 Kevin C. Fraser1*
Kevin C. Fraser1* Kimberley T. A. Davies2
Kimberley T. A. Davies2 Christina M. Davy3,4
Christina M. Davy3,4 Adam T. Ford5
Adam T. Ford5 D. T. Tyler Flockhart6
D. T. Tyler Flockhart6 Eduardo G. Martins7
Eduardo G. Martins7- 1Department of Biological Sciences, University of Manitoba, Winnipeg, MB, Canada
- 2Department of Oceanography, Dalhousie University, Halifax, NS, Canada
- 3Wildlife Research and Monitoring Section, Ontario Ministry of Natural Resources, Peterborough, ON, Canada
- 4Environmental and Life Sciences Graduate Program, Trent University, Peterborough, ON, Canada
- 5Department of Biology, The University of British Columbia - Okanagan Campus, Kelowna, BC, Canada
- 6Appalachian Laboratory, University of Maryland Center for Environmental Science, Frostburg, MD, United States
- 7Ecosystem Science and Management Program, University of Northern British Columbia, Prince George, BC, Canada
From butterflies to elephants, the rapidly developing science of movement ecology is providing increasingly detailed spatio-temporal data on a wide array of mobile animals. Thus, this discipline also holds great promise for improving the conservation of wildlife. To measure progress toward this promise, we investigated the degree to which movement ecology research is connected to conservation goals as well as the proportion of studies that were incorporated into federal and international status assessments for mobile species at risk. We examined 13,349 “movement ecology” papers published between 1990 and 2014 and found that explicit connections to conservation and management were made in 35% (n = 4, 672) of these papers, with the number of connections increasing over time. We then measured the uptake of movement ecology research into species status assessment and recovery plans (n = 72 documents) produced by three different governance agencies for 12 endangered mobile species. We found that on average 60% of available movement ecology research was used in the status assessment process, demonstrating that when movement ecology research is available, it is generally being utilized in conservation planning. However, for 25% of these species, there was little movement research available to be used, highlighting that knowledge gaps remain for some at-risk species despite the general growth of movement ecology research. We outline opportunities for movement ecology to promote more effective conservation of taxa that move.
Introduction
Humans have tracked the movements of animals for millennia—first for our survival, and more recently to better understand, manage, and protect wildlife and fisheries populations. The miniaturization and proliferation of new tracking technology has enabled the rapid growth of research aimed at studying movement across a variety of scales. In this “golden age of bio-logging” (sensu Wilmers et al., 2015) and biotelemetry (Hussey et al., 2015; Cooke et al., 2017; Taylor et al., 2017), accurate and precise estimates of movement are possible even for small and cryptic species such as insects (O'Neal et al., 2004), bats (Castle et al., 2015), and long-distance migrants such as songbirds, salmon, and sea turtles (Godley et al., 2007; McKinnon et al., 2013; Clark et al., 2016), giving rise to the rapidly developing field of movement ecology (Nathan, 2008). At the same time, the global impact of human activities that alter natural habitats is reducing the ability of animals to move (Tucker et al., 2018) and may also be inducing range shifts, thus underlining the critical importance of acquiring baseline data on home ranges and movement patterns.
Improvements in the accuracy and precision of animal tracking have increased knowledge of habitat associations, the extent and use of geographical ranges, migratory pathways, and phenology, as well as interactions between animals and their environment. For example, mule deer (Odocoileus hemionus) are one of the most commonly tagged animals in wildlife studies, yet only recently have researchers determined the details of what turns out to be the longest land-based migration in the conterminous United States (Sawyer et al., 2016). Similarly, (Hu et al., 2016) describe novel techniques to measure the cryptic flows of trillions of migrating insects at high attitude. In addition to helping to better understand how animals navigate and time their movements through space, movement ecology has the potential to inform management and policy decisions with conservation outcomes. By providing increasingly detailed information on the spatio-temporal movements of mobile species (from insects to whales), at the individual and/or population level, movement ecology research can provide data that could be used to inform conservation status assessments, for dynamic management (Allen and Singh, 2016), or to increase protected areas to better encapsulate the range and extent of animal movement (Coristine et al., 2018).
Realizing the promise of movement ecology requires effective integration of movement data at relevant scales to inform conservation policy (Hays et al., 2016). The field of movement ecology seeks to identify the scale at which movement occurs (Allen and Singh, 2016). Scale is often associated with a geography over which an organism may range but the geographic range is only interpretable over some timescale relevant to life history. For example, daily foraging movements can occur within a small breeding territory of less than a hectare relative to seasonal migrations that occur across continents (Runge et al., 2014). Conservation biologists link the scale and structure of movement to demography that ultimately contributes to population dynamics (Sample et al., 2018). Practitioners can then frame policy to prioritize actions (e.g., dynamic site closures across space and time; e.g., Hoop et al., 2015) to reach management and conservation objectives, identify intra-, and trans-boundary conservation partners (López-Hoffman et al., 2009).
Here, we evaluate the conservation promise of movement ecology by addressing three objectives. First, we quantify the degree to which researchers explicitly link their movement ecology research to conservation and management through the use of key words. Second, we quantify the incorporation of the movement ecology literature into species assessments for national and international organizations. Third, we consider future directions for the integration of movement ecology and conservation policy that could promote the effective management and recovery of threatened species.
How Explicitly Is Movement Ecology Research Linked to Conservation Applications?
There is a prevalent assertion that movement ecology research can inform conservation (Barton et al., 2015; Wilmers et al., 2015; Allen and Singh, 2016; Davy et al., 2017). To assess the validity of this claim, we conducted a literature search and found that the term “movement ecology” is mentioned in 13,349 publications between 1990 and 2014 (Web of Science search, Figure 1; see Supplemental Methods 1). Of these, 4,687 papers (35%) also referenced at least one of four key words related to conservation: “conservation,” “management,” “recovery,” or “protection.” Use of these key words increased over time, from inclusion in 4% of papers published in 1990, to inclusion in 44% of papers published in 2014 (Figure 1). However, none of these words appear in a key paper framing the discipline of movement ecology and developing a framework for future research (Nathan, 2008). Clearly, movement ecologists are increasingly seeing the conservation value of their work, so the question then remains: to what extent is this research actually being used by conservation practitioners?
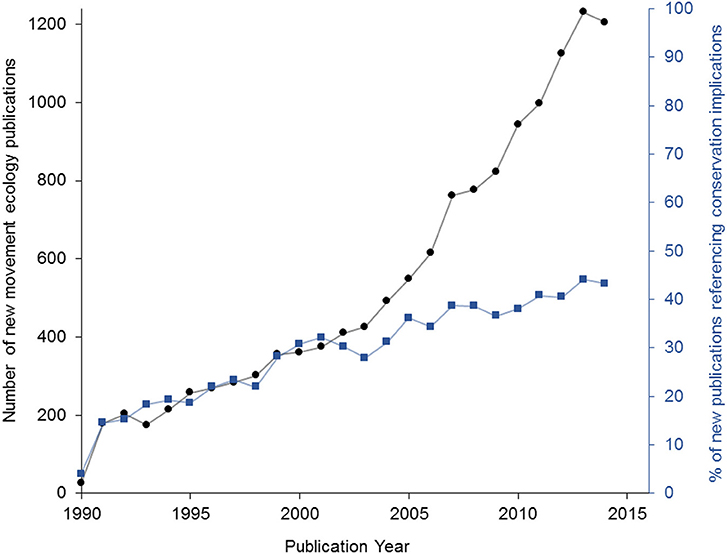
Figure 1. Annual number of peer-reviewed publications addressing movement ecology has increased steadily since 1990 (shown in black; y-axis on left), along with the percentage of those publications that also address species conservation, management, protection, and/or recovery (shown in blue; y-axis on right).
Is Movement Ecology Research Informing Science-Based Policy Processes?
We used a case study approach to investigate how movement ecology is fulfilling its potential to inform policy and conservation decision-making, and to highlight the effects of available research on uptake into conservation-related policy processes. One of the primary mechanisms by which movement ecology can become integrated into the policy process is through species assessment and recovery plans. Such plans typically summarize the available scientific information in peer-reviewed literature and then make recommendations based on that knowledge. In the case of transboundary species, multiple jurisdictions, and agencies may assess the same species. This multi-agency approach provides a level of replication to assess the uptake of movement ecology research and an opportunity to understand how different policy frameworks influence uptake.
Our case study focuses on transboundary species occurring across the Canada-USA border. Between 1985 and 2014, Canadian and US agencies and the International Union for Conservation of Nature (IUCN) each published status assessments for 12 of the same transboundary species, all of which are highly mobile (n = 38 documents, Table 1). Canada and the USA further developed recovery strategies for these 12 species, which in some cases included multiple plans for the same species by the same agency (n = 34 documents). The species list included a range of taxa (fish, birds, mammals, and reptiles) and ecosystems (aerial, terrestrial, and marine). Each of these 12 species faces threats such as habitat loss and fragmentation that can only be effectively mediated if movement data are available (Table 1). We expected that peer-reviewed studies that provided data or knowledge of value to conservation would be included in the assessment process and therefore in the resulting documents. We used the occurrence of movement ecology literature in the bibliographies of species assessment and recovery plans to calculate the uptake of the literature into the policy process.
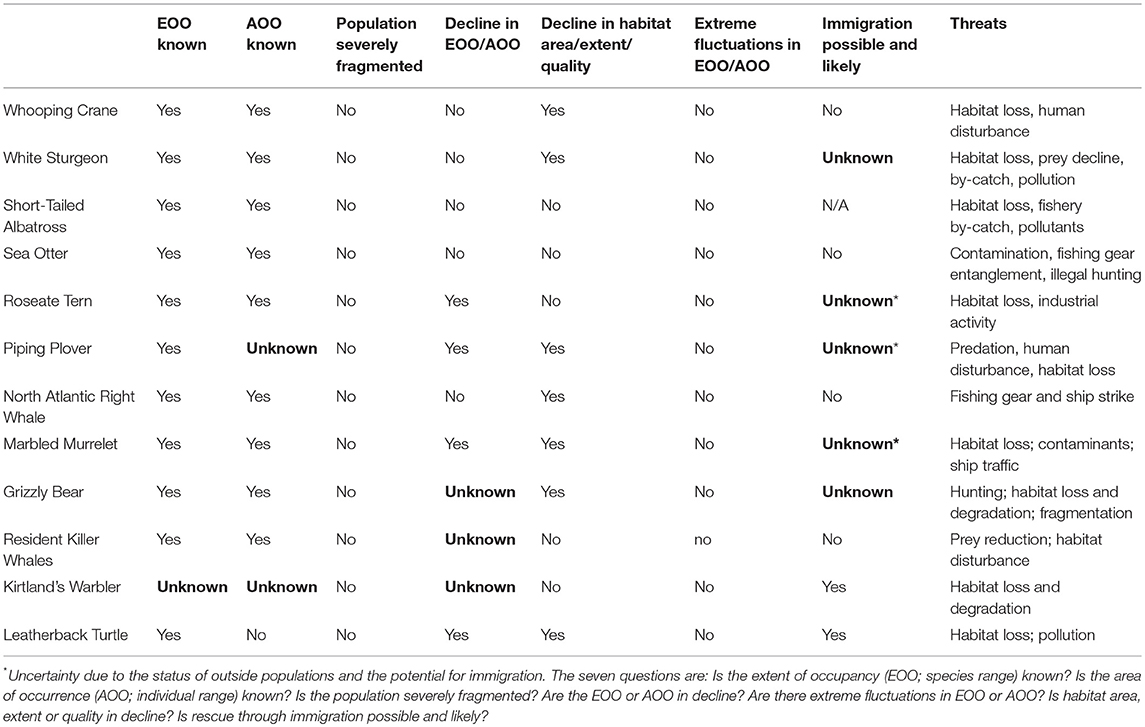
Table 1. Seven movement-related questions asked by federal (Canadian) conservation status assessment processes and level of knowledge (known (yes/no) or unknown) about each question for 12 federally listed species.
We quantified the availability of movement ecology literature for our 12 target species by finding the total number of papers listed on Web of Science up to the end of the year 2014 that included the species name and at least one of a suite of keywords related to movement ecology (see Supplemental Methods 2). We then assessed the proportion of available movement ecology literature used in the bibliographies of each recovery strategy or status assessment. We only included published studies up to the publication date of the latest status assessment or recovery strategy of each agency to calculate availability (Table 1). In the cases where multiple status assessments were published for a species by the same agency/jurisdiction (e.g., the USA published two updates to the original 1987 recovery plans for grizzly bears, Ursus arctos), we used all available assessments in the analysis. This analysis provided two measures for each species: (1) a minimum estimate of the number of movement ecology studies available (i.e., published by the year the assessments and recovery strategies were released), and (2) the proportion of movement ecology papers that were cited in status assessment and recovery strategies.
We found that on average 60% of movement ecology papers per species were used in at least one of the national status assessments, recovery strategies, or IUCN Red List assessments for each of the 12 species we examined (Figure 2). There was no significant difference between the proportion of studies included in the recovery plans and status assessments in Canada and the USA (Two-Way ANOVA, p > 0.05, Figure 3). A higher proportion of movement ecology studies were cited in status assessments from the USA and Canada (pooled mean 0.39 ± SD 0.27) than from the IUCN Red List assessments (0.11 ± 0.11; One-Way ANOVA, n = 12, F = 6.19, p = 0.006, R2 = 0.29, Figure 3). We suggest two potential explanations. First, the IUCN assessments consult fewer studies overall than the federal legislative bodies (e.g., for assessments of the North Atlantic right whales (Eubalaena glacialis), IUCN used 33 references whereas Committee on the Status of Endangered Wildlife in Canada [COSEWIC] status assessment used 158 references). Second, unlike assessment in Canada and the USA, IUCN assessments rely more heavily on changes in population size (under Criterion A: reduction in population size), rather than spatial factors such as range reduction (under Criterion B: geographic range), to which movement ecology is more directly associated.
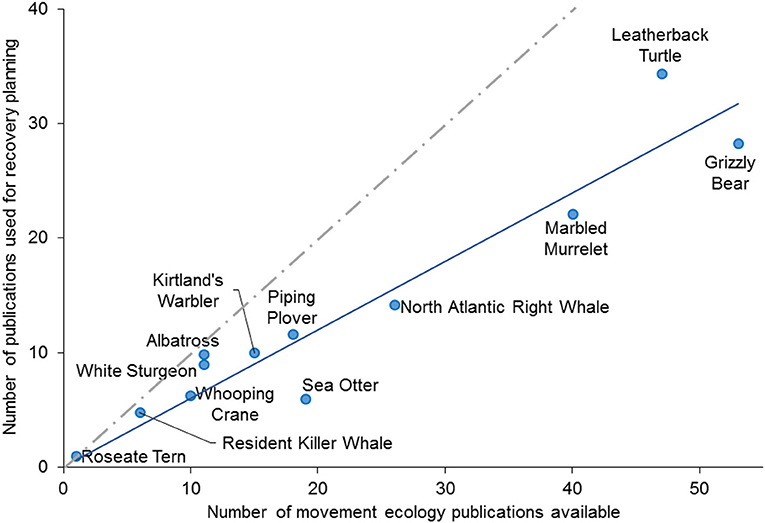
Figure 2. Conservation status assessments and recovery planning for threatened species are informed by only a proportion of the available research on movement ecology. Availability (y-axis) and use (x-axis) of research on movement ecology are strongly correlated (best-fit line, slope = 0.6, r2 = 0.91, p < 0.001) but recovery planning for 11/12 species did not use all available data. Gray dashed line shows a 1:1 correlation for comparison. Figure includes all species occurring in the United States and Canada that have trans-jurisdictional ranges, for which conservation status assessments and recovery plans have been completed, and that have each been assessed under three conservation governance bodies (United States Federal, Canada Federal, IUCN; N = 12).
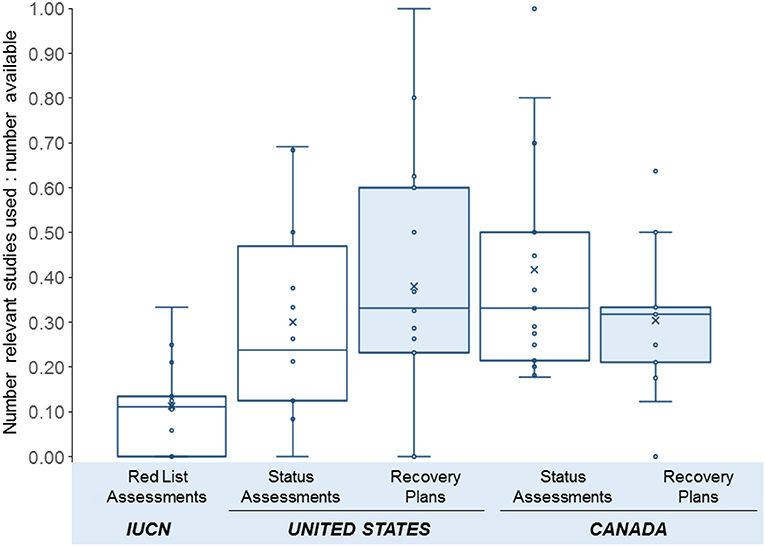
Figure 3. Conservation assessments (open boxes) and recovery plans (shaded boxes) for threatened, transboundary species. Y-axis indicates the proportion of peer-reviewed studies used in conservation assessments to relevant, peer-reviewed studies available, for a subset of 12 species (Figure 2) that have been assessed globally, by the IUCN, federally within the U.S.A., and federally within Canada. Boxes show the first (lower) and third (upper) quartiles; whiskers show adjacent values. Central lines indicate the median value; X indicates the mean.
We next reviewed the COSEWIC status assessments to determine whether the movement-related questions that the policy process explicitly asked were answered (Table 1). Status assessments included seven key questions related to movement:
• Is the extent of occupancy (EOO; species range) known?
• Is the area of occurrence (AOO; individual range) known?
• Is the population severely fragmented?
• Are the EOO or AOO in decline?
• Are there extreme fluctuations in EOO or AOO?
• Is habitat area, extent or quality in decline?
• Is rescue through immigration possible and likely?
The technical summaries of the COSEWIC status assessments explicitly address each question in the same format, clearly stating whether each criterion can be addressed, or represents a knowledge gap. Answers to these questions are critical to management because they were required to determine whether a species meets the criteria for listing as a species at risk.
We found evidence that the field of movement ecology is, with only a few exceptions, filling key knowledge demands in species status assessments, and recovery plans (Table 1). For every species, the EOO, the level of fragmentation (severe or not), level of fluctuation in EOO and AOO, and the trend in habitat area, extent or quality (declining or not), were known. Less certain were the trends in EOO and AOO and whether or not a rescue effect was possible. The questions were often answered based on one or a few papers [e.g., Roseate Tern (Sterna dougallii), Whooping Crane (Grus americana), White Sturgeon (Acipenser transmontanus), and Killer Whales (Orcinus orca) all had 11 or fewer studies] (Figure 2). The absence of an asymptote in Figure 2 suggests that saturation of movement ecology science in conservation has not yet been reached for any of the 12 species. For some species, critical knowledge gaps remained at the time of status assessment that could be addressed with new movement ecology data. For example, the Kirtland's Warbler assessment noted unknown EOO and AOO (as well as declines in these parameters; see Table 1) but recent direct tracking data may contribute to filling these knowledge gaps for this species in future (Cooper et al., 2017).
New knowledge may be particularly useful in defining the temporal trends in EOO and AOO. For example, the range of grizzly bears is thought to be shrinking in the U.S. and expanding in northern Canada, potentially due to climate change, but this range shift has not been accurately measured (Committee on the Status of Endangered Wildlife in Canada, 2012). A critical limitation of movement ecology data is the potential illusion of representativeness they create because few animals in the population are tagged (Hebblewhite and Haydon, 2010). For example, most studies of migratory birds have sample sizes of <20 individuals (McKinnon et al., 2013), thus identifying only a small proportion of critical habitats used by target species. As more individuals are monitored at multiple locations across their distribution, we can improve understanding of population dynamics and habitat use across large geographic scales (Fraser et al., 2012; Stanley et al., 2015).
Our analysis quantifies the fulfillment of the conservation promise and potential of movement ecology research through informing policy processes, such as the status assessment and recovery planning for species at risk. Research focused on regional (i.e., state, province) level wildlife management plans in North America suggested that the putative hallmarks of contemporary science are not being used by management, when this was examined in species that are harvested (Artelle et al., 2018). In contrast, our objective analysis of scientific uptake suggests that the relatively novel science of movement ecology is already and importantly informing the assessment process in wildlife management and conservation nationally and internationally.
How Can Movement Ecologists Improve the Integration of Their Research Into Applied Wildlife Conservation?
Two important factors can limit a deeper integration of movement ecology into conservation policy: (1) the findings present in the study are unavailable to the assessment group; and (2) the data are deemed inapplicable or irrelevant to the assessment process. Researchers can increase the uptake of their work by conservation practitioners and policy makers by directly addressing both factors. Accessibility of movement ecology research can be improved by publishing in open-access journals, by improving the science communication and social media surrounding such studies (Peoples et al., 2016), and by creating lay summaries of their work (Brisbois et al., 2018).
Language was not a factor in our case study, but making research results available in the language of relevance to location or species of interest may also increase data application. Promoting movement ecology research through social media (Peoples et al., 2016) to maximize exposure to conservation work may also improve uptake, although publications still need to be accessible for this strategy to be effective. Developing a variety of data products that facilitate the communication of results to broader audiences, such as interactive data products, and visualizations (Wszola et al., 2017), may increase the use and uptake of movement ecology data into conservation applications. Highly collaborative approaches formed at the beginning of movement ecology research projects that involve managers and multi-stakeholders may be highly effective and limit the need for uni-directional knowledge transfer systems, from science to management (Roux et al., 2006). The implementation of a variety of strategies aimed at increasing the speed and amount of uptake of movement ecology research into conservation applications through increasing dialogue and collaboration are bound to be most effective.
Ensuring that movement ecology research contributes directly to solving conservation challenges requires dialogue between researchers, wildlife managers, and policy makers. Improved dialogue can not only increase uptake of research by policy makers, but can also increase uptake of policy makers' science needs by researchers, who can better tailor new movement ecology research to address questions of the greatest value for conservation and management. For example, data collected at a scale that will most benefit management actions will better facilitate the application of movement ecology data to conservation and management (Allen and Singh, 2016).
Creating new frameworks for the application of movement ecology is another novel frontier in conservation. Human-wildlife conflicts are on the rise in the airspace, threatening a diverse array of aerial taxa (Lambertucci et al., 2015). For example, many threatened migratory songbird species are at risk of collision with buildings, hydro lines, and wind turbines as they journey across North America on their seasonal migrations each spring and fall. In recognition of this developing conservation need, (Davy et al., 2017) created a new framework for understanding how organisms move through and use aerial habitats. Applying classic terrestrial-based paradigms of “corridors” to these aerial habitats gives rise to a novel application of movement ecology: quantifying aerial corridors and defragmenting aerial habitats. The utility of this framework for achieving conservation outcomes depends, in part, on knowledge emerging from the movements of organisms through the airscape. For example, recent advances in combined GPS/altimeter data-logging have allowed 3-D tracking of bats moving through complex aerial habitats (Roeleke et al., 2016, 2018) and tracking small (<100 g) vertebrates in real-time from a space-mounted receiver will soon be possible through the implementation of the system (ICARUS, 2018). By providing data on the spatiotemporal movements of animals in the airspace, these new systems will generate innumerable opportunities for conservation and dynamic management that targets mass migrations and other diurnal and seasonal periods when numerous taxa are aloft.
Finally, movement ecology underlies existing policies and efforts to enhance population viability for a host of mobile organisms. Large-scale, collaborative conservation initiatives—such as the Yellowstone to Yukon Conservation Initiative [Y2Y], Algonquin to Adirondack, Baja to Bering [B2B], Migratory Birds Convention—have animal movement at the heart of their planning strategy. For example, the design of Y2Y is largely based on the movement of grizzly bears through the Rocky Mountains of North America, and B2B is based on the long-distance movements of several species, including gray whale (Eschrichtius robustus) migration along the Pacific Coast of the Americas. An outward-looking approach to movement ecology would link research to these initiatives to identify barriers to movement and opportunities for conservation. Likewise, as signatories to the Convention on Biological Diversity (Convention on Biological Diversity, 2017) strive to implement Aichi Target 11 (17% of terrestrial and inland water areas and 10% of coastal and marine areas be protected by 2020), greater attention is being paid to enhance connectivity through the protected areas network. Access to knowledge of how organisms move through disturbed landscapes to maintain ecological flows among protected areas will help ensure that global commitments to the CBD are biologically meaningful in an era of climate change (Coristine et al., 2018).
Conclusion
The lack of basic data on species distributions, habitat requirements, and migratory behavior or movements can be obstacles to status assessment and recovery planning for threatened species (Parsons, 2016). We have identified important linkages between the production of movement ecology science and the intent to inform the effective conservation and management of highly mobile animals. We found that much of the available movement ecology research for a given species at risk was used in the status assessments we examined, suggesting movement ecology is providing valuable data for improving the conservation of wildlife. However, we show a lack of available movement ecology data for some at-risk species, suggesting that despite a massive increase in movement ecology research, many gaps may remain even for our most at risk species. In a golden age of bio-logging, genetic tagging, and biotelemetry a growing variety of animals can now be tracked with ever-greater precision across vast spatial scales. At the same time, the current era has been characterized by immense anthropogenic change to the globe, with concomitant decline in species abundance (Dirzo et al., 2014) and the movement of individual animals (Tucker et al., 2018). There is an unprecedented opportunity for movement ecology to fulfill its conservation potential in an era of rapid, human-induced global change.
Author Contributions
KF, KD, CD, AF, DF, and EM conceived and developed the project, and wrote and edited the manuscript. KF, KD, CD, and AF analyzed the data.
Funding
This research was supported by the Natural Sciences and Engineering Research Council of Canada and the University of Manitoba and Liber Ero Foundation.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2018.00150/full#supplementary-material
References
Allen, A. M., and Singh, N. J. (2016). Linking movement ecology with wildlife management and conservation. Front. Ecol. Evol. 3:155. doi: 10.3389/fevo.2015.00155
Artelle, K. A., Reynolds, J. D., Treves, A., Walsh, J. C., Paquet, P. C., and Dairmont, C. T. (2018). Hallmarks of science missing from North American wildlife management. Sci. Adv. 4:eaao0167. doi: 10.1126/sciadv.aao0167
Barton, P. S., Lentini, P. E., Alacs, E., Bau, S., Buckley, Y. M., Burns, E. L., et al. (2015). Guidelines for using movement science to inform biodiversity policy. Environ. Manage. 56, 791–801. doi: 10.1007/s00267-015-0570-5
Brisbois, M. C., Girling, K., and Findlay, S. (2018). Academics should build rapport with government's policy analysts. Nature 555:165. doi: 10.1057/s41599-018-0144-2
Castle, K. T., Weller, T. J., Cryan, P. M., Hein, C. D., Schirmacher, M. R., et al. (2015). Using sutures to attach miniature tracking tags to small bats for multimonth movement and behavioral studies. Ecol. Evol. 5, 2980–2989. doi: 10.1002/ece3.1584
Clark, T. D., Furey, N. B., Rechisky, E. L., Gale, M. K., Jeffries, K. M., Porter, A. D., et al. (2016). Tracking wild sockeye salmon smolts to the ocean reveals distinct regions of nocturnal movement and high mortality. Ecol. Appl. 26, 959–978. doi: 10.1890/15-0632
Committee on the Status of Endangered Wildlife in Canada (2012). COSEWIC Assessment and Status Report on the Grizzly Bear Ursus Arctos in Canada. Ottawa, ON: COSEWIC. xiv + 84 pp. Available online at: www.registrelepsararegistry.gc.ca/default_e.cfm
Convention on Biological Diversity (2017). Available online at: https://www.cbd.int/sp/targets/
Cooke, S. J., Nguyen, V. M., Kessel, S. T., Hussey, N. E., Young, N., and Ford, A. T. (2017). Troubling issues at the frontier of animal tracking for conservation and management. Conserv. Biol. 31, 1205–1207. doi: 10.1111/cobi.12895
Cooper, N. W., Hallworth, M. T., and Marra, P. P. (2017). Light-level geolocation reveals wintering distribution, migration routes, and primary stopover locations of an endangered long-distance migratory songbird. J. Avian Biol. 48, 209–219. doi: 10.1111/jav.01096
Coristine, L. E., Jacob, A. L., Schuster, R., Otto, S. P., Baron, N. E., Bennett, N. J., et al. (2018). Facets 3, 531–562. doi: 10.1139/facets-2017-0102
Davy, C. M., Ford, A. T., and Fraser, K. C. (2017). Aeroconservation for the fragmented skies. Conserv. Lett. 10, 773–780. doi: 10.1111/conl.12347
Dirzo, R., Young, H. S., Galette, M., Ceballos, G., Isacc, N. J. B., and Collen, B. (2014). Defaunation in the Anthropocene. Science 345, 401–406. doi: 10.1126/science.1251817
Fraser, K. C., Stutchbury, B. J. M., Silverio, C., Kramer, P. M., Barrow, J., Newstead, D., et al. (2012). Continent-wide tracking to determine migratory connectivity and tropical habitat associations of a declining aerial insectivore. Proc. Biol. Sci. 279, 4901–4906. doi: 10.1098/rspb.2012.2207
Godley, B. J., Blumenthal, J. M., Broderick, A. C., Coyne, M. S., Godfrey, M. H., Hawkes, L. A., et al. (2007). Satellite tracking of sea turtles: where have we been and where do we go next? Endanger. Species Res. 3, 1–20.
Hays, G. C., Ferreira, L. C., Sequeira, A. M. M., Meekan, M. G., Duarte, C. M., Bailey, H., et al. (2016). Key questions in marine megafauna movement ecology. Trends Ecol. Evol. 31, 463–475. doi: 10.1016/j.tree.2016.02.015
Hebblewhite, M., and Haydon, D. T. (2010). Distinguishing technology from biology: a critical review of the use of gps telemetry data in ecology. Philos. T R. Soc. B 365, 2303–2312. doi: 10.1098/rstb.2010.0087
Hoop, J. M., Venderlaan, A. S., Cole, T. V., Henry, A. G., Hall, L., Mase-Guthrie, B., et al. (2015). Vessel strikes to large whales before and after the 2008 Ship Strike Rule. Conserv. Lett. 8, 24–32. doi: 10.1111/conl.12105
Hu, G., Lim, K. S., Horvitz, N., Clark, S. J., Reynolds, D. R., Sapir, N., et al. (2016). Mass seasonal bioflows of high-flying insect migrants. Science 354, 1584–1587. doi: 10.1126/science.aah4379
Hussey, N. E., Kessel, S. T., Aarestrup, K., Cooke, S. J., Cowley, P. D., Fisk, A. T., et al. (2015). Aquatic animal telemetry: a panoramic window into the underwater world. Science 348:1255642. doi: 10.1126/science.1255642
ICARUS (2018). International cooperation for animal research using space. Available online at: https://icarusinitiative.org/
Lambertucci, S. A., Shepard, E. L. C., and Wilson, R. P. (2015). Human-wildlife conflicts in a crowded airspace. Science 348, 502–504. doi: 10.1126/science.aaa6743
López-Hoffman, L., Varady, R. G., Flessa, K. W., and Balvanera, P. (2009). Ecosystem services across borders: a framework for transboundary conservation policy. Front. Ecol. Environ. 8:216. doi: 10.1890/070216
McKinnon, E. A., Fraser, K. C., and Stutchbury, B. J. M. (2013). New discoveries in landbird migration using geolocators, and a flight plan for the future. Auk 130, 211–222. doi: 10.1525/auk.2013.12226
Nathan, R. (2008). An emerging movement ecology paradigm. Proc. Natl. Acad. Sci. U. S. A. 105, 19050–19051. doi: 10.1073/pnas.0808918105
O'Neal, M. E., Landis, D. A., Rothwell, E., and Kempel, L. (2004). Tracking insects with harmonic radar: a case study American. Entomologist 50, 212–218.
Parsons, E. C. M. (2016). Why IUCN should replace “Data Deficient” conservation status with a precautionary “Assume Threatened” status—a cetacean case study. Front. Mar. Sci. 3:193. doi: 10.3389/fmars.2016.00193
Peoples, B. K., Midway, S. R., Sackett, D., Lynch, A., and Cooney, P. B. (2016). Twitter predicts citation rates of ecological research. PLoS ONE 11:e0166570. doi: 10.1371/journal.pone.0166570
Roeleke, M., Blohm, T., Kramer-Schadt, S., Yovel, Y., and Voigt, C. C. (2016). Habitat use of bats in relation to wind turbines revealed by GPS tracking. Sci. Rep. 6:28961. doi: 10.1038/srep28961
Roeleke, M., Teige, T., Hoffmeister, U., Klingler, F., and Voigt, C. C. (2018). Aerial-hawking bats adjust their use of space to the lunar cycle. Mov. Ecol. 6:11. doi: 10.1186/s40462-018-0131-7
Roux, D. J., Rogers, K. H., Biggs, H. C., Ashton, P. J., and Sergeant, A. (2006). Bridging the science–management divide: moving from unidirectional knowledge transfer to knowledge interfacing and sharing. Ecol. Soc. 11:4. Available online at: http://www.ecologyandsociety.org/vol11/iss1/art4/
Runge, C. A., Martin, T. G., Possingham, H. P., et al. (2014). Conserving mobile species. Front. Ecol. Environ. 12, 395–402. doi: 10.1890/130237
Sample, C., Fryxell, J. M., Bieri, J. A., Willis, S. G., and Fuller, R. A. (2018). A general modeling framework for describing spatially structured population dynamics. Ecol. Evol. 8:493–508. doi: 10.1002/ece3.3685
Sawyer, H., Middleton, A. D., Hayes, M. M., Kauffman, M. J., and Monteith, K. L. (2016). The extra mile: ungulate migration distance alters the use of seasonal range and exposure to anthropogenic risk. Ecosphere 7:e01534. doi: 10.1002/ecs2.1534
Stanley, C. Q., McKinnon, E. A., Fraser, K. C., Macpherson, M. P., Casbourn, G., Friesen, L., et al. (2015). Connectivity of wood thrush breeding, wintering, and migration sites based on range-wide tracking. Conserv. Biol. 29, 164–174. doi: 10.1111/cobi.12352
Taylor, P. D., Crewe, T. L., Mackenzie, S. A., Lepage, D., Aubry, Y., Crysler, Z., et al. (2017). The Motus Wildlife Tracking System: a collaborative research network to enhance the understanding of wildlife movement. Avian Conserv. Ecol. 12:8. doi: 10.5751/ACE-00953-120108
Tucker, M. A., Böhning-Gaese, K., Fagan, W. F., Fryxell, J. M., Van Moorter, B., Alberts, S. C., et al. (2018). Moving in the anthropocene: global reductions in terrestrial mammalian movements. Science 359, 466–469. doi: 10.1126/science.aam9712
Wilmers, C. C., Nickel, B., Bryce, C. M., Smith, J. A., Wheat, R. E., and Yovovich, V. (2015). The golden age of bio-logging: how animal-borne sensors are advancing the frontiers of ecology. Ecology 96, 1741–1753. doi: 10.1890/14-1401.1
Keywords: bio-logging, dispersal, migration, tracking, telemetry
Citation: Fraser KC, Davies KTA, Davy CM, Ford AT, Flockhart DTT and Martins EG (2018) Tracking the Conservation Promise of Movement Ecology. Front. Ecol. Evol. 6:150. doi: 10.3389/fevo.2018.00150
Received: 01 June 2018; Accepted: 10 September 2018;
Published: 01 October 2018.
Edited by:
Jordi Figuerola, Estación Biológica de Doñana (EBD), SpainReviewed by:
Melinda Grace Conners, Old Dominion University, United StatesYannis Peter Papastamatiou, Florida International University, United States
Copyright © 2018 Fraser, Davies, Davy, Ford, Flockhart and Martins. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kevin C. Fraser, a2V2aW4uZnJhc2VyQHVtYW5pdG9iYS5jYQ==