 Maurício S. Reis1*
Maurício S. Reis1* Tiago Montagna1
Tiago Montagna1 Andréa G. Mattos1
Andréa G. Mattos1 Samantha Filippon1
Samantha Filippon1 Ana H. Ladio2
Ana H. Ladio2 Anésio da Cunha Marques3
Anésio da Cunha Marques3 Alex A. Zechini1
Alex A. Zechini1 Nivaldo Peroni4
Nivaldo Peroni4 Adelar Mantovani5
Adelar Mantovani5- 1Núcleo de Pesquisas em Florestas Tropicais, Programa de Pós-Graduação em Recursos Genéticos Vegetais, Universidade Federal de Santa Catarina, Florianópolis, Brazil
- 2Grupo de Etnobiología, INIBIOMA, Universidad Nacional del Comahue, Bariloche, Argentina
- 3Floresta Nacional de Três Barras, Instituto Chico Mendes de Conservação da Biodiversidade, Três Barras, Brazil
- 4Laboratório de Ecologia Humana e Etnobotânica, Departamento de Ecologia e Zoologia, Universidade Federal de Santa Catarina, Florianópolis, Brazil
- 5Centro de Ciências Agroveterinárias, Universidade do Estado de Santa Catarina, Lages, Brazil
Araucaria forest is a sub-tropical component of the Atlantic Forest Biome, occurring between 18 and 30° S latitude, and 500–1,800 m altitude in Southern and Southwestern Brazil and Northwestern Argentina. In recent history (Twentieth century), this forest has undergone non-sustainable exploitation and reduction in area dedicated to agricultural and forestry use. However, smallholders continue using several resources from this forest, even adapting management systems. The main system is geared toward the production of yerba mate (Ilex paraguariensis) under araucaria trees (Araucaria angustifolia), which holds economic, cultural, and social relevance for thousands of farmers. Seeking evidence of domestication and conservation of the resources managed in this system, we compared different landscapes on the Northern Plateau of Santa Catarina in Southern Brazil. Focusing on three species-yerba mate, araucaria and caraguatá (Bromelia antiacantha)—we characterized management practices (interviews and guided tours), demographic structure (permanent plots in farming zones and in a protected area), and genetic diversity in populations from the same places. Demographic structure and genetic diversity from different landscapes were compared to evaluate the system potential for conservation. The results indicated that the three species are intentionally promoted with practices of protection, transplanting and/or selection, in different ways and with different functions (caraguatá hedges, yerba mate harvesting, and collection of pinhões—seed like nuts—of araucaria). Landscapes are managed for yerba mate harvesting and cattle raising, for both economic and cultural reasons, with a consequent reduction in the density of most other plant species. In all cases the genetic diversity of the species was high for most of the sampled properties, and the set of farmers' populations did not differ from the protected area. The set of populations of each species operates as a metapopulation and local management practices contribute to conservation. Thus, the farmers' management systems and practices maintain the landscape with productive forest fragments, favoring the conservation-by-use of these species. The system requires these management practices, which bring about changes in various species and are motivated by cultural and economic factors, in order to maintain the landscapes domesticated.
Introduction
Recent literature has presented several examples of a human signature imprinted on the landscape, reflecting cultural strength in the prehistoric/historical determination of composition and structure of forest formations (Denevan, 2001; Balée, 2006; Bitencourt and Krauspenhar, 2006; Balick, 2007; Behling and Pillar, 2007; Reis et al., 2014; Clement et al., 2015; Iriarte et al., 2017; Levis et al., 2017; Roberts et al., 2017). One example of such human action with the greatest accumulation of evidence is the Amazonian Dark Earth (Terra Preta de Índio). Several studies have revealed that Amazonian Dark Earth is a cultural mark in forest formation—eventually considered pristine (Clement and Junqueira, 2010)—perceived since the arrival of European botanists in the Americas (Clement and Junqueira, 2010; Clement et al., 2015; Levis et al., 2017).
These advances encourage a new perspective on the dynamics of supposedly natural ecosystems and the ecology of species, as well as possibilities for conservation and use. In particular, two aspects emerge from this new vision:
A. The marks of domestication processes on numerous species and landscapes, with models specific to different cultures in several environments.
B. The fact of landscape and species domestication in different environments can be present and currently active at the interface of agricultural and forest environments, with autochthonous species.
Thus, domestication systems involving different intensities of use/intervention initiated in the past remain in the present, associated with the distinct needs and perspectives of different peoples and cultures. Management practices entailing protection, promotion and selection are present in today's “extractivism” systems, and imply effective adjustments of landscapes and populations to the dimensions of current use (see Wiersum, 1996, 1997b; Martins, 2005; Emperaire and Peroni, 2007; Miranda and Hanazaki, 2008; Capparelli et al., 2011; Parra et al., 2012; Dawson et al., 2013; Steenbock and Reis, 2013; Roberts et al., 2017).
Plant domestication in forest environments begins with a process mediated by humans which favors the occurrence of the chosen species to the detriment of others (Wiersum, 1997a). Later, in the same environment, the implantation of selected species is common (Wiersum, 1997a). Therefore, from the conceptual reference of Clement (1999), domestication can be understood as a process acting on the landscape as a result of human intervention, and, influenced by cultural background. Thus, landscapes are produced where the degree of human interference can lead to different forest structures and different genotypic and phenotypic frequencies in plant populations of interest, some also being affected negatively. In the same landscape humans can target one or several resources, in different intensities and microenvironments, thus producing cultural landscapes (sensu Berkes et al., 2000).
These cultural landscapes (sensu Berkes et al., 2000; Berkes and Folke, 2002; Berkes and Turner, 2006; Ladio, 2011) are more common than previously imagined, and this perception brings the need for a change in posture regarding the possibility of conservation and use. The human presence and its cultural values can be determinant in the landscape perceived today, and favor both conservation and use at the same time. Furthermore, several recent studies (Heywood et al., 2007; Clement et al., 2010; Capparelli et al., 2011; Shepard and Ramirez, 2011; Steenbock et al., 2011; Parra et al., 2012; Dawson et al., 2013; Steenbock and Reis, 2013) demonstrate the importance of management systems and local practices associated with obtaining resources from forest environments, as discussed by Roberts et al. (2017). These studies have been carried out with a view to understanding different forms of landscape domestication, or in the sense of recognizing and valuing this strategy for obtaining income, and the diversification of family farming (Kubo et al., 2008; Vieira-da-Silva and Reis, 2009; Vieira-da-Silva and Miguel, 2014).
The main innovation of these kinds of studies is a more realistic understanding of how the composition of species and the structure of landscapes are determined. This understanding encompasses the active presence of humans and their interests, allowing more consistent actions for conservation and use of landscapes, including public policies (see Dawson et al., 2013; Roberts et al., 2017). Studies with this emphasis are recent and scarce in the Brazilian Atlantic Rainforest, despite its importance for the sustainable use of species domesticated by the native people, and for the establishment of public policies for conservation and use of biodiversity. In this context the Araucaria forest (Ombrophilous Mixed Forest), a forest formation of the Atlantic Rainforest, has been the object of several studies (see Reis et al., 2010; Zechini et al., 2012; Steenbock and Reis, 2013; Vieira-da-Silva and Miguel, 2014; Adan et al., 2016).
Before the arrival of Europeans in South America, the Araucaria forest occupied an estimated area of 200,000 km2 in Brazil and Argentina (Hueck, 1972). However, since the beginning of the Twentieth century this ecosystem has suffered more than 100 years of wood exploitation (Trajano, 1996; Nodari and de Carvalho, 2010), intensive agriculture and livestock production, and the effects of habitat fragmentation (Guerra et al., 2002; Ribeiro et al., 2009). This has left the Araucaria forest restricted to only 12% of its original area in Brazil (Ribeiro et al., 2009), and produced several legal restrictions to its use.
However, smallholders (such as family farmers) continue to use several resources from this forest, even adapting management systems. The main system is geared toward the production of yerba mate (Ilex paraguariensis) under araucaria trees (Araucaria angustifolia)—the native yerba mate system, NYMS—which holds economic, cultural, and social relevance for thousands of farmers (Marques, 2014). For instance, only in 2014, more than 8,700 tons of pinhões (seed-like nuts from A. angustifolia) were gathered in Brazil, representing a total product value of R$ 19,325,000.00 (IBGE, 2014; at that time over US$ 8.2 million).
According to the Brazilian Institute of Geography and Statistics (IBGE, 2014), in 2014 Brazil produced 333,017 tons of native yerba mate, and it was the main non-timber product in terms of production volume (IBGE, 2014). Production of native yerba mate is carried out in 5,150 establishments (predominately family farmers) in Santa Catarina State (EPAGRI/CEPA, 2016).
In this context we investigated landscapes with NYMS, characterizing the principal practices and motivation of family farmers, seeking evidence of landscape domestication and evaluating whether the system conserves the principal resources managed.
Our study has focused on three species in the NYMS: yerba mate (I. paraguariensis), araucaria (A. angustifolia), and caraguatá (Bromelia antiacantha). If on the one hand these species represent significant resources for the subsistence of family farmers, on the other hand they are interesting case studies to exemplify how these farmers play an active role in the conservation of plant populations. In addition, in order to evaluate whether the NYMS can conserve genetic diversity of the studied species, we analyzed genetic diversity in the yerba mate, araucaria and caraguatá populations from NYMS and from a protected area.
Materials and Methods
Study Area
The study was conducted in southern Brazil, in the Northern Plateau of Santa Catarina state. Plant populations and ethnobotanical studies were realized in farmers' properties situated in five different municipalities, and in a Protected Area - PA (Três Barras National Forest) (Figure 1). The climate in this region is described as subtropical humid (Cfa), according to Köeppen's classification, with mean annual temperature ranging from 15.5 to 17°C, and annual precipitation between 1,360 and 1,670 mm (EPAGRI, 2001). The soil in all sampled plots (further described) is classified as a sedimentary, nutrient-poor, deep and red latosol, and the relief is classified as smooth wavy, and the mean altitude is 750 m (Santa-Catarina, 1986).
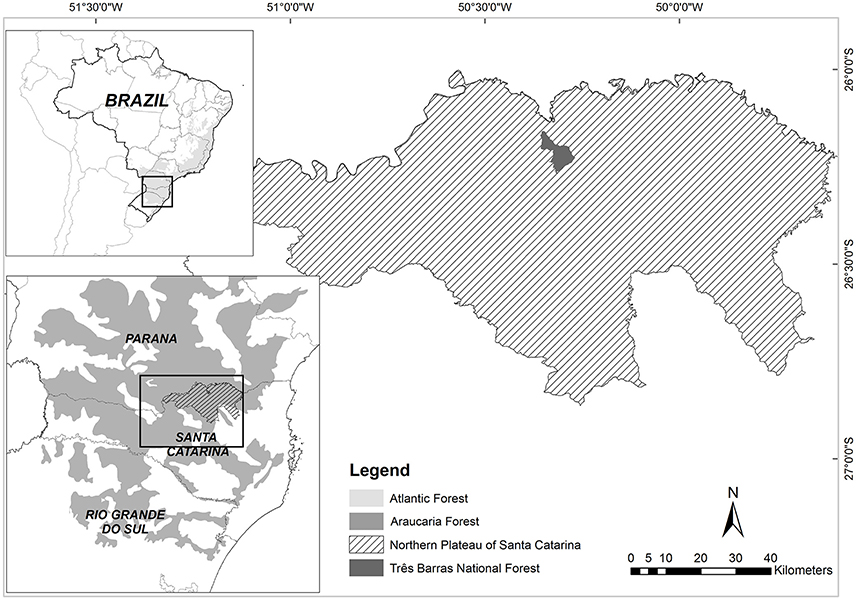
Figure 1. Study area – Northern Plateau of Santa Catarina State, Brazil, including localization of Três Barras National Forest (Protected Area).
The Northern Plateau of Santa Catarina covers 10,466.70 km2, is composed of 14 municipalities and has an average HDI of 0.79 (IBGE, 2006). The total population of the territory is 351,332 inhabitants, of which 23.76% live in the rural area (IBGE, 2014). The Northern Plateau produces 27.2% of Santa Catarina native yerba mate and 38.33% (1,974) of the state's productive rural properties which grow native yerba mate are situated here (EPAGRI/CEPA, 2016).
Study Species
Araucaria angustifolia, popularly known as Paraná pine, pinheiro-do-paraná, or pinheiro-brasileiro, is a conifer that has been intensively exploited since the beginning of the Twentieth century for wood production (Guerra et al., 2002; Nodari and de Carvalho, 2010), and is listed as critically endangered by the IUCN (IUCN, 2017). The species is dioecious and wind pollinated with a reproductive cycle of 2–3 years (Mantovani et al., 2004); it covered approximately 200,000 km2 in southern Brazil and northern Argentina (Reitz and Klein, 1966; Mattos, 1994; Guerra et al., 2002). Nowadays this species is economically and culturally important (Vieira-da-Silva and Reis, 2009; Assis et al., 2010; Zechini et al., 2012; Reis et al., 2014; Adan et al., 2016), especially for the production of seeds (pinhões), which are used as food and provide a source of income for many family farmers in southern Brazil (Guerra et al., 2002; Vieira-da-Silva and Reis, 2009; Vieira-da-Silva et al., 2011; Adan et al., 2016).
Ilex paraguariensis is a tree species native to South America and popularly known as yerba mate, erva-mate or mate (Edwin and Reitz, 1967). The leaves are used to make a traditional tea-like beverage (Chimarrão) in Brazil, Argentina, Uruguay, and Chile, and it is also a medicinal plant. It is a dioecious species with entomophilous pollination and zoocorical dispersal (mainly by birds) (Ferreira et al., 1983; Pires et al., 2014; Mattos, 2015). In Brazil it is the main non-timber product (in tons) from extractive activity (IBGE, 2014), with great economic and social importance in southern Brazil, contributing to the conservation of forest remnants through NYMS (ervais—landscapes with yerba mate) (Marques, 2014; Mattos, 2015). The NYMS, however, presents a great diversity of situations, due to the different management methods and meanings these systems can have for the farmers, resulting in different landscapes (Marques, 2014; Mattos, 2015).
Bromelia antiacantha (caraguatá) is a bromeliad native to the Atlantic Forest, and occurs in different forest formations, including Araucaria forest (Reitz, 1983). This plant has attracted the attention of researchers (Santos et al., 2004; Andrighetti-Fröhner et al., 2005; Brehmer, 2005; Duarte et al., 2007; Filippon et al., 2012a,b) due to its medicinal, ornamental and industrial potential. Studies show that the use of B. antiacantha forms part of the history of Northern Plateau communities (Duarte et al., 2007; Filippon et al., 2012a,b). The caraguatá is found in Araucaria forest fragments (Hanisch et al., 2006, 2010; Mattos, 2011; Mello, 2013), areas where cattle graze, areas where yerba mate is grown and in hedges (Filippon, 2009).
Field and Laboratory Methodology
This study integrates different field methodologies, including interviews with family farmers, as well as demographic and genetic studies of the above-mentioned species in a Protected Area and on farmers' properties. In all cases the plant populations sampled were the same as in the demographic and ethnobotanical studies. Below we detail each of the analyses:
Ethnobotanical Studies
Ethnobotanical studies were conducted through semi-structured interviews with family farmers that employ the NYMS on their properties (eight interviews focused on A. angustifolia; 93 on I. paraguariensis and 41 on B. antiacantha), which sought to identify and develop questions directly related to management of the species and local use. Before each interview we obtained a prior informed consent in accordance with the code of ethics of the International Society of Ethnobiology. The interviews addressed topics associated with the naming, use and management practices employed for the three species. Guided tours were also conducted (Albuquerque et al., 2008) with the farmers to describe the management systems and practices employed in each area. We intentionally selected the interviewees, considering only farmers who use and/or manage the target species. Interviewees were contacted through the snowball method described by Bailey (1994). The results are presented in a descriptive interpretative form, based on interviews and observations, and expressed as percentage of occurrence or average values with standard deviation, when necessary.
Demographic Studies
For characterization of the population structure of the three studied species, permanent plots (40 × 40 m) were marked out in forest fragments of farmers' properties, in distinct landscape units, and in the PA (in an area not managed in the last 50 years). All the trees and shrubs were marked and diameter at breast height (DBH) was measured. For the analysis we determined the density of I. paraguariensis, A. angustifolia and all species of the Myrtaceae family, considered the key structuring species (Assis et al., 2010; Mello and Peroni, 2015).
For the demographic characterization of I. paraguariensis and A. angustifolia, sampling was carried out in two different situations on the farmers' properties, according to the local management mentioned in the interviews, and in the Protected Area:
A. Landscapes without cattle: areas with I. paraguariensis gathering and few interventions (8 plots on 4 properties).
B. Landscapes with cattle: areas with I. paraguariensis gathering, frequent mowing, and cattle grazing (19 plots on 5 properties).
C. PA: area with no I. paraguariensis gathering and no other interventions (12 plots).
Additionally, for A. angustifolia, one sample of 60 female plants, 30 on farmers' properties and 30 in the PA, was evaluated annually (2010–2012) in terms of number of pinhas (reproductive structures) per plant, and productivity per pinha (only 2010).
For the demographic characterization of B. antiacantha, permanent plots of 20 × 40 m were used in forests and 20 linear meters to study the caraguatá hedges. All rosettes of the species were counted, classified as vegetative or reproductive (considering the presence of inflorescences or infructescences) and leaf length was measured. Bromelia antiacantha populations were studied in the following areas:
A. Managed: livestock management (cattle, sheep, pigs and horses) (3 plots in farmer 2), livestock management (cattle, sheep, pigs, and horses), harvesting of yerba and mowing (3 plots in Farmer 1 and 2 plots in another area in farmer 2),
B. Unmanaged: areas that have been characterized for not having undergone any of the activities previously mentioned for at least 10 years previously in the case of farmer properties and 50 years in the PA (3 plots in farmer 3 and 12 plots in PA).
C. Hedges: characterized as “strips”/linear bands of B. anticantha rosettes planted, established and administered by local farmers (26 plots in farmers 1 and 2). Considered the areas with most human interference.
The results are expressed as average frequencies estimated per hectare, with respective standard deviations.
Genetic Characterization
The three studied species were genetically characterized using allozyme markers, in starch gel (13%), following the recommendations of Kephart (1990) and Alfenas (1998). The extraction protocol was the same for all species, macerating fresh leaves with an automatic homogenizer and using extraction solution n° 1 (Alfenas, 1998). The enzymatic systems scored for each species, as well as the buffers are described in Table 1. Three buffers were utilized in the electrophoresis process: Tris Citrate pH 7.5 (Tris 27 g.L−1 and citric acid 16.52 g.L−1), Morpholine Citrate pH 6.1 (7.68 g.L−1 citric acid) and Histidine pH 8.0 (105.82 g.L−1 sodium citrate tribasic).
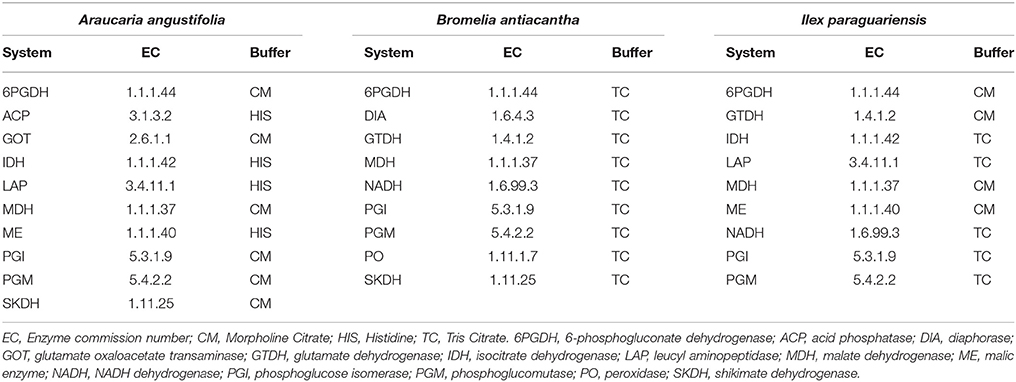
Table 1. Enzymatic systems used for genetic characterization of Araucaria angustifolia, Bromelia antiacantha, and Ilex paraguariensis populations.
Leaf tissues were collected from 50 individuals in each population. For A. angustifolia and I. paraguariensis sampling respected a minimum distance of 50 m between individuals, in order to minimize family structure. For B. antiacantha, adults/mature rosettes (leaves more than 2 m in length) were sampled, respecting at least 15 m distance between them and avoiding the collection of ramets from the same plant.
Genetic Data Analysis
Based on the genotypes obtained in gel, the following genetic descriptors were estimated: allelic frequencies (Supplementary Table 1), number of alleles per population/group (), number of unique alleles per population/group (Âun), mean allelic richness per loci based on the lowest sample size (Ân), observed (ĤO), and expected heterozygosity (ĤE) and fixation index (). Statistical significance (p < 0.05) of was tested, permuting alleles between individuals within populations/groups. All these analyses were performed using the FSTAT program, version 2.9.3.2 (Goudet, 2002).
In order to evaluate whether farmers conserve species genetic diversity, all these descriptors were estimated, in each species, for groups of populations inside the PA and on farmer's properties. In the case of B. antiacantha and I. paraguariensis, in order to evaluate possible differences between landscape units as described in the Demographic studies item, all genetic descriptors were also estimated for groups of populations in each landscape unit.
Possible genetic differences between farmers' properties and PA, and between landscape units were tested through confidence intervals (95%) for Ân, jackknifing the values across loci in R language (R Development Core Team, 2015) and for ĤE and ĤO through 1,000 bootstraps of individuals within population/group, also in R language, but using the “PopGenKit” package (Paquette, 2012).
Results and Discussion
Structural Characteristics of Landscapes with NYMS
The forest structure of the landscapes with NYMS in the region is presented in Table 2, which also includes information on the total number of species, density of the araucaria and the total number of individuals of Myrtaceae and yerba mate, considered as structuring species of these landscapes (Assis et al., 2010; Mello and Peroni, 2015).
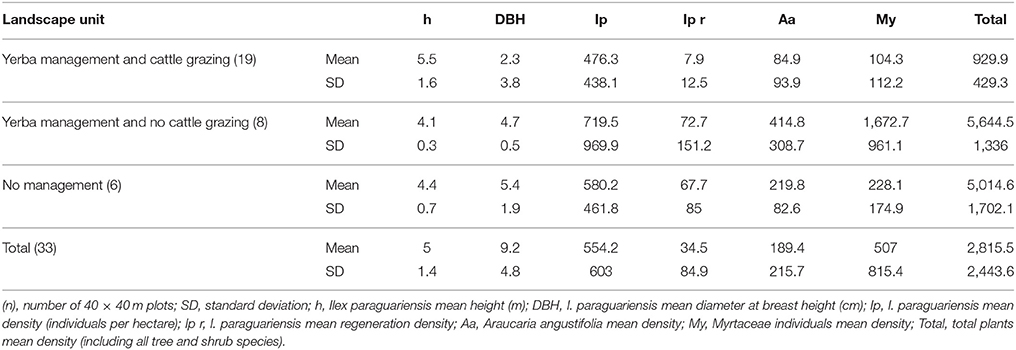
Table 2. Structural characteristics of landscapes with Ilex paraguariensis.
Demographic results presented great variation within landscape units (high standard deviations), showing peculiarities between and within the managed areas of each group (Table 2). Yerba mate represented 20.9% of the individuals (588.6 plants/ha); the Myrtaceae group represented 18% (507.0 plants/ha) and araucaria 6.7% (189.4 plants/ha). These species, considered structuring species of the Araucaria forests, represented up to 45.6% of the existing plants (1,285.0 plants/ha). This result reinforces the perspective that these species are key cultural species in the context of landscapes with management of yerba mate, as mentioned in Assis et al. (2010). The result also demonstrates the high number of other species in the system (see also Mello and Peroni, 2015).
The three structuring species (Myrtaceae, araucaria and yerba mate) have a strong historical/prehistoric relationship with the family farmers, caboclos (mestizos), and the indigenous peoples, in terms of food production and other uses in forest environments (Brandt and Campos, 2008; Gerhardt, 2009; Reis et al., 2014). This reinforces the recognition of human contributions to changes in vegetation patterns. Human activity influences species dynamics, which can lead to a new equilibrium and directional changes, or result in vegetation with different characteristics (Wiersum, 1997a; Clement, 1999; Trombulak et al., 2004; Casas et al., 2007; Steenbock et al., 2011).
Local Perceptions of Landscapes with NYMS
Historically on the Northern Plateau (SC) there is a link between native landscapes with the presence of yerba mate and harvesting practices carried out by farmers (Valentini, 2003; Barreto, 2009; Gerhardt, 2009; Grzebieluka and Sahr, 2009; Schuster and Sahr, 2009). This link should be considered in order to understand how farmers interpret their landscapes and make decisions with regard to their subsistence. These management practices are multi-dimensional and involve actions that are fundamental for maintenance of useful resources. This know-how constitutes part of the people's traditional ecological knowledge (TEK), which has been defined as a cumulative body of knowledge, beliefs and practices developed through adaptive processes, and involves cultural transmission over generations (Berkes et al., 2000).
For all the interviewed farmers the presence of A. angustifolia defines the landscape and holds connotations of regional identity associated with their ancestors. However, these farmers collect pinhão only for their own consumption, not for commercialization, since they consider yerba mate and cattle as the income-generating elements of the system. This differs from other regions, where gathering is carried out primarily for sale (Vieira-da-Silva and Reis, 2009; Vieira-da-Silva et al., 2011; Zechini et al., 2012; Adan et al., 2016).
Most of the informants (57%) referred to the forest environment with yerba mate harvesting using local terms, reflecting the close connection between farmers and their forest area. This form of naming consists of identification and classification of distinctive components of the managed environments. It contains empirical observations and information about the behavior and abundance of flora and domestic fauna. Farmers relied on such knowledge to ensure a supply of food, medicines and other subsistence resources. They mentioned four names: caíva (51.9%), potreiro (16.7%), invernada (16.7%), and mato nativo (14.7%). Caívas are ecosystems containing remnants of native Araucaria forests, with different levels of forest structure and grazing in the herbaceous stratum; pastures may be natural or naturalized (Marques et al., 2008; Reis et al., 2013). This landscape has been created due to the permanence, for decades, of a traditional productive system combining grazing of the herbaceous stratum with yerba mate and firewood extraction in the understory (Marques et al., 2008; Reis et al., 2013; Marques, 2014). Thus, the presence of cattle in many areas is a determining factor of this system, due to the value added by the sale of livestock. Mato nativo, potreiro and invernada are areas with some degree of Araucaria forest cover and yerba mate management; in the first there is no livestock production, in contrast to the other two.
Most of the NYMS areas (ervais) are located in places (soil, topography) of excellent to good agricultural aptitude, and consequently, with high pressure for conversion into crops. Yerba mate is cited as one of the main reasons for non-conversion in 80% of cases, and the multiple-use forest (timber, firewood, and yerba mate) with the livestock, in 55% of cases.
On the Northern Plateau (SC), yerba mate is managed in several situations; for instance, Marques (2014) identified 13 typologies (including situations with planted yerba mate). However, the main landscape is considered to be forest, occupying about 80% of the area, with yerba mate management, among which the caívas stand out.
All farmers considered yerba mate important mainly because it is a low-risk activity, with little investment and labor demand, associated with landscapes which have different uses. According to all interviewed farmers, the NYMS constitutes a stabilizing element on their property, capable of generating resources for forest areas and linked to strong cultural aspects, as well as being a pleasurable activity for the great majority of farmers, according to those who were interviewed. These aspects reinforce the role of these family farmers as maintainers of Araucaria forest fragments nowadays.
An additional relevant aspect is the preference shown by Brazilian consumers for a product (ground tea-like leaves for chimarrão) with a milder flavor (de Oliveira Suertegaray, 2002) from herbs grown in shade (native yerba mate), implying higher prices paid by the industry for the raw material originating from this system (EPAGRI/CEPA, 2017).
In these forest systems, rather than generating significant income in relation to total gross income, yerba mate constitutes an important reserve for 72% of the families, with the characteristics of a savings account, to be used in investments, emergencies and payment of debts.
Local Management Practices in NYMS
Yerba mate occurs spontaneously in the forest understory, but farmers (98.9%) promote it through removal of certain tree species and periodic mowing of shrub and arborescent vegetation. According to farmers, the presence of animal grazing also contributes to the control of this vegetation. In addition, yerba mate is planted (59.1%) in many situations using seedlings removed from other forest sites (5.4%), produced on the property (23.2%) or even bought from commercial nurseries (71.4%). However, the observed yerba mate natural regeneration is low (Table 2), since the farmers tend to prune all the plants without worrying about leaving seed trees.
For araucaria, besides the collection of pinhão no other practices were recorded for the species in the interviews. However, farmers recognize at least four varieties of female pine nut production plants (population variation as detected by Adan et al., 2016 in another region): São José, Comum, Cayová, and Macaco. The differences are associated with the maturation times of pinhão, an aspect already highlighted in other works (Vieira-da-Silva and Reis, 2009; Zechini et al., 2012; Adan et al., 2016) and they are even classified as botanical varieties (Reitz and Klein, 1966; Mattos, 1994) (see Adan et al., 2016 for further discussion).
The pinhões average yield, evaluated by a sample of 30 trees from farms, was 11.1 pinhas/tree between 2010 and 2012, with 3.6, 8.5, and 21.1 pinhas/tree for the years 2010, 2011, and 2012 respectively. Average productivity over the years was 71.1 kg/ha, with 23.0 kg/ha in 2010, 54.9 kg/ha in 2011 and 135.3 kg/ha in 2012. In the PA sample observations were made in the years 2011 and 2012, and the average production for these was 17.5 pinhas/tree, with averages of 11.8 in 2011 and 23.2 pinhas/tree in 2012. The average productivity of 2011 and 2012 was 333.0 kg/ha, varying from 224.3 kg/ha in 2011 to 441.7 kg/ha in 2012.
Pinhões productivity per hectare estimated for all populations was higher than that found by Vieira-da-Silva and Reis (2009) in 2006 (44.3 kg/ha) and by Mantovani et al. (2004) in 2001, and similar to that found by these authors in 2002 (117 and 160 kg/ha, respectively). In general, differences in productivity are attributed to the number of female individuals per hectare and the degree of forest evolution (Vieira-da-Silva and Reis, 2009; Figueiredo-Filho et al., 2011). The number of female plants producing pinhas varied between years; considering the entire period evaluated, the average number of female plants in production was 13.9 on farms, while in PA the this average was 29.6.
In economic terms, forests located in certain areas would yield (at the average price paid by regional markets of R$ 2.00/kg of pinhão) on average, R$ 142.00 per hectare, from commercialization of the pinhão. Although pinhão is less economically attractive than yerba mate in the region of Três Barras (Zechini et al., 2012), this resource represents an important source of income for many rural families in the state of Santa Catarina (Vieira-da-Silva and Reis, 2009; Adan et al., 2016).
An important aspect is the great variation in pinhão productivity in different years of production, as already mentioned by Mattos (1994), Mantovani et al. (2004), and Vieira-da-Silva and Reis (2009), and even between relatively close sites (Zechini et al., 2012). This variation reflects the unpredictability of production that is also used by farmers as an argument for the lower value of pinhão as a trade product in the region.
Considering the management practices carried out by farmers, the population structure of yerba mate and structure of the forest fragments (considering the set of structuring species—araucaria, yerba mate and several Myrtaceae), two main landscapes were detected:
A. Landscapes without cattle: areas used with few anthropic interventions (no livestock presence and no mowing), but with yerba mate management practices. These practices can be more drastic, such as pruning with shears every 3 years or less drastic, such as “hand” pruning every 4 years.
B. Landscapes with cattle: areas used with more intense interventions, such as the presence of cattle and mowing. Includes the same management characteristics for herbaceous plants as the previous group.
In both cases there are practices for protection of yerba mate plants, including possible transplanting and enrichment with plants from the same forest fragment or seedlings produced by farmers.
The Caraguatá Uses and Management in the NYSM
Caraguatá has been used for a long time in the region, for various purposes (Filippon, 2014). The fruits of this plant are used for medicinal treatment of, for example, pulmonary diseases (bronchitis/asthma) and influenza (Filippon, 2009; Filippon et al., 2012a). Uses of the species for making liquor and jelly were also cited in Filippon (2014) but the most frequently cited use (60%) was for hedges.
Caraguatá hedges were very common in the region from 1900 to 1960. According to farmers, this kind of fencing was used especially for pig farming; animals can't pass through the hedge due to the high density of rosettes and the thorns in their leaves. Caraguatá hedges are also used in other kinds of animal breeding like sheep, horses and cattle. Nowadays this type of pig breeding is no longer common; however, the hedges still remain on the properties. Some farmers still plant new caraguatá hedges, following tradition, and taking advantage of the efficiency of this type of fencing to hold animals in and establish property boundaries.
The density of B. antiacantha rosettes varies between studied areas. The unmanaged area presented the greatest density of rosettes and reproductive rosettes per hectare when compared to the managed areas (Table 3). Hedges possess, on average, the highest population density of all the sampled areas (316 times greater than that of the population in the unmanaged area), and also the highest number of the reproductive rosettes (Table 3).
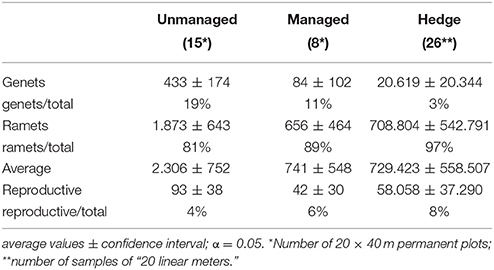
Table 3. Rosette density (per hectare) of Bromelia antiacantha in unmanaged, managed, and hedge populations.
Ramet proportions were high in all three situations; however, among these, the hedges stood out with up to 97% ramets (Table 3). In comparison to managed areas and hedges, the unmanaged area showed the highest average densities of genets per hectare. There was also variation in the proportion of reproductive rosettes. This variation is reflected in all landscape units including the hedges; that is, when the number of reproductive individuals was low in managed and unmanaged areas, it was also low in hedges (Table 3).
The caraguatá hedges were, and still are, made with rosettes harvested from the Araucaria forest fragments, where I. paraguariensis is extracted and where cattle graze. Sometimes the ramets used in hedges are donations from neighbors who want to “clean” the area to improve pasture for cattle, or want better conditions for working with yerba mate, since the caraguatá thorns make the farmers' work more difficult.
On these occasions the number of rosettes removed is generally high, and the farmers who receive them have an abundance of ramets to plant, so they can generally choose the strongest, lushest and youngest rosettes (0.8–1 m leaf length). This intention of making a hedge and the selection of rosettes allows us to highlight a domestication process. These management practices (choosing rosettes, building, and hedge maintenance) employed by local farmers generate a change in landscape, tending toward greater productivity and convenience.
In these areas caraguatá is not the main focus, but a means of adaptation of the landscape to facilitate the development of activities that generate income for the property, such as the harvesting of yerba mate and cattle grazing. Thus, from this perspective, management (mowing) of caraguatá can be seen as a consequence of this domesticated landscape for the production of yerba mate and/or cattle grazing.
Genetic Diversity and Conservation on Farms
Genetic descriptors for groups of populations in PA or on farmers' properties (the same for the three species) are presented in Table 4. In general, the data indicate that genetic diversity of the three species is being better maintained on the farmers' properties, compared to the PA. This result can be interpreted as arising from the common effect of past overexploitation; however, it is also an indication that the medium-term possibilities of maintaining diversity are similar in both situations, for the three species, reinforcing the possibility of conservation by use.
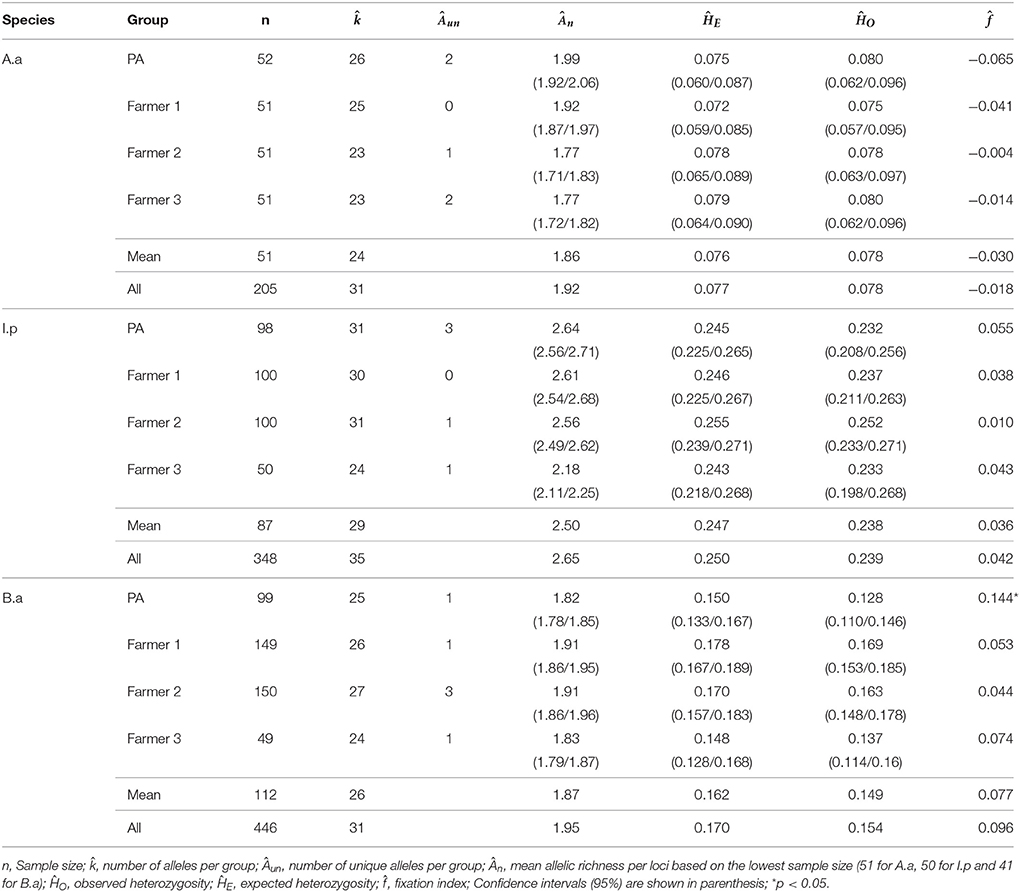
Table 4. Genetic descriptors estimated for Araucaria angustifolia (A.a), Ilex paraguariensis (I.p), and Bromelia antiacantha (B.a) populations grouped per occurrence in Protected Area (PA) or on Farmers' Properties.
The mean expected heterozygosity (genetic diversity - ĤE) estimated for A. angustifolia varied between 0.079 and 0.072 and did not differ in the four situations evaluated (Table 4). Thirty-one distinct alleles were detected, ranging from 23 to 26 in the different situations, with unique alleles present in both PA (2) and farmers' properties (3). It should be noted that the number of alleles found in PA (26) was higher, on average, than that found on each of the properties; however, the number found for the set of properties (28) was higher, including 3 unique alleles. In all situations the fixation index () was not different from zero (Table 4).
The estimated values were lower than the mean values obtained in a large study in the SC (Reis et al., 2012), which evaluated 31 A. angustifolia populations (13 allozymic loci). The values found were lower than the SC average for ĤE (0.124) and total number of alleles (51), but similar in terms of regional (Northern Plateau) ĤE mean (0.104) and total number of alleles detected (30). Thus, in general, for A. angustifolia, there were no differences between the diversity maintained in PA and on farmers' properties. In all the studied situations for A. angustifolia on agricultural properties, the practices associated with the species are similar, involving maintenance of the adult individuals and cone collection.
For yerba mate populations, the mean ĤE ranged from 0.255 to 0.243, with no difference between the situations evaluated (Table 4). For the set of populations 35 different alleles were detected, ranging from 24 to 31 in the different situations, with the presence of unique alleles in both PA (3) and farmers' properties (2). The set of populations on farm properties presented 32 alleles while 31 alleles were detected in PA. Estimated was not different from zero in all situations (Table 4).
Caraguatá populations presented mean ĤE ranging from 0.148 to 0.178, not differing between the studied situations (Table 4). Thirty-one different alleles were detected for the set of populations, varying from 24 to 27 in the different situations, and unique alleles were detected in both PA (1) and farmers' properties (5). Protected Area unit presented 25 alleles while the set of farm populations harbored 29 alleles. The fixation index was significant (0.144) only in PA, not differing from zero on farms (Table 4).
Genetic Diversity in Different Landscapes
For yerba mate, management practices and systems determine two distinct landscapes. Thus, Table 5 presents the diversity indexes considering population groups sampled in both landscapes, as well as the indexes referring to two populations sampled in the Três Barras National Forest (PA), representing unmanaged populations. The mean values for ĤE do not differ between situations; however, allelic richness (Â96) was higher in the unmanaged area. On the other hand, was significant only in the unmanaged area (Table 5). Unique alleles were found in two situations, 4 in non-managed populations and 3 in populations with yerba mate management and cattle grazing.
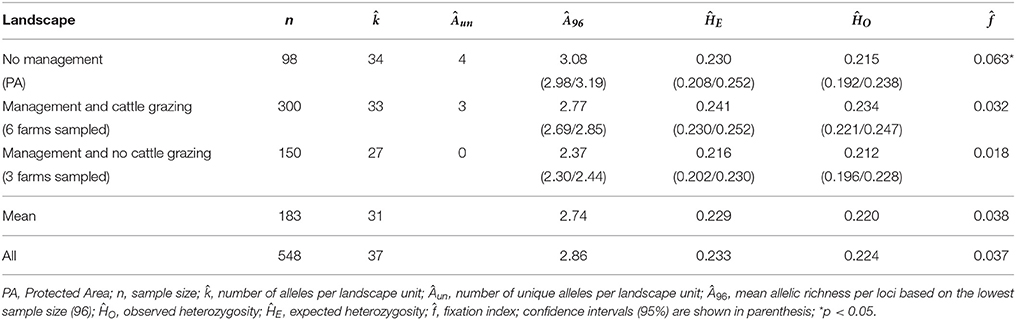
Table 5. Genetic descriptors estimated for Ilex paraguariensis in different landscape units.
Caraguatá populations presented some differences in heterozygosities (ĤO and ĤE) between landscapes. Mean values of ĤO and ĤE for hedges were significantly higher than those estimated for unmanaged landscape (Table 6). Hedges also presented higher numbers of alleles ( = 28), unique alleles (Âun = 3) and allelic richness (Ân = 2.15) than all other landscape units. Fixation indexes were significantly different from zero in the unmanaged landscape. In this case, the reduced number of rosettes when compared to the hedges may contribute to more crossing between relatives.
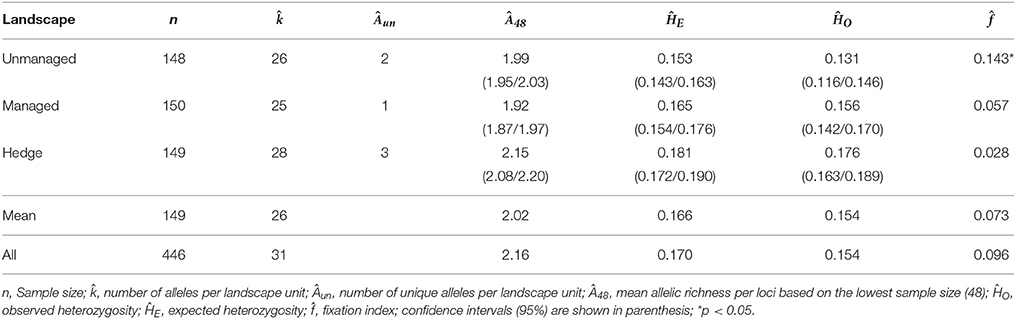
Table 6. Genetic descriptors estimated for Bromelia antiacantha in different landscape units.
The higher ĤO, ĤE, , Âun and Â48 values and the non-significant estimated for hedges, compared to unmanaged populations, could be related to the origin of the plants (from the same or different locations) used for hedge construction and to the selection of plants for hedges. As mentioned earlier, on many occasions neighbors donated the seedlings for hedges, mainly when the land was cleared, so any single hedge could have been made with seedlings from different populations, possibly increasing their number of alleles and heterozygotes.
In this context, observing the genetic diversity indexes obtained for B. antiacantha, it can be inferred that the hedges presented greater genetic diversity than populations that had not been managed for over 50 years, in accordance with results from other studies (Otero-Arnaiz et al., 2005; Zizumbo-Villarreal et al., 2013). This aspect is possibly due to the way the hedges are constructed, with rosettes from sites of natural occurrence of the species (forests).
Demographic studies showed that either by greater exposure to light in the hedge or other factors such as soil and temperature, the hedge had more reproductive rosettes (Table 3). The rosettes may come from various populations/farms with different genetic characteristics, which may contribute to greater diversity in hedges. In this sense, caraguatá fences can be seen as a metapopulation of the species, where each population of a landscape unit is a subpopulation.
Finally, it is possible to argue that the decrease in genetic diversity, as one of the indicators of domestication generally suggested in the literature, must be taken into account in relation to management practices. Thus, the fact that intensively managed populations present greater genetic diversity than unmanaged ones does not imply that they are considered less domesticated than the others, in the sense of human intervention in the population and in the landscape.
In addition, the farmers' way of life related to the gathering of rosettes in different populations is a result of management of the ervais (NYMS). In this sense there is an opportunity to “collect seedlings” from an activity directed by other managements which generate income for the property, like cattle and yerba mate. This fact differentiates the caraguatá from other plants cultivated and used in hedges: the use of these seedlings is desired, and in turn they are collected from different populations. Thus, the fact that the yerba mate areas are managed together with the existence of farmers interested in making caraguatá hedges increases species diversity. Therefore, this domestication process has a tendency to increase genetic diversity.
Both ethnobotanical and genetic studies show that caraguatá hedges can be seen as a form of on-farm conservation of genetic diversity. Traditionally, the focus of conservation has been the creation of reserves, and much has been debated about the size, form and number of reserves that must be created (Wiens, 1997). However, more recently this trend has been directed toward seeking alternatives that allow not only conservation, but also use, maintenance, and even an increase in population diversity (Clement et al., 2007). In this sense, on-farm conservation has been seen as a great opportunity for the conservation of species of human interest (Dawson et al., 2013).
Conclusion: Conservation-by-Use in Araucaria Forests
The management systems and practices conducted by farmers maintain the landscape with productive forest fragments, favoring the conservation of these species. The different landscape units, which include the three studied species at the same time, occur predominantly in fragments of less than 50 hectares (mean = 45.8 ± 33.4 ha), and represent, on average, 45.8% of the total property area. Several landscapes and management systems generally occur on the same property, forming a mosaic of situations (Marques, 2014; Mattos, 2015). These distinct fragments of a wider landscape form a unique situation that allows a high level of gene flow between the various units, favoring maintenance of the genetic diversity of each species and of the whole, as a metapopulation. Each landscape unit studied has structural peculiarities that influence ecological and genetic patterns. The units also have an interconnection with allele movement (pollen, seeds and individuals) mediated by fauna and man, allowing the maintenance of diversity in the metapopulation. However, such metapopulation is created and maintained for cultural and economic motives, in addition to ecological ones.
Thus it is possible to state that caívas are fundamental pieces in conservation of the species and landscapes with araucaria, mainly due to different management methods that result in mosaics of vegetation with different demographic, and consequently genetic, structures. This factor (management of areas) reinforces the idea of conservation-by-use of fragments, where in addition to conserving the species, the conditions that allow the development of new germplasm are also maintained. On-farm conservation is based not only on the conservation of existing germplasm; but the genetic variability maintained in this type of conservation (backyards, gardens and agroforestry systems) also allows the maintenance of biodiversity through evolutionary processes (FAO, 1996; Clement et al., 2007).
The system requires these management practices in the landscape, with changes in various species motivated by cultural and economic factors, in order to maintain domestication of these landscapes. However, contrary to expectation, domesticated landscapes for the production of yerba mate did not lead to an important reduction in genetic variability, since there is no conscious selection action in the species studied.
It is important to note that maintenance of Araucaria forest remnants in the region of native yerba mate exploitation is directly associated with the possibility of exploitation of this resource, which is related to the conservation-by-use of these fragments (Petersen et al., 2001; Mazuchowski, 2004; Mattos, 2011, 2015; Marques, 2014). However, there is also a potential risk that could jeopardize the system due to economic and socio-cultural reasons. The prices paid for yerba mate show significant fluctuations between years (EPAGRI/CEPA, 2017), which may be a reason to substitute the system for another agricultural or forestry activity. In addition, the new generations do not show interest in continuing farming activities, implying risks to the maintenance of regional cultural values (Marques, 2014).
Thus, the perspective of conservation-by-use of the multispecies system described here depends heavily on valorization of cultural aspects of the region, as well as on the economic valuation of yerba mate from this artisan system. Mechanisms of certification of origin could be an alternative of great regional importance, increasing the possibility of in situ, on-farm conservation of several autochthonous species from these Araucaria forests.
Author Contributions
MdR: Conceived the research, designed the research, was responsible for resource acquisition, directed AGM, SF thesis, AZ dissertation, codirected AdCM thesis and wrote the manuscript. TM: Conducted field and laboratory work, analyzed genetic data and wrote the manuscript. AGM: Conducted field work (including interviews), analyzed part of data and wrote the manuscript. SF: Conducted field work (including interviews), analyzed part of data and wrote the manuscript. AL: Conceived the research, provided comments on the manuscript. AdCM: Conducted fieldwork (interviews), analyzed part of data and provided comments on the manuscript. AZ: Conducted field and laboratory work and analyzed part of data. NP: Conceived the research, co-directed the AGM and SF thesis, and provided comments on the manuscript. AM: Conceived the research, co-directed the AZ master dissertation, was responsible for resource acquisition and provided comments on the manuscript. This research is the product of AZ master's thesis and AGM, AdCM, and SF Ph.D. thesis.
Funding
This study was supported by the Fundação de Amparo à Pesquisa e Inovação do Estado de Santa Catarina (FAPESC – process no. 4448/2010-2 and 11939/2009) and Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA – Macroprojetos 2/2009). Conselho Nacional de Desenvolvimento Científico e Tecnológico provided a Productivity Scholarship for MdR (CNPq – 309128/2014-5) and NP, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) provided doctoral scholarships for TM, AGM, and SF and a master's scholarship for AZ. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) supported AL.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are profoundly grateful to all farmers for their kindness in sharing their knowledge with us, and for their hospitality. We would also like to thank Núcleo de Pesquisas em Florestas Tropicais (NPFT), Laboratório de Fisiologia do Desenvolvimento e Genética Vegetal (LFDGV), and Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio) for logistical support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2018.00011/full#supplementary-material
References
Adan, N., Atchison, J., Reis, M. S., and Peroni, N. (2016). Local knowledge, use and management of ethnovarieties of Araucaria angustifolia (Bert.) Ktze. in the Plateau of Santa Catarina, Brazil. Econ. Bot. 70, 353–364. doi: 10.1007/s12231-016-9361-z
Albuquerque, U. P., Lucena, R. F. P., and Neto, E. M. F. L. (2008). “Seleção e escolha dos participantes da pesquisa,” in Métodos e Técnicas na Pesquisa Etnobotânica, eds U. P. Albuquerque, R. F. P. Lucena, and L. V. F. C. Cunha (Recife: Comunigraf), 21–40.
Alfenas, A. C. (1998). Eletroforese de Isoenzimas e Proteínas Afins: Fundamentos e Aplicações em Plantas e Microorganismos. Viçosa: Editora Universidade Federal de Viçosa.
Andrighetti-Fröhner, C. R., Sincero, T. C. M., da Silva, A. C., Savi, L. A., Gaido, C. M., Bettega, J. M. R., et al. (2005). Antiviral evaluation of plants from Brazilian Atlantic Tropical Forest. Fitoterapia 76, 374–378. doi: 10.1016/j.fitote.2005.03.010
Assis, A. L., Mattos, A., Peroni, N., Hanazaki, N., Reis, M. S., Mattos, A., et al. (2010). “Espécie-chave cultural: indicadores e aplicabilidade em etnoecologia,” in Etnoecologia em Perspectiva: Natureza, Cultura e Conservação, eds A. G. C. Alves, F. J. B. Souto, and N. Peroni (Recife: NUPEEA), 145–153.
Balée, W. (2006). The research program of historical ecology. Annu. Rev. Anthropol. 35, 75–98. doi: 10.1146/annurev.anthro.35.081705.123231
Balick, M. J. (2007). “Ethnobotany, drug development and biodiversity conservation—exploring the linkages,” in Ciba Foundation Symposium 185 - Ethnobotany and the Search for New Drugs, eds D. J. Chadwick and J. Marsh (Chichester: John Wiley & Sons, Ltd.), 4–24.
Barreto, M. (2009). “A produção para a vida e a produção para o capital, a lógica camponesa e a lógica capitalista na extração da erva-mate nas comunidades de faxinais do Centro-Sul do Paraná,” in Encontro Nacional de Geografia Agrária (São Paulo).
Behling, H., and Pillar, V. D. (2007). Late Quaternary vegetation, biodiversity and fire dynamics on the southern Brazilian highland and their implication for conservation and management of modern Araucaria forest and grassland ecosystems. Philos. Trans. R. Soc. B Biol. Sci. 362, 243–251. doi: 10.1098/rstb.2006.1984
Berkes, F., and Folke, C. (2002). “Back to the future: ecosystem dynamics and local knowledge,” in Panarchy: Understanding Transformations in Human and Natural Systems, eds L. H. Gunderson and C. S. Holling (Washington, DC: Island Press), 121–146.
Berkes, F., Folke, C., and Colding, J. (2000). Linking Social and Ecological Systems: Management Practices and Social Mechanisms for Building Resilience. Cambridge: Cambridge University Press.
Berkes, F., and Turner, N. J. (2006). Knowledge, learning and the volution of conservation practice for social-ecological system resilience. Hum. Ecol. 34:479. doi: 10.1007/s10745-006-9008-2
Bitencourt, L. A. V., and Krauspenhar, P. M. (2006). Possible prehistoric anthropogenic effect on Araucaria angustifolia (Bert.) O. Kuntze expansion during the late holocene. Rev. Bras. Paleontol. 9, 109–116. doi: 10.4072/rbp.2006.1.12
Brandt, M., and Campos, N. J. (2008). Uso comum da terra e práticas associativas da população cabocla do planalto catarinense. Geosul 23, 43–64. doi: 10.5007/2177-5230.2008v23n45p43
Brehmer, J. S. (2005). Estudo de Extratos de Plantas Medicinais no Desenvolvimento Do Tumor ascítico de Ehrlich. Master's thesis, Universidade do Vale do Itajaí, Itajaí.
Capparelli, A., Hilgert, N., Ladio, A., Lema, V. S., Llano, C., Molares, S., et al. (2011). Paisajes culturales de Argentina: pasado y presente desde las perspectivas etnobotánica y paleoetnobotánica. Rev. Asoc. Argentina Ecol. Paisajes 2, 67–79.
Casas, A., Otero-Arnaiz, A., Pérez-Negrón, E., and Valiente-Banuet, A. (2007). In situ management and domestication of plants in Mesoamerica. Ann. Bot. 100, 1101–1115. doi: 10.1093/aob/mcm126
Clement, C. R. (1999). 1492 and the loss of amazonian crop genetic resources. I. The relation between domestication and human population decline. Econ. Bot. 53, 188–202. doi: 10.1007/BF02866498
Clement, C. R., De Cristo-Araújo, M., D'Eeckenbrugge, G. C., Alves Pereira, A., and Picanço-Rodrigues, D. (2010). Origin and domestication of native amazonian crops. Diversity 2, 72–106. doi: 10.3390/d2010072
Clement, C. R., Denevan, W. M., Heckenberger, M. J., Junqueira, A. B., Neves, E. G., Teixeira, W. G., et al. (2015). The domestication of Amazonia before European conquest. Proc. R. Soc. B Biol. Sci. 282:20150813. doi: 10.1098/rspb.2015.0813
Clement, C. R., and Junqueira, A. B. (2010). Between a pristine myth and an impoverished future. Biotropica 42, 534–536. doi: 10.1111/j.1744-7429.2010.00674.x
Clement, C. R., Rocha, S. F. R., Cole, D. M., and Vivan, J. L. (2007). “Conservação on farm,” in Recursos Genéticos Vegetais, ed L. L. Nass (Brasília: Embrapa Recursos Genéticos e Biotecnologia), 511–544.
Dawson, I. K., Guariguata, M. R., Loo, J., Weber, J. C., Lengkeek, A., Bush, D., et al. (2013). What is the relevance of smallholders' agroforestry systems for conserving tropical tree species and genetic diversity in circa situm, in situ and ex situ settings? A review. Biodivers. Conserv. 22, 301–324. doi: 10.1007/s10531-012-0429-5
Denevan, W. M. (2001). Cultivated landscapes of native Amazonia and the Andes. Oxford: Oxford University Press.
de Oliveira Suertegaray, C. E. (2002). Dinâmica da Cultura Erva-mate (Ilex paraguariensis St. Hil.) em Sistemas Agroflorestais e Monocultivo. Master's thesis, Universidade Federal de Santa Catarina, Florianópolis.
Duarte, A. S., Vieira-da-Silva, C., Puchalski, A., Mantovani, M., Silva, J. Z., and Reis, M. S. (2007). Estrutura demográfica e produção de frutos de Bromelia antiacantha Bertol. Rev. Bras. Plantas Med. 9, 106–112.
Edwin, G., and Reitz, R. (1967). “Aquifoliáceas,” in Flora Ilustrada de Santa Catarina, ed R. Reitz (Itajaí: Herbário Barbosa Rodrigues), 47.
Emperaire, L., and Peroni, N. (2007). Traditional management of agrobiodiversity in Brazil: a case study of manioc. Hum. Ecol. 35, 761–768. doi: 10.1007/s10745-007-9121-x
EPAGRI (2001). Dados e Informações Biofísicas da Unidade de Planejamento Regional Planalto Norte Catarinense – UPR 4. Florianópolis: CD-rom.
EPAGRI/CEPA (2016). Síntese Anual da Agricultura de Santa Catarina 2015-2016. Available online at: http://docweb.epagri.sc.gov.br/website_cepa/publicacoes/Sintese_2016.pdf (Accessed May 22, 2017).
EPAGRI/CEPA (2017). Preço médio Mensal ao Produtor – Estadual. Preços agrícolas. Available online at: http://www.epagri.sc.gov.br/?page_id=2711 (Accessed June 19, 2017).
FAO (1996). The State of the World's Plant Genetic Resources for Food and Agriculture. Rome: Food and Agriculture Organization of the United Nations.
Ferreira, A. G., Kaspary, R., Ferreira, H. B., and Rosa, L. M. (1983). Proporção de sexo e polinização em Ilex paraguariensis St. Hil. Bras. Florest. 13, 29–33.
Figueiredo-Filho, A., Orellana, E., Nascimento, F., Dias, A. N., and Inoue, M. T. (2011). Produção de sementes de Araucaria angustifolia em plantio e em floresta natural no centro-sul do estado do Paraná. Floresta 41, 155–162. doi: 10.5380/rf.v41i1.21196
Filippon, S. (2009). Aspectos da Demografia, Fenologia e uso Tradicional do Caraguatá (Bromelia antiacantha Bertol.) no Planalto Norte Catarinense. Master's Thesis, Universidade Federal de Santa Catarina, Florianópolis.
Filippon, S. (2014). Uso e Manejo de Caraguatá (Bromelia antiacantha) no Planalto Norte Catarinense: está em Curso um Processo de Domesticação? Ph.D. thesis, Universidade Federal de Santa Catarina, Florianópolis.
Filippon, S., Fernandes, C. D., Ferreira, D. K., Duarte, A. S., and Reis, M. S. (2012a). Produção de frutos para uso medicinal em Bromelia antiancatha (caraguatá): fundamentos para um extrastivismo sustentável. Rev. Bras. Plantas Med. 14, 506–513. doi: 10.1590/S1516-05722012000300013
Filippon, S., Fernandes, C. D., Ferreira, D. K., Silva, D. L. S., Altrak, G., Duarte, A. S., et al. (2012b). Bromelia antiacantha Bertol. (Bromeliaceae): caracterização demográfica e potencial de manejo em uma população no Planalto Norte Catarinense. Biodiversidade Bras. 2, 83–91.
Gerhardt, M. (2009). A produção da erva-mate na perspectiva da história ambiental. Rev. Bras. Agroecol. 4, 2678–2680.
Goudet, J. (2002). FSTAT, A Program to Estimate and Test Gene Diversities and Fixation Indices. Version 2.9.3.2.
Grzebieluka, D., and Sahr, C. L. L. (2009). Comunidades de faxinal e suas dinâmicas sócio-espaciais: da formação à desagregação de uma tradição no município de Tibagi (PR) - um estudo sobre o Faxinal dos Empoçados. Rev. Geogr. 4, 34–58. doi: 10.5380/geografar.v4i1.14427
Guerra, M. P., Silveira, V., Reis, M. S., and Schneider, L. (2002). “Exploração, manejo e conservação da araucária (Araucaria angustifolia),” in Sustentável Mata Atlântica: A Exploração de Seus Recursos Florestais, eds C. F. Lino and L. L. Simões (São Paulo: Senac), 85–102.
Hanisch, A. L., Gaertner, F. C., Hirano, E., and Bonfleur, R. (2006). Plano Territorial de Desenvolvimento Rural Sustentável do Planalto Norte Catarinense. Brasília: Ministério do Desenvolvimento Agrário.
Hanisch, A. L., Vogt, G. A., Marques, A., da, C., Bona, L. C., and Bosse, D. D. (2010). Estrutura e composição florística de cinco áreas de caíva no planalto norte de Santa Catarina. Pesqui. Florest. Bras. 30, 303–310. doi: 10.4336/2010.pfb.30.64.303
Heywood, V., Casas, A., Ford-Lloyd, B., Kell, S., and Maxted, N. (2007). Conservation and sustainable use of crop wild relatives. Agric. Ecosyst. Environ. 121, 245–255. doi: 10.1016/j.agee.2006.12.014
Hueck, K. (1972). As Florestas da América do Sul: ecologia, Composição e Importância Econômica. São Paulo: Polígono.
IBGE (2006). Instituto Brasileiro de Geografia e Estatística - Número de Estabelecimentos Agropecuários e Area dos Estabelecimentos. Censo Agropecuário. Available online at: http://www.ibge.gov.br/estadosat/temas.php?sigla=sc&tema=censoagro (Accessed April 12, 2017).
IBGE (2014). Instituto Brasileiro de Geografia e Estatística - Produção da Extração Vegetal e Silvicultura. Available online at: http://biblioteca.ibge.gov.br/visualizacao/periodicos/74/pevs_2014_v29.pdf
Iriarte, J., Smith, R. J., Gregorio de Souza, J., Mayle, F. E., Whitney, B. S., Cárdenas, M. L., et al. (2017). Out of Amazonia: late-Holocene climate change and the Tupi–Guarani trans-continental expansion. Holocene 27, 967–975. doi: 10.1177/0959683616678461
IUCN (2017). The IUCN Red List of Threatened Species. Version 2017-1. Available online at: www.iucnredlist.org
Kephart, S. R. (1990). Starch gel electrophoresis of plant isozymes: a comparative analysis of techniques. Am. J. Bot. 77, 693–712. doi: 10.2307/2444817
Kubo, R. R., Souza, G. C., and Miguel, L. A. (2008). “O extrativismo da samambaia-preta e os dilemas da busca de uma estatégia de desenvolvimento sustentável para a área de Mata Atlântica do Rio Grande do Sul,” in Extrativismo de Samambaia-Preta no Rio Grande do Sul, eds G. C. Souza, R. R. Kubo, and L. A. Miguel (Porto Alegre: Editora da UFRGS), 234–258.
Ladio, A. H. (2011). Traditional knowledge of edible wild native and exotic plants in the context of cultural change in human populations of arid Patagonia. Bioremed. Biodiv. Bioavailab. 5, 60–64.
Levis, C., Costa, F. R. C., Bongers, F., Peña-Claros, M., Clement, C. R., Junqueira, A. B., et al. (2017). Persistent effects of pre-Columbian plant domestication on Amazonian forest composition. Science 355, 925–931. doi: 10.1126/science.aal0157
Mantovani, A., Morellato, L. P. C., and Reis, M. S. (2004). Fenologia reprodutiva e produção de sementes em Araucaria angustifolia (Bert.) O. Kuntze. Rev. Bras. Bot. 27, 787–796. doi: 10.1590/S0100-84042004000400017
Marques, A. C., Hanisch, A. L., and Bona, L. C. (2008). Uso sustentável de áreas de vegetação de caívas e sua relação com os aspectos fisiológicos da produção de leite a pasto. Rev. Estud. Val. do Iguaçu 11, 129–140.
Marques, A. d. C. (2014). As Paisagens do Mate e a Conservação Socioambiental: um Estudo Junto aos Agricultores Familiares do Planalto Norte Catarinense. Ph.D. thesis, Universidade Federal do Paraná, Curitiba.
Martins, P. S. (2005). Dinâmica evolutiva em roças de caboclos amazônicos. Estud. Avançados 19, 209–220. doi: 10.1590/S0103-40142005000100013
Mattos, A. G. (2011). Caracterização das Práticas de Manejo e das Populações de Erva-mate (Ilex Paraguariensis A. Sant. Hil) Nativa em Exploração no Planalto Norte Catarinense. Master's thesis, Universidade Federal de Santa Catarina, Florianópolis.
Mattos, A. G. (2015). Conservação Pelo Uso de Populações de IIex Paraguariensis A. St. Hil, em Sistemas Extrativistas no Planalto Norte Catarinense. Ph.D. thesis, Universidade Federal de Santa Catarina, Florianópolis.
Mazuchowski, J. Z. (2004). Influência de Níveis de Sombreamento e de Nitrogênio na Produção de Massa Foliar da Erva-Mate Ilex Paraguariensis St. Hil. Master's Thesis, Universidade Federal do Paraná, Curitiba.
Mello, A. J. M. (2013). Etnoecologia e Manejo Local de Paisagens Antrópicas da Floresta Ombrófila Mista, Santa Catarina, Brasil. Master's Thesis, Universidade Federal de Santa Catarina, Florianópolis.
Mello, A. J. M., and Peroni, N. (2015). Cultural landscapes of the Araucaria Forests in the northern plateau of Santa Catarina, Brazil. J. Ethnobiol. Ethnomed. 11:51. doi: 10.1186/s13002-015-0039-x
Miranda, T. M., and Hanazaki, N. (2008). Conhecimento e uso de recursos vegetais de restinga por comunidades das ilhas do Cardoso (SP) e de Santa Catarina (SC), Brasil. Acta Bot. Brasilica 22, 203–215. doi: 10.1590/S0102-33062008000100020
Nodari, E. S., and de Carvalho, M. M. X. (2010). “As fases da exploração madeireira na floresta com Araucária e os progressivos avanços da indústria madeireira sobre as florestas primárias (1870–1970),” in Simpósio Internacional de História Ambiental e Migrações, 707–726. Available online at: www.labimha.ufsc.br
Otero-Arnaiz, A., Casas, A., and Hamrick, J. L. (2005). Direct and indirect estimates of gene flow among wild and managed populations of Polaskia chichipe, an endemic columnar cactus in Central Mexico. Mol. Ecol. 14, 4313–4322. doi: 10.1111/j.1365-294X.2005.02762.x
Paquette, S. R. (2012). PopGenKit: Useful Functions for (Batch) File Conversion and Data Resampling in Microsatellite Datasets. R Package Version 1.0.
Parra, F., Blancas, J. J., and Casas, A. (2012). Landscape management and domestication of Stenocereus pruinosus (Cactaceae) in the Tehuacán Valley: human guided selection and gene flow. J. Ethnobiol. Ethnomed. 8:32. doi: 10.1186/1746-4269-8-32
Petersen, P., Tardín, J. M., and Marochi, F. (2001). Del manejo extractivo al regenerativo, el caso de los bosques de araucaria en el Paraná. LEISA Rev. Agroecol. 16, 8–9.
Pires, E. Z., Stedille, L. I. B., Machado, S., Mantovani, A., and Lopes da Costa, R. (2014). Biologia reprodutiva de erva-mate (Ilex paraguariensis A. St. Hil.) em remanescente de Floresta Ombrófila Mista Altomontana. Rev. Ciênc. Agrovet 13, 171–180.
R Development Core Team (2015). R: A Language and Environment for Statistical Computing.Vienna: R Foundation for Statistical Computing. Available online at: http://www.r-project.org/
Reis, M. S., Ladio, A., and Peroni, N. (2014). Landscapes with Araucaria in South America: evidence for a cultural dimension. Ecol. Soc. 19:art43. doi: 10.5751/ES-06163-190243
Reis, M. S., Mantovani, A., Silva, J. Z., Mariot, A., Bittencourt, R., Nazareno, A. G., et al. (2012). “Distribuição da diversidade genética e conservação de espécies arbóreas em remanescentes florestais de Santa Catarina,” in Inventário Florístico Florestal de Santa Catarina, Vol. 1, Diversidade e Conservação dos Remanescentes Florestais, eds A. C. Vibrans, L. Sevegnani, A. L. de Gasper, and D. V. Lingner (Blumenau: Edifurb), 143–169.
Reis, M. S., Peroni, N., Mariot, A., Steenbock, W., Filippon, S., Vieira-da-Silva, C., et al. (2010). “Uso sustentável e domesticação de espécies da Floresta Ombrófila Mista,” in Agrobiodiversidade no Brasil: Experiências e Caminhos da Pesquisa, eds C. M. Lin, C. M. Amorozo, and C. W. Kffuri (Recife: NUPEEA), 183–214.
Reis, M. S., Vieira-da-Silva, C., Mattos, A. G., Zechini, A. A., Mantovani, A., and Peroni, N. (2013). “Caívas and their contribution to the conservation of Atlantic forest landscapes in Brazil,” in Community Biodiversity Management: Promoting Resilience and the Conservation of Plant Genetic Resources, eds W. S. de Boef, A. Subedi, N. Peroni, M. Thijssen, and E. O'Keeffe (Routledge), 151–155. Available online at: http://www.scopus.com/inward/record.url?eid=2-s2.0-84917490753&partnerID=40&md5=6094c0f0aaaf47cfff0e4fcd6035746d
Reitz, R. (ed.). (1983). “Bromeliáceas e a malária - bromélia endemica,” in Flora Ilustrada Catarinense (Itajaí: Herbário Barbosa Rodrigues), 808.
Reitz, R., and Klein, R. M. (1966). “Araucariáceas,” in Flora Ilustrada Catarinense, ed R. Reitz (Itajaí: Herbário Barbosa Rodrigues), 62.
Ribeiro, M. C., Metzger, J. P., Martensen, A. C., Ponzoni, F. J., and Hirota, M. M. (2009). The Brazilian Atlantic Forest: how much is left, and how is the remaining forest distributed? Implications for conservation. Biol. Conserv. 142, 1141–1153. doi: 10.1016/j.biocon.2009.02.021
Roberts, P., Hunt, C., Arroyo-Kalin, M., Evans, D., and Boivin, N. (2017). The deep human prehistory of global tropical forests and its relevance for modern conservation. Nat. Plants 3:17093. doi: 10.1038/nplants.2017.93
Santos, D. S., Puchalski, A., Gomes, G. S., Mantovani, M., Silva, J. Z., and Reis, M. S. (2004). Variação no período de germinação de sementes em uma população natural de Bromelia antiacantha Bertol. Rev. Bras. Plantas Med. 6, 35–41.
Schuster, W. T., and Sahr, C. L. L. (2009). O faxinal do presente e o faxinal do passado: evolução do uso da terra no faxinal Saudade Santa Anita Turvo (PR). XIX Econtro Nac. Geogr. Agrária 1–21.
Shepard, G. H., and Ramirez, H. (2011). “Made in Brazil”: human dispersal of the Brazil nut (Bertholletia excelsa, Lecythidaceae) in ancient Amazonia. Econ. Bot. 65, 44–65. doi: 10.1007/s12231-011-9151-6
Steenbock, W., and Reis, M. S. (2013). Domesticação da paisagem em bracatingais de assentamentos rurais no Noroeste do Planalto Catarinense. Ciência Florest. 23, 427–437. doi: 10.5902/1980509810554
Steenbock, W., Siminski, A., Fantini, A. C., and Reis, M. S. (2011). Ocorrência da bracatinga (Mimosa scabrella Benth.) em bracatingais manejados e em florestas secundárias na região do Planalto Catarinense. Rev. Árvore 35, 845–857. doi: 10.1590/S0100-67622011000500010
Trajano, N. G. (1996). A constituição das cndústrias Klabin de papel e celulose S/A. História Ensino 2, 129–137. doi: 10.5433/2238-3018.1996v2n0p129
Trombulak, S. C., Omland, K. S., Robinson, J. A., Lusk, J. J., Fleischner, T. L., Brown, G., et al. (2004). Principles of conservation biology: recommended guidelines for conservation literacy from the education committee of the Society for Conservation Biology. Conserv. Biol. 18, 1180–1190. doi: 10.1111/j.1523-1739.2004.01851.x
Valentini, D. J. (2003). Da Cidade Santa à Corte Celeste: Memórias de Sertanejos e a Guerra do Contestado, 3rd Edn. Caçador: Universidade do Contestado.
Vieira-da-Silva, C., Martins, G., Steiner, N., Santos, K. L., Camargo, R., Mantovani, A., et al. (2011). “Araucaria angustifolia,” in Espécies Nativas da Flora Brasileira de Valor Econômico Atual ou Potencial: Plantas Para o Futuro – Região Sul, eds L. Coradin, A. Reis and A. Siminski (Brasília: Ministério do Meio Ambiente), 134–150.
Vieira-da-Silva, C., and Miguel, L. A. (2014). Extrativismo e Abordagem Sistêmica. Novos Cad. do NAEA 17, 189–217. doi: 10.5801/ncn.v17i2.1580
Vieira-da-Silva, C., and Reis, M. S. (2009). Produção de pinhão na região de Caçador, SC: aspectos da obtenção e sua importância para comunidades locais. Ciência Florest. 19, 363–374. doi: 10.5902/19805098892
Wiens, J. A. (1997). “Metapopulation dynamics and landscape ecology,” in Metapopulation Biology, eds L. Hanski and M. E. Gilpin (San Diego, CA: Academic Press), 43–62.
Wiersum, K. F. (1996). Domestication of valuable tree species in agroforestry systems: evolutionary stages from gathering to breeding. Domest. Commer. non-timber For. Prod. Agrofor. Syst. FAO Tech. Pap. Non-wood For. Prod. 9, 147–158.
Wiersum, K. F. (1997a). From natural forest to tree crops, co-domestication of forests and tree species, an overview. Netherlands J. Agric. Sci. 45, 425–438.
Wiersum, K. F. (1997b). Indigenous exploitation and management of tropical forest resources: an evolutionary continuum in forest-people interactions. Agric. Ecosyst. Environ. 63, 1–16. doi: 10.1016/S0167-8809(96)01124-3
Zechini, A. A., Schussler, G., Silva, J. Z., Mattos, A. G., Peroni, N., Mantovani, A., et al. (2012). Produção, comercialização e identificação de variedades de pinhão no entorno da Floresta Nacional de Três Barras – SC. Biodiversidade Bras. 2, 74–82.
Zizumbo-Villarreal, D., Vargas-Ponce, O., Rosales-Adame, J. J., and Colunga-GarcíaMarín, P. (2013). Sustainability of the traditional management of Agave genetic resources in the elaboration of mezcal and tequila spirits in western Mexico. Genet. Resour. Crop Evol. 60, 33–47. doi: 10.1007/s10722-012-9812-z
Keywords: cultural landscapes, Araucaria angustifolia, Ilex paraguariensis, Bromelia antiacantha, genetic diversity, ethnobotany, local management system, non-timber forest products
Citation: Reis MS, Montagna T, Mattos AG, Filippon S, Ladio AH, Marques AdC, Zechini AA, Peroni N and Mantovani A (2018) Domesticated Landscapes in Araucaria Forests, Southern Brazil: A Multispecies Local Conservation-by-Use System. Front. Ecol. Evol. 6:11. doi: 10.3389/fevo.2018.00011
Received: 29 July 2017; Accepted: 22 January 2018;
Published: 07 February 2018.
Edited by:
Pan Kaiwen, Chengdu Institute of Biology (CAS), ChinaReviewed by:
Fernando José Cebola Lidon, Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa, PortugalIndira Devi Puthussery, Kerala Agricultural University, India
Copyright © 2018 Reis, Montagna, Mattos, Filippon, Ladio, Marques, Zechini, Peroni and Mantovani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maurício S. Reis, bXNlZHJlekBnbWFpbC5jb20=