 Victor Manuel Bravo-Cuevas
Victor Manuel Bravo-Cuevas Eduardo Jiménez-Hidalgo
Eduardo Jiménez-Hidalgo- 1Museo de Paleontología, Área Académica de Biología, Universidad Autónoma del Estado de Hidalgo, Ciudad del Conocimiento, Pachuca, Mexico
- 2Laboratorio de Paleobiología, Instituto de Recursos, Campus Puerto Escondido, Universidad del Mar, Oaxaca, Mexico
North American Equinae integrates a monophyletic clade that consists of about 20 genera and 80 species. This group includes horses with hypsodont cheek teeth belonging to the merychippines and the tribes Hipparionini, Protohippini, and Equini. The primary adaptive radiation and diversification of Equinae occurred in the Neogene period (23.03–2.53 Ma). In Mexico, 11 genera and 20 species of Neogene Equinae have been reported, which correspond to 55 and 28% of the generic and specific diversity in North America, respectively. The Mexican record exhibited their major diversity during the late Miocene (10 species) and evidences part of the evolutionary history of North American Equinae, as it is stated in the following considerations: (1) The presence of merychippine species from the late Hemingfordian—early Barstovian (18–15 Ma) of southern Mexico (“Merychippus” cf. primus and “M.” cf. sejunctus) that are related with earliest representatives of Equinae in North America. (2) The occurrence of populations referable to Cormohipparion aff. quinni, Calippus sp., and Pliohippus sp. from the early—late Barstovian (15–14 Ma) of southern Mexico, which are synchronous with the first known appearances of those genera from the Great Plains and Gulf Coastal Plain in the United States. (3) The equine horse Dinohippus mexicanus from the late Hemphillian (4.8 Ma) of central and northern Mexico that is considered the closest sister species of primitive Equus. These records suggest that early differentiation of some hipparionines, protohippines, and equines may have had occurred also in areas of southern tropical North America during the middle Miocene; furthermore, the origin of primitive Equus could be traced from the Mexican record.
Introduction
Horses were one of the most common components of the Cenozoic faunas of North America (MacFadden, 1992, 1998). Because of their significant occurrence in the fossil record, they have been considered as a suitable model to explain rates and patterns of evolution (MacFadden, 1988, 1992; MacFadden and Hulbert, 1988; Carroll, 1998). These perissodactyls experienced one of their major adaptive radiations during the Miocene (18–15 Ma), resulting in the differentiation of members that comprise the subfamily Equinae (Hulbert and MacFadden, 1991).
The subfamily Equinae is defined as a group of subhypsodont to hypsodont horses (unworn molar crown height ≥25 mm), consisting of a set of paraphyletic species traditionally referred as the merychippines, as well as the monophyletic hipparionines (Hipparionini), protohippines (Protohippini) and equines (Equini) (Hulbert and MacFadden, 1991; Hulbert, 1993; Kelly, 1995, 1998).
In North America, the Equinae spanned from the early Miocene to the late Pleistocene, having a geochronological duration of about 18 million years. They became extinct at about 11 ka and survived in the Old World, although Europeans brought them back to the Americas in the sixteenth century (MacFadden, 1992). Extant members belong to the genus Equus, and include horses, asses, and zebras (MacFadden, 1992; Franzen, 2010).
Most of the evolutionary history of North American Equinae occurred during the Neogene (23–2.6 Ma). This group of horses experienced important changes in their species diversity related to speciation and extinction processes (Hulbert, 1993). Most of what is known regarding the evolution of North American Equinae derives from the study of fossil samples recovered from numerous localities in the United States (MacFadden, 1984a, 1992; Hulbert, 1989; Hulbert and MacFadden, 1991; Kelly, 1995, 1998; Maguire and Stigall, 2008). A small number of studies have considered Mexican samples in order to understand evolutionary aspects of some members of Equinae (e.g., MacFadden, 1984b; MacFadden and Carranza-Castañeda, 2002; Ferrusquía-Villafranca et al., 2014).
The North American record of Neogene Equinae consists of 20 genera and at least 70 species (Maguire and Stigall, 2008). The Mexican record includes 11 genera and 20 species, evidencing 55 and 28% of the generic and specific diversity respectively, from North America (Priego-Vargas et al., 2016).
Given the significant representation of members of Equinae throughout the Mexican Neogene the aims of this study are to present an overview of the known record of this group of horses and to provide some assumptions on their taxonomic evolution. Furthermore, comments on the significance of the Mexican record in relation to the adaptive radiation of North American Equinae and the origin of Equus are also provided.
Materials and Methods
Taxonomic information and geographic distribution of Equinae from the Mexican Neogene were collected from the comprehensive review of the Cenozoic horses from Mexico by Priego-Vargas et al. (2016) (Table 1). The Mexican horse record is described in a biochronological sequence from the Hemingfordian to the Blancan North American Land Mammal Ages (NALMAs). NALMAs are from Tedford et al. (2004) and Bell et al. (2004).
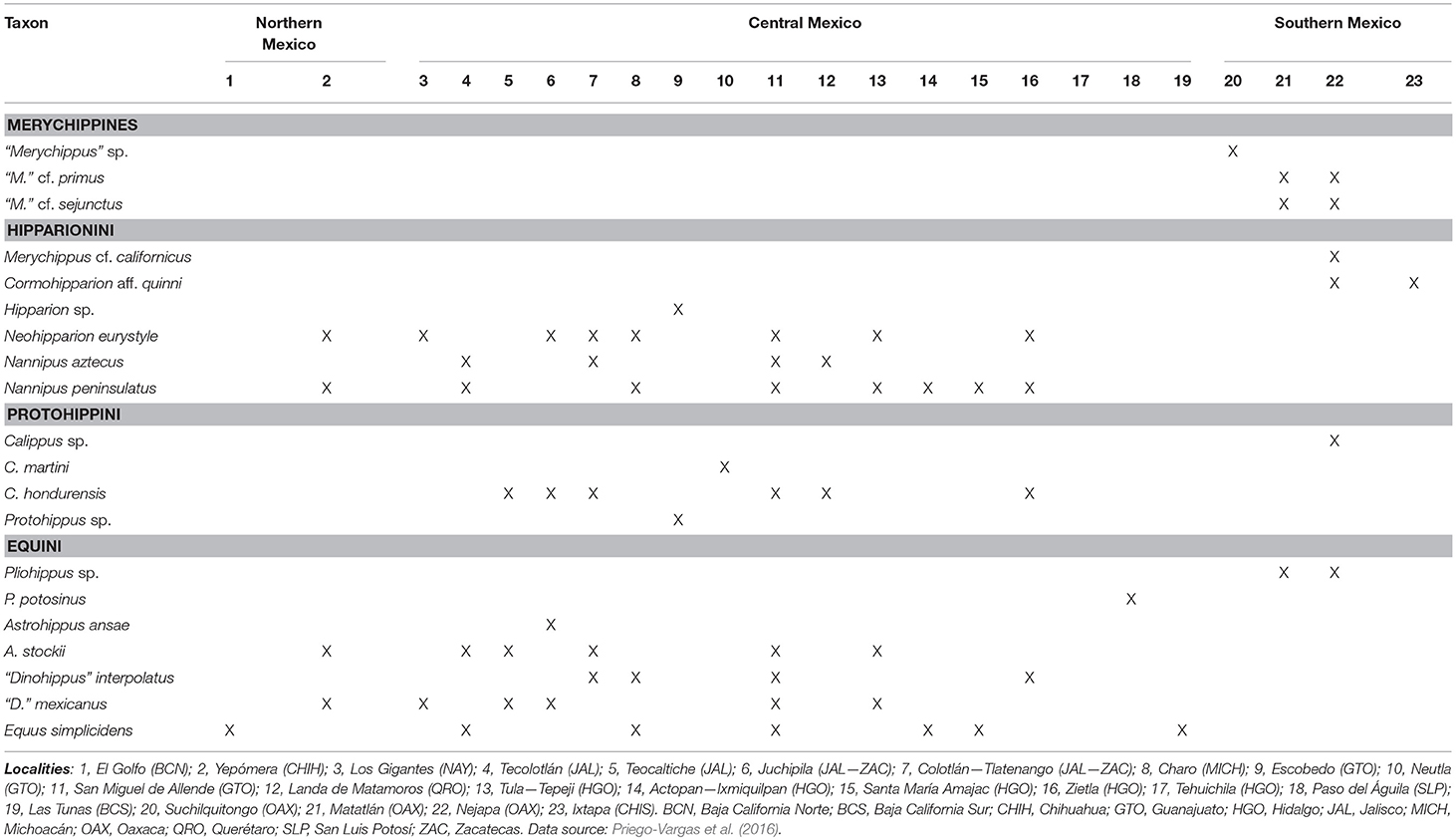
Table 1. List of localities that bear fossil material belonging to Equinae from the Mexican Neogene.
The pattern of species richness of the Mexican Equinae was described using a bar graph of the number of species reported throughout the Neogene. The change in the number of species has been related to the geology and climate of the Mexican Neogene. Geologic history is for the most part from Ortega-Gutiérrez et al. (1992) and Ferrusquía-Villafranca (1993), whereas climatic history is from González-Medrano (1998).
An approach of the taxonomic evolution of Equinae from the Mexican Neogene is described in terms of the chronological range of selected taxa (Hulbert, 1993; Carroll, 1998). The chronological range is defined by the first and last appearances of a particular horse species in the Mexican fossil record, considering its occurrence in all localities where it has been reported. It should be stated that species reported from a single locality were removed from this approach because cannot provide a suitable estimation of its temporal range (Robinson et al., 2018). The species “Merychippus” sp. from the Hemingfordian of Oaxaca and the majority of the protohippines have been not considered given that are known from single localities. It is noted that Calippus hondurensis is the well-known and documented protohippine from the late Miocene of central Mexico (Carranza-Castañeda, 2006; Carranza-Castañeda et al., 2013).
Temporal ranges of the selected species are derived from the dating of some of the rock bearing-units and the biochronology of Mexican mammalian faunas that contain Neogene horses (Lindsay et al., 1984; Ferrusquía-Villafranca, 1992, 1996, 2001; Kowallis et al., 1999; Carranza-Castañeda, 2006; Carranza-Castañeda et al., 2013). The species considered comprise 70% of the known record of the Mexican Neogene horses (Table 2).
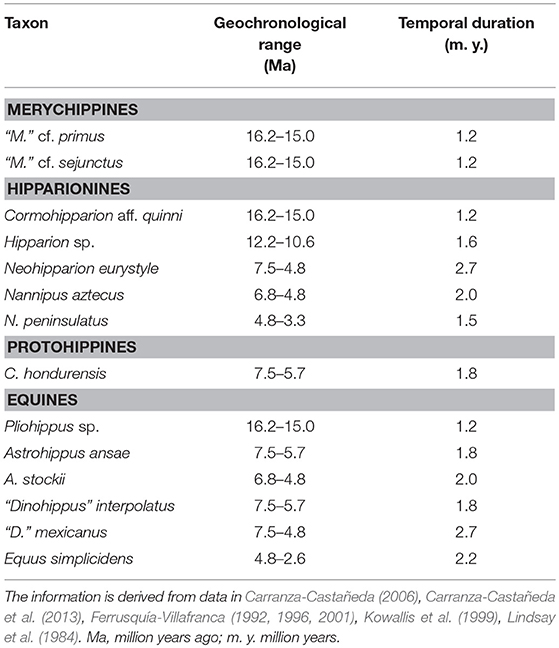
Table 2. Observed geochronological range and temporal duration of the selected species of Equinae from the Mexican Neogene.
We compared the temporal range of selected Mexican hipparionines and equines with that from the North American record. The comparison with the other groups of horses (merychippines and protohippines) is restricted given their information available from the Mexican Neogene. The temporal duration of the North American species is derived from the temporally calibrated cladogram of the Equinae of Maguire and Stigall (2008; Figure 1, p. 176) (Supplementary Data). A Mann-Whitney U-test was used to compare the temporal range of the Mexican and North American Equinae. This test was selected because the normality of the data is unwarranted. It should be stated that the results derived from the analysis represent a rough interpretation of the taxonomic evolution of the Equinae from the Mexican Neogene.
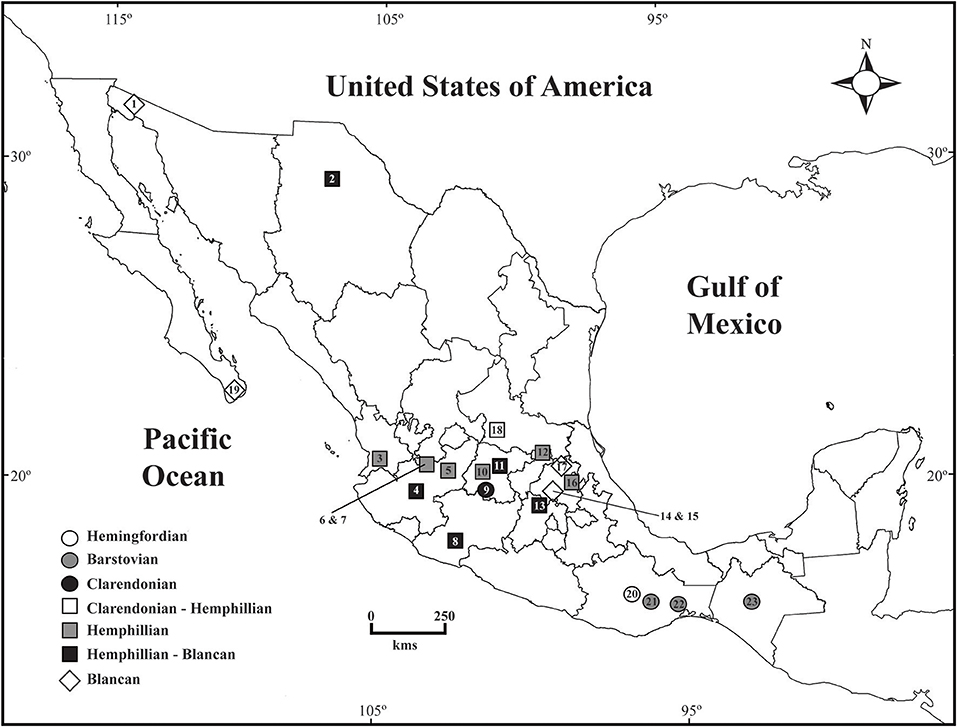
Figure 1. Geographic distribution of Mexican Equinae throughout the Neogene. The list of localities as in Table 1.
The interpretation of the species diversity and the taxonomic evolution of the Mexican Neogene horses is derived from the available information of this group of horses. It should be stated that the known record shows taxonomical, chronological, and geographical limitations. Taxonomically, there are an important number of taxa whose species identity is uncertain, such as the merychippines and the majority of the protohippines. Chronologically and geographically, there is a biased to the Hemphillian and Blancan faunas of central Mexico. Hence, considerations regarding these topics are susceptible to be modified and/or corroborated, as new evidence of Mexican Neogene horses is available. Nevertheless, this report represents the first attempt to provide information on the evolutionary significance of the Mexican Neogene horses.
The following terms have been used to refer to the groups of horses considered in the study:
Equinae.-A group of hypsodont horses consisting of merychippines, hipparionines, protohippines, and equines.
Merychippine.-A paraphyletic group of species referred as to “Merychippus,” including “Merychippus” gunteri, “M.” primus, “M.” sejunctus, “M.” intermontanus, “M.” coloradensis, “M.” republicanus, and “M.” goorisi.
Hipparionine.-A monophyletic group of species in the tribe Hipparionini, including Merychippus insignis, M. brevidontus, M. californicus, Neohipparion, Pseudhipparion, Hipparion, Nannipus, and Cormohipparion.
Protohippine.-A monophyletic group of species in the tribe Protohippini, including the genera Protohippus and Calippus.
Equine.-A monophyletic group of species in the Tribe Equini, including Acritohippus, Parapliohippus, Pliohippus, “Pliohippus,” Astrohippus, Dinohippus, “Dinohippus,” Equus, Onohippidion, and Boreohippidion.
Overview of Equinae From the Mexican Neogene
The record of Equinae from the Mexican Neogene consists of 20 species in 11 genera, including representatives of the merychippines, hipparionines, protohippines, and equines. The fossil sample that integrates this record includes skulls, mandibles, as well as numerous teeth and postcranial bones. The material has been recovered from about 23 areas unevenly distributed across the Mexican territory. Most of the sites are late Miocene to Pliocene in age and are distributed in central Mexico, except El Golfo (Baja California Norte) and Yepómera (Chihuahua) localities, northern Mexico. There are one early Miocene site and three middle Miocene sites in southern Mexico (Figure 1, Table 1).
Hemingfordian and Barstovian Mexican Horses
The horses of these faunal stages come from the Suchilquitongo, Matatlán, and Nejapa localities in Oaxaca, as well as the Ixtapa locality in Chiapas, all in southern Mexico. The record consists of seven species that comprise 35% of the Mexican Equinae, including the oldest members of this group of horses from the Neogene of Mexico and the southernmost occurrences of fossil populations referable to merychippines, hipparionines, protohippines, and equines from North America during the middle Miocene (Bravo-Cuevas, 2000; Bravo-Cuevas and Ferrusquía-Villafranca, 2006, 2008, 2010). The fossil bearing-units are part of the Suchilquitongo Formation (K-Ar and paleomagnetic dated in 18.2–19.6 Ma) (Ferrusquía-Villafranca, 1992; Urrutia-Fucugauchi and Ferrusquía-Villafranca, 2001), the Matatlán and El Camarón formations (both K-Ar dated 15.15 ± 0.8–16.35 ± 0.75 Ma), Oaxaca (Ferrusquía-Villafranca, 1992, 2001); as well as the Ixtapa Formation (K-Ar dated 15.02 ± 0.35–16.02 ± 0.53 Ma), Chiapas (Ferrusquía-Villafranca, 1996).
The Hemingfordian record consists of a single occurrence referable to “Merychippus” sp. from Suchilquitongo (Ferrusquía-Villafranca, 1990), which is (as far as we know) the oldest member of the Mexican Equinae. The sample includes a tooth fragment whose crown height and fossette plication degree are comparable to those observed in some Hemingfordian and Barstovian equines (e.g., Acritohippus tertius) and hipparionines (e.g., Merychippus californicus) (Ferrusquía-Villafranca, 1990).
Six Mexican species of Equinae are known during the Barstovian. The merychippines have been referred to “Merychippus” cf. primus and “Merychippus” cf. sejunctus from the Matatlán and Nejapa localities. The sample belonging to these species includes a mandibular fragment, a maxillary fragment, and several isolated teeth (Bravo-Cuevas and Ferrusquía-Villafranca, 2006).
“Merychippus” primus is known from the late Hemingfordian of Nebraska and Florida (Hulbert and MacFadden, 1991) and the early Barstovian of Florida (Bryant, 1991); it is one of the basal species of the North American Equinae (Hulbert, 1989; Hulbert and MacFadden, 1991). “Merychippus” sejunctus has been reported from the Barstovian of Colorado and Texas and it is a species that exhibits a combination of cranial and dental features related to the equines and hipparionines (Hulbert, 1989; Hulbert and MacFadden, 1991); however, recent phylogenetic information on the North American Equinae, places this merychippine as ancestral to the Protohippini—Hipparionini clade (Maguire and Stigall, 2008). The coexistence of related species of ancestral Equinae in the middle Miocene of Oaxaca, suggests that part of the differentiation of the advanced hypsodont horses also took place in southern areas of tropical North America.
The Barstovian record of hipparionines includes Merychippus cf. californicus from Nejapa and Cormohipparion aff. quinni from Nejapa and Ixtapa. Sets of isolated teeth and several postcranial remains have been referred to these species (Bravo-Cuevas, 2000; Bravo-Cuevas and Ferrusquía-Villafranca, 2008).
Merychippus insignis [Barstovian of Nebraska, Texas, Colorado, and California (Hulbert and MacFadden, 1991)], M. brevidontus [Barstovian of California (Kelly and Stewart, 2008)], and M. californicus [Barstovian of California (Downs, 1961; Kelly and Stewart, 2008)] are among the basal species of the Hipparionini (Maguire and Stigall, 2008). Therefore, Merychippus cf. californicus from the Barstovian of Oaxaca should be considered as one of the earliest hipparionines in North America.
Cormohipparion quinni is one of the oldest members of the genus and it is known from the late Barstovian (14–13 Ma) of Nebraska and Colorado (MacFadden, 1984a; Hulbert, 1987; Woodburne, 1996), together with C. sphenodus from the early late Barstovian (15–14 Ma) of northeastern Colorado (MacFadden, 1984a). Cormohipparion aff. quinni would extend the temporal range of C. quinni, its probable closest species, to the early late Barstovian (Bravo-Cuevas and Ferrusquía-Villafranca, 2008).
By the Barstovian, protohippines include Calippus sp. from Nejapa, which is characterized by having a crown height similar to that of the small-sized Calippus [= C. (Calippus) sensu (Hulbert, 1988a)], but with an occlusal pattern comparable to that of the medium-sized Calippus (= C. (Gramohippus) sensu Hulbert, 1988a; Bravo-Cuevas, 2000). The genus Calippus is recognized from the early late Barstovian to the late early Hemphillian (ca. 14.5–6 Ma) (Hulbert, 1988a). Calippus sp. is relatively synchronous to C. circulus from the middle late Barstovian (13.5–12.5 Ma) of Texas (Hulbert, 1988a), which is one of the most primitive species of the genus (Maguire and Stigall, 2008).
Barstovian equines include several isolated teeth assigned to Pliohippus sp. from Matatlán and Nejapa (Bravo-Cuevas and Ferrusquía-Villafranca, 2010). The occlusal pattern of this species resembles that of P. mirabilis from the late early Barstovian–late Barstovian (ca. 12–15 Ma) of Nebraska, Colorado, and Florida (Hulbert and MacFadden, 1991; Kelly, 1998), which is considered as the oldest and most plesiomorphic species of the genus (Kelly, 1998).
Cormohipparion aff. quinni, Calippus sp., and Pliohippus sp. had a biochronological range and geographic distribution that place these records among the most primitive populations of each genus, extending their geographic distribution from the Great Plains and the Gulf Coast to southern Mexico during the middle Miocene.
Clarendonian Mexican Horses
Clarendonian horses have been recently discovered from Escobedo, Guanajuato and Paso del Águila, San Luis Potosí, central Mexico (Robles-Rivera and Carranza-Castañeda, 2012; Ferrusquía-Villafranca et al., 2014; Robles-Rivera, 2015). The record includes three species that comprise 15% of the Mexican Equinae. The fossil horse sample from Escobedo comes from a fossil-bearing unit that underlies a sequence of andesitic lava flows Ar40/Ar39 dated 12.2 ± 04–10.6 ± 0.3 (Robles-Rivera, 2015). The fossil-bearing strata of El Paso de Águila are part of the San Nicolas Formation, whose age is bracketed between 12.33 and 7.41 Ma by the Ar40/Ar39 dating of volcanic ashes above and below the fossiliferous strata (Ferrusquía-Villafranca et al., 2014).
Isolated teeth have been referred to Protohippus sp. and Hipparion sp. from Escobedo, Guanajuato (Robles-Rivera and Carranza-Castañeda, 2012; Robles-Rivera, 2015). To extent of our knowledge, these are the only mentions of these genera from the Mexican Neogene. Members of Protohippus and Hipparion were more common in the United States, spanning from the late Barstovian to the early Hemphillian, whose records are known from different sites in the Great Plains, the Great Basin, the Gulf Coast, and the Coast of California (MacFadden, 1984a; Hulbert, 1988a,b). The protohippines and hipparionines from the Clarendonian of central Mexico evidence that representatives of Protohippus and Hipparion reached areas of southern temperate North America by the early late Miocene.
A rostral fragment has been assigned to a new species named as Pliohippus potosinus from El Paso del Águila, San Luis Potosí (Ferrusquía-Villafranca et al., 2014). This species is only known from the Mexican territory, exhibiting a cranial and dental configuration similar to that of Pliohippus fossulatus from the Clarendonian of Texas (Kelly, 1998). Pliohippus potosinus together with Pliohippus sp. from the Barstovian of Oaxaca are the only known representatives of the genus from the Mexican Neogene, spanning from the late early Barstovian to the earliest Hemphillian; a similar biochronological range for Pliohippus is reported from the United States (Kelly, 1998).
Hemphillian and Blancan Mexican Horses
The greatest number of fossil horses from the Mexican Neogene is from Hemphillian and/or Blancan localities. The record encompasses 50% of the known Mexican Equinae, including three hipparionines, two protohippines, and five equines. An important number of fossil-bearing localities in central Mexico have been dated by fission track, 40Ar/39Ar, and/or U/Pb radiometric techniques. Several localities in the San Miguel de Allende Basin, Guanajuato, have yielded ages ranging from about 4.8 to 3.9 Ma (Kowallis et al., 1999), whereas some localities in the states of Jalisco and Nayarit yield ages of 4.89 ± 0.16 Ma and 5.50 ± 0.05 Ma respectively (Carranza-Castañeda, 2006; Carranza-Castañeda et al., 2013). Recently, the dating of localities at the Juchipila Basin in Zacatecas indicates dates from 6.95 ± 0.27 to 5.59 ± 0.11 Ma (Carranza-Castañeda et al., 2013). The age of other localities in the states of Baja California, Hidalgo, Michoacán, and Querétaro is established by means of biochronology, indicating Hemphillian and Blancan faunas (Carranza-Castañeda, 2006). Particularly, the Yepómera locality in Chihuahua is placed between the magnetic chrons C3n2r and C3n2n (Gilbert Time), thus evidencing an age of about 4.6 Ma (Lindsay et al., 1984).
The species Neohipparion eurystyle and Nannipus aztecus were the typical hipparionines in the Hemphillian of Mexico, whereas N. peninsulatus was Blancan. These horses are represented by an important sample of cranial, dental, and/or postcranial remains (Priego-Vargas et al., 2016).
Neohipparion eurystyle is the best-known and well-documented Mexican hipparionine, which has been recorded through a major part of the Hemphillian (7.5–4.8 Ma) and had a wide distribution across the country (Stirton, 1955; Carranza-Castañeda, 2006; Carranza-Castañeda et al., 2013). This species exhibited the greatest temporal range (of about 2.7 million years) of all hipparionines from the Miocene and Pliocene of Mexico.
The genus Nannipus includes the late Hemphillian N. aztecus and the Blancan N. peninsulatus. Their record mainly comes from localities in central Mexico (Carranza-Castañeda, 2006; Carranza-Castañeda et al., 2013); however, there is evidence of both species from the Yepómera locality, Chihuahua, northern Mexico (Lance, 1950; MacFadden, 2006). The type localities of these hipparionines are in Mexico, including Rancho El Ocote in Guanajuato for N. aztecus (Mooser, 1968) and Tehuichila in Hidalgo for N. peninsulatus (Osborn, 1918). Nannipus aztecus has been considered as the putative ancestral species of the Blancan N. beckensis and N. peninsulatus of North America (MacFadden, 1984a; Hulbert, 1993). N. aztecus and N. peninsulatus are chronologically successive at several localities in the Tecolotlán Basin, Jalisco and San Miguel de Allende Basin, Guanajuato (Carranza-Castañeda, 2006). It seems that differentiation, probably by anagenesis, of some populations of Blancan Nannipus could have had occurred in areas of central Mexico. It is noted that anagenetic differentiation of Blancan Nannipus through N. aztecus has been previously suggested (MacFadden, 2006).
Protohippines include members of Calippus belonging to C. martini and C. hondurensis; both species are mainly known from dental material. Of these two species, C. hondurensis is better known and has been reported from several late early Hemphillian to early late Hemphillian (7.5–5.7 Ma) localities across central Mexico (Carranza-Castañeda, 2006; Carranza-Castañeda et al., 2013). In contrast, C. martini is represented by a single occurrence from the late early Hemphillian (7.5–6.8 Ma) of Guanajuato (Robles-Rivera, 2015). In the United States, C. martini is known from the early to middle Clarendonian (about 12.0 to 10.0 Ma) of the Great Plains and Gulf Coast, whereas C. hondurensis from the early Hemphillian (about 7.0–9.0 Ma) of Florida (Hulbert, 1988a). Hence, the Hemphillian records of these species in Mexico extend its biochronological range by about 3 million years, representing the youngest populations of North America.
Fossil material of equines includes skulls, mandibles, numerous teeth, and/or postcranial bones belonging to Astrohippus, Dinohippus, and Equus. The two species of Astrohippus from North America, namely A. ansae and A. stockii, have been reported in Mexico. The record of A. ansae consists of a single occurrence from the early Hemphillian of Zacatecas, central Mexico (Carranza-Castañeda et al., 2013). A. stockii is the most common species of the genus and it is known from several late Hemphillian localities in Chihuahua, Jalisco, Guanajuato, and Hidalgo (Lance, 1950; MacFadden, 1984b; Carranza-Castañeda, 2006).
Two species of “Dinohippus” are known from the Mexican Neogene, including “D.” interpolatus and “D.” mexicanus. The sample belonging to “D.” interpolatus comes from the early Hemphillian of Guanajuato, Hidalgo, and Jalisco (Carranza-Castañeda, 2006). The taxonomic status of this species is controversial, and it has been reported from the late early Hemphillian and early late Hemphillian (ca. 6.8 Ma) of Texas and California (Kelly, 1998 and references therein); other alleged records are from Colorado, Kansas, Oklahoma, Arizona, South Dakota, and New Mexico (MacFadden and Carranza-Castañeda, 2002). The Mexican record testifies that members of “D.” interpolatus reached areas of southern temperate North America during the late Miocene.
“Dinohippus” mexicanus is present in all the late Hemphillian faunas known from Mexico (Carranza-Castañeda et al., 2013). This species was originally described trough fossil material recovered from Yepómera, Chihuahua in northern Mexico (Lance, 1950). It has been shown that “D.” mexicanus gave rise to primitive species of Equus, such as E. simplicidens (Hulbert, 1989, 1993; MacFadden, 1992; MacFadden and Carranza-Castañeda, 2002).
Equus simplicidens is the youngest equine known from the Mexican Neogene. It is reported from several Blancan localities in Baja California, Guanajuato, Hidalgo, Michoacán, and Jalisco (Miller, 1980; Carranza-Castañeda and Miller, 1999; Carranza-Castañeda, 2006). This species is the oldest and most plesiomorphic of Equus and it is well-known from the early Blancan of Hagerman, Idaho and Broadwater, Nebraska (Azzaroli, 1992).
Diversity Changes in the Mexican Neogene Horses
The known Mexican horse record evidences a diversity that varied from one to eight contemporaneous species (Figure 2). Based on the available information, it appears that the greatest species diversity occurred during the early middle Miocene (16–14 Ma) and the latest Miocene (7–5 Ma) (Figure 3). The 70% of the total diversity of the Mexican Equinae is represented in these geochronological intervals. Hipparionines and equines were the most diverse with six and seven species respectively, whereas merychippines and protohippines were less diverse with three species in each group throughout the Neogene. Hipparionines and equines showed the greatest temporal range of about 13 million years followed by the protohippines with 8 million years and in a lesser extent the merychippines with 5 million years (Figure 2).
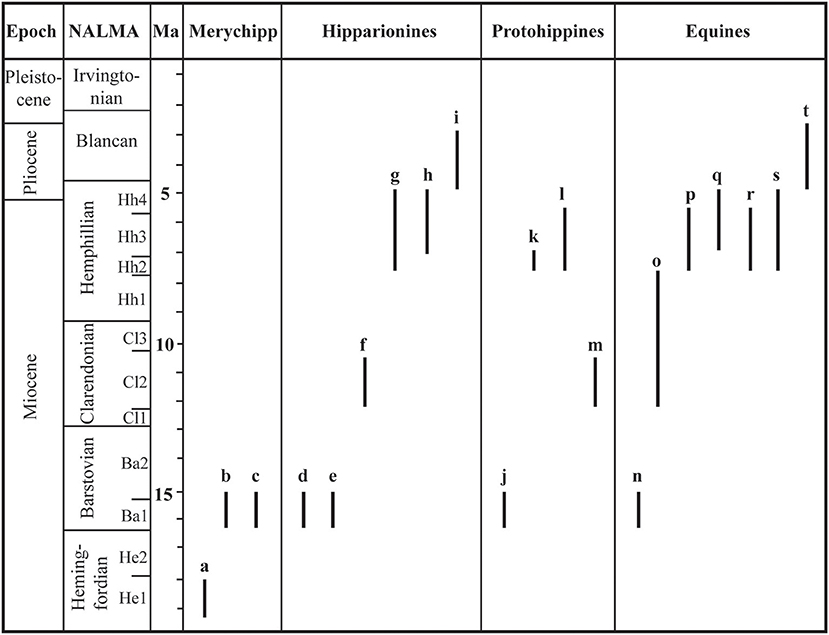
Figure 2. Temporally calibrated record of Equinae from the Mexican Neogene. Merychippines (Merychipp): a, “Merychippus” sp.; b, “M.” cf. primus; c, “M.” cf. sejunctus. Hipparionines: d, Merychippus californicus; e, Cormohipparion aff. quinni; f, Hipparion sp.; g, Neohipparion eurystyle; h, Nannipus aztecus; i, N. peninsulatus. Protohippines: j, Calippus sp.; k, C. martini; l, C. hondurensis; m, Protohippus sp. Equines: n, Pliohippus sp.; o, P. potosinus; p, Astrohippus ansae; q, A. stockii; r, “Dinohippus” interpolatus; s, “D.” mexicanus; t, Equus simplicidens. The first and last appearance datums are derived from Lindsay et al. (1984), Ferrusquía-Villafranca (1992, 1996, 2001), Kowallis et al. (1999), Urrutia-Fucugauchi and Ferrusquía-Villafranca (2001), Carranza-Castañeda (2006), Carranza-Castañeda et al. (2013), Ferrusquía-Villafranca et al. (2014), Robles-Rivera (2015). North American Land Mammal Ages (NALMAs) from Tedford et al. (2004).
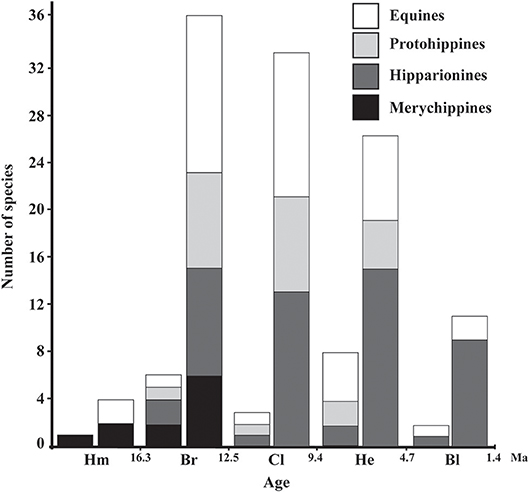
Figure 3. Pattern of species richness of Neogene Equinae from Mexico (left column) and North America (right column). Hm, Hemingfordian; Br, Barstovian; Cl, Clarendonian; He, Hemphillian; Bl, Blancan.
It seems that the species diversity of Mexican Equinae increased significantly from one species in the late Hemingfordian to six species in the late early Barstovian. In the late early Barstovian, merychippines and hipparionines were most diverse in comparison to the protohippines and equines. Then in the Clarendonian, merychippines are unknown and the other groups of Equinae included one species each.
In the late early Hemphillian it is observed the maximum species diversity known (so far) from the Mexican Neogene, including eight contemporaneous species. In this faunal stage, the equines were the most diverse with four species, whereas the protohippines are unknown. It is observed a significant reduction of species at the end of the Hemphillian, implying the loss of the 75% of the recorded species in this faunal stage. In the Blancan, one hipparionine and one equine species are observed.
The change in species diversity observed from the known record of the Mexican Neogene horses indicates two intervals of major diversity, the first at about 15 Ma (six species) and the second at about 7 Ma (eight species); later at about 5 Ma the group experienced an important reduction of species (two species). A similar pattern in species diversity is recovered from the record of the United States, showing a maximum of contemporary species at least twice during the Miocene (at 13.5 Ma and at 6.5 Ma) and a decline in species diversity by about 4.5 Ma (Figure 3). Hipparionines, protohippines, and most of the equines became extinct at about 2 Ma (Hulbert, 1993).
Diversity of Mexican Equinae in the Miocene
It has long been recognized that the evolution of North American Equinae was driven by environmental fluctuations during the Neogene (see Hulbert, 1993; Webb et al., 1995; Maguire and Stigall, 2008). The geological evolution of Mexico in the Cenozoic was very complex, resulting in its rugged geomorphology and diverse climatic conditions (Ortega-Gutiérrez et al., 1992; Ferrusquía-Villafranca, 1993). Evolution of the Trans-Mexican Volcanic Belt started from the early to the middle Miocene (20–10 Ma) (Ferrari et al., 2012), while that of the Sierra Madre del Sur started during the Oligocene and still was active during the early Miocene (Mastretta-Yanes et al., 2015). The development of these volcanic arcs promoted an intense volcanic activity that had important effects in the orography (e.g., transformation of continental basins), hydrology (e.g., alteration of fluvial currents), and climate (e.g., modification in plant communities) of the Mexican Neogene (Cevallos-Ferriz and González-Torres, 2006).
The horse record (reported so far) indicates that the first members of Equinae occurred in areas of southern Mexico during the first half of the Miocene. The climate of southern Mexico varied from temperate to humid tropical at the beginning of the Miocene (González-Medrano, 1998). The palynoflora of Simojovel from the late Oligocene—early Miocene of Chiapas evidence this climatic variation, consisting of oak-pine woodland associated with mangrove forest (Rhizophora) (Graham, 1994, 1999) (Figure 4). The warmest temperatures recorded in the Neogene were reached during the middle Miocene (ca. 15 Ma); this episode has been called the “Mid-Miocene climatic optimum” (Zachos et al., 2001). In this regard, the palynological data recovered from the Ixtapa Formation, middle Miocene of Chiapas indicates the presence of tropical plant communities (mangrove forest of Rhizophora and Pelliceria) with some grasses (Monoporites) and gymnosperms (Pinus) (Martínez-Hernández, 1992). A progressive decrease in temperature occurred at the end of the middle Miocene and continued until the Pliocene (González-Medrano, 1998; Graham, 1999).
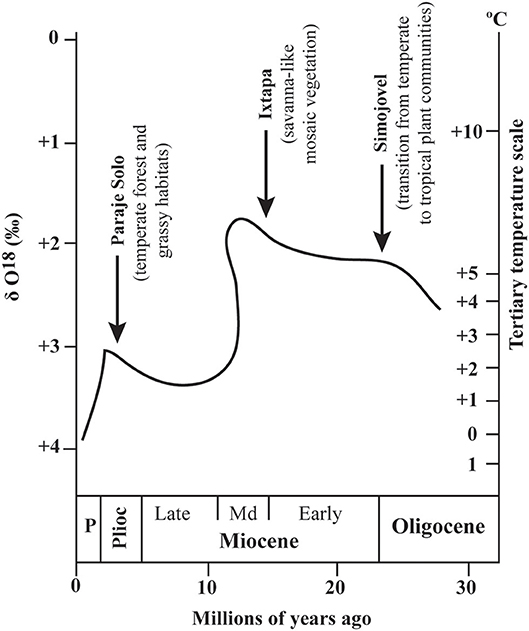
Figure 4. Neogene temperature change based on marine benthic δ O18 record. The arrows indicate representative palynofloras and related plant communities from the Mexican Neogene (modified from Graham, 1994; Figure 3).
The volcanoclastic deposits that preserved the Miocene horse communities of southern Mexico evidence at least two events of silicic explosive eruptions, at about 20 Ma (Suchilquitongo Formation) and at about 15 Ma (Matatlán, El Camarón, and Ixtapa formations). The volcanic activity spread from the Oaxaca Valley, the Tehuantepec Isthmian Region, and areas of central-western Chiapas, during the middle Miocene (Jiménez-Hidalgo et al., 2002). The volcanism has been associated with extensional tectonics that produced a complex basin and range topography, making up valleys (graben) flanked by highlands (horsts) (Ortega-Gutiérrez et al., 1992).
Climate conditions, plant communities, and geological features of southern Mexico in the first half of the Miocene (20–15 Ma) suggest that horses inhabited tropical lowland areas covered by heterogeneous vegetation. It is noted that mammalian composition of faunas from the middle Miocene of Oaxaca and Chiapas are related to savanna-like ecosystems, including browsers (e.g., gomphotheres, leptomerycids, and oreodonts), mixed-feeders (e.g., camelids and protoceratids), and grazers (e.g., rhinocerotids and antilocaprids) (Jiménez-Hidalgo et al., 2002).
The first peak of diversity in Equinae coincides with the warmest and humid conditions of the Mexican Neogene (Figure 4). The horse communities of the middle Miocene of southern Mexico include subhypsodont (mean molar crown height between 25 and 30 mm) species such as “Merychippus” cf. primus, “M.” cf. sejunctus, Merychippus cf. californicus, and Calippus sp. (Bravo-Cuevas, 2000; Bravo-Cuevas and Ferrusquía-Villafranca, 2006), as well as hypsodont (mean molar crown height >30 mm) species like Cormohipparion aff. quinni and Pliohippus sp. (Bravo-Cuevas, 2000; Bravo-Cuevas and Ferrusquía-Villafranca, 2008, 2010). This high species diversity is comparable to that reported from several Barstovian faunas in the United States, such as Norden Bridge of north-central Nebraska (Tedford et al., 2004), Eubanks of northeastern Colorado (Galbreath, 1953), and Cold Spring of Texas (Quinn, 1955).
Traditionally, the evolution of hypsodont molars in horses (and in other herbivore mammals) has been explained as a coevolutionary process in response to the expansion of grassland habitats during the middle Miocene (Webb, 1977, 1983). Nevertheless, isotopic evidence and mesowear data in North American fossil horses indicate that there is not a definitive correlation between the increase of the crown height with the shifting from browsing to grazing (Wang et al., 1994; Mihlbachler et al., 2011). Tooth enamel carbonate δ13C values in samples of “Merychippus” cf. sejunctus, Cormohipparion aff. quinni, and Pliohippus sp. from the middle Miocene of Oaxaca are characteristically <− 9 ‰ (Pérez-Crespo et al., 2016). This carbon isotopic evidence suggests that C3 plants were an important component of the terrestrial ecosystems of southern Mexico at about 15 Ma. Paleobotanical evidence and fossil horse communities have shown that C3 biomes were common in the United States during the radiation phase of the Equinae (from 18 to 15 Ma) (Hulbert, 1993). The information presented here indicates that comparable biomes also occurred in areas of southern tropical North America.
Given the above, the complicated physiography and the presence of savanna-like mosaic vegetation in a humid and tropical climate of southern Mexico would be an appropriate scenario for sustaining a diverse horse community (four to six sympatric species), allowing for suitable resource partitioning. By the same token, this high diversity coincides with the adaptive radiation of Miocene Equinae, suggesting that part of this evolutionary process could also have had occurred in areas of southern Mexico.
The first horses of the Mexican Neogene to become extinct were the merychippines. It seems that this group of horses preferred humid and forested environments and their disappearance probably was in response to the progressive cooling and aridification which occurred across North America from the late Miocene to the Pliocene (Thomasson and Voorhies, 1990; Potts and Behrensmeyer, 1992). Merychippines were eventually replaced by more hypsodont hipparionines, protohippines, and equines, as evidenced by the horse record from the Clarendonian of central Mexico, including members of the genera Hipparion, Protohippus, and Pliohippus (Figures 2, 3).
By the early late Miocene (ca. 9 Ma) a sudden temperature decrease in the Mexican territory occurred (González-Medrano, 1998) (Figure 4). This change in temperature is evidenced by the reduction of tropical plant communities and the arrival of boreal elements like the spruce (Picea) to areas of central Mexico at 18°N Lat. Furthermore, the presence of flora from temperate regions [e.g., pine (Podocarpus and Pinus), oak (Quercus), and sweetgum (Liquidambar)] in lowland areas (150–250 m.a.s.l.) of the Gulf Coastal Plain and southern Mexico was common (González-Medrano, 1998). The diversity dropped from six to three species together with the disappearance of the merychippines from the Barstovian to the Clarendonian (Figure 3). This decrease in diversity could have resulted from the climatic and vegetational changes at the beginning of the late Miocene, being the merychippines drastically affected. It should be stated that merychippines became extinct in temperate North America by the end of the Barstovian (Hulbert and MacFadden, 1991).
The aridification and cooling trend that occurred throughout the late Miocene certainly resulted in a progressive increase of open habitats in the Mexican territory, such as grasslands and prairies (Graham, 1993, 1994, 1999). In this regard, the mammalian communities from the Hemphillian of central Mexico were characterized by having hypsodont horses (e.g., Neohipparion eurystyle, Calippus hondurensis, and “Dinohippus” mexicanus). The presence of high crowned teeth in the Hemphillian Mexican horses allowed them to exploit abrasive food resources typical of grassy habitats.
In the Trans-Mexican Volcanic Belt of central Mexico, a mafic volcanic pulse gradually migrated eastwards from western Mexico, from the late Miocene to the early Pliocene (11–3 Ma), creating large plateaus and gradually increasing the elevation ranges due to isostatic movements (Ferrari et al., 2000; Orozco-Esquivel et al., 2003; Ferrari, 2004). From the latest Miocene to the early Pliocene (≈7.5–3.0 Ma), calderas and ignimbrites covered large areas, and domes complexes were formed in central-western Mexico (Ferrari et al., 2012).
The Trans-Mexican Volcanic Belt was affected by NW–SE trans-tensive regime during the Miocene and Pliocene, generating grabens and semigrabens in western Mexico; these depressions were filled with water, creating extensive lakes (Israde-Alcántara et al., 2010). Additionally, the formation of the Trans-Mexican Volcanic Belt isolated the Mexican Central Plateau, preventing the entry of humid air from the Pacific Ocean, thus promoting aridification of this area (Ferrari, 2017).
The species diversity of Mexican horses during the Clarendonian partially coincides with the early development of the Trans-Mexican Volcanic Belt during the early to the late Miocene (19–8 Ma) (Ferrari et al., 2012). During the late Miocene, a tropical climate with forested habitats, with a mean annual temperature above 20°C and a high mean annual precipitation were evidenced in some areas of central Mexico (Castañeda-Posadas et al., 2009); these favorable conditions could support a large amount of herbivore mammals, including horses. As it has been stated, the record of Clarendonian horses is poorly known, although it is probable that the true nature of the mammalian faunas that inhabited the Mexican territory during this faunal stage is biased, considering the dynamics of the Trans-Mexican Volcanic Belt. In this regard, the accretion of mafic lava across central Mexico, could have had promoted the uplift of sedimentary sequences deposited during the late Miocene, which have been eventually eroded.
The known record of Mexican Equinae indicates that reached their major diversity of five genera and eight contemporaneous species during the Hemphillian (at about 7 Ma). In this faunal stage, all groups of advanced hypsodont horses displayed their greatest species diversity, being the equines the most diverse. By the late early Hemphillian (≈7.0 Ma) humid conditions allowed the basins of the Trans-Mexican Volcanic Belt to begin filling with water, creating several lakes (Israde-Alcántara et al., 2010). The mammalian associations recovered from the localities of central Mexico of this age and microfossil proxies suggest a woodland landscape under warm conditions (Israde-Alcántara et al., 2010; Carranza-Castañeda et al., 2013). The available resources during this age allowed for a great diversity of horses, showing its second diversity peak in Mexico (Figure 3).
Diversity of Mexican Equinae in the Pliocene
Between 5.2 and 3.6 Ma there was an increase in oceanic levels and warm temperatures at the marine surface, which promoted a high precipitation levels on land; the climate was warm and there was a maximum north-south tectonic extension in several areas along the Trans-Mexican Volcanic Belt (Israde-Alcántara et al., 2010).
By the latest Hemphillian -around 4.8 Ma- there was a large mammalian extinction event in North America (Webb, 1984), which is also observed in several local faunas of central Mexico (Jimenez-Hidalgo, 2005), including horses (Figure 3). This extinction event could be related to a cooler and drier climate, and to the emergence of more open vegetation in temperate North America (Prothero, 2006). It seems that this extinction could be correlated with volcanic activity, which disrupted plant communities, and the further uplift of the Trans-Mexican Volcanic Belt, which resulted in further aridification and the spreading of more open vegetation compared to the Miocene.
In the Blancan, only two horse species are recorded after the disappearance of the majority of the Hemphillian hipparionines and equines (Figure 3). Between 3.3 and 2.0 Ma, the oceans became cooler, with stronger cycles by 2 Ma, and there were more arid conditions in central and northern Mexico. The lakes from central Mexico became shallower and some disappeared (Israde-Alcántara et al., 2010). In central Mexico, mammalian assemblages of Blancan age indicate more open vegetation when compared to Hemphillian ones (Carranza-Castañeda, 2006). In east-central Mexico, within the Trans-Mexican Volcanic Belt, some floral assemblages indicate mesothermal woodland with a temperate climate (Velasco-de León et al., 2010). Furthermore, the floristic composition of the Paraje Solo Formation from the Pliocene of Veracruz, east-central Mexico, evidences an increase in diversity and abundance of temperate to cold elements, including pine-oak forest (Abies, Picea, and Pinus), deciduous forest (Psilotum, Lycopodium, and Sphaeropteris), and perennial forest (Podocarpus and Alchornea) (Graham, 1993, 1994; González-Medrano, 1998) (Figure 4).
Taxonomic Evolution of Mexican Neogene Horses
The overall mean species duration of selected Mexican Neogene horses is 1.79 m.y., ranging from 1.2 m.y. (“Merychippus” cf. primus, “M.” cf. sejunctus, and Cormohipparion aff. quinni) to 2.7 m. y. (Neohipparion eurystyle and “Dinohippus” mexicanus) (Table 2). The equines showed the highest average species duration with 1.95 m.y., whereas the hipparionines have average species duration of 1.85 m.y. The considered merychippines have species durations of 1.2 m.y. (Table 2). The Mexican record of Calippus hondurensis extends the biochronological range of this species of about 2 million years, similar to that observed in other calippines, such as C. elachistus from the late Clarendonian—early Hemphillian of central Florida, as well as C. regulus from the late Barstovian—Clarendonian of the Great Plains and early Clarendonian of the Texas Gulf Coastal Plain (Hulbert, 1988a).
A comparison of the temporal range of selected hipparionines and equines from Mexico with those from the United States indicates that these horses showed similar durations in North America during the Neogene (Table 3). The average temporal duration of the selected Mexican Equinae is near the lower limit of the range of the mean species duration of North American Cenozoic mammals of ~2–3 million years and it is lower than the average temporal duration of Equidae that is of 2.9 million years (Prothero, 2014).
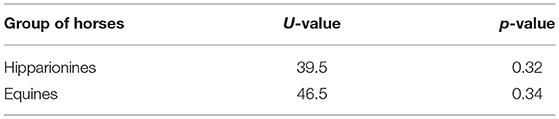
Table 3. Results of the Mann-Whitney U-Tests (p < 0.05) of the temporal duration of the Mexican and North American hipparionines and equines.
The merychippines “Merychippus” cf. primus and “M.” cf. sejunctus are characterized by having short temporal durations of 1.2 m.y., suggesting that they had a rapid evolution in the early to middle Miocene. The long temporal durations of the best documented hipparionines and equines of about 2.0 m.y., suggest that they could have evolved more slowly after their diversification in the middle to late Miocene and persisting until the end of the Neogene. It has been shown that horses have evolved rapidly during the Cenozoic (Simpson, 1984; MacFadden, 1992; Hulbert, 1993; Franzen, 2010). The data presented here provide additional information in this regard.
Comment on the Adaptive Radiation of North American Equinae
It has been largely recognized that the adaptive radiation of North American Equinae started at about 18 Ma in particular areas of the United States (MacFadden and Hulbert, 1988; Hulbert and MacFadden, 1991; MacFadden, 1992; Hulbert, 1993; Maguire and Stigall, 2008). At least four species including primitive merychippines (“Merychippus gunteri” and “M.” primus) and equines (Acritohippus tertius and Parapliohippus carrizoensis) have been reported from the late Hemingfordian of the Coast of California, the Great Plains, and the Gulf Coast (Bryant, 1988, 1991; Hulbert and MacFadden, 1991; Kelly, 1995, 1998). The adaptive radiation continued in the same areas during the Barstovian at about 15 Ma, leading to the differentiation of earliest representatives of the advanced hypsodont genera Pliohippus, Calippus, Protohippus, and Cormohipparion (Hulbert, 1993).
In Mexico, there is a record referable to “Merychippus” sp. from the late Hemingfordian of Oaxaca (Ferrusquía-Villafranca, 1990). By the late early Barstovian, the Mexican horse communities were composed by basal merychippines (“Merychippus” cf. “M.” primus and “M.” cf. “M.” sejunctus) and primitive populations of hipparionines (Cormohipparion aff. quinni), protohippines (Calippus sp.), and equines (Pliohippus sp.), which inhabited humid and tropical areas that now are part of the states of Oaxaca and Chiapas, southern Mexico (Bravo-Cuevas, 2000; Bravo-Cuevas and Ferrusquía-Villafranca, 2006, 2008, 2010).
This information suggests that the adaptive radiation of Equinae would have occurred in areas of temperate and tropical North America. In this scenario, the evolutionary process occurred in separate areas at least 2, 000 km apart and in different biogeographic regions. Two hypotheses may explain this pattern as it is indicated in the following paragraphs.
(1) The differentiation occurred throughout North America, including tropical areas of southern Mexico. This scenario is less probable because it requires genetically continuous but geographically disperse populations. Furthermore, the known record of Equinae from the Hemingfordian—Barstovian of North America is unevenly distributed in different areas of the United States (Maguire and Stigall, 2008), and southern Mexico as well.
(2) The differentiation took place in areas of temperate and tropical North America, implying short temporal dispersals after a speciation event. This hypothesis appears more likely because the geographic distribution of Equinae shows some provinciality and speciation by dispersal was common in the evolution of the group at the base of their adaptive radiation during the middle Miocene (Maguire and Stigall, 2008).
It seems that the Mexican record evidences part of the radiation phase of the North American Equinae, which in turn indicates a somewhat synchronous appearance of primitive hipparionines, protohippines, and equines in temperate and tropical regions of North America during the early—middle Miocene. The proposed scenario implies regional dispersal events within the North American subcontinent, similarly to the Hipparion and Equus datum planes that involved dispersal events between North America and Eurasia during the late Neogene (Woodburne et al., 1981; Lindsay et al., 1984; Bernor et al., 1989).
Comment on the Origin of Equus
The species “Dinohippus” mexicanus and Equus simplicidens are closely related taxa (MacFadden, 1984b, 1986). Hulbert (1993) considered that, by anagenetic evolution, Equus descended from “D.” mexicanus. Some years later, MacFadden and Carranza-Castañeda (2002) commented on the horse record from Anza-Borrego desert in California, where Downs and Miller (1994) described a cf. Dinohippus specimen of late Blancan age (≈2.7 Ma) associated with specimens of Equus simplicidens; they also reported some isolated teeth with a transitional morphology of equine horses pertaining to Dinohippus and/or Equus from the San Buenaventura Formation (late Blancan, 2.62 Ma) in the Tecolotlán area of Jalisco state, west-central Mexico (Kowallis et al., 2017); some of these teeth can be identified as D. mexicanus, while others can be referred to primitive Equus (E. cf. simplicidens). Both records indicate that primitive Equus originated from “Dinohippus” mexicanus by means of cladogenesis (MacFadden and Carranza-Castañeda, 2002). The geographic locations of both localities with these coetaneous taxa suggest that this cladogenetic event probably took place in western North America, including areas of central Mexico.
Conclusions
The record of Mexican Equinae provides additional support on their evolutionary history during the Neogene, characterized by having two peaks of diversity during the Miocene and a progressive decline by the Pliocene. The change in species diversity is largely related to the climatic and vegetational change occurred during the Neogene in areas of temperate and tropical North America.
The somewhat short temporal duration showed by some Mexican Equinae (ca. 1.6 m. y.) suggests that they evolved rapidly during the Miocene, although the hipparionines and equines probably evolved more slowly after their diversification in the middle to late Miocene, persisting to the end of the Pliocene.
The members of Equini and Hipparionini were dominant throughout much of the Mexican Neogene and, to a lesser extent, the Protohippini. The Mexican merychippines were restricted to the first half of the Miocene (from 18 to 15 Ma) at the base of the adaptive radiation of the North American Equinae.
The taxonomic composition of the diverse horse communities from the Barstovian of Oaxaca and Chiapas, southern Mexico, integrated by basal merychippines (“Merychippus” cf. primus and “M.” cf. sejunctus) and primitive populations of advanced hypsodont hipparionines (Cormohipparion aff. quinni), protohippines (Calippus sp.), and equines (Pliohippus sp.), suggests that part of the radiation phase of Equinae could also have occurred in areas of tropical North America.
The species “Dinohippus” mexicanus was erected by Mexican fossil material, and it has been considered as the species that gave rise by cladogenesis to primitive Equus. The coexistence of “D.” mexicanus and E. simplicidens in areas of central Mexico indicates that part of the differentiation of Equus took place in Mexican territory.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We appreciate the comments and suggestions of the three reviewers, which greatly improved the final version of the manuscript. Both authors thank the editors Gina Semprebon, Florent Rivals, Ray Bernor, Florent Rivals, Leo Avilla, and Eric Scott, for their kind invitation to participate in the present volume. Thanks to Nuria Melisa Morales García (University of Bristol, UK) for the linguistical review of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00287/full#supplementary-material
References
Azzaroli, A. (1992). Ascent and decline of monodactyl equids: a case for prehistoric overkill. Ann. Zool. Fennici 28, 151–163.
Bell, C. J., Lundelius, E. L. Jr., Barnosky, A. D., Graham, R. W., Lindsay, E. H., Ruez, D. R. Jr., et al. (2004). The Blancan, Irvingtonian, and Rancholabrean mammal ages, in Late Cretaceous and Cenozoic mammals of North America, ed. M. O. Woodburne (New York, NY: Columbia University Press), 232–314.
Bernor, R. L., Tobien, H., and Woodburne, M. (1989). “Patterns of the Old World hipparionine evolutionary diversification and biogeographic extension,” in European Neogene mammal chronology, eds. E. H. Lindsay, R. L. Bernor, V. Fahlbusch, and P. Mein (New York, NY: Plenum Press, 263–319.
Bravo-Cuevas, V. M. (2000). Los équidos mesomiocénicos del estado de Oaxaca: Taxonomía e implicaciones paleobiológicas. [dissertation/MSc. Thesis]. [Ciudad de México, México]: Facultad de Ciencias, Universidad Nacional Autónoma de México
Bravo-Cuevas, V. M., and Ferrusquía-Villafranca, I. (2006). Merychippus (Mammalia, Perissodactyla, Equidae) from the Middle Miocene of the State of Oaxaca, Southeastern Mexico. Geobios-Lyon 39, 771–784. doi: 10.1016/j.geobios.2005.10.006
Bravo-Cuevas, V. M., and Ferrusquía-Villafranca, I. (2008). Cormohipparion (Mammalia, Perissodactyla, Equidae) From the middle Miocene of Oaxaca, southeastern México. J. Vertebr. Paleontol. 28, 243–250. doi: 10.1671/0272-4634(2008)28[[243:CMPEFT]]2.0.CO;2
Bravo-Cuevas, V. M., and Ferrusquía-Villafranca, I. (2010). The oldest record of Equini (Mammalia: Equidae) from Mexico. Rev. Mex. Cienc. Geol. 27, 593–602.
Bryant, J. D. (1988). New Miocene vertebrate localities and marine-nonmarine correlations in the eastern Florida Panhandle. J. Vertebr. Paleontol. 8:10A.
Bryant, J. D. (1991). New Early Barstovian (Middle Miocene) verterbrates from the upper Torreya Formation, eastern Florida Panhandle. J. Vertebr. Paleontol. 11, 472–489. doi: 10.1080/02724634.1991.10011416
Carranza-Castañeda, O. (2006). “Late Tertiary fossil localities in central México between 19° 23°N,” in Advances in late Tertiary vertebrate Paleontology in Mexico and the Great American Biotic Interchange, eds. O. Carranza-Castañeda, and E. H. Lindsay (México: Universidad Nacional Autónoma de México, Instituto de Geología and Centro de Geociencias, 45–60.
Carranza-Castañeda, O., Aranda-Gómez, J. J., Wang, X., and Iriondo, A. (2013). The early-late Hemphillian (Hh2) Fauna assamblage from Juchipila Basin, State of Zacatecas, México, and its Biochronologic correlation with other Hemphillian Faunas in Central México. Contrib. Sci. 521, 13–49.
Carranza-Castañeda, O., and Miller, E. W. (1999). A new terrestrial vertebrate locality from Southern Baja California. J. Vertebr. Paleontol. 19:36.
Carroll, R. L. (1998). Patterns and processes of vertebrate evolution. New York: Cambrige University Press (Cambridge Paleobiology Series).
Castañeda-Posadas, C., Calvillo-Canadell, L., and Cevallos-Ferriz, S. R. S. (2009). Woods from Miocene sediments in Panotla, Tlaxcala, Mexico. Rev. Palaeobot. Palynol. 156, 494–506. doi: 10.1016/j.revpalbo.2009.04.013
Cevallos-Ferriz, S. R. S., and González-Torres, E. A. (2006). “Geological setting and phytodiversity in Mexico,” in Studies on Mexican Paleontology, eds. F. Vega, T. G. Nyborg, M. C. Perrillat, M. Montellano-Ballesteros, S. R. S. Cevallos-Ferris, and S. A. Quiroz-Barroso (Dordrecht: Springer, 1–18.
Downs, T. (1961). A study of variation and evolution in Miocene Merychippus. Contrib. Sci. 45, 1–75.
Downs, T., and Miller, G. J. (1994). Late Cenozoic equids from the Anza-Borrego Desert of California. Contrib. Sci. 440, 1–90.
Ferrari, L. (2004). Slab detachment control on mafic volcanic pulse and mantle heterogeneity in central Mexico. Geology 32, 77–80. doi: 10.1130/G19887.1
Ferrari, L. (2017). “Geología y biodiversidad. Una evolución paralela,” in Paleobiología, Interpretando procesos de la vida pasada, Coords. eds. S.R.S. Cevallos-Ferriz, and A. R. Huerta-Vergara (México: Universidad Nacional Autónoma de México, Facultad de Ciencias, Dirección General de Publicaciones y Fomento Editorial, Instituto de Geología, 237–253.
Ferrari, L., Conticelli, S., Vaggelli, C., Petrone, C., and Manetti, P. (2000). Late Miocene mafic volcanism and intra-arc tectonics during the early development of the Trans-Mexican Volcanic Belt. Tectonophysics 318, 161–185. doi: 10.1016/S0040-1951(99)00310-8
Ferrari, L., Orozco-Esquivel, T., Manea, V., and Manea, M. (2012). The dynamic history of the Trans-Mexican Volcanic Belt and the Mexico subduction zone. Tectonophysics 522–523, 122–149. doi: 10.1016/j.tecto.2011.09.018
Ferrusquía-Villafranca, I. (1990). Biostratigraphy of the Mexican continental Miocene: part II, The southeastern (Oaxacan) faunas. Paleontol. Mex. 56, 57–109.
Ferrusquía-Villafranca, I. (1992). Contribución al conocimiento del Cenozoico en el Sureste de México y de su relevancia en el entendimiento de la evolución geológica regional. VIII Congreso Geológico Latinoamericano Salamanca España 4, 40–44.
Ferrusquía-Villafranca, I. (1993). “Geology of Mexico. A synopsis,” in Biological Diversity of Mexico-Origins and Distribution, eds. T. P. Ramamoorthy, R. A. Bye, and A. Loct (New York, NY: Oxford Univeristy Press, 3–107.
Ferrusquía-Villafranca, I. (1996). Contribucion al conocimiento geológico de Chiapas: El área Ixtapa Soyaló. Inst. Geol. Bol. UNAM 109, 1–130.
Ferrusquía-Villafranca, I. (2001). Contribucion al conocimiento geológico de Oaxaca: El área Nejapa de Madero. Inst. Geol. Bol. UNAM 111, 1–110.
Ferrusquía-Villafranca, I., Ruiz-González, J. E., Martínez-Hernández, E., Torres-Hernández, J. R., and Woolrich-Piña, G. (2014). A new Miocene local fauna from the Sierra Madre Oriental at San Luis Potosí, Central-East Mexico, and its paleontologic significance. Geobios-Lyon 47, 199–220. doi: 10.1016/j.geobios.2014.06.004
Galbreath, E. C. (1953). A contribution to the Tertiary geology and Paleontology of northeastern Colorado. Paleontol. Contrib. Univ. Kansas 4, 1–120.
Graham, A. (1993). “Historical factors and biological diversity of México,” in Biological Diversity of Mexico-Origins and Distribution, eds. T. P. Ramamoorthy, R. A. Bye, and A. Loct (New York, NY: Oxford Univeristy Press, 109–117.
Graham, A. (1994). Neogene Palynofloras and Terrestrial Paleoenvironments in Northern Latin America. USGS Open-File Report.
Graham, A. (1999). Late Cretaceous and Cenozoic History of North American Vegetation. New York, NY: Oxford University Press.
Hulbert, R. C., and MacFadden, B. J. (1991). Morphological transformation and cladogenesis at the base of the adaptive radiation of Miocene hypsodont horses. Am. Mus. Novit. 3000, 1–61.
Hulbert, R. C. Jr. (1987). Late Neogene Neohipparion (Mammalia, Equidae) from the Gulf Coastal Plain of Florida and Texas. J. Paleontol. 61, 809–830. doi: 10.1017/S0022336000029152
Hulbert, R. C. Jr. (1988a). Calippus and Protohippus (Mammalia, Perisodactyla, Equidae) from the Miocene (Barstovian-Early Hemphillian) of the Gulf Coastal Plain. Bull. Fla. Mus. Nat. Hist. 32, 221–340.
Hulbert, R. C. Jr. (1988b). Cormohipparion and Hipparion (Mammalia, Perisodactyla, Equidae) from the late Neogene of Florida. Bull. Fla. Mus. Nat. Hist. 33, 229–338.
Hulbert, R. C. Jr. (1989). “Phylogenetic interrelationships and evolution of North American Late Neogene Equinae,” in The evolution of Perissodactyla, eds. D. R. Prothero, and R. M. Schoch (New York, NY: Oxford University Press, 176–193.
Hulbert, R. C. Jr. (1993). Taxonomic evolution in North American Neogene horses (Subfamily Equinae): the rise and fall of an adaptive radiation. Paleobiology 19, 216–234. doi: 10.1017/S0094837300015888
Israde-Alcántara, I., Miller, W. E., Garduño-Monroy, V. H., Barron, J., and Rodriguez-Pascua, M. A. (2010). Palaeoenvironmental significance of diatom and vertebrate fossils from Late Cenozoic tectonic basins in west-central México: a review. Quaternary Int. 219, 79–94. doi: 10.1016/j.quaint.2010.01.012
Jimenez-Hidalgo, E. (2005). Bioestratigrafía de los artiodáctilos (Tylopoda y Ruminantia) de las faunas del Terciario tardío de la Faja Volcánica Transmexicana [dissertation/ PhD. Thesis] [Cd. de México]: Universidad Nacional Autónoma de México.
Jiménez-Hidalgo, E., Ferrusquía-Villafranca, I., and Bravo-Cuevas, V. M. (2002). “El registro mastofaunístico miocénico de México y sus implicaciones paleobiológicas,” in Avances en los Estudios Paleomastozoológicos, eds. M. Montellano-Ballesteros, and J. Arroyo-Cabrales (México: Instituto Nacional de Antropología e Historia, 47–58.
Kelly, T. S. (1995). New Miocene horses from the Caliente Formation, Cuyama Valley Badlands, California. Contrib. Sci. 455, 1–33.
Kelly, T. S. (1998). New Middle Miocene equid crania from California and their implications for the phylogeny of equini. Contrib. Sci. 473, 1–44.
Kelly, T. S., and Stewart, J. D. (2008). New records of middle and late Miocene Perissodactyla and Artiodactyla from the Western border of the San Joaquin Valley, Diablo Range, Fresno County, California. Contrib. Sci. 516, 1–32.
Kowallis, B. J., Christiansen, E. H., Carranza-Castañeda, O., Miller, W. E., Ross, K. T., and Tingey, D. G. (2017). The geology of the tecolotlán Graben, Jalisco, Mexico. Geol. Soc. Am. Digital Map Chart. 22, 1–37. doi: 10.1130/2017.DMCH022.TXT
Kowallis, J. B., Swisher, C. C., Carranza-Castañeda, O., Miller, E. W., and Tingey, G. D. (1999). Fission-track and single crystal 40 AR/39AR laser fusion ages from volcanic ash layers in fossil-bearing Pliocene sediments in central México. Rev. Mex. Cienc. Geol. 15, 157–160.
Lance, J. F. (1950). Paleontología y estratigrafía del Plioceno de Yepómera Estado de Chihuahua. Parte 1: equidos, excepto Neohipparion. Inst. Geol. Bol. UNAM 54, 1–81.
Lindsay, E. H., Opdyke, N. D., and Johnson, N. M. (1984). Blancan- Hemphillian land mammal ages and late Cenozoic mammal dispersal events. Ann. Rev. Earth Pl. Sc. 12, 445–488. doi: 10.1146/annurev.ea.12.050184.002305
MacFadden, B. J. (1984a). Systematics and phylogeny of Hipparion, Neohipparion, Nannippus, and Cormohipparion (Mammalia, Equidae) from the Miocene and Pliocene of the New World. Am. Mus. Novit. 179, 1–196.
MacFadden, B. J. (1984b). Astrohippus and Dinohippus from the Yepómera local fauna (Hemphillian, Mexico) and implications for the phylogeny of one-toed horses. J. Vertebr. Paleontol. 4, 273–283. doi: 10.1080/02724634.1984.10012009
MacFadden, B. J. (1986). Late Hemphillian monodactyl horses (Mammalia, Equidae) from the Bone Valley Formation of Central Florida. J. Paleontol. 60, 466–475. doi: 10.1017/S0022336000021995
MacFadden, B. J. (1988). Horses, the fossil record, and evolution. A current perspective. Evol. Biol. 22, 131–158. doi: 10.1007/978-1-4613-0931-4_3
MacFadden, B. J. (1992). Fossil Horses: Systematics, Paleobiology, and Evolution of the Family Equidae. New York, NY: Cambridge University Press.
MacFadden, B. J. (1998). “Equidae,” in Evolution of Tertiary Mammals of North America (Vol. 1): Terrestrial carnivores, ungulates, and Ungulatelike mammals, eds. C. M. Janis, K. M. Scott, and L. L. Jacobs (New York, NY: Cambridge University Press, 537–559.
MacFadden, B. J. (2006). “Early Pliocene (latest Hemphillian) horses from the Yepómera Local Fauna, Chihuahua, Mexico,” in Advances in Late Tertiary Vertebrate Paleontology in Mexico and the Great American Biotic Interchange eds O. Carranza-Castañeda and E. H. Lindsay (México: Universidad Nacional Autónoma de México, Instituto de Geología and Centro de Geociencias, 33–43.
MacFadden, B. J., and Carranza-Castañeda, O. (2002). Cranium of Dinohippus mexicanus (Mammalia: Equidae): from the early Pliocene (latest Hemphillian) of central Mexico, and the origin of Equus. Bull. Fla. Mus. Nat. Hist. 43, 163–185.
MacFadden, B. J., and Hulbert, R. C. Jr. (1988). Explosive speciation at the base of the adaptive radiation of Miocene grazing horses. Nature 336, 466–468. doi: 10.1038/336466a0
Maguire, C. K., and Stigall, A. L. (2008). Paleobiogeography of Miocene Equinae of North America: a phylogenetic biogeographic analysis of the relative roles of climate, vicariance, and dispersal. Palaeogeogr. Palaeocl. 267, 175–184. doi: 10.1016/j.palaeo.2008.06.014
Martínez-Hernández, E. (1992). Caracterización ambiental del Terciario de la región de Ixtapa, Estado de Chiapas-un enfoque palinoestratigráfico. Inst. Geol. Rev. UNAM 10, 54–64.
Mastretta-Yanes, A., Moreno-Letelier, A., Piñero, D., Jorgensen, T. H., and Emerson, B. C. (2015). Biodiversity in the Mexican highlands and the interaction of geology, geography and climate within the Trans-Mexican Volcanic Belt. J. Biogeogr. 42, 1586–1600. doi: 10.1111/jbi.12546
Mihlbachler, M. C., Solounias, N., Rivals, F., and Semprebon, G. (2011). Dietary change and evolution of horses in North America. Science 331, 1178–1181. doi: 10.1126/science.1196166
Miller, W. E. (1980). The Late Pliocene Las Tunas local fauna from southernmost Baja California, Mexico. J. Paleontol. 54, 762–805
Mooser, O. (1968). Fossil equidae from the middle pliocene of the central plateau of Mexico. Southwest Nat. 13, 1–12 doi: 10.2307/3668809
Orozco-Esquivel, M. T., Ferrari, L., Tagami, T., Eguchi, M., and Petrone, C. (2003). The Eastern Alkaline Province (Mexico) Revised: Geology, Geochronologyand Geochemistry of Neogene Volcanism in Veracruz State. GSA Cordilleran Section Meeting, Paper No. 21–4, 58.
Ortega-Gutiérrez, F., Mitre Salazar, M. L., Roldán-Quintana, J., Aranda-Gómez, J. J., Morán-Zenteno, B., Alaniz-Alvarez, J. A., et al. (1992). Carta geológica de la Repúlica Mexicana, Esc. 1: 2 000 000, 5a. ed. y texto explicativo. Universidad Nacional Autónoma de México, Instituto de Geología y Secretaría de Minas Industria Paraestatal Consejo Recursos Minerales (Gainesville, FL), 74.
Osborn, H. F. (1918). Equidae of the Oligocene, Miocene and Pliocene of North America, iconographic type revision. Am. Mus. Nat. Hist. 2, 1–217. doi: 10.5962/bhl.title.84425
Pérez-Crespo, V. A., Ferrusquía-Villafranca, I., Bravo-Cuevas, V. M., Morales-Puente, P., and Ruiz-González, J. E. (2016). Dietary analysis of Late Cenozoic Mexican equids from three different geographic/geologic settings using stable carbon isotopes: coincidences, differences and paleobiologic significance. J. S. Am. Earth Sci. 66, 97–109. doi: 10.1016/j.jsames.2015.11.015
Potts, R., and Behrensmeyer, A. K. (1992). “Late Cenozoic terrestrial ecosystems,” in Terrestrial Ecosystems Through Time Evolutionary Paleoecology of Terrestrial Plants and Animals, eds. A. K. Behrensmeyer, J. D. Damuth, W. A. DiMichele, R. Potts, S. Hans-Dieter, and J. L. Wing (Chicago: University of Chicago Press, 419–519.
Priego-Vargas, J., Bravo-Cuevas, V. M., and Jiménez-Hidalgo, E. (2016). The record of Cenozoic horses in Mexico: current knowledge and palaeobiological implications. Palaeobio. Palaeoenv. 96, 305–331. doi: 10.1007/s12549-015-0223-y
Prothero, D. R. (2006). After the Dinosaurs: the Age of Mammals. Bloomington, IN: Indiana University Press.
Prothero, D. R. (2014). Species longevity in North American fossil mammals. Integr. Zool. 9, 383–393. doi: 10.1111/1749-4877.12054
Robinson, C., Campbell, T. L., Cote, S., and de Ruiter, D. J. (2018). Temporal ranges and ancestry in the hominin Fossil record: the case of Australopithecus sediba. S. Afr. J. Sci. 114, 1–7. doi: 10.17159/sajs.2018/20170327
Robles-Rivera, J. C. (2015). Mastofaunula del Clarendoniano tardío-Henfiliano temprano (Mioceno tardío), del área de Neutla, Municipio de Comonfort, estado de Guanajuato, México. [dissertation/MSc. Thesis]. [Cd. de México]: Universidad Nacional Autónoma de México.
Robles-Rivera, J. C., and Carranza-Castañeda, O. (2012). Primer registro de Mastofaunula del Clarendoniano tardío-Henfiliano temprano (Mioceno inferor) en el centro de México en el área de Neutla- Empalme de Escobedo, Municipio de Comonfort, Guanajuato. Geos 32, 96–98.
Stirton, R. A. (1955). Two new species of the equid genus Neohipparion from the Middle Pliocene, Chihuahua, Mexico. J. Paleontol. 29, 886–902.
Tedford, R. H., Albright, L. B., Barnosky, A. D., Ferrusquía-Villafranca, I., Hunt, R. M. Jr., Storer, J. E., Whistler, D. P., et al. (2004). “Mammalian biochronology of the Arikareean through Hemphillian interval (late Oligocene through early Pliocene epochs),” in Late cretaceous and Cenozoic mammals of North America: Biostratigraphy and geochronology, ed. M. Woodburne (New York, NY: Columbia University Press, 169–231.
Thomasson, J. R., and Voorhies, M. R. (1990). “Grasslands and grazers,” in Palaeobiology: A Synthesis, eds. D. E. G. Briggs and P. R. Crowther (Oxford: Blackwell Scientific, 84–87.
Urrutia-Fucugauchi, J., and Ferrusquía-Villafranca, I. (2001). Paleomagnetic results for the Middle-Miocene continental Suchilquitongo Formation, Valley of Oaxaca, southeastern Mexico. Geofis. Int. 40, 191–205.
Velasco-de León, M. P., Spicer, R. A., and Steart, D. C. (2010). Climatic reconstruction of two Pliocene floras from Mexico. Palaeobio. Palaeoenv. 90, 99–110 doi: 10.1007/s12549-010-0022-4
Wang, Y., Cerling, T. E., and MacFadden, B. J. (1994). Fossil horses and carbon isotopes: new evidence for Cenozoic dietary, habitat, and ecosystem changes in North America. Palaeogeogr. Palaeocl. 107, 269–280. doi: 10.1016/0031-0182(94)90099-X
Webb, S. D. (1977). A history of savanna vertebrates in the New World. Part I: North America. Annu. Rev. Ecol. Syst. 8, 355–380. doi: 10.1146/annurev.es.08.110177.002035
Webb, S. D. (1983). “The rise and fall of the Late Miocene ungulate fauna in North America,” in Coevolution, ed. M. H. Nitecki (Chicago: University Chigago Press, 267–306.
Webb, S. D. (1984). “Ten million years of mammal extinctions in North America,” in Quaternary Extinctions, eds. P. S., Martin, and R. G. Klain (Tucson: University of Arizona Press), 189–210.
Webb, S. D., Hulbert, R. C. Jr., and Lambert, W. D. (1995). “Climatic implications of large-herbivore distributions in the Miocene of North America,” in Paleoclimate and Evolution with Emphasis on Human Origins, eds. E. S. Vrba, G. H. Denton, T. C. Partridge, and L.H. Burckle (New Haven, CT: Yale University Press, 91–108.
Woodburne, M. O. (1996). Reappraisal of the Cormohipparion from the Valentine Formation, Nebraska. Am. Mus. Novit. 3163, 1–56.
Woodburne, M. O., MacFadden, B. J., and Skinner, M. F. (1981). The North American “Hipparion” datum, and implications for the Neogene of the New World. Geobios-Lyon 14, 1–32. doi: 10.1016/S0016-6995(81)80124-6
Keywords: species diversity, taxonomic evolution, adaptive radiation, horses, Neogene, Mexico
Citation: Bravo-Cuevas VM and Jiménez-Hidalgo E (2019) Evolutionary Significance of Equinae From the Mexican Neogene. Front. Ecol. Evol. 7:287. doi: 10.3389/fevo.2019.00287
Received: 17 January 2019; Accepted: 15 July 2019;
Published: 31 July 2019.
Edited by:
Leonardo Dos Santos Avilla, Universidade Federal do Estado do Rio de Janeiro, BrazilReviewed by:
Helena Machado, University of Oregon, United StatesKaroliny De Oliveira Nascimento, Universidade Federal do Estado do Rio de Janeiro, Brazil
Marisol Montellano, National Autonomous University of Mexico, Mexico
Copyright © 2019 Bravo-Cuevas and Jiménez-Hidalgo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Victor Manuel Bravo-Cuevas, dm1iYzFAeWFob28uY29t