 Andy J. Green
Andy J. Green Johan Elmberg2
Johan Elmberg2 Ádám Lovas-Kiss
Ádám Lovas-Kiss- 1Wetland Ecology Department, Estación Biológica de Doñana, Sevilla, Spain
- 2Department of Science and Bioscience, Kristianstad University, Kristianstad, Sweden
- 3Department of Tisza Research, MTA Centre for Ecological Research-DRI, Debrecen, Hungary
It is well-known that some members of the crow family (Corvidae) are important for seed dispersal either via frugivory (e.g., when feeding on berries) or by scatter hoarding (e.g., of nuts). Dispersal via gut passage of seeds within a fleshy fruit can be considered “classical endozoochory.” However, corvids are rarely recognized as vectors of plants lacking a fleshy fruit, or a large nut (such as plants with a dry achene, capsule or caryopsis). Dispersal of such seeds via gut passage can be considered “non-classical endozoochory.” A century ago, Heintze (1917a,b); Heintze (1918) reported on extensive field studies of seed dispersal by 11 species of European Corvidae. His work is overlooked in contemporary reviews of corvid biology. We resurrect his work, which suggests that contemporary views about seed dispersal by corvids are too narrow. Heintze identified 157 plant taxa from 42 families which were dispersed by corvids by endozoochory, as well as another nine taxa only dispersed by synzoochory (which includes scatter-hoarding). Most (54%) of the plant species dispersed by endozoochory lack a fleshy fruit and have previously been assigned to other dispersal syndromes, mainly associated with wind (10%), self-dispersal (22%) or epizoochory (18%). Plants lacking a fleshy fruit were particularly well-represented from the Caryophyllaceae (12 species), Poaceae (14 species), and Polygonaceae (8 species). Of 27 taxa germinated by Heintze from seeds extracted from corvid pellets or feces (71% of those tested), 20 lack a fleshy fruit. Similarly, of 32 taxa he recorded as seedlings having germinated from pellets in the field, 11 lacked a fleshy fruit. However, Heintze's quantitative data show that classical endozoochory is dominant in Magpies Pica pica and Hooded Crows Corvus cornix, for which 97% of seeds dispersed were fleshy-fruited. Corvids overlap with waterfowl as vectors of terrestrial plants dispersed by non-classical endozoochory, and 56 species are dispersed by both corvids and dabbling ducks according to the lists of Heintze and Soons et al. (2016). Finally, Heintze's data show that corvids were already dispersing alien plants in Europe a century ago, such as the North American Dwarf Serviceberry Amelanchier spicata.
Introduction
Plants disperse their diaspores (“seeds” from here on) by many means (Ridley, 1930), including via animal vectors (“zoochory”). By virtue of their flight, birds are excellent vectors, and their ability to disperse seeds via gut passage (“endozoochory”) is particularly well-known, and was already recognized by the ancient Greeks (Theophrast who died in 287 BC described how birds disperse mistletoe, Holmboe, 1900). Seed dispersal is considered one of the most important of avian ecological functions (Sekercioglu, 2006). Most contemporary research on avian endozoochory is focussed on frugivores, and fruit has been reported to make up 10% or more of the diet of 83 species in the crow family (Corvidae) (Table 5.1 in Wenny et al., 2016). Dispersal of seeds embedded in the fleshy pulp of an edible berry or fruit has been called “classic endozoochory” (McPartland and Naraine, 2018). This corresponds to the long-standing practice of assigning “dispersal syndromes” to plant species on the basis of diaspore morphology, in which only fleshy-fruited diaspores are assigned to a “endozoochory syndrome” (van der Pijl, 1982; Perez-Harguindeguy et al., 2013). On the other hand, the Corvidae are the bird family most associated with plant dispersal by seed-caching or scatter-hoarding. This dispersal mode (a form of “synzoochory”) occurs when jays, nutcrackers or other corvids carry large diaspores, such as nuts, acorns or pine seeds in their bill and then bury them in caches (Pesendorfer et al., 2016; Tomback, 2016).
Corvids are particularly familiar birds to people, and have influenced human culture (Marzluff and Angell, 2007). They provide many ecosystem services, for example through their value in forest regeneration from scatter-hoarding (Tomback, 2016). On the other hand, corvids are partly granivorous and have long had a reputation for causing damage to cereal crops (Barrows, 1888; Hadjisterkotis, 2003), hence the term “scarecrows.” In this paper, we highlight the importance of corvid endozoochory for a broader spectrum of plants, many of which are dispersed by granivory rather than by frugivory. We refer to dispersal of plants that do not have a fleshy fruit as “non-classical endozoochory.” Granivory by corvids can lead to endozoochory of plants assigned to other dispersal syndromes, such as barochory, and which are therefore assumed to disperse by other mechanisms with less capacity to disperse seeds over long distances (Czarnecka and Kitowski, 2013). In order to understand the broader importance of corvid endozoochory, we resurrect the work of Sven August Heintze, a Swedish pioneer in the study of avian seed dispersal a century ago (Figure 1). Heintze's contributions have long been overlooked and are not cited in major reviews of corvid biology or their role in seed dispersal (e.g., Coombs, 1978; Cramp and Perrins, 1994; Birkhead, 2010; Tomback, 2016).

Figure 1. Sven August Heintze (1881–1941) demonstrated the importance of corvids for seed dispersal in a wide range of plant species. Heintze was trained at Lund University, Sweden, where he defended his doctorate in botany in 1913. He published extensively about seed dispersal by birds and mammals in the years 1910-1918 (in Swedish), and later reached out for a wider audience in a two-volume synthesis about plant dispersal (in German; Heintze, 1932, 1935). Three of his papers provide data used for analyses in the present paper, resurrecting Heintze's legacy as a pioneer in this field.
Although the term “endozoochory” did not yet exist at the time, this dispersal process was considered by Holmberger (1785), a student of Linnaeus who reviewed dispersal strategies in plants, who mentioned seed consumption by Magpies Pica pica, and discussed seed survival after gut passage by birds in general. More than a century later, Holmboe (1900) applied the term “endozoochory” (which he attributed to A.P. de Candolle) to corvids. Holmboe recovered many seeds from the gizzards of Northern Nutcracker (Nucifraga caryocatactes caryocatactes) from Norway (see Table S1), but did not demonstrate that they survived gut passage. Soon after, Heintze (1917a,b); Heintze (1918) went much further than Holmboe in the study of endozoochory by corvids, and reported on his own field data for the following seven species: Magpie, Hooded Crow (Corvus cornix), Jackdaw (C. monedula), Rook (C. frugilegus), Common Raven (C. corax), European Jay (Garrulus glandarius), and Siberian Jay (Perisoreus infaustus). Heintze's dataset was most extensive for Magpies and Hooded Crows, allowing us to conduct detailed quantitative analysis of seeds dispersed by these two species. In addition to extensive field studies of plants and bird behavior during all seasons, Heintze carried out germination experiments with seeds extracted from pellets and feces of Magpies and Hooded Crows (Heintze, 1917a,b, 1918).
On top of his own thorough field and experimental work, he reviewed data previously provided by others on seed dispersal by additional corvid species [the seven listed above plus Carrion Crow (Corvus corone), Northern Nutcracker, Red-billed Chough (Pyrrhocorax pyrrhocorax), and Alpine Chough (P. graculus)] (Heintze, 1918). The latter paper ends in a synthetic discussion in which Heintze argues at length that European corvids have a wide role as dispersers of a long list of plant species in a wide range of habitats. Not only is his own research remarkable, the literature he reviewed 100 years ago shows that avian endozoochory was already a well-established research field at that time. Regrettably, these insights published in Swedish were largely neglected in his later seminal books written in German on plant dispersal sensu latu. Heintze (1932, 1935) mentioned the role of corvids only in passing, which is another reason his insights on this topic have been overlooked.
In this paper, we resurrect Heintze's work to show how important European corvids are for both classical and non-classical endozoochory. We identify the plant species he considered to be dispersed by endozoochory by 11 different European corvid species, quantifying the numbers of taxa recorded with different fruit types, morphological dispersal syndromes, habitat requirements and seed size. In the case of his more detailed datasets for pellets and feces of Magpies and Hooded Crows (Heintze, 1917a,b, 1918), we compare the relative abundance of intact seeds from each of the above categories, and compare seeds in pellets with those in feces.
Methods
Methods Used by Heintze
Sven August Heintze was born in Skurup in southern Sweden in 1881 and earned a doctorate in botany at Lund University in 1913. His thesis was on zonation in alpine plants in Swedish Lapland, but he soon changed focus to study seed dispersal by birds. In 1912, Heintze already had the ambition to study the general role of corvids as dispersers of seeds in “Holarctic and Neoboreal” regions. His main contributions to this field were made in 1910–1917, when he collected feces, pellets and gut samples from European corvids (Heintze, 1917a,b, 1918). He also carried out extensive literature compilations, using sources in at least eight languages.
A description of his objectives and methods is found (in Swedish) on page 210 in Heintze (1917a). He traveled extensively in Sweden to collect corvid feces and pellets and examined the gastrointestinal tract of shot corvids. In his exhaustive review of terms for different plant dispersal processes, Heintze (1932) even proposed separate terms for seeds dispersed by egestion in pellets (“hemiendozoochory”) and feces (“euendozoochory”). Heintze was well aware of the role of corvids as scatter-hoarders, which was already widely acknowledged in the literature in the early 1900's, and this is why he focused on endozoochory instead. He emphasized the importance of carrying out these studies “in all seasons,” as he recorded the occurrence of germinated plants in nature (in excreta, at corvid nests, roosts, and at feeding sites) which had been dispersed by corvids. In addition, Heintze made ambitious germination experiments to study the implications of endozoochory; “fresh regurgitates and excrements were subjected to a rinsing/washing process [in the lab] similar to that accomplished by rain- and snowmelt water in nature” (Heintze, 1917a, p. 210). He carried out germination experiments of the contents of regurgitated pellets and excrements from Magpie and Hooded Crow. Most germination trials were carried out in early spring, of seeds that had been collected the preceding autumn or winter. Although many seeds germinated within a few weeks, trials were run throughout summer and autumn if necessary to detect later germinations (e.g., in 1916 the germination experiments were run until October 16th, Heintze, 1917a, p. 240). Further details of these experiments (e.g., substrate, indoor/outdoor, water supply, etc.) are unknown.
Methods Used in Our Review of Heintze's Work
We extracted empirical data from the Swedish texts of Heintze explicitly focussing on seed dispersal by corvids (Heintze, 1917a,b, 1918). These sources list observations about regurgitated pellets, sampling of feces (mainly at roost and nest sites), and on contents of the upper (gizzard) and lower (intestines) gut. These data were originally reported in a rather unsystematic way, with variation in the level of detail and the extent to which different sample types were described. Interfoliated among Heintze's own data are references to research by colleagues, mainly in Central Europe, which are included in his summaries of the general role of corvids as seed dispersers. Many of his original sources are obscure and difficult to trace.
From Heintze's texts, we extracted details of those plant species with strong evidence of endozoochory, in particular cases where details of the bird species were provided, together with good evidence for endozoochory either because seeds were found in an intact state in pellets, feces or the alimentary canal, or they were germinated after removal from pellets or feces. Heintze stated that his list of species he considered to be dispersed by endozoochory “contains plant species that with full certainty or with high degree of certainty are spread endozooically by European corvids” (Heintze, 1918, p. 35). He also specified which plants were dispersed by synzoochory (i.e., scatter hoarding or for direct consumption at roosts without caching), especially by European Jays and Northern Nutcrackers, and we summarize these data for comparison.
We present detailed results for two different sets of Heintze's data. Firstly, a complete list of the plant taxa dispersed by endozoochory in Northern and Central Europe by any of the 11 species of corvids which he reviewed (Heintze, 1918). Secondly, his own detailed empirical dataset on pellets and fecal samples of Magpies and Hooded Crows in Sweden, for which he provided the number of intact seeds extracted for each plant species. The first dataset allows us to analyse what kinds of plant taxa were dispersed by endozoochory. The second dataset allows us to do a quantitative analysis of the relative abundance of seeds from each plant category (i.e., of seed size, fruit morphology, dispersal syndrome or habitat requirements) in pellets and feces for both species. The vast majority of this second dataset was collected from a restricted area on the west coast of southern Sweden.
The categories of fruit type and dispersal syndrome were assigned for each plant species from the Baseflor database (Julve, 1998). Seed length data were from the LEDA traitbase (Kleyer et al., 2008). When multiple measurements were available, we used their mean. Ellenberg moisture values were obtained from Hill et al. (1999).
For Heintze's detailed dataset for Magpies and Hooded Crows, we compared the frequency of occurrence of intact seeds of different plant species between pellets and feces using Fisher exact tests using online software (http://vassarstats.net/tab2x2.html). We then compared the frequency of occurrence of different plants in pellets of the two corvid species. We did not repeat this for feces because of small sample sizes. We also compared the paired frequencies of occurrence of each plant species between pellets (fp) and feces (fe) for each corvid species using a matched paired t-test (i.e., testing fp-fe), to see if there was a consistent difference between sample types (e.g., higher frequency of seeds in pellets). We then tested if the difference between the frequencies in pellets and feces (i.e., fp-fe) for individual plant taxa was related to seed size, using a non-parametric correlation (between fp-fe and seed length). T-tests and correlations were carried out in R (R Core Team, 2017).
Unfortunately, it was not possible for us to realize all the statistical tests of interest, as many of the details of which seeds were present in each individual sample were lacking, even for Magpies and Hooded Crows. Heintze did not always report which plant species were found together, or the combined total number of seeds, in each sample.
Results
Diversity of Plants Dispersed by Endozoochory and Their Relation With Dispersal Syndromes
When combining Heintze's reviews of the literature with his own data, a total of 157 plant species from 42 angiosperm families were recorded as dispersed by endozoochory by at least one corvid species (Table S1). In contrast, Heintze cited nine taxa as dispersed exclusively by synzoochory (including scatter-hoarding): Aesculus hippocastanum, Carpinus betulus, Castanea sativa, Corylus avellana, Fagus silvatica, Juglans regia, Pinus cembra, Quercus robur, and Q. sessiliflora.
Of those species dispersed by endozoochory, 91 (56%) do not have a fleshy fruit and hence do not belong to the “endozoochory syndrome,” and were dispersed by non-classical endozoochory (Table S1; Figure 2). Many taxa with epizoochory, barochory and anemochory syndromes were recorded (Figure 2). Hence, many plants generally assumed to disperse via other mechanisms, such as wind, attached to fur, or self-dispersal, were dispersed by non-classical endozoochory. Those plant species dispersed by the most corvid species were fleshy-fruited, with up to nine different vectors per plant (Table S1). Nevertheless, some species with dry achenes and associated with barochory or epizoochory syndromes were dispersed by as many as six different corvid species (e.g., Polygonum aviculare, Rumex acetosella, Urtica urens, Table S1). Amongst the corvid species, the Magpie (114 plant species) and Hooded Crow (105) were recorded as vectors for the most plant species, reflecting Heintze's extensive work with those corvid species. They were followed by the Raven (71), Carrion Crow (33) and Jackdaw (29), whereas the Red-billed Chough (4) dispersed the fewest plant species (Table S1). The relative importance of classical and non-classical endozoochory varied between species. For example, of 29 species dispersed by Jackdaws, 18 (62%) were via non-classical endozoochory, compared to 36 (51%) of 71 species dispersed by Ravens.
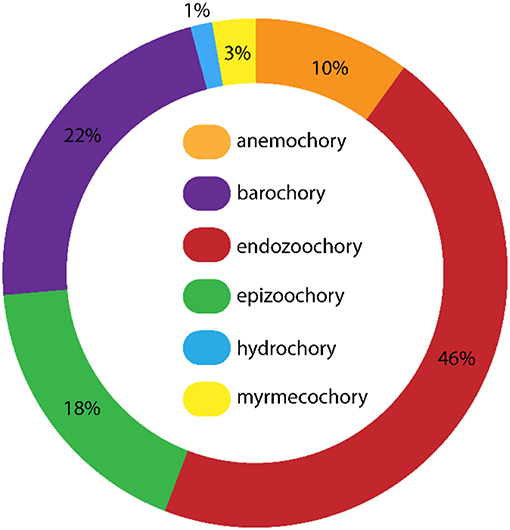
Figure 2. Relative frequencies of different dispersal syndromes (Baseflor, Julve, 1998) assigned to all the plant taxa dispersed by corvids through endozoochory, according to Heintze's complete list (n = 148 taxa, another nine species were missing because their dispersal syndrome was unknown, see Table S1). Anemochory is wind dispersal, barochory is self-dispersal, hydrochory is water dispersal, epizoochory is dispersal stuck on the outside of animals, myrmecochory is dispersal by ants (see Julve, 1998; Perez-Harguindeguy et al., 2013).
Five species alien to the European continent were included amongst the plants dispersed by corvid endozoochory (Table S1). Aliens to Europe included the Dwarf Serviceberry Amelanchier spicata which is native to North America, the other four being cultivated species, such as the Cucumber Cucumis sativus, White Mulberry Morus alba, and Buckwheat Fagopyrum esculentum. Amongst the species recorded in Heintze's samples from Sweden, there were also an additional 10 species alien to Sweden but native to other parts of Europe, such as the grass Bromus arvensis (Table S1).
For the quantitative dataset on pellets and feces of Magpie and Hooded Crow produced from intensive study by Heintze, intact seeds from a total of 65 plant species (plus 5 plants identified to genus and one to family) were recorded, of which 37 (56%) do not have a fleshy fruit and so do not belong to an “endozoochory syndrome” (Table 1). Many taxa assigned to epizoochory (24% of taxa), barochory (20%) and anemochory (10%) syndromes were dispersed by non-classical endozoochory (Table 1). Nevertheless, when considering the total abundance of seeds from each syndrome, and not merely the number of taxa, the pattern changed markedly, and 97% of seeds were from an endozoochory syndrome (Table 1).
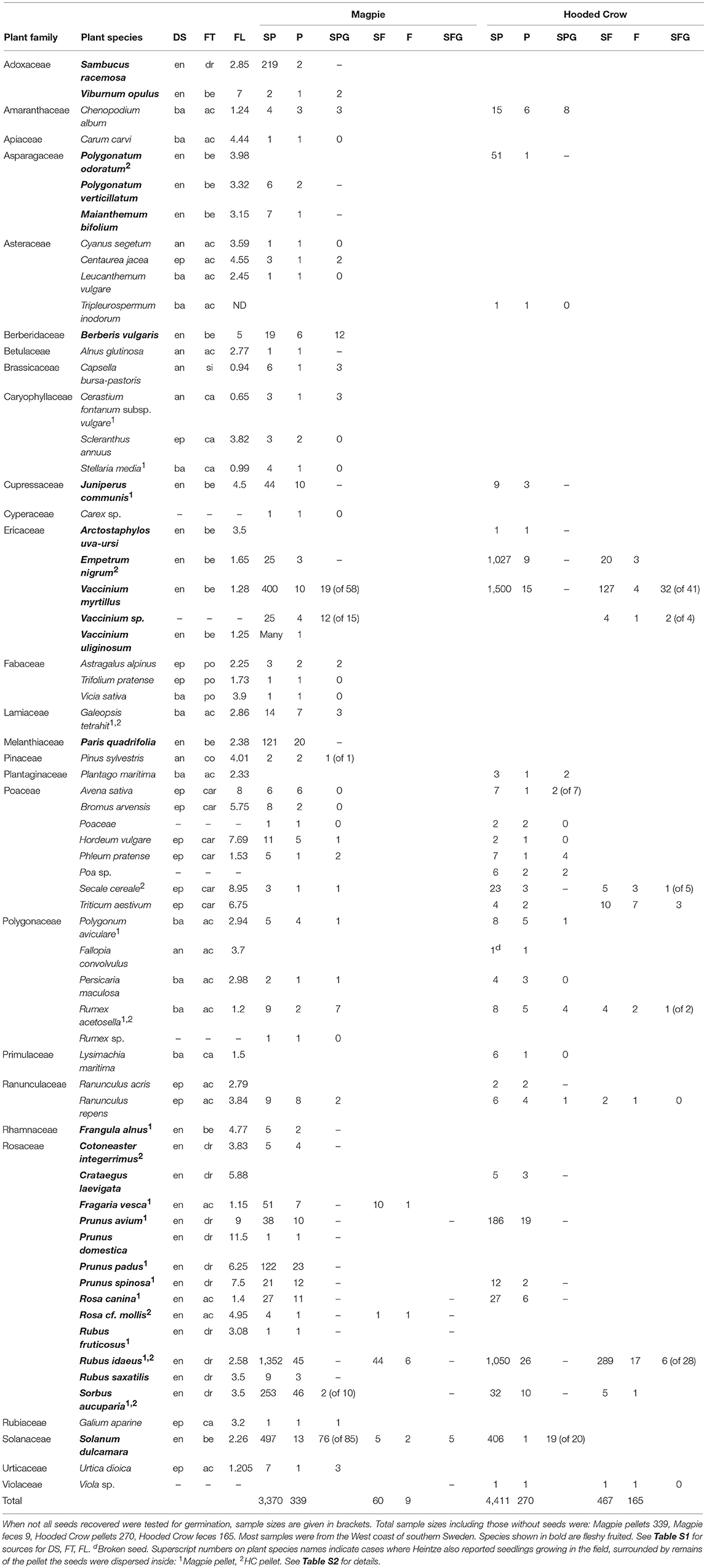
Table 1. Abundance and frequencies of different plant taxa recovered by Heintze from the pellets and feces of Magpies and Hooded Crows, showing: Dispersal syndrome (DS, an, anemochory; ba, barochory; en, endozoochory; ep, epizoochory), Fruit type (FT, ac, achene; be, berry; ca, capsule; car, caryopsis; co, cone; dr, drupe; po, pod; si, silique), Fruit length (FL), Number of intact seeds in pellets (SP), Number of pellets containing seeds (P), Number of seeds germinated from pellets (SPG), Number of intact seeds found in feces (SF), Number of fecal samples containing seeds (F), Number of seeds germinated from fecal samples (SFG).
Germination tests confirmed the viability of seeds from 25 plant species (from a total of 35 species tested, i.e., 71%) plus two taxa identified to genus whose seeds were recovered from pellets and feces of Magpies and Hooded Crows (Table 1). Viability was confirmed for taxa from four dispersal syndromes, including all of 6 species with an endozoochory syndrome, 6 of 12 with barochory, 3 of 4 with anemochory and 10 of 13 with epizoochory (note, the number of seeds tested per species varied, Table 1). Overall, two taxa were only germinated from feces, 16 were only germinated from pellets, and 5 species (20% of the total) were germinated from both pellets and feces (Table 1). In addition, Heintze germinated Polygonum aviculare from the rectum of a European Jay, and Vaccinium cf vitis-idea from the feces of a Raven.
In addition, Heintze (1917a, 1918) recorded seedlings of at least 32 plant species in the process of becoming established in nature after germinating within the remnants of excreta (27 plant species in Magpie pellets and at least 10 in Hooded Crow pellets). These species included at least 11 that lack fleshy fruits and have been assigned to epizoochory, anemochory and barochory syndromes (Table 1; Table S2), confirming the importance of non-classical endozoochory.
Fruit Types
Plant taxa dispersed by corvid endozoochory were from a broad range of fruit types, of which berries, drupes and achenes were the most frequent (Figure 3). Only seven of 45 taxa with achenes had a fleshy achene, corresponding to the “endozoochory syndrome.” When considering fruit types for plant taxa recovered from pellets and feces of Magpies and Hooded Crows (Figure 4), berries and drupes were more dominant, together with smaller numbers of achenes (only 11% of achenes recovered were fleshy). This is consistent with the dominance of seeds with an endozoochory syndrome in these samples (Table 1). For Hooded Crows, drupes appeared more likely to be recorded in feces and berries more likely to be recorded in pellets (Figure 4), although we were unable to test the statistical significance of this difference.
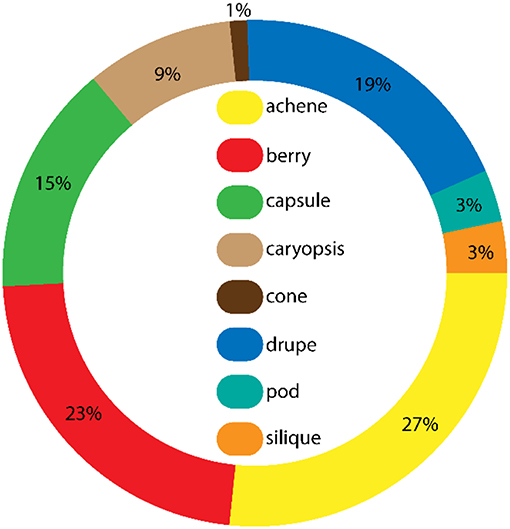
Figure 3. Relative frequency of different fruit types (Baseflor, Julve, 1998) for all the plant taxa dispersed by European corvids through endozoochory, according to Heintze's complete list (n = 150 taxa, excluding seven species with unknown fruit types, see Table S1 for details). Fleshy fruits are drupes, berries, and seven of the 45 species with achenes (such as the fig Ficus carica).
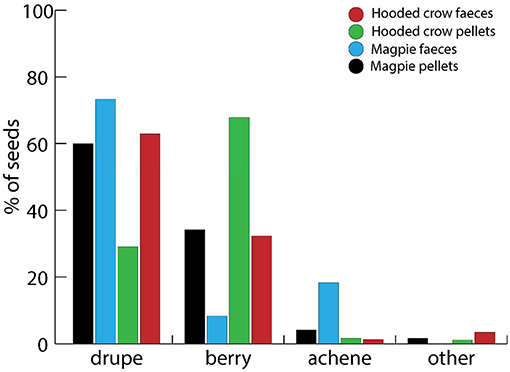
Figure 4. Relative abundance of different fruit types (Baseflor, Julve, 1998) among intact seeds recovered from pellet and fecal samples of Magpies and Hooded Crows by Heintze. Fleshy fruits are drupes, berries and three species with fleshy achenes (such as dog rose, Rosa canina). “Other” includes small numbers each of siliques, capsules, pods, cones, and caryopsis. Sample sizes: Magpie pellet: 3,370 seeds; Magpie feces: 60 seeds; Hooded Crow pellet: 4,411 seeds; Hooded crow feces: 467 seeds.
Seed Size
European corvids dispersed seeds with a broad size range by endozoochory, from a minimum seed length of 0.75 mm (Cerastium fontanum) to a maximum length of 11.5 mm (Prunus domestica), although most taxa had a length of <4 mm (Figure 5). In the detailed dataset for Magpies and Hooded Crows, the size distribution was more skewed toward smaller seeds (Figure 6). Comparing the size distribution for all taxa dispersed by corvids (Figure 5) with the data on relative abundance (Figure 6) suggests that taxa with relatively small seeds (<4 mm in length) were dispersed more often.
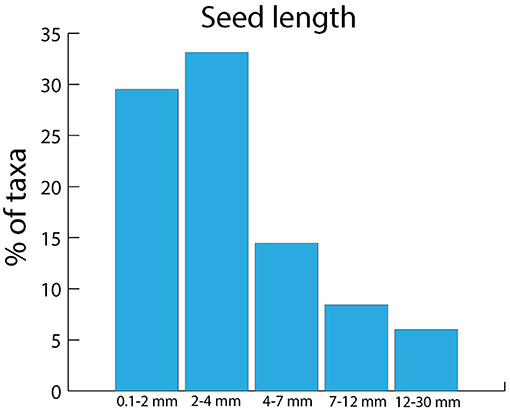
Figure 5. Seed length distribution for all the plant taxa dispersed by European corvids through endozoochory, according to Heintze's complete list (n = 143 taxa, 14 species were excluded because their seed length was unknown, see Table S1).
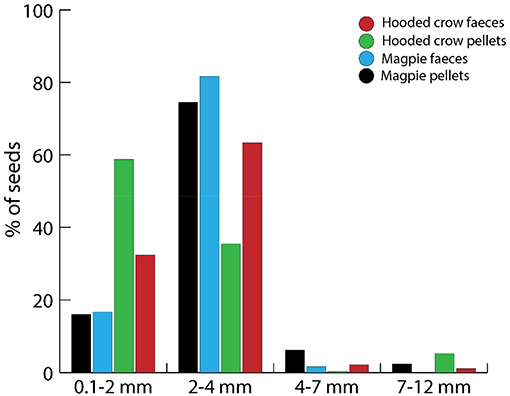
Figure 6. Size distribution of all intact seeds recovered from pellets and fecal samples of Magpies and Hooded Crows by Heintze. See Figure 4 for sample sizes.
Seeds dispersed by Magpies tended to be larger than those dispersed by Hooded Crows, with a higher proportion of seeds of 2–4 mm for Magpies. Surprisingly, seeds tended to be larger overall in the fecal samples of Hooded Crows than in their pellets, although the largest seeds of >4 mm were more frequent in pellets (Figure 6).
Plant Habitat Requirements
Plant taxa dispersed by corvid endozoochory were from a broad range of terrestrial habitats, as indicated by Ellenberg moisture values (Figure 7). When considering the numbers of seeds from pellets and feces of Magpies and Hooded Crows assigned to each Ellenberg value, there was a tendency for higher soil moisture, and values 6 and 8 were better represented (Figure 8). No aquatic plants were recorded.
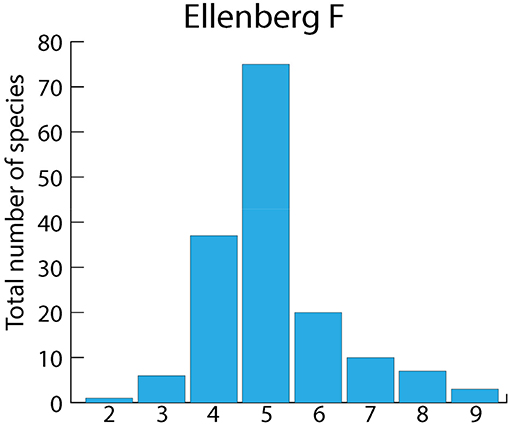
Figure 7. Relative frequencies of Ellenberg moisture indicator values (F) for plant taxa dispersed by corvids through endozoochory, according to Heintze's complete list. N = 150 taxa (excluding seven species with an unknown F value, see Table S1). This scale goes from 1 (dry soils) to 12 (submerged macrophytes). Plants with F ≥ 10 can be considered aquatic.
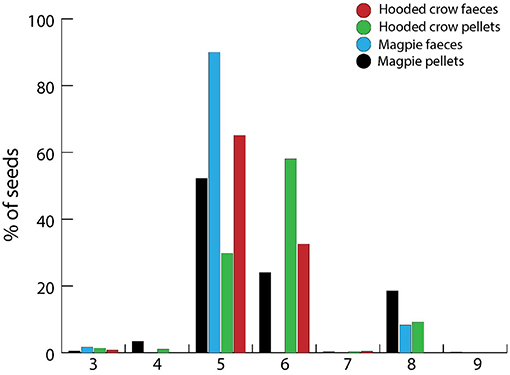
Figure 8. Relative abundance of intact seeds recovered from pellet and fecal samples of Magpies and Hooded Crows by Heintze assigned to different Ellenberg moisture indicator values (F). This scale goes from 1 (dry soils) to 12 (submerged macrophytes). For Ellenberg 9 there was only 1 seed (0.02% of the total) of alder Alnus glutinosa from a Magpie pellet. See Figure 7 for sample sizes.
Comparing the Detailed Composition of Pellets and Feces of Magpies and Hooded Crows
Overall, on average there were more intact seeds in Magpie pellets (9.9 seeds per sample) than their feces (6.6 seeds). Similarly, there were more seeds in Hooded Crow pellets (16.3 seeds per sample) than in their feces (2.8 seeds). However, since Heintze did not provide the combined total number of seeds in individual samples, we could not test these differences statistically.
For Magpies there were relatively few fecal samples, but Rubus idaeus was significantly more prevalent in feces than in pellets (Fisher exact, P = 0.0004). For Hooded Crows, Triticum aestivum was more prevalent in feces (P = 0.001), whereas Prunus avium was more prevalent in pellets (P = 0.0003). Eight plant species were significantly more frequent in Magpie pellets than in Hooded Crow pellets, and two species were more frequent in Hooded Crow pellets (Table 2).
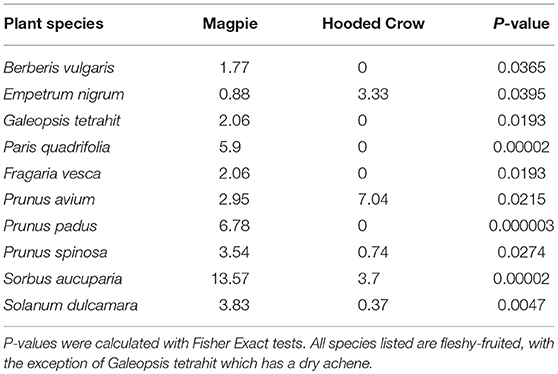
Table 2. Plant species with a significant difference in the frequency of occurrence (given as %) of intact seeds between the pellets of Magpies and Hooded Crows.
In Hooded Crows, there was a higher overall frequency of seeds in pellets than in feces (comparing the frequencies of occurrence for each plant taxon with a matched paired t-test: t = 2.73, df = 64, p = 0.0082). In Magpies, there were few fecal samples and no significant difference in frequencies with pellets (t = 0.373, df = 63, p = 0.71). There was no relationship between the seed size of a particular plant species and the difference in its frequency of occurrence between pellets and feces, whether for Magpies (ρ = 0.106, p = 0.48) or Hooded Crows (ρ = 0.074, p = 0.72).
Discussion
Heintze's work shows that corvids disperse a broad taxonomic and ecological range of plants through gut passage, and slightly more than half of the taxa are dispersed by non-classical endozoochory (i.e., through granivory). The importance of classical endozoochory by corvids has adequately been demonstrated by much contemporary research (Nogales et al., 1999; Wenny et al., 2016; Bai et al., 2017). Heintze (1918, p. 29) also wrote: “Magpies, Hooded Crows, Carrion Crows, Rooks, Jackdaws, Ravens, Choughs, Yellow-billed Choughs, and Siberian Jays disperse seeds of pretty much all European trees, bushes, shrubs, and herbs with berries [fleshy fruits].” The importance of synzoochory by corvids has also been demonstrated by much contemporary research (Pesendorfer et al., 2016; Tomback, 2016). Heintze's dataset identifies few plant species dispersed by synzoochory, but this was probably a consequence of his deliberate focus on endozoochory.
Non-classical endozoochory can also be considered a form of “non-standard dispersal” because the means of dispersal does not match the dispersal syndrome (Higgins et al., 2003; Nathan et al., 2008). However, syndromes are commonly assigned based solely on a botanist's interpretation of fruit morphology, and in the absence of empirical data on dispersal mechanisms. Hence, syndromes may not identify what mechanisms are “standard” in a sense that has any true ecological or evolutionary meaning, and they have proved to be poor predictors of mechanisms for long-distance dispersal (Higgins et al., 2003; Nogales et al., 2012; Lovas-Kiss et al., 2018a, 2019). Syndromes recognize the importance of frugivory (through the “endozoochory syndrome”), but ignore the role of granivory as a dispersal mechanism.
Our results are supported by a careful reading of other corvid literature showing that corvids can be important vectors by non-classical endozoochory. In his seminal review of plant dispersal, Ridley (1930) recognized the importance of different corvid species around the world for frugivory and scatter-hoarding, but he also provided a list of over 50 plant taxa with dry fruits and grains that may be dispersed within pellets. Ridley cited Heintze (misspelled as “Heinitz”) and listed some of the plant species he recorded in the diet of Hooded Crow and Magpie. Herb seeds were the most abundant food item in gut contents of adult Magpies from the former Soviet Union (Cramp and Perrins, 1994). Unique modern research shows the importance of non-classical endozoochory in Rooks (Czarnecka and Kitowski, 2010, 2013; Czarnecka et al., 2013). Other passerines with similar ecology to corvids can also be important vectors of non-classical endozoochory, as illustrated by the Australian Magpie Gymnorhina tibicen (Artamidae, Twigg et al., 2009).
Classical vs. Non-classical Endozoochory
Based on presence/absence of plant taxa dispersed by endozoochory, classical and non-classical endozoochory by corvids appear to be equally important. However, the quantitative dataset for Magpies and Hooded Crows suggest that classical endozoochory events are considerably more frequent. Heintze found a similar diversity of plant taxa with and without a fleshy-fruit to be dispersed by these two species, but seeds from the latter were much less abundant. Heintze's extensive dataset suggests important differences among different corvid species as plant vectors by endozoochory, although this dataset is influenced by differences in sampling effort and relative abundance of each corvid in the habitats where he and his colleagues worked. His data do not allow us to fully assess the relative importance of classical vs. non-classical endozoochory in the other nine species, but more recent research demonstrates that classical endozoochory is not always dominant. Czarnecka and Kitowski (2013) found for Rooks in Poland that seeds were present in 18% of pellets, and half of all seeds belonged to dry-fruited species.
Differences among corvid species in their roles as plant vectors are inevitable, given their differences in habitat use. Heintze argued that in landscapes transformed by agriculture in southern Sweden, the role of Magpie and Hooded Crow as plant vectors by endozoochory was equivalent to that played by the Raven and European Jay in forest landscapes. He also observed that some corvids disperse plants by secondary endozoochory, a form of diplochory (Vander Wall and Longland, 2004). Magpies, Hooded Crows, Rooks, and Jackdaws frequently extracted seeds from the dung of horses, cows, sheep, goats, and wild ungulates, and Ravens consumed seeds from dung of reindeer, cows, and horses (Heintze, 1917a,b, 1918). Similarly, seeds in the feces of herbivores, such as ungulates or elephants are often secondarily dispersed by rodents that extract them from the dung (Jaganathan et al., 2016).
Heintze recognized that differences between corvid species in the amounts of grit in the gizzard are likely to influence seed survival (as shown for Anatidae, Figuerola et al., 2002), and suggested that Magpie, Raven, Siberian Jay, and Northern Nutcracker all have little or no grit. In contrast, he found Eurasian Jays to contain the most grit, and combined with their habit of rarely producing pellets he concluded “the Jay surely has a much lesser importance as an endozoochoric species than do other corvids” (Heintze, 1917a, pp. 209, 225; Heintze, 1918, pp.7, 11, 15), although this species is a major plant vector by synzoochory (Pesendorfer et al., 2016).
Corvids Are Effective Dispersers Over Long Distances
For dispersal to be effective, seeds must be dispersed into a suitable habitat allowing germination and subsequent growth. Heintze frequently reported field observations on established plants growing near Magpie nests, which he concluded had been dispersed there by the birds (Heintze, 1917a). His extensive observations of seedlings growing among the remnants of Magpie and Hooded Crow pellets are particularly important (Table 1; Table S2), confirming that not only are viable seeds dispersed by classical and non-classical endozoochory, but that they are moved into suitable microhabitats. Heintze also considered Raven pellets to be important in the establishment of plants in nature, and cited Holmboe as recording many germinated plants under Raven nests, e.g., Actea spicata in Norway and Ficus caria and Olea europea at nests in Cyprus (Heintze, 1918, p. 7–8).
Central to the role of corvids as endozoochoric seed dispersers are their movements between ingestion and egestion of a viable seed. Heintze knew that the Magpies and Hooded Crows he studied mainly undertook daily, local movements rather than long-distance migrations. He wrote that Magpies “regularly transport seeds ca 2 km, rarely more than 5 km” (Heintze, 1917a, p. 229–230). He found seeds from seashore plants in Magpie pellets retrieved 5 km from the closest seashore (Heintze, 1917a, pp. 217 and 230). For Hooded Crow, he stated “they frequently fly 10–20 km, or even longer, to reach their regular night roost” (Heintze, 1918, p. 2). He also provided known dispersal distances for Raven; “often 20–30 km” (Heintze, 1918, p. 9) and Rook [50–60 km, citing Altum (Heintze, 1918, p. 6)]. Moreover, Heintze was interested in endozoochory over longer distances during seasonal migrations by the Hooded Crow. He suggested that this species has the capacity for overseas seed dispersal between land masses, particularly since his dissections showed that seeds may remain in the gizzard for up to 24 h (Heintze, 1918, pp. 34–35). Some northern populations of Hooded Crow still undertake seasonal migrations.
The likely dispersal distances identified by Heintze a century ago are remarkably similar to known movement distances in present-day corvids in northern Europe (see also Cramp and Perrins, 1994; e.g., Wernham et al., 2002; Fransson and Hall-Karlsson, 2008; Czarnecka and Kitowski, 2010; Pesendorfer et al., 2016; Marchand et al., 2018). This confirms that corvids are important vectors for long-distance dispersal of plants, and offer a maximum dispersal distance far greater than is generally recorded for other mechanisms including epizoochory, anemochory, and obviously barochory (Bullock et al., 2017).
Corvids can be important vectors for alien species (Nogales et al., 1999), especially in urban environments (Czarnecka et al., 2013). Interestingly, Heintze showed they were already dispersing numerous alien plants a century ago. Heintze also stated that Magpies were responsible for spreading “our cultivated garden trees, bushes and herbs with berries into the wild” (1917a, p. 230). Furthermore, many of the native species dispersed by corvids in Europe by non-classical endozoochory are alien species in other continents, where they may be spread by corvids in a manner not predicted by diaspore morphology. For example, the Hemp-nettle Galeopsis tetrahit has a dry achene that was recorded from six corvid species by Heintze (Table S1), was particularly frequent in Magpie pellets (Table 2), and is a widespread and problematic alien in North America (USDA and NRCS, 2019).
Seed Size
Plants with the largest seeds can be expected to be dispersed less often by corvid endozoochory since they are more likely to be destroyed in the foregut, and also because of the general negative relationship between seed size of a given species and the numbers of seeds produced by individual plants (Bruun and Poschlod, 2006; Green et al., 2016). Comparing the seed size distribution for all plants dispersed by corvid endozoochory (Figure 5) with the relative abundance of seeds dispersed by Magpies and Hooded Crows (Figure 6) does provide indirect support for a greater dispersal frequency of smaller seeds. Heintze (1917a, 1918) suggested that larger seeds were more likely to be egested and dispersed via pellets and smaller ones in the feces. Heintze stated that “all larger, most middle-sized, and more than half of the small-sized seeds are spread hemiendozoically [i.e., in pellets], whilst the rest comes out in the excrements” (1918, p. 30). This would be consistent with expectations based on gut morphology, since the sphincter passage between the gizzard and intestines should make it harder for larger seeds to pass intact. It is also consistent with literature for other birds, such as gulls and shorebirds, in which larger seeds were egested in pellets (Lovas-Kiss et al., 2018b, 2019). However, such a clear pattern is not supported from our analyses of Heintze's dataset for Magpies and Hooded Crows (Figure 6).
Heintze suggested (at least for Magpie and Hooded Crow) that seed dispersal via regurgitated pellets was more important than seed dispersal via feces (Heintze, 1917a, p. 228). We found evidence that seeds are more frequent in pellets than in feces, especially for Hooded Crows. However, rates of endozoochory events depend on the rate of egestion, and it is likely that each pellet sample contains a higher proportion of the daily regurgitated output than each feces sample contains of daily fecal output. Some birds produce <1 pellet a day (e.g., White Storks, Rosin and Kwiecinski, 2011). Based on dissections, Heintze concluded that seeds may remain in the gizzard of a Hooded Crow for up to 24 h (Heintze, 1918, pp. 34–35), suggesting they produce at least one pellet a day.
Comparison Between Corvids and Waterfowl
Owing to the focus on classical endozoochory in the literature, little information is available about the importance of non-classical endozoochory by other granivorous birds so as to enable a comparison with corvids. The best known vectors of non-classical endozoochory are the waterfowl Anatidae (Green et al., 2016). In particular, the dabbling ducks Anas spp. are likely to disperse over 500 plant species in Europe, and only a small fraction of seeds they disperse have an endozoochory syndrome (Soons et al., 2016; Lovas-Kiss et al., 2018a). Interestingly, there is a considerable overlap in plant species dispersed by corvids and waterfowl, mainly being cases of non-classical endozoochory (see Table S1 for full details). Of the 157 plant species listed by Heintze as dispersed by corvid endozoochory, 56 are also thought to be dispersed by dabbling ducks (Soons et al., 2016), and 20 were listed as dispersed by Greylag Geese Anser anser in the Stockholm archipelago in Sweden (Jerling et al., 2001). We found corvids to disperse more seeds from large size categories (most of which are of fleshy-fruited plants) than dabbling ducks (see Figure 2 in Soons et al., 2016). Plant species common to corvid and waterfowl endozoochory are terrestrial species. Not surprisingly, the distributions of Ellenberg moisture values differ among the plant taxa and seeds dispersed by corvids and dabbling ducks, because ducks disperse many moist soil and aquatic taxa with an Ellenberg value of 9–12 (see Figure 3 in Soons et al., 2016).
Conclusions
Based on the work of others and his own extensive research Heintze concluded (1917a, p. 210) that “It appears that within these zoogeographic areas [the Holarctic and the Neoboreal] corvids are the most important seed dispersers among all landbirds.” This article lends support to this statement, given the demonstrated importance of corvids as vectors for an extensive diversity of plants dispersed by both classical and non-classical endozoochory. Much has changed in European habitats over the past century, and it would be interesting to conduct similar research again in the same parts of Sweden where Heintze collected his own data.
It is unfortunate that the work of Heintze and co-workers a century ago or before has been overlooked for so long. The scientific community has effectively lost their understanding about the role of corvids as seed dispersers for a very wide range of terrestrial plants, and we hope our paper helps to restore this knowledge. The effort and knowledge required by Heintze to retrieve and study thousands of pellets and fecal samples is awe-inspiring, and should humble us all to pay more attention to the work of such pioneers. Increasing access to digitalized older literature is making this easier. His work also illustrates the immense value that natural history plays in ecology, and the serious consequences of its current decline (Tewksbury et al., 2014).
Many corvid species are likely to be vectors for a broad range of plants through both frugivory and granivory because they are opportunistic, generalist foragers able to exploit a range of habitats in both natural and human-created landscapes. In urban environments, Magpies, Jackdaws and other species can be amongst the most abundant birds. Corvids worldwide may be one of the most important bird groups in providing a vital seed dispersal service that maintains and restores a wide range of plants with a variety of growth forms and other traits, and in forested and open landscapes, as well as urban habitats. Corvids are heavily hunted in many countries, and this may negatively impact their important role as native plant vectors. On the other hand, corvids also have a role in unwanted spread of alien plants.
As shown previously for waterfowl, shorebirds and gulls (Jerling et al., 2001; Lovas-Kiss et al., 2018a,b, 2019), non-classical endozoochory by corvids is a major dispersal process that requires more attention if we are to fully understand plant dispersal processes and plant-animal interactions. We hope this paper will inspire others to look beyond frugivory and synzoochory and pay more attention to the importance of non-classical endozoochory by corvids, as well by other little-studied granivorous birds, such as galliformes and small passerines (Swank, 1944; Orlowski et al., 2016).
Author Contributions
AG, JE, and ÁL-K conceived the study. JE and ÁL-K collected the data. AG and ÁL-K analyzed the data. AG, JE, and ÁL-K wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
AG was supported by Spanish National Plan project CGL2016-76067-P (AEI/FEDER, EU). Peter Bengtsson at Kristianstad University library kindly provided bibliographic assistance. ÁL-K was supported by NKFIH FK-127939 project. We acknowledge support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00133/full#supplementary-material
Table S1. Complete list of plant taxa whose seeds are dispersed by endozoochory according to data in Heintze (1917a,b); Heintze (1918).
Table S2. Complete list of plant taxa recorded as seedlings in the field within remnants of pellets of Magpies and Hooded Crows, and considered by Heintze to have been dispersed within those pellets.
References
Bai, B., Li, N., Li, X. H., and Lu, C. H. (2017). Bulbuls and crows provide complementary seed dispersal for China's endangered trees. Avian Res. 8:31. doi: 10.1186/s40657-017-0089-y
Barrows, W. B. (1888). The food of crows. U.S. Dept. Agri. Report. 1888, 498–535. Available online at: https://ia801309.us.archive.org/35/items/CAT30951786026/ros1888.pdf
Birkhead, T. (2010). The Magpies: The Ecology and Behaviour of Black-Billed and Yellow-Billed Magpies. A&C Black.
Bruun, H. H., and Poschlod, P. (2006). Why are small seeds dispersed through animal guts: large numbers or seed size per se? Oikos. 113, 402–411. doi: 10.1111/j.2006.0030-1299.14114.x
Bullock, J. M., Gonzalez, L. M., Tamme, R., et al. (2017). A synthesis of empirical plant dispersal kernels. J Ecol. 105, 6–19. doi: 10.1111/1365-2745.12666
Cramp, S., and Perrins, C. M. (Eds.). (1994). The Birds of the Western Palearctic, Volume 8: Crows to Finches. Oxford: Oxford University Press.
Czarnecka, J., and Kitowski, I. (2010). Seed dispersal by the rook corvus frugilegus l. In agricultural landscape–mechanisms and ecological importance. Pol. J. Ecol. 58, 511–523.
Czarnecka, J., and Kitowski, I. (2013). Rook spring seed dispersal in the agricultural landscape–frugivory, granivory or accidental transport? Folia Geobot. 48, 55–73. doi: 10.1007/s12224-012-9134-0
Czarnecka, J., Kitowski, I., Sugier, P., et al. (2013). Seed dispersal in urban green space–does the rook Corvus frugilegus l. Contribute to urban flora homogenization? Urban Urban Gree. 12, 359–366. doi: 10.1016/j.ufug.2013.03.007
Figuerola, J., Green, A. J., and Santamaria, L. (2002). Comparative dispersal effectiveness of wigeongrass seeds by waterfowl wintering in south-west spain: quantitative and qualitative aspects. J. Ecol. 90, 989–1001. doi: 10.1046/j.1365-2745.2002.00734.x
Fransson, T., and Hall-Karlsson, S. (2008). Svensk ringmärkningsatlas. Vol. 3. [Swedish Bird Ringing Atlas. Vol 3.]. Stockholm: Naturhistoriska Riksmuseet & Sveriges Ornitologiska Förening.
Green, A.J., Brochet, A.L., Kleyheeg, E., and Soons, M.B. (2016). “Dispersal of plants by waterbirds,” in Why Birds Matter: Avian Ecological Function and Ecosystem Services, eds C. H. Sekercioglu, D. G. Wenny, and C. J. Whelan (Chicago: University of Chicago Press), 147–195.
Hadjisterkotis, E. (2003). The effect of corvid shooting on the populations of owls, kestrels and cuckoos in Cyprus, with notes on corvid diet. Z. Jagdwiss. 49, 50–60. doi: 10.1007/BF02192013
Heintze, A. (1917a). Om endo- och synzoisk fröspridning genom europeiska kråkfåglar [On endo- and synzoic seed dispersal by European corvids Part 1.]. Bot. Notiser. 1917, 209–240.
Heintze, A. (1917b). Om endo- och synzoisk fröspridning genom europeiska kråkfåglar [On endo- and synzoic seed dispersal by European corvids Part 2.]. Bot. Notiser. 1917, 297–300.
Heintze, A. (1918). Om endo- och synzoisk fröspridning genom europeiska kråkfåglar [On endo- and synzoic seed dispersal by European corvids Part 3.]. Bot. Notiser. 1918, 1–47.
Heintze, A. (1932). Handbuch der Verbreitungsökologie der Pflanzen. Lieferung 1. [Handbook of Dispersal Ecology of Plants. Part 1.]. Stockholm.
Heintze, A. (1935). Handbuch der Verbreitungsökologie der Pflanzen. Lieferung 2. [Handbook of Dispersal Ecology of Plants. Part 2.]. Stockholm.
Higgins, S. I., Nathan, R., and Cain, M. L. (2003). Are long-distance dispersal events in plants usually caused by nonstandard means of dispersal? Ecology. 84, 1945–1956. doi: 10.1890/01-0616
Hill, M. O., Mountford, J. O., Roy, D. B., and Bunce, R. G. H. (1999). Ellenberg's Indicator Values for British Plants. ECOFACT Volume 2 Technical Annex (Vol. 2). Huntingdon: Institute of Terrestrial Ecology.
Holmberger, P. (1785). Theorie om Växternas kringspridande på Jorden [Theory on the dispersal of plants over the world]. Kongliga Götheborgska Wetenskaps och Witterhets Samhällets handlingar 3, 7–26.
Holmboe, J. (1900). Notizen über die endozoische Samenverbreitrung der Vögel [Notes about endozoic seed dispersal in birds]. Nyt Mag. Nat. Videnska. 38, 303–320. Den physiografiske forening i Christiania. Christiania [Oslo].
Jaganathan, G. K., Yule, K., and Liu, B. L. (2016). On the evolutionary and ecological value of breaking physical dormancy by endozoochory. Perspect. Plant Ecol. 22, 11–22. doi: 10.1016/j.ppees.2016.07.001
Jerling, L., Löfgren, A., and Lannek, J. (2001). Växtlivet i Stockholms skärgård: mönster i tid och rum. [Plant life in the Stockholm archipelago: patterns in time and space]. Svensk Bot. Tidskr. 95, 212–226.
Julve, P. (1998). Baseflor. Index botanique, écologique et chorologique de la flore de France. Lille: Institut Catholique de Lille. Retrieved from: http://perso.wanadoo.fr/philippe.julve/catminat.htm (accessed September 25, 2018).
Kleyer, M., Bekker, R. M., Knevel, I. C., Bakker, J. P., Thompson, K., Sonnenschein, M., et al. (2008). The LEDA traitbase: a database of life-history traits of Northwest European flora. J. Ecol. 96, 1266–1274. doi: 10.1111/j.1365-2745.2008.01430.x
Lovas-Kiss, Á., Sanchez, M.I., Wilkinson, D.M., Coughlan, N. E., Alves, J. A., and Green, A. J. (2019). Shorebirds as important vectors for plant dispersal in Europe. Ecography. doi: 10.1111/ecog.04065
Lovas-Kiss, A., Sanchez, M. I., Molnar, V. A., Valls, L., Armengol, X., Mesquita-Joanes, F., et al. (2018b). Crayfish invasion facilitates dispersal of plants and invertebrates by gulls. Freshw. Biol. 63, 392–404. doi: 10.1111/fwb.13080
Lovas-Kiss, A., Vizi, B., Vincze, O., Molnar, V. A., and Green, A. J. (2018a). Endozoochory of aquatic ferns and angiosperms by mallards in central Europe. J. Ecol. 106, 1714–1723. doi: 10.1111/1365-2745.12913
Marchand, P., Loretto, M. C., Henry, P. Y., Duriez, O., Jiguet, F., Bugnyar, T., et al. (2018). Relocations and one-time disturbance fail to sustainably disperse non-breeding common ravens Corvus corax due to homing behaviour and extensive home ranges. Eur. J. Wildlife Res. 64:57. doi: 10.1007/s10344-018-1217-7
Marzluff, J. M., and Angell, T. (2007). In the Company of Crows and Ravens. New Haven, CT: Yale University Press.
McPartland, J. M., and Naraine, S. G. (2018). Experimental endozoochory of Cannabis sativa achenes. Med. Cannabis Cannabinoids. 1, 69–103. doi: 10.1159/000492971
Nathan, R., Schurr, F. M., Spiegel, O., Steinitz, O., Trakhtenbrot, A., and Tsoar, A. (2008). Mechanisms of long-distance seed dispersal. Trends Ecol. Evol. 23, 638–647. doi: 10.1016/j.tree.2008.08.003
Nogales, M., Heleno, R., Traveset, A., and Vargas, P. (2012). Evidence for overlooked mechanisms of long-distance seed dispersal to and between oceanic islands. New Phytol. 194, 313–317. doi: 10.1111/j.1469-8137.2011.04051.x
Nogales, M., Hernandez, E. C., and Valdes, F. (1999). Seed dispersal by common ravens corvus corax among island habitats (Canarian archipelago). Ecoscience. 6, 56–61. doi: 10.1080/11956860.1999.11952193
Orlowski, G., Czarnecka, J., Golawski, A., Karg, J., and Panek, M. (2016). The effectiveness of endozoochory in three avian seed predators. J. Ornithol. 157, 61–73. doi: 10.1007/s10336-015-1248-8
Perez-Harguindeguy, N., Diaz, S., Garnier, E., Lavorel, S., Poorter, H., Jaureguiberry, P., et al. (2013). New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 61, 167–234. doi: 10.1071/BT12225
Pesendorfer, M. B., Sillett, T. S., Koenig, W. D., and Morrison, S. A. (2016). Scatter-hoarding corvids as seed dispersers for oaks and pines: a review of a widely distributed mutualism and its utility to habitat restoration. Condor. 118, 215–237. doi: 10.1650/CONDOR-15-125.1
R Core Team. (2017). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: https://www.R-project.org/.
Rosin, Z. M., and Kwiecinski, Z. (2011). Digestibility of prey by the White Stork (Ciconia ciconia) under experimental conditions. Ornis Fennica. 88, 40–50. doi: 10.3184/175815515X14232310459990
Sekercioglu, C. H. (2006). Increasing awareness of avian ecological function. Trends Ecol. Evol. 21, 464–471. doi: 10.1016/j.tree.2006.05.007
Soons, M. B., Brochet, A. L., Kleyheeg, E., and Green, A. J. (2016). Seed dispersal by dabbling ducks: an overlooked dispersal pathway for a broad spectrum of plant species. J. Ecol. 104, 443–455. doi: 10.1111/1365-2745.12531
Swank, W. G. (1944). Germination of seeds after ingestion by ring-necked pheasants. J. Wildlife Manage. 8, 223–231. doi: 10.2307/3795702
Tewksbury, J. J., Anderson, J. G., Bakker, J. D., Billo, T. J., Dunwiddie, P. W., Groom, M. J., et al. (2014). Natural history's place in science and society. Bioscience. 64, 300–310. doi: 10.1093/biosci/biu032
Tomback, D. F. (2016). “Seed dispersal by corvids: Birds that build forests,” in Why Birds Matter, eds C. H. Sekercioglu, D. G. Wenny, and C. J. Whelan (Chicago, IL: University of Chicago Press), 196–234.
Twigg, L. E., Lowe, T. J., Taylor, C. M., Calver, M. C., Martin, G. R., Stevenson, C., et al. (2009). The potential of seed-eating birds to spread viable seeds of weeds and other undesirable plants. Austral. Ecol. 34, 805–820. doi: 10.1111/j.1442-9993.2009.01992.x
USDA NRCS. (2019). The PLANTS Database. Greensboro, NC: National Plant Data Team. Available online at: http://plants.usda.gov (accessed February 22, 2019).
van der Pijl, L. (1982). Principles of Dispersal in Higher Plants. 3rd Edn. Berlin: Springer-Verlag.
Vander Wall, S. B., and Longland, W. S. (2004). Diplochory: are two seed dispersers better than one? Trends Ecol. Evol. 19, 155–161. doi: 10.1016/j.tree.2003.12.004
Wenny, D. G., Sekercioglu, C., Cordeiro, N. J., Rogers, H. S., and Kelly, D. (2016). “Seed dispersal by fruit-eating birds,” in Why Birds Matter: Avian Ecological Function and Ecosystem Services, eds C. H. Sekercioglu, D. G. Wenny, and C. J. Whelan (Chicago: University of Chicago Press), 107–145.
Keywords: Corvidae, gut-passage, Hooded Crow, Magpie, non-classical endozoochory, non-standard dispersal, pellets, seed dispersal
Citation: Green AJ, Elmberg J and Lovas-Kiss Á (2019) Beyond Scatter-Hoarding and Frugivory: European Corvids as Overlooked Vectors for a Broad Range of Plants. Front. Ecol. Evol. 7:133. doi: 10.3389/fevo.2019.00133
Received: 03 January 2019; Accepted: 03 April 2019;
Published: 18 April 2019.
Edited by:
Anouschka R. Hof, Wageningen University & Research, NetherlandsReviewed by:
Wolfgang Fiedler, Max Planck Institute of Ornithology, GermanyJoanna Czarnecka, Maria Curie-Sklodowska University, Poland
Mario B. Pesendorfer, Cornell Lab of Ornithology, United States
Copyright © 2019 Green, Elmberg and Lovas-Kiss. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andy J. Green, YWpncmVlbkBlYmQuY3NpYy5lcw==