 Carolina Levis1,2*
Carolina Levis1,2* Bernardo M. Flores3
Bernardo M. Flores3 Priscila A. Moreira4
Priscila A. Moreira4 Bruno G. Luize5
Bruno G. Luize5 Rubana P. Alves1
Rubana P. Alves1 Juliano Franco-Moraes6
Juliano Franco-Moraes6 Juliana Lins7Evelien Konings2Marielos Peña-Claros2Frans Bongers2Flavia R. C. Costa8
Juliana Lins7Evelien Konings2Marielos Peña-Claros2Frans Bongers2Flavia R. C. Costa8 Charles R. Clement9
Charles R. Clement9- 1Programa de Pós-graduação em Ecologia, Instituto Nacional de Pesquisas da Amazônia, Manaus, Brazil
- 2Forest Ecology and Forest Management Group, Wageningen University & Research, Wageningen, Netherlands
- 3Departamento de Biologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, Campinas, Brazil
- 4Programa de Pós-graduação em Botânica, Instituto Nacional de Pesquisas da Amazônia, Manaus, Brazil
- 5Programa de Pós-graduação em Ecologia e Biodiversidade, Instituto de Biociências, Universidade Estadual Paulista (UNESP), Rio Claro, Brazil
- 6Programa de Pós-graduação em Ecologia, Instituto de Biociências, Universidade de São Paulo, São Paulo, Brazil
- 7Instituto Socioambiental, São Gabriel da Cachoeira, Brazil
- 8Coordenação de Pesquisas em Biodiversidade, Instituto Nacional de Pesquisas da Amazônia, Manaus, Brazil
- 9Coordenação de Tecnologia e Inovação, Instituto Nacional de Pesquisas da Amazônia, Manaus, Brazil
For millennia, Amazonian peoples have managed forest resources, modifying the natural environment in subtle and persistent ways. Legacies of past human occupation are striking near archaeological sites, yet we still lack a clear picture of how human management practices resulted in the domestication of Amazonian forests. The general view is that domesticated forests are recognizable by the presence of forest patches dominated by one or a few useful species favored by long-term human activities. Here, we used three complementary approaches to understand the long-term domestication of Amazonian forests. First, we compiled information from the literature about how indigenous and traditional Amazonian peoples manage forest resources to promote useful plant species that are mainly used as food resources. Then, we developed an interdisciplinary conceptual model of how interactions between these management practices across space and time may form domesticated forests. Finally, we collected field data from 30 contemporary villages located on and near archaeological sites, along four major Amazonian rivers, to compare with the management practices synthesized in our conceptual model. We identified eight distinct categories of management practices that contribute to form forest patches of useful plants: (1) removal of non-useful plants, (2) protection of useful plants, (3) attraction of non-human animal dispersers, (4) transportation of useful plants, (5) selection of phenotypes, (6) fire management, (7) planting of useful plants, and (8) soil improvement. Our conceptual model, when ethnographically projected into the past, reveals how the interaction of these multiple management practices interferes with natural ecological processes, resulting in the domestication of Amazonian forest patches dominated by useful species. Our model suggests that management practices became more frequent as human population increased during the Holocene. In the field, we found that useful perennial plants occur in multi-species patches around archaeological sites, and that the dominant species are still managed by local people, suggesting long-term persistence of ancient cultural practices. The management practices we identified have transformed plant species abundance and floristic composition through the creation of diverse forest patches rich in edible perennial plants that enhanced food production and food security in Amazonia.
Introduction
The notion of pristine rainforests has been questioned by increasing archaeological and ecological evidence suggesting long-term human activities across even the most intact forests worldwide (Denevan, 1992; Van Gemerden et al., 2003; Willis et al., 2004; Ross, 2011; Boivin et al., 2016; Roberts et al., 2017). Amazonia is no exception — over thousands of years with humans living in the region, forest composition has been altered significantly (Clement et al., 2015; Levis et al., 2017b). Many dominant species in Amazonian forests are widely used as food resources by native indigenous peoples (ter Steege et al., 2013), and at least 85 tree and palm species were domesticated to some degree during pre-Columbian times (Clement, 1999; Levis et al., 2017b). Plant domestication is a long-term process that results from the capacity of humans to overcome environmental selection pressures with the purpose of managing and cultivating useful plants (Kennedy, 2012; Boivin et al., 2016; Levis et al., 2017b), leading to significant changes in natural ecosystems and plant communities across landscapes (Clement, 1999; Terrell et al., 2003). First, useful individuals are managed in situ (Rindos, 1984; Wiersum, 1997a) and later humans select the best varieties with more desirable morphological traits for cultivation (Darwin, 1859; Rindos, 1984; Clement, 1999). Over time, humans create a mosaic of domesticated landscapes to favor numerous useful plant populations, each domesticated with different intensities and outcomes (Wiersum, 1997b). In modern Amazonian forests, legacies of past human societies are evident in the surroundings of archaeological sites, where humans enriched the forest with useful, especially edible, and domesticated plants (Balée, 1989; Erickson and Balée, 2006; Junqueira et al., 2010; Levis et al., 2017b). These pre-Columbian legacies suggest that Native Amazonians interacted with natural ecological processes and shaped the distribution of plants and entire forest landscapes across the region (Balée, 2013).
In Amazonia, as in any other ecosystem, natural ecological processes drive the formation of plant assemblages and communities (Keddy, 1992; Zobel, 1997; Lortie et al., 2004; ter Steege et al., 2006). The first ecological process described to structure plant communities is the plant's capacity to disperse its seeds across landscapes (Ricklefs, 1987; Lortie et al., 2004), which depends on the regional species pool and multiple dispersal strategies, including occasional events of long distance dispersal (Ricklefs, 1987; Nathan et al., 2008). In wet Neotropical forests, animal dispersal is used by 75–98% of the tree species (Howe and Smallwood, 1982; Muller-Landau et al., 2008) and mammals disperse large-seeded species over long distances (Jordano, 2017). Once a propagule arrives in a given location, the second ecological process is related to how plants are able to overcome local environmental filters to successfully germinate and survive (Lortie et al., 2004). Plants compete with their neighbors for limited amounts of resources, such as light, nutrients and water (Moles and Westoby, 2006). The understory of a tropical forest is typically light-limited, forcing trees to either grow tall or survive in shady conditions (Poorter et al., 2003). Soils are also limited in water and nutrients, and plants need to compete in the rooting zone (Barberis and Tanner, 2005; Schnitzer et al., 2005). The third ecological process structuring plant assemblages is interaction with other organisms, such as herbivores and pathogens (Lortie et al., 2004; Bagchi et al., 2014). These multiple environmental and biological filters act simultaneously, resulting in trade-offs. For instance, species that grow fast under high light conditions tend to produce leaves that are less protected from herbivores, compared to the tougher and more resistant leaves of shade-tolerant species (Coley, 1983). In the long run, these ecological processes result in the selection of numerous adaptive plant traits (Reich et al., 2003), allowing species to thrive in complex and highly diverse systems, such as Amazonian forests. The high diversity of tropical ecosystems is in part maintained by natural disturbances and local biotic interactions, sometimes promoted by herbivores and pathogens that reduce the abundance of the most effective competitors, creating space for other species (Connell, 1978; LaManna et al., 2017).
Nonetheless, a few tree species often dominate plant assemblages forming oligarchic forests in diverse tropical forests (Connell and Lowman, 1989; Peh et al., 2011), including Amazonia (Peters et al., 1989; Pitman et al., 2001, 2013; ter Steege et al., 2013), Africa (Hart et al., 1989; Hart, 1990; Peh et al., 2011), Mesoamerica (Campbell et al., 2006), and Asia (Connell and Lowman, 1989; Peh et al., 2011). Natural and anthropogenic origins for the hyperdominance of tree species in Amazonian forests have been proposed. Aggregated patches of a few pioneer species occur after human or natural disturbance, while aggregated patches of a few shade-tolerant species may occur due to dispersal limitations (Valencia et al., 2004). Other hypotheses to explain why some species dominate large areas of Amazonian forests include: the species' ability to tolerate multiple environmental conditions, and to disperse over long distances (Pitman et al., 2001, 2013); and, in the case of useful species, the intentional or non-intentional enrichment promoted by past and contemporary human societies (Balée, 1989, 2013; Peters et al., 1989; ter Steege et al., 2013; Levis et al., 2017b).
During the Holocene, useful plant populations benefited from a new set of interactions when humans started to transform landscapes (Denevan, 1995; Smith, 2011; Boivin et al., 2016), and manage plant populations, consciously or not (Rindos, 1984; Wiersum, 1997a,b; Peters, 2000). Indigenous management practices were formally defined by Wiersum (1997a, p. 7) as “the process of making and effectuating decisions about the use and conservation of forest resources within a local territory.” When humans consciously manage forest resources, the underlying intention of their actions is not to domesticate forests, but to achieve certain short-term objectives, for instance to favor individual plants in the forest and promote their regeneration. Although changes in forest composition may not be the main goal of human actions, management practices also modify forest composition and structure beyond the targeted species in a long-term process. In tropical and subtropical forests worldwide, native societies have managed plants and landscapes, promoting oligarchic forests dominated by useful plant species, also defined as cultural or domesticated forests (Balée, 1989, 2013; Peters et al., 1989; Campbell et al., 2006; Michon et al., 2007; Reis et al., 2014; Morin-Rivat et al., 2017).
Today, many indigenous and traditional peoples recognize the handprints of their ancestors in the landscape (Frikel, 1978). Indigenous people are defined here as the descendants of native ethnic groups and members of an indigenous community that retains historical and cultural connections with the social organization of pre-Columbian indigenous societies (https://pib.socioambiental.org)1. Traditional peoples can be understood as culturally differentiated and recognizable groups that have their own forms of social organization using knowledge, innovations and practices generated and transmitted by tradition, but they are not recognized as a member of indigenous communities (Brazilian Federal Decree No. 6.040)2. In Amazonia, traditional peoples are generally descendants of migrants who intermarried with local indigenous peoples and they often exchange practices, objects and knowledge with members of indigenous communities. Although contemporary indigenous and traditional societies both cultivate fruit trees in their territory, they also take advantage of the aggregated patches of fruit trees created by the practices of previous generations (Frikel, 1978; Balée, 1989, 2013). These ancient cultivated landscapes were probably created by integrated agroforestry systems that included homegardens, swiddens and managed fallows in which tree and non-tree crops were intertwined (Denevan et al., 1984; Stahl, 2015). Such integrated systems were likely more efficient, in terms of food production, than long-fallow shifting cultivation systems when only stone axes were used to clear the forest in the past (Denevan, 1992). This is supported by the fact that past indigenous tree cultivation (arboriculture) was a common and widespread practice covering large areas of forest-savanna transition zones in Amazonia (Frikel, 1978).
Because trees persist in the forest following management (Levis et al., 2017b) and annual crops disappear after human abandonment (Clement, 1999), contemporary indigenous and traditional people commonly attribute the aggregated distribution of useful perennial plants to the action of their ancestors. Based on this knowledge, they sometimes select a new place to settle in the forest (Frikel, 1978; Politis, 2007; Rival, 2007; Zurita-Benavides et al., 2016). For instance, the Nukak Indians in Colombian Amazonia prefer camping around sororoca plants (Phenakospermum guyannense), because they believe that these plants were brought by their ancestors to “their living world,” and they discard a large quantity of seeds around their temporary camps, contributing to form new patches (Politis, 2007). Given that multiple human generations have moved around through time, places like riverine settings and archaeological sites were frequent dispersal routes of people and their cultures, and consequently of useful plants in pre- and post-Columbian times (Denevan, 1996; Hornborg, 2005; Guix, 2009; Heckenberger and Neves, 2009; Clement et al., 2010; Levis et al., 2017a,b). The intimate connections between Native Amazonians, their ancestors and their plants can reveal how persistent pre-Columbian forest management practices (Balée, 2000) contributed to the large-scale vegetation patterns we observe in modern forests (Pitman et al., 2011; Levis et al., 2017a,b).
Our study aimed to unravel how people interacted with natural ecological processes to transform pristine forests into domesticated forests with different degrees of human intervention through unintentional and intentional management practices. How indigenous and traditional peoples have used and shaped Amazonian forests is described in ethnographical, ethnobotanical, archaeological, paleoethnobotanical, paleoecological, and ecological publications. Here we used a historical-ecological perspective to evaluate the available information about how Native Amazonians have affected the distribution of plant species used mainly as food resources. Based on the information gathered from the literature, we developed an interdisciplinary conceptual model of how multiple management practices transformed pristine forests into domesticated forests, considering temporal and spatial contexts. In the field, we collected data about management practices and the composition of forest patches dominated by useful plants surrounding 30 contemporary villages, settled on or near archaeological sites. We compared field and literature data by documenting the multiple management practices known by 33 informants from two villages along the lower Tapajós River, and by relating these practices to the distribution and composition of the forest patches surrounding all 30 villages.
Materials and Methods
Construction of the Conceptual Model of Forest Domestication
We reviewed the scientific literature for evidence of management practices of 22 useful perennial species (mainly used as food resources) that occur in forest patches in different parts of the Amazon basin (see Supplementary Table 1 for information about the species). These species were also chosen because the authors had previous field knowledge about them and they include a variety of useful plants with wild, cultivated and domesticated populations. Although our review focused on edible perennial plants, we used the general concept of useful plants to define plant species that are currently used for any purpose or have been used by any human group in the past. Eighty-one studies in ethnographical, ethnobotanical, archaeological, paleoethnobotanical, paleoecological and ecological publications, including books, scientific articles and dissertations, were analyzed (Supplementary Data). The literature review was conducted using the scientific name, English name and Portuguese name of each species as keywords in Web of Science and as title in Google Scholar.
Based on the information gathered for the 22 species, we classified the multiple management practices into eight categories that consist of a summary of all practices reported in the literature (Table 1): (1) removal of non-useful plants, (2) protection of useful plants, (3) attraction of non-human dispersers of useful plants, (4) human transportation of useful plants, (5) selection of phenotypes useful to humans, (6) fire management, (7) planting, and (8) soil improvement. The literature review provides examples to identify the role of—in many cases—multiple management practices in the formation and persistence of domesticated forests in Amazonia.
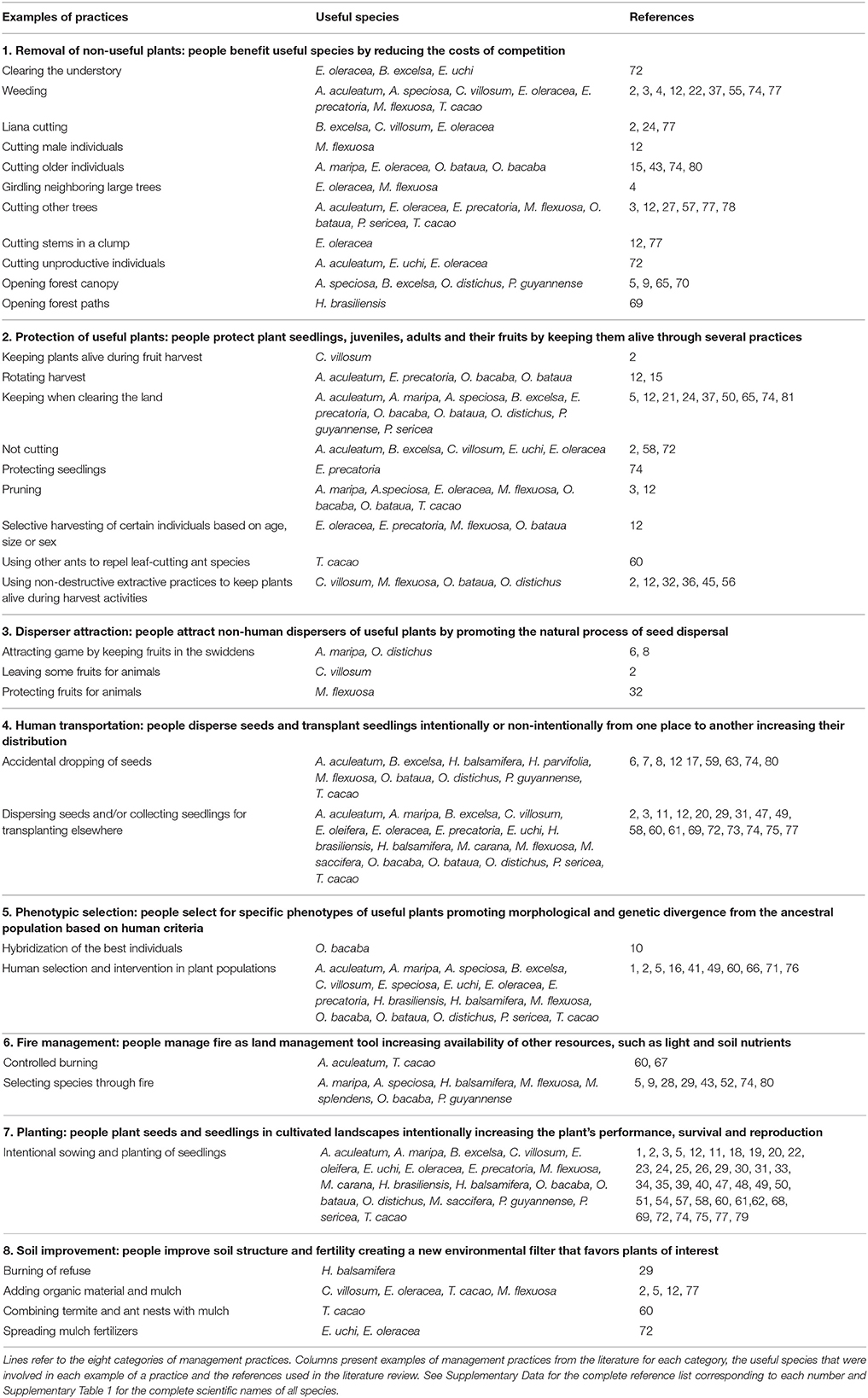
Table 1. Examples of all management practices classified into eight categories.
We combined different management practices into a category depending on: (1) what people want to achieve; (2) whether the effects of the practice are directional or not in the way they fundamentally shape plant species assemblages; and (3) whether the practices result in similarities in terms of forest composition, abundance and distribution of useful species. For instance, practices that remove non-useful plants in the forest, such as opening the canopy, clearing the understory, weeding and cutting lianas, are used to selectively benefit useful species or enhance their growth rate by reducing the competition of non-useful plants around the targeted plants. As a side effect, humans increase light availability in the forest and tend to favor light demanding species that may therefore be protected if useful. More similarities are expected inside each category than between them because each category leads to a unique type of interference in natural ecological processes. Nonetheless, their interactions may result in a diverse composition of useful species with different or even contrasting adaptations. Below we detail each of these eight categories, providing a definition, interaction with ecological processes and some examples.
Removal of Non-useful Plants
The most common practices used to remove non-useful plants in the forest are: opening the canopy; clearing the understory; weeding; cutting lianas; and removing unproductive individuals of useful species. These practices are used to selectively benefit useful species by reducing the costs of competition, and are expected to increase the performance of the selected useful plants. Competition can be reduced either by controlling the abundance of non-useful species (directly excluding them), or increasing the amount of available resources (e.g., light or space). Practices that reduce leaf and root density of lianas, for example, can release the growth of some trees (Schnitzer et al., 2005), and increase fruit production (Kainer et al., 2014). Similar to other small-scale natural disturbances (Connell, 1978), these long-term management practices may increase the diversity of plants between plant communities at a regional scale (beta-diversity) (Balée, 2006). The Hotï Indians from northern Amazonia act as ecological disturbance agents by constantly creating and managing gaps that increase the amount of light inside the forest necessary to cultivate light-demanding useful plants (Zent and Zent, 2004). In southern Amazonia, the Kayapó Indians create forest islands by managing savanna landscapes, increasing the heterogeneity of the landscape and the resource abundance for humans, game animals and plants (Posey, 1985). The Nukak Indians from western Amazonia constantly move between old camps for hunting and gathering activities; when returning to old camps, they selectively clear the understory and canopy, altering plant composition and benefiting useful and domesticated plants by promoting their growth and reproduction (Politis, 1996).
Protection of Useful Plants
Humans protect plant seedlings, juveniles, adults and their fruits by keeping them alive through several practices: taking care of fruits, seedlings and adult plants; using non-destructive extractive practices; avoiding fire near useful trees; pruning; and repelling leaf-cutting ant species. Protection can be targeted to individuals with specific traits or to whole plant populations, by reducing the abundance of herbivores, predators, and natural disturbances. For instance, the Kayapó Indians in southern Amazonia use Azteca ants to repel leaf-cutting ants that eat useful species' leaves (Posey, 1987). The Huaorani Indians in western Amazonia and Hotï Indians in northern Amazonia increase the abundance of several useful plant species by keeping fruit trees alive in their territory (Rival, 1998; Zent and Zent, 2012). Aggregated patches of many useful plants are spared when clearing the forest for crop cultivation (Shanley et al., 2016), increasing the survival rates of these plants. This practice protects useful plant populations of Amazon nut trees (Bertholletia excelsa), uxí trees (Endopleura uchi), tucumã palms (Astrocaryum aculeatum), and açaí palms (Euterpe oleracea) in different parts of Amazonia (Shanley et al., 2016). Babaçu palms (Attalea speciosa) with more inflorescences are also protected in agroforestry systems of eastern Amazonia (Anderson et al., 1991).
Attraction of Non-human Dispersers of Useful Plants
The natural process of seed dispersal can be enhanced by human practices. Leaving some fruits under the mother tree for animals in domesticated landscapes and cultivating large-seeded species to attract game are common practices in traditional communities of Amazonia (Shanley et al., 2010). Although humans were responsible for population declines, and even local extinctions of large vertebrates across Neotropical forests (Guimarães Jr. et al., 2008), humans have also positively interacted with terrestrial animals by increasing their food availability via cultivation and protection of fruit trees in domesticated landscapes (Balée, 1993), thus increasing the dispersal capacity and distribution of useful plant species. Dispersal strategies among large-seeded species and their dispersers may result in aggregated distributions of Amazonian plant species. For instance, forest patches of inajá palm (Attalea maripa) are associated with tapir latrines, suggesting that tapirs are partly responsible for the aggregated distribution of this palm in Amazonian forests (Fragoso et al., 2003). Seeds of bacaba palm (Oenocarpus distichus) persist in secondary forests of Ka'apor Indians after abandonment, because game is attracted to these food resources and disperse even more seeds within these forests (Balée, 1993, 2013). Attracting animals to domesticated landscapes may indirectly contribute to form and maintain multi-species patches of useful plants from ancient homegardens and swiddens (Balée, 2013).
Human Transportation of Useful Plants
Human transportation is the intentional or non-intentional movement of seeds and plants by humans from one place to another, outside or within the geographical limits of the plant population. For instance, planting seedlings or dispersing seeds intentionally and non-intentionally along forest trails, in swiddens and homegardens. During the Holocene, humans may have acted as primary long-distance dispersal vectors by transporting seeds of useful plants over long distances, often surpassing natural evolutionary barriers (Hodkinson and Thompson, 1997; Nathan et al., 2008). Past humans intentionally transported seeds, seedlings and clones of useful plants over long distances across the world (Boivin et al., 2016). As a consequence, the expansion of sedentary farming populations in Amazonia is associated with the dispersal of important native crops across the basin, such as manioc (Manihot esculenta) (Arroyo-Kalin, 2012), Amazon nut trees (Shepard and Ramirez, 2011; Thomas et al., 2015), and cacao trees (Theobroma cacao) (Thomas et al., 2012). Over short distances, human seed dispersal occurs when plants are exchanged among groups (Eloy and Emperaire, 2011), during periodic movements of groups to new areas (Posey, 1993), systematic movements between forests and settlements (Ribeiro et al., 2014), and between temporary camps (Politis, 2007). Short distance dispersal within a plant population's range is also reported, when seeds are scattered along trails during hunting and gathering activities, often non-intentionally (Zent and Zent, 2004; Ribeiro et al., 2014). The Hotï spend days in the forest to collect large quantities of umirí (Humiria balsamifera) fruits, many of which drop from baskets on the way back to the village, explaining its high abundance surrounding their villages (Zent and Zent, 2004). Similarly, the Kayapó transport large amounts of Amazon nut seeds, suggesting that the high density of seedlings along trail margins results from seeds accidentally dropped during transport (Ribeiro et al., 2014). Extensive trail systems were described in the Kayapó territory where they intentionally plant, transplant and spread useful species (Posey, 1993), forming landscapes full of useful plant species.
Phenotypic Selection of Useful Plants
Trait selection practices are motivated by human preferences for specific phenotypes, for instance, fruits with larger sizes or larger contents of desirable properties, such as sugar, starch and oil. Humans often protect individuals previously selected for their preferred traits and they propagate these individuals outside their original population (see section Human Transportation of Useful Plants), resulting in plant domestication (Rindos, 1984; Clement, 1999). Phenotypic selection promotes morphological and genetic divergence from the ancestral population based on human criteria (Clement, 1999). The set of phenotypic traits that distinguish domesticated from wild plant populations is called the domestication syndrome (Hammer, 1984; Harlan, 1992; Meyer et al., 2012). Selection does not necessarily imply intentionality; however, if unconscious practices lead to changes in plant traits, followed by selection and propagation, these actions start to be systematically repeated (Rindos, 1984; Zeder, 2006). Human criteria for selecting plant traits vary across geographical regions, through time and with cultural interests (Meyer et al., 2012), and depend on the availability of useful populations in the landscape and the knowledge to interpret and manage morphological variation (Terrell et al., 2003). In Amazonia, some studies have described domestication syndromes for useful plants: variation in the toxicity of manioc roots that were selected for different soil types (McKey et al., 2010; Fraser et al., 2012); peach palm (Bactris gasipaes) may have been first selected for its small oily fruits or wood, and later for large starchy fruits with better fermentation qualities (Clement et al., 2009); the selection of annatto (Bixa orellana) with increased pigment yield from its seeds, and changed fruit dehiscence (Moreira et al., 2015); the high morphological variation of pequí fruit (Caryocar brasiliense) varieties selected by the Kuikuro Indians of the upper Xingu River (Smith and Fausto, 2016); selection of varieties of Virola elongata with exudates of different hallucinogenic qualities, and varieties of Cyperus articulatus with rhizomes having different medicinal properties selected by Yanomami groups in Northwestern Brazil (Albert and Milliken, 2009). Along the lower Tapajós River, traditional people selected non-bitter fruits of Caryocar villosum, domesticating them accidentally or intentionally (Alves et al., 2016). The importance of selection for promoting agrobiodiversity in Amazonia is underscored in ethnographies of cultivated plants, such as manioc (Boster, 1984; Rival and McKey, 2008) and pequí (Smith and Fausto, 2016).
Fire Management
Fire has been a land management tool since pre-historical times (Pausas and Keeley, 2009). People have used prescribed fire in forests or swiddens mainly for cultivation, and also highly controlled fire for waste management near their houses. People manage fire for hunting activities, group communication, rituals, and to prevent uncontrollable fires (Mistry et al., 2016). Fire was intensely managed by pre-Columbian peoples in homegardens or settlement areas for domestic activities, such as cooking and burning waste. This domestic use may have contributed in the long run to fertilize the soil, producing the Terra Preta de Índio (TPI or Amazonian Dark Earths – ADE) (Smith, 1980; Schmidt et al., 2014) found throughout the Amazon basin (McMichael et al., 2014). Fire was also managed in swiddens to improve soil fertility with intensive cultivation techniques in ancient times, forming fertile dark brown soils, a soil slightly less fertile than TPI (Denevan, 2001; Woods et al., 2013). Management practices involving fire also increase availability of other resources, such as light, by reducing the abundance of competitors, and promoting useful species that are more nutrient demanding, such as chili peppers (Capsicum spp.) (Junqueira et al., 2016a). Patches of burití palms (Mauritia flexuosa), for instance, are associated with fire history in the Gran Savana, where people have used fire to prevent forest re-expansion into savannas (Montoya et al., 2011). When people manage fire to reduce competition for cultivated plants, fire-adapted species are often selected (Jakovac et al., 2016a). Many plants, useful or not, have evolved to tolerate contact with fire, allowing them to persist through time in frequently burnt places (Bond and Midgley, 2001). Some examples are the light-demanding sororoca (P. guyanense) that resprout after fire, cumatí trees (Myrcia splendens) that form patches in gaps managed with fire (Elias et al., 2013) and babaçu palms that persist in burnt sites due to cryptogeal germination (Jackson, 1974). The ancient connection between fire and humans (Bowman et al., 2011) and the intense fire history in Amazonian forests is revealed by the high charcoal abundance in forests around old settlements (Bush et al., 2015), which are expected to be dominated by fire-adapted species.
Planting
Planting is defined here as the intentional planting, sowing and transplanting of seeds and seedlings to cultivated landscapes. It is important to note that when seeds and seedlings are transported by humans (see section Human Transportation of Useful Plants) with the intention of planting, these categories overlap. When humans disperse seed without this intention (e.g., when gathering fruits in the forest) the overlap between planting and human transportation doesn't exist, which justifies separating these categories of practices. Planting practices may increase a useful plant's performance, survival and reproduction because people usually take care of seedlings after planting. In Amazonia, several tree and palm species are planted mostly in agroforestry systems, forest gardens and forest gaps surrounding settlements (Denevan et al., 1984; Balée, 1993; Zent and Zent, 2012). In the past, indigenous groups also planted several perennial species, originating patches of useful trees and palm species across the basin (Frikel, 1978). Therefore, the presence and abundance of edible trees and palms in Amazonian forests and their proximity to ancient settlements may indicate past indigenous planting activities (Balée, 2013; Levis et al., 2017b). Some examples in Amazonia are forest patches of Poraqueiba sericea (Padoch and De Jong, 1987; Franco-Moraes, 2016) in western Amazonia, C. brasiliense in the upper Xingu River (Smith and Fausto, 2016), C. villosum in the lower Tapajós River (Alves et al., 2016), and B. excelsa in Amapá (Paiva et al., 2011) that are all associated with past indigenous planting.
Soil Improvement
In some parts of the Amazon basin, terra-firme forests are poor in nutrients, which selected for plants with efficient nutrient-conservation mechanisms (Herrera et al., 1978). Amerindians, however, interfered with these processes by changing soil structure and increasing soil fertility (Kleinman et al., 1995). Soil improvement involves several practices, such as the addition of charcoal and ashes that release nutrients and carbon in the soil; the use of organic additives, such as human and animal wastes, ash, garbage, crop residues, leaves, compost, cleared weeds, seaweed, mulch, urine, ant nest refuse, turf, muck, and water; and also by building mounds in floodable landscapes (Denevan, 1995, 2001). The improvement of soil conditions was observed for piquiá trees inside the forest, in which local people accumulate leaf litter under the trees (Alves et al., 2016), and for açaí, uxí, and peach palm through organic additives (Shanley et al., 2016). Also, extremely fertile TPI were probably created in pre-Columbian refuse heaps in which ash and charcoal, human and animal wastes, and ceramics accumulated (Woods and McCann, 1999; Schmidt et al., 2014). Although TPI soils were a product of sedentary human settlement and cannot be classified as a management practice, modern people usually take advantage of these fertile soils to cultivate crops (Junqueira et al., 2016b). Brown soils were probably formed in cultivation zones with ash and charcoal that originated from frequent burning, and by composting and mulching the soil (Denevan, 1995). Unintentional and sometimes intentional soil improvement practices that resulted in the creation of TPI and brown soils were probably common in the past, since anthropogenic soils occur across most of the Amazon basin (Woods et al., 2013). The improvement of soil structure and fertility creates a new environmental filter that favors plants of interest and excludes species not adapted to the new soil conditions. Species with adaptations to resist or tolerate fire or to benefit from fertile soils may become dominant in improved soils. As a consequence, useful species adapted to fertile soils can form aggregated patches in TPI sites across the basin (Balée, 1989). This is may be case H. balsamifera trees, dominant in soils previously burned in the upper Negro River (Franco-Moraes, 2016), and palm species, such as Elaeis oleifera, Attalea phalerata, and Astrocaryum murumuru, which are indicators of anthropogenic soils along the Madeira River (Junqueira et al., 2011).
Synthesis
As a synthesis of the information obtained about these eight management practices, their interactions and how each practice affects natural ecological processes, we present a new conceptual model that explains the process of Amazonian forest domestication. Following Goldberg et al. (2016), we describe a temporal continuum from the late Pleistocene until today. We also present spatial gradients from settlements through swiddens to domesticated forests, and from old-growth forests to domesticated forests, illustrating at which distances from settlements these different practices operate to form domesticated forests with different degrees of human intervention. Although Goldberg et al. (2016) modeled human population dynamics during the Holocene without data from Central Amazonia, this model is the only one available describing a temporal continuum of past human population in South America. We considered a temporal dynamic that starts in the Pleistocene when humans arrived, and follows human population growth rates during the Holocene (Goldberg et al., 2016). In our conceptual model, we considered pristine forests to exist when humans had not yet altered natural ecological processes (Denevan, 1992). Pristine forests were the norm during the Pleistocene and, with at least 13,000 years of growing human populations across the Amazon basin, pristine forests gradually disappeared (Clement et al., 2015) and old-growth forests—mature forests without recent human interference, but not necessarily pristine (Wirth et al., 2009)—cover most of the basin today.
Field Surveys
All authorizations to conduct the study were obtained before field work. The study was approved by the Brazilian Ethics Committee for Research with Human Beings (Process n° 10926212.6.3001.5020, 2013), the Federation of the Indigenous Organizations of the Negro River–FOIRN and the Regional Coordinator of the Brazilian National Indigenous Foundation - FUNAI, and the Brazilian System of Protected Areas (SISBIO, process n° 47373-1, 2014). In each village, we obtained the informed consent of each local traditional or indigenous leadership at the beginning of the study.
In the field, we studied 30 contemporary villages settled on river banks distributed in nine sub-basins of four major rivers (Madeira, Solimões, Negro, Tapajós) across Brazilian Amazonia (see Supplementary Table 2 for names of the villages visited and their distances to archaeological sites). We visited from 2 to 10 villages in each sub-basin and selected villages located on or near archaeological sites with TPI. Archaeological sites with anthropogenic soils are ancient sedentary settlements (Neves et al., 2003), and they were chosen for our study because they indicate long-term human occupation, where rich soils, new landforms and domesticated plants accumulated through time in response to human agency (Clement et al., 2015). In each village, from March 2013 until March 2015 (3 months per year during the rainy season), we searched for indigenous and traditional ecological knowledge about the forest patches dominated by useful plant species in the surroundings of these villages.
Of the 30 contemporary villages along river banks, 27 are currently inhabited by traditional peoples (ribeirinhos) that have lived there for at least one generation; most of them are descendants of migrants who intermarried with local indigenous peoples. Their daily activities include farming, fishing, hunting, timber, and non-timber forest product extraction, and two villages are involved in community-based tourism. Three villages in the upper Negro River are inhabited by members of the Baré indigenous group, descendants of Arawak speaking groups, who lost their original language and adopted the Tupi-based Nheengatu, taught by the missionaries.
In each village, we searched for patches of native forest species used mainly as food resources. We focused on edible fruits because previous studies showed that these resources accumulated around ancient indigenous villages (Frikel, 1978; Balée, 1989, 1993). We interviewed 56 local people (on average 2 per village) regarding the occurrence and distribution of these forest patches, and used participatory mapping techniques (Gilmore and Young, 2012) to locate these patches around the villages. We used the suffix “zal” or “al,” which means abundance, aggregation or patches in Portuguese, and “tíwa” (in the Nheengatu language) to communicate with local people. These terms are used by contemporary people that associate the suffix with the name of the dominant species and identify a forest patch of useful species based on their traditional knowledge. For instance, a patch of bacaba palm (Oenocarpus bacaba/O. distichus) is named a bacabal in Portuguese and a iwakátíwa in Nheengatu. All patches of useful species were mapped with participatory mapping and complemented with the information collected during guided tour (Gilmore and Young, 2012; Albuquerque et al., 2014). Participatory mapping techniques are used to map local knowledge about the landscape, and to translate indigenous and local representations into techno-scientific language (Chapin et al., 2005; Heckenberger, 2009; Gilmore and Young, 2012). All local residents were invited to participate in a participatory mapping workshop that occurred during one morning or afternoon in each village. People were encouraged to draw and identify first the main local rivers, second TPI sites, and third different patches of useful species on maps made with georeferenced grids on top of recent cloud-free LANDSAT TM images of the area. With participatory mapping, we obtained the approximate location and size of TPI sites, and patches of useful species surrounding the villages. With guided tour we validated the location of at least one TPI site and/or one patch of useful species per village. Village members chose one person to guide us and visit the most accessible forest and TPI site. During the guided tour, we collected geographical coordinates of TPI sites and useful forest patches, and documented all useful species observed according to local knowledge. The botanical species were pre-identified in the field using some books of fruit trees and palms (Henderson, 1995; Cavalcante, 2010), and when possible, botanical material was also collected for final identification. The botanical identification was confirmed by José Ramos, a parataxonomist at INPA (Instituto Nacional de Pesquisas da Amazônia). Some plants were only identified to genus level in the field due to logistical limitations. The distribution of all forest patches identified around the villages was documented during the interviews, participatory mapping and guided tour. In total, we studied 21 patches visited with local informants dominated by 14 different useful species, as some patches visited concentrate the same dominant species. Forest patches are located up to 5 km from archaeological sites, and we documented a minimum of four useful species, a maximum of 21, and median of seven useful species per patch. In each of the nine sub-basins visited in the field, we documented a minimum of six useful forest patches dominated by different species, a maximum of 14 and a median of nine patches.
We compared our results obtained from field surveys and the literature review with field data from two villages along the right margin of the lower Tapajós River, where we documented all management practices performed by local people with the species that dominate local forest patches. This comparison served as ground-truth for our conceptual model. During free listing interviews and guided tour (Albuquerque et al., 2014) local informants described practices with which they benefit useful species found in patches of this sub-basin. In January and February of 2015, we interviewed 33 informants who know and use forest species in Maguarí and Jamaraquá villages in the Tapajós National Forest (FLONA). We also walked approximately 80 km along trails in the FLONA Tapajós with the seven most experienced informants to identify useful species in the forest. During these guided tour, the informants explained how they manage the useful species found in forest patches. With information about how local residents manage useful species, we compared the number and frequency of the practices obtained in the field with the same information obtained from the literature review.
We used ArcGis software to map the information collected in the field with participatory mapping and GPS. The closest (minimum distance) and longest (maximum distance) linear distances from each patch of useful species to the closest TPI were calculated manually using a digital ruler. We calculated the frequency of forest patches that occur at intervals of a minimum distance of 1 km to the nearest TPI. Using the minimum distance from forest patches to the closest TPI sites, we compared the spatial gradient of our conceptual model (settlements, swiddens or old-growth forests) with the location of the forest patches found in the field: patches on top of TPI sites were associated with pre-Columbian settlements, those located in fallows close to TPI sites were associated with past swiddens, and forest patches more distant from TPI sites were associated with old-growth forests, and confirmed by local knowledge and the presence of large trees.
Results
A Conceptual Model of Forest Domestication in Amazonia
Our conceptual model shows how pristine forests were converted into domesticated forests by a long-term process involving the interaction between eight human management practices (Figure 1). The conceptual model presents three general aspects of the forest domestication process: (1) a time span since the Pleistocene (Figure 1A); (2) interactions among human practices (arrows in Figure 1B); and (3) a spatial zone of influence for each management practice (arrows in Figure 1C). First, our model proposes that the frequency of these management practices increases with human population in South America (Goldberg et al., 2016), resulting in more extensive domestication of Amazonian forests through the Holocene (Figure 1A). Second, each arrow presented in our conceptual model indicates interactions among a pair of categories of management, showing that one practice can positively affect others (Figure 1B). For instance, humans remove non-useful plants (Practice 1–P1) while often selectively protecting useful individuals with desirable phenotypes (P5), or plant selected individuals (P5) in forest gaps (natural or created by humans–P1), swiddens and homegardens (P7). Native Amazonians protect plants (P2) as sources of seeds for future planting (P7) and selection (P5), and also to attract animal dispersers (P3). A gradual transformation of the forest is expected to occur by the interaction between humans (P4) and non-human dispersers (P3). Seeds and seedlings of selected useful plants (P5) are transported by humans from natural to domesticated landscapes (P4), guaranteeing their planting and propagation (P7). Fire management (P6) is often used in association with protection of species (P2) with plants previously selected for traits of interest (P5). The combination of fire management (P6) with the protection of certain species (P2) in domesticated landscapes may allow even useful fire-sensitive plants to form patches in ancient cultivated systems. Ancient planting practices (P7) attract dispersers (humans and non-humans; P3 and P4) and improve soil conditions (P8). The planting of useful edible trees (P7) attracts game animals that may disperse their seeds throughout the area (P3), thus increasing the abundance of the species locally. Indigenous people disperse seeds of plants (P4) and plant them in agroforestry systems and along forest trails (P7) when they move from one place to another, increasing food availability during long walks in the forests. Trees planted in agroforestry systems (P7) may enrich soil fertility (P8), reproducing the nutrient-conservation mechanism observed in the forest. By improving naturally nutrient-poor soils (P8), pre-Columbian societies enhanced food production in Amazonian landscapes, also allowing their population expansion.
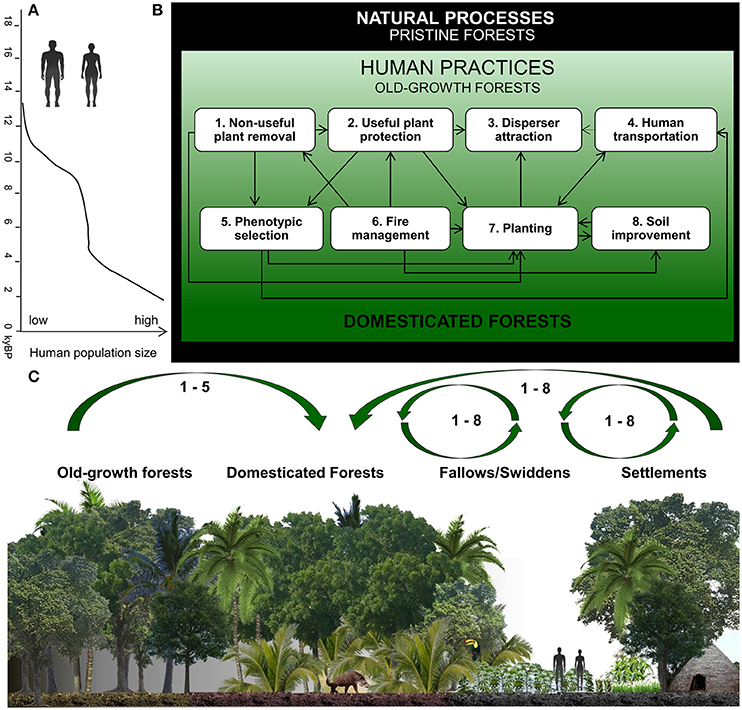
Figure 1. Conceptual model illustrating the interaction of eight management practices and their effects on the domestication of forests through time. (A) Expected trends in human population growth rate in Amazonia from fourteen to two thousand years ago before present (kyBP) based on published data for South America outside of Amazonia (adapted from Goldberg et al., 2016). (B) Management practices (1–8), their interactions and their effects on the forest domestication process through time [from top (16 kyBP) to bottom (0 kyBP)]. Natural ecological processes operate during all moments in time and along a domestication gradient from pristine to domesticated forests. Management practices may have a positive direct effect (dark arrows) or hypothetical positive effect (light arrow) on other practices that intensify as human population increases (from light green to dark green). (C) The forest domestication process in a spatial context of human influence from settlements, through swiddens, domesticated forests to old-growth forests, which may have been domesticated in the past, but lack recent human intervention. Domesticated forests can originate (arrows) from settlements and swiddens, or from old-growth forests. Our model describes an open-ended process.
Third, the gradient of soil improvement is illustrated in the spatial representation in our conceptual model (Figure 1C). Five practices, removal of non-useful plants (P1), protection of useful plants (P2), attraction of non-human dispersers of useful plants (P3), human transportation of useful plants (P4), and selection of phenotypes useful to humans (P5) occur across the entire gradient of human influence from settlements, through swiddens, to domesticated forests to old-growth forests. Fire management (P6), direct planting (P7), and soil improvement (P8) are practices mainly used in swidden/fallows and settlements, giving rise to domesticated forests with useful plants related to these activities.
Relationships among Management Practices: Evidence from the Literature and Field
We found that all eight categories of management practices described in the literature (Table 1) are also known by traditional people in the two villages along the lower Tapajós River that we studied (Figure 2). Transportation of plants by humans, planting of useful plants and selection of desirable phenotypes were the most frequent practices in the literature, whereas clearing the understory, cutting lianas and weeding (P1-removal of non-useful plants) and not cutting useful plants (P2-protection of the useful) were the most cited practices in field interviews (Figure 2). Attraction of dispersers and soil improvement were the least frequent practices in the literature and field interviews, documented for less than 40% of the species investigated.
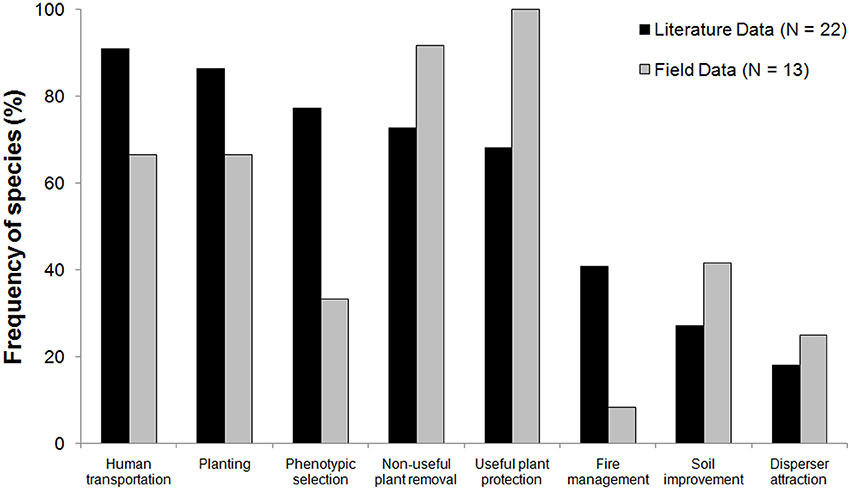
Figure 2. Frequency of useful species involved in each management practice based on the literature (black bars) and field interviews (gray bars). Information for 22 species was obtained from the literature and for 13 species in the field in two villages.
More than half of the useful plant species investigated in the literature and the field are managed with at least five practices. Based on the literature, four species (A. maripa, C. villosum, M. flexuosa, T. cacao) are managed with seven practices, and for these species at least five different uses were reported (Supplementary Table 1). Based on field data, two species (C. villosum and E. uchi) are managed with seven practices and used for several purposes, such as food, medicine and hunting (Supplementary Table 1). Local people reported that they do not clear the land or use fire in places where aggregated patches of these species occur, with the purpose of protecting the whole population. One species, M. splendens, with only two uses reported in the literature (manufacturing and fuel), is managed with only one practice (P6 - fire management) based on the literature.
Multi-species Patches of Useful Plants
We found multiple forest patches of useful species surrounding the 30 contemporary villages visited in Amazonia (Figure 3). In total, people cited 35 patches with different names and corresponding to 38 useful species (Supplementary Table 1). The most common patches were açaízal (E. precatoria), babacal (O. bacaba), castanhal (B. excelsa), piquiázal (C. villosum), patauázal (O. bataua), and uxízal (E. uchi) (Figure 4). Most patches are common in more than one sub-basin visited and a few patches are only common in one sub-basin visited; some examples of localized patches are cf. Neoxythece elegans in the lower Madeira River basin, Duguetia stenantha in the upper Solimões River basin, H. balsamifera in the upper Negro River basin, and Hymenea parvifolia in the lower Tapajós River basin. Detailed information of the regional differences of forest patches across Amazonia is given in Supplementary Tables 1, 3. Of all species that dominate the patches, 90% are used for more than one purpose (Figure 3).
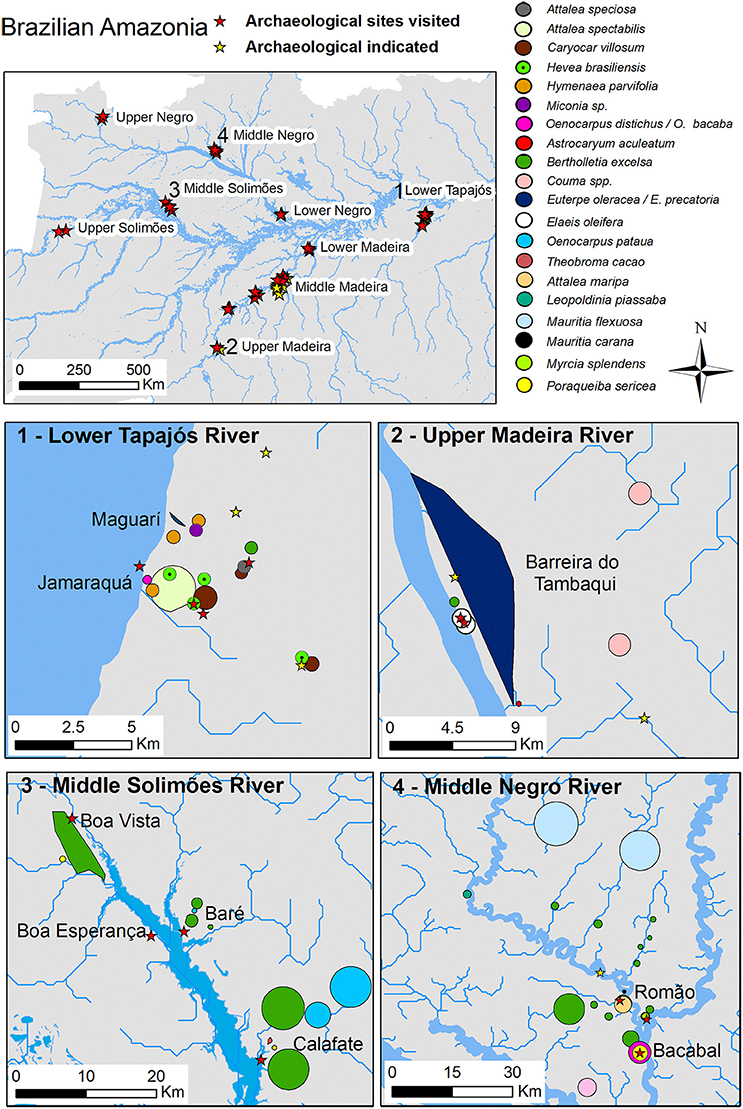
Figure 3. Maps of examples of useful forest patches around archaeological sites in four sub-basins of Brazilian Amazonia. Different sizes and shapes of forest patches presented in the figures are based on local knowledge descriptions and local drawings. See Supplementary Table 1 for more information about the forest patches presented in this figure. Archaeological sites are ancient sedentary settlements with anthropogenic soils (TPI) and have been re-occupied by contemporary peoples.

Figure 4. Forest patches of useful species found in nine Amazonian sub-basins. Shades of gray indicate the frequency of citation in each sub-basin (very light gray–1 to black–7 citations). The total number of uses was obtained from both the literature review and field interviews. See Supplementary Table 1 for more information on the forest patches and uses attributed to each species.
Although forest patches are dominated by one species after which they are named, they concentrated multiple useful species that dominate forest patches in different sub-basins of Brazilian Amazonia (Table 2). We visited 21 patches that are dominated by 14 out of 38 useful species that form patches across the basin. Palm species of the genus Oenocarpus occur in 75% of the 21 forest patches visited across the basin. We found regional differences in the composition of useful palm species that occur in the forest patches: A. maripa were found in most patches of the Madeira River basin, E. precatoria of the Solimões River basin, O. bataua of the Negro River basin and O. distichus of the Tapajós River basin. Forest patches dominated by B. excelsa species are the most common and the most diverse patches: they concentrate 5–8 useful species that also are dominant species in other forest patches in different parts of the basin (Figure 4 and Table 2). In total, 87 useful species were cited in the patches visited (Supplementary Table 3) and the number of useful species cited increases with the number of patches visited (Supplementary Figure 1).
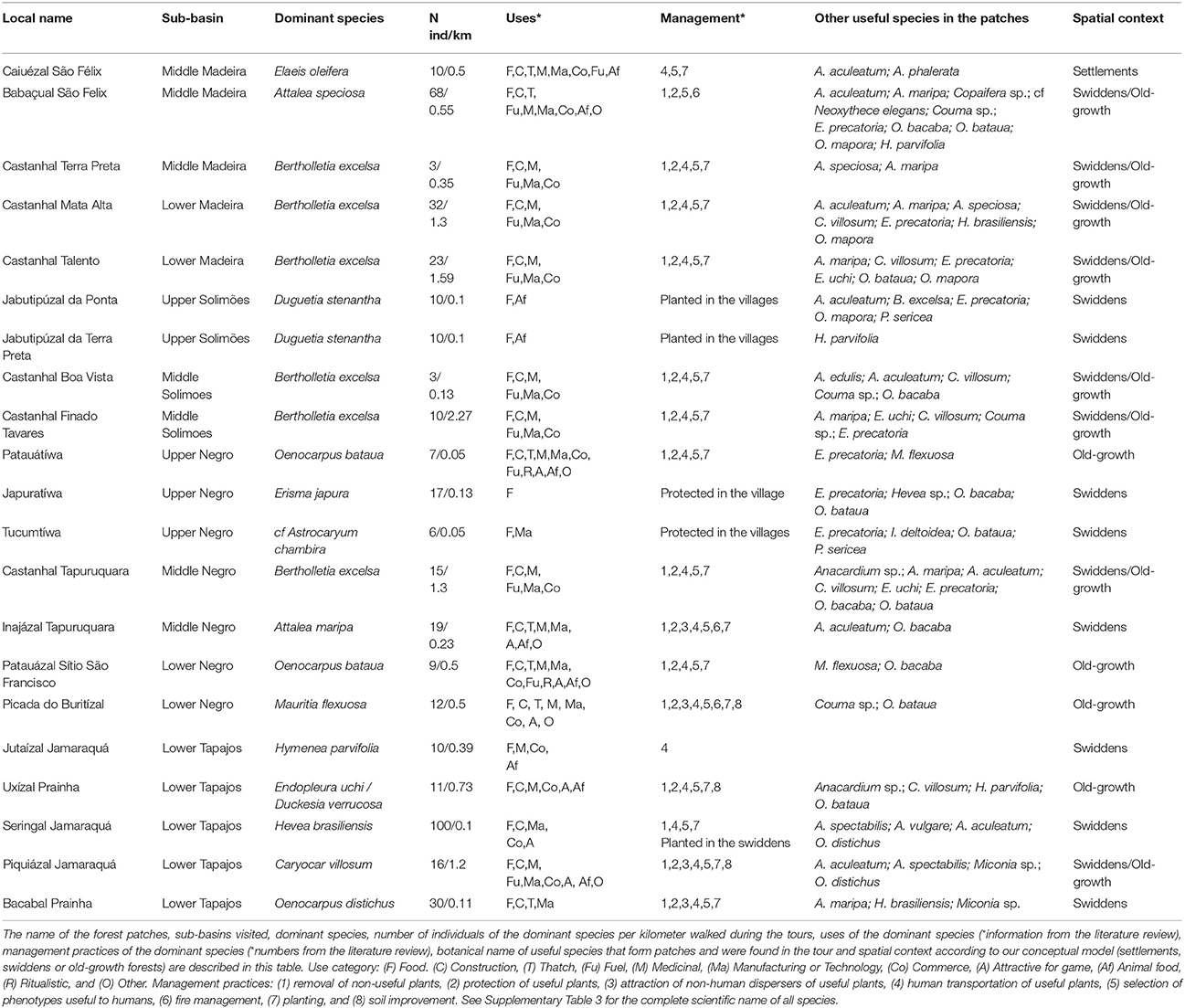
Table 2. List of useful species that occur in the 21 forest patches visited during guided tour.
Most patches are small in size (less than 1 km2), and occur at various distances from archaeological sites (0–40 km), implying that they may have originated from all spatial contexts: settlements, old swiddens, or old-growth forests (Figure 5). Few patches are restricted to TPI sites and old villages. Half of all patches are located up to 1 km from the archaeological sites, although some patches can be found up to 40 km away from these sites (Figure 5 and Supplementary Figure 2). As a common pattern and according to local people, patches dominated by useful palm species are more common in valley forests, whereas patches dominated by tree species occur commonly in other environmental settings, such as plateau forests and white-sand forests (campinaranas).
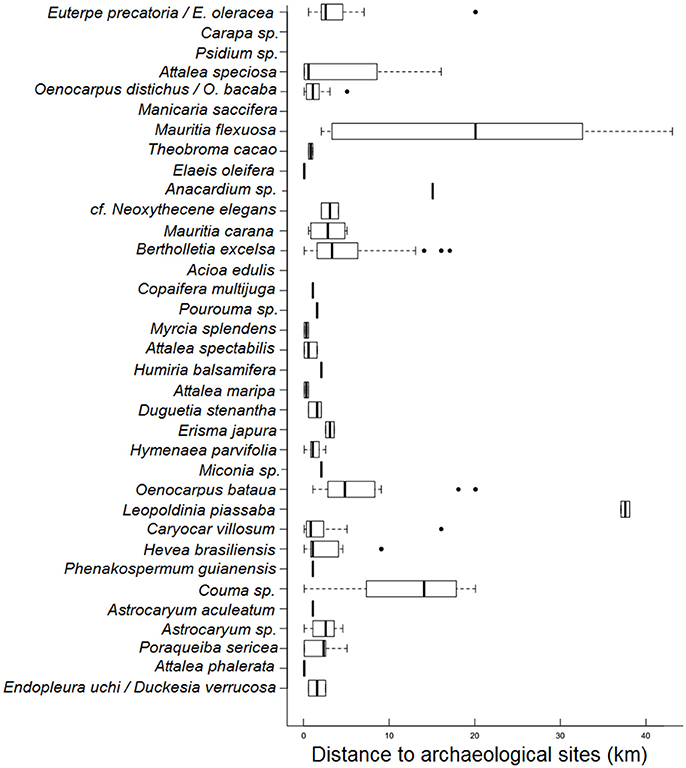
Figure 5. Occurrences of patches of useful species along a distance gradient from archaeological sites. Median (dark line), first and third quartile (rectangles), minimum and maximum distances (dotted line) from the forest patches to the closest archaeological site are presented. Archaeological sites are ancient sedentary settlements with anthropogenic soils (TPI) and have been re-occupied by contemporary peoples. Black dots are extreme values (outliers). No data is available for three species because people couldn't determine the location of these patches in the maps we used.
Discussion
Based on our multidisciplinary approach, we provide a framework for understanding how human practices have led to the formation of patches of useful perennial plant species across Amazonian forests. Our conceptual model portrays how Amazonian peoples manage forests in multiple ways through eight categories of management practices that interfere with natural ecological processes and promote domesticated forests around human settlements. The similarities between ethnographic descriptions of management practices across the basin and our field observations of two villages indicate the commonness of these practices, suggesting that pre-Columbian and contemporary peoples transformed forest composition at varying distances from their settlements by multiple management practices. In the field, we confirmed that multiple diverse patches of useful species, currently managed by indigenous and traditional peoples, occur mainly near these settlements. Overall, our results support the view that these diverse patches of useful plant species were created and maintained by human actions.
Our conceptual model also reflects positive long-term interactions between humans and plants (Smith, 2011), as described in other tropical regions worldwide (Wiersum, 1997a; Michon, 2005; Kennedy, 2012; Reis et al., 2014; Boivin et al., 2016; Roberts et al., 2017). Previous models had suggested that the plant and forest domestication processes are associated with the cultivation of domesticated tree crops (Wiersum, 1997a,b). Although our model is inspired by previous studies (Harris, 1989; Wiersum, 1997a,b), we present a new framework to understand the domestication of Amazonian forests that simplifies the complex network of interactions between human actions and natural ecological processes. Because these interactions cannot be understood by separately assessing only individual management practices or species, the intricate groups of management practices shown in our model illustrate how multiple human actions interact to shape Amazonian forests. Species-specific details are scattered in the literature, and here we synthesized this information into a single model that can be tested with individual site-specific situations.
In our model, forest domestication is defined as an open-ended process (Rival, 2007; Kennedy, 2012), in which domesticated forests can originate through varying degrees of human intervention from settlements and swiddens, and also from old-growth forests. This perspective makes the typical distinction between hunter-gatherers vs. farming groups inappropriate for the Amazonian context (Terrell et al., 2003; Kennedy, 2012), as most ancient Native Amazonians (often characterized as hunter-gatherers) were actually practicing many activities, including planting tree species (Frikel, 1978). Amazonian forests that were once cultivated and domesticated are often transformed into swiddens or settlements as a cyclic pattern that has also been observed in Indonesian forests (Michon, 2005). Because early successional species usually depend on forest gaps for recruiting, they are maintained with management practices, similar to fully domesticated plant populations that require human care for survival and reproduction (Clement, 1999).
Although it is likely that current management practices maintain the legacy of past societies (Junqueira et al., 2017), the effects of past forest domestication have been detected in forests even without recent management activities (Van Gemerden et al., 2003; Dambrine et al., 2007; Ross, 2011; Levis et al., 2017b). The persistent effect of pre-Columbian plant domestication on modern forest composition has been revealed in Amazonian old-growth forests (Junqueira et al., 2017; Levis et al., 2017b), secondary forests (Junqueira et al., 2010) and even in highly dynamic homegardens growing in archaeological sites (Lins et al., 2015). Domesticated species adapted to stable soil conditions created by management practices, such as TPI, may persist for a long time after abandonment (Quintero-Vallejo et al., 2015). This may explain why domesticated palms dominate modern forests growing on pre-Columbian mounds, anthropogenic soils and geoglyphs abandoned more than 400 years ago (Erickson and Balée, 2006; Quintero-Vallejo et al., 2015; Watling et al., 2017b). Another possible explanation for this persistence is the continuous recruitment of useful and domesticated plants present in the forest seed bank (Lins et al., 2015). Pre-Columbian peoples may also have played a major role in disseminating large multi-seeded fruits within and across Neotropical biomes during the Holocene, resulting in the spread of diverse patches of useful plants associated with human settlements and trails (Guix, 2005). Human-mediated dispersal of invasive plants is well-documented (Hodkinson and Thompson, 1997; Nathan et al., 2008); however, ecological studies frequently overlook this mechanism when considering native species (Levis et al., 2017a).
Modern Amazonian peoples who live on pre-Columbian settlements seem to have inherited indigenous knowledge, including these management practices that benefit useful and domesticated plant populations. Our field data show that most useful species dominant in forest patches occur in more than one sub-basin visited, suggesting a widespread use and management of forest resources by past and contemporary peoples. The forest domestication process was assimilated by contemporary societies through the transmission of indigenous knowledge from one generation to another, as described for indigenous groups from Ecuadorian Amazonia (Zurita-Benavides et al., 2016) and traditional people in Brazilian Amazonia (Alves et al., 2016). Villages with homegardens that were occupied by several pre-Columbian cultures contain a higher beta diversity of useful plants compared to villages with homegardens occupied by a single culture (Lins et al., 2015), suggesting that previously existing useful plants were incorporated into new agroforestry systems when old villages are re-occupied (Miller and Nair, 2006). Some practices, however, have changed in intensity and extension through time. Slash-and-burn agriculture, for instance, has increased since the arrival of European societies that introduced metal tools to cut down the forest (Denevan, 2001). In pre-Columbian times, sedentary societies frequently improved soil conditions by managing fire in their habitation and cultivation zones (Denevan, 2001; Neves et al., 2003; Woods et al., 2013). Sedentary societies with high human population densities were responsible for the formation of anthropogenic soils that are no longer being created on a broad scale (Neves et al., 2003). These same anthropogenic soils, however, are widely used by modern societies to cultivate crops, allowing the diversification and intensification of food production in Amazonia (Woods et al., 2013; Junqueira et al., 2016b).
Amazonian societies managed fire, planted useful species and improved soils that resulted in substantial transformation in forests close to their homes. Although some scholars argue for a localized impact involving these three practices in pre-Columbian Amazonia, associating them with the margins of the main rivers (McMichael et al., 2012, 2014; Bush et al., 2015; Piperno et al., 2015), the impact of long-term management practices has been detected in the forests of interfluvial areas (Levis et al., 2012; Franco-Moraes, 2016; Watling et al., 2017b) and across the Amazon basin (Levis et al., 2017b). These findings suggest that even in remote areas, far from known archaeological sites, contemporary people also manage the forest, protecting useful species and removing the non-useful, which are the most frequent practices reported by contemporary societies. Logistical limitations constrain our ability to detect the long-term effects of these practices away from current human settlements (Stahl, 2015), and even the participatory techniques used in this study are based on current knowledge about the forest, requiring ethnographic projection to infer the impact of past peoples. For instance, patches of rubber tree (Hevea brasiliensis) have been managed by modern societies driven by economic interest since the mid-nineteenth century (Schroth et al., 2003), but were probably managed differently before that time. Although several socio-economic factors push contemporary peoples to concentrate their activities on market-oriented forest resources (Jakovac et al., 2016b), they occasionally use and manage forest patches located up to 40 km from their villages for hunting animals and gathering fruits (Figure 5; Franco-Moraes, 2016). As an alternative approach, the abundance and richness of useful plants, especially of domesticated species, might be used to predict the location of ancient human settlements in these remote Amazonian areas (Levis et al., 2017b).
Future multidisciplinary studies that combine alternative methods may help to reconstruct forest composition dynamics (Stahl, 2015), as Watling et al. (2017b) did in the geoglyph region of Acre, revealing more details of the influence of past peoples in Amazonian forests. The integration of paleoecology, archaeology, archaeobotany and forest ecology is a promising combination (Mayle and Iriarte, 2014; Iriarte, 2016; Watling et al., 2017a,b). In southwestern Amazonia, archaeobotanical remains have revealed that past peoples consumed a rich diet, including many palm fruits (Dickau et al., 2012). The increase in palm abundance is also visible in soil profiles of archaeological sites across the region (McMichael et al., 2015; Watling et al., 2017b), suggesting that past societies enriched the forest with useful palms to improve food production. Today, useful and domesticated palms are dominant in southwestern Amazonian forests (Levis et al., 2017b), growing on abandoned pre-Columbian mounds, anthropogenic soils and geoglyphs created by past management practices (Erickson and Balée, 2006; Quintero-Vallejo et al., 2015; Levis et al., 2017b; Watling et al., 2017b). Many palm species were found in most of the forest patches investigated here, suggesting long-term human management. Regional contrasts in palm and other plant species composition across Amazonia may reveal different human practices or specific environmental conditions that should be investigated in detail.
We conclude that our literature review, conceptual model and field results contribute to explain how domesticated forests were formed in Amazonia, in part by revealing how integrated categories of management practices interfere with natural ecological processes that shape plant communities in tropical forests. Different degrees and types of management, cultural preferences and environmental conditions may lead to a wide variety of outcomes and explain why diverse combinations of useful species were found in Amazonian forest patches. Insights from agroforestry systems in tropical and sub-tropical regions confirm that indigenous management practices have been used worldwide to domesticate plant species and entire forest landscapes (Wiersum, 1997a,b; Michon, 2005; Kennedy, 2012; Reis et al., 2014). Learning about indigenous knowledge of forest management is important not only to understand the plant and landscape domestication processes, but also to guide policies for forest conservation, local people's empowerment, and food production (Michon et al., 2007; Roberts et al., 2017). In Amazonia today, millions of people live in rural landscapes, with partial dependence on forest resources for their well-being, and with profound local knowledge that should be incorporated in environmental conservation and management plans.
Author Contributions
CL conceived the study; CL, BF, PM, BL, RA, JF-M, JL, and EK collected data; CL, BF, PM, BL, RA, JF-M, JL, EK, FB, MP-C, FC, and CC designed the analyses; CL, BF, PM, BL, RA, JF-M, JL, and EK performed the analyses; CL, BF, PM, BL, RA, JF-M, JL, EK, FB, MP-C, FC, and CC discussed further analyses; CL, BF, PM, BL, RA, JF-M, JL, FB, MP-C, FC, and CC wrote the manuscript.
Funding
Fundação de Amparo a Pesquisa do Estado do Amazonas - FAPEAM Universal proc. no. 3137/2012 and 062.03137/2012; Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq Universal proc. no. 473422/2012-3 and 458210/2014-5.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer BS and handling Editor declared their shared affiliation.
Acknowledgments
We thank local residents for their participation, the Instituto de Desenvolvimento Agropecuário e Florestal do Amazonas, the Centro Estadual de Unidades de Conservação do Amazonas, the Instituto Chico Mendes de Conservação da Biodiversidade, the Instituto de Desenvolvimento Sustentável Mamirauá, the Instituto Socioambiental de São Gabriel da Cachoeira, the Cooperativa Mista da Flona do Tapajós and the Federação das Organizações Indígenas do Rio Negro for field assistance, and Sara Deambrozi Coelho for information about the uses of species. CL thanks CNPq for a doctoral scholarship, RA and JL thank INPA and CNPq for research scholarships, JF-M thanks CNPq for a master's scholarship, FC and CC thank CNPq for research fellowships, BF thanks São Paulo Research Foundation (FAPESP) for grant #2016/25086-3. BL thanks FAPEAM for research fellowship and FAPESP for grant #2015/24554-0.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2017.00171/full#supplementary-material
Footnotes
1. ^https://pib.socioambiental.org/files/file/PIB_institucional/No_Brasil_todo_mundo_é_índio.pdf
2. ^http://www.planalto.gov.br/ccivil_03/_Ato2007-2010/2007/Decreto/D6040.htm
References
Albert, B., and Milliken, W. (2009). Urihi A: a Terra-floresta Yanomami. São Paulo: Instituto Socioambiental.
Albuquerque, U. P., Ramos, M. A., Lucena, R. F. P., and de Alencar, N. L. (2014). “Methods and techniques used to collect ethnobiological data,” in Methods and Techniques in Ethnobiology and Ethnoecology, eds U. P. Albuquerque, L. V. F. C. da Cunha, R. F. P. de Lucena, and R. R. N. Alves (New York, NY: Springer), 15–37.
Alves, R. P., Levis, C., and Clement, C. R. (2016). Use and management of piquiá suggest in situ domestication a long the lower Tapajós river, Brazilian Amazonia. Econ. Bot. 70, 198–202. doi: 10.1007/s12231-016-9340-4
Anderson, A. B., May, P. H., and Balick, M. J. (1991). The Subsidy from Nature: Palm Forests, Peasantry, and Development on an Amazon Frontier. New York, NY: Columbia University Press.
Arroyo-Kalin, M. (2012). Slash-burn-and-churn: landscape history and crop cultivation in pre-Columbian Amazonia. Quat. Int. 249, 4–18. doi: 10.1016/j.quaint.2011.08.004
Bagchi, R., Gallery, R. E., Gripenberg, S., Gurr, S. J., Narayan, L., Addis, C. E., et al. (2014). Pathogens and insect herbivores drive rainforest plant diversity and composition. Nature 506, 85–88. doi: 10.1038/nature12911
Balée, W. (1989). “The Culture of Amazonian Forests,” in Resource Management in Amazonia: Indigenous and Folk Strategies, eds D. A. Posey and W. Balée (Bronx, NY: NYGB Press), 1–21.
Balée, W. (1993). Indigenous transformation of Amazonian forests: an example from Maranhão, Brazil. L'Homme 33, 231–254. doi: 10.3406/hom.1993.369639
Balée, W. (2000). Antiquity of traditional ethnobiological knowledge in Amazonia: the Tupí-Guaraní family and time. Ethnohistory 47, 399–422. doi: 10.1215/00141801-47-2-399
Balée, W. (2006). The research program of historical ecology. Annu. Rev. Anthr. 35, 75–98. doi: 10.1146/annurev.anthro.35.081705.123231
Balée, W. (2013). Cultural Forests of the Amazon: a Historical Ecology of People and their Landscapes. Tuscaloosa, AL: The University of Alabama Press.
Barberis, I. M., and Tanner, E. V. J. (2005). Gaps and root trenching increase tree seedling growth in Panamanian semi-evergreen forest. Ecology 86, 667–674. doi: 10.1890/04-0677
Boivin, N. L., Zeder, M. A., Fuller, D. Q., Crowther, A., Larsong, G., Erlandsonh, J. M., et al. (2016). Ecological consequences of human niche construction: examining long-term anthropogenic shaping of global species distributions. Proc. Natl. Acad. Sci. U.S.A. 113, 6388–6396. doi: 10.1073/pnas.1525200113
Bond, W. J., and Midgley, J. J. (2001). Ecology of sprouting in woody plants: the persistence niche. Trends Ecol. Evol. 16, 45–51. doi: 10.1016/S0169-5347(00)02033-4
Boster, J. S. (1984). “Classification, cultivation and selection of Aguaruna cultivars of Manihot esculenta (Euphorbiaceae),” in Ethnobotany in the Neotropics, eds G. T. Prance and J. A. Kallunki (Bronx, NY: NYBG Press), 34–47.
Bowman, D. M. J. S., Balch, J., Artaxo, P., Bond, W. J., Cochrane, M. A., D'Antonio, C. M., et al. (2011). The human dimension of fire regimes on Earth. J. Biogeogr. 38, 2223–2236. doi: 10.1111/j.1365-2699.2011.02595.x
Bush, M. B., McMichael, C. H., Piperno, D. R., Silman, M. R., Barlow, J., Peres, C. A., et al. (2015). Anthropogenic influence on Amazonian forests in pre-history: an ecological perspective. J. Biogeogr. 42, 2277–2288. doi: 10.1111/jbi.12638
Campbell, D. G., Ford, A., Lowell, K. S., Walker, J., Lake, J. K., Ocampo-Raeder, C., et al. (2006). “The feral forests of the eastern Petén,” in Time and Complexity in Historical Ecology: Studies in the Neotropical Lowlands, eds W. L. Balée and C. L. Erickson (New York, NY: Columbia University Press), 21–55.
Chapin, M., Lamb, Z., and Threlkeld, B. (2005). Mapping indigenous lands. Annu. Rev. Anthropol. 34, 619–638. doi: 10.1146/annurev.anthro.34.081804.120429
Clement, C. R. (1999). 1492 and the loss of Amazonian crop genetic resources. I. The relation between domestication and human population decline. Econ. Bot. 53, 188–202. doi: 10.1007/BF02866498
Clement, C. R., de Cristo-Araújo, M., Coppens D'Eeckenbrugge, G., Alves Pereira, A., and Picanço-Rodrigues, D. (2010). Origin and domestication of native Amazonian crops. Diversity 2, 72–106. doi: 10.3390/d2010072
Clement, C. R., Denevan, W. M., Heckenberger, M. J., Junqueira, A. B., Neves, E. G., Teixeira, W. G., et al. (2015). The domestication of Amazonia before European conquest. Proc. R Soc. B 282:20150813. doi: 10.1098/rspb.2015.0813
Clement, C. R., Klüppel, M. P., German, L. A., Almeida, S. S., de Major, J., Aragão, L. eC de, et al. (2009). “Diversidade Vegetal em Solos Antrópicos da Amazônia,” in As Terras Pretas de Índio da Amazônia: Sua Caracterização e uso Deste Conhecimento na Criação de Novas Áreas, eds W. G. Teixeira, D. C. Kern, B. E. Madari, H. N. Lima, and W. I. Woods (Manaus: Embrapa Amazônia Ocidental), 146–161.
Coley, P. D. (1983). Herbivory and defensive characteristics of tree species in a lowland tropical forest. Ecol. Monogr. 53, 209–234. doi: 10.2307/1942495
Connell, J. H. (1978). Diversity in tropical rain forests and coral reefs. Science 199, 1302–1310. doi: 10.1126/science.199.4335.1302
Connell, J. H., and Lowman, M. D. (1989). Low-diversity tropical rain forests: some possible mechanisms for their existence. Am. Nat. 134, 88–119. doi: 10.1086/284967
Dambrine, E., Dupouey, J. L., Laüt, L., Humbert, L., Thinon, M., Beaufils, T., et al. (2007). Present forest biodiversity patterns in France related to former Roman agriculture. Ecology 88, 1430–1439. doi: 10.1890/05-1314
Denevan, W. M. (1992). The pristine myth: the landscape of the Americas in 1492. Ann. Assoc. Am. Geogr. 82, 369–385. doi: 10.1111/j.1467-8306.1992.tb01965.x.
Denevan, W. M. (1995). 2 Prehistoric agricultural methods as models for sustainability. Adv. Plant Pathol. 11, 21–43. doi: 10.1016/S0736-4539(06)80004-8
Denevan, W. M. (1996). A bluff model of riverine settlement in prehistoric Amazonia. Ann. Assoc. Am. Geogr. 86, 654–681. doi: 10.1111/j.1467-8306.1996.tb01771.x
Denevan, W. M. (2001). Cultivated Landscapes of Native Amazonia and the Andes. New York, NY: Oxford University Press.
Denevan, W. M., Treacy, J. M., Alcorn, J. B., Padoch, C., Denslow, J., and Paitan, S. F. (1984). Indigenous agroforestry in the Bora Indian management of swidden fallows. Interciencia 9, 346–357.
Dickau, R., Bruno, M. C., Iriarte, J., Prümers, H., Betancourt, C. J., Holst, I., et al. (2012). Diversity of cultivars and other plant resources used at habitation sites in the Llanos de Mojos, Beni, Bolivia: evidence from macrobotanical remains, starch grains, and phytoliths. J. Archaeol. Sci. 39, 357–370. doi: 10.1016/j.jas.2011.09.021
Elias, F., Marimon, B. S., de Almeida Reis, S. M., Forsthofer, M., Gomes, L., Morandi, P. S., et al. (2013). Dinâmica da distribuição espacial de populações arbóreas, ao longo de uma década, em cerradão na transição Cerrado-Amazônia, Mato Grosso. Biota Amaz. 3, 1–14. doi: 10.18561/2179-5746/biotaamazonia.v3n3p1-14
Eloy, L., and Emperaire, L. (2011). La circulation de l'agrobiodiversité sur les fronts pionniers d'Amazonie (région de Cruzeiro do Sul, état de l'Acre, Brésil). L'esp. Géographique 40, 62–74. doi: 10.3917/eg.401.0062
Erickson, C. L., and Balée, W. L. (2006). “The historical ecology of a complex landscape in Bolivia,” in Time and Complexity in Historical Ecology: Studies in the Neotropical Lowlands, eds W. L. Balée and C. L. Erickson (New York, NY: Columbia University Press), 187–233.
Fragoso, J., Silvius, K. M., and Correa, J. A. (2003). Long-distance seed dispersal by tapirs increases seed survival and aggregates tropical trees. Ecology 84, 1998–2006. doi: 10.1890/01-0621
Franco-Moraes, J. (2016). Ecologia Histórica De Florestas Da Bacia Do Rio Içana, Alto Rio Negro, Amazonas: Um Legado Baniwa Nas Paisagens. Master's Thesis, INPA, Manaus.
Fraser, J. A., Alves-Pereira, A., Junqueira, A. B., Peroni, N., and Clement, C. R. (2012). Convergent adaptations: bitter manioc cultivation systems in fertile anthropogenic dark earths and floodplain soils in Central Amazonia. PLoS ONE 7:e43636. doi: 10.1371/journal.pone.0043636
Frikel, P. (1978). Áreas de arboricultura pré agrícola na Amazônia: notas preliminares. Rev. Antropol. 21, 45–52.
Gilmore, M. P., and Young, J. C. (2012). The use of participatory mapping in ethnobiological research, biocultural conservation, and community empowerment: a case study from the Peruvian Amazon. J. Ethnobiol. 32, 6–29. doi: 10.2993/0278-0771-32.1.6
Goldberg, A., Mychajliw, A. M., and Hadly, E. A. (2016). Post-invasion demography of prehistoric humans in South America. Nature 532, 232–235. doi: 10.1038/nature17176
Guimarães, P. R. Jr., Galetti, M., and Jordano, P. (2008). Seed dispersal anachronisms: rethinking the fruits extinct megafauna ate. PLoS ONE 3:e1745. doi: 10.1371/journal.pone.0001745
Guix, J. C. (2005). Evidence of old anthropic effects in forests at the confluence of the Caurés and Negro Rivers, NW Amazonia: the role of indians and caboclos. Grup. Estud. Ecológicos 8, 1–27.
Harris, D. R. (1989). “An evolutionary continuum of people-plant interaction,” in Foraging and Farming: the Evolution of Plant Exploitation, eds D. R. Harris and G. Hilman (London: Unwin Hyman), 11–26.
Hart, T. B. (1990). Monospecific dominance in tropical rain forests. Trends Ecol. Evol. 5, 6–11. doi: 10.1016/0169-5347(90)90005-X
Hart, T. B., Hart, J. A., and Murphy, P. G. (1989). Monodominant and species-rich forests of the humid tropics: causes for their co-occurrence. Am. Nat. 133, 613–633. doi: 10.1086/284941
Heckenberger, M. J. (2009). Mapping indigenous histories: collaboration, cultural heritage, and conservation in the Amazon. Collab. Anthropol. 2, 9–32. doi: 10.1353/cla.0.0017
Heckenberger, M., and Neves, E. G. (2009). Amazonian archaeology. Annu. Rev. Anthropol. 38, 251–266. doi: 10.1146/annurev-anthro-091908-164310
Herrera, R., Jordan, C. F., Klinge, H., and Medina, E. (1978). Amazon ecosystems. Their structure and functioning with particular emphasis on nutrients. Interciencia 3, 223–231.
Hodkinson, D. J., and Thompson, K. (1997). Plant dispersal: the role of man. J. Appl. Ecol. 34, 1484–1496. doi: 10.2307/2405264
Hornborg, A. (2005). Ethnogenesis, regional integration, and ecology in prehistoric Amazonia: toward a system perspective. Curr. Anthropol. 46, 589–620. doi: 10.1086/431530
Howe, H. F., and Smallwood, J. (1982). Ecology of seed dispersal. Annu. Rev. Ecol. Syst. 13, 201–228. doi: 10.1146/annurev.es.13.110182.001221
Iriarte, J. (2016). “Investigating Amazonian dark earths as agro-ecosystems and their impact on the regional landscapes of the lower Amazon,” in Beyond Waters Archaeology and Environmental History of the Amazonian Inland, ed P. Stenborg (Gothenburg: University of Gothenburg), 71–86.
Jackson, G. (1974). Cryptogeal germination and other seedling adaptions to the burning of vegetation in savanna regions: the origin of the pyrophytic habit. New Phytol. 73, 771–780. doi: 10.1111/j.1469-8137.1974.tb01305.x
Jakovac, C. C., Bongers, F., Kuyper, T. W., Mesquita, R. C., and Peña-Claros, M. (2016a). Land use as a filter for species composition in Amazonian secondary forests. J. Veg. Sci. 27, 1104–1116. doi: 10.1111/jvs.12457
Jakovac, C. C., Peña-claros, M., Mesquita, R., Bongers, F., and Kuyper, T. W. (2016b). Swiddens under transition: consequences of agricultural intensification in the Amazon. Agr. Ecosyst. Environ. 218, 116–125. doi: 10.1016/j.agee.2015.11.013
Jordano, P. (2017). What is long-distance dispersal? And a taxonomy of dispersal events. J. Ecol. 105, 75–84. doi: 10.1111/1365-2745.12690
Junqueira, A. B., Levis, C., Bongers, F., Peña-Claros, M., Clement, C. R., Costa, F., and ter Steege, H. (2017). Response to comment on “persistent effects of pre-Columbian plant domestication on Amazonian forest composition”. Science 358. doi: 10.1126/science.aan8837
Junqueira, A. B., Shepard, G. H., and Clement, C. R. (2010). Secondary forests on anthropogenic soils in Brazilian Amazonia conserve agrobiodiversity. Biodivers. Conserv. 19, 1933–1961. doi: 10.1007/s10531-010-9813-1
Junqueira, A. B., Shepard, G. H., and Clement, C. R. (2011). Secondary forests on anthropogenic soils of the middle Madeira river: valuation, local knowledge, and landscape domestication in Brazilian Amazonia. Econ. Bot. 65, 85–99. doi: 10.1007/s12231-010-9138-8
Junqueira, A. B., Souza, N. B., Stomph, T. J., Almekinders, C. J., Clement, C. R., and Struik, P. C. (2016a). Soil fertility gradients shape the agrobiodiversity of Amazonian homegardens. Agric. Ecosyst. Environ. 221, 270–281. doi: 10.1016/j.agee.2016.01.002
Junqueira, A. B., Stomph, T. J., Clement, C. R., and Struik, P. C. (2016b). Variation in soil fertility influences cycle dynamics and crop diversity in shifting cultivation systems. Agric. Ecosyst. Environ. 215, 122–132. doi: 10.1016/j.agee.2015.09.015
Kainer, K. A., Wadt, L. H., and Staudhammer, C. L. (2014). Testing a silvicultural recommendation: Brazil nut responses 10 years after liana cutting. J. Appl. Ecol. 51, 655–663. doi: 10.1111/1365-2664.12231
Keddy, P. A. (1992). A pragmatic approach to functional ecology. Funct. Ecol. 6, 621–626. doi: 10.2307/2389954
Kennedy, J. (2012). Agricultural systems in the tropical forest: a critique framed by tree crops of Papua New Guinea. Quat. Int. 249, 140–150. doi: 10.1016/j.quaint.2011.06.020
Kleinman, P. J. A., Pimentel, D., and Bryant, R. B. (1995). The ecological sustainability of slash-and-burn agriculture. Agric. Ecosyst. Environ. 52, 235–249. doi: 10.1016/0167-8809(94)00531-I
LaManna, J. A., Mangan, S. A., Alonso, A., Bourg, N. A., Brockelman, W. Y., Bunyavejchewin, S., et al. (2017). Plant diversity increases with the strength of negative density dependence at the global scale. Science 356, 1389–1392. doi: 10.1126/science.aam5678
Levis, C., Clement, C. R., Ter Steege, H., Bongers, F., Junqueira, A. B., Pitman, N., et al. (2017a). Forest conservation: humans' handprints. Science 355, 466–467. doi: 10.1126/science.aal2175
Levis, C., Costa, F. R. C., Bongers, F., Peña-Claros, M., Clement, C. R., Junqueira, A. B., et al. (2017b). Persistent effects of pre-Columbian plant domestication on Amazonian forest composition. Science 355, 925–931. doi: 10.1126/science.aal0157
Levis, C., de Souza, P. F., Schietti, J., Emilio, T., da Veiga Pinto, J. L. P., Clement, C. R., et al. (2012). Historical human footprint on modern tree species composition in the Purus-Madeira interfluve, central Amazoni. PLoS ONE 7:e48559. doi: 10.1371/journal.pone.0048559
Lins, J., Lima, H. P., Baccaro, F. B., Kinupp, V. F., Shepard, G. H. Jr., and Clement, C. R. (2015). Pre-Columbian floristic legacies in modern homegardens of Central Amazonia. PLoS ONE 10:e0127067. doi: 10.1371/journal.pone.0127067
Lortie, C. J., Brooker, R. W., Choler, P., Kikvidze, Z., Michalet, R., Pugnaire, F. I., et al. (2004). Rethinking plant community theory. Oikos 107 433–438. doi: 10.1111/j.0030-1299.2004.13250.x
Mayle, F. E., and Iriarte, J. (2014). Integrated palaeoecology and archaeology–a powerful approach for understanding pre-Columbian Amazonia. J. Archaeol. Sci. 51, 54–64. doi: 10.1016/j.jas.2012.08.038
McKey, D., Elias, M., Pujol, B., and Duputié, A. (2010). The evolutionary ecology of clonally propagated domesticated plants. New Phytol. 186, 318–332. doi: 10.1111/j.1469-8137.2010.03210.x
McMichael, C. H., Palace, M. W., Bush, M. B., Braswell, B., Hagen, S., Neves, E. G., et al. (2014). Predicting pre-Columbian anthropogenic soils in Amazonia. Proc. R. Soc. B 281:20132475. doi: 10.1098/rspb.2013.2475
McMichael, C. H., Piperno, D. R., Bush, M. B., Silman, M. R., Zimmerman, A. R., Raczka, M. F., et al. (2012). Sparse pre-Columbian human habitation in western Amazonia. Science 336, 1429–1431. doi: 10.1126/science.1219982
McMichael, C. H., Piperno, D. R., Neves, E. G., Bush, M. B., Almeida, F. O., Mongeló, G., et al. (2015). Phytolith assemblages along a gradient of ancient human disturbance in western Amazonia. Front. Ecol. Evol. 3, 1–15. doi: 10.3389/fevo.2015.00141
Meyer, R. S., DuVal, A. E., and Jensen, H. R. (2012). Patterns and processes in crop domestication: an historical review and quantitative analysis of 203 global food crops. New Phytol. 196, 29–48. doi: 10.1111/j.1469-8137.2012.04253.x
Michon, G. (2005). Domesticating Forests: How Farmers Manage Forest Resources. Bogor: IRD/CIFOR/ICRAF.
Michon, G., de Foresta, H., Levang, P., and Verdeaux, F. (2007). Domestic forests: a new paradigm for integrating local communities' forestry into tropical forest science. Ecol. Soc. 12:1. doi: 10.5751/ES-02058-120201
Miller, R. P., and Nair, P. R. (2006). Indigenous agroforestry systems in Amazonia: from prehistory to today. Agrofor. Syst. 66, 151–164. doi: 10.1007/s10457-005-6074-1
Mistry, J., Bilbao, B. A., and Berardi, A. (2016). Community owned solutions for fire management in tropical ecosystems: case studies from indigenous communities of South America. Phil. Trans. R. Soc. B 371:20150174. doi: 10.1098/rstb.2015.0174
Moles, A. T., and Westoby, M. (2006). Seed size and plant strategy across the whole life cycle. Oikos 113, 91–105. doi: 10.1111/j.0030-1299.2006.14194.x
Montoya, E., Rull, V., Stansell, N. D., Abbott, M. B., Nogué, S., Bird, B. W., et al. (2011). Forest–savanna–morichal dynamics in relation to fire and human occupation in the southern Gran Sabana (SE Venezuela) during the last millennia. Quat. Res. 76, 335–344. doi: 10.1016/j.yqres.2011.06.014
Moreira, P. A., Lins, J., Dequigiovanni, G., Veasey, E. A., and Clement, C. R. (2015). The domestication of annatto (Bixa orellana) from Bixa urucurana in Amazonia. Econ. Bot. 69, 127–135. doi: 10.1007/s12231-015-9304-0
Morin-Rivat, J., Fayolle, A., Favier, C., Bremond, L., Gourlet-Fleury, S., Bayol, N., et al. (2017). Present-day central African forest is a legacy of the 19th century human history. Elife 6:e20343. doi: 10.7554/eLife.20343
Muller-Landau, H. C., Wright, S. J., Calderón, O., Condit, R., and Hubbell, S. P. (2008). Interspecific variation in primary seed dispersal in a tropical forest. J. Ecol. 96, 653–667. doi: 10.1111/j.1365-2745.2008.01399.x
Nathan, R., Schurr, F. M., Spiegel, O., Steinitz, O., Trakhtenbrot, A., and Tsoar, A. (2008). Mechanisms of long-distance seed dispersal. Trends Ecol. Evol. 23, 638–647. doi: 10.1016/j.tree.2008.08.003
Neves, E. G., Petersen, J. B., Bartone, R. N., and Silva, C. D. (2003). “Historical and socio-cultural origins of Amazonian Dark Earths,” in Amazonian Dark Earths: Origin, Properties, Management, eds J. Lehmann, D. C. Kern, B. Glaser, and W. I. Woods (Berlin: Springer Science and Business Media), 29–50.
Padoch, C., and De Jong, W. (1987). “Traditional agroforestry practices of native and ribereno farmers in the lowland Peruvian Amazon,” in Agroforestry: Realities, Possibilities and Potentials, ed H. L. Gholz (Dordrecht: Martinus Nijhoff Publishers), 179–194.
Paiva, P. M., Guedes, M. C., and Funi, C. (2011). Brazil nut conservation through shifting cultivation. For. Ecol. Manage. 261, 508–514. doi: 10.1016/j.foreco.2010.11.001
Pausas, J. G., and Keeley, J. E. (2009). A burning story: the role of fire in the history of life. Bioscience 59, 593–601. doi: 10.1525/bio.2009.59.7.10
Peh, K. S. H., Lewis, S. L., and Lloyd, J. (2011). Mechanisms of monodominance in diverse tropical tree-dominated systems. J. Ecol. 99, 891–898. doi: 10.1111/j.1365-2745.2011.01827.x
Peters, C. M. (2000). “Precolumbian silviculture and indigenous management of neotropical forests,” in Imperfect Balance: Landscape Transformations in the Precolumbian Americas, ed D. L. Lentz (New York, NY: Columbia University Press), 203–223.
Peters, C. M., Balick, M. J., Kahn, F., and Anderson, A. B. (1989). Oligarchic forests of economic plants in Amazonia: utilization and conservation of an important tropical resource. Conserv. Biol. 3, 341–349. doi: 10.1111/j.1523-1739.1989.tb00240.x
Piperno, D. R., McMichael, C., and Bush, M. B. (2015). Amazonia and the Anthropocene: what was the spatial extent and intensity of human landscape modification in the Amazon Basin at the end of prehistory? Holocene 25, 1588–1597. doi: 10.1177/0959683615588374
Pitman, N. C. A., Terborgh, J. W., Silman, M. R., Núñez, V. P., Neill, D. A., Cerón, C. E., et al. (2001). Dominance and distribution of tree species in upper Amazonian terra firme forests. Ecology 82, 2101–2117. doi: 10.1890/0012-9658(2001)082[2101:DADOTS]2.0.CO;2
Pitman, N. C., Cecilio, M. P., Pudicho, M. P., Graham, J. G., Núñez V, M. P., Valenzuela, M., et al. (2011). Indigenous perceptions of tree species abundance across an upper Amazonian landscape. J. Ethnobiol. 31, 233–243. doi: 10.2993/0278-0771-31.2.233
Pitman, N. C., Silman, M. R., and Terborgh, J. W. (2013). Oligarchies in Amazonian tree communities: a ten-year review. Ecography 36, 114–123. doi: 10.1111/j.1600-0587.2012.00083.x
Politis, G. G. (1996). Moving to produce: Nukak mobility and settlement patterns in Amazonia. World Archaeol. 27, 492–511. doi: 10.1080/00438243.1996.9980322
Politis, G. G. (2007). Nukak: Ethnoarchaeology of an Amazonian People. Walnut Creek, CA: Left Coast Press.
Poorter, L., Bongers, F., Sterck, F. J., and Wöll, H. (2003). Architecture of 53 rain forest tree species differing in adult stature and shade tolerance. Ecology 84, 602–608. doi: 10.1890/0012-9658(2003)084[0602:AORFTS]2.0.CO;2
Posey, D. A. (1985). Indigenous management of tropical forest ecosystems: the case of the Kayapo Indians of the Brazilian Amazon. Agrofor. Syst. 3, 139–158. doi: 10.1007/BF00122640
Posey, D. A. (1987). Ethnoentomological survey of Brazilian indians. Entomol. Gen. 12, 191–202. doi: 10.1127/entom.gen/12/1987/191
Posey, D. A. (1993). “The importance of semi-domesticated species in post-contact Amazonia: effects of the Kayapo Indians on the dispersal of flora and fauna,” in Tropical Forests, People and Food: Biocultural interactions and applications to development, eds C. M. Hladik, A. Hladik, O. F. Linares, H. Pagezy, A. Semple, and M. Hadley (Paris: UNESCO-Parthenon), 63–71.
Quintero-Vallejo, E., Klomberg, Y., Bongers, F., Poorter, L., Toledo, M., and Peña-Claros, M. (2015). Amazonian dark earth shapes the understory plant community in a Bolivian forest. Biotropica 47, 152–161. doi: 10.1111/btp.12193
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., et al. (2003). The evolution of plant functional variation: traits, spectra, and strategies. Int. J. Plant Sci. 164, S143–S164. doi: 10.1086/374368
Reis, M., Ladio, A., and Peroni, N. (2014). Landscapes with Araucaria in South America: evidence for a cultural dimension. Ecol. Soc. 19:43. doi: 10.5751/ES-06163-190243
Ribeiro, M. B. N., Jerozolimski, A., de Robert, P., Salles, N. V., Kayapó, B., Pimentel, T. P., et al. (2014). Anthropogenic landscape in southeastern Amazonia: contemporary impacts of low-intensity harvesting and dispersal of Brazil nuts by the Kayapó Indigenous people. PLoS ONE 9:e102187. doi: 10.1371/journal.pone.0102187
Ricklefs, R. E. (1987). Community diversity: relative roles of local and regional processes. Science 235, 167–171. doi: 10.1126/science.235.4785.167
Rival, L. (1998). “Domestication as a historical and symbolic process: wild gardens and cultivated forest in the Ecuatorian Amazon,” in Advances in Historical Ecology, ed W. Balée (New York, NY: Columbia University Press), 232–250.
Rival, L. (2007). “Domesticating the landscape, producing crops and reproducing society in Amazonia,” in Convergence and Emergence: Towards a New Holistic Anthropology? eds D. Parkin and S. Ulijaszek (Oxford: Berghahn Books), 72–90.
Rival, L., and McKey, D. (2008). Domestication and diversity in manioc (Manihot esculenta Crantz ssp. esculenta, Euphorbiaceae). Curr. Anthropol. 49, 1119–1128. doi: 10.1086/593119
Roberts, P., Hunt, C., Arroyo-Kalin, M., Evans, D., and Boivin, N. (2017). The deep human prehistory of global tropical forests and its relevance for modern conservation. Nature Plants 3:201793. doi: 10.1038/nplants.2017.93
Ross, N. J. (2011). Modern tree species composition reflects ancient Maya “forest gardens” in northwest Belize. Ecol. Appl. 21, 75–84. doi: 10.1890/09-0662.1
Schmidt, M. J., Py-Daniel, A. R., de Paula Moraes, C., Valle, R. B. M., Caromano, C. F., Texeira, W. G., et al. (2014). Dark earths and the human built landscape in Amazonia: a widespread pattern of anthrosol formation. J. Archaeol. Sci. 42, 152–165. doi: 10.1016/j.jas.2013.11.002
Schnitzer, S. A., Kuzee, M. E., and Bongers, F. (2005). Disentangling above-and below-ground competition between lianas and trees in a tropical forest. J. Ecol. 93, 1115–1125. doi: 10.1111/j.1365-2745.2005.01056.x
Schroth, G., Coutinho, P., Moraes, V. H., and amd Albernaz, A. L. (2003). Rubber agroforests at the Tapajós river, Brazilian Amazon—environmentally benign land use systems in an old forest frontier region. Agric. Ecosyst. Environ. 97, 151–165. doi: 10.1016/S0167-8809(03)00116-6
Shanley, P., Clement, C. R., Carvalho, J. E. U., de Homma, A. K. O., Menezes, A. J. E. A., and de (2016). “Amazonian fruits: How farmers nurture nutritional diversity on farm and in the forest,” in Tropical Fruit Tree Diversity: Good Practices for in situ and on-Farm Conservation, eds B. Sthapit, H. A. H. Lamers, V. R. Rao, and A. Bailey (Abingdon: Routledge), 147–160.
Shanley, P., Serra, M., and Medina, G. (2010). Frutíferas e Plantas úteis na Vida Amazônica. Bogor: CIFOR.
Shepard, J. R. G., and Ramirez, H. (2011). “Made in Brazil”: human dispersal of the Brazil nut (Bertholletia excelsa, Lecythidaceae) in ancient Amazonia. Econ. Bot. 65, 44–65. doi: 10.1007/s12231-011-9151-6
Smith, B. D. (2011). General patterns of niche construction and the management of ‘wild’ plant and animal resources by small-scale pre-industrial societies. Philos. Trans. R. Soc. B 366, 836–848. doi: 10.1098/rstb.2010.0253
Smith, M., and Fausto, C. (2016). Socialidade e diversidade de pequis (Caryocar brasiliense, Caryocaraceae) entre os Kuikuro do alto rio Xingu (Brasil). Bol. Mus. Para. Emílio Goeldi. Cienc. Hum. 11, 87–113. doi: 10.1590/1981.81222016000100006
Smith, N. J. H. (1980). Anthosols and human carrying capacity in Amazonia. Ann. Assoc. Am. Geogr. 70, 553–566. doi: 10.1111/j.1467-8306.1980.tb01332.x
Stahl, P. W. (2015). Interpreting interfluvial landscape transformations in the pre-Columbian Amazon. Holocene 25, 1598–1603. doi: 10.1177/0959683615588372
ter Steege, H., Pitman, N. C. A., Sabatier, D., Baraloto, C., Salomão, R. P., Guevara, J. E., et al. (2013). Hyperdominance in the Amazonian tree flora. Science 342:1243092. doi: 10.1126/science.1243092
ter Steege, H., Pitman, N. C., Phillips, O. L., Chave, J., Sabatier, D., Duque, A., et al. (2006). Continental-scale patterns of canopy tree composition and function across Amazonia. Nature 443, 444–447. doi: 10.1038/nature05134
Terrell, J. E., Hart, J. P., Barut, S., Cellinese, N., Curet, A., Denham, T., et al. (2003). Domesticated landscapes: the subsistence ecology of plant and animal domestication. JAMT 10, 323–368. doi: 10.1023/B:JARM.0000005510.54214.57
Thomas, E., Alcázar Caicedo, C., McMichael, C. H., Corvera, R., and Loo, J. (2015). Uncovering spatial patterns in the natural and human history of Brazil nut (Bertholletia excelsa) across the Amazon Basin. J. Biogeog. 42, 1367–1382. doi: 10.1111/jbi.12540
Thomas, E., van Zonneveld, M., Loo, J., Hodgkin, T., Galluzzi, G., and van Etten, J. (2012). Present spatial diversity patterns of Theobroma cacao L. in the neotropics reflect genetic differentiation in Pleistocene refugia followed by human-influenced dispersal. PLoS ONE 7:e47676. doi: 10.1371/journal.pone.0047676
Valencia, R., Foster, R. B., Villa, G., Condit, R., Svenning, J.-C., Hernández, C., et al. (2004). Tree species distributions and local habitat variation in the Amazon: large forest plot in eastern Ecuador. J. Ecol. 92, 214–229. doi: 10.1111/j.0022-0477.2004.00876.x
Van Gemerden, B. S., Olff, H., Parren, M. P., and Bongers, F. (2003). The pristine rain forest? Remnants of historical human impacts on current tree species composition and diversity. J Biogeogr. 30, 1381–1390. doi: 10.1046/j.1365-2699.2003.00937.x
Watling, J., Iriarte, J., Maylec, F. E., Schaand, D., Pessendae, L. C. R., Loaderf, N. J., et al. (2017a). Reply to Piperno et al.: it is too soon to argue for localized, short-term human impacts in interfluvial Amazonia. Proc. Natl. Acad. Sci. U.S.A. 114:201705697. doi: 10.1073/pnas.1705697114
Watling, J., Iriarte, J., Mayle, F. E., Schaan, D., Pessenda, L. C. R., Loader, N. J., et al. (2017b). Impact of pre-Columbian “geoglyph” builders on Amazonian forests. Proc. Natl. Acad. Sci. U.S.A. 114, 1868–1873. doi: 10.1073/pnas.1614359114
Wiersum, K. F. (1997a). Indigenous exploitation and management of tropical forest resources: an evolutionary continuum in forest-people interactions. Agric. Ecosyst. Environ. 63, 1–16. doi: 10.1016/S0167-8809(96)01124-3
Wiersum, K. F. (1997b). From natural forest to tree crops, co-domestication of forests and tree species, an overview. NJAS 45, 425–438.
Willis, K. J., Gillson, L., and Brncic, T. M. (2004). How“ virgin” is virgin rainforest? Science 304, 402–403. doi: 10.1126/science.1093991
Wirth, C., Messier, C., Bergeron, Y., Frank, D., and Fankhänel, A. (2009). “Old-growth forest definitions: a pragmatic view,” in Old-Growth Forests: Function, Fate and Value, eds C. Wirth, G. Gleixner, and M. Heimann (New York, NY; Berlin; Heidelberg: Springer), 11–33.
Woods, W. I., Denevan, W. M., and Rebellato, L. (2013). “How many years do you get for couterfeitting a paradise?” in Soils, Climate and Society Archaeological Investigations in Ancient America, eds S. E. Hayes and J. D. Wingard (Denver: University Press of Colorado), 1–20.
Woods, W. I., and McCann, J. M. (1999). “The Anthropogenic origin and persistence of amazonian dark earths,” in Yearbook (Conference of Latin Americanist Geographers) (Austin, TX: University of Texas Press), 7–14.
Zeder, M. A. (2006). Central questions in the domestication of plants and animals. Evol. Anthr. 15, 105–117. doi: 10.1002/evan.20101
Zent, E. L., and Zent, S. (2004). “Amazonian Indians as ecological disturbance agents: the Hotï of the Sierra de Maigualida, Venezuelan Guayana,” in Etnobotany and Conservation of Biocultural Diversity, eds T. S. Carlson and L. Maffi (Bronx, NY: NYBG Press), 79–112.
Zent, S., and Zent, E. (2012). Jodï horticultural belief, knowledge and practice: incipient or integral cultivation? Bol. Mus. Para. Emílio Goeldi. Cienc. Hum. 7, 293–338. doi: 10.1590/S1981-81222012000200003
Zobel, M. (1997). The relative of species pools in determining plant species richness: an alternative explanation of species coexistence? Trends Ecol. Evol. 12, 266–269. doi: 10.1016/S0169-5347(97)01096-3
Keywords: cultural forests, patch formation, dominance, Amazonian useful species, indigenous management, landscape domestication, Terra Preta de Índio
Citation: Levis C, Flores BM, Moreira PA, Luize BG, Alves RP, Franco-Moraes J, Lins J, Konings E, Peña-Claros M, Bongers F, Costa FRC and Clement CR (2018) How People Domesticated Amazonian Forests. Front. Ecol. Evol. 5:171. doi: 10.3389/fevo.2017.00171
Received: 31 July 2017; Accepted: 13 December 2017;
Published: 17 January 2018.
Edited by:
B. Mohan Kumar, Nalanda University, IndiaReviewed by:
Bharath Sundaram, Nalanda University, IndiaJean Kennedy, Australian National University, Australia
Louis S. Santiago, University of California, Riverside, United States
Copyright © 2018 Levis, Flores, Moreira, Luize, Alves, Franco-Moraes, Lins, Konings, Peña-Claros, Bongers, Costa and Clement. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carolina Levis, Y2Fyb2xsZXZpc0BnbWFpbC5jb20=