 Jose de Jesus Luna-Ruiz
Jose de Jesus Luna-Ruiz Gary P. Nabhan
Gary P. Nabhan Araceli Aguilar-Meléndez
Araceli Aguilar-Meléndez- 1Centro de Ciencias Agropecuarias, Universidad Autonoma de Aguascalientes, Aguascalientes, Mexico
- 2The Southwest Center, University of Arizona, Tucson, AZ, United States
- 3Centro de Investigaciones Tropicales, Universidad Veracruzana, Xalapa, Mexico
We propose that comparisons of wild and domesticated Capsicum species can serve as a model system for elucidating how crop domestication influences biotic and abiotic interactions mediated by plant chemical defenses. Perhaps no set of secondary metabolites (SMs) used for plant defenses and human health have been better studied in the wild and in milpa agro-habitats than those found in Capsicum species. However, very few scientific studies on SM variation have been conducted in both the domesticated landraces of chile peppers and in their wild relatives in the Neotropics. In particular, capsaicinoids in Capsicum fruits and on their seeds differ in the specificity of their ecological effects from broad-spectrum toxins in other members of the Solanaceae. They do so in a manner that mediates specific ecological interactions with a variety of sympatric Neotropical vertebrates, invertebrates, nurse plants and microbes. Specifically, capsaicin is a secondary metabolite (SM) in the placental tissues of the chile fruit that mediates interactions with seed dispersers such as birds, and with seed predators, ranging from fungi to insects and rodents. As with other Solanaceae, a wide range of SMs in Capsicum spp. function to ecologically mediate the effects of a variety of biotic and abiotic stresses on wild chile peppers in certain tropical and subtropical habitats. However, species in the genus Capsicum are the only ones found within any solanaceous genus that utilize capsaicinoids as their primary means of chemical defense. We demonstrate how exploring in tandem the evolutionary ecology and the ethnobotany of human-chile interactions can generate and test novel hypotheses with regard to how the domestication process shifts plant chemical defense strategies in a variety of tropical crops. To do so, we draw upon recent advances regarding the chemical ecology of a number of wild Capsicum species found in the Neotropics. We articulate three hypotheses regarding the ways in which incipient domestication through “balancing selection” in wild Capsicum annuum populations may have led to the release of selective biotic and abiotic pressures. We then analyze which shifts under cultivation generated the emergence of Capsicum chemotypes, morphotypes and ecotypes not found in high frequencies in the wild. We hypothesize that this “competitive release” can lead to a diversification of the domesticate's investment in a greater diversity of SM potency across different cultural uses, cropping systems and ecogeographic regions. While most studies of plant domestication processes focus on morphological changes that confer greater utility or productivity in human-managed environments, we conclude that changes in the chemical ecology of a useful plant can be of paramount importance to their cultivators. The genus Capsicum can therefore provide an unprecedented opportunity to compare the roles of SMs in wild plants grown in natural Neotropical ecosystems with their domesticated relatives in the milpa agro-ecosystems of Mesoamerica. Even with the current depth of knowledge available for crop species in the genus Capsicum and Solanum, our understanding of how particular SMs affect the reproduction and survival of wild vs. domesticated solanaceous plants remains in its infancy.
Introduction
What changes occur in a Neotropical plant's chemical defenses when it is domesticated for crop production as a food, medicine, vermifuge or condiment, or for all four of these uses? There is remarkably little tested ecological theory regarding how domestication affects plant chemical defenses (Rindos, 1984; Johns, 1990; Casas et al., 2015). This may be because most phytogeographic, agroecological, and archaeobotanical studies of plant domestication have largely used morphological indicators to track the domestication process rather than identifying phytochemical indicators of changes in ecological interactions. As recently argued by Zeder (2017), ecologists need to identify tractable model systems that allows for an assessment of the core assumptions of the Extended Evolutionary Synthesis (EES).
The domestication of crop plants by human cultures provides one such case study opportunity. That is why we propose that the genus Capsicum can serve as important model system for discerning how changes in secondary metabolites (SMs) reveal shifts in plant chemical defenses that have occurred with domestication. In the case of domesticated chile peppers, these shifts influenced both (1) antagonistic interactions with predators and abiotic pressures, and (2) facilitated (including mutualistic) interactions among chiles, their avian dispersers, nurse plants and human cultures. The integration of ethnobotanical, paleoecological, archeological, linguistic, genetic and evolutionary perspectives on chile domestication that has been in process for the last two decades (Tewksbury and Nabhan, 2001; Pickersgill, 2007; Tewksbury et al., 2008b; Aguilar-Meléndez et al., 2009; Haak et al., 2012; Kraft et al., 2014; Carrizo-García et al., 2016) has already contributed substantively to the possibilities of such an EES.
For these reasons, we have chosen to integrate ecological studies of wild Capsicum species in natural habitats of the Neotropics with ethnobotanical, agroecological and nutritional studies of domesticated Capsicum landraces in culturally-managed milpa habitats and indigenous diets of Mesoamerica. By doing so, we wish to further test hypotheses underpinning the theory that a major trend in crop evolution in the Neotropics has been a dramatic shift in the ecological functionality of plant chemical defenses (Johns, 1990). We propose that testing the following three hypotheses can help identify the most parsimonious fit with data and trends involving the evolutionary transition from wild Capsicum annuum var. grabriusculum to domesticated Capsicum annuum var. annuum:
(H1) a reduction and simplification of the potency of plant chemical defenses against seed predators, foliage herbivores and disease microbes with greater reliance on human intervention to protect the plants;
(H2) a diversification of the levels of potency and mixes of defense chemicals, given the wider range of habitats, cultural management and uses, and broader geography to which the crop chile plants are exposed;
(H3) an intensification of the potency of certain plant chemical defenses, given the need to protect the plants in agro-habitats where they occur at higher density and without as much beta diversity of neighboring plant species to slow the spread of predators, herbivores, competing weeds or diseases.
Crop plants in the Solanaceae (including Capsicum chile peppers) may be extremely useful models for looking at changes in potency, diversity or effectiveness of plant chemical defenses which occur with domestication. This is because their SMs and the ecological roles which these plant defenses play have been intensively investigated in the field and in the laboratory for well over two centuries (Johns, 1990; Eich, 2008). Neverthless, it remains clear that we lack the detailed knowledge needed to determine how particular plant chemical defenses (e.g., specific capsaicinoids) function in repelling (or attracting) various sets of vertebrates, invertebrates and fungi which serve as seed predators, seed dispersers, fruit and foliage consumers or root parasites on various solanaceous crops. Even with the current depth of knowledge available for crop species in the genus Capsicum and Solanum, our understanding of how particular SMs affect the reproduction and survival of wild vs. domesticated solanaceous plants remains in its infancy.
Of the 97–102 genera represented by 2300–2460 distinct species documented in the Solanaceae (Hunziker, 2001; Eich, 2008), SMs (such as the ornithine-derived alkaloids which function as the primary chemical defenses of most of these species) have so far been documented in more than 61 genera (Eich, 2008). Many of the SMs commonly found in the Solanaceae—such as tropane, nicotinoid, pyridine and terpenoid alkaloids—can be toxic or at least repellant to a broad variety of insects as well as to vertebrate herbivores; some also reduce fungal or bacterial infestations of seeds, fruit or foliage. We will focus the rest of this inquiry on the ecological and ethnobotanical consequences of these chemical defenses as found in seeds and fruits of solanaceous crops, with particular focus on chile peppers (Capsicum annuum).
These broad-spectrum alkaloids function as primary chemical defenses in a number of solanaceous crop plants, and in their wild relatives as well. We have therefore placed the domestication of Capsicum species in the context of other domestication studies for the following genera: Jaltomata (xaltomatl, sogorome); Lycium (goji berry); Nicotiana (tobacco); Solanum (potato, tomato, eggplant, garden huckleberry/chichiquelite); Physalis (tomatillo/ground cherry, cape gooseberry/uchuva) (e.g., Johns, 1990; Pickersgill, 2007 among many others). While some of the same alkaloids characteristic of many species in the Solanaceace are present in extremely low concentrations in the foliage of Capsicum species, nearly all the species in this genus have taken up an altogether different strategy—Capsaicinoids, for defending their seeds and fruits from biotic stresses.
Departing from the norm in the Solanaceae—where species principally use broad-spectrum and highly toxic glyco-alkaloids for defense—most Capsicum species instead employ another, unique set of SMs that are not appreciably toxic to animals. In contrast to all other genera and species in the nightshade family, both wild and domesticated chile peppers produce several of the 22 known capsaicinoids, with capsaicin, dihydrocapsaicin and nordihydrocapsaicin being the most prevalent, widely-studied and economically important ones. However, it is likely that each distinct capsaicinoid functions in varying degrees to direct avian seed dispersal or to repel and reduce damage by insects, mammals, bacteria and fungi (esp. Fusarium) (Tewksbury and Nabhan, 2001; Tewksbury et al., 2008b; Haak et al., 2012). Unfortunately, to arrive at a comprehensive EES (Zeder, 2017), we will require more detailed knowledge on the specific ecological effects of 19 of those distinctive capsaicinoids on various faunal and fungal species found in Neotropical habitats.
The ability to produce capsaicinoids is a monophyletic synapomorphic carácter shared by most of the 35 + wild Capsicum species. The exceptions are few, and are found in the wild Andean clade (C. ciliatum = C. rhomboideum, C. scolnikianum, C. geminifolium, C. lanceolatum, and C. dimorphum), and the Longidentatum clade (C. longidentatum) (Eich, 2008; Haak et al., 2012; Carrizo-García et al., 2016).
Pungency in all other wild chile peppers has a simple genetic basis that is expressed only in glands within the placental fruit, where it serves to protect viable seeds from predation by granivorous mammals, or from microbial infestation. It also facilitates the directed dispersal of seeds by frugivorous birds such as thrashers, cardinals, and finches to safe sites for germination and recruitment under nurse plants, providing an unusually direct ecological link to changes in reproductive fitness that is often missing from studies of chemical ecology (Nabhan, 2004; Tewksbury et al., 2008a). Pungency is polymorphic in several wild chile species (Carrizo-García et al., 2016), and such polymorphic populations have been identified along natural environmental gradients (Haak et al., 2012; Carrizo-García et al., 2016). These polymorphisms provide unique opportunities to advance an extended evolutionary synthesis from field comparisons of wild and domesticated subspecies in the same crop species and economic genus (Hernández-Verdugo et al., 2001a; Haak et al., 2012; Chen et al., 2015).
These attributes make chile peppers excellent systems through which to investigate the evolution of adaptive constraints found under various levels of domestication.
Ironically, consumption of the very same capsaicinoids that function as chemical defenses for chile plants have long been used by Mesoamerican cultures as defenses against microbial and invertebrates challenging human health (Nabhan, 2004). Their many indigenous uses as food or medicine has likely benefited overall human health and reproductive fitness in Neotropical environments for well over six millennia (Perry and Flannery, 2007; Kraft et al., 2014); these biomedically-significant ethnobotanical uses mediated by SMs (Mostafa-Kamal et al., 2015) possibly triggered the domestication and diversification of chile peppers.
Capsaicinoids are now the most widely used SMs in the world, even though their commercial production is dominated by landraces of just five species in the genus Capsicum. Now culturally-dispersed far beyond the Neotropics, each continent and its biomes favors different ecotypes of place-based landraces such as the tabasco pepper, ghost pepper, piri-piri, aji, habanero, jalapeño, and long green New Mexican chile. Today, more than a third of the world's human population daily consume food products derived from 2500+ landraces, standard varieties and modern hybrids of chile peppers (Tewksbury et al., 2008b). In fact, we predict that if one includes the number of human daily ingesting and topically-applying chile peppers as pharmaceuticals and folk medicinals then over half the world's population are currently consuming some form of chile peppers for nourishment, health and ultimately, reproductive fitness.
We will focus most of our analysis on discerning historic shifts in plant chemical defenses in the most widely-used Capsicum species – C. annuum L., domesticated in the dry subtropical habitats of Mesoamerica over 6,500 years ago (Kraft et al., 2014). We posit that these shifts in SM enhanced, or at least diversified, the mutualistic relationships among chile peppers and indigenous Mesoamerican cultures, as a result of relatively rapid selection and linguistically-traceable diffussion, that intensified around 6,500 years B.P. (Brown, 2010; Kraft et al., 2014).
It appears that Homo sapiens is one of the few mammalian species which routinely overcome a deep-seated aversion to the consumption of pungent chile peppers (Rozin and Schiller, 1980; Nabhan, 2004), perhaps because the evolutionary benefits of consuming chile fruits outweighed the costs when exposed to environmental challenges, commonly exhibited in certain Neotropical habitats.
Crop Domestication
Domestication is the outcome of both conscious and unconcious selection processes that lead to increased co-evolutionary adaptation of plants to cultivation and utilization by humans in managed environments (Gepts, 2010). Paleolithic cultures developed tools, food preparation and plant selection techniques for detoxifying certain plant foods rich in SMs (Johns and Kubo, 1988; Johns, 1990). As such, the coevolutionary response of Mesoamerican cultures to chile peppers certainly included memes, but may also have included the selection of “non-taster” genes in humans for organoleptic tolerance of pungency and bitterness (Nabhan, 2004).
On the other hand, the suite of traits that marks the divergence from its wild ancestor(s) has been defined as the “domestication syndrome” (Harlan, 1992). A domestication syndrome may include selection for combinations of several different morphological and phytochemical traits, including seed retention (non-shattering), increased fruit and/or seed size, changes in branching and stature, changes in reproductive strategy, and, importantly, changes in SMs (Pickersgill, 2007; Gepts, 2010; Meyer et al., 2012).
Often, domestication selects against traits that formerly increased the plant's defensive or reproductive successes in natural environments (Meyer et al., 2012). However, this generalization may not completely fit for SMs such as capsaicinoids in C. annuum in the Neotropics, where a high diversity of landraces and wild populations express some degree of pungency as a natural defense against predators.
Cultural selection can therefore work in opposition to natural selection, and certain domesticated crops may exhibit reduced fitness, or, in some cases, an inability to survive outside of cultivation (Pickersgill, 2007; Gepts, 2010). The very act of moving plants from natural habitats into culturally-managed habitats such as milpas alters the mix of selection pressures, leading to increased adaptation to cultivation, and to actual physical protection from pests and predators by cultural managers, potentially at the expense of traits conferring fitness in the natural environment (Meyer et al., 2012). In the very least, selection pressures for plant chemical defenses against predators might be relaxed if human intervention with the same predators (eg., rodents) is consistently offered to the crop variety over multiple generations.
Secondary Metabolites in Plants
Plant chemicals can be divided into two major categories: primary metabolites (PMs) and secondary metabolites (SMs). PMs are substances produced by all plant cells that are directly involved in growth, development, or reproduction (sugars, proteins, amino acids, and nucleic acids). PMs function in basic anabolic and catabolic processes required for respiration, nutrient assimilation, and growth/development (Kliebenstein, 2004; Freeman and Beattie, 2008).
SMs may not be directly involved in growth or reproduction, but they are often involved with plant defense (Freeman and Beattie, 2008), particularly in the case of Capsicum species (Tewksbury et al., 2008b). SMs are considered the major mediators of ecological interactions of plants as a result of their large and diverse biological functions in nature. SMs are produced in response to certain biotic and/or abiotic stress signals or stimuli. They function in the defense against herbivores, microbes, viruses or competing plants, and also as signal compounds to attract pollinating or seed dispersing animals (Wink, 2003). Thus, SMs are very important for plant's survival and reproductive fitness. This complex multirole of SM has led plants to synthesize many different chemical compounds in nature during evolution (Kliebenstein, 2004).
According to their role in plant's defense, SMs have been classified on the basis of their host protection and fostering of beneficial biotic interactions. According to Freeman and Beattie (2008), SMs usually belong to one of three large chemical classes: terpenoids, phenolics, and alkaloids.
Terpenoids include a series of toxic and non-toxic phytochemicals produced in different plant organs that inhibit, repel, or attract other living organisms, such as predators (plant pathogens, herbivores invertebrates, vertebrates) and non-predators (dispersers, pollinators, pest-enemies).
Phenolics include a series of toxic and non-toxic compounds such as flavonoids, isoflavonoids, and phenolic monomers produced in different organs (roots, stems, leaves, flowers, fruits, and seeds). Phenolics and their derivatives have different functions in nature (UV-protectan, antifungal, antibiotic, insecticidal, and others).
Alkaloids are N-compounds produced and aggregated in different organs such as roots, leaves, fruits and seeds. Alkaloid-based SMs may function as bactericides, fungicides, insecticides and allelopathics. Alkaloids may have degrading and digestive effects on different tissues of predators and pathogens. Examples of this type of SM include cafeine, cocaine, morphine, nicotine, atrophine, plus capsaicine and other capsaicinoids. Other N-compounds important for plant chemical defense include cyanogenic glucosides, defensins, lectins, and hydrolitic enzymes.
Therefore, SMs in chile peppers and other solanaceous plants in Neotropical habitats have evolved as defense mechanisms against microorganisms (viruses, bacteria, fungi), herbivores (molluscs, hemipteran insects, vertebrates), and competing plants. They may also function to attraction of pollinators and seed dispersers by virtue of their fragrances and colors they express in the plants. Regardless of the efficacy of such benefits, SMs require a great deal of plant resources and energy to be produced. Consequently, they may be synthesized and translocated after a pathogen or pest has attacked the plant and triggered their activation. Once activated, these chemical defensive compounds are usually very effective inhibitors of fungi, bacteria, nematodes, and hemipteran insect herbivores.
Chemical Ecology of Wild Capsicum in Neotropical Habitats
To address the changes in plant chemical defenses that have occurred with the domestication of Capsicum annuum, we must briefly establish the context through which wild chile peppers and other solanaceous plants deal with biotic and abiotic stresses prevalent in the Neotropics. In particular, we will focus on the biotic interactions as well as the biotic and abiotic stresses that wild chile plants may particularly respond to in dry subtropical thornscrub and tropical deciduous forest vegetation types, characteristic of the Sierra Madre Oriental and the Trans-Volcanic Belt in Mesoamerica. At least one EES-style integration has determined that these vegetation types are among the most likely Neotropical habitats where C. annuum domestication and diffusion may have occurred (Kraft et al., 2014). However, because there has been considerable change in the areas covered by these habitat types over the last 6500 years (Kraft et al., 2014), other proposed geographic areas such as the Yucatan peninsula remain viable enough as putative centers of chile pepper domestication that we do not wish to rule them out (Aguilar-Meléndez et al., 2009).
In contrast, the pungency of wild chile pepper fruit repels small mammals that function as seed predators, but directs their dispersal to safe sites under nurse trees where germination, recruitment and establishment have higher probabilities (Tewksbury and Nabhan, 2001; Carlo and Tewksbury, 2014). The seeds from these pungent wild chiles are also protected from “predation” by Fusarium fungi that might otherwise leave the infected seeds inviable (as evidence shows for C. chacoense). Thus, the directed dispersal adaptations of wild chile peppers afforded to them by the pungency of their specialized SMs–their capsaicinoids—have conferred to them a level of reproductive fitness that has incidentally allowed them to be present in abundance and accessible to human foragers in the Neotropics for millennia.
Changes in Secondary Metabolite Intensity With Chile Domestication
What are the traits that have been modified as a result of selection under cultivation that have made modern and fully domesticated varieties of chile peppers so poorly adapted to the natural Neotropical habitats? We propose that the morphological and/or phenotypic changes which occurred during cultural selection and domestication of C. annuum have been accompanied by (if not surpassed in importance by) corresponding changes in SMs that regulate ecological interactions of chile peppers with their surrounding abiotic and biotic environments. The complexity and specificity of SMs as chemical mediators of biotic interactions of both wild and domesticated C. annuum in the Neotropics are summarized in Figure 1.
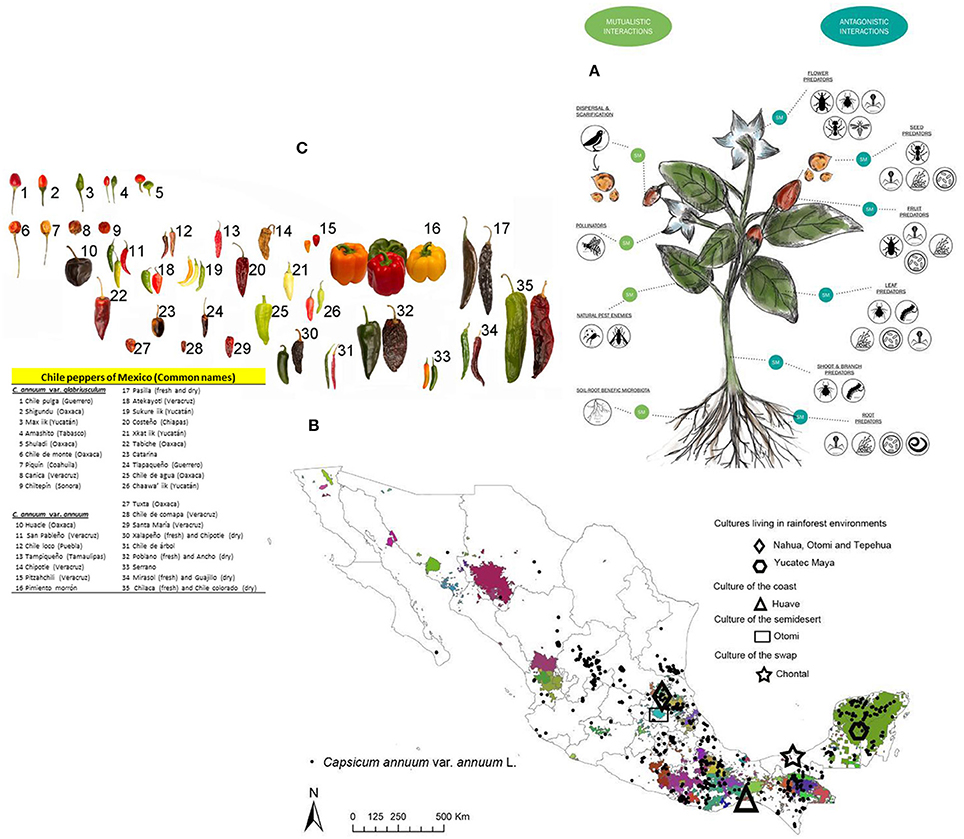
Figure 1. Ecological and cultural interactions shaping diversity of chile peppers (Capsicum annuum L.) across Mexico. (A) Graphic illustration of SMs as chemical mediators of ecological interactions with wild C. annuum in natural and semi-managed habitats. (Illustration designed by Frida Isabel Luna-Vallejo). (B) Map of Mexico showing indigenous territories, contrasted by colors. The symbols identify particular ecological zones where certain indigenous groups have persisted in modern times. All indigenous groups represented here have documented uses of chile peppers. (Map elaborated by Andres Lira Noriega and Araceli Aguilar-Melendez based on data from the authors, SINAREFI-SNICS-SAGARPA and SNIB/CONABIO 2016; the layer of indigenous territories was provided by Eckard Boege). (C) A representative sample of the wider array current morpho-typic diversity and levels of domestication of chile peppers across Mexico. (Photos by Ivan Montes de Oca Cacheux and Miguel Angel Sicilia Manzo/Image repository CONABIO).
Wild populations of chile pepper have coexisted and coevolved with many different organisms of tropical origin. Figure 1A focuses on two types of biotic interactions with wild Capsicum species: mutualistic and antagonistic. Every particular plant interaction is regulated by some SM produced and expressed in a particular organ, at a certain phenological stage, in response to specific biotic or abiotic signals. Chile pepper interactions have been strongly influenced by humans and cultural diversity in Mesoamerica over the last 10,000 years. The cultural diversity present in modern Mexico, and a sample of the wide morphological variation and levels of domestication that are currently found in Mexican chile peppers are shown in Figures 1B,C. The variation in Mexican chile peppers also applies to the chemical compounds, which may help explain the wide differences in fruit taste and flavor for different purposes and uses across Mexico.
Chen et al. (2015) indicated that among their various functions, SMs play particularly important roles in insect-plant interactions. Studies that have compared chemical defense traits in wild crop relatives and their cultivated counterparts are increasing in number, and their outcomes consistently show that domesticated plants provide a better food resource for herbivores than their more toxic wild progenitors. Several studies provide evidence of such changes in the chemical ecology and biotic interactions along a domestication gradient (Holt and Birch, 1984; Benrey et al., 1998; Rodriguez-Saona et al., 2011; Dávila-Flores et al., 2013). These widely-observed trends seem to contextualize, if not explain, shifts in the chemical defenses of C. annuum during its domestication in certain but not all, Neotropical habitats of Mesoamerica.
To date, most studies of SMs in C. annuum in Mesoamerica have been focused on fruits of fully domesticated commercial varieties for consumption as fresh fruits (jalapeño, serrano, ancho and sweet pepper morphotypes). In addition, there are few ecological field studies of how capsaicinoids in wild Capsicum species of arid North America and tropical South America mediate relationships with native fauna, but they do not specify which capsaicinoid(s) drive those interactions (Tewksbury and Nabhan, 2001; Tewksbury et al., 2008a; Carlo and Tewksbury, 2014; Haak et al., 2014). Most analyses have concentrated on capsaicinoids and few have included other SMs, such as phenolics and carotenoids. The literature available on SMs in chile peppers is focused on their presence in both, vegetative organs and in fruits and seeds (Do Rêgo et al., 2012; Kim et al., 2014). The presence of SMs in different organs and genotypic backgrounds may help explain the existence of natural sources of genetic resistance in Capsicum to particular herbivorous pests and seed predators.
The identities of most SMs remains incomplete among wild C. annuum var. glabriusculum from the Neotropics. However, genetic resistance to Huasteco pepper virus has been documented for wild C. annuum from Nortwest Mexico (Hernández-Verdugo et al., 2001b; Retes-Manjarrez, 2016). Of the known cases of genetic resistance among domesticated chile peppers are their tolerance to Phytophthora capsici and root knot nematodes, first documented in the Criollo de Morelos landrace—CM-334 (Pegard et al., 2005) also, leaf phenolic extracts from domesticated chile landraces have been used to control Alternaria altata in tomatoes.
Crop domestication can lead to a decrease in SMs associated with pest resistance, a trend corroborated by Meyer et al. (2012); they found a decline in levels of some SMs across 203 separate crop varieties, relative to levels in their wild progenitors, including C. annuum. However, other SMs, such as capsaicinoids, have dramatically increased within some natural and domesticated chile pepper landraces (e.g., Bhut Jolokia; Bosland and Baral, 2007), so that these changes are not unidirectional.
Given that “original” contexts for how wild Capsicum species function and survive in the Neotropics, Table S1 proposes a set of differences that may have been triggered by “balancing selection” during the domestication process. Balancing selection operated in ways that transformed some wild polymorphic populations into fully-domesticated but still heterogeneous C. annuum landraces. We place particular emphasis on levels of SMs and other adaptations that appear to confer reproductive fitness to Capsicum populations in Neotropical habitats.
Other Changes Occurring With Domestication of Chile Peppers
We do not wish to presume that shifts in SMs were the only changes which have occurred with the domestication of Capsicum species in Neotropical habitats. We wish to briefly mention several other traits of adaptive significance in Neotropical habitats.
Loss of Dispersal Mechanisms
Wild chile peppers are naturally dispersed by frugivorous birds to the understory of selected nurse plants (Tewksbury and Nabhan, 2001; Carlo and Tewksbury, 2014), while domesticated chiles depend on human intervention for dispersal. Seed dispersal often involves lost of an abscission zone from some part of the plant. Fruits of wild chile peppers separate easily from the receptacle at maturity. Fruits of domesticated peppers remain firmly attached to the plant. Mature wild chile pepper fruits are consumed and effectively dispersed by a variety of frugivorous Neotropical birds. Domesticated peppers are either too large, or are not attractive to nor dispersed by most Neotropical birds. Different SMs may mediate seed dispersal in wild chiles, but carotenoids in the fruit pulp probably are likely the most important due to bird attraction by their red color. The pyrazine fragrances of chile peppers may also serve to attract certain birds.
Loss of Seed Dormancy
Most wild chile pepper seeds have staggered seed dormancy, which allows germination and recruitment when optimal conditions occur in a more variable and uncertain environment. Domesticated chiles do not exhibit any seed dormancy (Pickersgill, 2007). Therefore, domesticated chiles would likely have poor recruitment, survival and fitness if placed in most naturally wild environments. Seed dormancy in most wild Capsicum species is mediated by SMs such as ABA, a plant regulator that inhibits seed germination (Marrush et al., 1998; Sariyildiz et al., 2005; Nambara et al., 2010), and lignin, a structurally protective and hydrophobic compound of the seed coat (Randle and Honma, 1981; Tewksbury et al., 2008b; Nambara et al., 2010).
Wild chile pepper seeds with thick lignified testas become increasingly impermeable to water on drying. This feature is disadvantageous for—if not absent from—most domesticated crop seeds, not only because these seeds germinate slowly, but also because they may require prolonged soaking to remove inhibitors from the seed coat (Randle and Honma, 1981; Pickersgill, 2007; Carlo and Tewksbury, 2014). Therefore, domesticated chile peppers generally have thinner testae than their wild progenitors.
Changes in Organ Size and Quantity
As part of the domestication syndrome, changes in secondary metabolite content may be correlated with other physical and chemical traits, such as nutrient content, size, or biomass (Chen et al., 2015). Compared to most domesticated landraces, wild Capsicum species exhibit smaller leaves, flowers, fruits and seeds, but a larger number of these organs per plant (Pickersgill, 2007). These characteristics—such small but numerous leaves and seeds—confer adaptability, stress reduction, survivability, and bet-hedging strategies to wild chile peppers for the production and dispersal of their seeds in Neotropical habitats (Tewksbury et al., 2008b).
Increased Morphological Variation
According to Chen et al. (2015), morphological changes arising from domestication can disrupt plant-herbivore-natural enemy interactions, however domesticated chile landraces now exhibit enormous inter-varietal and some intra-varietal heterogeneity in morphological traits.
This factor also is especially marked in the parts of the chile pepper plant used by Mesoamerican cultures. While domesticated chile peppers vary greatly in fruit size and shape, and to a lesser extent in color, wild C. annuum var. glabriusculum populations show little morphological variation in fruit size, shape, and color. In certain coastal Neotropical habitats, chile pepper fruits are selected for particular colors and shapes, said to be the best for seasoning turtle meat, while others, of different color and shape, are known as perfume peppers because they have a fragrant aroma as well as pungency. Pickersgill (2007) and Boster (1985) suggest that such traits result from cultural “selection for perceptual distinctiveness.”
In short, the different landraces of chile peppers grown and consumed across Mesoamerica display an astounding range of morphological variation in plant architecture and fruit shape, as well as in fruit color, pungency, and particular cultural uses (Bosland and Votava, 2000). All SMs in Capsicum species, including carotenoids, flavonoids, capsaicinoids, and ascorbic acid, are to some extent, linked with these morphological traits. Boster (1985) has deftly summarized the many references documenting the pronounced differences in morphology between wild and domesticated peppers.
Changes in Plant Habit Related to Resource Partitioning
Selection for increased harvest index (ratio of harvested to total biomass produced per plant) may result in reduced or suppressed lateral branching (Pickersgill, 2007). Reduced number of inflorescences per plant and producing more synchronous fruit ripening on an individual plant and within a stand, facilitating harvesting of the stand as a whole. Fewer nodes and shorter internodes, greater synchronization of maturation of vegetative branches and fruit ripening is also favored by a determinate habit.
The transition from the perennial indeterminate habit of wild chile peppers to the annualized compact habit of domesticated peppers has been triggered by selection for earliness, larger fruits, compact growth/reduced branching with reduced number of fruits per plant, and more synchronous fruit ripening. Loss of perennial plant habit may be the final/accumulated result of human selection for non-dormant seed, which probably modified fruit and seed morphologies, and SM potencies.
Changes in Reproduction
In Capsicum species, floral phenology and pollination, as well as fruit and seed development are influenced by different SMs. For example, carotenoid and flavonoid derivatives are secondary metabolites in the flower that attract pollinators. Similarly, fruit and seed dispersal are mediated by SMs which serve to attract seed dispersers. Simultaneously, fruit and seed protection is mediated by particular SMs (capsaicinoids and phenolics) that repel predators of fruits and seeds.
Wild C. annuum is an autogamous plant with protaginous flowers (exerted stigmas) and high rates of outcrossing by insect pollinators, and indeterminate growth in neotropical Mesoamerica. Flower initiation is late, but once initiated is persistent and very prolific, with overlapping stages of flower and fruit development over the season. Fully domesticated C. annuum land races can also be autogamous, but exhibit much lower rates of outcrossing, probably due to more synchrony in anther and stigma maturation. Most of the fully domesticated chile pepper land races exhibit determinate growth under cultivation, with more rapid onset of flower initiation, fruit development and ripening. For such reasons, fruit and seed production of fully domesticated chile landraces would be almost impossible under natural wild environments in the Neotropics.
Loss of Chemical or Physical Protection Against Biotic and Abiotic Stresses
Many other domesticated crops have partially or completely lost the SMs that protect their wild relatives against predators (herbivores, plant pests and pathogens), and abiotic stresses (drought, salinity, heat, frost, daming radiation, etc.). However, this trend does not necessarily hold true for most domesticated C. annuum land races. Capsaicinoids and other SMs are synthesized in the placental tissue of domesticated chile fruits after flowering as part of fruit development. In other words, in domesticated chiles, SMs may play a small role in chemical defense of plant tissues before fruit and seed development (Meyer et al., 2012; Fernández-Marín et al., 2014).
Protection of wild chile pepper fruits in populations against predators is mostly conferred by capsaicinoids, although flavonoids and phenolics may also play protective roles against predators. However, protection against hervibory in wild chile plants (prior to their flowering) is also facilitated by the “prey refugia” offered by the dense thorny canopies of certain nurse plants. Where they lack nurse plant protection in Mesomerican milpas, domesticated chile peppers must rely on farmers themselves to evict (or to reduce the damage potentially wreaked by) mammalian predators and browsers (Pickersgill, 2007; Gepts, 2010; Padilha and Barbieri, 2016).
With regard to protection against abiotic stresses, wild chile pepper plants employ SMs such as flavonoids, phenolics and vitamin C for protection against drought, heat and daming radiation. In particular, carotenoid derivatives confer protection against plant cell oxidative reactions caused by lethal radiation, such as direct sunlight and UV light (Wahyuni et al., 2013).
Fully domesticated C. annuum landraces express widely varying concentrations of capsaicinoids compared to pungency levels in wild populations. Today, the mildest to most pungent domesticated chiles vary in the capsaicin and pungency content (~5,000–300,000 SHU); with most (but not all) wild populations being in the medium-to-high range (~100,000 SHU) of pungency (Eich, 2008). The hottest chile peppers belong to C. chinense and currently there are some cultivars of this species such as “Bhut Jolokia” and “Trinidad Scorpion” which have around 1.0 million SHU (Bosland and Baral, 2007), and “Carolina Reaper,” the hottest pepper in the world exceeding 1.5 million SHU (Padilha and Barbieri, 2016). Domesticated landraces of C. annuum may also have larger but more variable amounts of other SMs, including more antioxidant capacity (Wahyuni et al., 2011).
Agroecological Context of Milpa Cultivation as a Selective Pressure
Lack of both seed dormancy and a facultatively perennial plant habit probably enabled the shift from avian dispersal of fruits under nurse plant canopies in the wild to open cultivation of annual plants with non-dormant seeds in milpa agro-ecosystems. The loss of ecological interactions with birds and nurse plants due to intentional seed-saving and dispersal by humans must have generated incidental changes in SMs. Shifting the patterns of SMs through such selection could explain, in part, the emergence of new chemotypes, genotypes and morphotype landraces under cultivation in milpas within the Neotropics. The Mesoamerican milpa agroecosystem may have gradually replaced the nurse plants in agroforestry systems during the early domestication of C. annuum, but as it did, it likely accelerated unconscious selection away from wild chemotypes and morphotypes.
Synthesis of Coevolutionary Shifts Occurring With Domestication
We suggest that incipient cultivation and “re-balancing” selection of seed germinability in polymorphic founder populations of C. annuum var. glabrisculum in Mesoamerica around 6500 BP rapidly led to changes in gene frequencies associated with other adaptive traits. Curiously, this is roughly the time period when a new meme –a chile-processing technology and associated culinary techniques–first became evident in the prehistoric cultures of south-central Mexico. This technology was called mollicaxtli in Nahuatl (now molcajete today in Spanish, and consists of a round three-legged, grinding bowl and pestle for crushing dried spices, made out of fired clay or volcanic stone (Vela, 2009).
The molcajete's sudden emergence and wide diffusion suggests that domesticated chile pepper were not merely being eaten fresh, but surplus harvests were being dried and stored between growing seasons for use as a dried spice, condiment, medicine or vermifuge. Undoubtedly, these multiple uses of small, dried chile “pods” emerged long before the selection for larger fleshier fruits, which could be used as a vegetable that was stuffed with meats, fruits or other spices. Thus, a new technology (molcajetes) and its associated culinary uses, as well as seed saving and trade beyond their ancestral habitats may have accelerated selection for a wider range of Neotropical habitats and overall diversification of domesticated chile pepper landraces.
Most remarkably, chile pepper fruits of some cultivated landraces are many times hotter or milder than those of wild populations, suggesting that domestication has not only diversified, but shifted total pungency in both directions—to higher “heat levels” in some varieties (e.g., ghost peppers), and to lesser levels in nearly non-pungent varieties (e.g., bell peppers). There is limited evidence that the mixes of capsaicinoids found in cultivated chile varieties are also more variable than those in wild populations, but comparable sampling has been poor. Neverthless, we see evidence for both (H2)—a diversification of the levels of potency—and (H3)—an intensification of potency of selected SMs with chile pepper domestication.
In the case of milder (less pungent) chile peppers, we assume that farmers' protection of the plants compensates to some extent for lower levels of chemical defenses. Haak et al. (2012) have confirmed tradeoffs between expression of capsaicinoid pungency, and yield under water-stressed conditions. While capsaicinoids remain the most important plant chemical defenses in most domesticated chiles as they are in wild peppers, the roles of other secondary metabolites found in lower concentrations should not be dismissed.
Mesoamerican Human/Chile Pepper Coevolution in Relation to Benefits of Chemical Defenses
According to paleobiolinguistic reconstructions of the presumed origins and diffusion of domesticated chile peppers in Mesoamerica, the oldest reconstructed term for cultivated chiles is found in proto-Otomanguean from south-central Mexico, estimated to be in transcultural circulation by 6592 B.P. (Brown et al., 2013; Kraft et al., 2014). This evidence is supported by archeological analyses that confirm the presence of domesticated chile fruit and spice-grinding molcajetes at sites along the Sierra Madre Oriental/Trans-Volcanic by 6000 years ago, especially in seasonally dry subtropical thornscrub (Kraft et al., 2014).
Nevertheless, several lines of research agree that the origin of the domesticated C. annuum landraces may have also occurred elsewhere within the broader Mesoamerican region (Eshbaugh, 1970; Hernández-Verdugo et al., 2001a; Perry and Flannery, 2007; Pickersgill, 2007; Aguilar-Meléndez et al., 2009). In other words, the precise location or locations of domestication of C. annuum in Mesoamerica still remains unknown.
Based on linguistic analyses, Brown (2010) suggests that the earliest plant management in Mesoamerica was of grain, succulent and oil crops; they became cultivated as staples no later than 7000 years ago. The earliest cultivation of spices (including chiles) for seasoning these staples came centuries later.
In short, staples such as maize, maguey, nopal and avocado were probably cultivated to provide seasonal surpluses for storage and consumption at least a thousand years before the earliest detectable onset of chile pepper cultivation as a spice, anthelmentic medicine, vermifuge or condiment (but most likely not as a fresh green vegetable).
The pervasiveness of the use of chile peppers in treating illnesses in Mesoamerica and Aridoamerica (N Mexico and SW USA) is without peer, among any of the other crops domesticated in these regions. This fact alone suggests that the culinary uses of Capsicum were not the only catalysts to domestication. Table S2 shows several ancient medicinal uses derived from extensive studies of indigenous farming cultures in Mesoamerica. Collectively, this information suggests that a “Mesoamerican intellectual tradition” of indigenous medicinal-culinary knowledge (López Austin, 2001; Good, 2005) may have guided the selection of SMs and other traits in chile pepper landraces. The very cultural persistence of chile plants (as well as maize, etc.) within milpas and dooryard gardens in this modern globalized world, is clear evidence that ancestral cultural traditions spanning 6000–7000 years, still have adaptive value today.
In addition, the milpa management traditions have been culturally maintained to keep alive what is culturally perceived as a sacred agroecosystem that maintains and regenerates everyday life, community values and collective identities among many Mesoamerican societies (Bonfil-Batalla, 2012; Good, 2015). The medicinal, ceremonial and culinary uses of chile peppers by over 60 native cultures in Mesoamerica are embedded a small but inseparable and integral part of a broader cosmovision, one that persists up through this present moment (Alcorn, 1984; Long-Solís, 1986; López Austin, 2001; de Avila, 2008). Any true EES that attempts to use chile pepper domestication as a model system must inevitably take these cultural memes into account.
There is no reason to assume that chiles were first gathered, then cultivated, for a single use, given that tobacco, cacao and other early crops also had multiple uses. However, as staple crops grew in yields and diets became more redundant, chile peppers may have played critically-important roles in protecting grains and legumes aggregated in storage facilities from post-harvest consumption by insect pests and fungi common in the Neotropics. Some of these same chemical defenses in chile peppers may have protected humans who were aggregated into increasingly dense habitations from intestinal parasites, and from body lice or fleas. Finally, the SMs in chile peppers may also have become increasingly necessary elements of the traditional diets and pharmacopeia as “nutraceuticals” that counteracted the greater redundancy in agricultural diets.
The pharmacological utility of SMs in chile peppers is not restricted to the control of fleas, lice and intestinal microbes. They have recently been demonstrated to be effective in reducing intestinal infections by aquatic helminthes of the same group as the intestinal worms that cause ill health and sluggishness among one third of the world's population, especially children in tropical climes (Mostafa-Kamal et al., 2015). This is a clear example of how plant chemical defenses have proven efficacy for “defending” human health against various biotic stresses among those who consume the same plant as a food, a medicine or both (Mostafa-Kamal et al., 2015).
In Table S2, we wish to underscore the myriad medicinal uses retrieved from historical documents that persist to this day in Mesoamerican intellectual traditions. Out of 47 ailments to which chile peppers were applied, 24 of these were recorded among Maya communities. In 2000, fieldwork in Yucatecan Mayan communities documented the persistence of medicinal uses of at least seven different types of chiles (Aguilar-Meléndez and Lira-Noriega, 2018), suggesting that the diversification of chile peppers may continue to generate direct benefits to human health.
Conclusions
In this paper three hypothesis were evaluated and discussed:
(H1) A reduction and simplification of the potency of plant chemical defenses against seed predators, foliage herbivores and disease microbes with greater reliance on human intervention to protect the plants. This assumes that fully domesticated modern and commercial varieties of peppers under intense monoculture are more susceptible to predators (insect pests and diseases), than their wild progenitors, because they produce less number and concentration of SM in fruits, seeds, and leaves.
(H2) A diversification of the levels of potency and mixes of defense chemicals, given the wider range of habitats and broader geography to which the crop plants are exposed. This assumes that different C. annuum landraces in different agroecosystems produce variable amounts and types of SM.
(H3) An intensification of the potency of certain plant chemical defenses, given the need to protect the plants in agro-habitats where they occur at higher density and without as much beta diversity of neighboring plant species to slow the spread of predators, herbivores, competing weeds or diseases. This assumes that some domesticated landraces and modern varieties produce larger concentrations of valued SMs (capsaicinoids and carotenoids) under intense monoculture, compared to their wild progenitors.
Of these three hypotheses, we see more evidence supporting both H2 and H3, with respect to the diversification and heightening of pungency through chile pepper domestication. H2– the diversification of levels in SMs under domestication– seems to fit with the mechanism of “balancing selection,” in the sense of maintaining polymorphisms in Mesoamerican chile pepper landraces. The H3 trend has mostly been in more recently advanced cultivars of chile peppers outside their area of Neotropical origins. The H1 trend toward a reduction in pungency and other SMs such as phenolics and carotenoids in fruits and other organs is most evident in the recently advanced “bell pepper” group of chile landraces and cultivars, which are also most popular outside of the Neotropics. There is no question that sweet bell pepper cultivars of C. annuum must rely on human protection to survive against different predators that may prey on roots, leaves, fruits, and seeds. While birds may damage bell peppers grown in temperate climates outside of the Neotropics, they are virtually ineffective in dispersing the fruit (or most seeds within the fruit) to safe sites for germination and recruitment.
We conclude that contrary to trends in other crops, domestication has not necessarily reduced potency or homogenized the levels of chemical defenses—or at least of capsaicinoids—in chile pepper fruits. It has diversified capsaicinoid potency levels among and across domesticated varieties, compared to those found in most wild chile peppers. However, scientists still lack sufficient evidence to conclude that such diversification has occurred in any other SMs involved in chile pepper plant defense.
The likely diversification of SM production and/or concentration in domesticated C. annuum is the result of differential human selection of different allelic combinations—including selection of many recessive genes, under different environments and managed ecosystems—that are only rarely expressed in truly wild populations (Haak et al., 2014).
Higher concentrations of pungent compounds such as capsaicin may confer better adaptation and fitness to chile pepper crops under novel environments. These highly pungent varieties are now finding new uses in pharmacological and culinary uses, but the majority of the world's human inhabitants continues to directly use wild or domesticated landraces of chile peppers medicinally and gastronomically as they have for centuries.
There is plausible evidence from diverse cultures in Mexico that the SMs expressed in C. annuum fruits have been efficacious in reducing human diseases as well as infestations of internal and external parasites. This may in part explain why so many of the distinctive medicinal uses of chiles persist in nearly every Mesoamerican and Aridoamerican culture today. The nutritional and medicinal benefits of chiles may initially appear diffuse or minor to evolutionary ecologists, but their collective benefits as perceived by their “co-evolved” Mesoamerican cultivators, curanderas, cooks and consumers are impressive.
The extraordinary potency and the current intensity of gastronomic and pharmacological uses of chile peppers (Bosland and Votava, 2000) suggest that chile peppers should no longer be relegated the status of a “minor crop” as standard economic botany references and global agricultural statistics have done in the past. By 2010, global production of domesticated Capsicum fruits had reached 1.8 million ha, with more than 29 million metric tons annually harvested (Wahyuni et al., 2013). Their production continues to expand, while their culinary as well as medicinal and pest-repellent uses continue to diversify.
We should acknowledge that the current efficacy and economic significance of chile peppers' secondary metabolities in our diets and pharmocopieas is not merely due to the historic inventiveness of and mutualistic interactions with our own kind. It has benefited from the selective pressures by fungi, hemipteran insects, nematodes and rodents, as well as the directed dispersal of chile seeds by numerous bird species in the Neotropics. As such, there remains much to be learned by further advancing analyses of chile domestication to serve as a model for extended evolutionary synthesis.
Author Contributions
JL-R: designed research and wrote the paper; GN: designed research and wrote the paper; AA-M: wrote the paper.
Funding
Funding for this publication comes in part from Programa de Fortalecimiento a la Calidad Educativa (PFCE) of the Universidad Autónoma de Aguascalientes, México.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer, DP, and handling Editor declared their shared affiliation.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2018.00048/full#supplementary-material
References
Aguilar-Meléndez, A., and Lira-Noriega, E. (2018). ”Dónde crecen los chiles en México?” in Los Chiles que le dan Sabor al Mundo. Contribuciones Multidisciplinarias, eds A. Aguilar-Meléndez, M. A. Vásquez Dávila, E. Katz, and M. R. Hernández Colorado (Xalapa: Universidad Veracruzana and IRD), 61–79.
Aguilar-Meléndez, A., Morrell, P. L., Roose, M. L., and Kim, S. C. (2009). Genetic diversity and structure in semiwild and domesticated chiles (Capsicum annuum; Solanaceae) from Mexico. Am. J. Bot. 96, 1190–1202. doi: 10.3732/ajb.0800155
Benrey, B., Callejas, A., Rios, L., Oyama, K., and Denno, R. F. (1998). The effects of domestication of brassica and phaseolus on the interaction between phytophagous insects and parasitoids. Biol. Control 11, 130–140. doi: 10.1006/bcon.1997.0590
Bosland, P. W., and Baral, J. B. (2007). “Bhut Jolokia”—The world's hottest known chile pepper is a putative naturally occurring interspecific hybrid. HortScience 42, 222–224.
Bosland, P. W., and Votava, E. J. (2000). Peppers: Vegetable and Spice Capsicums, Vol. 22. Wallingford, UK: CABI.
Boster, J. S. (1985). Selection for perceptual distinctiveness: evidence from aguaruna cultivars of Manihot esculenta. Econ. Bot. 39, 310–325. doi: 10.1007/BF02858802
Brown, C. H. (2010). “Development of agriculture in prehistoric mesoamerica: the linguistic evidence,” in Pre-Columbian Foodways: Interdisciplinary Approaches to Food, Culture, and Markets in Ancient Mesoamerica, eds J. Staller and M. Carrasco (New York, NY: Springer), 71–107.
Brown, C. H., Clement, C. R., Epps, P., Luedeling, E., and Wichmann, S. (2013). The paleobiolinguistics of domesticated chili pepper (Capsicum spp.). Ethnobiol. Lett. 4, 1–11. doi: 10.14237/ebl.4.2013.2
Carlo, T. A., and Tewksbury, J. J. (2014). Directness and tempo of avian seed dispersal increases emergence of wild chiltepins in desert grasslands. J. Ecol. 102, 248–255. doi: 10.1111/1365-2745.12180
Carrizo-García, C., Barfuss, M. H. J., Sehr, E. M., Barboza, G. E., Samuel, R., Moscone, E. A., et al. (2016). Phylogenetic relationships, diversification and expansion of chili peppers (Capsicum, Solanaceae). Ann. Bot. 118, 35–51. doi: 10.1093/aob/mcw079
Casas, A., Parra, F., Rangel, S., Guillén, S., Blancas, J., and Figueredo, C. J. (2015). “Evolutionary ecology and ethnobiology,” in Evolutionary Ethnobiology, eds U. P. Albuquerque, P. Muniz de Madeiros, and A. Casas (Cham: Springer), 37–57.
Chen, Y. H., Gols, R., and Benrey, B. (2015). Crop domestication and its impact on naturally selected trophic interactions. Annu. Rev. Entomol. 60, 35–58. doi: 10.1146/annurev-ento-010814-020601
Dávila-Flores, A. M., DeWitt, T. J., and Bernal, J. S. (2013). Facilitated by nature and agriculture: performance of a specialist herbivore improves with host-plant life history evolution, domestication, and breeding. Oecologia 173, 1425–1437. doi: 10.1007/s00442-013-2728-2
de Avila, B. A. (2008). “La diversidad lingüística y el conocimiento etnobiológico,” in Capital Natural de México. Vol. I: Conocimiento Actual de la Biodiversidad, ed J. Sarukhán (México, DF: CONABIO), 497–556.
Do Rêgo, E. R., Finger, F. L., and do Rêgo, M. M. (2012). “Consumption of pepper in Brazil and its implications on nutrition and health of humans and animals,” in Pepper: Nutrition, Consumption and Health, Vol. 1, eds M. A. Salazar and J. M. Ortega (New York, NY: Nova Science), 159–170.
Eich, E. (2008). Solanaceae and Convolvulaceae: Secondary Metabolites: Biosynthesis, Chemotaxonomy, Biological and Economic Significance (A Handbook). Berlin, Heidelberg: Springer.
Eshbaugh, W. H. (1970). A biosystematic and evolutionary study of Capsicum baccatum (Solanaceae). Brittonia 22, 31–43. doi: 10.2307/2805720
Fernández-Marín, B., Milla, R., Martín-Robles, N., Arc, E., Kranner, I., Becerril, J. M., et al. (2014). Side-effects of domestication: cultivated legume seeds contain similar tocopherols and fatty acids but less carotenoids than their wild counterparts. BMC Plant Biol. 14:1599. doi: 10.1186/s12870-014-0385-1
Freeman, B., and Beattie, G. (2008). An overview of plant defenses against pathogens and herbivores. Plant Health Instr. doi: 10.1094/PHI-I-2008-0226-01
Gepts, P. (2010). Crop domestication as a long-term selection experiment. Plant Breed. Rev. 24, 1–44. doi: 10.1002/9780470650288.ch1
Good, E. C. (2005). Ejes conceptuales entre los nahuas de Guerrero: expresión de un modelo fenomenológico mesoamericano. Estud. Cult. Náhuatl 36, 87–113.
Good, E. C. (2015). “Las cosmovisiones, la historia y la tradición intelectual en Mesoamérica,” in Cosmovisión Mesoamericana. Reflexiones, polémicas y etnografías. Fideicomiso historia de las Americas. (México, DF: Serie Ensayos), 139–160.
Haak, D. C., Kostyun, J. L., and Moyle, L. C. (2014). “Merging ecology and genomics to dissect diversity in wild tomatoes and their relatives,” in Ecological Genomics: Ecology and the Evolution of Genes and Genomes, eds C. R. Landry and N. Aubin-Horth (Dordrecht: Springer), 273–298.
Haak, D. C., McGinnis, L. A., Levey, D. J., and Tewksbury, J. J. (2012). Why are not all chilies hot? A trade-off limits pungency. Proc. R. Soc. B Biol. Sci. 279, 2012–2017. doi: 10.1098/rspb.2011.2091
Hernández-Verdugo, S., Guevara-González, R. G., Rivera-Bustamante, R. F., and Oyama, K. (2001b). Screening wild plants of Capsicum annuum for resistance to pepper huasteco virus (PHV): presence of viral DNA and differentiation among populations. Euphytica 122, 31–36. doi: 10.1023/A:1012624830340
Hernández-Verdugo, S., Luna-Reyes, R., and Oyama, K. (2001a). Genetic structure and differentiation of wild and domesticated populations of Capsicum annuum (Solanaceae) from Mexico. Plant Syst. Evol. 226, 129–142. doi: 10.1007/s006060170061
Holt, J., and Birch, N. (1984). Taxonomy, evolution and domestication of Vicia in relation to aphid resistance. Ann. Appl. Biol. 105, 547–556. doi: 10.1111/j.1744-7348.1984.tb03081.x
Hunziker, A. T. (2001). Genera Solanacearum: The Genera of Solanaceae Illustrated, Arranged According to a New System. Ruggell: A.R.G. Gantner.
Johns, T. (1990). With Bitter Herbs They Shall Eat it: Chemical Ecology and The origins of Human Diet and Medicine. Tucson: University of Arizona Press.
Johns, T., and Kubo, I. (1988). A survey of traditional methods employed for the detoxification of plant foods. J. Ethnobiol. 8, 81–129
Kim, W.-R., Kim, E. O., Kang, K., Oidovsambuu, S., Jung, S. H., Kim, B. S., et al. (2014). Antioxidant activity of phenolics in leaves of three red pepper (Capsicum annuum) cultivars. J. Agric. Food Chem. 62, 850–859. doi: 10.1021/jf403006c
Kliebenstein, D. J. (2004). Secondary metabolites and plant/environment interactions: a view through Arabidopsis thaliana tinged glasses. Plant Cell Environ. 27, 675–684. doi: 10.1111/j.1365-3040.2004.01180.x
Kraft, K. H., Brown, C. H., Nabhan, G. P., Luedeling, E., Ruiz, J., de J. L., d'Eeckenbrugge, G. C., et al. (2014). Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc. Natl. Acad. Sci. U.S.A. 111, 6165–6170. doi: 10.1073/pnas.1308933111
Long-Solís, J. (1986). Capsicum y Cultura: la Historia del Chilli. México: Fondo de Cultura Económica.
López Austin, A. (2001). “El núcleo duro, la cosmovisión y la tradición mesoamericana,”, in Cosmovisión, Ritual e Identidad de los Pueblos Indígenas de México, eds J. Broda and F. Báez-Jorge (México: CONACULTA y Fondo de Cultura Económica), 47–65.
Marrush, M., Yamaguchi, M., and Saltveit, M. E. (1998). Effect of potassium nutrition during bell pepper seed development on vivipary and endogenous levels of abscisic acid (ABA). J. Am. Soc. Horticult. Sci. 123, 925–930.
Meyer, R. S., DuVal, A. E., and Jensen, H. R. (2012). Patterns and processes in crop domestication: an historical review and quantitative analysis of 203 global food crops. New Phytol. 196, 29–48. doi: 10.1111/j.1469-8137.2012.04253.x
Mostafa-Kamal, A., Chowdhury, K. A. A., Moazzam, M., Chy, H., Shill, L. K., Chowdhury, S., et al. (2015). Evaluation of anthelmintic activity of seeds of Sesamum indicum L. and fruits of Capsicum frutescens L. J. Pharmacogn. Phytochem. 3, 256–259.
Nabhan, G. P. (2004). Why Some Like it Hot: Food, Genes, and Cultural Diversity. Washington, DC: Island Press.
Nambara, E., Okamoto, M., Tatematsu, K., Yano, R., Seo, M., and Kamiya, Y. (2010). Abscisic acid and the control of seed dormancy and germination. Seed Sci. Res. 20, 55–67. doi: 10.1017/S0960258510000012
Padilha, H. K. M., and Barbieri, R. L. (2016). Plant Breeding of Chili Peppers (Capsicum, Solanaceae)-A Review. Embrapa Clima Temperado-Artigo em periódico indexado (ALICE). Availabele online at: https://www.alice.cnptia.embrapa.br/bitstream/doc/1060246/1/RosaLiaartigo2016plantbreedingofchilipepperareview.pdf
Pegard, A., Brizzard, G., Fazari, A., Soucaze, O., Abad, P., and Djian-Caporalino, C. (2005). Histological characterization of resistance to different root-knot nematode species related to phenolics accumulation in Capsicum annuum. Phytopathology 95, 158–165. doi: 10.1094/PHYTO-95-0158
Perry, L., and Flannery, K. V. (2007). Precolumbian use of chili peppers in the Valley of Oaxaca, Mexico. Proc. Natl. Acad. Sci. U.S.A. 104, 11905–11909. doi: 10.1073/pnas.0704936104
Pickersgill, B. (2007). Domestication of plants in the Americas: insights from Mendelian and molecular genetics. Ann. Bot. 100, 925–940. doi: 10.1093/aob/mcm193
Randle, W. M., and Honma, S. (1981). Dormancy in peppers [Capsicum annuum]. Sci. Hortic. 14, 19–25. doi: 10.1016/0304-4238(81)90074-1
Retes-Manjarrez, J. E. (2016). Detección de resistencia al virus huasteco vena amarilla del chile y su heredabilidad en genotipos silvestres de Capsicum annuum L. Interciencia 41, 541–547.
Rindos, D. (1984). The Origins of Agriculture: An Evolutionary Perspective. San Diego, CA: Academic Press.
Rodriguez-Saona, C., Vorsa, N., Singh, A. P., Johnson-Cicalese, J., Szendrei, Z., Mescher, M. C., et al. (2011). Tracing the history of plant traits under domestication in cranberries: potential consequences on anti-herbivore defences. J. Exp. Bot.. 62, 2633–2644. doi: 10.1093/jxb/erq466
Rozin, P., and Schiller, D. (1980). The nature and acquisition of a preference for chili pepper by humans. Motiv. Emot. 4, 77–101. doi: 10.1007/BF00995932
Sariyildiz, Z., Demir, I., and Halloran, N. (2005). Germination at stress temperatures and ABA content of pepper (Capsicum annuum L.) seeds in relation to seed development. Eur. J. Horticult. Sci. 70, 67–70.
Tewksbury, J. J., Levey, D. J., Huizinga, M., Haak, D. C., and Traveset, A. (2008a). Costs and benefits of capsaicin-mediated control of gut retention in dispersers of wild chilies. Ecology 89, 107–117. doi: 10.1890/07-0445.1
Tewksbury, J. J., and Nabhan, G. P. (2001). Seed dispersal: directed deterrence by capsaicin in chillies. Nature 412, 403-404. doi: 10.1038/35086653
Tewksbury, J. J., Reagan, K. M., Machnicki, N. J., Carlo, T. A., Haak, D. C., Pe-aloza, A. L. C., et al. (2008b). Evolutionary ecology of pungency in wild chilies. Proc. Natl. Acad. Sci. U.S.A. 105, 11808–11811. doi: 10.1073/pnas.0802691105
Wahyuni, Y., Ballester, A. R., Sudarmonowati, E., Bino, R. J., and Bovy, A. G. (2011). Metabolite biodiversity in pepper (Capsicum) fruits of thirty-two diverse accessions: variation in health-related compounds and implications for breeding. Phytochemistry 72, 1358–1370. doi: 10.1016/j.phytochem.2011.03.016
Wahyuni, Y., Ballester, A.-R., Sudarmonowati, E., Bino, R. J., and Bovy, A. G. (2013). Secondary metabolites of Capsicum species and their importance in the human diet. J. Nat. Prod. 76, 783–793. doi: 10.1021/np300898z
Wink, M. (2003). Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 64, 3–19. doi: 10.1016/S0031-9422(03)00300-5
Keywords: Capsicum annuum, plant domestication, secondary metabolites, plant chemical defenses, Neotropics, Mesoamerica
Citation: Luna-Ruiz JdJ, Nabhan GP and Aguilar-Meléndez A (2018) Shifts in Plant Chemical Defenses of Chile Pepper (Capsicum annuum L.) Due to Domestication in Mesoamerica. Front. Ecol. Evol. 6:48. doi: 10.3389/fevo.2018.00048
Received: 31 August 2017; Accepted: 05 April 2018;
Published: 24 April 2018.
Edited by:
Alejandro Casas, Instituto de Investigaciones en Ecosistemas y Sustentabilidad, Universidad Nacional Autónoma de México, MexicoReviewed by:
Daniel Pinero, Universidad Nacional Autónoma de México, MexicoRosa Lia Barbieri, Embrapa Clima Temperado, Brazil
Copyright © 2018 Luna-Ruiz, Nabhan and Aguilar-Meléndez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jose de Jesus Luna-Ruiz, am9zZWx1bmFydWl6MTFAeWFob28uY29tLm14