2013), while it is possible a nearby roost may have skewed activity results. Call activity among all species appears to be positively associated with creek presence and open areas.
Current literature recognizes that roads act as barriers to bat populations (Kerth and Melber, 2009; Berthinussen and Altringham, 2012; Bennett and Zurcher, 2013) by restricting dispersal (Abbott et al., 2012) or forcing changes in behavior, such as road avoidance (Zurcher et al., 2010; Bennett and Zurcher, 2013), although the degree to which this occurs is debated. Berthinussen and Altringham (2011) observed that roads act as considerable, if not complete, barriers to microbat species, while Abbott et al. (2012) suggested that roads of similar construction to Compton Road are not absolute barriers to bat movement. How severely bat communities are affected by road barriers often depends on their morphology (Rydell, 1992; Kerth and Melber, 2009; Bhardwaj et al., 2017). Therefore, observing activity patterns between bats with different foraging behaviors may assist in determining whether, and how, Compton Road affects bat communities living in the surrounding forest. The high levels of bat activity, richness and detectability on the overpass (apex and sides) compared with the cleared roadside suggests that the overpass does provide a level of relief of road presence; however this same trend is notably present when comparing the sides of the overpass with the inner forest. One explanation for this may be the hourglass shape of the overpass creates a bottleneck, restricting bat activity to a narrow area, and therefore concentrating activity to within range of the EMTs. This particular point would benefit from a more intensive monitoring regime which may include monitoring further into each forest.
Bats are known to alter their behavior in response to the structure and availability of vegetation (Bennett and Zurcher, 2013) with suitable vegetation facilitating higher bat diversity (Abbott et al., 2012, 2015). The presence of comparatively high bat activity on the overpass, by all species and groups detected, suggests that the reason for high detectability, richness and activity on the overpass is the presence of complexity in the form of vegetation. The overpass appears to provide habitat sufficient enough to support, and indeed encourage, diverse bat species. This is evident in the comparative concentration of gleaning species activity on the overpass apex, suggesting appropriately complex and suitable habitat for forest specialists. The vegetation on the overpass may be sufficient to reduce negative aspects of the road which are known to affect bats, for example artificial light (Stone et al., 2009) and traffic noise (Schaub et al., 2008), two elements of the road that are noticeably reduced on the apex, although these were not measured in the present study. In addition, the presence of open and edge foragers suggests a planting regime which does not exclude these species, a known consequence of dense vegetation (Medinas et al., 2013). The planting regime employed on the overpass aimed to replicate the local open woodland forest, which may have encouraged overpass use by species rich microbats. To fully understand the benefit of the overpass to bats, ascertaining crossing data by tagging and tracking individual bats would be of great benefit. This would specifically determine whether individuals are using the overpass as a crossing route.
Limitations of the study area size restricted the size of the transects which, due to the potential for road effects to extend far into forest habitat, may have been influenced by the road effect zone. The limited call activity from gleaning species throughout the forest, such as long–eared Nyctophilus which are recognized as sensitive forest specialists (Lumsden et al., 2002; Threlfall et al., 2013), compared with far higher call activity from edge feeding free–tailed and sheath–tailed bats, suggests that the road effect zone may extend into the forest, beyond the length of the 75 m transects. This possible outcome is consistent with taxa-diverse studies which suggest road effect zones can extend from 100 to 1,000 m from the road (Reijnen et al., 1995; Eigenbrod et al., 2009), which includes aspects of disturbance (such as noise and light) known to affect bat activity (Schaub et al., 2008; Stone et al., 2009). The broad spread and high call activity from edge foraging species may also suggest that edge effects extend well into the forest, while the dominant presence of Molossidae species, as open-space foragers (Jung et al., 2014), along the roadside indicates substantial influence of road. These patterns suggest that Compton Road may be affecting more sensitive bat species, and is likely to present a complete barrier if this is the case. The presence of Nyctophilus/Myotis sp. activity on the overpass may provide evidence of increased habitat permeability, and most likely some level of road effect mitigation for these species. It is unlikely that Compton Road impacts open and edge foraging species to the same intensity, although it does not prevent them from being at risk of barrier effects, traffic injury or other road related impacts (Kerth and Melber, 2009).
A variety of management practices have been suggested to reduce road impacts on bat species and potentially prevent further fragmentation. Bennett and Zurcher (2013) recommend management practices that involve interlinking tree canopies across roads, as well as restoring and enhancing vegetation along roadsides. One goal of habitat restoration is to enhance landscape permeability, which is also a primary goal of overpass construction (Forman et al., 2003). Although vegetated overpasses have not traditionally been built or considered for bat communities, the overlap between what an overpass achieves, and what benefits bat communities, appears to be substantial. Bach et al. (2004) concluded that over– road crossing features are more restrictive to bat species than are under–road (typical underpasses as well as culverts) crossings, while underpasses have been suggested as a potential solution to reducing road impacts on some bat species (Abson and Lawrence, 2003; Kerth and Melber, 2009; Boonman, 2011; Abbott et al., 2012). The ecological value of the underpasses at Compton Road has not been determined; however, call activity and species richness on the overpass suggests that the overpass is a substantial asset to the facilitation of habitat continuity across Compton Road.
Current research has highlighted the necessity of maintaining remnant forests within urbanized environments for the long–term persistence of microbats (Avila-Flores and Fenton, 2005; Oprea et al., 2009; Threlfall et al., 2013). The increasing reduction and isolation of patches of remnant forest in urban areas worldwide continues to jeopardize local biodiversity, particularly for species sensitive to habitat fragmentation. Retention of tree cover and linear vegetation, in particular, are critical to the sustainable protection and long–term persistence of urban forests for use by bat communities (Threlfall et al., 2013). Additionally, the presence of overpasses and similar infrastructure that enhances and reintroduces habitat connectivity needs to be appreciated in areas that are already highly fragmented. The Compton Road fauna overpass has shown to be successful in facilitating road permeability for a wide variety of taxa (Bond and Jones, 2008; McGregor et al., 2015; Pell and Jones, 2015); this may now include microbats, with the potential for maintaining habitat connectivity and enabling crossing between Karawatha and Kuraby. Without determining whether bats are crossing the road without the aid of the overpass, it is difficult to quantify “effectiveness” (as defined in Forman et al., 2003; van der Ree et al., 2007; van der Grift and van der Ree, 2015). However, this study shows that the overpass itself is being frequented by a diverse range of microbat species. This study is the first indication that a vegetated fauna overpass can facilitate activity and provide road permeability for diverse microbats in Australia, and one of very few worldwide. It is, therefore, significant in demonstrating the potentially important role of fauna passages in facilitating the long–term persistence of microbat communities in urban landscapes.
Author Contributions
MM devised, managed and conducted field research and wrote the majority of the final manuscript as well as facilitating the editing and submission processes. KM contributed significantly to expertise required in the field and assisted in correctly writing the technical aspects of the manuscript as well as editing. DJ supervised the field research and writing process, contributing substantially to editing, research design and execution of the project.
Funding
Wildlife Queensland student research grant ($1,000).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer CT and handling Editor declared their shared affiliation.
Acknowledgments
We would like to thank Wildlife Queensland for providing equipment funding and Brisbane City Council for their support and facilitation of this research. We are grateful to everyone who assisted in the field and to Bruce Thompson who provided expertise in identifying difficult bat call recordings. Comments by reviewers improved the quality of the manuscript, as did constructive comment from Rob Appleby. We recognize the association between this manuscript and the Infra-Eco Network of Ecology (IENE) Conference in 2016. This research was undertaken with the permission of Brisbane City Council and the Griffith University Animal Ethics Committee (ENV 08/14 AEC).
Footnotes
1. ^SPSS (V. 22, IBM) IBM Corp. Released 2013. IBM SPSS Statistics for Windows, Version 19.0. Armonk, NY: IBM Corp.
2. ^Microsoft Excel 2016 (Released in 2016). Microsoft Corporation, Washington DC, United States of America.
References
Abbott, I. M., Berthinussen, A., Stone, E., Boonman, M., Melber, M., and Altringham, J. (2015). Bats and roads, in Handbook of Road Ecology, eds R. van der Ree, C. Grilo, and D. J. Smith (West Sussex: John Wiley and Sons), 293–302.
Abbott, I. M., Butler, F., and Harrison, S. (2012). When flyways meet highways – the relative permeability of different motorway crossing sites to functionally diverse bat species. Landsc. Urban Plan. 106, 293–302. doi: 10.1016/j.landurbplan.2012.03.015
Abson, R. N., and Lawrence, R. E. (2003). Monitoring the use of the Slaty Creek wildlife underpass, Calder Freeway, Black Forest, Macedon, Victoria, Australia, in Proceedings of the 2003 International Conference on Ecology and Transportation, eds C. L. Irwin, P. Garrett, and K. P. McDermott (Raleigh, NC: Center for Transportation and the Environment, North Carolina State University), 303–308.
Altringham, J., and Kerth, G. (2015). Bats and roads, in Bats in the Anthropocene: Conservation of Bats in a Changing World, Vol. 606, eds C. Voigt and T. Kingston (New York, NY: Springer), 35–62. doi: 10.1007/978-3-319-25220-9_3
Avila-Flores, R., and Fenton, M. (2005). Use of spatial features by foraging insectivorous bats in a large urban landscape. J. Mammal. 86, 1193–1204. doi: 10.1644/04-MAMM-A-085R1.1
Bach, L., Burkhardt, P., and Limpens, H. (2004). Tunnels as a possibility to connect bat habitats. Mammalia 68, 411–420. doi: 10.1515/mamm.2004.041
Bach, L., and Muller-Steiss, H. (2005). Fachbeitrag Fledermause an ausgewahlten Grunbrucken. Effizienzkontrolle von Wildtierpassagen in Baden-Wurttemberg.
Bennett, V. J., and Zurcher, A. A. (2013). When corridors collide: road-related disturbance in commuting bats. J. Wildl. Manage. 77, 93–101. doi: 10.1002/jwmg.467
Bennett, V., Sparks, D., and Zollner, P. (2013). Modeling the indirect effects of road networks on the foraging activities of bats. Landsc. Ecol. 28, 979–991. doi: 10.1007/s10980-013-9874-0
Berthinussen, A., and Altringham, J. (2011). The effect of a major road on bat activity and diversity. J. Appl. Ecol. 49, 82–89. doi: 10.1111/j.1365-2664.2011.02068.x
Berthinussen, A., and Altringham, J. (2012). Do bat gantries and underpasses help bats cross roads safely? PLoS ONE 7:e38775. doi: 10.1371/journal.pone.0038775
Bhardwaj, M., Soans, K., Straka, T. M., Lahoz-Monfort, J. J., Lumsden, L. F., and van der Ree, R. (2017). Differential use of highway underpasses by bats. Biol. Conserv. 212, 22–28 doi: 10.1016/j.biocon.2017.05.022
Bond, A. R., and Jones, D. N. (2008). Temporal trends in use of fauna-friendly underpasses and overpasses. Wildl. Res. 35, 103–112. doi: 10.1071/WR07027
Boonman, M. (2011). Factors determining the use of culverts underneath highways and railway tracks by bats in lowland areas. Lutra 54, 3–16.
Brisbane City Council (2014). Traffic Data: Compton Road. Brisbane: Brisbane City Council. Available online at: www.brisbane.qld.gov.au
Clevenger, A. P., and Ford, A. T. (2010). Wildlife crossing structures, fencing, and other highway design considerations, in Safe Passages. Highways, Wildlife, and Habitat Connectivity, eds J. P. Beckmann, A. P. Clevenger, M. Huijser, and J. A. Hilty (Washington, DC: Island Press), 17–49.
Eigenbrod, F., Hecner, S. J., and Fahrig, L. (2009). Quantifying the road-effect zone: threshold effects of a motorway on Anuran populations in Ontario, Canada. Ecol. Soc. 14:24. doi: 10.5751/ES-02691-140124
Forman, R. T. T., Sperling, D., Bissonette, J. A., Clevenger, A. P., Cutshall, C. D., Dale, V. H., et al. (2003). Road Ecology: Science and Solutions. Washington, DC: Island Press.
Gaisler, J., Řehák, Z., and Bartonička, T. (2009). Bat casualties by road traffic (Brno-Vienna). Acta Theriol. 54, 147–155. doi: 10.1007/BF03193170
Glista, D. J., DeVault, T. L., and DeWoody, J. A. (2009). A review of mitigation measures for reducing wildlife mortality on roadways. Landsc. Urban Plan 91, 1–7. doi: 10.1016/j.landurbplan.2008.11.001
Google Earth version 7.1.5 (2015). Available online at: https://earth.google.com (Accessed June 18, 2015).
Hourigan, C. (2011). Insectivorous Bat Diversity and Habitat Use in a Subtropical Mosaic Urban Landscape. Ph.D. thesis, Griffith University.
Hourigan, C., Catterall, C., Jones, D., and Rhodes, M. (2010). The diversity of insectivorous bat assemblages among habitats within a subtropical urban landscape. Aust. Ecol. 35, 849–857. doi: 10.1111/j.1442-9993.2009.02086.x
Jones, D. (2010). Vegetation structure on overpasses is critical in overcoming the road barrier effect for small birds, in IENE 2010 International Conference on Ecology and Transportation (Scope Ltd.).
Jones, D. N., Bakker, M., Bichet, O., Coutts, R., and Wearing, T. (2011). Restoring habitat connectivity over the road: vegetation on a fauna land-bridge in south-east Queensland. Ecol. Manage. Restor. 12, 76–79. doi: 10.1111/j.1442-8903.2011.00574.x
Jones, D. N., and Pickvance, J. (2013). Forest birds use vegetated fauna overpass to cross multi-lane road. Oecol. Aust. 17, 42–51. doi: 10.4257/oeco.2013.1701.12
Jung, K., Molinari, J., and Kalko, E. (2014). Driving factors for the evolution of species-specific echolocation call design in new world free-tailed bats (Molossidae). PLoS ONE 9:e85279. doi: 10.1371/journal.pone.0085279
Kerth, G., and Melber, M. (2009). Species-specific barrier effects of a motorway on the habitat use of two threatened forest-living bat species. Biol. Conserv. 142, 270–279. doi: 10.1016/j.biocon.2008.10.022
Lesinski, G. (2008). Linear landscape elements and bat casualties on roads - an example. Ann. Zool. Fenn. 45, 277–280. doi: 10.5735/086.045.0406
Lesinski, G., Sikora, A., and Olszewski, A. (2011). Bat casualties on a road crossing a mosaic landscape. Eur. J. Wildl. Res. 57, 217–223. doi: 10.1007/s10344-010-0414-9
Longcore, T., and Rich, C. (2004). Ecological light pollution. Front. Ecol. Environ. 2, 191–198. doi: 10.1890/1540-9295(2004)002[0191:ELP]2.0.CO;2
Lumsden, L., Bennett, A., and Silins, J. (2002). Location of roosts of the lesser long-eared bat Nyctophilus geoffroyi and Gould's wattled bat Chalinolobus gouldii in a fragmented landscape in south-eastern Australia. Biol. Conserv. 106, 237–249. doi: 10.1016/S0006-3207(01)00250-6
Mack, P. (2005). When Fauna Corridors and Arterial Road Corridors Intersect. Brisbane: City Design, Brisbane City Council.
Mata, C., Hervás, I., Herranz, J., Suárez, F., and Malo, J. E. (2008). Are motorway wildlife passages worth building? Vertebrate use of road-crossing structures on a Spanish motorway. J. Environ. Manage. 88, 407–415. doi: 10.1016/j.jenvman.2007.03.014
McGregor, M. E., Wilson, S. K., and Jones, D. N. (2015). Vegetated fauna overpass enhances habitat connectivity for forest dwelling herpetofauna, Global Ecol. Conserv. 4, 221–231. doi: 10.1016/j.gecco.2015.07.002
Medinas, D., Marques, J., and Mira, A. (2013). Assessing road effects on bats: the role of landscape, road features, and bat activity on road-kills. Ecol. Res. 28, 227–237. doi: 10.1007/s11284-012-1009-6
Oprea, M., Mendes, P., Vieira, T., and Ditchfield, A. (2009). Do wooded streets provide connectivity for bats in an urban landscape? Biodivers. Conserv. 18, 2361–2371. doi: 10.1007/s10531-009-9593-7
Pell, S., and Jones, D. (2015). Are wildlife overpasses of conservation value for birds? A study in Australian sub-tropical forest, with wider implications. Biol. Conserv. 184, 300–309. doi: 10.1016/j.biocon.2015.02.005
Pennay, M., Law, B., and Reinhold, L. (2004). Bat Calls of New South Wales: Region Based Guide to the Echolocation Calls of Microchiropteran Bats. Hustville, NSW: New South Wales Department of Environment and Conservation.
Reijnen, R., Foppen, R., Braak, C. T., and Thissen, J. (1995). The effects of car traffic on breeding bird populations in woodland. III. Reduction of density in relation to the proximity of main roads. J. Appl. Ecol. 32, 187–202 doi: 10.2307/2404428
Reinhold, L., Law, B., Ford, G., and Pennay, M. (2001). Key to the Bat Calls of South-East Queensland and North-Eastern New South Wales. Brisbane, QLD: Queensland Department of Natural Resources and Mines.
Russo, D., and Ancillotto, L. (2014). Sensitivity of bats to urbanization: a review. Mamm. Biol. 80, 205–212. doi: 10.1016/j.mambio.2014.10.003
Rydell, J. (1992). Exploitation of insects around streetlamps by bats in Sweden. Funct. Ecol. 6, 744–750. doi: 10.2307/2389972
Schaub, A., Ostwald, J., and Siemers, B. M. (2008). Foraging bats avoid noise. J. Exp. Biol. 211, 3174–3180. doi: 10.1242/jeb.022863
Stone, E. L., Jones, G., and Harris, S. (2009). Street lighting disturbs commuting bats. Curr. Biol. 19, 1123–1127. doi: 10.1016/j.cub.2009.05.058
Stone, E. L., Jones, G., and Harris, S. (2015). Impacts of artificial lighting on bats: a review of challenges and solutions. Mamm. Biol. 80, 213–219. doi: 10.1016/j.mambio.2015.02.004
Taylor, B., and Goldingay, R. (2012). Restoring connectivity in landscapes fragmented by major roads: a case study using wooden poles as “stepping stones” for gliding mammals. Restor. Ecol. 20, 671–678. doi: 10.1111/j.1526-100X.2011.00847.x
Threlfall, C. G., Law, B., and Banks, P. B. (2013). The urban matrix and artificial light restricts the nightly ranging behaviour of Gould's long-eared bat (Nyctophilus gouldi). Austral Ecol. 38, 921–930. doi: 10.1111/aec.12034
van der Grift, E. A., and van der Ree, R. (2015). Guidelines for evaluating use of wildlife crossing structures, in Handbook of Road Ecology, eds R. van der Ree, C. Grilo, and D. J. Smith (West Sussex: John Wiley and Sons), 119–128.
van der Ree, R., Smith, D. J., and Grilo, C. (2015). The Ecological Effects of Linear Infrastructure and Traffic: 1-9, in Handbook of Road Ecology, eds R. van der Ree, C. Grilo, and D. J. Smith (West Sussex: John Wiley and Sons).
van der Ree, R., van der Grift, E., Gulle, N., Holland, K., Mata, C., and Suarez, F. (2007). Overcoming the barrier effect of roads – how effective are mitigation strategies? An international review of the use and effectiveness of underpasses and overpasses designed to increase the permeability of roads for wildlife, in Proceedings of the 2007 International Conference on Ecology and Transportation (Raleigh, NC: Center for Transportation and Environment, North Carolina State University), 423–431.
Veage, L., and Jones, D. N. (2007). Breaking the barrier: assessing the value of fauna-friendly crossing structures at Compton Road, in Report to Brisbane City Council, Centre for Innovative Conservation Strategies (Griffith University), 112.
Verboom, B., and Huitema, H. (1997). The importance of linear landscape elements for the pipistrelle Pipistrellus pipistrellus and the serotine bat Eptesicus serotinus. Landsc. Ecol. 12, 117–125. doi: 10.1007/BF02698211
Keywords: fauna passage, overpass, land bridge, Yangochiroptera, road ecology, habitat connectivity
Citation: McGregor M, Matthews K and Jones D (2017) Vegetated Fauna Overpass Disguises Road Presence and Facilitates Permeability for Forest Microbats in Brisbane, Australia. Front. Ecol. Evol. 5:153. doi: 10.3389/fevo.2017.00153
Received: 03 January 2017; Accepted: 17 November 2017;
Published: 30 November 2017.
Edited by:
Rodney Van Der Ree, University of Melbourne, AustraliaReviewed by:
Caragh Grace Threlfall, University of Melbourne, AustraliaDaniel Lewanzik, Max Planck Institute for Ornithology (MPG), Germany
Copyright © 2017 McGregor, Matthews and Jones. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mel McGregor, bWVsLm1jZ3JlZ29yQGdyaWZmaXRodW5pLmVkdS5hdQ==